
The Promoter Region of the Human PMCA1 Gene Mediates Transcriptional Downregulation by...
7
The Promoter Region of the Human PMCA1 Gene Mediates Transcriptional Downregulation by 1,25-Dihydroxyvitamin D 3 Paul Glendenning,* , ² ,1 Thomas Ratajczak,² , ‡ , § Richard L. Prince,* , ² , ‡ Nandor Garamszegi, ¶ and Emanuel E. Strehler ¶ *Department of Medicine and ‡Department of Pharmacology, University of Western Australia, Western Australia, Australia; §Department of Endocrinology and Diabetes, Sir Charles Gairdner Hospital, Perth, Western Australia, Australia; ¶ Department of Biochemistry and Molecular Biology, Mayo Foundation, Rochester, Minnesota; and ²West Australian Institute for Medical Research, Queen Elizabeth II Medical Centre, Nedlands, Western Australia 6009, Australia Received September 30, 2000 The gene for plasma membrane calcium pump isoform 1 (PMCA1) is expressed in calcium-trans- porting epithelia and bone mesenchymal cells and is upregulated to 1,25-(OH) 2 D 3 in those tissues. A can- didate sequence for a vitamin D response element (VDRE) is present within a 1.7-kb promoter region of the human PMCA1 (hPMCA1) gene. We studied hPMCA1 promoter activity in MDBK and ROS 17/2.8 cell lines as PMCA1 mRNA expression is upregulated by 1,25-(OH) 2 D 3 in both. Structural analysis of the putative hPMCA1 VDRE sequence was performed using mobility shift analysis (EMSA) and nuclear extracts from COS-1 cells expressing human VDR (hVDR) and RXRa (hRXRa). 1,25-(OH) 2 D 3 induced transrepression of the entire 1.7-kb hPMCA1 pro- moter and of one promoter deletion construct in ROS 17/2.8 cells but not MDBK cells when assayed by luciferase reporter gene assays. Three additional hPMCA1 promoter deletion constructs were unaf- fected by 1,25-(OH) 2 D 3 in either cell line. While hVDR and hRXRa were capable of complexing with a rat osteocalcin DR3 VDRE, EMSA analysis of the poten- tial VDRE from the hPMCA1 gene did not show in- teraction of either nuclear receptor. Our results in- dicate tissue-specific sensitivity of the promoter region of the hPMCA1 gene to direct transcriptional downregulation by 1,25-(OH) 2 D 3 and suggest that any positive regulatory VDRE must lie outside of the 1.7-kb core promoter. © 2000 Academic Press Key Words: plasma membrane calcium ATPase; pro- moter; calcium transport; calcitriol; osteoblast; kidney distal tubule. The plasma membrane calcium ATPase (PMCA) is an energy requiring, ATP-dependent, membrane-bound protein playing a major role in Ca 21 efflux. In mammals, four non-allelic genes code for PMCA isoforms 1– 4. Quantitatively, PMCA1 is by far the major isoform in most tissues, including osteoblasts and kidney (1– 4). The active vitamin D metabolite, 1,25-(OH) 2 D 3 en- hances transcellular calcium transport in tissues such as the kidney and proximal intestine (5, 6). 1,25- (OH) 2 D 3 influences calcium transport, at least in part, via upregulation of PMCA1 expression (7–11) and this regulation appears to occur by a genomic effect of 1,25- (OH) 2 D 3 on PMCA1 gene transcription (12). Transcrip- tional activation by 1,25-(OH) 2 D 3 classically involves vitamin D receptor (VDR) and retinoic acid receptor (RXR) interacting with cis-acting elements in the 59 region of target genes (13). Positive effects on tran- scription can be mediated by the heterodimeric com- plex interacting with a direct tandem repeat of two 6 bp half elements, separated by 3 nucleotides (DR3 VDRE). The human PMCA1 (hPMCA1) gene 59-flanking region revealed the presence of two alternative ter- minal exons termed exon 1 and exon 1* (14) suggest- ing the existence of alternative promoters in the gene. The putative promoter region of the hPMCA1 gene also displays multiple start sites for the initia- tion of transcription, the absence of a canonical TATA box element, and a high G 1 C content. In 1 To whom correspondence should be addressed at Department of Endocrinology and Metabolism, 7th Floor, Services and Teaching Block, Royal Adelaide Hospital, North Terrace, Adelaide, South Aus- tralia 5000, Australia. Fax: 618 8222 5908. E-mail: pglenden@mail. rah.sa.gov.au. Biochemical and Biophysical Research Communications 277, 722–728 (2000) doi:10.1006/bbrc.2000.3745, available online at http://www.idealibrary.com on 722 0006-291X/00 $35.00 Copyright © 2000 by Academic Press All rights of reproduction in any form reserved.
Transcript of The Promoter Region of the Human PMCA1 Gene Mediates Transcriptional Downregulation by...

