
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY: METABOLIC MECHANISMS OF TOXICITY
30
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY: METABOLIC MECHANISMS OF TOXICITY A. Buckpitt, * B. Boland, M. Isbell, D. Morin, M. Shultz, R. Baldwin, K. Chan, A. Karlsson, C. Lin, A. Taff, J. West, M. Fanucchi, L. Van Winkle, and C. Plopper Departments of Molecular Biosciences and Anatomy, Physiology, and Cell Biology, School of Veterinary Medicine, University of California, Davis, CA 95616 ABSTRACT The lung, which is in intimate contact with the external environment, is exposed to a number of toxicants both by virtue of its large surface area and because it receives 100% of the cardiac output. Lung diseases are a major disease entity in the U.S. population ranking third in terms of morbidity and mortality. Despite the importance of these diseases, key issues remain to be resolved regarding the interactions of chemicals with lung tissue and the factors that are critical determinants of chemical-induced lung injury. The importance of cytochrome P450 monooxygenase dependent metabolism in chemical-induced lung injury in animal models was established over 25 years ago with the furan, 4-ipomeanol. Since then, the significance of biotransformation and the reasons for the high degree of pulmonary selectivity for a myriad of different chemicals has been well documented, mainly in rodent models. However, with many of these chemicals there are substantial differences in the susceptibility of rats vs. mice. Even within the same species, varied levels of the respiratory tract respond differently. Thus, key pieces of 791 DOI: 10.1081/DMR-120015694 0360-2532 (Print); 1097-9883 (Online) Copyright q 2002 by Marcel Dekker, Inc. www.dekker.com * Corresponding author. Alan Buckpitt, Department of Molecular Biosciences, School of Veterinary Medicine, University of California, Davis, CA 95616. E-mail: [email protected] DRUG METABOLISM REVIEWS Vol. 34, No. 4, pp. 791–820, 2002 ©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc. MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016 Drug Metabolism Reviews Downloaded from informahealthcare.com by CDL-UC Davis on 12/19/14 For personal use only.
-
Upload
ua-birmingham -
Category
Documents
-
view
0 -
download
0
Transcript of NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY: METABOLIC MECHANISMS OF TOXICITY

NAPHTHALENE-INDUCED RESPIRATORY TRACTTOXICITY: METABOLIC MECHANISMS OF
TOXICITY
A. Buckpitt,* B. Boland, M. Isbell, D. Morin, M. Shultz,
R. Baldwin, K. Chan, A. Karlsson, C. Lin, A. Taff, J. West,
M. Fanucchi, L. Van Winkle, and C. Plopper
Departments of Molecular Biosciences and Anatomy, Physiology, and
Cell Biology, School of Veterinary Medicine, University of California,
Davis, CA 95616
ABSTRACT
The lung, which is in intimate contact with the external environment, is
exposed to a number of toxicants both by virtue of its large surface area and
because it receives 100% of the cardiac output. Lung diseases are a major
disease entity in the U.S. population ranking third in terms of morbidity and
mortality. Despite the importance of these diseases, key issues remain to be
resolved regarding the interactions of chemicals with lung tissue and the
factors that are critical determinants of chemical-induced lung injury. The
importance of cytochrome P450 monooxygenase dependent metabolism in
chemical-induced lung injury in animal models was established over 25 years
ago with the furan, 4-ipomeanol. Since then, the significance of
biotransformation and the reasons for the high degree of pulmonary selectivity
for a myriad of different chemicals has been well documented, mainly in
rodent models. However, with many of these chemicals there are substantial
differences in the susceptibility of rats vs. mice. Even within the same species,
varied levels of the respiratory tract respond differently. Thus, key pieces of
791
DOI: 10.1081/DMR-120015694 0360-2532 (Print); 1097-9883 (Online)Copyright q 2002 by Marcel Dekker, Inc. www.dekker.com
*Corresponding author. Alan Buckpitt, Department of Molecular Biosciences, School of Veterinary
Medicine, University of California, Davis, CA 95616. E-mail: [email protected]
DRUG METABOLISM REVIEWS
Vol. 34, No. 4, pp. 791–820, 2002
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

data are still missing when evaluating the applicability of data generated in
rodents to primates, and as a result of this, there are substantial uncertainties
within the regulatory community with regards to assessing the risks to humans
for exposure to some of these chemicals. For example, all of the available data
suggest that the levels of cytochrome P450 monooxygenases in rodent lungs
are 10–100 times greater than those measured in the lungs of nonhuman
primates or in man. At first glance, this suggests that a significant margin of
safety exists when evaluating the applicability of rodent studies in the human,
but the issues are more complex. The intent of this review is to outline some of
the work conducted on the site and species selective toxicity and metabolism
of the volatile lung toxic aromatic hydrocarbon, naphthalene. We argue that a
complete understanding of the cellular and biochemical mechanisms by which
this and other lung toxic compounds generate their effects in rodent models
with subsequent measurement of these cellular and biochemical events in
primate and human tissues in vitro will provide a far better basis for judging
whether the results of studies done in rodent models are applicable to humans.
Key Words: Naphthalene; Clara cell toxicity; CYP2F; Covalent binding;
Naphthalene epoxide; Naphthoguinone; Lung; Human; Rodent cytochrome
P450
INTRODUCTION
The respiratory system provides a critical interface with the environment, and as
a result, it is an important portal of entry for volatile chemicals and gases. In addition,
the lung receives the entire cardiac output from the right side of the heart and thus is
exposed to chemicals via the circulation. Nononcogenic and oncogenic pulmonary
diseases are a leading cause of death and are a major factor in morbidity and disability
in the United States.[1] While cigarette smoking is an important contributor to these
diseases, other environmental factors including exposure to chemicals in the air, food,
and drinking water are likely to play a role in these disease processes. The
identification of specific factors important in the etiology of human pulmonary
diseases is a difficult and uncertain process for several reasons. The known variations
in response of different rodent species to pulmonary toxicants requiring bioactivation
results in significant uncertainties in the extrapolation of the experimental studies
conducted in animal models to the human. In addition, good methods for
distinguishing between exposures that are likely to disrupt cellular homeostasis in the
respiratory tract from those that are inconsequential are not currently available. There
are several potential approaches to address these issues all of which require a good
understanding of biochemical and cellular events that occur in target cells and that are
critical to the injury process. Once these are well understood, biomarker approaches,
which indicate that an exposure has occurred at sufficient intensity and of sufficient
duration to result in events that are likely to result in untoward effects in the cell, can
be developed and carefully validated in animal models. The overall intent would be to
BUCKPITT ET AL.792
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

then apply these biomarkers to exposed human populations. A second approach
would involve the development of in vitro methods capable of monitoring those
biochemical/cellular events that are clearly tied to toxicity in animal models with the
intent of applying these to human or other primate tissues. Although our current
understanding of the mechanisms involved in the toxicity and potential oncogenicity
of naphthalene is not complete, there has been considerable progress in delineating
some of the key issues involved in these processes in animal models. Accordingly, the
intent of this manuscript is to outline some of these approaches and to review what is
known about the metabolic/cellular mechanisms of toxicity of naphthalene. In the
process, we will identify critical gaps in our knowledge where additional data would
be useful in refining our assessments for potential toxicity in the human.
ENVIRONMENTAL OCCURRENCE—POTENTIAL FOR HUMANEXPOSURE
Naphthalene, a slightly water soluble (32 mg/L) volatile aromatic hydrocarbon,
is present in both groundwater and air emissions from a variety of sources.[2]
Naphthalene and close structural congeners are prominent contaminants of old coal
and oil gasification sites; the concentrations of naphthalene are more than 10 fold
higher than any of the other polycyclic aromatic hydrocarbons (PAH) identified in
ambient air above the site.[3] Over 3000 such sites have been identified and many are
in heavily populated urban areas.[4] In a recent survey of hazardous waste sites,
naphthalene was listed as present in 47 of the 300 sites surveyed. Naphthalene has
been detected in urban air (average concentration of 0.9mg/m3)[5] and is also a
prominent pyrolysis product of both mainstream and side stream tobacco smoke.[6,7]
The fact that naphthalene has been detected in nearly 40% of the fat samples tested[8]
and in 75% of breast milk samples[9] obtained from humans argues that the U.S.
population is exposed to this compound. Data demonstrating the relatively rapid
metabolism of naphthalene in human liver microsomes[10] argue that the clearance of
this compound in humans is likely to be quite rapid. Despite the high fat/blood
partition ratio for naphthalene, these findings suggest that a considerable portion of
the population is likely exposed to low concentrations of this compound on a frequent
basis. Additional information on the sources of potential human exposure to
naphthalene can be obtained in a very thorough document released by the Agency for
Toxic Substances Disease Registry (ATSDR).[2]
VARIATION IN THE RESPONSE OF RODENTS TO CHEMICAL-INDUCED INJURY TO THE RESPIRATORY TRACT
A number of compounds including dichloroethylene,[11,12] methylene
chloride,[13,14] naphthalene, and close structural analogs of naphthalene (2-
methylnaphthalene),[15] when administered intraperitoneally (ip), result in highly
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 793
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

selective injury to mouse but not rat lung. Again, this clearly underscores the need
to understand the mechanisms by which each of these chemicals elicits untoward
effects in the organism. For example, the selective carcinogenic actions of
methylene chloride in mouse lung have been related to the metabolic activation of
the parent compound by theta class glutathione S-transferase. The finding of very
low theta glutathione S-transferase activities in human lung[16] and that very little
of this theta class enzyme is present in the nucleus of cells in human lung[13] has
been used to argue that this compound is unlikely to be a carcinogen in human
pulmonary tissue.[13] Although these views have been challenged,[14] the fact that
the mouse is highly susceptible to methylene chloride carcinogenicity and that the
differential susceptibility between mice and rats is related to metabolism certainly
raises some concerns regarding the applicability of data derived in mouse
bioassays with methylene chloride to the human.
Acute Pulmonary Injury by Naphthalene
The pulmonary toxicity of naphthalene has been studied extensively by both
our laboratories[17 – 19] and by others.[20 – 22] Clara cells (nonciliated bronchiolar
epithelial cells) lining the airway epithelium of the mouse are the primary target
cells for naphthalene toxicity, irrespective of the route of administration. One of
the most striking features of naphthalene cytotoxicity is the high degree of tissue
and species selectivity. After parenteral administration of low doses of the
compound, the only tissue affected is the respiratory tract (Table 1). Hepatic
necrosis is not observed at any dose of naphthalene tested, while proximal tubular
cells of the kidney are injured only in some mouse strains and only at very high
doses (400 and 600 mg/kg, ip).[21] Swelling of Clara cells in terminal airways is
detected in mice at doses as low as 50 mg/kg, ip. In contrast, in rats even at LD50
doses (1600 mg/kg, ip), airway Clara cells are apparently normal. There is some
swelling of Clara cells in the hamster at the LD50 dose (800 mg/kg, ip). Thus, there
are fundamental differences between these species that can be used as a tool to
determine the relevance of particular metabolites and metabolite-macromolecular
adducts in the processes leading to cytotoxicity.[17] In all of the species tested, no
injury to the alveolar type I or II cells has been observed. In addition to the lesions
observed in the lung, the olfactory region of the nose is sensitive to naphthalene
after intraperitoneal administration[17] in both the mouse and the rat. The rat nasal
olfactory epithelium is injured at lower doses of naphthalene (200 mg/kg, ip) than
the doses required to produce injury in the nasal epithelium of the mouse
(400 mg/kg, ip) (Table 1). Finally, more recent studies investigating the sex and
strain differences in susceptibility to naphthalene toxicity indicate that the female
Swiss Webster mouse is more susceptible to the cytotoxicity of naphthalene than is
the male.[23] As discussed later, the National Toxicology Program (NTP) bioassay
investigating the possible neoplastic effects of naphthalene showed a sex
difference in susceptibility with female mice showing a slight increase in
bronchioloalveolar neoplasms over control whereas in males there was no effect.
BUCKPITT ET AL.794
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

