
Downregulation of a putative tumor suppressor BMP4 by SOX2 promotes growth of lung squamous cell...
11
Downregulation of a putative tumor suppressor BMP4 by SOX2 promotes growth of lung squamous cell carcinoma Wen-Tsen Fang 1 , Chi-Chen Fan 2,3 , Shih-Miao Li 1,4,5 , Te-Hsuan Jang 6 , Hsiu-Ping Lin 1 , Neng-Yao Shih 1 , Chung-Hsing Chen 1 , Tao-Yeuen Wang 7,8,9 , Shiu-Feng Huang 6 , Alan Yueh-Luen Lee 1 , Ying-Lan Liu 1 , Fang-Yu Tsai 1 , Chih-Ting Huang 1 , Su Jing Yang 10 , Lin-Ju Yen 11 , Chih-Pin Chuu 11 , Chih-Yi Chen 12 , Chao A. Hsiung 5 , Jang-Yang Chang 1,13 , Lu-Hai Wang 6 , I-Shou Chang 1 and Shih Sheng Jiang 1 1 National Institute of Cancer Research, NHRI, Zhunan, Taiwan 2 Department of Physiology, Mackay Memorial Hospital, Taipei, Taiwan 3 Department of Medical Laboratory Science and Biotechnology, Yuanpei University, Hsinchu, Taiwan 4 Institute of Bioinformatics and Structural Biology, National Tsing Hua University, Hsinchu, Taiwan 5 Institute of Population Health Sciences, NHRI, Zhunan, Taiwan 6 Institute of Molecular and Genomic Medicine, NHRI, Zhunan, Taiwan 7 Department of Pathology, Mackay Memorial Hospital, Taipei, Taiwan 8 Mackay Medical College, Taipei, Taiwan 9 Mackay Medicine, Nursing and Management College, Taipei, Taiwan 10 Department of Medical Imaging and Radiological Science, Central Taiwan University of Science and Technology, Taichung, Taiwan 11 Institute of Cellular and System Medicine, NHRI, Zhunan, Taiwan 12 Cancer Center, China Medical University and Hospital, Taichung, Taiwan 13 Division of Hematology/Oncology, Department of Internal Medicine, National Cheng Kung University Hospital, College of Medicine, National Cheng Kung University, Tainan, Taiwan SOX2 is a transcription factor essential for self-renewal and pluripotency of embryonic stem cells. Recently, SOX2 was found overexpressed in the majority of the lung squamous cell carcinoma (SQC), in which it acts as a lineage-survival oncogene. However, downstream targets/pathways of SOX2 in lung SQC cells remain to be identified. Here, we show that BMP4 is a downstream target of SOX2 in lung SQC. We found that SOX2-silencing-mediated inhibition of cell growth was accompanied by upregulation of BMP4 mRNA and its protein expression. Meta-analysis with 293 samples and qRT-PCR validation with 73 clinical samples revealed an inversely correlated relationship between levels of SOX2 and BMP4 mRNA, and significantly lower mRNA levels in tumor than in adjacent normal tissues. This was corroborated by immunohistochemistry analysis of 35 lung SQC samples showing lower BMP4 protein expression in tumor tissues. Cell-based experiments including siRNA transfection, growth assay and flow cytometry assay, further combined with a xenograft tumor model in mice, revealed that reactivation of BMP4 signaling could partially account for growth inhibition and cell cycle arrest in lung SQC cells upon silencing SOX2. Finally, chromatin immunoprecipitation analysis and luciferase reporter assay revealed that SOX2 could negatively regulate BMP4 promoter activity, possibly through binding to the promoter located in the first intron region of BMP4. Collectively, our findings suggest that BMP4 could act as a tumor suppressor and its downregulation by elevated SOX2 resulting in enhanced growth of lung SQC cells. Key words: SOX2, BMP4, lung cancer, lung squamous cell carcinoma Additional Supporting Information may be found in the online version of this article. Conflict of interest: Nothing to report W.-T.F. and C.-C.F. contribute equally to this study Grant sponsor: National Science Council (National Core Facility Program for Biotechnology Grants); Grant number: NSC 102-2319-B-400- 001; Grant sponsor: Department of Health, Taiwan; Grant number: DOH102-TD-C-111-004; Grant sponsor: NHRI DOI: 10.1002/ijc.28734 History: Received 17 July 2013; Accepted 13 Jan 2014; Online 25 Jan 2014 Correspondence to: I-Shou Chang, National Institute of Cancer Research, National Health Research Institutes, 35 Keyan Road, Zhunan Town, Miaoli County 350, Taiwan, Tel.: 1886-37-246166, ext. 36130, Fax: 1886-37-586463, E-mail: [email protected] or Shih Sheng Jiang, National Institute of Cancer Research, National Health Research Institutes, 35 Keyan Road, Zhunan Town, Miaoli County 350, Taiwan, Tel.: 1886-37-246166, ext. 31708, Fax: 1886-37-586463, E-mail: [email protected] Cancer Cell Biology Int. J. Cancer: 00, 00–00 (2014) V C 2014 UICC International Journal of Cancer IJC
Transcript of Downregulation of a putative tumor suppressor BMP4 by SOX2 promotes growth of lung squamous cell...

Downregulation of a putative tumor suppressor BMP4 bySOX2 promotes growth of lung squamous cell carcinoma
Wen-Tsen Fang1, Chi-Chen Fan2,3, Shih-Miao Li1,4,5, Te-Hsuan Jang6, Hsiu-Ping Lin1, Neng-Yao Shih1, Chung-Hsing Chen1,
Tao-Yeuen Wang7,8,9, Shiu-Feng Huang6, Alan Yueh-Luen Lee1, Ying-Lan Liu1, Fang-Yu Tsai1, Chih-Ting Huang1,
Su Jing Yang10, Lin-Ju Yen11, Chih-Pin Chuu11, Chih-Yi Chen12, Chao A. Hsiung5, Jang-Yang Chang1,13,
Lu-Hai Wang6, I-Shou Chang1 and Shih Sheng Jiang1
1 National Institute of Cancer Research, NHRI, Zhunan, Taiwan2 Department of Physiology, Mackay Memorial Hospital, Taipei, Taiwan3 Department of Medical Laboratory Science and Biotechnology, Yuanpei University, Hsinchu, Taiwan4 Institute of Bioinformatics and Structural Biology, National Tsing Hua University, Hsinchu, Taiwan5 Institute of Population Health Sciences, NHRI, Zhunan, Taiwan6 Institute of Molecular and Genomic Medicine, NHRI, Zhunan, Taiwan7 Department of Pathology, Mackay Memorial Hospital, Taipei, Taiwan8 Mackay Medical College, Taipei, Taiwan9 Mackay Medicine, Nursing and Management College, Taipei, Taiwan10 Department of Medical Imaging and Radiological Science, Central Taiwan University of Science and Technology, Taichung, Taiwan11 Institute of Cellular and System Medicine, NHRI, Zhunan, Taiwan12 Cancer Center, China Medical University and Hospital, Taichung, Taiwan13 Division of Hematology/Oncology, Department of Internal Medicine, National Cheng Kung University Hospital, College of Medicine, National Cheng Kung
University, Tainan, Taiwan
SOX2 is a transcription factor essential for self-renewal and pluripotency of embryonic stem cells. Recently, SOX2 was found
overexpressed in the majority of the lung squamous cell carcinoma (SQC), in which it acts as a lineage-survival oncogene.
