
Downregulation and Nuclear Relocation of MLP During the Progression of Right Ventricular Hypertrophy...
11
J Mol Cell Cardiol 32, 2385–2395 (2000) doi:10.1006/jmcc.2000.1269, available online at http://www.idealibrary.com on Downregulation and Nuclear Relocation of MLP During the Progression of Right Ventricular Hypertrophy Induced by Chronic Pressure Overload Aline Ecarnot-Laubriet 1 , Karelle De Luca 1 , David Vandroux 1 , Maryvonne Moisant 2 , Colette Bernard 2 , Mahfoud Assem 1 , Luc Rochette 1 and Jean-Raymond Teyssier 1,3 1 Laboratoire de Physiopathologie et Pharmacologie Cardiovasculaires Expe ´rimentales, Faculte ´ de Me ´decine, Universite ´ de Bourgogne, Dijon, France, 2 Laboratoire d’Histologie, Faculte ´ de Me ´decine, Universite ´ de Bourgogne, Dijon, France, 3 Laboratoire de Ge ´ne ´tique Mole ´culaire, Faculte ´ de Me ´decine, Universite ´ de Bourgogne, Dijon, France (Received 31 July 2000, accepted in revised form 25 September 2000, published electronically 26 October 2000) A. E-L, K. D L, D. V, M. M, C. B, M. A, L. R J.-R. T . Downregulation and Nuclear Relocation of MLP During the Progression of Right Ventricular Hypertrophy Induced by Chronic Pressure Overload. Journal of Molecular and Cellular Cardiology (2000) 32, 2385–2395. The cardiac LIM domain protein MLP plays a crucial role in the architecture and mechanical function of cardiac myocytes. Mice lacking the MLP gene develop cardiac hypertrophy, dilated cardiopathy and heart failure. We investigated whether downregulation of MLP is induced by pressure overload and contributes to the physio- pathology of cardiac hypertrophy and failure. We studied this mechanism in rat right ventricles submitted to pulmonary arterial hypertension, because it is known that this ventricle is very vulnerable to the deleterious effects of pressure overload. During the progression of cardiac hypertrophy to failure over a 31 days period there was a dramatic decrease by 50% of the MLP transcripts level. Consistently, immunohistochemistry detected very weak protein signals in the cytoplasms of cardiomyocytes at the failing stage, but myocytes nuclei were heavily labeled. The nuclear relocation was confirmed by the immunodetection of MLP on the nuclear and cytosolic fractions. This nuclear localization is the hallmark of a retro-differentiated phenotype, since it has been observed only in differentiating myoblasts. These changes were associated with ultrastructural disorganization of the myofibrils similar to that observed in MLP −/− mice. Therefore, MLP dowregulation occurring during gene reprogramming may critically contribute to mechanical failure of the myocardium. 2000 Academic Press K W: Heart failure; Gene; LIM proteins; MLP; Pulmonary hypertension; Cor pulmone. in cell growth, cell death, and functional dif- Introduction ferentiation are either activated or depressed in the hypertrophic and failing heart. 2 How cumulative Sustained hemodynamic overload causes pro- gressive contractile dysfunction of the hypertrophic alterations of these systems contribute to the pro- gressive deterioration of the cardiac performance myocardium, which eventually ends with ir- reversible congestive heart failure. 1 Genes involved remains poorly understood. 3 The study of human Please address all correspondence to: J. R. Teyssier, Laboratoire de Ge ´ne ´tique Mole ´culaire, Faculte ´ de Me ´decine, 7 Boulevard Jeanne d’Arc, 21033, Dijon, France. 0022–2828/00/122385+11 $35.00/0 2000 Academic Press
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Downregulation and Nuclear Relocation of MLP During the Progression of Right Ventricular Hypertrophy...