Additional work examining the acute toxicity of naphthalene administered
by inhalation has been completed.[24] Four hour exposures at concentrations as
low as 2 ppm results in detectable Clara cell injury in adult mice[24] but no injury to
Clara cells in rat lung. In mice, 4-hr exposures conducted at concentrations equal
to the current 8-hr human occupational exposure standard (10 ppm, TWA)[25]
produce substantial injury to cells in both the upper and lower respiratory tract of
the mouse (Table 1).
In addition to the species, cell, and tissue variations in susceptibility to
naphthalene induced injury, there are regional differences in susceptibility of the
airway epithelium that are dependent upon route of administration. Clara cells of
murine terminal airways are most susceptible to low doses of naphthalene
administered parenterally (Table 1).[17] Doses of 200 mg/kg and above are required
to observe toxicity in more proximal airways. Although this airway level difference
in susceptibility has been attributed to variations in rates of metabolism to the
epoxide,[26,27] data obtained from the inhalation experiments suggest that this
differential susceptibility may be due to differences in distribution of the parent
compound. After inhalation exposures, cells of the more proximal airways are
injured at lower concentrations than cells of the distal airways (bronchioles) while
higher inhaled concentrations are required to produce injury in terminal airways.
These findings are consistent with the slight water solubility of naphthalene.
While the parenteral doses used in these studies far exceed those likely to be
encountered by human populations via other routes of exposure, the fact that the
injury is localized to the epithelial cells of the respiratory tract after systemic
administration of the compound strongly supports the view that there is an
underlying biochemical basis for this injury. Numerous studies conducted over the
past 15 years have provided considerable insight regarding the initial metabolic
steps necessary for toxicity, but it is clear that additional work is needed to fully
understand the intracellular events that lead to cell death. With inhalation exposures,
Clara cell toxicity is observed in the mouse at exposure concentrations that are
plausible for human occupational exposures. Again, whether these exposure
concentrations are capable of producing any untoward effects in humans is not
known and a clear understanding of the mechanism of injury along with the
development of biomarkers that are capable of indicating exposures that are at levels
likely to result in untoward effects, is important. In addition, testing of naphthalene
for acute toxicity in animal models, which more closely resemble the human also
may provide data useful in assessing the potential toxicity of naphthalene as well as
its close structural analogs (methylnaphthalenes and nitronaphthalenes).
Tolerance to Multiple Naphthalene Exposures
In contrast to the Clara cell toxicity observed after single doses of
naphthalene, multiple daily treatments with naphthalene by both the
intraperitoneal and inhalation routes result in tolerance to high challenge doses
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 795
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

Table 1. Species, Tissue, and Regional Differences in Naphthalene Toxicity
Lung Nasal Epithelium
Species Dose Route
Trachea/Lobar
Bronchus
Terminal
Bronchiole Parenchyma Olfactory Respiratory Comments
Mouse, adult male
LD50 ¼ 380 mg=kg
50 mg/kg ip 0 þ 0 0 0 No toxicity noted in liver or
kidney of male Swiss Webster
mice; ICR mice showed
lesions of proximal tubule at
highest dose (400 and
600 mg/kg)[17,21]
100 mg/kg ip 0 þþ 0 0 0
200 mg/kg ip þ /0 þþþ 0 0 0
300 mg/kg ip þþ þþþþ 0 ND ND
400 mg/kg ip þþþ þþþþ 0 þþþ 0
2–5 ppm ip þ /0 0 0 ND ND Ref. [24]
8.5–11.5 ppm inh þ þ /0 0 ND ND
25–31 ppm inh þþ þ 0 ND ND
72–77 ppm inh þþþ þþþ 0 ND ND
96–111 ppm inh þþþ þþþ 0 ND ND
Mouse, adult
tolerance
200 mg/kg £ 7a ip ND 0 ND ND ND Areas of bronchiolar epithelial
cell hyper-plasia observed
after 7 days, data from
Refs. [28,29]
BU
CK
PIT
TE
TA
L.
79
6
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.
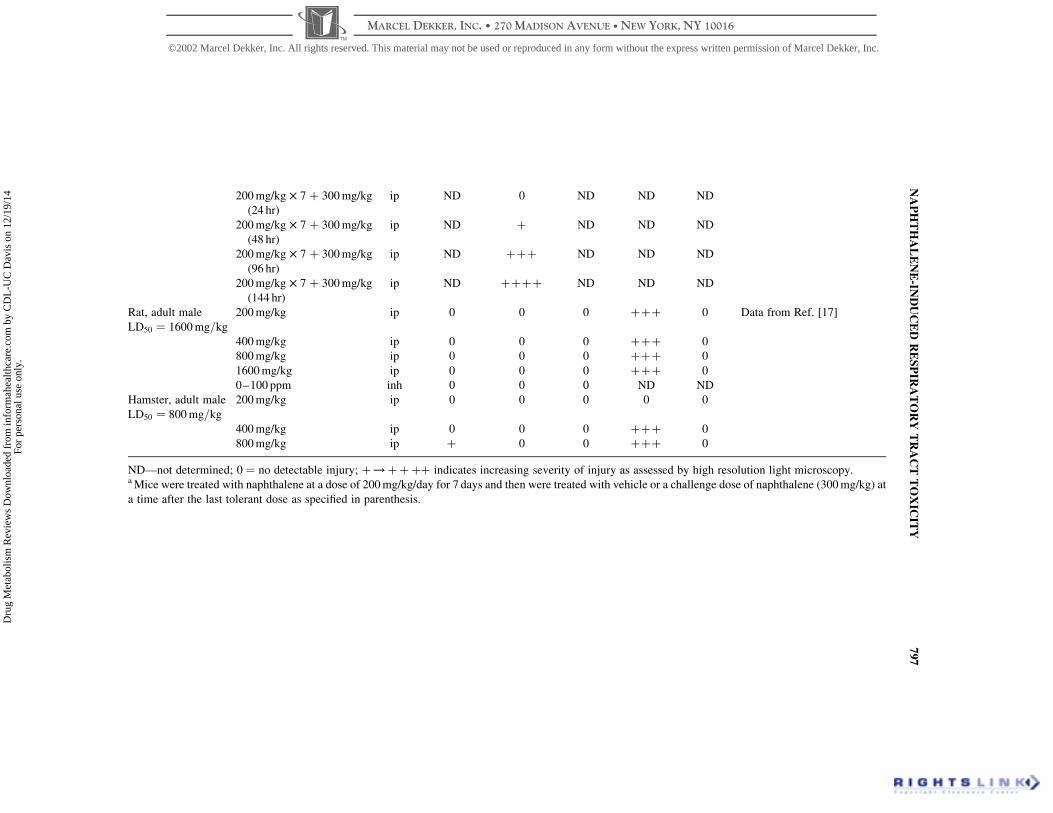
200 mg/kg £ 7 þ 300 mg/kg
(24 hr)
ip ND 0 ND ND ND
200 mg/kg £ 7 þ 300 mg/kg
(48 hr)
ip ND þ ND ND ND
200 mg/kg £ 7 þ 300 mg/kg
(96 hr)
ip ND þþþ ND ND ND
200 mg/kg £ 7 þ 300 mg/kg
(144 hr)
ip ND þþþþ ND ND ND
Rat, adult male
LD50 ¼ 1600 mg=kg
200 mg/kg ip 0 0 0 þþþ 0 Data from Ref. [17]
400 mg/kg ip 0 0 0 þþþ 0
800 mg/kg ip 0 0 0 þþþ 0
1600 mg/kg ip 0 0 0 þþþ 0
0–100 ppm inh 0 0 0 ND ND
Hamster, adult male
LD50 ¼ 800 mg=kg
200 mg/kg ip 0 0 0 0 0
400 mg/kg ip 0 0 0 þþþ 0
800 mg/kg ip þ 0 0 þþþ 0
ND—not determined; 0 ¼ no detectable injury; þ!þþþþ indicates increasing severity of injury as assessed by high resolution light microscopy.a Mice were treated with naphthalene at a dose of 200 mg/kg/day for 7 days and then were treated with vehicle or a challenge dose of naphthalene (300 mg/kg) at
a time after the last tolerant dose as specified in parenthesis.
NA
PH
TH
AL
EN
E-IN
DU
CE
DR
ES
PIR
AT
OR
YT
RA
CT
TO
XIC
ITY
79
7
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

of compound.[28 – 30] As discussed earlier, substantial injury is observed in Clara
cells of mice treated with a single 200 mg/kg dose, ip. In comparison, light
microscopic and morphometric evaluation of airway epithelium from mice treated
daily with 200 mg/kg naphthalene, ip for 7 days showed only slight hyperplasia of
the epithelium in comparison to animals treated with vehicle alone[29] (Table 1). In
addition, this pretreatment regimen markedly decreased the susceptibility of the
epithelium to a large challenge dose of naphthalene (300 mg/kg, ip) when the
challenge dose was administered 24 hr after the last of the seven 200 mg/kg doses
(Table 1). As the time was extended between the last 200 mg/kg dose and the
challenge dose from 24 to 96 hr, the lungs regained a portion of their sensitivity to
the 300 mg/kg challenge dose. These data, showing that the lung becomes tolerant
to multiple doses of naphthalene at dose levels that produced substantial toxicity in
airway epithelial cells after single administration are consistent with the 14-day
and 90-day oral gavage studies. This work demonstrated no significant alterations
in serum enzyme levels, body weight, organ weight, or various indices of immune
function in CD-1 mice treated daily with doses up to 267 mg/kg (14 day) or
133 mg/kg (90 day).[31]
Carcinogenesis Studies
Chronic inhalation exposure of mice to naphthalene (10 or 30 ppm) results in
inflammation in the nose, metaplasia of the olfactory epithelium, and hyperplasia
of the respiratory epithelium.[32,33] No neoplastic effects were noted in male mice,
but a slight increase in alveolar/bronchiolar adenomas and alveolar/bronchiolar
carcinomas were noted at the highest exposure level in female mice. These results
are consistent with our findings that nasal olfactory epithelial cells and bronchiolar
epithelial cells are targets in the mouse after acute exposures to naphthalene and
that administration of the compound daily at high doses over a 7-day period results
in hyperplasia of the respiratory epithelium in the terminal airways.[29]
Recently, the NTP 2-year bioassay has been completed in rats. Animals were
exposed to vapor concentrations of 0, 10, 30, and 60 ppm. The nasal epithelium
was found to be a primary target for these exposures.[34,35] A concentration-
dependent increase in adenomas of the respiratory epithelium of the nose and of
neuroblastomas of the olfactory epithelium are considered to be significant in
these rat studies and have raised concerns about naphthalene as a potential human
carcinogen. Slight hyperplasia of the alveolar epithelium was noted but only in
female rats. Finally, inflammation of the olfactory epithelium was observed in
both males and females at all concentrations of naphthalene studied.
The species and regional differences in susceptibility to naphthalene
carcinogenicity noted in the cancer bioassays in both mice and rats correspond to
the differences noted in sensitivity during the acute toxicity studies. Mouse lung
airway epithelium is considerably more susceptible than rat lung airway
epithelium after single ip doses of naphthalene, and female mice are more
BUCKPITT ET AL.798
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