However, downstream targets/pathways of SOX2 in lung SQC cells remain to be identified. Here, we show that BMP4 is a
downstream target of SOX2 in lung SQC. We found that SOX2-silencing-mediated inhibition of cell growth was accompanied
by upregulation of BMP4 mRNA and its protein expression. Meta-analysis with 293 samples and qRT-PCR validation with 73
clinical samples revealed an inversely correlated relationship between levels of SOX2 and BMP4 mRNA, and significantly lower
mRNA levels in tumor than in adjacent normal tissues. This was corroborated by immunohistochemistry analysis of 35 lung
SQC samples showing lower BMP4 protein expression in tumor tissues. Cell-based experiments including siRNA transfection,
growth assay and flow cytometry assay, further combined with a xenograft tumor model in mice, revealed that reactivation of
BMP4 signaling could partially account for growth inhibition and cell cycle arrest in lung SQC cells upon silencing SOX2.
Finally, chromatin immunoprecipitation analysis and luciferase reporter assay revealed that SOX2 could negatively regulate
BMP4 promoter activity, possibly through binding to the promoter located in the first intron region of BMP4. Collectively, our
findings suggest that BMP4 could act as a tumor suppressor and its downregulation by elevated SOX2 resulting in enhanced
growth of lung SQC cells.
Key words: SOX2, BMP4, lung cancer, lung squamous cell carcinoma
Additional Supporting Information may be found in the online version of this article.
Conflict of interest: Nothing to report
W.-T.F. and C.-C.F. contribute equally to this study
Grant sponsor: National Science Council (National Core Facility Program for Biotechnology Grants); Grant number: NSC 102-2319-B-400-
001; Grant sponsor: Department of Health, Taiwan; Grant number: DOH102-TD-C-111-004; Grant sponsor: NHRI
DOI: 10.1002/ijc.28734
History: Received 17 July 2013; Accepted 13 Jan 2014; Online 25 Jan 2014
Correspondence to: I-Shou Chang, National Institute of Cancer Research, National Health Research Institutes, 35 Keyan Road, Zhunan
Town, Miaoli County 350, Taiwan, Tel.: 1886-37-246166, ext. 36130, Fax: 1886-37-586463, E-mail: [email protected] or Shih Sheng
Jiang, National Institute of Cancer Research, National Health Research Institutes, 35 Keyan Road, Zhunan Town, Miaoli County 350, Taiwan,
Tel.: 1886-37-246166, ext. 31708, Fax: 1886-37-586463, E-mail: [email protected]
Can
cerCellBiology
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC
International Journal of Cancer
IJC

Lung cancer is the leading cause of cancer-related death inmany countries in the world.1 Among all subtypes of lungcancer, non-small cell lung carcinoma (NSCLC) is the mostfrequent one, which accounts for greater than 80% of lungcancers.2 The prognosis of NSCLC patients is poor, with the5-year survival rate of only about 18%,2 and the high mortal-ity of NSCLC is thought to be attributed to difficulties inearly diagnosis and the lack of effective therapeutic methodsin general. NSCLC can be further classified into three histo-logical subtypes, i.e., squamous cell carcinoma (SQC), adeno-carcinoma (ADC) and large cell carcinoma (LCC). Amongthese subtypes, SQC and ADC are the major ones, whichtogether represent �70% of NSCLC.2 SQC was once themost common subtype of NSCLC during the past century.However, it was noted later that proportion of SQC relativeto ADC declined gradually,3,4 most probably owing tochange in smoking behavior and improved diagnostic meth-ods. Today, ADC is the most common subtype in manycountries, while SQC is estimated to account for 20–25% ofNSCLCs.2
Compared with ADC of the lung, in which many drivermutations have been identified and targeted therapies are appli-cable to a good proportion of patients especially in the Far East-ern countries, neoplasms of SQC of the lung were once regardedas tumors without readily targetable genetic abnormalities.5
However, recent studies have identified new lung SQC-associated genetic changes,6,7 including the amplification andoverexpression of SOX2. SOX2 is located on chromosome 3q26,a region amplified in about 20% of lung SQC.6,8 At a muchhigher rate than gene amplification, overexpression of SOX2mRNA was observed in about 90% of the lung SQC,8 indicatingthat SOX2 might play oncogenic role(s) in the tumorigenesis oflung SQC, which is supported by studies reporting that SOX2 isa lineage-survival oncogene of SQC of the lung.6,9
As SOX2 functions upstream of the hierarchy of geneexpression network, it is likely that its aberrant expression inlung epithelia could cause profound change in a wide varietyof molecular pathways, which may contribute to growth andsurvival of the lung SQC cells. Although the oncogenic roleof SOX2 in lung SQC has been identified, there is still a pau-city in the understanding of the mechanisms regarding howSOX2-mediated signaling network affects the formation orprogression of lung SQC cells.
In an initiative, using SOX2-abundant lung SQC cell lines,we screened for genes whose expression was not only affectedby silencing of SOX2 but can also account for the inhibition
of cell growth upon SOX2 silencing. Among the other candi-date genes, we found that BMP4, a member of the TGF-bsuperfamily genes, was significantly affected by silencingSOX2. As genes of TGF-b family are well known for theirinvolvement in the regulation of cell proliferation and differ-entiation, we postulated that BMP4 is a plausible target ofSOX2. In our study, several lines of evidence from in vitroand in vivo observation were provided showing that BMP4expression may be transcriptionally suppressed by SOX2 topromote growth of lung SQC cells.
