
Transient reactivation of CSF in parthenogenetic one-cell mouse embryos

+ Models
THE-10884; No of Pages 12
Ultrastructural characterization of fresh and cryopreserved
in vivo produced ovine embryos
E.M.V. Bettencourt a,*, C.M. Bettencourt b, J.N.C.E. Silva c, P. Ferreira d,C.P. de Matos b, E. Oliveira e, R.J. Romao f, A. Rocha d, M. Sousa e
a Department of Veterinary and Institute of Mediterranean Agricultural Sciences (ICAM), University of Evora, 7000-092 Evora, Portugalb Regional Agriculture Direction of Alentejo (DRAAL), Herdade da Abobada, V.N.S. Bento, Portugal
c Ex-Division for Animal Selection and Reproduction (DSRA), and Veterinary General Direction (DGV), Amadora, Portugald Campus Agrario de Vairao, University of Porto, Portugal
e ICBAS-Lab Cell Biology, Institute of Biomedical Sciences Abel Salazar, University of Porto, Portugalf Department of Zootechny, and Institute of Mediterranean Agricultural Sciences (ICAM), University of Evora, Evora, Portugal
Received 23 May 2008; received in revised form 22 September 2008; accepted 27 October 2008
Abstract
Controlled slow freezing and vitrification have been successfully used for ovine embryo cryopreservation. Selection of embryos
for transfer is based on stereomicroscopical embryo scoring after thawing, but the subjectivity inherent to this selection step has
been demonstrated by ultrastructural studies of controlled slow frozen, in vivo produced ovine morulae and blastocysts. These
studies have shown that certain abnormalities remain undetected by stereomicroscopy only. In the present study, using ovine in vivo
produced morulae and blastocysts, we have studied the ultrastructural alterations induced by open pulled straw vitrification (OPS)
and controlled slow freezing, compared stereomicroscopical embryo scoring with light microscopy evaluation of embryo’s semithin
sections, and related the ultrastructural cellular damage with the embryo classification by stereomicroscopical embryo scoring of
embryos’ and semithin section evaluation by light microscopy. The ultrastructural lesions found for OPS-vitrified and controlled
slow frozen embryos were similar, independently of embryo stage. A significant higher number of grade 3 embryos was found at
stereomicroscopical scoring after controlled slow freezing (P = 0.02), and a significant higher number of grade 3 blastocysts was
found at semithin sectioning after OPS vitrification (P = 0.037). The extension of ultrastructural damage, especially of
mitochondria and cytoskeleton, was related to the semithin classification but not to stereomicroscopical scoring at thawing. This
suggests that semithin scoring is a useful tool for predicting ultrastructural lesions and new improvements in cryopreservation and
thawing methods of ovine embryos are still warranted, including in the case of blastocysts cryopreserved by OPS vitrification.
# 2008 Elsevier Inc. All rights reserved.
Keywords: Cryopreservation; OPS vitrification; Ovine embryo morphology; Ultrastructure
www.theriojournal.com
Available online at www.sciencedirect.com
Theriogenology xxx (2008) xxx–xxx
Please cite this article in press as: Bettencourt EMV, et al. Ultras
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.t
* Corresponding author. Tel.: +351 266760809;
fax: +351 266760944.
E-mail address: [email protected] (E.M.V. Bettencourt).
0093-691X/$ – see front matter # 2008 Elsevier Inc. All rights reserved.
doi:10.1016/j.theriogenology.2008.10.019
1. Introduction
In order to increase the genetic gain resulting from
selection, efficient cryopreservation protocols are
required to allow delayed transfer of embryos and for
incrementing the use of multiple ovulation and embryo
transfer techniques [1,2]. Cryopreservation of embryos
tructural characterization of fresh and cryopreserved in vivo
heriogenology.2008.10.019

E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx2
+ Models
THE-10884; No of Pages 12
from many mammalian species have seen a widespread
use in reproductive research and in animal breeding in
recent years [2–4], but the technique has a much lower
utilization in sheep than, for example, in cattle [5]. This
fact may be related to the relatively high cost of the
technique in sheep when compared to the value of the
animals. Reducing costs at any stage of embryo
production and cryopreservation, including through
an increase in the efficiency of cryopreservation
methods, is likely to augment multiple ovulation and
embryo transfer techniques utilization in the ovine
species [1,4].
Controlled slow freezing and vitrification have been
the two major techniques used for embryo cryopre-
servation [4,6]. Controlled slow freezing was intro-
duced firstly and remains the mostly common technique
used, both with domestic animal and human embryos
[6]. However, slow-freezing requires expensive biolo-
gical freezers and is time consuming. The use of
ultrarapid techniques, such as vitrification, has reduced
part of the costs, since it does not require any special
equipment and is, therefore, well adapted to routine
field use [1]. The open pulled straw (OPS) method was
later developed [7] as a modification of embryo
vitrification, allowing increased cooling rates without
affecting embryo survival.
Controlled slow freezing and OPS have been used for
the cryopreservation of ovine morulae and blastocysts
produced in vivo [1,8–11] and in vitro [10,11]. The loss
of viability associated with freezing can be influenced
by the type and concentration of the cryoprotectant
used, the freezing protocol, the species and genotype of
the animal, the developmental stage of the embryos and
the type (in vivo vs in vitro) of embryo production
[2,3,6]. Comparisons between the different techniques
have relied on lambing rates after embryo transfer.
However, selection of embryos for transfer is based on
the stereomicroscopical evaluation of embryo morphol-
ogy after thawing [12] in accordance to the guidelines of
the International Embryo Transfer Society [13]. The
subjectivity inherent to this selection step has been
demonstrated by ultrastructural studies of vitrified in
vitro and in vivo produced bovine blastocysts [14], and
of controlled slow frozen in vivo produced ovine
morulae and blastocysts [15], which have shown that
certain abnormalities remain undetected by stereomi-
croscopy.
