
Evaluation of epirubicin-induced acute oxidative stress toxicity in rat liver cells and...
7
Experimental and Toxicologic Pathology 61 (2009) 161–167 Evaluation of epirubicin-induced acute oxidative stress toxicity in rat liver cells and mitochondria, and the prevention of toxicity through quercetin administration M. Kebieche a, , Z. Lakroun a , M. Lahouel a , J. Bouayed b , Z. Meraihi c , R. Soulimani b, a Phytochemistry and Pharmacology Laboratory, Faculty of Science, University of Jijel, Jijel 18000, Algeria b Ethnobotanique et pharmacologie/Anxie´te´, stress Oxydant et bioactivite´, Universite´P. Verlaine-Metz, Bridoux, Rue du ge´ne´ral Delestraint, 57070 Metz, France c Department of Natural Science, Faculty of Science, University of Constantine,Constantine 25000, Algeria Received 15 November 2007; accepted 18 June 2008 Abstract Anticancer therapy with epirubicin (EPI) results in acute hepatotoxicity, likely due to the generation of free radicals. However, the oxidative status of rat liver cells and mitochondria after EPI toxicity has not been investigated. In the present study, we first investigated the pro-oxidant effect of EPI on both hepatic cells and mitochondrial function. Injection of EPI into rats at a dose of 9 mg/kg (cumulative dose in human chemotherapy), induced hepatic dysfunction, as revealed by a significant increase in serum glutamate oxaloacetate transaminases (SGOT) and glutamate pyruvate transaminases (SGPT). Oxidative stress in liver cells and mitochondria was provoked by EPI because a statistically significant reduction of catalase (CAT), superoxide dismutase (SOD) and cytosolic glutathione (GSH) levels, and a significant increase in malonedialdehyde (MDA) levels – an indicator of lipid peroxidation that can perforate biological membranes – were observed. Second, the protective effect of quercetin (QE) (0.33 mg/kg) against EPI-induced oxidative stress was also investigated. Indeed, the pretreatment of rats with QE protected liver cells and mitochondria from oxidative stress. This treatment prevented hepatic dysfunction by maintaining normal levels of serum transaminases following the inhibition of their hepatic leakage by preventing lipid peroxidation. Thus, QE works through the prevention of cellular membrane perforation and the antioxidant defense system of mitochondria from liver cells, which represent compartments for the permanent production of reactive oxygen species (ROS) through the respiratory chain. r 2008 Elsevier GmbH. All rights reserved. Keywords: Epirubicin; Hepatotoxicity; Oxidative stress; Quercetin; Hepatoprotective effect Introduction Redox homeostasis is a delicate equilibrium between antioxidants and pro-oxidants in a living system. In the human body, the principal reactive oxygen species (ROS) like the superoxide anion (O 2 d ), hydrogen peroxide (H 2 O 2 ) and the hydroxyl radical ( d OH) are ARTICLE IN PRESS www.elsevier.de/etp 0940-2993/$ - see front matter r 2008 Elsevier GmbH. All rights reserved. doi:10.1016/j.etp.2008.06.002 Corresponding author. Tel./fax: +21334474896. Also corresponding author. E-mail addresses: [email protected] (M. Kebieche), [email protected] (R. Soulimani).
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Evaluation of epirubicin-induced acute oxidative stress toxicity in rat liver cells and...

ARTICLE IN PRESS
0940-2993/$ - se
doi:10.1016/j.et
�Correspond��Also corre
E-mail addr
soulimani@uni
Experimental and Toxicologic Pathology 61 (2009) 161–167
www.elsevier.de/etp
Evaluation of epirubicin-induced acute oxidative stress toxicity in rat
liver cells and mitochondria, and the prevention of toxicity through
quercetin administration
M. Kebiechea,�, Z. Lakrouna, M. Lahouela, J. Bouayedb, Z. Meraihic, R. Soulimanib,��
aPhytochemistry and Pharmacology Laboratory, Faculty of Science, University of Jijel, Jijel 18000, AlgeriabEthnobotanique et pharmacologie/Anxiete, stress Oxydant et bioactivite, Universite P. Verlaine-Metz, Bridoux,
Rue du general Delestraint, 57070 Metz, FrancecDepartment of Natural Science, Faculty of Science, University of Constantine,Constantine 25000, Algeria
Received 15 November 2007; accepted 18 June 2008
Abstract
Anticancer therapy with epirubicin (EPI) results in acute hepatotoxicity, likely due to the generation of free radicals.However, the oxidative status of rat liver cells and mitochondria after EPI toxicity has not been investigated. In thepresent study, we first investigated the pro-oxidant effect of EPI on both hepatic cells and mitochondrial function.Injection of EPI into rats at a dose of 9mg/kg (cumulative dose in human chemotherapy), induced hepatic dysfunction,as revealed by a significant increase in serum glutamate oxaloacetate transaminases (SGOT) and glutamate pyruvatetransaminases (SGPT). Oxidative stress in liver cells and mitochondria was provoked by EPI because a statisticallysignificant reduction of catalase (CAT), superoxide dismutase (SOD) and cytosolic glutathione (GSH) levels, and asignificant increase in malonedialdehyde (MDA) levels – an indicator of lipid peroxidation that can perforatebiological membranes – were observed. Second, the protective effect of quercetin (QE) (0.33mg/kg) againstEPI-induced oxidative stress was also investigated. Indeed, the pretreatment of rats with QE protected liver cells andmitochondria from oxidative stress. This treatment prevented hepatic dysfunction by maintaining normal levels ofserum transaminases following the inhibition of their hepatic leakage by preventing lipid peroxidation. Thus, QEworks through the prevention of cellular membrane perforation and the antioxidant defense system of mitochondriafrom liver cells, which represent compartments for the permanent production of reactive oxygen species (ROS)through the respiratory chain.r 2008 Elsevier GmbH. All rights reserved.