Material and MethodsCell cultures
Human lung SQC cell lines NCI-H520 (H520 thereafter) wereobtained from ATCC (Manassas, VA), while EBC-1 and LK2were obtained from RIKEN (Ibaraki, Japan). All cell lines wereauthenticated by using the StemElight ID system (Promega,Madison, WI) and analyzed by ABI 3730 Genetic Analyzer andGeneMapper software v3.7 (Applied Biosystems, Carlsbad,CA). All cell lines were examined to exclude mycoplasma con-tamination. Cells were cultured in RPMI-1640 medium (Invi-trogen, Carlsbad, CA) with 10% fetal bovine serum(Invitrogen, Carlsbad, CA) at 37�C in 5% of CO2.
Clinical samples
Two independent sets of total RNA or cDNA derived fromclinical lung tissue samples were used for validation of meta-analysis study. The first set was from Chang Gung LungCancer collection. In this set total RNA samples were derivedfrom 55 clinical lung tissue samples (24 SQC and 31 normallung) from 43 individuals, among whom 12 individuals hadmatched pair of normal and tumor samples from the samepatient. These samples were collected from lung cancerpatients receiving operation of surgical removal at ChangGung Memorial Hospital, Taoyuan, Taiwan. All samples werecollected with approval from the Institutional Review Boardand informed consents from all participants. The other setwas commercially available from TissueScan tissue qPCRarray HLRT102 (OriGene, Rockville, MD), which contained13 lung SQC and five normal lung tissue samples. For immu-nohistochemistry (IHC) analysis, 35 lung SQC tissue samplesin tissue sections were obtained from 35 patients diagnosedof lung SQC and receiving operation of surgical resection atMacKay Memorial Hospital, Taipei, Taiwan; all of these sam-ples were collected with approval from the InstitutionalReview Board and informed consents from all participants.
What’s new?
The oncogene SOX2 is overexpressed in lung cancer. To find out how it contributes to cancer development, these authors
investigated which genes were affected by silencing SOX2. They discovered that a lack of SOX2 significantly boosted the
expression of the gene BMP4. As a member of the TGF-b family, BMP4 seemed a good candidate for a key player in the cancer
process. Further investigation revealed that tumor cells contain less BMP4, and that reactivation of BMP4 could slow tumor
growth and halt cell proliferation. These findings may explain how SOX2 promotes tumor development.
Can
cerCellBiology
2 SOX2 downregulates BMP4 in lung SQC cells
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

Gene expression microarray experiment
Half of each cell line (H520 and LK2) was treated with SOX2siRNA, whereas the other half was treated with controlsiRNA. After 48 hr, cells were harvested and total RNA wasextracted for gene expression microarray analysis. The proto-col for gene expression profiling experiments using Illumina’sBeadChip Human HT-12v3 array (Illumina, San Diego, CA)was as described previously10 with minor modification. Illu-mina GenomeStudio software (Illumina, San Diego, CA) wasused to analyze the raw data. Quantile normalization usingGeneSpringGX 11 software was further applied before analy-sis to obtain differentially expressed genes in testing samples.These microarray data have been deposited to GEO with theaccession number GSE48871.
Enzyme-linked immunosorbent assay
The level of BMP4 protein secreted into conditioned mediumwas determined by enzyme-linked immunosorbent assay(ELISA) Kit (R&D Systems, Minneapolis, MN) using mousemonoclonal anti-BMP4 antibody (MAB7571, R&D Systems,Minneapolis, MN) as capture antibody and biotinylated mousemonoclonal antibody (BAM7572, R&D Systems, Minneapolis,MN) as detection antibody according to the manufacturer’sinstructions. The conditioned medium from each experimentalcondition was concentrated by YM-10 Centricon (Millipore,Billerica, MA) for 2- to 20-fold before the assay. Solutions ofvarious concentrations (0.125, 0.25, 0.5, 1.0, 2.0 and 4.0 ng/ml)of recombinant human BMP4 (R&D Systems, Minneapolis,MN) in phosphate-buffered saline were generated by serialdilution for construction of standard curve.
Clonogenic assay
For transfection experiments, cells were collected, limiteddiluted and reseeded at 48 hr post-transfection. To test theeffect of BMP4 on cell growth, recombinant human BMP4(R&D Systems, Minneapolis, MN) of indicated concentrationwas directly added in the culture medium for the clonogenicassay. For each condition, 100 cells were seeded with culturemedium in triplicates in six-well plates. After 14–20 days,depending on growth rate, foci formed by least 50 cells werestained with crystal violet and counted.
Luciferase reporter assay
Luciferase reporter assay was performed using Duo-Glo sys-tem (Promega, Madison, WI) according to the manufac-turer’s instruction. All primers used for amplification ofputative BMP4 promoter sequences (Fig. 4a) and subcloninginto pGL3-based reporter plasmids (Promega, Madison, WI)contained KpnI site in the forward primer and HindIII sitein the reverse primer. The primer sequences (all in 50 to 30
direction) were as follows. Forward primes: GGTACCAAGGATGTATGAGCACAAAA CAA for pGL3-BP1, GGTACCGAGAAAGTGAACTCAG GAGTGAC for pGL3-BP2, GGTACCAAGAGGAGGAAGGAAGATGCGAG for pGL3-BP3,
GGTACCTTTTAGAACC GCGCTCTCCCGCC for pGL3-BP4,GGTACCCTGCTTTGC ATGGGGAGAGG for pGL3-BP5 andGGTACCCCTCTCG GTG GTGAGGCTTTGGG for pGL3-BP6. Reverse primer used for all constructs: AAGCTTGAAGCCAGAGGATTT GGGGCGC.
Statistical analysis
Parametric Student’s t-test or nonparametric Wilcoxon testwas used to assess the significance of difference between dif-ferent conditions of treatment depending on whether data ofinterest was apparently normally distributed. In all analyses,p value< 0.05 was considered as statistically significant.
ResultsA negatively correlated relationship between mRNA levels
of SOX2 and BMP4 in lung SQC cells/tissues
To identify genes/pathways likely to be downstream of SOX2,we sought for SOX2-expressing human lung SQC cell lines toconduct SOX2-silencing experiments and analyze associatedglobal gene transcription changes by gene expression microar-ray assay. Among lung SQC cell lines surveyed, we found thatboth LK2 and H520 were SOX2 abundant, and their SOX2expression can be almost abolished by treatment with SOX2siRNAs (Fig. 1a, inset). Consistent with previous reports,6 wenoted that growth of both cell lines was inhibited when SOX2was knocked down (to be described later). To identify candi-date SOX2 target genes, we screened for genes having at least1.5-fold change in transcript level in both cell lines uponsilencing of SOX2. We then looked for genes whose functionsare known to be involved in growth regulation. With a 1.6-foldand 4.4-fold upregulation in LK2 and H520 lines, respectively,BMP4 was one of the candidate genes that met the above crite-ria. Notably, the 4.4-fold upregulation of BMP4 was among thetop three most upregulated genes in the H520 cell line (datanot shown). Besides, BMP4 is well known for its roles in theregulation of cell proliferation, differentiation and apoptosis.The upregulation of the BMP4 expression in gene expressionarray assay was confirmed by a quantitative reversetranscription PCR (qRT-PCR, cf. Supporting Information PartI) analysis (Fig. 1a), which revealed a 3.5-fold and 9.0-foldupregulation in LK2 and H520 cells, respectively, suggestingthat BMP4 transcription could be negatively regulated bySOX2 in lung SQC cells.