J Mol Cell Cardiol 32, 2385–2395 (2000)
doi:10.1006/jmcc.2000.1269, available online at http://www.idealibrary.com on
Downregulation and Nuclear Relocationof MLP During the Progression of RightVentricular Hypertrophy Induced byChronic Pressure OverloadAline Ecarnot-Laubriet1, Karelle De Luca1, David Vandroux1,Maryvonne Moisant2, Colette Bernard2, Mahfoud Assem1, Luc Rochette1
and Jean-Raymond Teyssier1,3
1Laboratoire de Physiopathologie et Pharmacologie Cardiovasculaires Experimentales, Faculte deMedecine, Universite de Bourgogne, Dijon, France, 2Laboratoire d’Histologie, Faculte de Medecine,Universite de Bourgogne, Dijon, France, 3Laboratoire de Genetique Moleculaire, Faculte de Medecine,Universite de Bourgogne, Dijon, France
(Received 31 July 2000, accepted in revised form 25 September 2000, published electronically 26 October 2000)
A. E-L, K. D L, D. V, M. M, C. B, M. A, L. R J.-R.T. Downregulation and Nuclear Relocation of MLP During the Progression of Right Ventricular HypertrophyInduced by Chronic Pressure Overload. Journal of Molecular and Cellular Cardiology (2000) 32, 2385–2395. Thecardiac LIM domain protein MLP plays a crucial role in the architecture and mechanical function of cardiacmyocytes. Mice lacking the MLP gene develop cardiac hypertrophy, dilated cardiopathy and heart failure. Weinvestigated whether downregulation of MLP is induced by pressure overload and contributes to the physio-pathology of cardiac hypertrophy and failure. We studied this mechanism in rat right ventricles submitted topulmonary arterial hypertension, because it is known that this ventricle is very vulnerable to the deleteriouseffects of pressure overload. During the progression of cardiac hypertrophy to failure over a 31 days period therewas a dramatic decrease by 50% of the MLP transcripts level. Consistently, immunohistochemistry detected veryweak protein signals in the cytoplasms of cardiomyocytes at the failing stage, but myocytes nuclei were heavilylabeled. The nuclear relocation was confirmed by the immunodetection of MLP on the nuclear and cytosolicfractions. This nuclear localization is the hallmark of a retro-differentiated phenotype, since it has been observedonly in differentiating myoblasts. These changes were associated with ultrastructural disorganization of themyofibrils similar to that observed in MLP −/− mice. Therefore, MLP dowregulation occurring during genereprogramming may critically contribute to mechanical failure of the myocardium.
2000 Academic Press
K W: Heart failure; Gene; LIM proteins; MLP; Pulmonary hypertension; Cor pulmone.
in cell growth, cell death, and functional dif-Introductionferentiation are either activated or depressed in thehypertrophic and failing heart.2 How cumulativeSustained hemodynamic overload causes pro-
gressive contractile dysfunction of the hypertrophic alterations of these systems contribute to the pro-gressive deterioration of the cardiac performancemyocardium, which eventually ends with ir-
reversible congestive heart failure.1 Genes involved remains poorly understood.3 The study of human
Please address all correspondence to: J. R. Teyssier, Laboratoire de Genetique Moleculaire, Faculte de Medecine, 7 Boulevard Jeanned’Arc, 21033, Dijon, France.
0022–2828/00/122385+11 $35.00/0 2000 Academic Press

A. Ecarnot-Laubriet et al.2386
monogenic cardiomyopathies has provided evid- the stroke volume and output. We hypothesizedthat this sensitivity to high intracavity pressuresence that structural defects of the cytoskeleton
are intrinsic causes of myocardium hypertrophy, may be the consequence of chamber-specific mo-lecular defect in the mechanotransduction pathway,chamber dilatation and failure.4–6 However,
whether quantitative and qualitative alterations of because the compliant, thin, right ventricle wallcannot respond to stretch by sufficient tension con-these critical scaffold proteins are caused by ex-
trinsic, overload-dependent pathological conditions, trol. Supporting this hypothesis, our results showthat in the hypertrophic right ventricle myocardiumand whether they contribute to the transition to
congestive heart failure in the common form of reactivation of the fetal genetic program, down-regulation of MLP expression, nuclear relocationcardiopathies remain unknown. The knockout mice
model suggests that pathway involving the striated of this protein and structural alterations of themyofibrillar apparatus occurred concomitantly.muscle-specific LIM-only protein (MLP) plays a
