
Transcriptional Regulation of BMP4 in the XenopusEmbryo: Analysis of Genomic BMP4 and Its Promoter
15
Transcriptional Regulation of BMP-4 in the Xenopus Embryo: Analysis of Genomic BMP-4 and Its Promoter Jaebong Kim,* Katherine Tidman Ault,² Hung-Dar Chen,* Ren-He Xu,² Dong-Hyun Roh,‡ Marie C. Lin,² Mae-Ja Park,§ and Hsiang-Fu Kung* ,1 *Laboratory of Biochemical Physiology, Frederick Cancer Research and Development Center, National Cancer Institute, Frederick, Maryland 21702-1201; ²Intramural Research Support Program, Science Applications International Corporation- Frederick, Frederick, Maryland 21702-1201; ‡ National Institute of Diabetes and Digestive and Kidney Diseases, Laboratory of Biochemistry and Genetics, Bethesda, Maryland 20892; and §Department of Anatomy, Kyung-Pook National University, Taegu, Korea Received August 6, 1998 Recent experiments in the Xenopus embryo suggest that proper regulation of BMP-4 signaling is critical to the dorsal ventral specification of both mesoderm and ectoderm. Regulation of BMP-4 signaling is known to occur extracellularly by direct binding with chordin, noggin, and follistatin, and intracellularly through the antagonistic signal interaction with dorsalizing TGF-b family member activin. However, the tight repres- sional regulation of BMP transcription may also be required to sustain the dorsal and neural status of the induced cells. Here we demonstrate that the dominant negative mutant of the BMP receptor (DN-BR) or the BMP-4 antagonizers, chordin and noggin, negatively regulate BMP-4 transcription in animal cap explants. We suggest that repression of BMP-4 transcription is important in the maintenance of dorsal fate and that continuous input of BMP-4 signaling is required to sustain the expression of BMP-4 transcription in the maintenance of epidermal/ventral fate. Consistent with this postulation, we found that the promoter re- gion of the isolated BMP-4 genomic DNA includes sev- eral consensus binding sites for transcriptional regu- lators functioning under BMP-4 signaling such as GATA binding and ventralizing homeobox genes. In a functional assay we found that the GATA binding and ventral homeobox proteins can positively modulate BMP-4 promoter activity. We also observed that DN-BR decreases BMP-4 promoter activity. This was likely due to a repression of the above-mentioned tran- scription factors. The significance of these observa- tions to embryonic patterning is discussed. Recent experiments in the Xenopus embryo suggest that proper regulation of the BMP-4 signaling pathway is critical to the dorsal ventral specification of both mesoderm and ectoderm. In the mesoderm, BMP-4 is a potent ventralizing factor that is able to overcome the dorsalizing effects of activin and the organizer itself (1-2). At the onset of gastrulation, BMP-4 transcripts are uniformly dis- tributed in the animal cap and ventral marginal zone, but are excluded from the dorsal-most meso- dermal region (3-4). Injection of mRNA encoding BMP-4 into the dorsal marginal zone of intact em- bryos results in the suppression of dorsoanterior structures (1-2). Conversely, ventral expression of the dominant negative form of the BMP-4 receptor, which specifically blocks BMP-4 signaling, converts ventral mesoderm to dorsal mesoderm resulting in the formation of a secondary axis (5-7). Likewise, inhibition of BMP-4 signaling by dominant negative ligands or antisense BMP-4 mRNA results in meso- derm dorsalization (8-9). BMP-4 is thought to act as a morphogen, patterning the mesoderm along the dorsal to ventral axis. Dosch et al. has observed that increasing levels of BMP-4 signaling generate in- creasingly ventral derivatives of mesoderm (10). Ectodermal patterning also requires an intact BMP-4 signaling pathway. BMP-4 is thought to act endogenously to specify epidermis, and neural induc- tion may involve a BMP-4 antagonist (11-12). Two lines of evidence support this notion. First, BMP-4 suppresses autoneuralization and restores dispersed ectodermal cells to the epidermal fate of intact ex- plants (12). Second, ectopic expression of a dominant negative BMP-4 receptor (DN-BR) (11,13) of domi- nant negative ligands (8) or of antisense BMP-4 RNA (11) results in direct neural induction, suggesting 1 To whom correspondence and reprint requests should be ad- dressed. The U.S. Government’s right to retain a nonexclusive royalty-free license in and to the copyright covering this paper, for governmental purposes, is acknowledged. BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 1998, 516 –530 (1998) ARTICLE NO. RC989280 516 0006-291X/98
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Transcriptional Regulation of BMP4 in the XenopusEmbryo: Analysis of Genomic BMP4 and Its Promoter

Transcriptional Regulation of BMP-4 in the XenopusEmbryo: Analysis of Genomic BMP-4 and Its Promoter
Jaebong Kim,* Katherine Tidman Ault,† Hung-Dar Chen,* Ren-He Xu,† Dong-Hyun Roh,‡Marie C. Lin,† Mae-Ja Park,§ and Hsiang-Fu Kung*,1
*Laboratory of Biochemical Physiology, Frederick Cancer Research and Development Center, National Cancer Institute,Frederick, Maryland 21702-1201; †Intramural Research Support Program, Science Applications International Corporation-Frederick, Frederick, Maryland 21702-1201; ‡National Institute of Diabetes and Digestive and Kidney Diseases,Laboratory of Biochemistry and Genetics, Bethesda, Maryland 20892; and §Department of Anatomy,Kyung-Pook National University, Taegu, Korea
Received August 6, 1998
Recent experiments in the Xenopus embryo suggestthat proper regulation of BMP-4 signaling is critical tothe dorsal ventral specification of both mesoderm andectoderm. Regulation of BMP-4 signaling is known tooccur extracellularly by direct binding with chordin,noggin, and follistatin, and intracellularly through theantagonistic signal interaction with dorsalizing TGF-bfamily member activin. However, the tight repres-sional regulation of BMP transcription may also berequired to sustain the dorsal and neural status of theinduced cells. Here we demonstrate that the dominantnegative mutant of the BMP receptor (DN-BR) or theBMP-4 antagonizers, chordin and noggin, negativelyregulate BMP-4 transcription in animal cap explants.We suggest that repression of BMP-4 transcription isimportant in the maintenance of dorsal fate and thatcontinuous input of BMP-4 signaling is required tosustain the expression of BMP-4 transcription in themaintenance of epidermal/ventral fate. Consistentwith this postulation, we found that the promoter re-gion of the isolated BMP-4 genomic DNA includes sev-eral consensus binding sites for transcriptional regu-lators functioning under BMP-4 signaling such asGATA binding and ventralizing homeobox genes. In afunctional assay we found that the GATA binding andventral homeobox proteins can positively modulateBMP-4 promoter activity. We also observed thatDN-BR decreases BMP-4 promoter activity. This waslikely due to a repression of the above-mentioned tran-scription factors. The significance of these observa-tions to embryonic patterning is discussed.