susceptible than males.[23] The olfactory nasal epithelium of both species is highly
susceptible to naphthalene after parenteral administration of the compound with
the rat being more susceptible than the mouse. Since the regional and species
differences in susceptibility to both the cytotoxic and neoplastic effects associated
with naphthalene exposure appear to match, the underlying biochemical
mechanisms that lead to these toxicities may be similar. In the following sections,
the importance of metabolism and the role of various reactive metabolites in
cytotoxicity will be discussed.
METABOLISM AS A KEY DETERMINANT IN NAPHTHALENETOXICITY
Importance of P450-Dependent Metabolism
The Clara cell is an important target cell in the lung for cytotoxic and
genotoxic agents requiring metabolic activation. These include aromatic
hydrocarbons such as naphthalene (and several close structural analogs such as
1-nitronaphthalene), furans such as 4-ipomeanol, and nitrosamines such as 4-
(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK).[18,36,37] The sensitivity of
the Clara cell to these agents is presumed to be related to high rates of metabolic
activation catalyzed by P450 monooxygenases localized within this cell.[38]
Interaction of Reactive Metabolites with Critical Cellular Proteins as aDeterminant in Toxicity
The pioneering work of the Millers on the relationship between electrophiles
and cancer was extended to cytotoxicants by Mitchell, Jollow, and their coworkers
in their studies of the hepatotoxicants, acetaminophen and bromobenzene, in the
early 1970s.[39] Later work by Boyd and others[40] expanded these concepts to
include lung toxicants as well. All of this work has been based on the apparent
close association between overall levels of reactive metabolite binding in a
particular tissue with the incidence and severity of toxicity in that tissue. Reactive
metabolite binding does not a priori result in tissue necrosis, and there are
excellent examples, such as with 3-hydroxyacetanilide, where there is substantial
formation of reactive metabolites in the absence of any toxicity.[41]
Early in vivo studies demonstrated an interrelationship between the covalent
binding of reactive naphthalene metabolites and glutathione depletion with the
extent and severity of cytotoxicity in the lung.[42,43] These studies, like those with
acetaminophen and bromobenzene, showed that only at doses resulting in
significant glutathione depletion were significant covalent binding of reactive
metabolites and toxicity evident. All pretreatments that resulted in alterations in
the severity of bronchiolar necrosis after administration of naphthalene also
altered the levels of reactive metabolite(s) bound covalently to protein in the lung
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 799
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

in vivo. Specifically, inhibition of the cytochrome P450 monooxygenases
decreased binding and toxicity while glutathione depletion increased toxicity and
covalent binding of reactive metabolites.[42] Moreover, the rates of metabolic
activation of naphthalene to covalently bound metabolites are substantially higher
in preparations of target cells or in isolated tracheobronchial airways from the
mouse than they are in mouse hepatocytes or in rat airways.[44] Likewise, the
extent of glutathione depletion in the lung in vivo and the rates of metabolism of
naphthalene to reactive metabolites in lung microsomal preparations in vitro are
substantially greater in the mouse than in the rat.[21] The importance of glutathione
as a detoxication pathway for naphthalene metabolism is supported by recent
studies showing that increased glutathione synthesis is one of the underlying
reasons why mice treated with high doses of naphthalene daily become tolerant to
large challenge doses of this compound.[30] However, the tissue selective toxicity
of naphthalene is not reflected by tissue selective binding of reactive metabolites
in vivo.[42] The levels of bound metabolite in the liver and kidney were as high as
in the lung. There are several possibilities to explain these data which are
consistent with a role of reactive metabolites in lung injury including: (1) there are
several different reactive metabolites generated from naphthalene and differences
in target and nontarget tissue are related to the chemical nature of bound
metabolites; (2) binding only occurs in a small population of lung cells and thus
measurements of the level of bound metabolites in vivo considerably
underestimate the levels of reactive metabolite bound in target cell populations;
and (3) macromolecules adducted by reactive metabolites in target and nontarget
tissues differ. The corollary to this third possibility is that a protein that is adducted
in the lung but not the liver, is key to the loss of homeostasis in the lung. We have
recently identified several proteins adducted by reactive naphthalene metabolites
and are in the process of determining both the chemical nature of the intermediate
bound and the peptide residues adducted (Isbell et al., unpublished). It is clear
from the work that we have done thus far that there is a high degree of selectivity
for reactive metabolite binding. Although some of the proteins adducted by
reactive metabolites of naphthalene are high abundance proteins, others, which are
present at very low levels in the cell, are also highly labeled.
In summary, a clear understanding of the catalytic activities of the enzymes
involved in both the metabolic activation of naphthalene and detoxification of
naphthalene metabolites in both rodents and in primates will provide a far better
basis for assessing the potential risks of exposure to naphthalene in the human.
Moreover, delineation of the roles of reactive metabolites in processes that lead to
toxicity should provide targets for the development of biomarkers that can be
validated in rodent models and subsequently applied in humans. In the following,
the enzymes responsible for the formation of both the primary and several of the
secondary metabolites of naphthalene will be discussed individually along with
what is known about their potential to result in cytotoxicity. In each case, the
current state of knowledge of species differences in these processes will be
discussed.
BUCKPITT ET AL.800
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

NAPHTHALENE METABOLISM
Overview
The initial and obligate first reaction in naphthalene metabolism involves the
formation of an unstable 1,2-epoxide[45] (Fig. 1). In buffer at pH 7.4, this
metabolite has a half-life of 2–3 min[46] whereas in solutions of albumin the half-
life is considerably greater (11 min).[47] A number of further metabolites can be
generated directly from the epoxide including 1-naphthol (nonenzymatic),
naphthalene dihydrodiol (microsomal epoxide hydrolase), naphthalene diep-
oxides,[48] and glutathione conjugates (glutathione S-transferase).[45] Each of these
secondary metabolites can undergo further biotransformation, and in two of three
cases (1-naphthol and naphthalene dihydrodiol), these biotransformations can
result in more reactive metabolites that have been implicated in both the cytotoxic
and the potential oncogenic actions associated with naphthalene exposure in
animals. The potential for each of these metabolites to produce cytotoxic and/or
oncogenic effects will be discussed in additional detail along with their further
metabolism.
Naphthalene-1,2-epoxide
Recognition of the importance of regio- and stereochemistry in the formation
and toxicology of epoxides and diol epoxides of larger PAH led to development of
methods to assess both the regio- and stereochemistry of formation of these
unstable chemical entities.[49] As with the larger PAH, some of the cytochrome
P450 monooxygenases show remarkable stereoselectivity in the formation of
naphthalene epoxides.[46,50] Cytochrome P450 2B shows a slight preference for
the formation of the (1S,2R )-naphthalene epoxide (74%) whereas cytochrome
P450 1A1 preferentially generates (1R,2S )-epoxide (73–95%) (Fig. 2). Studies
showing marked differences in the formation of glutathione conjugates in lung vs.
liver microsomal incubations containing naphthalene, glutathione, and glutathione
transferases suggested that there might be differences in the stereoselectivity of
naphthalene epoxide formation,[51] but definitive demonstration of this required
additional structural information.[52] Further studies showed the preferential
formation of (1R,2S )-naphthalene epoxide at high rates in lung microsomes and in
isolated dissected airways of the mouse in comparison to the rat[27,53] and
suggested that the stereochemistry of epoxidation might be important in the target
tissue and species selectivity of naphthalene toxicity (Table 2). Although it is
possible that the toxicologic potency of the naphthalene epoxide enantiomers
differ, we felt it was far more likely that differential susceptibility was related to
differences in the rates of formation of the dihydrodiol or glutathione conjugates
(i.e., possible detoxication reactions), which, in turn, would control the residence
time of the epoxide in the cell. Work conducted in isolated hepatocytes appeared
to support this view. Addition of (1S,2R )-naphthalene epoxide to isolated murine
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 801
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

hepatocytes resulted in significant losses in cell viability while addition of the
(1R,2S )-naphthalene epoxide enantiomer did not.[54] (1R,2S )-Naphthalene
epoxide was metabolized to the dihydrodiol in hepatocytes at much higher rates
than the (1S,2R )-enantiomer suggesting that the longer cellular half-life of the
(1S,2R )-naphthalene epoxide contributes to the higher cytotoxicity observed with
this enantiomer. Definitive analysis of the importance of the stereochemistry of
epoxidation of naphthalene in the lung is problematic because of the instability of
the epoxide. However, short incubations of racemic naphthalene epoxide with
dissected airways of both the rat and mouse and in proximal vs. distal airways
Figure 1. Overview of naphthalene metabolism showing the formation of multiple, reactive
metabolites.
BUCKPITT ET AL.802
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

showed very little difference in the rates of formation of glutathione conjugates or
in the diastereomers produced. Likewise, there were no discernable differences in
the rates of dihydrodiol production between rat and mouse airways that appeared
to relate to the species differences in response to naphthalene.[27] Thus, it appeared
that the primary difference between species might have less to do with the
stereochemistry of epoxide formation than to the rates at which this intermediate
was generated.
Considerable progress has been made in the past 10 years in understanding
the importance of specific pulmonary cytochrome P450 monooxygenase isoforms
in the metabolic activation of lung toxicants. The contribution of each of these
proteins to this process is dependent not only on the amounts of protein present but
also on the catalytic activities of each of the proteins. Unfortunately, quantifying
the amounts of each of the cytochrome P450 isoforms present in various
subcompartments of the lung is difficult and in only a few cases has purified
Figure 2. Stereochemistry of naphthalene epoxide formation and elimination of thioethers in
mouse urine.
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 803
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