crucial role in the compensatory hypertrophic re-sponse. 4–6 MLP promotes myogenic differentiation,is associated with the actin-based extrasarcomeric Methodscytoskeleton and is essential to the organization ofcardiomyocyte cytoarchitecture.7,8 MLP is a mem-
Experimental procedureber of the conserved LIM-only protein family, con-tains two LIM functional zinc-finger domains, and
Adult male Wistar rats (Depre Breedings, France)is exclusively expressed in muscle cells. The LIMweighing 310–320 g were given an intraperitonealfamily, which includes the LIM-only and LIM-hom-injection of 60 mg/kg of monocrotaline (Sigma-oebox proteins, are implicated in tissue pattering,Aldrich, France). Control rats were injected withgene regulation and cell differentiation. MLP, whichthe same volume (1 ml) of saline. At days 7, 14,is essential for the expression of the myogenic21, and 31 after injection matched series of treatedphenotype,7 accumulates during formation of stri-and control rats were anesthetized with 90 mg/kgated muscle fibers, is downregulated in adult skeletalof sodium pentobarbital.muscle tissue but is highly expressed in atrial and
ventricular cardiomyocytes. Deficient (MLP−/−)transgenic mice developed hypertrophy, dilated car-diopathy, and heart failure which mimic the human Determination of the right ventricular hypertrophyclinical picture and are associated with disarrayof myofibrils.9 Therefore, it is conceivable that in At each time point hearts and lungs of six control
and six MCT-treated rats were quickly removed,acquired hypertrophic cardiopathy changes in theMLP expression pattern induced by mechanical washed and weighed. The septum and the right
and left ventricle walls were dissected, weighed,stimuli (especially a downregulation resulting fromreactivation of the fetal gene program) may con- and frozen in liquid nitrogen until extraction of RNA
for the RT-PCR assay. The ratio of right ventricle totribute to irreversible cardiac failure by causingdisorganization of cardiomyocyte ultrastructure. To left ventricle plus septum weights (RV/LV+S) was
used as an index of the hypertrophy. The load-test this hypothesis we have analyzed expression ofMLP in a model of rat right ventricular hypertrophy induced hypertrophic phenotype was also sub-
stantiated by measuring the transcripts levels ofproduced by pulmonary hypertension, induced bythe pyrrolizidine alkaloid monocrotaline (MCT).10,11 gene encoding ANF and the � and � myosin heavy
chains (MHC). The right intraventricular pressuresThis model was chosen for the following reasons:(i) the slow developing monocrotaline-induced right were monitored in a parallel series of six treated
and control rats anesthetized by intraperitonealventricular hypertrophy reproduces the functionalchanges observed in the corresponding human injection of thiopental and artificially ventilated.
The left carotid artery was catheterized for mon-pathological condition chronic cor pulmonale, whichaffects 40% of the 70 000 patients who die each itoring the mean arterial pressure by a Mediserve
transducer (C. LGL, France) connected to a chartyear with chronic obstructive pulmonary diseasein the USA;12 (ii) nothing is known about the role recorder (Easy Graf TA 240, Gould, USA). After a
stabilization period of 10 min, the thorax wasof the above-mentioned molecular systems in thispressure-induced right ventricular pathology; (iii) opened and a heparin-filled needle, connected to
the pressure transducer, was inserted into the rightin contrast with the left ventricle, the right ventricleresponds to chronic pressure overload by massive ventricle chamber. Finally, histological changes in
the pulmonary and myocardial tissues have beendilatation responsible for a profound decrease in

MLP in Right Ventricle Hypertrophy 2387
observed after routine hematoxylin-eosin and Mas- dilution in 5 �l of PCR buffer (Eurobio, France),1 m of each dNTP, 200 ng of primers, with 2.5 Uson’s trichrome staining.of the Eurobiotaq Taq polymerase (Eurobio, France)and an adjusted volume of sterile water. After10 min denaturation at 95°C, 26 cycles of am-Immunohistochemistryplification were carried out on a PHC3 thermocycler(Techne, UK), followed by a final extension ofThe midregion of the hearts was cut perpendicular
to the major axis, fixed in buffered formalin, em- 10 min at 72°C. The cycling parameters were: de-naturation at 94°C, annealing at 60°C and ex-bedded in paraffin, then cut in 5 �m sections which
were labeled by the immunoperoxidase technique. tension at 72°C (each step for 1 min). Afteramplification, 15 �l of the PCR mixture was elec-The slides were incubated for 1 h at room tem-