Recent experiments in the Xenopus embryo suggestthat proper regulation of the BMP-4 signaling pathwayis critical to the dorsal ventral specification of bothmesoderm and ectoderm.
In the mesoderm, BMP-4 is a potent ventralizingfactor that is able to overcome the dorsalizing effectsof activin and the organizer itself (1-2). At the onsetof gastrulation, BMP-4 transcripts are uniformly dis-tributed in the animal cap and ventral marginalzone, but are excluded from the dorsal-most meso-dermal region (3-4). Injection of mRNA encodingBMP-4 into the dorsal marginal zone of intact em-bryos results in the suppression of dorsoanteriorstructures (1-2). Conversely, ventral expression ofthe dominant negative form of the BMP-4 receptor,which specifically blocks BMP-4 signaling, convertsventral mesoderm to dorsal mesoderm resulting inthe formation of a secondary axis (5-7). Likewise,inhibition of BMP-4 signaling by dominant negativeligands or antisense BMP-4 mRNA results in meso-derm dorsalization (8-9). BMP-4 is thought to act asa morphogen, patterning the mesoderm along thedorsal to ventral axis. Dosch et al. has observed thatincreasing levels of BMP-4 signaling generate in-creasingly ventral derivatives of mesoderm (10).
Ectodermal patterning also requires an intactBMP-4 signaling pathway. BMP-4 is thought to actendogenously to specify epidermis, and neural induc-tion may involve a BMP-4 antagonist (11-12). Twolines of evidence support this notion. First, BMP-4suppresses autoneuralization and restores dispersedectodermal cells to the epidermal fate of intact ex-plants (12). Second, ectopic expression of a dominantnegative BMP-4 receptor (DN-BR) (11,13) of domi-nant negative ligands (8) or of antisense BMP-4 RNA(11) results in direct neural induction, suggesting
1 To whom correspondence and reprint requests should be ad-dressed.
The U.S. Government’s right to retain a nonexclusive royalty-freelicense in and to the copyright covering this paper, for governmentalpurposes, is acknowledged.
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 1998, 516–530 (1998)ARTICLE NO. RC989280
5160006-291X/98

that neural induction requires the removal of endog-enous BMP signals in the ectoderm.
Thus, in Xenopus, a dorsal to ventral ascending gra-dient of BMP-4 signaling is required for the formationof distinct mesodermal and ectodermal derivatives.BMP-4 appears to play a similarly crucial role in thedorsal ventral patterning of both mouse (14) and Dro-sophila embryos (15-17). In the mouse embryo, BMP-4is required for the differentiation of mesoderm derivedfrom the posterior primitive streak and for develop-ment of the ventral lateral mesoderm (14). This obser-vation is compatible with the function of BMP-4 in theXenopus embryo. In Drosophila, DPP (regarded as ahomologue of BMP-4) acts as a potent dorsalizing mor-phogen, in keeping with the reversal in the dorsal toventral axis from invertebrates to vertebrates. All ofthese findings underscore the idea that proper regula-tion of BMP-4 signaling plays a crucial and highlyconserved role in embryonic patterning.
In Xenopus, recent studies have shown that a gradi-ent of BMP-4 signaling is maintained by the opposingaction of the organizer-specific factors noggin, chordinand follistatin. Noggin and chordin have both beenshown to directly bind mature BMP protein in theextracellular space, thus preventing interaction ofBMPs with their receptors (18-19). Follistatin has alsobeen shown to bind BMP in vitro (20). BMP-4 signalingis also regulated intracellularly through an antagonis-tic interaction with the dorsalizing TGF-b signalingmolecule activin (21).
There is some evidence that BMP-4 signalingmight also be controlled at the transcriptional level.Indeed, the spatial expression pattern of BMP-4 inthe gastrula stage Xenopus embryo (3) suggests thatsuch a mechanism is involved. It has been observedthat overexpression of BMP-4, or two related down-stream effectors results in the enhancement ofBMP-4 transcription suggesting the presence of anautoregulatory positive feedback loop (2, 22-23). Inaddition, recent studies in Xenopus and Zebrafishhave provided some evidence that the dorsalizingfactors, gsc, noggin and chordin, negatively regulateBMP-4 transcription (3, 24-25). All of these observa-tions support the idea that proper regulation ofBMP-4 transcription is an important factor in gen-erating and maintaining a dorsal-ventral gradient ofBMP-4 signaling.
Understanding the transcriptional regulation of anygene requires analysis of its promoter region. The Dro-sophila, mouse and human promoters for BMP-4 havebeen isolated (26-29). However, we still know littleabout the transcriptional regulation of BMP-4 in earlypatterning of vertebrate embryo. Furthermore, no in-formation has been reported concerning the XenopusBMP-4 promoter.