Next, we wondered if a similar relationship existedbetween SOX2 and BMP4 mRNA levels in lung SQC tissuesor other cell lines. To answer this, we conducted a meta-analysis in the public lung cancer datasets derived from geneexpression microarray analysis on clinical samples or celllines. Interestingly, a moderate to significant negative correla-tion (r5 20.23 to 20.57) between the mRNA levels of SOX2and BMP4 was observed in six datasets containing lung SQCtissue samples or cell lines (Fig. 1b, Supporting InformationFigs. S1A–S1E and Table S1). These results indicated thatBMP4 expression could also be negatively regulated by SOX2in lung SQC tissues.
Can
cerCellBiology
Fang et al. 3
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

As previous studies have indicated that the majority oflung SQC tissues had elevated SOX2 mRNAs when comparedwith the adjacent normal tissues, we further hypothesizedthat BMP4 mRNA could be downregulated in lung SQC tis-sues, relative to the adjacent normal lung tissues. To verifythis hypothesis, we conducted another meta-analysis in data-sets containing lung SQC and normal lung tissues simultane-ously. Based on such criteria, three datasets (Boston,GSE19188 and GSE 18842) from independent studies, whichin total contain 93 lung SQC tissues and 127 normal lung tis-sues, were selected. As expected, the results invariably showeda significant upregulation of SOX2 expression but downregu-lation of BMP4 expression in the majority of lung SQC tissuesamples when compared with normal lung tissues (Fig. 1c,Supporting Information Figs. S2A and S2B and Table S2). Toverify the downregulation of BMP4 in lung SQC tissuesrevealed by meta-analysis, a validation assay using qRT-PCRwith our own samples from two independent cohorts (seeMaterial and Methods section) was conducted. Consistentwith the results of meta-analysis, both the Chang Gung LungCancer tissue samples (Fig. 1d) and another panel of com-
mercial cDNA samples (Supporting Information Fig. S2C)showed significant upregulation of SOX2 mRNA and down-regulation of BMP4 mRNA, as summarized in SupportingInformation Table S2.
SOX2 negatively regulates BMP4 protein expression in
lung SQC cells
To know whether BMP4 protein expression in lung SQC cells isalso subject to regulation of SOX2, we conducted the followingexperiments. In the first one, a Western blotting analysis wasused to examine the BMP4 protein expression under SOX2-knockdown condition. Under the SOX2-knockdown condition,a roughly 1.4- and 2.0-fold increase in the levels of BMP4 pre-cursor protein (Fig. 2a) could be reached in LK2 and H520 cells,respectively. By contrast, in EBC-1, another lung SQC cell linewithout detectable SOX2 protein expression, its forced expres-sion by transfection of SOX2 plasmid DNA caused a significantdecrease in the level of BMP4 precursor protein (Fig. 2b).
In the second experiment, as BMP4 is known as a secretedprotein, we applied ELISA to examine whether the amount ofmature BMP4 secreted to the extracellular space was also
Figure 1. siRNA transfection experiment and meta-analysis revealed negatively correlated relationship between the mRNA levels of SOX2
and BMP4 in lung SQC cells/tissues. (a) Upregulation of BMP4 mRNA in LK2 and H520 by SOX2 silencing. Cells were transfected with siRNA
of SOX2 (siSOX2) or nontargeting siRNA (control) for 2 days and subjected to quantification of mRNAs expression using qRT-PCR. Shown in
inset is a Western blotting analysis validating silencing of SOX2 protein expression by siRNA treatment. Values represent mean 6 SD of trip-
licate measurements. (b) A representative result of meta-analysis on GSE6253 dataset showing a negative correlation between transcript
levels of SOX2 and BMP4 in lung SQC tissues. (c and d) Box plots showing relative SOX2 and BMP4 mRNA expression levels of lung SQC
and adjacent normal lung (N) tissues in a public dataset (BOSTON) and in our Chang Gung Lung Cancer validation cohort.
Can
cerCellBiology
4 SOX2 downregulates BMP4 in lung SQC cells
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

affected by silencing of SOX2. As shown in Figure 2c, a markedincrease in the level of secreted BMP4 in the culture media wasobserved in both LK2 and H520 upon SOX2 silencing. By con-trast, transfection of SOX2 plasmid DNA into EBC-1 cellsresulted in a decrease in the amount of secreted BMP4, eventhough the basal level of BMP4 in the conditioned medium ofEBC-1 cells was already low (Supporting Information Fig. S3).Interestingly, we also noted that owing to unknown reason, thebasal levels of secreted BMP4 differed between H520 and LK2by about two orders of magnitude (Fig. 2c).
The upregulation of BMP4 expression in those lung SQCcells upon SOX2 silencing strongly implicates that BMP4 sig-naling could be activated by SOX2 silencing. To validate this,we tested if the level of phosphorylated SMAD1/5 (pSMAD1/5), an indicative of activation of BMP signaling,11 could beaffected by SOX2. As shown in Figure 2d, a significant eleva-tion in the level of pSMAD1/5 was observed in both LK2and H520 cells with knockdown SOX2, indicating that BMP4signaling could be reactivated and involved in growth inhibi-tion upon SOX2 silencing.