The ultrastructure of fresh and controlled slow frozen
in vivo produced ovine morulae and blastocysts has
been studied [15,16], but no reports have described the
ultrastructural features of OPS vitrified embryos nor
compared the two methods. The aims of the current
Please cite this article in press as: Bettencourt EMV, et al. Ultras
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.t
study were: (a) to investigate the ultrastructural
alterations induced by OPS vitrification of in vivo
produced ovine embryos at the morula and blastocyst
stages, (b) compare streomicroscopical embryo scoring
with light microscopy scoring of semithin sections, (c)
relate ultrastructural cellular damage with embryo
classification by stereomicroscopical scoring and light
microscopy semithin evaluation.
2. Materials and methods
2.1. Embryo production and embryo score
In vivo produced embryos (n = 51) were obtained
from Portuguese Black Merino ewes, superovulated
during spring. Donors were synchronized with
intravaginal sponges containing 40 mg flurogestone
acetate (Chronogest, Intervet Laboratories, Boxmeer,
Holland) for 12 days. Starting on day 9 of sponge
treatment and for 4 consecutive days, ewes were
treated (im) with 1.25 ml oFSH (Ovagen, Immuno-
logical Products Ltd, Aukland, New Zealand), twice a
day time (9:00 and 21:00). Sponges were removed at
the time of the 7th oFSH administration. Laparo-
scopic intrauterine insemination was performed with
fresh diluted semen in all ewes, 48 h after sponge
removal, as described [17]. A minimum of 50 � 106
motile spermatozoa was placed in each uterine horn.
Additionally, ewes were hand mated at 36 and 48 h
after sponge withdrawal and left with rams for 4–6 h
at a ewe:ram ratio of 3:1. Embryos were recovered by
abdominal laparotomy, under general anesthesia, on
day 6 or 7 after sponge removal. Each uterine horn
was flushed using a Foley catheter as described [18].
The collection medium (40 ml for each uterine horn)
was PBS containing 2% BSA.
Stereomicroscopical scoring of embryos was per-
formed based on morphological criteria and develop-
mental stage in accordance to International Embryo
Transfer Society guidelines [13]. Only fresh embryos of
grade 1 or grade 2 were selected for freezing. Fresh
embryos classified as grade 1 or 2 (n = 11) were directly
processed for transmission electron microscopy.
Collected grade 1 and 2 embryos at the morula and
blastocyst stages were immediately fixed or processed
for controlled slow freezing and OPS vitrification.
Frozen embryos were stereomicroscopical scored 1 h
after thawing and rescored at semithin sectioning. In the
smithin sections, embryos were scored as grade 1
(excellent or good) when presenting a symmetrical and
spherical cell mass with�85% of intact cells (<25% of
degenerated cells/cell fragments), with individual cells
tructural characterization of fresh and cryopreserved in vivo
heriogenology.2008.10.019
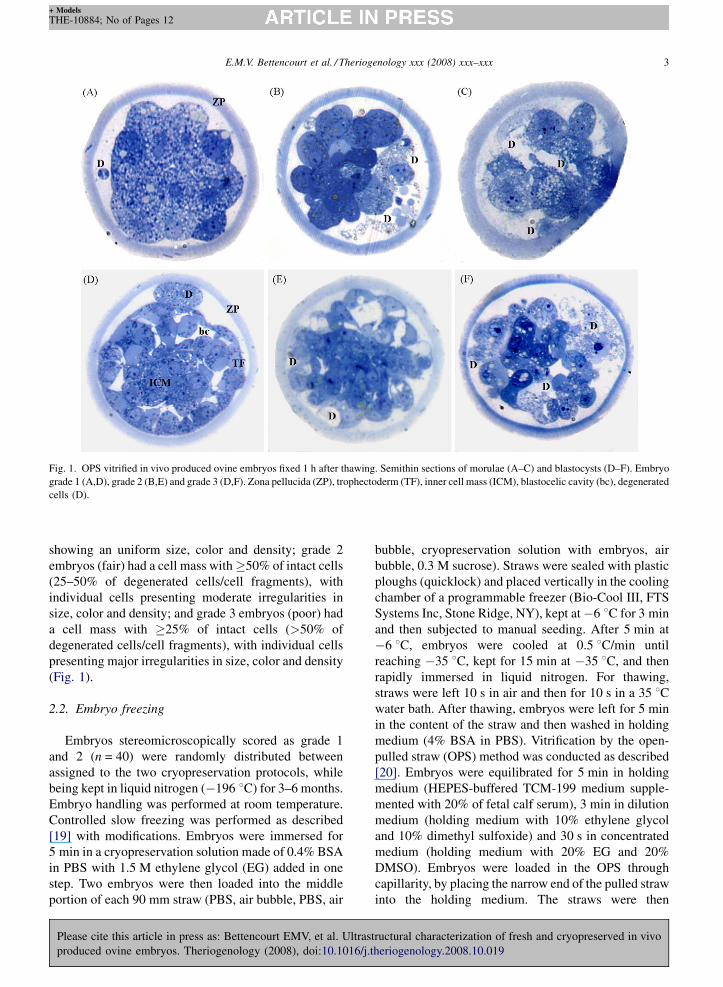
E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx 3
+ Models
THE-10884; No of Pages 12
Fig. 1. OPS vitrified in vivo produced ovine embryos fixed 1 h after thawing. Semithin sections of morulae (A–C) and blastocysts (D–F). Embryo
grade 1 (A,D), grade 2 (B,E) and grade 3 (D,F). Zona pellucida (ZP), trophectoderm (TF), inner cell mass (ICM), blastocelic cavity (bc), degenerated
cells (D).
showing an uniform size, color and density; grade 2
embryos (fair) had a cell mass with�50% of intact cells
(25–50% of degenerated cells/cell fragments), with
individual cells presenting moderate irregularities in
size, color and density; and grade 3 embryos (poor) had
a cell mass with �25% of intact cells (>50% of
degenerated cells/cell fragments), with individual cells
presenting major irregularities in size, color and density
(Fig. 1).