Here we demonstrate that expression of the domi-nant negative BMP-4 receptor or of the diffusible or-
ganizer signals, chordin and noggin, negatively regu-late BMP-4 transcription in the Xenopus animal capexplants. We suggest that repression of BMP-4 tran-scription by these endogenous factors is important inthe maintenance of dorsal fate. In addition, we reportthe isolation of the BMP-4 promoter region and iden-tify consensus binding sites for several transcriptionalregulators. In a functional assay we show that thesetranscriptional regulators can modulate BMP-4 pro-moter activity. The significance of these observationsto embryonic patterning is discussed.
MATERIALS AND METHODS
DNA and RNA preparation. CDNA clones encoding noggin, chor-din, BMP-4, DN-BR, 3m, b-galactosidase (b-gal), GATA-1, GATA-2,PV.1 and AP.1 have all been described elsewhere (30-33). Each of thecDNAs were linearized and used for in vitro synthesis of cappedmRNA using an Ambion transcription kit in accordance with themanufacturer’s instructions. The synthetic RNA was quantitated byethidium bromide staining in comparison with a standard RNA.
Embryo injection and explant culture. Xenopus laevis embryoswere obtained by in vitro fertilization (13). Developmental stageswere designated according to Nieuwkoop and Faber (34). Embryos atthe two-cell stage were injected in the animal pole with messengerRNA or cDNA as described in the figure legends. Animal caps weredissected from the injected embryos at stages 8.5-9 and cultured tovarious stages for further analysis as described in the figure legends.
Reverse transcription-polymerase chain reaction (RT-PCR). TotalRNA was extracted from cultured explants with TRIzol reagent (LifeTechnologies, Inc.) following the manufacturer’s instructions. RT-PCR was performed with a Superscript preamplification system (LifeTechnologies, Inc.). Primer sets and PCR conditions were as de-scribed previously (30).
FIG. 1. Negative transcriptional regulation of BMP-4 by DN-BR.RT-PCR analysis of BMP-4 transcripts in animal cap explants. An-imal caps injected with mRNA encoding b-galactosidase (1 ng),BMP-4 (1 ng), DN-BR (0.5 ng), Xlim-1 (1.0 ng), noggin (0.5 ng), andchordin (1 ng) were dissected at stage 8.5-9 and cultured until sib-lings reached stage 13. Total RNA was isolated from 10 animal capsand assayed for expression of BMP-4 by RT-PCR. EF-1a was used tonormalize between samples. No RT control reactions were performedwith total RNAs obtained from b-galactosidase-injected animal capsto confirm the absence of contaminating genomic DNA. Ectopic over-expression of any of these dorsalizing agents significantly repressedBMP-4 expression.
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
517

Isolation of clones encoding BMP-4 genomic DNA. Genomicclones for BMP-4 were isolated by screening a Xenopus lEMBL4genomic library (gift from Dr. Igor Dawid, 35, Rosa et al., 1988) witha PCR amplified cDNA probe which corresponds to the segment frombase pair 11 to 413 of Xenopus BMP-4 cDNA (Nishimatsu et al.1992). Approximately 7 3 106 clones were screened with a 32P-labeled probe or using ECL system in accordance with the manufac-turer’s (Amersham) instruction. Forty putative clones were isolatedand analyzed by PCR using the same primers (11-413 nt) used forprobe generation. Seven clones were selected, which produced adistinct band of about 1 kb on the gel. The 1 kb PCR product from theputative BMP-4 clones was analyzed by southern hybridization witha BMP-4 cDNA probe, and by digestion with the restriction enzymesEcoRI, BamHI, HindIII, XbaI, Not 1, XhoI or SmaI. Two clones werefound to be identical, and one of the clones was used for furtheranalysis. All the fragments generated by EcoRI were subcloned intopBluescript SK(2) (Stratagene). Four fragments, 5.2 Kb (clone B24),4.7 kb (B16), 1.3 kb (B19) and 1.9 kb (B5) were further analyzed bycomplete DNA sequence analysis after subcloning into pBluescriptSK(2). Both strands of DNA have been sequenced using automatedsequencing (accession number AF058764).
RACE-PCR cloning of 59 region of mRNA. The 59 rapid amplifi-cation of cDNA ends (RACE) method was conducted according to theprocedure described in the Marathon cDNA Amplification Kit (Clon-tech, Palo Alto). Briefly, 1 mg of poly A1RNA isolated from Xenopusembryo at stage 17 was used to prepare full length double-strand (ds)cDNA, which was then used to prepare a library of adaptor-ligated dscDNA. The cDNA library was amplified by polymerase chain reac-tion (PCR) using an adaptor-specific primer (AP1) supplied by themanufacturer and a gene specific primer (GSP). A second PCR reac-tion was then conducted using a nested (AP2) and primer and a genespecific (GSP) primer. The PCR program included a 1 min 94°Cdenaturation; 5 cycles of 94°C 30 s and 70°C 3 min; 5 cycles of 94°C30 s and 68°C 3 min; 30 cycles of 94°C 30 s and 66°C 3 min; with afinal extension at 68°C for 5 min. PCR products were analyzed byagarose gel electrophoresis, purified by glass milk extraction, clonedinto a TA cloning vector (Invitrogen) and sequenced by automatedsequencing. The antisense GSP used were designed from the re-ported cDNA sequence (36) R402 (nt number 402-425,59TATCGTGCAGTTCATCCTCCTCCTC-39), R295 (nt number 295-
323, 59-AACATCTGCAGCAGCGTCACCTC-39), and R189 (nt num-ber 18-226, 59-CAGGTATCAGGCTGGCATAGTTAGTG-39).
Plasmid constructs. The fragment (5.2 kb) from clone B24 con-taining 3.5 kb of 59-flanking region of N-terminal noncoding exonwas subcloned into pBluescript SK(2) (Stratagene, La Jolla, CA,USA). A HindIII site was created in the noncoding exon by PCR. TheEcoRI-HindIII fragment (which contains the region up to nt number3611) was subcloned into the luciferase basic plasmid (Promega,Madison, WI, USA, see Fig. 2A). The clone was designated BMP-4-luc.