Lung SQC tumor tissues have high SOX2 and low BMP4
protein expression
To examine the relationship of protein levels between SOX2and BMP4 in lung SQC tissues, we analyzed the IHC staining
of the two proteins in a panel of 35 lung SQC tumor tissuesamples, as summarized in Figure 3a. Similar to the previousfindings, we found that the majority (74%, n5 26) of samplesshowed strong (Fig. 3b) or moderate SOX2 expression, andonly few of them (26%, n5 9) showed weak SOX2 expression.By contrast, most (89%, n5 31) displayed negative (Fig. 3c) orweak BMP4 expression, while only few (11%, n5 4) showedmoderate BMP4 expression in tumor parts of lung SQC tis-sues. In adjacent normal lung tissues, however, the aboveSOX2-high and BMP4-low expression pattern of lung SQC tis-sues was completely reversed. SOX2 expression in adjacentnormal lung tissues was negative (Fig. 3b, upper part) in themajority (86%, n5 30) and moderate in merely few (14%,n5 5) of samples. By contrast, BMP4 expression in adjacentnormal lung tissues was moderate (Fig. 3c, indicated byarrows) or strong (Figs. 3d and 3e) in most samples (80%,n5 28), and was weak or negative in few (20%, n5 7) sam-ples. In general, BMP4 expression was rare in tumor parts oflung SQC tissues, but could be frequently detected in the adja-cent normal tissues, possibly including stromal fibroblasts (Fig.3c, indicated by arrows), alveolar macrophages (Fig. 3d, Sup-porting Information Fig. S4), bronchiolar epithelium (Fig. 3e)or pneumocytes. These results corroborated with our aboveobservations and implied that BMP4 protein expression couldalso be negatively regulated by SOX2 in lung SQC tissues.
Figure 2. SOX2 and BMP4 protein expression in lung SQC cell lines. (a) Western blotting analyses of LK2 and H520 cells transfected with
SOX2 siRNA (1) or nontargeting control siRNA (2) harvested at indicated time intervals post-siRNA transfection. Shown in the top are the
results of densitometry analysis of BMP4. (b) Western blotting analysis of SOX2 plasmid DNA-transfected EBC-1 cells harvested at 72 hr
post-transfection. Results of densitometry analyses of BMP4 are also shown in the top. (c) ELISA analysis showing BMP4 levels in the con-
ditioned media collected at indicated time intervals post-transfection with siSOX2 or nontargeting control siRNA. Values represent
mean 6 SD of triplicate measurements. (d) A Western blotting analysis of phospho-SMAD1/5 (pSMAD1/5) protein in cells transfected with
siSOX2 (1) or nontargeting control siRNA (2). Also shown in the top are results of densitometry analysis of pSMAD/1/5 normalized by a-
TUBULIN and further by control experiment (2).
Can
cerCellBiology
Fang et al. 5
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

Despite the obvious patterns of differential expression ofSOX2 and BMP4 in tumor versus adjacent normal tissues, nosignificant correlation between them and clinicopathologicaldata was observed (Supporting Information Table S3). Per-haps, this is due to limited sample size of this cohort or it islikely that dysregulated expression of both proteins isinvolved primarily in the tumorigenesis instead of progres-sion of lung SQC.
SOX2 negatively regulates BMP4 promoter activity
As SOX2 is a transcription factor, we next asked whether theupregulation of BMP4 mRNA expression upon silencing
SOX2 could be due to loss of transcriptional repression bySOX2. To assess such possibility, we applied luciferasereporter assay to search in the upstream or putative regula-tory loci of the BMP4 gene for regions that could response tothe regulation of SOX2. Among the target regions surveyed(Fig. 4a), pGL3-BP4 and pGL3-BP5, two reporter constructswith sequences corresponding to intron 1 of BMP4 gene,showed most significant upregulation in reporter activitiesupon SOX2 knockdown (Fig. 4b). As one of the two pro-moters of BMP4 is known to be localized within the firstintron,12,13 our results suggest the possibility of binding ofSOX2 to DNA within intron 1. In support of this notion, our
Figure 3. Immunohistochemistry of SOX2 and BMP4 in lung SQC tissues. (a) A summary of IHC analyses of BMP4 and SOX2 of 35 lung SQC
tissues and their adjacent normal lung tissues of our McKay Memorial Hospital cohort. (b) A representative image of IHC analysis showing
strong nuclear SOX2 expression in tumor cells (area inside dash line) but negative in surrounding normal fibroblast tissues. (c) A represen-
tative image of IHC analysis showing negative BMP4 expression in tumor cells and moderate BMP4 expression in surrounding normal fibro-
blast tissues (indicated by arrows). Tissue slides used in (b) and (c) were nearby tissue sections from the same tumor block. (d and e)
Images of IHC analyses showing strong BMP4 expression in alveolar macrophages (d) or bronchiolar epithelia cells (e, indicated by arrows)
within the adjacent normal parts of a lung SQC tissue section. Histological judgment of alveolar macrophages was based on their granular
shape with indented nucleus, and bronchiolar epithelium was based on their ciliated pseudostratified columnar structure with goblet cells.
Can
cerCellBiology
6 SOX2 downregulates BMP4 in lung SQC cells
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC
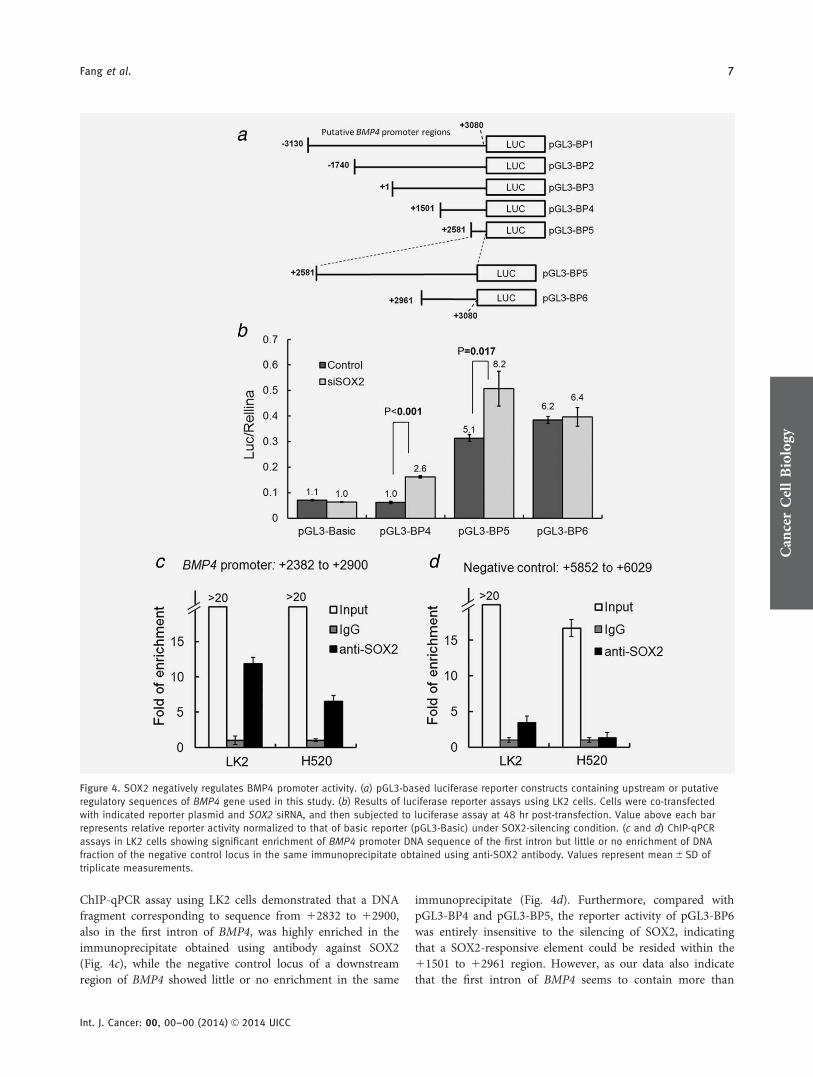
ChIP-qPCR assay using LK2 cells demonstrated that a DNAfragment corresponding to sequence from 12832 to 12900,also in the first intron of BMP4, was highly enriched in theimmunoprecipitate obtained using antibody against SOX2(Fig. 4c), while the negative control locus of a downstreamregion of BMP4 showed little or no enrichment in the same
immunoprecipitate (Fig. 4d). Furthermore, compared withpGL3-BP4 and pGL3-BP5, the reporter activity of pGL3-BP6was entirely insensitive to the silencing of SOX2, indicatingthat a SOX2-responsive element could be resided within the11501 to 12961 region. However, as our data also indicatethat the first intron of BMP4 seems to contain more than
Figure 4. SOX2 negatively regulates BMP4 promoter activity. (a) pGL3-based luciferase reporter constructs containing upstream or putative
regulatory sequences of BMP4 gene used in this study. (b) Results of luciferase reporter assays using LK2 cells. Cells were co-transfected
with indicated reporter plasmid and SOX2 siRNA, and then subjected to luciferase assay at 48 hr post-transfection. Value above each bar
represents relative reporter activity normalized to that of basic reporter (pGL3-Basic) under SOX2-silencing condition. (c and d) ChIP-qPCR
assays in LK2 cells showing significant enrichment of BMP4 promoter DNA sequence of the first intron but little or no enrichment of DNA
fraction of the negative control locus in the same immunoprecipitate obtained using anti-SOX2 antibody. Values represent mean 6 SD of
triplicate measurements.
Can
cerCellBiology
Fang et al. 7
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

Figure 5. Involvement of BMP4 signaling in SOX2-silencing-mediated inhibition of cell/tumor growth. (a) An xenograft tumor model based
on LK2 cells stably transfected with shRNA of SOX2 (shSOX2, open circle) or empty vector control (control, filled circle). The Western blot-
ting analysis in inset shows reduced expression of SOX2 in shSOX2 transfectant. All mice in this experiment grew tumor. Values represent
mean tumor volume 6 SD of tumors from eight mice in each group. (b) A box plot showing relative BMP4 mRNA expression in xenograft
tumors. Four tumors from shSOX2 group and five from control group obtained from mice sacrificed on day 33 were subjected to RNA extrac-
tion and qRT-PCR assays for measurement of BMP4 mRNA level. Values are average of triplicate measurements and then normalized by
result of a randomly selected tumor of control group. (c) A bar chart showing plasma BMP4 levels of mice of the xenograft tumor model
determined by ELISA assay. Values represent mean 6 SD of eight mice for each group. (d) A plot showing inhibition of clonogenic growth of
lung SQC cells by rhBMP4. (e) Representative images of clonogenic assay for both LK2 and H520 cells showing significant reduction in
number and size of colonies in the presence of rhBMP4 (100 ng/ml). (f and g) Bar charts showing clonogenic growth of cells transiently
transfected with indicated siRNAs and/or LDN-193189. Values represent mean 6 SD of triplicate measurements. (h) Mice xenograft tumor
model showing injection of LDN-193189 can partially rescue SOX2-silencing induced inhibition of tumor growth. The numbers of tumor-
bearing mice in control, siSOX2 and siSOX21LDN groups were 3, 4 and 3 mice, respectively. Values represent mean volume 6 SD of tumors
from tumor-bearing mice. * represent p<0.05 from t-test comparing tumor size between siSOX2 and siSOX2 1 LDN groups.
Can
cerCellBiology
8 SOX2 downregulates BMP4 in lung SQC cells
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

one SOX2-binding sites from other analyses (data notshown), further investigations are required to fine-map theregulatory site(s) in response to trans-acting regulationimposed by SOX2.
Upregulation of BMP4 expression is associated with
reduction of tumor growth mediated by SOX2 silencing
In addition to the cell-based experiments, we have establisheda LK2 cell-based xenograft tumor model in mice to furtherunderstand the significance of SOX2-BMP4 interaction invivo. We found that inoculation of SOX2 shRNA (shSOX2)transfectant or vector control cells could induce xenograftformation in all mice inoculated. However, tumors ofshSOX2 transfectant group exhibited significantly reducedkinetics of growth and size of tumors formed than those ofcontrol group (Fig. 5a and Supporting Information Figs. S5Aand S5C), indicating silencing SOX2 has tumor-inhibitoryeffect in vivo. In this model, we then asked whether theBMP4 mRNA levels of xenograft tumors could also beaffected by SOX2 silencing. The results showed that alltumors of shSOX2 transfectant group had significantly higherlevel of BMP4 mRNA and protein expression than those ofcontrol group (Fig. 5b and Supporting Information Figs. S5Band S5D). Very intriguingly, for both groups, the plasmaBMP4 level in mice (detected by ELISA) decreased as tumorsgrew (Fig. 5c). Moreover, a significant elevated plasma BMP4level of mice of the shSOX2 transfectant group relative tothat of control group (p5 0.037) was observed on day 21post-injection, suggesting that a higher level of plasma BMP4could be associated with inhibited growth of tumor.