Keywords: Epirubicin; Hepatotoxicity; Oxidative stress; Quercetin; Hepatoprotective effect
e front matter r 2008 Elsevier GmbH. All rights reserved.
p.2008.06.002
ing author. Tel./fax: +21334474896.
sponding author.
esses: [email protected] (M. Kebieche),
v-metz.fr (R. Soulimani).
Introduction
Redox homeostasis is a delicate equilibrium betweenantioxidants and pro-oxidants in a living system. In thehuman body, the principal reactive oxygen species(ROS) like the superoxide anion (O2
d), hydrogenperoxide (H2O2) and the hydroxyl radical (dOH) are

ARTICLE IN PRESSM. Kebieche et al. / Experimental and Toxicologic Pathology 61 (2009) 161–167162
continuously generated inside cells. Their generationresults from a number of endogenous metabolic path-ways as well as from exposure to xenobiotics in theambient environment (Chen et al., 2007; Kaur et al.,2006). Exposure of organisms to exogenous andendogenous factors induces the overproduction of awide range of ROS, resulting in the loss of equili-brium between pro-oxidants and antioxidant systemswithin the cells (Halliwell and Gutteridge, 1999;Kaur et al., 2006). The ROS may also oxidize thesurrounding molecules in the biological medium(nucleic acids, lipids, glucids and proteins) (Farber,1994), and induce cellular lesions, potentially leading tocell death by necrosis or apoptosis (Halliwell andGutteridge, 1999). The oxidative damage of variousbiomolecules is considered as an essential etiologicfactor that has been implicated in several diseases,including the chronic diseases of cancer, diabetes,atherosclerosis, cirrhosis and Alzheimer’s (Pong, 2003;Chen et al., 2007).
Several studies have reported that drug and toxicsubstance-induced hepatopathy has increased in thepast years, especially in developing countries (Brind,2007; Watkins and Seef, 2006). In this context, the toxiceffects of the derivatives of anthracyclines such asepirubicin (EPI), daunorubicin and doxorubicin on theheart and the liver have been widely recognized (Le Botet al., 1988; Nielsen et al., 1990; Goebel, 1993; Mazueet al., 1995). In fact, toxicological studies in humanshave demonstrated that EPI induces severe cardiotoxi-city. Nevertheless, EPI was considered less cardiotoxicthan adriamycin and has therefore been used inEurope in anticancer treatments (Lanza et al., 1989;Yeung et al., 1989; Germain et al., 2003). Currently, theavailable information suggests only that EPI-inducedtoxicity in the liver and heart can be mediated throughoxygen-free radicals produced during EPI metabolism(Le Bot et al., 1988; Germain et al., 2003). However,the biochemical mechanisms involved in the hepatotoxi-city and cardiotoxicity of EPI have not been clearlyidentified and must be established. Furthermore, theprotective effect of flavonoids on EPI-induced hepato-toxicity in vivo has not been investigated. It has beenreported that flavonoids offer protection from severaldiseases due to their potential antioxidant activity(Bors et al., 1994; Pannala et al., 1997; Lahouel et al.,2004; Raja et al., 2007). In the current study, ourobjectives were to first investigate the implicationsof oxidative stress in EPI-induced hepatotoxicity byassessing the redox status in liver cells in rats treatedwith EPI. Moreover, this study was extended tomitochondria by monitoring the subcellular range ofthis anticancer toxicity. Finally, the hepatoprotectiveeffect of quercetin (QE) – a nutraceutical flavonoid – onEPI-induced hepatotoxicity was also studied in a ratmodel.
Materials and methods
Chemicals
The majority of chemicals were procured from SigmaAldrich, Germany. Assay Kits for serum enzymes werepurchased from Biomerieux and EPI was purchasedfrom Pharmacia, St. Quentin in Yvelines, France.