2.2. Embryo freezing
Embryos stereomicroscopically scored as grade 1
and 2 (n = 40) were randomly distributed between
assigned to the two cryopreservation protocols, while
being kept in liquid nitrogen (�196 8C) for 3–6 months.
Embryo handling was performed at room temperature.
Controlled slow freezing was performed as described
[19] with modifications. Embryos were immersed for
5 min in a cryopreservation solution made of 0.4% BSA
in PBS with 1.5 M ethylene glycol (EG) added in one
step. Two embryos were then loaded into the middle
portion of each 90 mm straw (PBS, air bubble, PBS, air
Please cite this article in press as: Bettencourt EMV, et al. Ultras
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.t
bubble, cryopreservation solution with embryos, air
bubble, 0.3 M sucrose). Straws were sealed with plastic
ploughs (quicklock) and placed vertically in the cooling
chamber of a programmable freezer (Bio-Cool III, FTS
Systems Inc, Stone Ridge, NY), kept at�6 8C for 3 min
and then subjected to manual seeding. After 5 min at
�6 8C, embryos were cooled at 0.5 8C/min until
reaching �35 8C, kept for 15 min at �35 8C, and then
rapidly immersed in liquid nitrogen. For thawing,
straws were left 10 s in air and then for 10 s in a 35 8Cwater bath. After thawing, embryos were left for 5 min
in the content of the straw and then washed in holding
medium (4% BSA in PBS). Vitrification by the open-
pulled straw (OPS) method was conducted as described
[20]. Embryos were equilibrated for 5 min in holding
medium (HEPES-buffered TCM-199 medium supple-
mented with 20% of fetal calf serum), 3 min in dilution
medium (holding medium with 10% ethylene glycol
and 10% dimethyl sulfoxide) and 30 s in concentrated
medium (holding medium with 20% EG and 20%
DMSO). Embryos were loaded in the OPS through
capillarity, by placing the narrow end of the pulled straw
into the holding medium. The straws were then
tructural characterization of fresh and cryopreserved in vivo
heriogenology.2008.10.019
E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx4
+ Models
THE-10884; No of Pages 12
immediately submerged into liquid nitrogen. For
thawing, the end of the straw was directly immersed
in holding medium containing 0.25 M sucrose.
Embryos were left for 1 min in this medium, then
5 min in holding medium containing 0.15 M sucrose
and washed twice with holding medium.
2.3. Electron microscopy
Fresh (n = 11) and thawed embryos after controlled
slow freezing (n = 20) and OPS vitrification (n = 20)
were scored and then fixed, within 1 h after collection or
thawing, in Karnowsky (2 h, 4 8C), washed with 0.15 M
sodium cacodylate buffer, pH 7.3 (overnight, 4 8C),
post-fixed (1 h, 4 8C) in 1% OsO4 in buffer containing
0.8% hexanocyanoferrate potassium [K3Fe3+(CN)6],
and washed for 15 min in buffer. They were then
dehydrated through a graded series of ethanol (50, 70,
90, 2 � 100%; 30 min each), followed by propylene
oxide (15 min), impregnated with propylene oxide:epon
(3:1, 1 h; 1:1, overnight, 4 8C; 1:3, 4 h) and embedded
in Epon (7 h at room temperature, 3 days at 60 8C).
Sections were cut in a Leica ultramicrotome with a
Diatome knife. Semithin sections were stained with
aqueous azur II:methylene blue (1:1). Ultrathin sections
were collected on 200 mesh copper grids (Taab) and
stained with 3% aqueous uranyl acetate (20 min) and
Reynolds lead citrate (10 min). Sections were observed
in a transmission electron microscope JEOL 100CXII
operated at 60 kV.
Except where otherwise indicated, all chemicals
were obtained from Sigma Chemicals Company
(Barcelone, Spain).
2.4. Experimental design
Embryo scores were compared with 2-sided Fisher‘s
exact test (SPSS 15 for Windows).
Please cite this article in press as: Bettencourt EMV, et al. Ultras
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.t
Table 1
Morphological classification of ovine embryos.
Cryopreservation method N Controlled Slow Freez
Embryo score (grade) 1 2
Thawing
Morulae 6 0 4
Blastocysts 14 8 2
Totals 20 8 6
Semithin sectioning
Morulae 6 0 6
Blastocysts 14 6 6
Totals 20 6 12
Numbers with lower case letters represent significant differences along lines,
3. Results
All fresh embryos were rescored at semithin
sectioning as grade 1 (2 embryos) or grade 2 (9
embryos). The majority of the thawed embryos were
classified either as grade 1 or grade 2 at both
evaluations, with the zona pellucida being intact in
all cases (Table 1). A significant higher number of grade
3 embryos was found at steromicroscopical scoring in
the controlled slow freezing method (P = 0.02), without
significant differences between morulae and blasto-
cysts. Although no significant differences were found in
the classification of the semithin sections (light
microscopy) between the two methods of cryopreserva-
tion, there was a significant higher number of grade 3
blastocysts in the OPS method (P = 0.037), as a higher
number of grade 3 embryos, especially of blastocysts
were found at the semithin sectioning classification than
at the stereomicroscopical classification, in the OPS
method (P = 0.003).
For the same grade at semithin scoring, no
differences (P > 0.05) were found regarding ultra-
structural characteristics between fresh, controlled slow
frozen and OPS vitrified embryos.