Table 2. Species Comparisons in the Rates of Conversion of Naphthalene to Naphthalene Oxides: Pulmonary and Nasal Tissue
Lung Microsomes Species
Rate of Metabolism (nmol/-
min/mg Protein)
Stereoselectivity Ratio of
(1R,2S ) to (1S,2R )
Vmaxa
(nmol/mg/min)
Kma
(mM )
Diol as a %
Total References
Mouse 13.8b 10 (at saturating substrate
concentrations); 30 at low
concentrations
14 40 8 53
Rat 1.69b 1.3 or less 0.88 10 5 53
Hamster 5.12b 0.5–0.9 1.45 20 25 53
Rhesus
macaque
0.15 0.12 ND ND 21 53
Human 0.12 0.85 ND ND n/ac 86
Nasal post mitochondrial
supernatant (olfactory)
Mouse 87 12.7 ND ND 7.4 53
Rat 43.5 RELSP , 36 ND ND 4.1 53
Hamster 3.9 ND ND ND 7.8 53
ND—not determined.a Apparent Michaelis constants were calculated by using the total rate of formation of diol plus glutathione conjugates.b Incubations were conducted at the same time for comparative purposes and contained microsomes, 0.5 mM naphthalene, 5 mM glutathione, glutathione S-
transferase, and NADPH regenerating system.c The amount of diol generated cannot be calculated because cyclohexene oxide was added to the incubations as an inhibitor of epoxide hydrolase.
BU
CK
PIT
TE
TA
L.
80
4
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

protein been available as standard.[55] More information is available on the
catalytic properties of some of the P450 monooxygenases through the use of
recombinant proteins. Since most toxicant exposure is unintended, toxicants (as
compared to most drug entities) are present in very small quantities. We argue that
a P450 monooxygenase with a Km for a protoxicant in the mM range is unlikely to
contribute to the metabolism of these substrates in vivo, and therefore,
understanding the catalytic efficiencies of these proteins with different substrates
is an important part of understanding the potential for these toxicants to result in
cellular injury. Accordingly, in the following sections we provide a description of
what is known about the catalytic activities of the proteins thought to be important
in the formation of naphthalene epoxides with particular emphasis on the
differences in activity among orthologues across species (Table 3).
Enzymology of Naphthalene Epoxide Formation
CYP2F1 (Human)
Nagata and his coworkers[56] purified a cytochrome P450 monooxygenase
from mouse liver that metabolized naphthalene rapidly and with high
stereoselectivity. Antibodies raised to this protein were used to clone and
sequence the cDNA coding for a 50-kDa protein that was present at high levels in
mouse lung and liver and which had 82% sequence homology[57] to a cDNA that
had been cloned earlier from human lung. The human (CYP2F1) and mouse
(CYP2F2) proteins were expressed in HEPG2 cells and yeast, respectively.
Recombinant CYP2F1 metabolized a number of substrates including ethoxycou-
marin and propoxycoumarin thus indicating that the expressed protein was
catalytically active.[58] Other studies in which CYP2F1 was expressed in
lymphoblastoid cells[59] demonstrated a naphthalene turnover (,0.035 nmol
conjugate/min/nmol P450) that was less than 0.1% the rate of metabolism
observed with the mouse orthologue.[60] The recombinant human CYP2F1 showed
slight stereopreference in the generation of (1S,2R )-naphthalene epoxide. In
contrast, 3-methylindole, another substrate requiring metabolic activation for
pulmonary toxicity, was metabolized by CYP2F1 with relatively high catalytic
efficiency (Vmax 1.3 nmol/nmol P450/min; Km 60mM ) indicating that the protein
is fully functional. Since the amounts of CYP2F1 are limited, kinetic studies have
not been conducted with naphthalene.
CYP2F2 (Mouse)
Naphthalene is metabolized with a high degree of stereoselectivity by
recombinant mouse CYP2F2 expressed in either yeast[57] or in SF-21 insect
cells.[60] A very high Vmax (107 nmol product/nmol P450/min) and low Km (3mM )
for the metabolism of naphthalene by recombinant CYP2F2 is consistent with the
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 805
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.
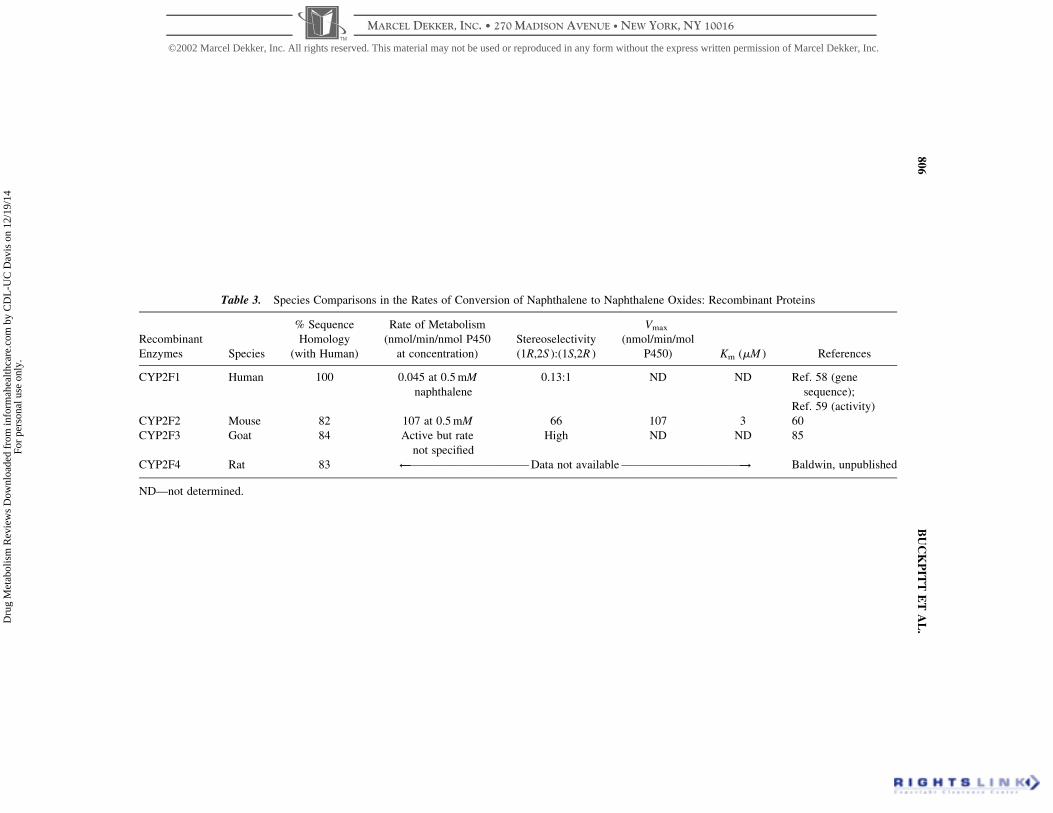
Table 3. Species Comparisons in the Rates of Conversion of Naphthalene to Naphthalene Oxides: Recombinant Proteins
Recombinant
Enzymes Species
% Sequence
Homology
(with Human)
Rate of Metabolism
(nmol/min/nmol P450
at concentration)
Stereoselectivity
(1R,2S ):(1S,2R )
Vmax
(nmol/min/mol
P450) Km (mM ) References
CYP2F1 Human 100 0.045 at 0.5 mM
naphthalene
0.13:1 ND ND Ref. 58 (gene
sequence);
Ref. 59 (activity)
CYP2F2 Mouse 82 107 at 0.5 mM 66 107 3 60
CYP2F3 Goat 84 Active but rate
not specified
High ND ND 85
CYP2F4 Rat 83 ˆ—————————— Data not available ——————————! Baldwin, unpublished
ND—not determined.
BU
CK
PIT
TE
TA
L.
80
6
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

importance of this protein in the metabolic activation and toxicity of naphthalene
in mouse lung. Previous work from our laboratories demonstrated a lung:blood
partition coefficient of 2.25 (unpublished observations). The levels of naphthalene
in the blood after a 7-hr inhalation exposure at 10 ppm in mice are 0.59 and
0.27mg/mL in male and female mice, respectively.[35] A 3mM Km is equivalent to
0.38mg/mL, and this suggests that the levels of naphthalene in target tissues easily
exceed the Km of CYP2F2 (mouse). Although the recently published
physiologically based pharmacokinetic (PBPK) model suggested that the lung
levels of naphthalene would be slightly lower (0.1mg/mL), these levels are still
within a range where CYP2F2 would be catalytically efficient. Similarly, in a
limited unpublished study (Cho and Buckpitt), unchanged naphthalene, measured
in whole lungs of mice treated with naphthalene (200 mg/kg) by gavage was 8.2
and 3.9mg/g at 30 and 60 min after administration, respectively. These
concentrations far exceed the Km of CYP2F2.
CYP2F4 (Rat)
Earlier work[57] failed to demonstrate the presence of transcript for CYP2F in
rat lung. In addition, while immunocytochemistry with antibodies generated to the
mouse 2F showed high staining intensities for this protein in mouse lung airway
epithelial cells, no detectable staining was observed in either the rat or primate.[27]
This suggested that the differences observed in susceptibility between these two
species might simply be related to the presence of a P450 isoform with high
catalytic activities toward naphthalene and similar substrates[61] in sensitive
species that are either not present or are present at low concentrations in resistant
species. Later studies demonstrated significant levels of CYP2F transcript by
Northern blot analysis, and a CYP has been cloned and sequenced from the rat
(Baldwin, NM_019303). The rat sequence has 93% identity with the mouse and
83% with the human (Baldwin, unpublished). Characterization of the catalytic
activities of the expressed protein and determination of the quantitative
distribution of this protein in mouse vs. rat lungs may indicate whether the
mouse/rat differences in susceptibility are related to lower protein levels or to
altered catalytic activities of the protein.
Clearly, further characterization of the catalytic activities of human CYP2F1
and possibly nonhuman primate orthologues will be essential to understand the
potential importance of these proteins in the metabolic activation of low molecular
weight aromatic hydrocarbons. In our view, it is important to note that the cellular
concentration of each of these P450 proteins will be a key factor in determining the
outcome of exposure. High concentrations of protein in a few cells will likely have
far greater chances of resulting in deleterious effects in response to exposure than
low concentrations of protein spread over many cells. Thus, any assessment of the
importance of these isoforms will have to take into account the localization of the
protein.
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 807
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