Animal maintenance
Male albino Wistar rats (body weight 220–280 g) thatwere originally from the Pasteur institute in Algiers,Algeria, were used in these experiments. Rats were bredin our animal facility in stainless metallic cages. Theroom housing the rats was temperature controlled(average of 22 1C, 50–60% relative humidity) and keptunder a daily 12 h light/dark cycle. Rats were fed foodand water ad libitum. Fasted rats were deprived of foodfor at least 16 h, but were allowed free access to water.Rats were adapted for 1 week before the indicatedtreatments. All experimental assays were carried out inconformity with international guidelines for the care anduse of laboratory animals.
Animal treatment protocol
The animals were grouped as follows:
�
Group 1, control rats: rats were administered 1ml ofphysiological saline 24 h prior to and after per os(p.o.) and by i.v. through the caudal vein. � Group 2, EPI-treated: rats were administered 1ml ofphysiological saline 24 h prior to and after p. o.,followed by a single i.v. injection of 1ml of EPI at9mg/kg. (Germain et al., 2003).
� Group 3, preventative group: rats were admini-stered 1ml of QE (0.33mg/kg) 24 h prior to andafter p. o., followed by a single i.v. dose of 1ml ofEPI at 9mg/kg.
EPI and QE were reconstituted in physiologicalsaline.
Serum sample selection and hepatic function test
After administration of the last treatments, the ratswere fasted for 24 h and were then sacrificed bydecapitation to collect blood samples. Blood wasallowed to clot and was centrifuged at 3000 rpm for10min at 4 1C to obtain sera (Bioblock scientific, Sigma1-15K). Serum glutamate oxaloacetate transaminase(SGOT) and serum glutamate pyruvate transaminase(SGPT) activities as markers of hepatic function weremeasured spectrophotometrically at 505 nm by the

ARTICLE IN PRESSM. Kebieche et al. / Experimental and Toxicologic Pathology 61 (2009) 161–167 163
method of Reitman et al. (1957) (UV mini 1240 UV–visspectrophotometer Shimadzu, China).
Preparation of cytosol and mitochondria matrix
fractions
Cytosol fractions were extracted as described bySanmugapriya and Venkataraman (2006). Hepatictissues were cut into small pieces and the requiredquantity was weighed and homogenized with threevolumes of phosphate buffer (0.1M; pH 7.4) containingKCl (1.17%) using a Dounce homogenizer (The KontesGlass company, an ISO-9001 certified firm, New Jersey,USA). Supernatant was centrifuged at 10,000 rpm for45min at 4 1C. The supernatant obtained from the spinwas used as a source of catalase (CAT), superoxidedesmutase (SOD), malonedialdehyde (MDA) and glu-tathione (GSH). Protein estimation was performed byLowry et al. (1951) method.
Mitochondrial matrix (stroma) was prepared byapplying the method described by Rustin et al. (1994)and Fan et al. (2005). Livers were quickly removed andperfused with 0.86% cold saline to completely drain allthe red blood cells, chopped into small pieces and placedinto ice-cold isolation buffer for mitochondria (10mMtris–HCl, pH 7.4, 250mM Sucrose, 0.5mM ethylenediamine tetra acetic acid (EDTA) and 0.5% bovineserum albumin). After being homogenized, the homo-genate was centrifuged at 10,000 rpm for 10min at 4 1C.Mitochondrial pellets were washed twice with and thenresuspended in isolation buffer. The mitochondrialmatrix was extracted from freshly prepared mitochon-dria by freezing and defrosting with repeated homo-genization in order to burst mitochondria. Aftercentrifugation at 10,000 rpm for 10min, the supernatantwas the source of CAT, SOD and MDA. Proteinestimation was performed by Lowry et al. (1951)method.
Biochemical evaluation of MDA, GSH, CAT and
SOD in rat liver cells and mitochondria
MDA levels in both the cytosol and the mitochondriawere evaluated as follows (Ohkawa et al., 1979). First,0.5ml of cytosol fraction or mitochondria matrix wasadded to 0.5ml of trichloroacetic acid (TCA) (20%) and1ml of thiobarbituric acid reactive substances (TBARS)(0.67%). The mixture was placed in boiling water.The tubes were moved to an ice-bath, received 4ml ofn-butanol and were centrifuged at 3000 rpm for 15min.The optical density of the supernatant was then assessedat 532 nm (UV mini 1240 UV–vis spectrophotometerSHIMADZU, China). MDA amounts are expressed asnMl/g of liver and were calculated using a standardcurve prepared under the same conditions with a
solution of 1,1,3,3-tetraetoxypropane that producesMDA after hydrolysis.