Luciferase assays. The level of luciferase enzyme activity wasmeasured using Luciferase assay systems (Promega) according tomanufacturer’s instructions. Five to ten whole embryos for eachsample were harvested for luciferase activity assay. Transcriptionalactivity was assayed after injection of 40 pg plasmid DNA into theanimal region of two-cell embryos. Animal explants or marginalzones were dissected at stage 8.5-9.5 and cultured until the stagesdescribed in the figure legends. Samples were homogenized in 50 mlof lysis buffer per embryo and in 20 ml per explant. Luciferaseactivity was assayed with 200 ml of luciferase substrate with 50 ml or20 ml of whole embryo or explant lysate, respectively. All luciferaseactivity was assayed in triplicate.
RESULTS AND DISCUSSION
Negative transcriptional regulation of BMP-4 by DN-BR, Xlim-1, noggin, and chordin. Xenopus dorsaliz-ing factors function by binding BMP growth factor inthe extracellular space thereby inhibiting intracellu-lar BMP signaling. Dorsalizing factors might alsofunction to repress BMP-4 transcription in an effortto maintain the dorsal state. Conversely, active BMPtranscription may be required to maintain ventralfates. It has previously been observed that injectionof mRNA encoding BMP-4, or of mRNA encoding twoclosely related downstream transcription factors
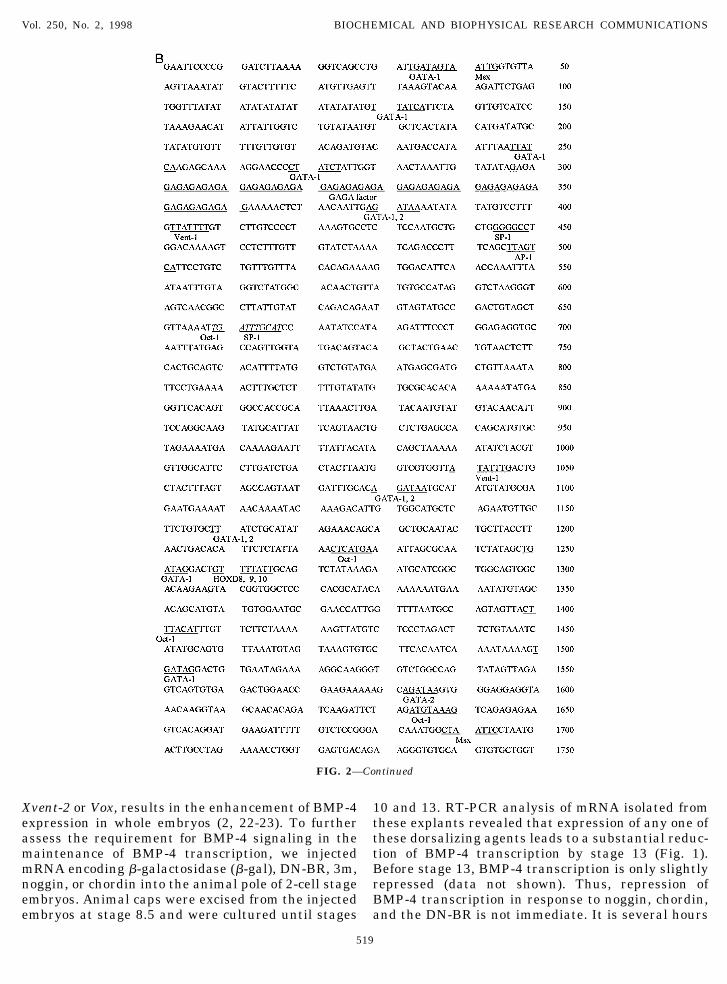
FIG. 2. Genomic organization of the Xenopus BMP-4 gene. (A) Schematic diagram of restriction enzyme map of 59 region of the BMP-4gene and over-all structure of genomic DNA. Numbers above the bar indicate the nucleotide sequence relative to 59-most end. 59 and39-flanking regions each have one repeated sequence, respectively. BMP-4 gene includes one noncoding exon, N-terminal exon with startcodon, and C-terminal exon with stop codon. (B) The BMP-4 genomic DNA sequence. Repeated sequences located in 59- and 39-flankingregions of the BMP-4 gene are indicated by italicized letters and the sequence for exons is shown as bold letters. Sequence motifs for theputative transcription binding sites are indicated as underlined letters.
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
518
Xvent-2 or Vox, results in the enhancement of BMP-4expression in whole embryos (2, 22-23). To furtherassess the requirement for BMP-4 signaling in themaintenance of BMP-4 transcription, we injectedmRNA encoding b-galactosidase (b-gal), DN-BR, 3m,noggin, or chordin into the animal pole of 2-cell stageembryos. Animal caps were excised from the injectedembryos at stage 8.5 and were cultured until stages
10 and 13. RT-PCR analysis of mRNA isolated fromthese explants revealed that expression of any one ofthese dorsalizing agents leads to a substantial reduc-tion of BMP-4 transcription by stage 13 (Fig. 1).Before stage 13, BMP-4 transcription is only slightlyrepressed (data not shown). Thus, repression ofBMP-4 transcription in response to noggin, chordin,and the DN-BR is not immediate. It is several hours
FIG. 2—Continued
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
519

after MBT before a decrease in BMP-4 expressionbecomes apparent.