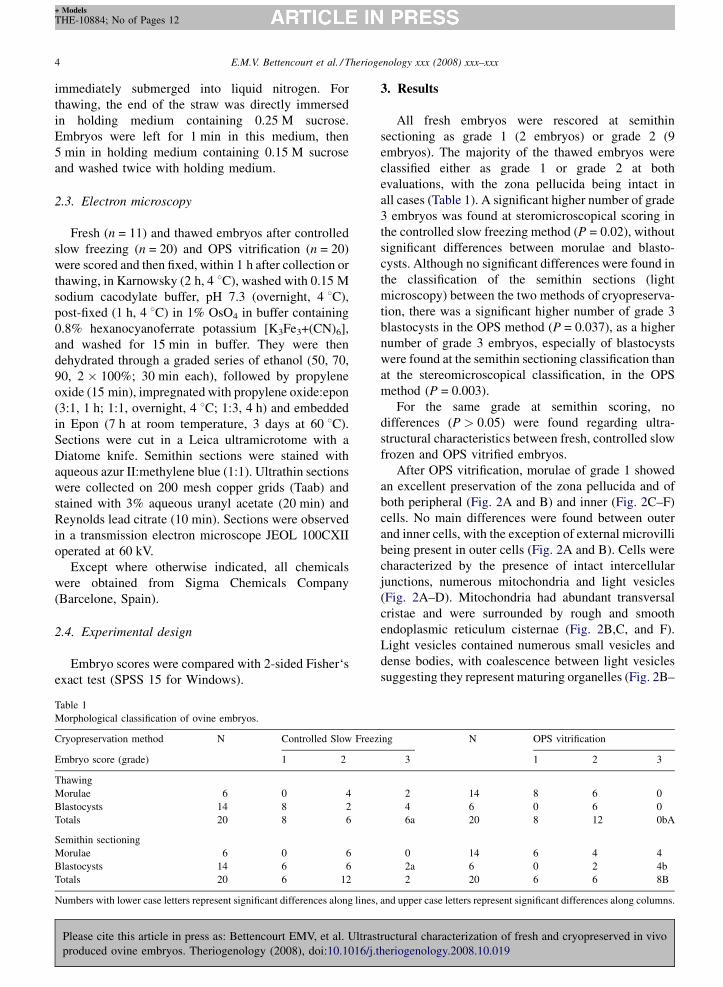
After OPS vitrification, morulae of grade 1 showed
an excellent preservation of the zona pellucida and of
both peripheral (Fig. 2A and B) and inner (Fig. 2C–F)
cells. No main differences were found between outer
and inner cells, with the exception of external microvilli
being present in outer cells (Fig. 2A and B). Cells were
characterized by the presence of intact intercellular
junctions, numerous mitochondria and light vesicles
(Fig. 2A–D). Mitochondria had abundant transversal
cristae and were surrounded by rough and smooth
endoplasmic reticulum cisternae (Fig. 2B,C, and F).
Light vesicles contained numerous small vesicles and
dense bodies, with coalescence between light vesicles
suggesting they represent maturing organelles (Fig. 2B–
tructural characterization of fresh and cryopreserved in vivo
heriogenology.2008.10.019
ing N OPS vitrification
3 1 2 3
2 14 8 6 0
4 6 0 6 0
6a 20 8 12 0bA
0 14 6 4 4
2a 6 0 2 4b
2 20 6 6 8B
and upper case letters represent significant differences along columns.
E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx 5
+ Models
THE-10884; No of Pages 12
Please cite this article in press as: Bettencourt EMV, et al. Ultrastructural characterization of fresh and cryopreserved in vivo
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.theriogenology.2008.10.019
Fig. 2. Ultrastructure of OPS vitrified in vivo produced ovine morulae fixed 1 h after thawing. Morulae were classified as grade 1 in semithin
sections. Outer (A,B) and inner (C–F) cells. Zona pellucida (ZP), microvilli (Mv), intercellular junctions (black arrows), mitochondria (m), light
vesicles (v), secondary lysosomes (LY), lipid droplets (L), Golgi complexes (G), smooth endoplasmic reticulum (white arrowheads), rough
endoplasmic reticulum (black arrowheads), nucleus (N), nuclear pore complexes (white open arrows), nucleolus (Nc), nucleolar fibrillar centers
(FC), nucleolar dense fibrillar component (DFC), nucleolar granular component (GC), polyribossomes (pr).
E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx6
+ Models
THE-10884; No of Pages 12
Please cite this article in press as: Bettencourt EMV, et al. Ultrastructural characterization of fresh and cryopreserved in vivo
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.theriogenology.2008.10.019
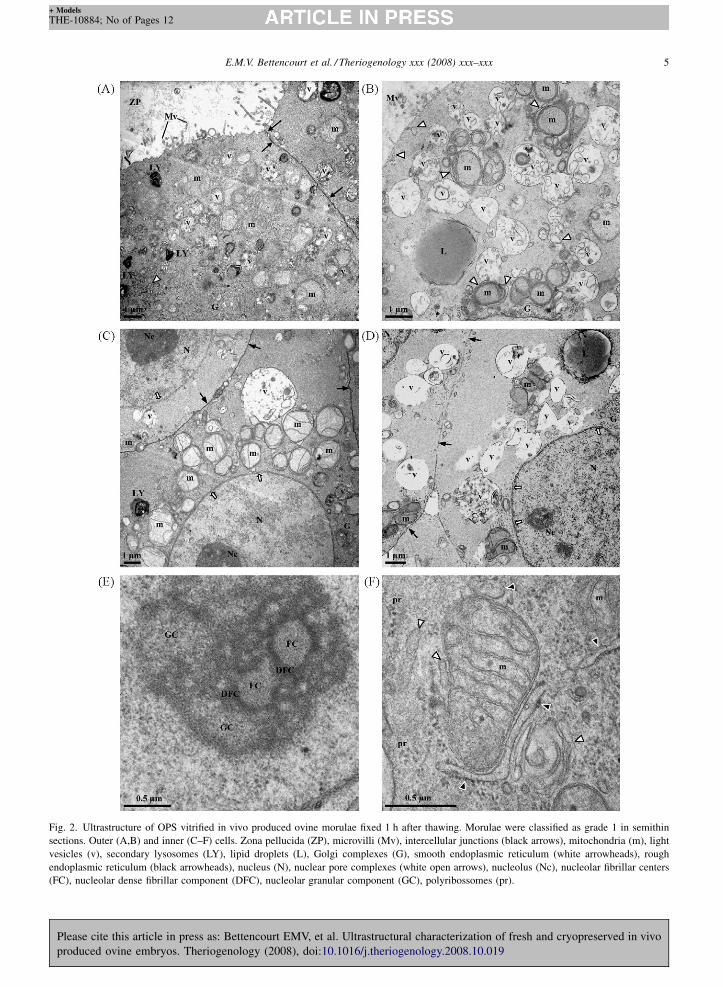
Fig. 3. Ultrastructure of OPS vitrified in vivo produced ovine morulae fixed 1 h after thawing. Morulae were classified as grade 2 (A,B) and grade 3
(C–F) in semithin sections. Outer (A,C) and inner (B,D–F) cells. (A,B) Presence of extracellular cell debris (F), loss of microvilli and intercellular
junctions (black arrows), moderate swelling of the mitochondrial matrix (*), endoplasmic reticulum (white arrowheads) and light vesicles (v), and
occasional degenerated mitochondria (d). Lipid droplets (L). (C) Discontinuities of the cytoplasmic membrane (black open arrows), loss of