Levels of GSH were assessed only in the cytosol usingEllman assay (1959). Then, 50 mL of the cytosol fractionwere diluted in 10ml of phosphate buffer (0.1M,pH 8.0). Twenty microliter of 5, 50-dithiobis 2-nitro-benzoic acid (0.01M) (DTNB) were added to 3ml of themixture dilution. After 15min, the absorbance ofthionitrobenzoic acid (TNB) produced after oxidationof GSH by DTNB was evaluated at 412 nm against ablank prepared by TCA (5%) under the same condi-tions. The GSH amounts were calculated using astandard curve of GSH, and were expressed in mM/g.
Cytosolic and mitochondrial CAT assessment wasperformed by Clairborne (1985) method. This assay isbased on the disappearance of H2O2 at 25 1C in thepresence of a cytosolic or mitochondrial enzyme source.Briefly, the assay mixture contained 1ml of phosphatebuffer (KH2PO4 0.1M, pH 7.2), 0.975ml of freshlyprepared H2O2 (0.091M) and 0.025ml of the enzymaticsource. The absorbance was measured at 240 nm eachminute, for 2min. The enzymatic activity was calculatedin terms of IU/mg of protein.
Cytosolic and mitochondrial SOD assessment wasperformed by Beauchamp and Fridovich (1971) techni-que. The assay mixture contained 2ml of reactive milieu(cyanide of sodium 10�2M, NBT solution 1.76� 10�4,EDTA 66mM, methionin 10�2M and riboflavin 2 mM,pH 7.8) and 5 mL of the enzymatic source (cytosol ormitochondrial matrix). This solution mixture wasexposed to a 15W lamp for 10min to induce thephotoreaction of riboflavin and O2. The reduction ofNBT by a superoxide anion on formazan was measuredspectrophotometrically at 560 nm. The enzymatic activ-ity was calculated in terms of IU/mg of protein.
Statistical studies
The numerical and graphical results are presented asmean7standard error (SE). The significance of thedifference between two treatment groups was verified bythe Student’s t-test. The degree of statistical significancewas set at a level of po0.05. Statistical calculations werecarried out using the Statviews 4.5 statistical package(Abacus Concept, Int) and the Exel6.0 (Microsoft, Inc.).
Results
Biochemical evaluation of hepatic dysfunction
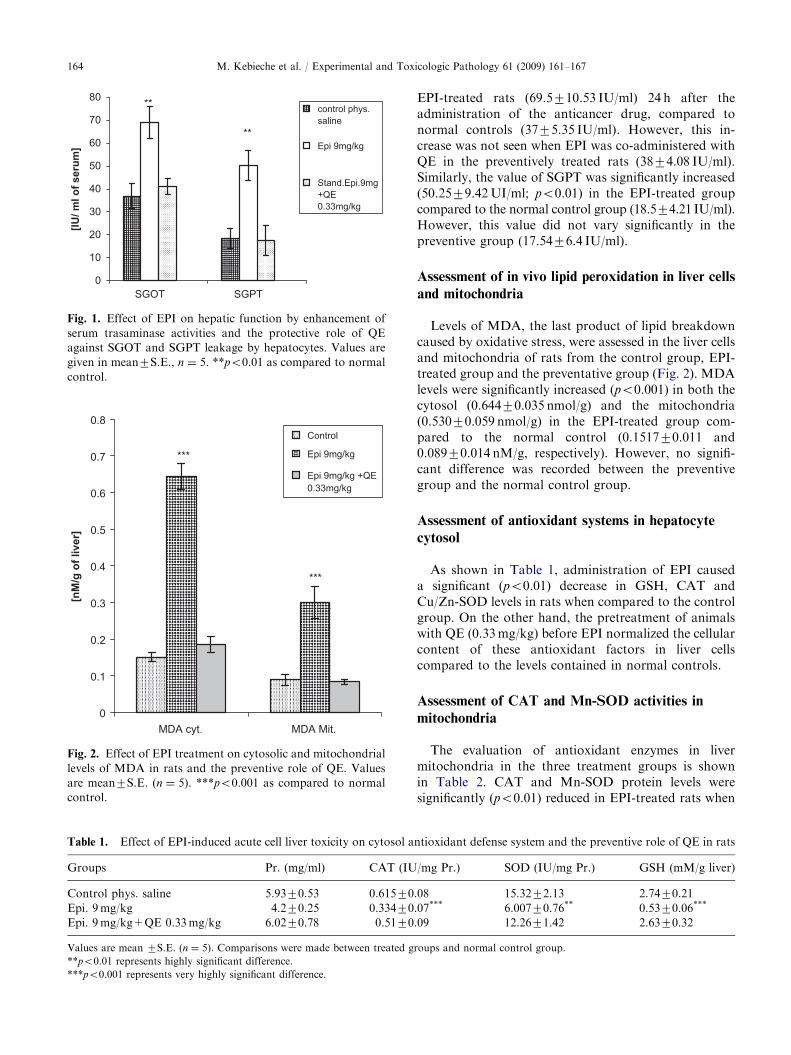
The serum SGOT and SGPT levels in the normalgroup, the EPI-treated group and the preventativegroup that received QE are shown in Fig. 1. There wasa significant increase (po0.05) in SGOT activity levels in
ARTICLE IN PRESS
0
10
20
30
40
50
60
70
80
SGOT
[IU/ m
l of s
erum
]
control phys.saline
Epi 9mg/kg
Stand.Epi.9mg+QE0.33mg/kg
**
**
SGPT
Fig. 1. Effect of EPI on hepatic function by enhancement of
serum trasaminase activities and the protective role of QE
against SGOT and SGPT leakage by hepatocytes. Values are
given in mean7S.E., n ¼ 5. **po0.01 as compared to normal
control.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
MDA cyt.