As mentioned, several downstream transcription fac-tors have been shown to positively regulate BMP-4expression. Therefore, we conducted a stage-dependentanalysis of transcript levels of Xvent-1,-2 and GATA-2in DN-BR- and noggin-expressing cells. We reasonedthat the observed decrease in BMP-4 transcript levels
may occur as a result of removing these positive regu-lators. Animal caps were excised from stage 8.5 em-bryos expressing the DN-BR, noggin or b-gal. Injectedcaps were cultured until stages 11 and 13, and totalRNA was isolated for analysis by RT-PCR. As expected,we observed that expression of these transcription fac-tors was significantly decreased by stage 11 (notshown, 22-23). At this stage, no significant change in
FIG. 2—Continued
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
520

BMP-4 transcripts was observed. However by stage 13,BMP-4 transcription was depressed. This sequence ofevents supports the notion that in the presence ofXvent-1,-2 and GATA-2, BMP-4 expression is sus-tained. However, as their transcript levels decline,BMP-4 expression is suppressed. However, we can notrule out the involvement of other transcription factorsin the regulation of BMP-4 transcription.
Isolation and characterization of the Xenopus BMP-4gene. To further investigate the regulation of BMP-4transcription, we isolated its genomic clone. A 400 basepair fragment encoding the Xenopus BMP-4 cDNA (11-413nt, 36) was used to screen a Xenopus genomic li-brary. Approximately 73106 clones were screened andputative clones were isolated and analyzed by PCR andSouthern hybridization. Seven clones that produced
FIG. 2—Continued
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
521

distinct bands (expected size of 1 kb) on a gel andhybridized with the BMP-4 cDNA fragment (nt11-413)were selected. Two of the seven clones were furtheranalyzed by restriction enzyme digestion and southernhybridization to generate a physical map and to deter-mine the approximate position of the exons. Four frag-ments were generated by restriction enzyme digestionof the genomic clone with EcoR1. A 5.2 Kb fragment
(clone 24), a 4.7 kb fragment (B16), a 1.3 kb fragment(B19) and a 1.9 kb fragment (B5) were subcloned intopBluescript SK(2) (Stratagene), and were analyzed bysequence analysis (Gene bank accession numberAF058764). Figure 2A shows a schematic diagram ofthe genomic organization of the Xenopus BMP-4 gene.The structural organization of the Xenopus BMP-4gene is similar to both the Drosophila homologue and
FIG. 2—Continued
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
522

mammalian genes. The Xenopus BMP-4 gene includesone 59 non-coding exon, an N-terminal exon with astart codon and a C-terminal exon with a stop codon. Inaddition, we identified 2 putative Xenopus repeatedsequences (shown in Figs. 2A and 2B in italic) frombase pair 2096 to 2389 in the 59-flanking region and10276 to 10553 in the 39-flanking region. This putativerepeated sequence is present in many identified Xeno-
pus genes (37). The Xenopus BMP-4 protein sequencepredicted from genomic DNA sequences differs by oneamino acid from that of published (nt 7103 in Fig. 2)when compared with XBMP-4II (38) or XBMP-4.1 (1).Since the sequence change was confirmed in at least 2different readings in both directions, this differencemay represent a polymorphism.
We used the 59 RACE method to determine the
FIG. 2—Continued
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
523

location of the transcription initiation site. Reactionsfor BMP-4 were performed with adaptor-ligated dou-ble stranded (ds) cDNA made from stage 17 Xenopuspoly A1 RNA (see Materials and Methods). Theadaptor-ligated ds cDNA library was initially ampli-fied by pairing the adaptor primer (AP) with theBMP-4 specific antisense primer R402. Multiple PCRproducts were observed with the major product at
approximately 460 bp in size. A second PCR reactionusing a nested AP (AP2) and gene specific primers,R295 or R189, produced PCR products approxi-mately 350 or 250 base pairs respectively (data notshown). Sequence analysis of PCR products indicatethat the initiation site is spread over the region from228 to 1 1 (corresponding to nt 3547-3576 in Fig.2B) with the major site around 22 (Fig. 2A, in bold).
FIG. 2—Continued
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
524

The initiation site around this 59-noncoding exonwas somewhat unexpected since this exon containsclearly a predicted exon-boundary at nt 3576, andhuman and mouse BMP-4 genes use upstream exonsfor their initiation (27-29).
Many putative transcriptional regulatory elementswere identified within the 59 region of our clone (un-derlined in Fig. 2B). Interestingly, we identified mul-
tiple putative binding sites for the GATA-1, GATA-2,Xvent-1 and AP-1 transcriptional regulators. All ofthese are reported to be downstream effectors of theBMP-4 signaling pathway (39-42). Eight putativeGATA-1 binding sites were identified at positions 43,131, 247, 269, 1251, 1501, 2385, and 2782. Six moresites which potentially bind both GATA-1 and -2 werefound at positions 1081, 1169, 1780, 2181, 2353, and
FIG. 2—Continued
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
525
2451. Finally, a single GATA-2 binding site was ob-served at nucleotide position 1582.
The binding site for Xvent-1 was recently reportedby Friedle et al. (43). Six such sequences were ob-
served at positions 402, 1041, 2025, 2769, 2807 and3483. In addition we identified 5 putative AP-1 bind-ing sites at nucleotide positions 496, 2245, 2863,3191 and 3244.
FIG. 3. DN-BR suppresses BMP-4 promoter activity. The BMP-4 promoter reporter construct (BMP-4-luc, 40 pg) was co-injected with 1ng of b-galactosidase or 0.5 ng of DN-BR RNA into 2-cell stage embryos. Embryos were collected at stages 13, 15, and 20, and then luciferaseassays were carried out as described under Materials and Methods.
FIG. 2—Continued
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
526

GATA-1 and -2, Xvent-1 and AP-1 have all beenshown to be strongly induced or activated by BMP-4(39-42). In addition, each of these transcription factorshas been observed to be involved in mediating theventralizing effects of BMP-4 in the ectoderm and/ormesoderm. As mentioned, Xvent-2 and its closely re-lated protein Vox have been observed to be involved ina feedback loop positively regulating BMP-4 transcrip-tion (22-23). The presence of putative binding sites forthe transcription supports that certain downstreameffectors of BMP-4 may be involved in a positive feed-back loop regulating BMP-4 expression.