microvilli, disruption of intercellular junctions (black arrows), severe swelling of the mitochondrial matrix (**), mitochondria degeneration (d),
swelling of cytosolic areas (c) and swelling of the endoplasmic reticulum (white arrowheads). Zona pellucida (ZP), Golgi complex (G). (D) Large
aggregate of light vesicles (v), swollen endoplasmic reticulum (white arrowheads), and degenerated mitochondria (d). (E) Severe swelling of
mitochondria, with organelle rupture (**) and envelope degeneration (black open arrow). (F) Loss of intercellular junctions (black arrows), swelling
E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx 7
+ Models
THE-10884; No of Pages 12
Please cite this article in press as: Bettencourt EMV, et al. Ultrastructural characterization of fresh and cryopreserved in vivo
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.theriogenology.2008.10.019
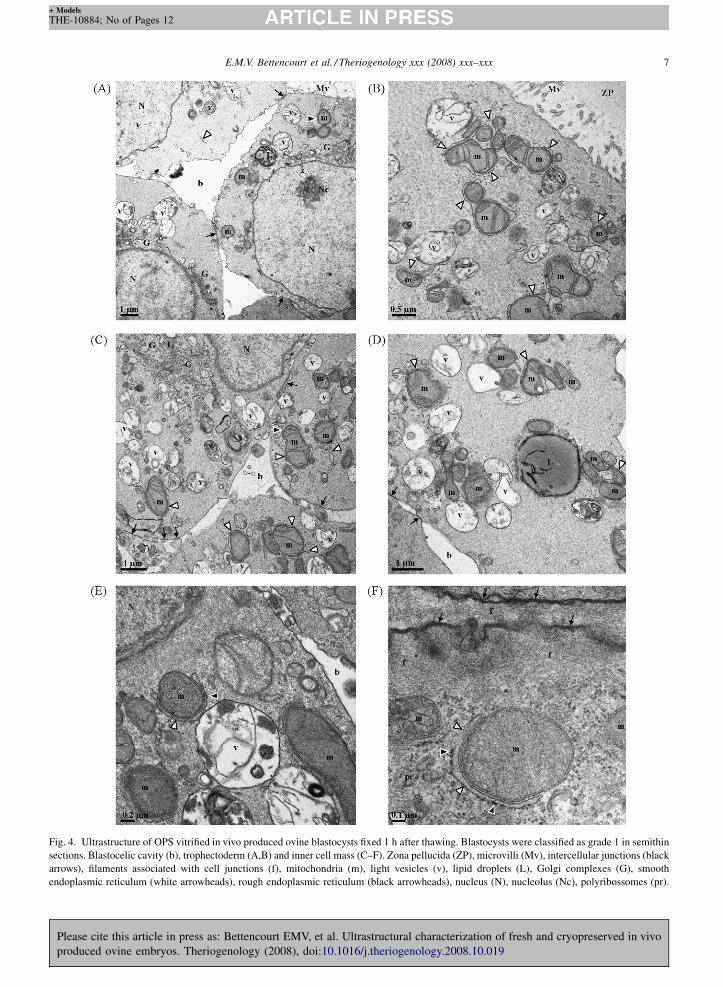
Fig. 4. Ultrastructure of OPS vitrified in vivo produced ovine blastocysts fixed 1 h after thawing. Blastocysts were classified as grade 1 in semithin
sections. Blastocelic cavity (b), trophectoderm (A,B) and inner cell mass (C–F). Zona pellucida (ZP), microvilli (Mv), intercellular junctions (black
arrows), filaments associated with cell junctions (f), mitochondria (m), light vesicles (v), lipid droplets (L), Golgi complexes (G), smooth
endoplasmic reticulum (white arrowheads), rough endoplasmic reticulum (black arrowheads), nucleus (N), nucleolus (Nc), polyribossomes (pr).

E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx8
+ Models
THE-10884; No of Pages 12
Please cite this article in press as: Bettencourt EMV, et al. Ultrastructural characterization of fresh and cryopreserved in vivo
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.theriogenology.2008.10.019
Fig. 5. Ultrastructure of OPS vitrified in vivo produced ovine blastocysts fixed 1 h after thawing. Blastocysts were classified as grade 2 (A,B) and
grade 3 (C–F) in semithin sections. Blastocelic cavity (b), trofectoderm (A,C,D) and inner cell mass (B,E,F). (A) Presence of cell debris (F),
cytoplasmic membrane discontinuities (black open arrow), decreased number and length of microvilli (Mv), moderate swelling of the endoplasmic

E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx 9
+ Models
THE-10884; No of Pages 12
D). In the remainder of the cytoplasm there were a few
Golgi complexes, secondary lysosomes, lipid droplets
and polyribosomes (Fig. 2A–D,F). The nucleus was
well defined and organized, with an intact nuclear
envelope, rich in nuclear pore complexes, and one or
more reticular nucleoli with evident fibrillar centers
surrounded by the dense fibrillar and granular compo-
nents (Fig. 2C–E).
At semithin scoring OPS vitrified morulae scored as
grade 2 also showed intact cellular structures in most of
the blastomeres, as described for grade 1 embryos.