[nM
/g o
f liv
er]
Control
Epi 9mg/kg
Epi 9mg/kg +QE0.33mg/kg
***
***
MDA Mit.
Fig. 2. Effect of EPI treatment on cytosolic and mitochondrial
levels of MDA in rats and the preventive role of QE. Values
are mean7S.E. (n ¼ 5). ***po0.001 as compared to normal
control.
Table 1. Effect of EPI-induced acute cell liver toxicity on cytosol a
Groups Pr. (mg/ml) CAT (IU
Control phys. saline 5.9370.53 0.61570
Epi. 9mg/kg 4.270.25 0.33470
Epi. 9mg/kg+QE 0.33mg/kg 6.0270.78 0.5170
Values are mean 7S.E. (n ¼ 5). Comparisons were made between treated g
**po0.01 represents highly significant difference.
***po0.001 represents very highly significant difference.
M. Kebieche et al. / Experimental and Toxicologic Pathology 61 (2009) 161–167164
EPI-treated rats (69.5710.53 IU/ml) 24 h after theadministration of the anticancer drug, compared tonormal controls (3775.35 IU/ml). However, this in-crease was not seen when EPI was co-administered withQE in the preventively treated rats (3874.08 IU/ml).Similarly, the value of SGPT was significantly increased(50.2579.42UI/ml; po0.01) in the EPI-treated groupcompared to the normal control group (18.574.21 IU/ml).However, this value did not vary significantly in thepreventive group (17.5476.4 IU/ml).
Assessment of in vivo lipid peroxidation in liver cells
and mitochondria
Levels of MDA, the last product of lipid breakdowncaused by oxidative stress, were assessed in the liver cellsand mitochondria of rats from the control group, EPI-treated group and the preventative group (Fig. 2). MDAlevels were significantly increased (po0.001) in both thecytosol (0.64470.035 nmol/g) and the mitochondria(0.53070.059 nmol/g) in the EPI-treated group com-pared to the normal control (0.151770.011 and0.08970.014 nM/g, respectively). However, no signifi-cant difference was recorded between the preventivegroup and the normal control group.
Assessment of antioxidant systems in hepatocyte
cytosol
As shown in Table 1, administration of EPI causeda significant (po0.01) decrease in GSH, CAT andCu/Zn-SOD levels in rats when compared to the controlgroup. On the other hand, the pretreatment of animalswith QE (0.33mg/kg) before EPI normalized the cellularcontent of these antioxidant factors in liver cellscompared to the levels contained in normal controls.
Assessment of CAT and Mn-SOD activities in
mitochondria
The evaluation of antioxidant enzymes in livermitochondria in the three treatment groups is shownin Table 2. CAT and Mn-SOD protein levels weresignificantly (po0.01) reduced in EPI-treated rats when
ntioxidant defense system and the preventive role of QE in rats
/mg Pr.) SOD (IU/mg Pr.) GSH (mM/g liver)
.08 15.3272.13 2.7470.21
.07*** 6.00770.76** 0.5370.06***
.09 12.2671.42 2.6370.32
roups and normal control group.

ARTICLE IN PRESS
Table 2. Effect of EPI-induced acute liver cell toxicity on mitochondrial antioxidant defense system, CAT and Mn-SOD activities,
and the preventive role of QE in rats
Groups Pr. (mg/ml) CAT (IU/mg Pr.) SOD (IU/mg Pr.)
Control, phys. saline 0.29170.05 6.26471.428 190.56716.72
Epi.9mg/kg 0.08870.022* 1.09970.207** 56.2379.69***
Epi.9mg/kg+QE0.33mg/kg 0.24270.038 5.82971.277 5.82971.277
Values are mean7S.E. (n ¼ 5). Comparisons were made between treated groups and normal control group.
*po0.05 represents significant difference.
**po0.01 represents highly significant difference.
***po0.001 represents very highly significant difference.
M. Kebieche et al. / Experimental and Toxicologic Pathology 61 (2009) 161–167 165
compared with the normal control group. The activitiesof these tranaminases in the preventive group did notvary significantly when compared to those measured inthe normal control group.