perature with a 1:100 dilution of the specific anti- trophoresed in ethidium bromide stained agarosegel for quantification of the fluorescent bands byMLP rabbit antiserum to the unique C-terminal
peptide GGLTHQVEKKE (kindly provided by Dr Pico image analysis. Densitometric quantification offluorescent signals was performed by the imageCaroni, Friedrich Miescher Institute, Basel, Swit-
zerland).9 After washing the slides were exposed for analyzer Gel Doc1000 system driven by the Mo-lecular Analyst software (BioRad, France). The PCR30 min to a biotinylated secondary antibody (1:
400) (E432, Dako S.A., France). The fixed anti- amplification yield was expressed in arbitrary units(AU) as the ratio of the target/�-actin ampliconsbodies were exposed for 30 min to a 1:600 dilution
of a rabbit streptavidin-biotin-peroxidase complex optical densities. At each experimental time pointthe results obtained in control and MCT-treated rats(Dako S.A., France), and the peroxydase reaction
was developed using the AEC chromogen (AEC kit, was expressed as the mean±.. of 24 densitometricreadings recorded from four PCR products, sampledSigma, France). The sections were slightly stained
with hematoxylin, mounted and observed. In- during the exponential phase of two independentamplification reactions, carried out in the six rightcubation omitting the specific antibody and using
unrelated antibodies was used as control. ventricle and left ventricle samples. In a preliminaryexperiment the reliability of the �-actin internalstandard has been established by determining theratio of the 28 S RNA/PCR bands fluorescence in-Quantification of genetic transcripts by RT-PCRtensity.14 Triplicate PCR reactions were carried outon ten samples (five controls and five treated rats)The steady state level of the MLP, ANF, �-MHC and
�-MHC gene transcripts has been evaluated by a randomly chosen. These ratios varied from 0.80 to0.85 (6%) [see Fig. 2(E) in Results], demonstratingcomparative RT-PCR assay using a �-actin sequence
internal standard as previously described.13 The that the �-actin mRNA levels in the RV of controland MCT-treated rats were similar and proportionalsequences of the forward (F) and reverse (R) primers
were as follows: MLP (Genbank #X81193): (F) to the RNA input. The cDNA products were alsoused to check the linearity of amplification efficiencyCTCTGGACAGCACCACAGTG (nt 167–186), (R)
GTAAGCCCTCCAAACCCAAT (nt 580–599); �-ac- according to the number of cycles and the volumeof cDNA mixture. To avoid amplification of genomictin (Genbank #J00691): (F) TTGTAACCAAC-
TGGGACGATATGG (nt 224–247), (R) GATCTT- processed �-actin pseudogenes, which produce PCRfragments from contaminating DNA identical inGATCTTCATGGTGCTAGG (nt 964–988); ANF
(Genbank #M27498): (F) TATTGGAGCAAATC- size to the cDNA amplification products, total RNAsamples were pretreated with DNase I.15 The dataCCGTAT (nt 127–146), (R) TTTGGCTGTTATCTT-
CGGTA (nt 512–531); �-MHC (Genbank were compared by the variance analysis test (ANO-VA).#G56654): (F) ACCAAGCAGCCACGCCAGTA (nt
1382–1401), (R) TCCAGCCAGCCCAAGATGTT (nt1802–1821); �-MHC (Genbank #X15939): (F) AC-CAAGCAGCCACGCCAGTA (nt 1355–1374), (R) Western blot analysis of cytosolic and nuclear proteinsTGCTTTGCCTTTGCCCTTGT (nt 1905–2014). Thesizes of corresponding amplicons were: MLP: At each time point three right ventricles from MCT-
treated and matched control rats were dissected433 bp, ANF: 405 bp, �-actin: 764 bp, �-MHC:440 bp, and �-MHC: 569 bp. The specificity of the and the proteins were extracted from the cytosolic
and nuclear fractions exactly as described by Cal-PCR products has been checked by restriction map-ping. The PCR reaction was set up in 50 �l of a lejas et al.16 Thirty �g of the nuclear and cytosolic
proteins were electrophoresed in a 12.5% (w/v)reaction mixture consisting in 5 �l of a 1/5 cDNA

A. Ecarnot-Laubriet et al.2388
polyacrylamide gel and blotted onto Hybond-ECLmembrane (Amersham Pharmacia Biotech, France)using the Mini-Protean 3 and Mini Trans-Blot de-vices (BioRad, France). Following blockage with 5%non-fat milk the membranes were exposed for 1 hto a 1:1000 dilution of the anti-MLP antiserum.Then the membranes were incubated with a 1:2000 dilution of a goat anti-rabbit horseradishperoxydase labeled secondary antibody (P044,Dako SA, France). The immunocomplex was visu-alized by the chemoluminescence kit ECL (Amer-sham Pharmacia Biotech, France) and exposure toa Kodak X-ray film. Band densities were quantifiedby densitometric scanning (GS-670 imaging dens-itometer, BioRad, France). Results were expressedby the variations in % compared with matchedcontrols of the means of optical density values.