However, there were several signs of cellular injury in
totally or partially detached cells. These ranged from
presence of cell debris, resulting from total cell lysis
(Fig. 3A), to decreased number and length of microvilli,
loss of intercellular junctions, and moderate swelling of
the mitochondrial matrix, endoplasmic reticulum and
light vesicles (Fig. 3A and B).
At semithin scoring, morulae vitrified by OPS and
classified as grade 3 presented additional degenerated
cells in both peripheral and internal cells, with badly
defined cellular limits, poor intracellular preservation,
disruption of the plasma membrane and presence of cell
debris, due to complete cell lysis. Less attained outer
cells showed discontinuities of the cytoplasmic mem-
brane, loss of microvilli, disruption of intercellular
junctions, severe swelling of the mitochondrial matrix,
degenerated mitochondria, focal swelling of cytosolic
regions and swelling of the endoplasmic reticulum
(Fig. 3C). Less attained inner cells showed vacuolar
degeneration, characterized by the presence of large
aggregates of light vesicles, swollen endoplasmic
reticulum and degenerated mitochondria (Fig. 3D),
while other cells presented severe swelling of mito-
chondria, with organelle rupture and envelope degen-
eration (Fig. 3E), loss of intercellular junctions,
swelling of the nucleoplasm and cytosol, disorganiza-
tion of the nucleolar structure, and cytoplasmic
membrane and nuclear envelope discontinuities
(Fig. 3F).
At semithin scoring, after OPS vitrification, blas-
tocysts of grade 1, showed an excellent preservation of
the zona pellucida, trophectoderm (Fig. 4A and B) and
Please cite this article in press as: Bettencourt EMV, et al. Ultras
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.t
reticulum (white arrowheads) and light vesicles (v). Zona pellucida (ZP), mito
loss of cristae (*), swelling of the endoplasmic reticulum (white arrowhead
extracellular cell debris (F). Nucleus (N), light vesicles (v), lipid droplets (L)
discontinuities (black open arrows), disruption of intercellular junctions (bla
light vesicles (v). Smooth endoplasmic reticulum (white arrowheads), rough
cytosol (c) and nucleoplasm (n), with disruption of intercellular junctions (bl
Degenerative cells, with loss of membrane limits (black arrows) and organel
open arrows), cytosol (c), degenerated mitochondria (d), light vesicles (v).
inner cell mass (Fig. 4C–F). No main differences were
found between cells of the trophectoderm and inner cell
mass, with the exception of the presence of external
microvilli in trophectoderm cells (Fig. 4A and B). Intact
intercellular junctions, consisting of tight junctions,
desmosomes and gap junctions, were observed in all
cells, including between the trophectoderm and inner
cell mass (Fig. 4A,C,D, and F). The cytoplasm
contained the same set of organelles previously found
in morulae, mitochondria with numerous transversal
cristae surrounded by cisternae of the smooth and rough
endoplasmic reticulum (Fig. 4B,C–F), more abundant
and well developed Golgi complexes (Fig. 4A and C),
light vesicles, lipid droplets, secondary lysosomes, and
polyribosomes (Fig. 4F). Cells displayed a large
euchromatic nucleus, with an intact nuclear envelope
rich in nuclear pore complexes and one or more reticular
nucleoli, with evident fibrillar centers surrounded by the
dense fibrillar and granular components (Fig. 4A,C, and
E).
At semithin scoring, OPS vitrified blastocysts scored
as grade 2 showed well preserved cellular structures but,
as described for grade 2 morulae, there were some signs
of cellular degeneration, such as extracellular cell
debris, decreased number of trophectoderm outer
microvilli, cytoplasmic membrane discontinuities,
swelling of light vesicles (Fig. 5A), mitochondria and
endoplasmic reticulum, and increased areas of the
cytoplasm without organelles (Fig. 5B).
At semithin scoring, blastocysts vitrified by OPS and
classified as grade 3 presented additional degenerated
cells in both trophectoderm and inner cell mass, with
badly defined cellular limits, poor intracellular preserva-
tion, disruption of the plasma membrane, nuclear
envelope and other organelle membranes, and presence
of cell debris due to complete cell lysis (Fig. 5F). Less
attained cells showed loss of microvilli (Fig. 5C),
disruption of intercellular junctions (Fig. 5D and E),
severe degenerative swelling of mitochondria (Fig. 5C
and D), discontinuities in the cytoplasmic membrane
(Fig. 5C and D) and nuclear envelope (Fig. 5E), as well as
swelling of the cytosol and nucleoplasm, with disorga-
nization of the nucleolar structure (Fig. 5E and F).
tructural characterization of fresh and cryopreserved in vivo
heriogenology.2008.10.019
chondria (m). (B) Moderate swelling of the mitochondrial matrix with
s), presence of large cytosolic areas devoid of organelles (c) and of
, intercellular junctions (black arrows). (C,D) Cytoplasmic membrane
ck arrows), and severe degenerative swelling of mitochondria (**) and
endoplasmic reticulum (black arrowheads). (E) Severe swelling of the
ack arrow) and rupture of the nuclear envelope (black open arrow). (F)
le structure. Nucleoplasm (n), nucleolus (Nc), nuclear envelope (white

E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx10
+ Models
THE-10884; No of Pages 12
4. Discussion
Ovine embryos have been subjected to cryopre-
servation using both controlled slow freezing and OPS
vitrification [1,9,10,11]. With the present native sheep
breed, the Portuguese Black Merino, we have found no
differences in lambing (55–68%) and embryo survival
(36–45%) rates after the transfer of embryos cryopre-
served by both methods (data not shown), which is in
agreement with previous studies in other sheep breeds
[1,9,11]. The relatively high pregnancy rates obtained
after transfer of cryopreserved embryos using both
techniques, suggests that cryopreservation and warm-
ing do not seriously affect embryo viability [1].
However, in most of the studies that compared
different cryopreservation methods, embryo reselec-
tion after thawing has been performed [9,11], and in
some cases only blastocysts that had reexpanded were
transferred [10], which may result in some embryos
being wrongly discarded based on post-thawing
morphological scorings [1]. On the other hand, a
few embryos appearing as morphologically good after
thawing fail to produce pregnancies [8], which also
might be due to the transfer of embryos with serious
ultrastructural lesions not detected at the lower
resolution given by stereomicroscopy.