Overall, these studies indicate that there may be major catalytic differences in
CYP2F proteins in human vs. mouse lungs. These differences, if well established
experimentally, could have considerable influence on our assessments of the
likelihood that naphthalene produces lung injury in the human. What has not been
determined at this stage is whether CYP2F is responsible for catalyzing the metabolic
activation of naphthalene in the nasal epithelium. The data in Table 3 shows that the
apparent turnover of naphthalene in postmitochondrial supernatants prepared from
olfactory nasal mucosa are much higher in the mouse and rat than they are in the
hamster. This corresponds with the differences noted in susceptibility of the olfactory
epithelium in these 3 rodent species to naphthalene toxicity. There are several
cytochrome P450 isoforms present in the nasal mucosa including CYP1A2, 2A, 2B,
2C, 2G1, 2J, and 3A,[62,63] but to date, the catalytic activities with naphthalene have
not been established. The critical issues that need to be addressed in order to better
understand the potential for naphthalene to cause nasal epithelial cell toxicity in the
human are the same as those for the lung. The catalytic efficiencies of each of the
P450s in naphthalene metabolism along with an assessment of the amounts of P450
protein present in nasal mucosa will need to be determined.
Role of Naphthalene Oxide in the Toxicity of Naphthalene and as anIntermediate Which Binds Covalently to Cellular Proteins
Work conducted in both the isolated perfused mouse lung and in isolated
murine Clara cells indicates that of the naphthalene metabolites tested,
naphthalene oxide produces Clara cell necrosis at the lowest concentrations of
any of the metabolites. Perfusion of isolated murine lungs with 0.25–1mmol
naphthalene oxide over a period of 1 hr produced a dose-dependent decrease in the
mass of airway epithelial cells (volume/surface area), a decrease in the mass of
nonciliated cells, and an increase in the mass of vacuolated cells.[47] The
magnitude of changes in airway epithelial cell morphometry elicited by perfusion
of the lung with 1mmol racemic naphthalene oxide were similar to those observed
after perfusion with 10mmol naphthalene. In comparison, 10mmol 1,2-
naphthoquinone resulted in only slight increases in vacuolated epithelial cell
mass and in no significant changes in either total cell mass or in nonciliated cell
mass. No alterations in bronchiolar epithelial cell morphology were observed in
lungs perfused with 6.6mmol 1-naphthol or 10mmol 1,4-naphthoquinone over a
period of 1 hr. While these data are consistent with the view that naphthalene oxide
mediates the Clara cell toxicity of naphthalene, contributions to this toxicity from
the quinones cannot be ruled out. The further conversion of naphthalene oxide to
other metabolites was not measured directly, and it is possible that some of the
epoxide is converted to quinone. Pulmonary glutathione was depleted by perfusion
with naphthalene oxide indicating that the epoxide was capable of reaching
intracellular compartments. However, the loss of glutathione only accounted for
approximately half of the naphthalene oxide entering the lung. The quinones are
BUCKPITT ET AL.808
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

chemically unstable, and it is not clear how much of these metabolites actually
reached the epithelial cells of the lung. It is also possible that the toxicity of
naphthalene is mediated by both naphthalene oxide and naphthoquinones.
Naphthalene oxide depletes glutathione thus enhancing the susceptibility of target
cells to small amounts of naphthoquinone which either bind covalently to critical
macromolecules or which redox cycle leading to the generation of reactive oxygen
species eventually leading to cell necrosis. Studies that are currently underway are
designed to identify those metabolites that are bound covalently to proteins.
In other studies using isolated murine Clara cells, Chichester and her
coworkers[64] demonstrated not only that naphthalene resulted in concentration-
and time-dependent losses in cell viability but that naphthalene oxide was the most
potent of the toxicants added to the incubation. Nearly complete loss of Clara cell
viability was noted after 4-hr incubations where naphthalene oxide (0.5 mM final
concentration) was added slowly over a period of 60 min. Addition of 1,4-
naphthoquinone (0.5 mM ) to isolated Clara cells resulted in similar losses of
viability while the 1,2-quinone, 1-naphthol, and naphthalene dihydrodiol were
considerably less cytotoxic. Again, these data must be interpreted cautiously
because there may be dramatic differences in the amounts of each of these
metabolites that reach the intracellular compartment of the Clara cell.
Data showing that the cytochrome P450 monooxygenase inhibitor, piperonyl
butoxide, blocks the loss of cell viability caused by naphthalene but not
naphthalene oxide, further supported the view that naphthalene oxide plays a
major role in naphthalene-induced Clara cell necrosis. Again, these studies cannot
rule out the possibility that the cytotoxicity of naphthalene represents the
combined actions of naphthalene oxide thus making the cell considerably more
vulnerable to 1,2-naphthoquinone that could be generated from the dihydrodiol
through dihydrodiol dehydrogenase.
Finally, additional data that support the importance of the epoxide in
mediating both the toxicity and the protein binding of reactive metabolites comes
from work in isolated murine hepatocytes.[54] Incubation of isolated hepatocytes
with (1S,2R )-naphthalene oxide resulted in nearly complete loss of cell viability
whereas incubation with the (1R,2S )-epoxide enantiomer caused no loss in trypan
blue dye exclusion. These differences have been attributed to the fact that the
(1R,2S )-epoxide is converted much more rapidly to the dihydrodiol than the
(1S,2R )-enantiomer presumably resulting in higher intracellular concentrations of
(1S,2R )-epoxide. Addition of radiolabeled naphthalene oxide to isolated
hepatocytes resulted in time-dependent covalent binding of radiolabel to proteins
which increased rapidly from 1 to 5 min and slowed considerably thereafter. The
fact that radioactivity from [3H]-naphthalene oxide rapidly bound to proteins with
no apparent lag period and that the rate of increase dropped dramatically after
5 min (at a time when most of the naphthalene oxide would have either been
biotransformed or would have undergone spontaneous hydration) argues against
the involvement of secondary metabolites, arising from either further metabolism
of 1-naphthol or the dihydrodiol, in this process.
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 809
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

1-Naphthol: A Possible Precursor for Reactive and Cytotoxic
Metabolites
A number of studies suggested that phenols that are products generated
spontaneously from the precursor aromatic hydrocarbon epoxides, could undergo
further metabolism to reactive, cytotoxic metabolites. For example, Hesse and
Metzger[65] demonstrated not only that 1-naphthol was efficiently metabolized by
rat liver microsomes to reactive metabolites but that 1-naphthol was likely a key
intermediate in the formation of covalently bound metabolites from naphthalene.
The evidence for this included the fact that inhibition of epoxide hydrolase did not
increase reactive metabolite binding in microsomal incubations with naphthalene,
addition of UDP glucuronic acid decreased binding, and addition of unlabeled 1-
naphthol to microsomal incubations containing [14C]-naphthalene decreased the
overall amounts of radiolabel covalently bound to microsomal proteins. Further
work in hepatocyte incubations confirmed these data showing that inhibitors of
either sulfation or glucuronidation markedly increased reactive metabolite binding
to proteins.[66] Later studies, which compared the rates of metabolism of
naphthalene to 1-naphthol and to covalently bound metabolites in murine lung vs.
liver microsomal incubations, demonstrated that the rates of 1-naphthol formation
were much higher in lung than in liver but that the rates of generation of reactive
metabolites in microsomal incubations from these two tissues were nearly
identical.[67] Addition of an epoxide hydrolase inhibitor to co-incubations of
human liver microsomes with peripheral mononuclear leukocytes increased both
the cytotoxicity and formation of covalent protein adducts.[10] In vivo, reactive
metabolites from [14C]-1-naphthol become bound covalently to proteins in lung,
liver, and kidney, but there was virtually no difference in the amounts of reactive
metabolite bound after administration of 1-naphthol compared to naphthalene.[67]
Since only a portion of the administered naphthalene would be expected to be
converted to 1-naphthol, we felt that this argued against a role of this metabolite in
covalent binding to proteins in vivo. Additional studies by O’Brien and her
coworkers[21] and Buckpitt and his colleagues[67] showed that while 1-naphthol
has a lower LD50 than naphthalene, it did not result in injury in any of the tissues
studied.
More recent work has again raised the issue of the importance of secondary
metabolites in naphthalene toxicity. Human peripheral mononuclear cells were
considerably less sensitive to the cytotoxic effects of naphthalene oxide than to 1-
naphthol (in the presence of human liver microsomes) and both 1,2- and 1,4-
naphthoquinone.[68] Finally, alkaline permethylation techniques developed for
examining the nature of reactive bromobenzene metabolites were applied to
studies in which murine Clara cells were incubated with naphthalene.[69] The
ratios of adducts generated from the 1,2-quinone vs. the 1,2-epoxide were 32:1,
and there was no evidence for the formation of 1,4-naphthoquinone. The results of
these studies are not in agreement with earlier work published by Doherty and her
coworkers.[70] These investigators showed that reactive metabolites generated in
BUCKPITT ET AL.810
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

microsomal incubations from 1-naphthol were not trapped with ethylene diamine,
a compound that reacts rapidly with 1,2-naphthoquinone. Based on these results,
1,4-naphthoquinone was the likely precursor to reactive metabolites generated
from 1-naphthol.
1,2-Dihydroxy-1,2-dihydronaphthalene (Naphthalene Dihydrodiol)
Although the dihydrodiols generated from aromatic hydrocarbon epoxides
are generally less reactive than the parent epoxide, these metabolites are the
precursors to the diol epoxide metabolites thought to be the penultimate
carcinogens from numerous PAH. Strong support for the in vivo formation of the
diol epoxide and/or diepoxide has been obtained from identification of urinary
naphthalene metabolites. A number of trihydroxytetrahydromethylthio deriva-
tives[48] and a trihydroxytetrahydromercapturate[71] have been recovered from the
urine of mice and rats treated with naphthalene. To our knowledge, there is no
quantitative information available on the rates of formation of these metabolites in
target tissues of susceptible and nonsusceptible species; thus, their role in the
toxicity of naphthalene is not known.
In addition to the reactions described earlier, naphthalene dihydrodiol can
undergo further biotransformation by cytosolic dihydrodiol dehydrogenase to
generate the catechol (1,2-dihydroxynaphthalene) that is easily oxidized to 1,2-
naphthoquinone, a compound that is both reactive and will undergo redox
cycling.[72,73] As discussed earlier, work in isolated Clara cells indicated that the
1,2-quinone was the major species bound to proteins covalently.[69] Antibodies to
the 1,2-naphthoquinone adducts have been utilized in a Western blot assay to show
that there are numerous protein adducts generated from the quinone in mouse liver
homogenate incubations.[74] 1,2-Naphthoquinone is mutagenic in the Ames
assay[73] and has been shown to form N-7 adducts with deoxyguanosine in
vitro.[75] These interactions could lead to depurination although this was not
demonstrated with 1,2-naphthoquinone. In addition, these adducts have not been
demonstrated in cell-based systems or in vivo. It is important to note that if 1,2-
naphthoquinone is shown to be an important intermediate in naphthalene toxicity,
the human could actually be more susceptible than some of the rodent models that
have been studied. This is based on the fact that both epoxide hydrolase[76] and
dihydrodiol dehydrogenase[72] have been shown to be highly active in human
tissues in comparison to rodents.
BIOMARKERS OF EXPOSURE AND EFFECT
As stated earlier, one of the approaches that can be used to provide further
information regarding rodent-to-human comparisons in terms of their response to
a chemical can be to develop biomarkers that not only indicate an exposure has
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 811
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