Transmission electron microscopy
At each time point tissue specimens from the rightventricular free wall of three control and threeMCT-treated rats were cut at midventricular level Figure 1 Parameters of hypertrophy. (A) Lung weights.into blocks 0.5 mm thick and 0.5 mm long. Tissue (B) Hypertrophic index: ratio of the RV to LV+S (septum)blocks were fixed in 2.5% (v/v) cacodylate-buffered weights. Values are mean±... (n=6). ∗ P<0.01 and
∗∗ P<0.001 v control.glutaraldehyde, postfixed in osmium tetroxide andembedded in epon. Ultrathin sections were stainedwith uranyl acetate and lead citrate and examinedusing a Hitachi 600 electron microscope. For each not be measured at day 31 because of lethalityheart at least 10 electron micrographs per section during the surgical procedure. At day 31 life-threat-were taken. Areas showing myofibrils were ex- ening congestive heart failure caused ascites andamined in greater detail. pleural fluid effusion. The anatomical hypertrophy
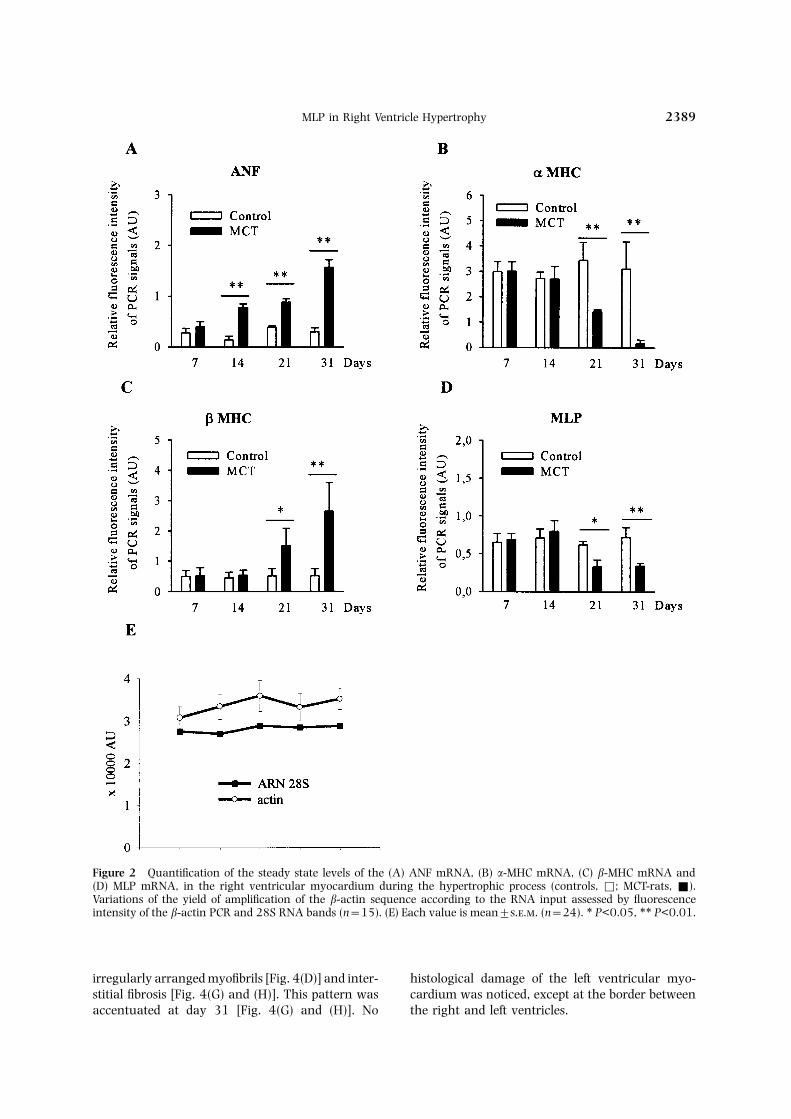
was associated with the characteristic reactivationof the fetal genetic program: there was a 200–400%increase in the content of ANF [Fig. 2(A)], and theratio of the �/� MHC RNAs levels dropped fromResults0.94 to 0.06 [Fig. 2(B) and (C)]. No change wasnoticed in the left ventricle, demonstrating that theDevelopment of right ventricular hypertrophypathological process was restricted to the rightchamber [Fig. 3(A)–(C)].The lung weight became significantly higher than
the control value by day 14 (56%) [Fig. 1(A)].Histological examination disclosed the well de-scribed pathological changes in the pulmonary vas-culature,11 including lesions of arteritis and Histopathologyocclusion of pulmonary arterioles and capillariesby fibrinous eosinophilic material. The RV/(LV+S) The right ventricular myocardium of control [Fig.
4(A), (B), (E) and (F)] and MCT-treated rats (notindex subsequently increased from day 21 to 31,indicating the development of right ventricular illustrated) showed a similar normal histology until
2 weeks after injection. Focal perivascular ed-hypertrophy in the MCT-treated rats [Fig. 1(B)].Consistently the intra-chamber pressures recorded ematous and fibrotic reactions were observed in the
right ventricle of MCT-treated from day 21. At thisat day 21 rose by 60% to 34.3(±3.6) mmHg (con-trol: 20.5±1.25, P<0.01). Pressure could time point the hypertrophic myocardium displayed
MLP in Right Ventricle Hypertrophy 2389
Figure 2 Quantification of the steady state levels of the (A) ANF mRNA, (B) �-MHC mRNA, (C) �-MHC mRNA and(D) MLP mRNA, in the right ventricular myocardium during the hypertrophic process (controls, Φ; MCT-rats, Ε).Variations of the yield of amplification of the �-actin sequence according to the RNA input assessed by fluorescenceintensity of the �-actin PCR and 28S RNA bands (n=15). (E) Each value is mean±... (n=24). ∗ P<0.05, ∗∗ P<0.01.
irregularly arranged myofibrils [Fig. 4(D)] and inter- histological damage of the left ventricular myo-cardium was noticed, except at the border betweenstitial fibrosis [Fig. 4(G) and (H)]. This pattern was
accentuated at day 31 [Fig. 4(G) and (H)]. No the right and left ventricles.