Although evaluation of embryo morphology in
semithin sections is not a viable method to select
embryos for transfer, it allows the identification of cell
and subcellular injuries that might not be detected by
stereomicroscopy of living embryos at thawing and so,
it can be a useful tool to improve our knowledge of the
lesions resulting from different cryopreservation
methods [8,21]. In the present study we found
dissimilar embryo grading classifications after semi-
thin evaluation of embryos previously graded by
stereomicroscopical scoring, which confirms the
usefulness of the semithin evaluations. These differ-
ences support previous reports [1,8] wherein some
embryos with a poor/fair grade at stereomicroscopical
scoring are still viable while others with a good score
are developmentally compromised [14,15]. This may
be due, at least in part, to misclassification after
stereomicroscopical scoring.
The ultrastructural analysis of cryopreserved–
thawed embryos is limited to ovine and bovine embryos
cryopreserved by controlled slow freezing [15,22],
and vitrified bovine and swine embryos [14,21,23]. In
the present study, we have described the ultrastructural
characteristics and the cryoinjuries observed in
grades 1, 2 and 3 in vivo produced ovine morulae
and blastocysts, after OPS vitrification and thawing.
Please cite this article in press as: Bettencourt EMV, et al. Ultras
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.t
Independently of the stage of development considered
and cryopreservation method used, embryos scored as
grade 1 at semithin evaluation showed well preserved
blastomeres with an intact structure, while embryos
scored as grade 3 presented degenerated cells, disrupted
cell membranes, severe mitochondrial injury and
cytoskeleton disorganization, which confirm results
from previous reports using controlled slow frozen
ovine and bovine embryos [15,24], vitrified bovine
embryos [14], and swine embryos vitrified by OPS
[21,25]. At the ultrastructural level, there were no
differences between both cryopreservation methods for
embryos with the same grade after semithin scoring.
Despite the absence of studies specifically comparing
the ultrastructure of embryos cryopreserved by con-
trolled slow freezing or vitrification, similar cellular
lesions were described for bovine, ovine and porcine
embryos cryopreserved by both methods
[14,15,21,22,23]. In addition, OPS vitrified swine
embryos classified as degenerated (grade 3) following
thawing presented similar cellular lesions, namely
badly defined cellular limits, poor intracellular pre-
servation and presence of cell debris [21].
Although cryoprotectants are known to affect
cytoskeleton dynamics, the interaction between cytos-
keleton dynamics and the cellular organelles of
embryos are not consistent between species and stages
of embryo development [3,24]. Nevertheless, the early
injuries here described, such as loss of microvilli,
intercellular junctions and cytoplasmic membrane
stability, could have a disruptive impact over the
cytoskeleton, which would then lead to a severely
disturbed cell organization such as that found in
degenerated blastomeres [25]. The present data also
suggests an early loss of the mitochondrial structure,
which leads to disruption of energy supply to cell
boundaries. Damaged mitochondria after cryopreserva-
tion had been reported in previous studies [14,15,21],
suggesting that the preservation level of the mitochon-
drial matrix and envelope can be used as a reliable sign
of cellular damage. Accordingly, stages showing higher
degrees of cell damage were here associated with more
severe mitochondrial injuries that ranged from severe
matrix swelling, with loss of cristae, to membrane
degeneration and rupture. This was accompanied by a
progressive degenerative process that involved swelling
of light vesicles and endoplasmic reticulum cisternae,
followed by swelling of the nucleoplasm, loss of nuclear
pore complexes and of the nucleolar structure, and
nuclear envelope disruption.
Semithin and transmission electron microscopy
evaluation of OPS vitrified bovine and swine blastocysts
tructural characterization of fresh and cryopreserved in vivo
heriogenology.2008.10.019

E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx 11
+ Models
THE-10884; No of Pages 12
were performed with embryos that were cultured in
vitro for 4 and 24 h after thawing [14,21,22]. With these
in vitro culture periods, clear signs of regeneration were
observed at the ultrastructural level, even in embryos
whose stereomicroscopy evaluation at thawing indi-
cated signs of severe damage. These signs of
regeneration included the reestablishment of junctional
contacts between trophectoderm cells and normal-
ization of the mitochondrial morphology [14,25]. The
present study was performed only after 1 h of culture
after thawing, to better simulate the clinical field
situation. It is possible that long term embryo recovery
then occurred in the uterine cavity, as it may happen in
vitro [14,25], which would help to explain the similar
rates of lambing and embryo survival after transfer of
embryos cryopreserved by both methods [1,9,11].
However, this is not necessarily always the case, as
some of the lesions found in this study, in particular for
embryos scored as grade 3, were hardly susceptible of
repair.
In conclusion, this study describes for the first time
the semithin and ultrastructural evaluation of OPS
vitrified ovine embryos, classified with different scores
at thawing (from grades 1 to 3). The utrastructural
lesions found for OPS-vitrified and controlled slow
frozen embryos were similar, independently of embryo
stage, and affected determinant cell components, such
as mitochondria and the cytoskeleton, which can
influence vital cell functioning and thus compromised
embryo viability after cryopreservation. The extension
and gravity of the cellular damage observed at the
ultrastructural level were proportional to those observed
at semithin sections performed on cryopreserved
embryos fixed after thawing. Furthermore, embryo
grading was dissimilar between stereomicroscopical
and smithin embryo scoring. Therefore, the semithin
scoring can be a useful tool to better evaluate
ultrastructural lesions in cryopreserved embryos. Most
of the cryopreserved embryos presented minor cellular
lesions. However, others displayed severe and extensive
injuries (grade 3 embryos), especially blastocysts
submitted to OPS vitrification. Therefore, cryopreser-
vation methods must be chosen according to the species
and embryonic stage and improvements in cryopreser-
vation and thawing methods to decrease cellular
damage of ovine embryos are still warranted.