occurred but also that the exposure has occurred at a level sufficient to cause
toxicity. Although biomarkers that are capable of providing this information for
naphthalene have not been developed and validated in animal models, there are
two urinary metabolites, which have been used to demonstrate that an exposure
has occurred and that at least the initial step in the metabolism of naphthalene has
taken place. These biomarkers include the measurement of mercapturic acid
metabolites and conjugates of naphthol in the urine.
Mercapturic acids were identified as urinary metabolites of naphthalene
more than 40 years ago.[77] Although the methodology used to examine the
amounts of mercapturate in urine of a number of rodent species and man was
semiquantitative (staining intensities of spots isolated by paper chromatog-
raphy), Boyland and Sims[77] presented convincing evidence that all species
treated with naphthalene including rodents and humans were capable of
generating and eliminating these metabolites in the urine. Rodent species were
treated with doses that varied from 200 to nearly 1000 mg/kg, ip while human
volunteers took doses of 500 mg orally (,10 mg/kg). Subsequent work, by
Summer, Rozman and their colleagues,[78,79] showing no detectable increase in
urinary thioethers in either the chimpanzee or Rhesus macaque after
administration of naphthalene at doses as high as 200 mg/kg are at variance
with the findings in man. However, these studies involved the alkaline
hydrolysis of the urine containing mercapturate followed by measurement of
the free thiols using Ellman’s reagent, and it is not clear whether these assays
yield reliable data since standards were not available.
More recent work has investigated the elimination of diastereomeric
mercapturates as a means of measuring the rates and formation of naphthalene
epoxides in mice compared to rats after administration of naphthalene by both
parenteral and inhalation routes.[71] These studies showed that there were no
significant differences observed in the percentage of the dose eliminated as
mercapturate in urine between mice (25–34%) and rats (24–35%) and that the
amount of mercapturate eliminated as a percentage of dose did not vary after ip
administration. The amounts of urinary mercapturate eliminated following 4-hr
inhalation exposures were substantially greater in mice than rats on a per gram
body weight basis, and this is consistent with the higher rates of epoxide formation
in lung microsomes and in dissected airways from mice compared to rats.[27,52]
These studies also explored the possibility that measurement of diastereomeric
mercapturates could be used to assess the stereochemistry of epoxidation in vivo.
Earlier studies had demonstrated that relatively consistent amounts of the 2-
glutathionyl adducts derived from both the (1R,2S )- and the (1S,2R )-naphthalene
epoxides were eliminated in the urine as the corresponding mercapturic acids after
intravenous administration of diastereomeric glutathione conjugates.[80] However,
the adduct formed by glutathione conjugation at C-1 underwent significant
metabolism to the pyruvic acid derivative. Nevertheless, measurement of the
ratios of mercapturates derived from thioether formation at C-2 would provide a
means for measuring the stereochemistry of naphthalene oxide metabolism in
BUCKPITT ET AL.812
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

vivo. After parenteral administration of naphthalene to mice, the ratio of
diastereomeric urinary mercapturates derived from the (1R,2S )- to (1S,2R )-
epoxide was 1:1 at low doses (1–3 mg/kg), increased to 3:1 at intermediate doses
(50 mg/kg), and decreased to 2:1 at high doses (100/200 mg/kg). In rats, these
ratios remained less than 1:1 at all doses.[71] After inhalation exposures, ratios of
mercapturates derived from the (1R,2S )-epoxide were 5–6:1 at low concen-
trations (less than 15 ppm) and decreased to 3:1 at higher concentrations (15–
100 ppm) in mice, while in rats, the ratios were 1:1 or less for all concentrations.
Again these data are consistent with the ratios of naphthalene epoxide enantiomers
generated in vitro.
Measurement of conjugates of 1-naphthol in the urine has been shown to be
well correlated with naphthalene exposures in workplace settings (naphthalene
and tar distillation, creosote impregnation) even at low exposure concen-
trations.[81,82] Unfortunately, neither of these studies controlled for smoking, and it
is clear that this is a major contributor to 1-naphthol levels in the urine. Recent
work showed that urinary 1-naphthol levels were three fold higher in smokers than
in nonsmokers.[83] The other consideration necessary in monitoring urinary 1-
naphthol is that this is a key metabolite of the insecticide carbaryl.[84]
Although the presence of either the mercapturic acid or 1-naphthol in the
urine correlate with exposure to naphthalene and the presence of either metabolite
is indicative that P450-dependent metabolism of naphthalene has occurred, neither
provides much information with respect to events that are closely tied to toxicity.
It is possible that once the importance of various protein adducts in events that lead
to cellular injury is well understood, and the pathways by which these adducts are
eliminated from the organism are delineated that biomarkers based on adducts
with peptides derived from the target proteins would be excellent markers of both
exposure and effect.
FUTURE DIRECTIONS
Key issues remain to be resolved in assessing the potential for
naphthalene to result in toxicity or oncogenicity in humans. As discussed in
this review, there are substantial differences in the metabolism of naphthalene
among rodent species, and these variations appear to be tied to the extent and
severity of toxicity. It is clear from the work conducted to date that the rates of
metabolism of naphthalene in the lung are substantially different in rodents,
humans, and nonhuman primates. Preliminary studies appear to indicate that
differences in the catalytic activities of the proteins involved in naphthalene
metabolism as well as the amounts of protein present might account for these
differences. Although these data suggest that the respiratory tract of the human
would be far less susceptible to naphthalene than the respiratory tissue of
rodents, careful studies are needed to define the localization of the primary
metabolic enzymes involved in both the activation and detoxication of
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 813
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

naphthalene. As pointed out in this review, measurement of rates of
metabolism in homogenate of a very heterogenous tissue such as the lung can
yield data that are not informative with respect to reactions taking place in a
very small proportion of target cells. Thus, additional efforts are underway to
localize some of the proteins in the respiratory tract and to study metabolism
in target cell populations of both humans and nonhuman primates. Since the
NTP bioassays have identified the nasal epithelium as an important target for
naphthalene, additional work is needed to examine rates of metabolism of
naphthalene in this portion of the respiratory tract.
Although many of the areas of the respiratory tract targeted acutely by
naphthalene are the same as those where tumors develop, there are some
important differences, and the two responses, namely cytotoxicity and
carcinogenicity, might be totally separate. Additional work examining the
importance of specific protein adducts in cytotoxicity and potentially
carcinogenicity (alteration of DNA repair enzymes for example) could be
very helpful in development of biomarkers that would clearly test the possibility
that humans are susceptible to naphthalene and/or close structural analogs.
Identification of the precursor reactive metabolite responsible for the covalent
binding would help focus the research on those enzyme pathways responsible
for generating that metabolite as well as those that prevent the formation or
detoxify it once it is formed.
More work is needed to identify which of the cytochrome P450
monooxygenases are responsible for the initial step in naphthalene metabolism
in the nasal epithelium, and work comparing the catalytic efficiencies and
measurement of amounts of CYP2F in rodent and primate nasal epithelium needs
to be completed. Finally, the development of in vitro approaches that can be used
to determine the susceptibility of target and nontarget regions of rodent respiratory
tract along with extension to the respiratory system of nonhuman primates or
tissue from humans, would provide more definitive information regarding cross
species comparisons for cytotoxicity. The availability of these data should be
helpful not only in determining whether naphthalene exposures represent possible
health risks in the human but also, on a larger scale, should be useful in
determining whether the mouse, which is far more susceptible to a number of
respiratory tract toxicants than other rodents, is a good experimental model for
assessing the potential deleterious effects of these agents in the human.
ACKNOWLEDGMENTS
Work from the authors laboratories has been supported by funding from the National
Institutes of Environmental Health Sciences (ES 04311, ES 04699, ES 08408, ES 09681).
Support for core facilities used in this work has come from the Center for Environmental
Health (ES 05707). We gratefully acknowledge the American Chemical Council’s
Naphthalene and Hydrocarbon Solvents Panels for financial support for the preparation of
this review.
BUCKPITT ET AL.814
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

REFERENCES
1. American Lung Association, Estimated Prevalence of Lung Disease by Lung
Association Report, http://www.lungusa.org/ Nov 2001.
2. Agency for Toxic Substance and Disease Registry (ATSDR), Sciences International
Inc., Toxicological Profile for Naphthalene, 1-Methylnaphthalene and 2-
Methylnaphthalene; U.S. Department of Health and Human Services: Washington,
DC, 1995; 0–196.
3. Environ Corporation, Potential Health Risks from Former Manufactured-Gas Plant
Sites: Risk Assessment Methodology, Report Prepared for Pacific Gas and Electric,
Southern California Edison and Southern California Gas Co., May 1989.
4. Environmental Research and Technology Inc., Handbook on Manufactured Gas
Plant Sites, Report Prepared for Utility Solid Waste Activities Group, Superfund
committee, Pittsburgh, PA, Sept 1984.
5. Howard, P.H. Handbook of Environmental Fate and Exposure Data for Organic
Chemicals; Lewis Publishers, 1989; Vol. 1, 408–421.
6. Schmeltz, I.; Tosk, J.; Hoffman, D. Formation and Determination of Naphthalenes in
Cigarette Smoke. Anal. Chem. 1976, 48 (0), 645–650.
7. Witschi, H.; Espiritu, I.; Maronpot, R.R.; Pinkerton, K.E.; Jones, A.D. The
Carcinogenic Potential of the Gas Phase of Environmental Tobacco Smoke.
Carcinogenesis 1997, 18, 2035–2042.
8. Stanley, J. Broad Scan Analysis of the FY 82 National Human Adipose Tissue Survey
Specimens, Executive Summary; EPA, Office of Toxic Substances: Washington,
DC, 1986; Vol. 1.
9. Pellizzari, E.; Hartwell, T.; Harris, B.; Waddell, R.; Whi, D.; Erickson, M. Purgeable
Organic Compounds in Mother’s Milk. Bull. Environ. Contam. Toxicol. 1986, 28,
322–328.
10. Tingle, M.D.; Pirmohamed, M.; Templeton, E.; Wilson, A.S.; Madden, S.;
Kitteringham, N.R.; Park, B.K. An Investigation of the Formation of Cytotoxic,
Genotoxic, Protein-Reactive and Stable Metabolites from Naphthalene by Human
Liver Microsomes. Biochem. Pharmacol. 1993, 46, 1529–1538.
11. Chieco, P.; Moslen, M.; Reynolds, E. Effect of Administrative Vehicle on Oral 1, 1-
Dichloroethylene Toxicity. Toxicol. Appl. Pharmacol. 1981, 57 (0), 146–155.
12. Forkert, P.G.; Reynolds, E.S. 1,1-Dichloroethylene-Induced Pulmonary Injury. Exp.
Lung Res. 1982, 3, 57–68.
13. Green, T. Methylene Chloride Induced Mouse Liver and Lung Tumors: An
Overview of the Role of Mechanistic Studies in Human Safety Assessment. Hum.
Exp. Toxicol. 1997, 16, 3–13.
14. Liteplo, R.G.; Long, G.W.; Meek, M.E. Relevance of Carcinogenicity Bioassays in
Mice in Assessing Potential Health Risks Associated with Exposure to Methylene
Chloride. Hum. Exp. Toxicol. 1998, 17, 84–87.
15. Griffin, K.; Johnson, C.; Breger, R.; Franklin, R. Pulmonary Toxicity, Hepatic, and
Extrahepatic Metabolism of 2- Methylnaphthalene in Mice. Toxicol. Appl.
Pharmacol. 1981, 61, 185–196.
16. Sherratt, P.J.; Pulford, D.J.; Harrison, D.J.; Green, T.; Hayes, J.D. Evidence that
Human Class Theta Glutathione S-Transferase T1-1 Can Catalyse the Activation of
Dichloromethane, a Liver and Lung Carcinogen in the Mouse. Comparison of the
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 815
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