A. Ecarnot-Laubriet et al.2390
Figure 3 Quantification of the steady state levels of the (A) ANF mRNA, (B) �-MHC mRNA, (C) �-MHC mRNA and(D) MLP mRNA, in the left ventricular myocardium during the hypertrophic process (controls, Φ; MCT-rats, Ε).
Ultrastructure Changes in MLP expression
The electron microscopy study focused on the myo- A significant drop by 47% and 53% of the MLPmRNA levels was detected at days 21 and 31cytes fibrillar apparatus. There was no marked
modification of the right ventricular cyto- respectively [Fig. 2(D)]. It followed the onset of ANFoverexpression induced by the pressure overload,architecture until day 14. At days 21 and 31
focal disorganization and rupture of myofibrils were and occurred during hypertophic growth as evid-enced by the �/� MHC switch. No significant changeobserved. They were associated with disruption of
Z-lines and appearance of non-myofibrillar spaces in the amount of gene transcripts was recorded inthe LV [Fig. 3(D)]. The results of immuno-(Fig. 5). In contrast the cytoarchitecture of the left
ventricular myocardium remained unaltered. histochemistry were consistent with those of
Figure 4 Histology of the control and MCT-rats right ventricular myocardium stained by (A) to (D) hematoxylin-eosinand (E) and (H) Masson’s trichrome (interstitial fibrosis stained in blue). Tissue sections of the right ventricle of a controlrat showing (A) to (E) normal vascular areas and (B) and (F) myocardial fibers. Similar views of the right ventricle inMCT-treated rats at day 21 showing (C) perivascular fibrosis and (D) irregular orientation of myocardial fibers (arrow).(G) and (H) The myocardium of MCT-treated rats at day 31 stained by Masson’s trichrome and observed at highermagnification shows the extent of the interstitial fibrous reaction (arrow). Magnification: (A) to (F) ×400 and (G) and(H) ×1000.

MLP in Right Ventricle Hypertrophy 2391

A. Ecarnot-Laubriet et al.2392
[Fig. 6(E) and (F)]. The specificity of this nuclearMLP localization was checked by using differentsecondary antibodies alone, as indicated in theMethods section. No change was observed in the leftventricle. The reciprocal changes in the cytoplasmicand nuclear expression of MLP was demonstratedby the Western blot. At the onset of hypertrophicgrowth (day 14) the amount of cytoplasmic MLPincreased four-fold, although the mRNA level re-mained unchanged. At this time the nuclear frac-tion was not modified. In contrast, at days 21 and31 the amount of MLP decreased by 50% in thecytoplasm, while it increased nine and 16 times,respectively, in the nucleus (Fig. 7).
Discussion
Our results demonstrate for the first time that theextrinsic pressure overload stimulus downregulatesMLP expression and induces its nuclear relocationin the hypertrophied right ventricular myocytes.It is significant that the onset of the drop wasconcomitant with the switch of MHC isogenes,and was associated with the development of overtanatomical hypertrophy. In contrast it was notdetected at the early phase, when the rise in ANFmRNA level attested the activation of the pressure-dependent pathway. This asynchrony supports theexperimental evidence that the increased vent-ricular ANF expression is not a univocal marker ofhypertrophy, but is primarily a sensitive indicatorof the early tissue response to hemodynamic patho-logical stimuli.17 It is also consistent with the hypo-thesis that the changes in expression of the �/� MHCFigure 5 Transmission electron microscopy of the right
ventricular myocardium tissue from (A) a control rat and genes correlate with the degree of hypertrophy,(B) MCT-rats showing disarray and rupture of myofibrils and the severity and/or duration of the pressure(arrows) at (B) day 21 and (C) day 31. Magnification: overload.18 It has been previously established that in×20 000.