Acknowledgements
This work was partially supported by funds from the
Portuguese AGRO projects (AGRO 438 Portuguese
Animal Germplasm Bank), FEDER and FCT (SFRH/
Please cite this article in press as: Bettencourt EMV, et al. Ultras
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.t
BD/30575/2006; POCI/SAU-MMO/60709,/60555 and/
59997/04; UMIB).
References
[1] Baril G, Traldi AL, Cognie Y, Leboeuf B, Beckers JF, Mermillod
P. Successful direct transfer of vitrified sheep embryos. Ther-
iogenology 2001;56:299–305.
[2] Guinot F. Cryoconservation des embryons des especes domes-
tiques. INRA Prod Anim 2005;18:27–35.
[3] Dobrinsky JR. Advancements in cryopreservation of domestic
embryos. Theriogenology 2002;57:285–302.
[4] Vajta G, Nagy ZP. Are programmable freezers still needed in the
embryo laboratory? Review on vitrification. Reproductive Bio-
Medicine Online 2006;12:779–96.
[5] Thibier M. Data Retrieval Committee annual report. IETS
Newsletter 2006;24:12–18.
[6] Vajta G, Kuwayama M. Improving cyopreservation systems.
Theriogenology 2006;65:236–44.
[7] Vajta G, Booth PJ, Holm P, Greve T, Callesen H. Successful
vitrification of early stage bovine in vitro produced embryos with
the Open Pulled Straw (OPS) method. Cryo-Lett 1997;18:191–5.
[8] Cocero MJ, Lopez Sebastian A, Barragan ML, Picazo RA.
Differences on post-thawing survival between ovine morulae
and blastocysts cryopreserved with ethylene glycol or glycerol.
Criobiology 1996;33:502–7.
[9] Martinez AG, Matkovic M. Cryopreservation of ovine embryos:
slow freezing and vitrification. Theriogenology 1998;49:
1039–49.
[10] Dattena M, Accardo C, Pilchi S, Isachenko V, Mara L, Chessa B,
et al. Comparison of different vitrification protocols on viability
after transfer of ovine blastocysts in vitro produced and in vivo
derived. Theriogenology 2004;62:481–93.
[11] Martinez AG, Valcarcel A, Furnus CC, de Matos DG, Iorio G, de
las Heras MA. Cryopreservation of in vitro-produced ovine
embryos. Small Rumin Res 2006;63:288–96.
[12] Abe H, Matsuzaki S, Hoshi H. Ultrastructural differences in
bovine morulae classified as high and low qualities by morpho-
logical evaluation. Theriogenology 2002;57:1273–83.
[13] Stringfellow DA, Seidel SM, editors. Manual of the International
Embryo Transfer Society Savoy II. USA: International Embryo
Transfer Society, Inc.; 1998.
[14] Vajta G, Hyttel P, Callensen H. Morphological changes of in
vitro produced bovine blastocysts after vitrification, in-straw
direct rehydratation, and culture. Mol Reprod Dev 1997;48:
9–17.
[15] Cocero MJ, Dıaz de la Espina SM, Aguilar B. Ultrastructural
characteristics of fresh and frozen-thawed ovine embryos using
two cryoprotectants. Biol Reprod 2002;66:1244–58.
[16] Ferrer F, Garcıa C, Villar J, Arias M. Ultrastructural study of the
early development of the sheep embryo. Anat Histol Embryol
1995;24:191–6.
[17] McKelvey AC, Robinson JJ, Aitken RP, Henderson G. The
evaluation of a laparoscopic insemination technique in ewes.
Theriogenology 1985;24:519–35.
[18] Smith CL, Murphy CA. An antegrade surgical uterine flush
technique for ova collection in the ewe. Am J Vet Res
1987;48:1129–31.
[19] Voelkel SA, Hu YX. Use of ethylene glycol as a cryoprotectant
for bovine embryos allowing direct transfer of frozen-thawed
embryos to recipient females. Theriogenology 1992;37:687–97.
tructural characterization of fresh and cryopreserved in vivo
heriogenology.2008.10.019

E.M.V. Bettencourt et al. / Theriogenology xxx (2008) xxx–xxx12
+ Models
THE-10884; No of Pages 12
[20] Vajta G, Holm P, Kuwayama M, Booth PJ, Jacobsen H, Greve T,
et al. Open pulled straw (OPS) vitrification: a new way to reduce
cryoinjuries of bovine ova and embryos. Mol Reprod Dev
1998;51:53–8.
[21] Cuello C, Berthelot F, Delaleu B, Venturi E, Pastor LM, Vazquez
JM, et al. The effectiveness of the stereomicroscopic evaluation of
embryo quality in vitrified-warmed porcine blastocysts: an ultra-
structural and cell death study. Theriogenoloy 2007;67:970–82.
[22] Fair T, Lonergan P, Dinnyes A, Cottell DC, Hyttel P, Ward FA,
et al. Ultrastructure of bovine blastocysts following cryopreser-
Please cite this article in press as: Bettencourt EMV, et al. Ultras
produced ovine embryos. Theriogenology (2008), doi:10.1016/j.t
vation: effect of method of blastocyst production. Mol Reprod
Dev 2001;58:186–95.
[23] Fabian D, Gjorret JO, Berthelot F, Martinat-Botte F, Maddox-
Hyttel P. Ultrastructure and cell death of in vivo derived and
vitrified porcine blastocysts. Mol Reprod Dev 2005;70:155–65.
[24] Dobrinsky JR. Cellular approach to cryopreservation of
embryos. Theriogenology 1996;45:17–26.
[25] Overstrom EW, Duby RT, Dobrinsky JR, Robl JM, Baguisi A,
Lonergan P, et al. Cytoskeletal damage in vitrified or frozen
bovine embryos. Theriogenology 1993;39:276.
tructural characterization of fresh and cryopreserved in vivo
heriogenology.2008.10.019
Copyright © 2022 FDOKUMEN




