Growth inhibition induced by SOX2 silencing was partially
mediated through BMP4 signaling
BMP4 has been known to be involved in the regulation ofcell proliferation. We further asked whether BMP4 also had asimilar role in lung SQC cells. As shown in Figures 5d and5e, the clonogenic growth of both LK2 and H520 cells, interms of size and number of colonies formed, was decreasedby the presence of recombinant human BMP4 (rhBMP4)protein in a dose-dependent manner, indicating a growth-inhibitory effect. Remarkably, the sensitivity of the cells torhBMP4 treatment differed significantly, a phenomenon evi-dently associated with the previously described difference inthe basal levels of BMP4 secreted in conditioned mediabetween cell lines (Fig. 2c). This could be because that lungSQC cells of various origins have evolved with different capa-bilities to evade the regulation by BMP4. Regardless of thedifference in sensitivity to BMP4 inhibition, these resultsimplied that knockdown of SOX2 might inhibit proliferationthrough upregulation of BMP4. To verify such assumption,we tested whether silencing BMP4 could rescue the growthinhibition upon silencing of SOX2. We found that transfec-tion of both LK2 and H520 cells with BMP4 siRNA(siBMP4) alone did not affect cell growth, perhaps due topredominant inhibition of BMP4 expression by endogenous
SOX2. However, co-transfection of siBMP4 and siSOX2 sig-nificantly suppressed SOX2-silencing-mediated upregulationof BMP4 mRNA (Supporting Information Fig. S6), and couldrescue 30 and 60% of inhibition (Fig. 5f) in LK2 and H520cells, respectively, from SOX2-silencing-induced reduction ofcolony formation. In another similar experiment, the siBMP4
Figure 6. Co-transfection of SOX2 and BMP4 siRNAs partially
released arrest of cell cycle induced by silencing SOX2. (a–d) Flow
cytometry cell cycle analysis of LK2 cells transiently transfected
with nontargeting control siRNA (control), BMP4 siRNA (siBMP4),
SOX2 siRNA (siSOX2) and the mixture of SOX2 and BMP4 siRNAs
(siSOX2 1 siBMP4). For each condition, cells were harvested at 72
hr post-transfection. FL2-A represents fluorescence intensity of pro-
pidium iodide (PI) staining. Indicated in the inset of each panel
were percentages of each phase of cell cycle, where values repre-
sent mean 6 SD of triplicate measurements.
Can
cerCellBiology
Fang et al. 9
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

was replaced by LDN-193189, a specific inhibitor of BMPreceptor. As expected, although LDN-193189 treatment alonehad little or no effect on cell growth, co-treatment of cellswith LDN-193189 and SOX2 siRNA could rescue 40 and60% inhibition (Fig. 5g) in LK2 and H520 cells, respectively,from SOX2-silencing-induced growth inhibition. Consistently,when MTT assay was applied, similar results were observed(Supporting Information Fig. S7). Also in agreement with thecell-based assays, in another xenograft tumor model, routineinjection of LDN-193189 to mice inoculated with SOX2-knockdown LK2 cells (siSOX21 LDN) can partially rescueSOX2-silencing-induced inhibition of tumor growth (Fig. 5h).These results as a whole indicated that BMP4 signaling reac-tivation in lung SQC cells may partially account for the inhi-bition of cell growth caused by knocking down SOX2.
Finally, as BMP4 has been known to regulate cell cycle pro-gression and modulate cell growth, we performed flow cytom-etry analysis to determine cell cycle phases under conditionssimilar to the co-transfection experiments above. In LK2 cells,we found that siBMP4 alone caused little or no effect on distri-bution of cell cycle phases (Figs. 6a and 6b), whereas transfec-tion of siSOX2 in LK2 cells led to significant accumulation ofcells in G1 phase (Fig. 6c) accompanied by upregulation ofp21 (Supporting Information Fig. S8). However, co-transfection of siSOX2 and siBMP4 could significantly releasecells from SOX2-silencing-mediated G1 arrest (Fig. 6d) anddownregulate p21 expression (Supporting Information Fig.S8), suggesting that SOX2 might promote cell cycle progres-sion via inhibiting antiproliferative BMP4 signaling.
DiscussionBMP4 was originally identified as a regulator of cartilage andbone formation, and is in general known for its critical rolesin embryonic development, mesenchymal development andorganogenesis of a variety of organs including the kidney,lung, heart, teeth, gut and skin (cf. Ref. 14 for review), andhas been implicated in adult lung homeostasis and tissueinjury repair.15,16 With special interest to development of thelung and associated branching morphogenesis, BMP4 hasbeen known as one of the key mediators of interactive signal-ing between endodermal epithelium and mesodermal mesen-chyme, in which cell proliferation and differentiation areregulated by BMP4.17 Our findings of BMP4 signaling as aSOX2 downstream and its suppression by oncogenic SOX2 inlung SQC suggest that it is worthy to further investigate therole(s) of BMP4 signaling in the development of lung SQCfrom normal lung tissue and associated pathogenesis.
Another important finding that BMP4 has a tumor-suppressive effect in lung SQC cells has been revealed for thefirst time in this study. This is, however, not surprising becausemany members of BMP protein family have been known tohave broad spectrum of anticancer capabilities, such as antipro-liferative or growth-inhibitory activities,18,19 metastasis-suppressive potential20,21 or apoptosis-inducing capability.22 Inthis regard, BMP4 has been known to inhibit cell growth or pro-liferation of prostate cancer23 or induce apoptosis in myelomacells.24 Furthermore, BMP4 has even been implicated as a spe-cific inhibitor of cancer metastases with the potential to beapplied via direct administration to treat breast cancer.25
Related to lung cancer, BMP4 was shown to induce senes-cence26 or mediate adriamycin-induced premature senescence27
in A549 cells, a cell lines derived from lung ADC. Our findingthat BMP4 signaling may be suppressed in lung SQC cells byoverrepresented SOX2 makes BMP4 a promising candidate tobe evaluated for its therapeutic effect in lung SQC treatment.
Our study provides evidences for transcriptional regula-tion of BMP4 expression by SOX2. However, we cannotexclude possibilities of alternative mechanism of the interac-tion between SOX2 and BMP4. For example, we found thatSOX2 had cytoplasmic expression pattern in tumor cells ofseveral cases of lung SQC (Supporting Information Fig. S9A),which is consistent with previous reports in lung SQC tissuesand glioma cell lines.8,28 Interestingly, even for such nontypi-cal localization of SOX2, the mutually exclusive expressionpattern between SOX2 and BMP4 remained obvious withintumor tissues having heterogeneous levels of SOX2 expres-sion (Supporting Information Fig. S9). This implies thatSOX2 protein might have unknown cytosolic function(s) asproposed elsewhere,29 or that BMP4 could be indirectly regu-lated by SOX2 via other unknown mechanism.
Target genes of SOX2 in various types of cancer cells havebeen linked to multiple pathways important for developmentof normal cells, and their dysregulation is very likely to con-tribute to tumorigenesis or cancer progression. It is, there-fore, appealing to further dissect other pathways, e.g.,apoptosis signaling,30 that are regulated by SOX2 in lungcancer and identify other downstream targets with therapeu-tic potential. Delineating the associated pathways and mecha-nisms would help us to understand the formation of lungSQC and identify new targets for lung cancer therapy.
AcknowledgementsThe authors thank Dr. Ya-Wen Chen and Dr. Wen-Chun Hung for criticalcomments.