non-myogenic cells and in differentiating myogeniccells MLP is exclusively nuclear, but that in maturemyotubes and myocytes it is f-actin-associated.7,8RT-PCR. A similar homogenous intense cytoplasmic
striated labeling was detected in the right and left In both primary and adult ventricular car-diomyocytes it was detected in a cytosolic striatedventricular myocardium of control [Fig. 6(A)] and
MCT-treated rats at days 7 and 14 (not illustrated). pattern in the vicinity of the Z lines.9 It is part of alarge glycoprotein network connecting the motorThis immunostaining showed 2 �m-spaced broad
bands similar to those identified in MLP (+/+) unit to the cell membrane, nuclear envelope andextracellular matrix. MLP, which is not found inmice cardiomyocytes [Fig. 6(B)].10 At days 21 and
31 this pattern was conserved but the intensity of proliferating myoblasts, drastically and specificallyreduces the S fraction of myogenic cells in dif-the signal was dramatically weakened, patches of
myocardium being virtually devoid of immuno- ferentiation-promoting conditions, and efficientlyprevents the inhibition of cell cycle exit exerted bystaining [Fig. 6(C) and (D)]. Furthermore, whereas
the cellular labeling was exclusively cytoplasmic in the transforming growth factor �1.7 Therefore, theobservation that downregulation of MLP gene ex-control and MCT-treated rats at days 7–14, at days
21–31 about 30% of nuclei were heavily labeled pression was associated with cytosolic depletion

MLP in Right Ventricle Hypertrophy 2393
Figure 6 MLP immunohistochemistry. MLP labeling of the right ventricular myocardium of control rat showing (A)homogenous intense immunoreactivity and (B) a typical striated cytoplasmic pattern. Weak MLP expression in theright ventricular myocardium of MCT-rats at (C) day 21 and (D) day 31. Absence of cytoplasmic MLP in the rightventricular myocardium of MCT-rats at day 31, but (E) and (F) detection of strong nuclear signals in a cluster ofmyocytes. Magnification: (A), (C) and (D) ×400 and (B), (E) and (F) ×1000.
and nuclear relocation of the protein further sup- regulation and nuclear addressing of MLP resultingfrom re-expression of the predifferentiated pheno-ports the paradigm linking the hypertrophic growth
mechanisms to the reversion of the cardiac muscle- type, contributed to the transition of compensatedhypertrophy to heart failure by impairing the mech-specific differentiation program, the so-called pheno-
conversion.19 It can be assumed that down- anical properties of cadiomyocytes. In cardiac myo-

A. Ecarnot-Laubriet et al.2394
Figure 7 Western blot quantification of the nuclear and cytosolic content of MLP in the right ventricular myocardium.(A) Homogeneity of the amount of proteins in the four samples used for the Western blot (Coomassie blue staining).(B) Example of immunoblot illustrating the reciprocal changes in the cytosolic and nuclear content of MLP in the rightventricle (control: 0; MCT-treated rats: days 14, 21, and 31). Variation of immunoblot densitometric values in thecytosolic and nuclear fractions of the right ventricle of MCT-treated rats expressed as a fraction of the control (0). (C)Each histogram represents the mean of the values from three right ventricles.
cytes MLP is associated with the actin-based cyto- a dramatic reduction of the resistance to mechanicalstress. It is significant that the histological andskeleton and plays a crucial role as a scaffold
protein regulating the cellular response to mech- ultrastructural alterations of the MLP(−/−) myo-cardium including fibrosis, increase in non-myo-anical stress.4 Transgenic MLP (−/−) mice dis-
played a striking nearly 100% penetrant phenotype fibrillar space, and fibrillar disarray were similar tothose we observed. In the transgenic-deficient micecombining marked enlargement of the two cardiac
chambers, myocardial hypertrophy and wall thin- these tissue changes were already detectable innewborn hearts supporting the causal role of MLPning.9 Ventricular failure was attested by im-
pairment of contractility and relaxation, and by deletion in the initiation of the pathological process.However, in this model mice heterozygous for theelevation of the end-diastolic pressure. These patho-
logical changes mimicked the features of dilated disrupted MLP had no detectable phenotype undernormal physiological hemodynamic conditions.cardiopathy in both human syndromes and animal
models. In deleted cardiomyocytes the absence of This feature indicates that 50% of the MLP proteinefficiently prevents hypertrophy and dilatationMLP was associated with an irregular cell shape and

MLP in Right Ventricle Hypertrophy 2395
5. C JC, C KR. Complexity in simplicity: mono-under normal hemodynamic load. Conversely, ingenic disorders and complex cardiomyopathies. Jour model the 50% decrease in the cytosolic MLPClin Invest 1999; 103: 1483–1485.