The effects of DN-BR and BMP-4 on BMP-4 promoteractivity. To further investigate the transcriptionalregulation of the BMP-4 promoter, we linked 3.6 Kb ofupstream sequence to the luciferase reporter gene(BMP-4-luc). From the above experiments shown inFig. 1 and from the reports of others, there appears tobe an autoregulatory loop enhancing BMP-4 expres-sion. Therefore, we investigated the effect of modulat-ing BMP-4 signaling levels on transcriptional activa-tion of the luciferase gene from the 3.6 Kb fragment ofthe BMP-4 promoter. MRNAs encoding DN-BR, BMP-4or b-gal were co-injected with the BMP-4/luciferasepromoter plasmids into the animal pole region of 2-cellstage embryos. Ten embryos were collected at differentstages (13, 15 and 20). Figure 3 shows the results of 1
of 3 representative experiments. At all stages exam-ined, DN-BR-injected embryos showed a significant de-crease in luciferase reporter activity. These observa-tions support the idea that BMP-4 promoter activity ismodulated by the level of BMP-4 signaling.
Dorsal–ventral modulation of BMP-4 promoter activ-ity. BMP-4 is expressed in the ventral mesoderm ofanimal pole gastrula stage embryos. BMP-4 tran-scripts are conspicuously absent from the dorsal meso-derm. To determine whether our BMP-4 promoter frag-ment contains elements which might contribute to theregulation of BMP-4 expression along the dorsal toventral axis, we injected 40 pg of BMP-4-luc into thetwo dorsal or two ventral cells of 4-cell stage embryos.Embryos were allowed to develop until stage 10 andventral marginal zones (VMZs) or dorsal marginalzones (DMZs) were explanted and cultured until stage13. At this time the explants were collected and as-sayed for luciferase activity. As shown in Fig. 4, lucif-erase activity in VMZ explants was approximately two-fold higher than that measured in DMZs suggestingthe presence of functional elements in the BMP-4 pro-moter which confer spatial selectivity.
As shown in Fig. 2B, putative binding sites wereidentified for AP-1, GATA-1 and -2 and Xvent-1. Wetherefore investigated the regulation of BMP-4 pro-moter activity by each of these transcription factors.
FIG. 4. Spatial regulation of BMP-4 promoter activity. BMP-4-luc (40 pg) was injected into the dorsal or ventral marginal zone of 4-cellstage embryos. Ventral or dorsal marginal zones were explanted at stage 10 and cultured until stage 13. Luciferase assays were performedas described under Materials and Methods.
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
527

Since the Xvent-1 clone was not available to us, weemployed the closely related homeobox gene PV.1.PV.1 is Xvent-1’s closest relative in both sequence andfunction. MRNAs encoding AP-1 (c-fos plus c-jun),GATA-1b, GATA-2, PV.1, Xvent-2 or b-gal. were coin-jected with the BMP/luc promoter plasmid into two-cellstage embryos. Injected embryos were allowed to de-velop until stage 11 at which time luciferase activitywas measured. The results of the assay are shown inFig. 5.
GATA-1b increased luciferase activity 3-fold overcontrol activity. We observed a 3.2-fold increase inBMP-4 promoter activity in the presence of GATA-2.PV.1 also significantly increased BMP-4 promoter ac-tivity (about 7-fold) as did Xvent-2 (2-fold). Coinjectionof mRNA encoding AP-1 resulted in a dramatic (17-fold) increase in luciferase activity relative to controls.These observations support that certain transcrip-tional regulators functioning downstream of BMP-4are involved in an autocatalytic loop positively regu-lating BMP-4 transcription. Thus, the gradual declineof BMP-4 transcription in response to dorsalizing sig-nals or the DN-BR is likely to occur, at least in part, asa result of the removal of positive regulators such asGATA-1,-2, PV.1, Xvent-2 and AP-1. However, it re-mains to be determined whether these transcription
factors regulate BMP-4 transcription directly or indi-rectly (through the activity of other transcriptionalregulators). Putative binding sites for several othertranscription factors were also identified. Binding sitesfor the ubiquitous transcription factors Oct1 and Sp1,for the erythroid-specific transcription factors CCAATand CACCC binding factors (44-45), for the long re-peated sequence GAGA, and for the essential region forthe murine BMP-4 promoter, the E-box (46) were iden-tified (Fig 2). However, we have not investigated thefunctional significance of these putative binding siteson the Xenopus promoter.
Since the maintenance of a gradient of BMP-4 sig-naling is crucial to the proper patterning of the earlyembryo, the transcriptional regulation of BMP-4 islikely to be very complicated. The Xenopus BMP-4 geneand its promoter will be a useful tool to further inves-tigate the transcriptional regulation of BMP-4 duringthe early stages of vertebrate development.
ACKNOWLEDGMENTS
We thank Annie Rogers for her editing and wordprocessing skills,Lynn Rasmussen for assistance in DNA sequencing, and TheresaKosack and James Wirth for their technical support.
FIG. 5. The effect of BMP-4 downstream transcription factors on BMP-4 promoter activity. The BMP-4 promoter reporter construct(BMP-4-luc, 40 pg) was co-injected with 1 ng of b-galactosidase, 0.5 ng of PV-1, 1 ng of GATA-1b, 1 ng of GATA-2, or 0.5 ng of c-Jun plus 0.5ng c-Fos RNA into fertilized eggs at 1-cell stage. Embryos were collected at stage 11 and luciferase assays were carried out as described underMaterials and Methods.
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
528

REFERENCES
1. Dale, L., Howes, G., Price, B. M., and Smith, J. C. (1992) Bonemorphogenetic protein 4: A ventralizing factor in early Xenopusdevelopment. Development 115, 573–585.
2. Jones, C. M., Lyons, K. M., Lapan, P. M., Wright, C. V., andHogan, B. L. (1992) DVR-4 (bone morphogenetic protein-4) as aposterior-ventralizing factor in Xenopus mesoderm induction.Development 115, 639–647.