Tissue Distribution of GST T1-1 with that of Classes Alpha, Mu and Pi GST in
Human. Biochem. J. 1997, 326, 837–846.
17. Plopper, C.G.; Suverkropp, C.; Morin, D.; Nishio, S.; Buckpitt, A. Relationship of
Cytochrome P-450 Activity to Clara Cell Cytotoxicity. I. Histopathologic
Comparison of the Respiratory Tract of Mice, Rats and Hamsters After Parenteral
Administration of Naphthalene. J. Pharmacol. Exp. Ther. 1992, 261, 353–363.
18. Plopper, C.G. Pulmonary Bronchiolar Epithelial Cytotoxicity: Microanatomical
Considerations. In Metabolic Activation and Toxicity of Chemical Agents to Lung
Tissue and Cells; Gram, T.E., Ed.; Pergamon Press: New York, 1993; Chap. 1, 1–24.
19. Van Winkle, L.; Johnson, Z.; Nishio, S.; Brown, C.; Plopper, C. Early Events in
Naphthalene-Induced Acute Clara Cell Toxicity: Comparison of Membrane
Permeability and Ultrastructure. Am. J. Respir. Cell Mol. Biol. 1999, 21, 44–53.
20. Mahvi, D.; Bank, H.; Harley, R. Morphology of a Naphthalene-Induced Bronchiolar
Lesion. Am. J. Pathol. 1977, 86, 559–572.
21. O’Brien, K.A.; Smith, L.L.; Cohen, G.M. Differences in Naphthalene-Induced
Toxicity in the Mouse and Rat. Chem. Biol. Interact. 1985, 55, 109–122.
22. Rasmussen, R.E.; Do, D.H.; Kim, T.S.; Dearden, L.C. Comparative Cytotoxicity of
Naphthalene and Its Monomethyl- and Mononitro-Derivatives in the Mouse Lung.
J. Appl. Toxicol. 1986, 6, 13–20.
23. Van Winkle, L.; Gunderson, A.; Shimizu, J.; Brown, C. Gender Differences in both
Naphthalene Metabolism and Naphthalene-Induced Acute Lung Injury. Am.
J. Physiol. Lung 2002, 282, L1182–L1134.
24. West, J.; Pakenham, G.; Morin, D.; Fleschner, C.; Buckpitt, A.; Plopper, C. Inhaled
Naphthalene Causes Dose Dependent Clara Cell Toxicity in Mice But Not in Rats.
Toxicol. Appl. Pharmacol. 2001, 173, 114–119.
25. American Conference of Governmental Industrial Hygenists (ACGIH), Threshold
Limit Values for Chemical Substances and Physical Agents and Biological Exposure
Indices; Cincinnati, Ohio, 1999.
26. Plopper, C.; Chang, A.; Pang, A.; Buckpitt, A. Use of Microdissected Airways to
Better Define Metabolism and Cytotoxicity in Murine Bronchiolar Epithelium. Exp.
Lung Res. 1991, 17, 181–196.
27. Buckpitt, A.; Chang, A.; Weir, A.; Van Winkle, L.; Duan, X.; Philpot, R.; Plopper,
C. Relationship of Cytochrome P450 Activity to Clara Cell Cytotoxicity. IV.
Metabolism of Naphthalene and Naphthalene Oxide in Microdissected Airways
from Mice, Rats, and Hamsters. Mol. Pharmacol. 1995, 47, 74–81.
28. O’Brien, K.; Suverkropp, K.; Kanekal, S.; Plopper, C.; Buckpitt, A. Tolerance to
Multiple Doses of the Pulmonary Toxicant, Naphthalene. Toxicol. Appl. Pharmacol.
1989, 99, 487–500.
29. Lakritz, J.; Chang, A.; Weir, A.; Nishio, S.; Hyde, D.; Philpot, R.; Buckpitt, A.;
Plopper, C. Cellular and Metabolic Basis of Clara Cell Tolerance to Multiple Doses
of Cytochrome P450-Activated Cytotoxicants. I: Bronchiolar Epithelial Reorgan-
ization and Expression of Cytochrome P450 Monooxygenases in Mice Exposed to
Multiple Doses of Naphthalene. J. Pharmacol. Exp. Ther. 1996, 278, 1408–1418.
30. West, J.; Buckpitt, A.; Plopper, C. Elevated Airway GSH Resynthesis Confers
Protection to Clara Cells from Naphthalene Injury in Mice Made Tolerant by
Repeated Exposures. J. Pharmacol. Exp. Ther. 2000, 294, 516–523.
BUCKPITT ET AL.816
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

31. Shopp, G.; White, K.; Holsapple, M.; Barnes, D.; Duke, S.; Anderson, A.; Condie,
L.; Hayes, J.; Borzelleca, J. Naphthalene Toxicity in CD-1 Mice: General
Toxicology and Immunotoxocology. Fundam. Appl. Toxicol. 1984, 4, 406–419.
32. National Toxicology Program (NTP), Toxicology and Carcinogenesis Studies of
Naphthalene (CAS no. 91-20-3) in B6C3F1 Mice (Inhalation Studies), Technical
Report Series No. 410, NIH Publication No. 92-3141; U.S. Department of Health and
Human Services, Public Health Service, National Institutes of Health: Research
Triangle Park, NC, 1992.
33. Abdo, K.; Eustis, S.; McDonald, M.; Joiken, M.; Adkins, B.; Haseman, J.
Naphthalene: A Respiratory Tract Toxicant and Carcinogen for Mice. Inhal.
Toxicol. 1992, 4, 393–409.
34. Abdo, K.M.; Grumbein, S.; Chou, B.J.; Herbert, R. Toxicity and Carcinogenicity
Study in f344 Rats Following 2 Years of Whole-Body Exposure to Naphthalene
Vapors. Inhal. Toxicol. 2001, 13 (10), 931–950.
35. National Toxicology Program (NTP), Toxicology and Carcinogenesis Studies of
Naphthalene (CAS no. 91-20-3) in F344/N Rats (Inhalation Studies), Technical
Report Series No. 500. NIH Publication No. 01-4434; U.S. Department of Health and
Human Services, Public Health Service, National Institutes of Health: Research
Triangle Park, NC, 2000.
36. Gram, T. Chemically Reactive Intermediates and Pulmonary Xenobiotic Toxicity.
Pharmacol. Rev. 1997, 49, 297–341.
37. Yost, G.S. Selected, Nontherapeutic Agents. In Toxicology of the Respiratory
System; Roth, R., Ed.; Pergamon Press: New York, 1997; Chap. 38, 591–610.
38. Buckpitt, A.; Cruikshank, M. Biochemical Function of the Lungs: Metabolism of
Xenobiotic Agents. In Toxicology of the Respiratory System; Roth, R., Ed.;
Pergamon Press: New York, 1997; Chap. 12, 159–186.
39. Hinson, J.; Pumford, N.R.; Nelson, S.D. The Role of Metabolic Activation in Drug
Toxicity. Drug Metab. Rev. 1994, 26, 395–412.
40. Boyd, M.R. Biochemical Mechanisms in Chemical Induced Lung Injury. CRC Crit.
Rev. Toxicol. 1980, 7, 103–176.
41. Roberts, S.A.; Price, V.F.; Jollow, D.J. Acetaminophen Structure–Toxicity Studies:
In Vivo Covalent Binding of a Nonhepatotoxic Analog, 3-Hydroxyacetanilide.
Toxicol. Appl. Pharmacol. 1990, 105, 195–208.
42. Warren, D.L.; Brown, D., Jr.; Buckpitt, A. Evidence for Cytochrome P450 Mediated
Metabolism in the Bronchiolar Damage by Naphthalene. Chem. Biol. Interact. 1982,
40, 287–303.
43. Buckpitt, A.R.; Warren, D.L. Evidence for Hepatic Formation, Export and Covalent
Binding of Reactive Naphthalene Metabolites in Extrahepatic Tissues In Vivo.
J. Pharmacol. Exp. Ther. 1983, 225, 8–16.
44. Cho, M.; Chichester, C.; Morin, D.; Plopper, C.; Buckpitt, A. Covalent Interactions
of Reactive Naphthalene Metabolites with Proteins. J. Pharmacol. Exp. Ther. 1994,
269, 881–889.
45. Jerina, D.; Daly, J.; Witkop, B.; Zaltzman-Nirenberg, P.; Udenfriend, S. 1,2-
Naphthalene Oxide as an Intermediate in the Microsomal Hydroxylation of
Naphthalene. Biochem. J. 1970, 9, 147–156.
46. Van Bladeren, P.; Vyas, K.; Sayer, J.; Ryan, D.; Thomas, P.; Levins, W.; Jerina, D.
Stereoselectivity of Cytochrome P-450c in the Formation of Naphthalene and
Anthracene 1,2-Oxides. J. Biol. Chem. 1984, 259, 8966–8973.
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 817
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