Discussion
EPI is an anti-tumour derivative of doxorubicin. Thisanthracycline analog is widely used in clinical oncologypractice due to its good therapeutic efficacy andlow cardiotoxicity (Li et al., 2005; Allen, 1992). It isprimarily metabolized by the liver and producesmetabolites including epirubicinol and EPI glucoronide(Weenen et al., 1984). In hepatocytes, doxorubicin andEPI are considered the most toxic anthracyclines (LeBot et al., 1988; Andersson et al., 1999). Despite thistoxicity, EPI has been used extensively in Europe as ananticancer drug (Ganzina, 1983).
Since oxidative stress is considered to be the majorevent responsible for anthracycline toxicity, extensiveinterest has arisen in the investigation of the range oftheir pro-oxidant power and their harmfulness to organsand cells. Interest has also arisen in the identification ofcompounds that are capable of modulating theseinjuries. Studies with cardiomyocyte cultures and EPIhave reported decreased cell viability related to mod-ifications in the biophysical and biochemical propertiesof the cellular membrane by the generation of ROS(Chan et al., 1996; Germain et al., 2003). Indeed, acorrelation between EPI clearance and serum transami-nase activities indicating hepatic dysfunction has beenreported in patients with cancer following administra-tion of this drug (Le Bot et al., 1988; Dobbs et al., 2003).
Since biochemical data concerning the oxidativestatus in EPI-induced hepatotoxicity in vivo are notwell documented, we investigated stress markers of EPIin both liver cells and mitochondria. To assess EPIhepatotoxicity, we intravenously administered EPI tothe rats at a cumulative dose of 9mg/kg, according toGermain et al. (2003). Twenty-four hours later, weobserved a significant enhancement in serum tranami-nase activities as well as the loss of pro-oxidant/
antioxidant system equilibrium due to oxidative stress.Indeed, the results of the current study demonstrated asignificant increase in serum SGOT and STGP, indicat-ing considerable hepatocellular necrosis in EPI-treatedgroup liver cells (Fig. 1). This leads to the elevation ofserum enzyme markers that are released from the liverinto the blood (Twelves et al., 1992; Ashork Shenoyet al., 2002). Preventive administration of QE at a doseof 0.33mg/kg in association with EPI attenuated theincreased quantities of serum transaminases measuredafter EPI treatment and normalized the activity levels ofthese enzymes to those observed in the control group.Unfortunately, EPI hepatotoxicity rat models have notbeen well documented in the literature. However, thehepatoprotective effect of flavonoids was found in otherdrugs and chemicals in hepatotoxic rat models usingcyclophosphamid, vinblastin or CCl4 (Lahouel et al.,2004; Sanmugapriya and Venkataraman, 2006; Kauret al., 2006; Arora et al., 2003; Raja et al., 2007).
In this study, the significant increase in MDA levels inthe cytosol and liver mitochondria after EPI treatmentsuggests enhanced lipid peroxidation causes tissuedamage. This also suggests that the failure of antiox-idant defense mechanisms prevents the formation ofexcessive free radicals (Sanmugapriya and Venkatara-man, 2006). This lipid peroxidation may explain the cellleakage of transaminase enzymes from liver cells into theblood. On the other hand, preventive administration ofQE in association with EPI significantly reversed thesechanges. These results are similar to those obtainedwhen flavonoids of propolis were used against cyclopho-sphamid- and vinblastin-induced liver injury (Lahouelet al., 2004).
Regarding non-enzymatic antioxidants, GSH-reducedGSH-constitutes the first line of defense against freeradicals. The depletion of GSH has been shown to beassociated with chemical-induced hepatotoxicity (Rajaet al., 2007; Hewawasam et al., 2003) and chemother-apeutic drugs such as cyclophosphamid and vinblastine(Lahouel et al., 2004). In this study, the results revealeda significant depletion of hepatic GSH in EPI-treatedrats. On the other hand, hepatic GSH was increasedwhen the animals were treated preventatively with QE,

ARTICLE IN PRESSM. Kebieche et al. / Experimental and Toxicologic Pathology 61 (2009) 161–167166
possibly due to de novo GSH synthesis or GSHregeneration following ROS neutralization by thephenolic and flavonoid compounds. Because of theirhigh reactivity and short life, ROS in this study havebeen analysed indirectly in vivo by measuring thechanges in antioxidases including SOD and CAT.SOD catalyses the dismutation of O2
d radicals intoH2O2 and O2. In parallel, CAT decomposes H2O2 intoO2 and H2O. These reactions constitute a mutuallysupportive team of defense against ROS (Venukumarand Latha, 2002; Halliwell and Gutteridge, 1999). Thereduced activity of SOD and CAT was observed notonly in the cytosol, but also in mitochondria for Mn-SOD when the rats were administered EPI. This couldexplain the intense production of superoxide anion inthe respiratory chain following the probable alterationin its electron transport system. The observed reductionin proteins in liver cells could also explain the loss ofantioxidase activities. These events may be generalizedfor all cell types.