References
1. Siegel R, Naishadham D, Jemal A. Cancer statis-tics, 2012. CA Cancer J Clin 2012;62:10–29.
2. Howlader N, Noone AM, Krapcho M, et al.SEER Cancer Statistics Review, 1975–2010 [Inter-net]. Bethesda, MD: National Cancer Institute.Available at: http://seer.cancer.gov/csr/1975_2010.Last accessed October 20, 2013.
3. Dodds L, Davis S, Polissar L. A population-basedstudy of lung cancer incidence trends by histologictype, 1974-81. J Natl Cancer Inst 1986;76:21–9.
4. Janssen-Heijnen ML, Nab HW, van Reek J, et al.Striking changes in smoking behaviour and lungcancer incidence by histological type in south-
east Netherlands, 1960–1991. Eur J Cancer 1995;31A:949–52.
5. Drilon A, Rekhtman N, Ladanyi M, et al. Squa-mous-cell carcinomas of the lung: emerging biol-ogy, controversies, and the promise of targetedtherapy. Lancet Oncol 2012;13:e418–e426.
Can
cerCellBiology
10 SOX2 downregulates BMP4 in lung SQC cells
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

6. Bass AJ, Watanabe H, Mermel CH, et al. SOX2 isan amplified lineage-survival oncogene in lungand esophageal squamous cell carcinomas. NatGenet 2009;41:1238–42.
7. Lockwood WW, Chari R, Coe BP, et al. Integra-tive genomic analyses identify BRF2 as a novellineage-specific oncogene in lung squamous cellcarcinoma. PLoS Med 2010;7:e1000315.
8. Hussenet T, Dali S, Exinger J, et al. SOX2 is anoncogene activated by recurrent 3q26.3 amplifica-tions in human lung squamous cell carcinomas.PLoS One 2010;5:e8960.
9. Yuan P, Kadara H, Behrens C, et al. Sex deter-mining region Y-Box 2 (SOX2) is a potential cell-lineage gene highly expressed in the pathogenesisof squamous cell carcinomas of the lung. PLoSOne 2010;5:e9112.
10. Jiang SS, Fang WT, Hou YH, et al. Upregulationof SOX9 in lung adenocarcinoma and its involve-ment in the regulation of cell growth and tumori-genicity. Clin Cancer Res 2010;16:4363–73.
11. Hoodless PA, Haerry T, Abdollah S, et al. MADR1,a MAD-related protein that functions in BMP2 sig-naling pathways. Cell 1996;85:489–500.
12. van den Wijngaard A, van Kraay M, van ZoelenEJ, et al. Genomic organization of the humanbone morphogenetic protein-4 gene: molecularbasis for multiple transcripts. Biochem BiophysRes Commun 1996;219:789–94.
13. Kurihara T, Kitamura K, Takaoka K, et al.Murine bone morphogenetic protein-4 gene: exis-tence of multiple promoters and exons for the 5’-untranslated region. Biochem Biophys Res Com-mun 1993;192:1049–56.
14. Hogan BL. Bone morphogenetic proteins: multi-functional regulators of vertebrate development.Genes Dev 1996;10:1580–94.
15. Koli K, Myllarniemi M, Vuorinen K, et al. Bonemorphogenetic protein-4 inhibitor gremlin isoverexpressed in idiopathic pulmonary fibrosis.Am J Pathol 2006;169:61–71.
16. Rosendahl A, Pardali E, Speletas M, et al. Activationof bone morphogenetic protein/Smad signaling inbronchial epithelial cells during airway inflamma-tion. Am J Respir Cell Mol Biol 2002;27:160–9.
17. Shannon JM, Hyatt BA. Epithelial-mesenchymalinteractions in the developing lung. Annu RevPhysiol 2004;66:625–45.
18. Ghosh-Choudhury N, Ghosh-Choudhury G,Celeste A, et al. Bone morphogenetic protein-2induces cyclin kinase inhibitor p21 and hypo-phosphorylation of retinoblastoma protein inestradiol-treated MCF-7 human breast cancercells. Biochim Biophys Acta 2000;1497:186–96.
19. Arnold SF, Tims E, McGrath BE. Identification ofbone morphogenetic proteins and their receptorsin human breast cancer cell lines: importance ofBMP2. Cytokine 1999;11:1031–7.
20. Rees JR, Onwuegbusi BA, Save VE, et al. In vivoand in vitro evidence for transforming growthfactor-beta1-mediated epithelial to mesenchymaltransition in esophageal adenocarcinoma. CancerRes 2006;66:9583–90.
21. Yang S, Du J, Wang Z, et al. BMP-6 promotes E-cadherin expression through repressing deltaEF1in breast cancer cells. BMC Cancer 2007;7:211.
22. Hallahan AR, Pritchard JI, Chandraratna RA,et al. BMP-2 mediates retinoid-induced apoptosis
in medulloblastoma cells through a paracrineeffect. Nat Med 2003;9:1033–8.
23. Brubaker KD, Corey E, Brown LG, et al.Bone morphogenetic protein signaling inprostate cancer cell lines. J Cell Biochem 2004;91:151–60.
24. Hjertner O, Hjorth-Hansen H, Borset M, et al.Bone morphogenetic protein-4 inhibits prolifera-tion and induces apoptosis of multiple myelomacells. Blood 2001;97:516–22.
25. Smith SC, Theodorescu D. Learning therapeuticlessons from metastasis suppressor proteins. NatRev Cancer 2009;9:253–64.
26. Buckley S, Shi W, Driscoll B, et al. BMP4 signal-ing induces senescence and modulates the onco-genic phenotype of A549 lung adenocarcinomacells. Am J Physiol Lung Cell Mol Physiol 2004;286:L81–L86.
27. Su D, Zhu S, Han X, et al. BMP4-Smad signalingpathway mediates adriamycin-induced prematuresenescence in lung cancer cells. J Biol Chem 2009;284:12153–64.
28. Annovazzi L, Mellai M, Caldera V, et al. SOX2expression and amplification in gliomas and gli-oma cell lines. Cancer Genomics Proteomics 2011;8:139–47.
29. Jia X, Li X, Xu Y, et al. SOX2 promotes tumori-genesis and increases the anti-apoptotic propertyof human prostate cancer cell. J Mol Cell Biol2011;3:230–8.
30. Chen S, Li X, Lu D, et al. SOX2 regulatesapoptosis through MAP4K4-Survivin signalingpathway in human lung cancer cells. Carcinogene-sis 2014 Jan 22 [Epub ahead of print].
Can
cerCellBiology
Fang et al. 11
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC




