content was likely to be a deleterious consequence 6. T JA, B KR, B NE. Etiologies ofof a molecular switch resulting from the reduced cardiomyopathy and heart failure. Evidence for a
final common pathway for disorders of the myo-capacity of the right ventricular myocardium tocardium. Nature Med 1999; 5: 266.buffer pressure-induced wall stress. It can be as-
7. A S, H G, C P. Muscle LIM Protein,sumed that MLP deficiency circularly worseneda novel essential regulator of myogenesis promotes
myocytes stretching and enhanced the ensuing myogenic differentiation. Cell 1994; 79: 213–221.hypertrophic signals, which contributed to pro- 8. A S, C P. Specificity of single LIM motifs
in targeting and LIM/LIM interactions in situ. Genesgressive chamber dilatation and irreversible mech-Dev 1996; 10: 289–300.anical failure. These data suggest that activation of
9. A S, H JJ, R J, H M, S G,a MLP-defective hypertrophic pathway representsB J, P JC, C KR, C P. MLP-
an intrinsic phenotypic flaw of the right ventricular deficient mice exhibited a disruption of cardiacmyocytes. It may be related to the known de- cytoarchitectural organization, dilated cardio-
myopathy, and heart failure. Cell 1997; 88: 393–leterious effects of pressure overload on the right403.ventricle, often responsible of clinically intractable
10. D SA, B L. Rat models of hypertension,failure in the cor pulmone syndrome.12 This hypo-cardiac hypertrophy and failure. Cardiovasc Res
thesis is supported by unpublished observation from 1998; 39: 89–105.our laboratory, showing that in rats with aortic 11. B L, M J, D A, S C. Cardiac
and vascular responses after monocrotaline-inducedbanding expression of MLP was not modified inhypertrophy in rats. J Cardiovasc Pharmacol 1998;the hypertrophic left ventricle, but that it was31: 108–115.remarkably decreased in the right ventricle affected
12. R VH, R PA, W JS, H JK,by the hemodynamic consequences of left ventricle B TM. The forgotten chamber: the importancedysfunction. The decrease in the MLP content of of the right ventricle. Catheter Cardiovasc Diag 1995;
35: 18–28.the left ventricles of human failing hearts recently13. A M, T JR, B M, T J,reported provided evidence that at the end stage of
L A, J A, D M, R L.mechanical failure downregulation of MLP alsoPattern of superoxide dismutase enzymatic activity
occurred in the left ventricular myocytes.20 How- and RNA changes in rat heart ventricles after myo-ever, the lack of change in the level of transcripts cardial infarction. Am J Pathol 1997; 151: 549–555.
14. K YK, Z Z, R J, R R, M B,and the absence of nuclear relocation of the proteinD B, L JS, O T, S-S M,observed by the authors indicated that even at thisJ ES, S D, W WH, F A, Bstage there was no full phenoconversion of the leftSE. Expression of mdr-1 in refractory lymphoma:
ventricular myocytes. Quantification by polymerase chain reaction andvalidation of the assay. Blood 1995; 86: 1515–1524.
15. S A, O’D SW, B ME, SG. An effective method of completely removing con-Acknowledgementstaminating genomic DNA from an RNA sample tobe used for PCR. Mol Biotech 1997; 8: 135–137.
A. Ecarnot-Laubriet was supported by grants from 16. C NA, C M, B L, M-S P.the Conseil Regional de Bourgogne and by the Requirement of nuclear kappaB for the constitutive
expression of nitric oxide synthase-2 and cyclo-Faculties of Medicine and Pharmacy of Dijon.oxygenase-2 in rat trophoblasts. J Cell Sci 1999;112: 3147–3155.
17. V KL, B T, F SM, LReferences LA. Hypertrophy, pathology, and molecular markers
of cardiac pathogenesis. Circ Res 1998; 82: 773–1. S PH. Signaling in myocardial hypertrophy. 778.
Life after calcineurin? Circ Res 1999; 84: 633–646. 18. S K, B KR, D L B D, L2. H JJ, G A, C KR. Molecular and cellu- AM, M JJ. Switches in cardiac muscle gene
lar biology of cardiac hypertrophy and failure. In: J. expression as a result of pressure and volume over-Breslow, J. Leiden, R. Rosenberg and C. Seidman load. Am J Physiol 1992; 262: R363–R369.(eds). Molecular Basis of Cardiovascular Disease. Phila- 19. A P, K J. Ventricular myocytes are notdelphia: W.B. Saunders Company, 1999: 211–250. terminally differentiated in the adult mammalian
3. M DL. Mechanisms and models in heart failure. heart. Circ Res 1998; 83: 1–14.A combinatorial approach. Circulation 1999; 100: 20. Z O, C P, B M. Decreased expression of999–1008. the cardiac LIM domain protein MLP in chronic
4. C KR. Stress pathways and heart failure. Cell human heart failure. Circulation 2000; 101: 2674–2677.1999; 98: 555–558.




