In the current study, the treatment of animalswith QE maintained the normal content of theantioxidant enzymes analysed. Therefore, QE showedantioxidant properties against ROS in vivo. Accord-ing to the literature, a diet rich in flavonoids reducesthe risk for oxidative stress-related chronic diseasessuch as diabetes, cancer and coronary heart disease(Skibola and Smith, 2000; Brooks et al., 2002). QE isone of the most frequently studied dietary flavonoids,ubiquitously present in various vegetables, fruits, seeds,nuts, tea and red wine and possesses excellent freeradical scavenging power (Beecher, 1999; Bors et al.,1994).
The protection of the liver from injuries after co-administration of EPI and QE in rats was established atmultiple levels. Lipid peroxidation was inhibited andconsequently the transaminases leakage from liver cellsinto the blood was normalized. The maintenance ofGSH storage, the more important reducing potential inthe cell. Finally, in the ultimate stage, QE was able toconserve the first line of antioxidant defense, theantioxidases, in both the cytosol and mitochondria.These results suggest that the side effects observedduring human chemotherapy treatment with EPI wouldbe reduced by the simultaneous administration ofnutraceutical products rich in QE.
References
Allen A. The cardiotoxicity of chemotherapeutic drugs.
Semiological Oncol 1992;19:529–42.
Andersson B, Eksborg S, Vidal R, Sundberg M, Carlberg M.
Anthraquinone-induced cell injury: acute toxicity of carmi-
nomycin, epirubicin, idarubicin and mitoxantrone in
isolated cardiomyocytes. Toxicology 1999;135:11–20.
Arora S, Kaur K, Kaur S. Indian medicinal plants as
a reservoir of protective phytochemicals. Teratogenesis
Carcinogenesis Mutagenesis Suppl 2003;1:301–12.
Ashork Shenoy K, Somayaji SN, Bairy KL. Evaluation of
hepatoprotective activity of Gingo bilobain rats. Indian
J Pharmacol 2002;46:170–4.
Beauchamp C, Fridovich I. Assay of superoxide dismutase.
Anal Biochem 1971;44:276–87.
Beecher GR. Flavonoids in foods. Antioxidant food supple-
ments in human health; 1999. p. 269–81.
Bors W, Michel C, Saran M. Flavonoid antioxidants: rate
constants for reactions with oxygen radicals. Methods
Enzymol 1994;234:420–9.Brind AM. Drugs that damage the liver. Medicin 2007;35:26–30.
Brooks PS, Digernes SB, Parks DA, Darley-Usmar V.
Mitochondrial function in response to cardiac ischemia-
reperfusion after treatment with quercetin. Free Radical
Bio Med 2002;32:1220–8.
Chan EM, Thomas MJ, Bady B, Tibbits GF. Effects of
doxorubicin, 40-epirubicin and antioxidant enzymes on the
contractibility of isolated cardiomyocytes. Can J Physiol
Pharmacol 1996;74:904–10.
Chen HY, Lin YC, Hsieh CL. Evaluation of antioxidant
activity of aqueous extract of some selected nutraceutical
herbs. Food Chem 2007;104:1418–24.
Clairborne A. Catalase activity. In: Greenwald RA, editor.
CRC Handbook of Methods for Oxygen Radical Research.
Boca Raton, FL: CRC Press; 1985. p. 283–4.
Dobbs NA, Twelves CJ, Gregory W, Cruikshanka C, Richards
MA, Rubens RD. Epirubicin in patients with liver
dysfunction: development and evaluation of a novel dose
modification scheme. Eur J Cancer 2003;39:580–6.
Ellman GL. Tissue sulfhidryl groups. Arch Biochem Biophys
1959;82:70–7.
Fan Q, Huang CG, Jin Y, Feng B, Miao H, Li W, et al. Effects
of shark hepatic stimulator substance on the function and
antioxidant capacity of liver mitochondria in an animal
model of acute liver injury. Acta Biochem Biophys Sin
2005;37:507–14.
Farber JL. Mechanism of cell injury by activated oxygen
species. Environ Health Perspect 1994;102:17–24.
Ganzina F. 4-Epi-doxorubicin, a new analogue of doxorubi-
cin: a preliminary overview of preclinical and clinical data.
Cancer Treat Rev 1983;10:1–22.
Germain E, Bonnet P, Aubourg L, Grangeponte MC, Chajes
V, Bougnoux P. Anthracycline-induced cardiac toxicity is
not increased by dietary omega-3 fatty acids. Pharmacol
Res 2003;47:111–7.
Goebel M. Oral idarubicin an anthracycline derivative with
unique properties. Ann Hematol 1993;66:33–43.
Halliwell B, Gutteridge JMC. Free radicals in biology and
medicine, 3rd ed. Oxford: Clarendone press; 1999. p. 22.