3. Fainsod, A., Steinbeisser, H., and De Robertis, E. M. (1994) Onthe function of BMP-4 in patterning the marginal zone of theXenopus embryo. EMBO J. 13, 5015–5025.
4. Schmidt, J. E., Suzuki, A., Ueno, N., and Kimelman, D. (1995)Localized BMP-4 mediates dorsal/ventral patterning in the earlyXenopus embryo. Dev. Biol. 169, 37–50.
5. Suzuki, A., Thies, R. S., Yamaji, N., Song, J. J., Wozney, J. M.,Murakami, K., and Ueno, N. (1994) A truncated bone morpho-genetic protein receptor affects dorsal-ventralpatterning in theearly Xenopus embryo. Proc. Natl. Acad. Sci. USA 91, 10255–10259.
6. Maeno, M., Ong, R. C., Suzuki, A., Ueno, N., and Kung, H-F.(1994) A truncated bone morphogenetic protein 4 receptor altersthe fate of ventral mesoderm to dorsal mesoderm: Roles of ani-mal pole tissue in the development of ventral mesoderm. Proc.Natl. Acad. Sci. USA 91, 10260–10264.
7. Graff, J. M., Thies, R. S., Song, J. J., Celeste, A. J., and Melton,D. A. (1994) Studies with a Xenopus BMP receptor suggest thatventral mesoderm-inducing signals override dorsal signals invivo. Cell 79, 169–179.
8. Hawley, S. H. B., Winnenberg-Stapleton, K., Hashimoto, C.,Laurent, M. N., Watabe, T., Blumberg, B. W., and Cho, K. W. Y.(1995) Disruption of BMP signals in embryonic Xenopus ecto-derm leads to direct neural induction. Genes Dev. 9, 2923–2935.
9. Steinbeisser, H., Fainsod, A., Niehrs, C., Sasai, Y., and De Ro-bertis, E. M. (1995) The role of gsc and BMP-4 in dorsal-ventralpatterning of the marginal zone in Xenopus: A loss-of-functionstudy using antisense RNA. EMBO J. 14, 5230–5243.
10. Dosch, R., Gawantka, V., Delius, H., Blumenstock, C., and Nie-hrs, C. (1997) BMP-4 acts as a morphogen in dorsoventral me-soderm patterning in Xenopus. Development 124, 2325–2334.
11. Sasai, Y., Lu, B., Steinbeisser, H., and De Robertis, E. M. (1995)Regulation of neural induction by the Chd and BMP-4 antago-nistic patterning signals in Xenopus. Nature 376, 333–389.
12. Wilson, P. A., and Hemmati-Brivanlou, A. (1995) Induction ofepidermis and inhibition of neural fate by BMP-4. Nature 371,331–333.
13. Xu, R-H., Kim, J., Taira, M., Zhan, S., Sredni, D., and Kung, H-F.(1995) A dominant negative bone morphogenetic protein 4 recep-tor causes neuralization in Xenopus ectoderm. BBRC 212, 212–219.
14. Winnier, G., Blessing, M., Labosky, P. A., and Hogan, B. L. M.(1995) Bone morphogenetic protein-4 is required for mesodermformation and patterning in the mouse. Genes Dev. 9, 2105–2116.
15. Irish, V. F., and Gelbar, W. M. (1987) The decapentaplegic geneis required for dorsal-ventral patterning in the Drosophila em-bryo. Genes Dev. 1, 868–879.
16. Ferguson, E. L., and Anderson, K. V. (1992) Localized enhance-ment and repression of the activity of the TGF-b family member,decapentaplegic, is necessary for dorsal-ventral pattern forma-tion in the Drosophila embryo. Development 114, 583–597.
17. Wharton, K. A., Ray, R. P., and Gelbart, W. M. (1993) An activitygradient of decapentaplegic, is necessary for specification of dor-sal pattern elements in the Drosophila embryo. Development117, 807–822.
18. Zimmerman, L. B., De Jesus-Escobar, J. M., and Harland, R. M.(1996) The Spemann organizer signal noggin binds and inacti-vates bone morphogenetic protein 4. Cell 86, 599–606.
19. Piccolo, S., Sasai, Y., Lu, B., and De Robertis, E. M. (1996)Dorso-ventral patterning in Xenopus: Inhibition of ventral sig-nals by direct binding of chordin to BMP-4. Cell 84, 589–598.
20. Fainsod, A. H., Steinbeisser, and De Robertis, E. M. (1997) Thedorsalizing and neural inducing gene follistatin is an antagonistof BMP-4. Mech. Dev. 63, 39–50.
21. Candia, A. F., Watabe, T., Hawley, S. H., Onichtchouk, D.,Zhang, Y., Derynck, R., Niehrs, C., and Cho, K. W. (1997) Cel-lular interpretation of multiple TGF-beta signals: Intracellularantagonism between activin/BVg1 and BMP-2/4 signaling medi-ated by Smads. Development 124, 4467–4480.
22. Schmidt, J. E., von Dassow, G., and Kimelman, D. (1996) Regu-lation of dorsal-ventral patterning: The ventralizing effects ofthe novel Xenopus homeobox gene Vox. Development 122, 1711–1721.
23. Onichtchouk, D., Gawantka, V., Dosch, R., Delius, H., Hir-schfeld, K., Blumenstock, C., and Niehrs, C. (1996) The Xvent-2homeobox gene is part of the BMP-4 signalling pathway control-ling [correction of controling] dorsoventral patterning of Xeno-pus mesoderm. Development 122, 3045–3053.
24. Reem-Kalma, Y., Lamb, T., and Frank, D. (1995) Competitionbetween noggin and bone morphogenetic protein 4 activities mayregulate dorsalization during Xenopus development. Proc. Natl.Acad. Sci. USA 92, 12141–12145.