47. Kanekal, S.; Plopper, C.; Morin, D.; Buckpitt, A. Metabolism and Cytotoxicity of
Naphthalene Oxide in the Isolated Perfused Mouse Lung. J. Pharmacol. Exp. Ther.
1991, 256, 391–401.
48. Horning, M.; Stillwell, W.; Griffin, G.; Tsang, W. Epoxide Intermediates in the
Metabolism of Naphthalene by the Rat. Drug Metab. Dispos. 1980, 8, 404–414.
49. Thakker, D.; Yagi, H.; Levin, W.; Wood, A.; Conney, A.; Jerina, D. Polycyclic
Aromatic Hydrocarbons: Metabolic Activation to Ultimate Carcinogens. In
Bioactivation of Foreign Compounds; Anders, M., Ed.; Academic Press: New
York, 1985; Chap. 7, 178–242.
50. Van Bladeren, P.; Sayer, J.; Ryan, D.; Thomas, P.; Levin, W.; Jerina, D. Differential
Stereoselectivity of Cytochrome P450b and P450c in the Formation of Naphthalene
and Anthracene 1,2-Oxides. J. Biol. Chem. 1985, 260, 10226–10235.
51. Smart, G.; Buckpitt, A.R. Formation of Reactive Naphthalene Metabolites by Target
vs Non-target Microsomes: Methods for the Separation of Three Glutathione
Adducts. Biochem. Pharmacol. 1983, 32 (5), 943–946.
52. Buckpitt, A.R.; Castagnoli, N.; Nelson, S.D.; Jones, A.D.; Bahnson, L.S.
Stereoselectivity of Naphthalene Epoxidation by Mouse, Rat and Hamster
Pulmonary, Hepatic and Renal Microsomal Enzymes. Drug Metab. Dispos. 1987,
15, 491–498.
53. Buckpitt, A.; Buonarati, M.; Avey, L.B.; Chang, A.M.; Morin, D.; Plopper, C.G.
Relationship of Cytochrome P450 Activity to Clara Cell Cytotoxicity. II.
Comparison of Stereoselectivity of Naphthalene Epoxidation in Lung and Nasal
Mucosa of Mouse, Hamster, Rat and Rhesus Monkey. J. Pharmacol. Exp. Ther.
1992, 261, 364–372.
54. Buonarati, M.; Morin, D.; Plopper, C.; Buckpitt, A. Glutathione Depletion and
Cytotoxicity by Naphthalene 1,2-Oxide in Isolated Hepatocytes. Chem. Biol.
Interact. 1989, 71, 147–165.
55. Domin, B.; Devereux, T.; Philpot, R.M. The Cytochrome P450 Monooxygenase
System of Rabbit Lung: Enzyme Components, Activities and Induction in the
Nonciliated Bronchiolar Epithelial (Clara) Cell, Alveolar Type II Cell and Alveolar
Macrophage. Mol. Pharmacol. 1986, 30, 296–303.
56. Nagata, K.; Martin, B.M.; Gillette, J.R.; Sasame, H.A. Isozymes of Cytochrome P-
450 That Metabolize Naphthalene in Liver and Lung of Untreated Mice. Drug
Metab. Dispos. 1990, 18, 557–564.
57. Ritter, J.K.; Owens, I.S.; Negishi, M.; Nagata, K.; Sheen, Y.Y.; Gillette, J.R.;
Sasame, H.A. Mouse Pulmonary Cytochrome P-450 Naphthalene Hydroxylase:
cDNA Cloning, Sequence, and Expression in Saccharomyces cerevisiae.
Biochemistry 1991, 30, 11430–11437.
58. Nhamburo, P.T.; Kimura, S.; Mcbride, O.W.; Kozak, C.A.; Gelboin, H.V.;
Gonzalez, F.J. The Human CYP2F Gene Subfamily: Identification of a cDNA
Encoding a New Cytochrome P450, cDNA-Directed Expression, and Chromosome
Mapping. Biochemistry 1990, 29, 5491–5499.
59. Lanza, D.L.; Code, E.; Crespi, C.L.; Gonzalez, F.J.; Yost, G.S. Specific
Dehydrogenation of 3-Methylindole and Epoxidation of Naphthalene by
Recombinant Human CYP2F1 Expressed in Lymphoblastoid Cells. Drug Metab.
Dispos. 1999, 27, 798–803.
BUCKPITT ET AL.818
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

60. Shultz, M.A.; Choudary, P.V.; Buckpitt, A.R. Role of Murine Cytochrome P-450
2F2 in Metabolic Activation of Naphthalene and Metabolism of Other Xenobiotics.
J. Pharmacol. Exp. Ther. 1999, 290, 281–288.
61. Shultz, M.A.; Morin, D.; Chang, A.M.; Buckpitt, A. Metabolic Capabilities of
CYP2F2 with Various Pulmonary Toxicants and Its Relative Abundance in Mouse
Lung Subcompartments. J. Pharmacol. Exp. Ther. 2001, 296 (2), 510–519.
62. Gu, J.; Dudley, C.; Su, T.; Spink, D.C.; Zhang, Q.Y.; Moss, R.L.; Ding, X.
Cytochrome P450 and Steroid Hydroxylase Activity in Mouse Olfactory and
Vomeronasal Mucosa. Biochem. Biophys. Res. Commun. 1999, 266, 262–267.
63. Xie, Q.; Zhang, Q.; Zhang, Y.; Su, T.; Gu, J.; Kaminsky, L.; Ding, X. Induction of
Mouse CYP2J by Pyrazole in the Eye, Kidney, Liver, Lung, Olfactory Mucosa and
Small Intestine, But Not in Heart. Drug Metab. Dispos. 2000, 28, 1311–1326.
64. Chichester, C.H.; Buckpitt, A.R.; Chang, A.; Plopper, C.G. Metabolism and
Cytotoxicity of Naphthalene and Its Metabolites in Isolated Murine Clara Cells. Mol.
Pharmacol. 1994, 45, 664–672.
65. Hesse, S.; Mezger, M. Involvement of Phenolic Metabolites in the Irreversible
Protein- Binding of Aromatic Hydrocarbons: Reactive Metabolites of (14C)
Naphthalene and (14C)1-Naphthol Formed by Rat Liver Microsomes. Mol.
Pharmacol. 1979, 16, 667–675.
66. Schwarz, L.; Mezger, M.; Hesse, S. Effects of Decreased Glucuronidation and
Sulfation on Covalent Binding of Naphthalene in Isolated Rat Hepatocytes.
Toxicology 1980, 17, 119–120.
67. Buckpitt, A.R.; Bahnson, L.S.; Franklin, R.B. Evidence that 1-Naphthol Is Not an
Obligate Intermediate in the Covalent Binding and the Pulmonary Bronchiolar
Necrosis by Naphthalene. Biochem. Biophys. Res. Commun. 1986, 126 (3),
1097–1103.
68. Wilson, A.S.; Davis, C.D.; Williams, D.P.; Buckpitt, A.R.; Pirmohamed, M.; Park,
B.K. Characterization of the Toxic Metabolite(s) of Naphthalene. Toxicology 1996,
114, 233–242.
69. Zheng, J.; Cho, M.; Jones, A.D.; Hammock, B.D. Evidence of Quinone Metabolites
of Naphthalene Covalently Bound to Sulfur Nucleophiles of Proteins of Murine
Clara Cells After Exposure to Naphthalene. Chem. Res. Toxicol. 1997, 10,
1008–1014.
70. Doherty, M.A.; Makowski, R.; Gibson, G.G.; Cohen, G.M. Cytochrome P-450
Dependent Metabolic Activation of 1-Naphthol to Naphthoquinones and Covalent
Binding Species. Biochem. Pharmacol. 1985, 34 (13), 2261–2267.
71. Pakenham, G.; Morin, D.; Lango, J.; Buonarati, M.H.; Buckpitt, A.R. Biomarkers for
Metabolically Activated Cytotoxic Agents: Studies on Urinary Diastereomeric
Mercapturic Acids of Naphthalene. Drug Metab. Dispos. 2002, 30, 1–7.
72. Penning, T.; Burczynski, M.; Hung, C.; McCoull, K.; Palackal, N.; Tsuruda, L.S.
Dihydrodiol Dehydrogenases and Polycyclic Aromatic Hydrocarbon Activation:
Generation of Reactive and Redox Active o-Quinones. Chem. Res. Toxicol. 1999,
12, 1–18.
73. Bolton, J.L.; Trush, M.A.; Penning, T.M.; Dryhurst, G.; Monks, T.J. Role of
Quinones in Toxicology. Chem. Res. Toxicol. 2000, 13 (3), 135–160.
74. Zheng, J.; Hammock, B.D. Development of Polyclonal Antibodies for Detection of
Protein Modification by 1,2-Naphthoquinone. Chem. Res. Toxicol. 1996, 9,
904–909.
NAPHTHALENE-INDUCED RESPIRATORY TRACT TOXICITY 819
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.

75. McCoull, K.; Rindgen, D.; Blair, I.; Penning, T.M. Synthesis and Characterization of
Polycyclic Aromatic Hydrocarbon o-Quinone Depurinating N7-Guanine Adducts.
Chem. Res. Toxicol. 1999, 12, 237–246.
76. Kitteringham, N.R.; Davis, C.; Howard, N.; Pirmohamed, M.; Park, B.K.
Interindividual and Interspecies Variation in Hepatic Microsomal Epoxide
Hydrolase Activity: Studies with cis-Stilbene Oxide, Carbamazepine 10,11-Epoxide
and Naphthalene. J. Pharmacol. Exp. Ther. 1996, 278, 1018–1027.
77. Boyland, E.; Sims, P. Metabolism of Polycyclic Compounds 12. An Acid Labile
Precursor of 1-Naphthylmercapturic Acid and Naphthol: An N-Acetyl-S-(1,2-
Dihydrohydroxynaphthyl)-L-Cysteine. Biochem. J. 1958, 68, 440–447.
78. Summer, K.; Rozman, K.; Coulston, F.; Greim, H. Urinary Excretion of Mercapturic
Acids in Chimpanzees and Rats. Toxicol. Appl. Pharmacol. 1979, 50, 207–212.
79. Rozman, K.; Summer, K.; Rozman, T.; Greim, H. Elimination of Thioethers
Following Administration of Naphthalene and Diethylmaleate to the Rhesus
Monkey. Drug Chem. Toxicol. 1982, 5, 265–275.
80. Buonarati, M.; Jones, A.D.; Buckpitt, A. In Vivo Metabolism of Isomeric
Naphthalene Oxide Glutathione Conjugates. Drug Metab. Dispos. 1990, 18,
183–189.
81. Heikkila, P.R.; Luotamo, M.; Riihimaki, V. Urinary 1-Naphthol Excretion in the
Assessment of Exposure to Creosote in an Impregnation Facility. Scand. J. Work
Environ. Health 1997, 23, 199–205.
82. Bieniek, G. Urinary Naphthols as an Indicator of Exposure to Naphthalene. Scand.
J. Work Environ. Health 1997, 23, 414–420.
83. Yang, M.; Koga, M.; Katoh, T.; Kawamoto, T. A Study for the Proper Application of
Urinary Naphthols, New Biomarkers for Airborne Polycyclic Aromatic Hydro-
carbons. Arch. Environ. Contam. Toxicol. 1999, 36, 99–108.
84. Shealy, D.B.; Barr, J.R.; Ashley, D.L.; Patterson, D.G., Jr.; Camann, D.E.; Bond,
A.E. Correlation of Environmental Carbaryl Measurements with Serum and Urinary
1-Naphthol Measurements in a Farmer Applicator and His Family. Environ. Health
Perspect. 1997, 105, 510–513.
85. Wang, H.; Lanza, D.; Yost, G. Cloning and Expression of CYP2F3, a Cytochrome
P450 that Bioactivates the Selective Pneumotoxins 3-Methylindole and Naphtha-
lene. Arch. Biochem. Biophys. 1998, 349, 329–340.
86. Buckpitt, A.R.; Bahnson, L.S. Naphthalene Metabolism by Human Lung
Microsomal Enzymes. Toxicology 1986, 41, 333–341.
BUCKPITT ET AL.820
©2002 Marcel Dekker, Inc. All rights reserved. This material may not be used or reproduced in any form without the express written permission of Marcel Dekker, Inc.
MARCEL DEKKER, INC. • 270 MADISON AVENUE • NEW YORK, NY 10016
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C D
avis
on
12/1
9/14
For
pers
onal
use
onl
y.




