Hewawasam RP, Jayatilaka KAPW, Pathirana C, Mudduwa
LKB. Protectiveeffect of Asteracantha longifolia extracts
mouse liver injury induced by carbon tetrachloride and
paracetamol. J Pharm Pharmacol 2003;55:1413–8.
Kaur G, Alam MS, Jabbar Z, Javed K, Athar M. Evalua-
tion of antioxidant activity of Cassia siamea flowers.
J Ethnopharmacol 2006;108:240–348.
Lahouel M, Boulkour S, Segueni N, Fillastre JP. The
flavonoids effect against vinblastine, cyclophosphamid

ARTICLE IN PRESSM. Kebieche et al. / Experimental and Toxicologic Pathology 61 (2009) 161–167 167
and paracetamol toxicity by inhibition of lipid-peroxidation
and increasing liver glutathione concentration. Pathol Biol
2004;52:314–22.
Lanza E, Rozza A, Favalli L, Monti E, Poggi P, Villani F. The
rat model in the comparative evaluation of anthracyclines
cardiotoxicity. Tumori 1989;75:533–6.
Le Bot MA, Begue JM, Kernaleguen D, Robert J, Ratanasa-
vanh D, Riche C, et al. Different cutotoxicity and
metabolism of doxorubicin, daunorubicin, epirubicin,
esorubicin and idarubicin in cultured human and rat
hepatocytes. Biochem Pharmacol 1988;37:3877–87.
Li R, Dong L, Huang J. Ultra performance liquid chromato-
graphy–tandem mass spectrometry for the determination of
epirubicin in human plasma. Anal Chem Acta 2005;546:
167–73.
Lowry OH, Rosegrough NJ, Farr AL, Randall RJ. Protein
measurement with the folin phenol reagent. J Biol Chem
1951;193:265–75.
Mazue G, Iatropoulos M, Imondi A, Castellino S, Brughera
M, Podesta A, et al. Anthracyclines: a review of general and
special toxicity studies. Int J Oncol 1995;7:713–26.
Nielsen D, Jensen JB, Dom Bernowski P, Munck O, Fogh J,
Brynjoff L, et al. Epirubicin cardiotoxicity: a study of 135
patients with advanced breast cancer. J Clin Oncol 1990;
8:1806–10.
Ohkawa H, Ohishi N, Yagi K. Assay of lipid peroxides in
animal tissue by thiobarbituric reaction. Anal Biochem
1979;95:351–8.
Pannala AS, Rice-Evans CA, Haliwell B, Singh S. Inhibition of
peroxynitrite-mediated tyrosine nitration by catechin poly-
phenols. Biochem Biophys Res Commun 1997;232:164–8.
Pong K. Oxidative stress in neurodegenerative diseases:
therapeutic implications for superoxide dismutase mi-
metics. Expert Opinion Biol Ther 2003;3:127–39.
Raja S, Nazeer Ahamed KFH, Kumar V, Mukherjee K,
Bandyopadyay A, Mukherjee PK. Antioxidant effect of
Cytisus scoparius against carbon tetrachloride treated liver
injury in rat. J Ethnopharmacol 2007;109:41–7.
Reitman S, Frankel S, Amer J. A colorimetric method for the
determination of serum glutamic oxaloacetic and glutamic
pyruvic transaminases. Am J Clin Pathol 1957;28:56–63.
Rustin P, Chretien D, Bourgeron T, Gerard B, Rotig A,
Munnich A. Biochemical and molecular investigation in
respiratory chain deficiences. Clin Chem Acta 1994;228:
35–51.
Sanmugapriya E, Venkataraman S. Studies of hepatoprotec-
tive and antioxidant actions of Strychonos potatorum Linn.
Seeds on CCl4 induced acute hepatic injury in experimental
rats. J Ethnopharmacol 2006;105:154–60.
Skibola CF, Smith MT. Potential health impacts of exces-
sive flavonoid intake. Free Radical Biol Med 2000;29:
375–83.
Twelves CJ, Dobbs NA, Michael V. Clinical pharmacokinetics
of epirubicin: the importance of liver biochemistry tests. Br
J Cancer 1992;66:765–9.
Venukumar MR, Latha MS. Antioxidant activity of Curculigo
orchioides in carbon tetrachloride induced hepatopathy in
rats. Indian J Clin Biochem 2002;17:80–7.
Weenen H, Van Maanen JMS, de Planque MM. Metabolism
of 40 modified of analogs of doxorubicin. Unique glucur-
onidation pathway for 40-epidoxorubicin. Eur J Cancer
Clin Oncol 1984;20:919–26.
Watkins PB, Seef LB. Drug-induced liver injury: summary of a
single topic clinical research committee. Hepatology 2006;
43:618–31.
Yeung TK, Simmonds RH, Hopewell JW. The relative toxicity
of intravenous and intraperitoneal doses of epirubicin.
Cancer Chemother Pharmacol 1989;24:211–8.




