25. Schulte-Merker, S., Lee, K. J., McMahon, A. P., and Hammer-schmidt, M. (1997) The zebrafish organizer requires chordino.Nature 387, 862–863.
26. St Johnston, R. D., Hoffmann, F. M., Blackman, R. K., Segal, D.,Grimaila, R., Padgett, R. W., Irick, H. A., and Gelbart, W. M.(1990) Molecular organization of the decapentaplegic gene inDrosophila melanogaster. Genes Dev. 4, 1114–1127.
27. Kurihara, T., Kitamura, K., Takaoka, K., and Nakazato, H.(1993) Murine bone morphogenetic protein-4 gene: Existence ofmultiple promoters and exons for the 59-untranslated region.Biochem. Biophys. Res. Commun. 192, 1049–1056.
28. Feng, J. Q., Chen, D., Cooney, J. J., Tsai, M.-J., Harris, M. A.,Tsai, S. Y., Feng, M., Mundy, G. R., and Harris, S. W. (1995) Themouse bone morphogenetic protein-4 gene. J. Biol. Chem. 270,28364–28373.
29. van den Wijngaard, A., van Kraay, M., van Zoelen, E. J. J.,Olijve, W., and Boersma, C. J. C. (1996) Genomic organization ofthe human bone morphogenetic protein-4 gene: Molecular basisfor multiple transcripts. Biochem. Biophys. Res. Commun. 219,789–794.
30. Xu, R-H., Kim, J., Taira, M., Lin, J. J., Zhang, C. H., Sredni, D.,Evans, T., and Kung, H-F. (1997) Differential regulation of neu-rogenesis by the two Xenopus GATA-1 genes. Mol. Cell Biol. 17,436–443.
31. Rauscher, F. J., III, Voulalas, P. J., Franza, B. R., Jr., andCurran, T. (1988) Genes Dev. 2, 1687–1699.
32. Curran, T., Gordon, M. B., Rubino, K. L., and Sambucetti, L.(1987) Oncogene 2, 79–84.
33. Ault, K. T., Dirksen, M. L., and Jamrich, M. (1996) A novelhomeobox gene PV.1 mediates induction of ventral mesoderm inXenopus embryos. Proc. Natl. Acad. Sci. USA 93, 6415–6420.
34. Nieuwkoop, P. D., and Faber, J. (1967) Normal Table of Xenopuslaevis (Daudin), North-Holland, Amsterdam.
35. Rosa, F., Sargent, T. D., Rebbert, M. L., Michaels, G. S., Jamrich,M., Grunz, H., Jonas, E., Winkles, J. A., and Dawid, I. B. (1988)Accumulation and decay of DG42 gene products follow a gradientpattern during Xenopus embryogenesis. Dev. Biol. 129, 114–123.
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
529

36. Nishimatsu, S., Suzuki, A., Shoda, A., Murakami, K., and Ueno,N. (1992) Genes for bone morphogenetic proteins are differen-tially transcribed in early amphibian embryos. Biochem. Bio-phys. Res. Commun. 186, 1487–1495.
37. Riggs, C. D., and Taylor, J. H. (1987) Sequence organization anddevelopmentally regulated transcription of a family of repetitiveDNA sequences of Xenopus laevis. Nucleic Acids Res. 15, 9551–9565.
38. Koster, M., Plessow, S., Clement, J. H., Lorenz, A., Tiedemann,H., and Knochel, W. (1991) Bone morphogenetic protein 4 (BMP-4), a member of the TGF-beta family, in early embryos of Xeno-pus laevis: Analysis of mesoderm inducing activity. Mech. Dev.33, 191–199.
39. Maeno, M., Mead, P. E., Kelley, C., Xu, R. H., Kung, H. F.,Suzuki, A., Ueno, N., and Zon, L. I. (1996) The role of BMP-4 andGATA-2 in the induction and differentiation of hematopoieticmesoderm in Xenopus laevis. Blood 88, 1965–1972.
40. Zhang, C., and Evans, T. (1996) BMP-like signals are requiredafter the midblastula transition for blood cell development. Dev.Genet. 18, 267–278.
41. Gawantka, V., Delius, H., Hirschfeld, K., Blumenstock, C., andNiehrs, C. (1995) Antagonizing the Spemann organizer: Role ofthe homeobox gene Xvent-1. EMBO J. 14, 6268–6279.
42. Xu, R. H., Dong, Z., Maeno, M., Kim, J., Suzuki, A., Ueno, N.,Sredni, D., Colburn, N. H., and Kung, H. F. (1996) Involve-ment of Ras/Raf/AP-1 in BMP-4 signaling during Xenopusembryonic development. Proc. Natl. Acad. Sci. USA 93, 834 –838.
43. Friedle, H., Rastegar, S., Paul, H., Kaufmann, E., and Knochel,W. (1998) Xvent-1 mediates BMP-4-induced suppression of thedorsal-lip-specific early response gene XFD-1’ in Xenopus em-bryos. EMBO J. 17, 2298–2307.
44. Brewer, A. C., Guille, M. J., Fear, D. J., Partington, G. A., andPatient, R. K. (1995) Nuclear translocation of a maternal CCAATfactor at the start of gastrulation activates Xenopus GATA-2transcription. EMBO J. 14, 757–766.
45. Miller, I. J., and Bieker, J. J. (1993) A novel, erythroid cell-specific murine transcription factor that binds to the CACCCelement and is related to the Kruppel family of nuclear proteins.Mol. Cell Biol. 13, 2776–2786.
46. Ebara, S., Kawasaki, S., Nakamura, I., Tsutsumimoto, T., Na-kayama, K., Nikaido, T., and Takaoka, K. (1997) Transcriptionalregulation of the mBMP-4 gene through an E-box in the 59-flanking promoter region involving USF. Biochem. Biophys. Res.Commun. 240, 136–141.
Vol. 250, No. 2, 1998 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
530




