TMb
PN*AA†N
R
ipud(ohcbpue(tmRlhfaottdrda1
EBtr
Biochemical and Biophysical Research Communications 277, 722–728 (2000)
doi:10.1006/bbrc.2000.3745, available online at http://www.idealibrary.com on
0CA
he Promoter Region of the Human PMCA1 Geneediates Transcriptional Downregulation
y 1,25-Dihydroxyvitamin D3
aul Glendenning,*,†,1 Thomas Ratajczak,†,‡,§ Richard L. Prince,*,†,‡andor Garamszegi,¶ and Emanuel E. Strehler¶
Department of Medicine and ‡Department of Pharmacology, University of Western Australia, Western Australia,ustralia; §Department of Endocrinology and Diabetes, Sir Charles Gairdner Hospital, Perth, Western Australia,ustralia; ¶Department of Biochemistry and Molecular Biology, Mayo Foundation, Rochester, Minnesota; andWest Australian Institute for Medical Research, Queen Elizabeth II Medical Centre,edlands, Western Australia 6009, Australia
eceived September 30, 2000
md
apfQm
ha(vr(tv(rspbV
rmiggtT
The gene for plasma membrane calcium pumpsoform 1 (PMCA1) is expressed in calcium-trans-orting epithelia and bone mesenchymal cells and ispregulated to 1,25-(OH)2D3 in those tissues. A can-idate sequence for a vitamin D response elementVDRE) is present within a 1.7-kb promoter regionf the human PMCA1 (hPMCA1) gene. We studiedPMCA1 promoter activity in MDBK and ROS 17/2.8ell lines as PMCA1 mRNA expression is upregulatedy 1,25-(OH)2D3 in both. Structural analysis of theutative hPMCA1 VDRE sequence was performedsing mobility shift analysis (EMSA) and nuclearxtracts from COS-1 cells expressing human VDRhVDR) and RXRa (hRXRa). 1,25-(OH)2D3 inducedransrepression of the entire 1.7-kb hPMCA1 pro-oter and of one promoter deletion construct inOS 17/2.8 cells but not MDBK cells when assayed by
uciferase reporter gene assays. Three additionalPMCA1 promoter deletion constructs were unaf-
ected by 1,25-(OH)2D3 in either cell line. While hVDRnd hRXRa were capable of complexing with a ratsteocalcin DR3 VDRE, EMSA analysis of the poten-ial VDRE from the hPMCA1 gene did not show in-eraction of either nuclear receptor. Our results in-icate tissue-specific sensitivity of the promoteregion of the hPMCA1 gene to direct transcriptionalownregulation by 1,25-(OH)2D3 and suggest thatny positive regulatory VDRE must lie outside of the.7-kb core promoter. © 2000 Academic Press
1 To whom correspondence should be addressed at Department ofndocrinology and Metabolism, 7th Floor, Services and Teachinglock, Royal Adelaide Hospital, North Terrace, Adelaide, South Aus-
ralia 5000, Australia. Fax: 618 8222 5908. E-mail: [email protected].
722006-291X/00 $35.00opyright © 2000 by Academic Pressll rights of reproduction in any form reserved.
Key Words: plasma membrane calcium ATPase; pro-oter; calcium transport; calcitriol; osteoblast; kidney
istal tubule.
The plasma membrane calcium ATPase (PMCA) isn energy requiring, ATP-dependent, membrane-boundrotein playing a major role in Ca21 efflux. In mammals,our non-allelic genes code for PMCA isoforms 1–4.uantitatively, PMCA1 is by far the major isoform inost tissues, including osteoblasts and kidney (1–4).The active vitamin D metabolite, 1,25-(OH)2D3 en-
ances transcellular calcium transport in tissues suchs the kidney and proximal intestine (5, 6). 1,25-OH)2D3 influences calcium transport, at least in part,ia upregulation of PMCA1 expression (7–11) and thisegulation appears to occur by a genomic effect of 1,25-OH)2D3 on PMCA1 gene transcription (12). Transcrip-ional activation by 1,25-(OH)2D3 classically involvesitamin D receptor (VDR) and retinoic acid receptorRXR) interacting with cis-acting elements in the 59egion of target genes (13). Positive effects on tran-cription can be mediated by the heterodimeric com-lex interacting with a direct tandem repeat of two 6p half elements, separated by 3 nucleotides (DR3DRE).The human PMCA1 (hPMCA1) gene 59-flanking
egion revealed the presence of two alternative ter-inal exons termed exon 1 and exon 1* (14) suggest-
ng the existence of alternative promoters in theene. The putative promoter region of the hPMCA1ene also displays multiple start sites for the initia-ion of transcription, the absence of a canonicalATA box element, and a high G 1 C content. In

aDhqD
mvifaDpmpPcfbwms
M
to(bmliocae
vphwtu
Nuclear protein extracts were prepared according to previously de-s
uithB(mFwsimaWDseca
tft(ttpap(
lbacoa
R
1
wpt1mpsfRcwitt
m
Rp
lss
Vol. 277, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ddition, a sequence resembling a potential vitaminregulatory element (VDRE) is found in the
PMCA1 gene between exons 1 and 1*. This se-uence (AGGCCA CCC GGTGCA) is comparable to aR3 VDRE (11, 15).We have recently shown that PMCA1 steady-stateRNA expression is upregulated in Madin–Darby bo-
ine kidney (MDBK) cells, a cell line that is phenotyp-cally similar to the distal kidney tubule epithelial cellrom which it was derived (16). ROS 17/2.8 cells werelso chosen for study as this cell line expresses vitaminreceptor (17) and PMCA1 (18, 19) and has been used
reviously to define transcriptional response of vita-in D responsive genes (20). Based on our data and
revious evidence that 1,25-(OH)2D3 may increaseMCA1 transcription in the intestine (12), we pro-eeded to examine the promoter activity of a 1.7 kbragment of the 59 region of the human PMCA1 gene inoth cell lines to determine if the effect of 1,25-(OH)2D3
as transcriptionally mediated by this promoter frag-ent and dependent on the putative VDRE found up-
tream of exon 1*.
ATERIALS AND METHODS
Cell culture. MDBK and COS-1 kidney cells were obtained fromhe American Type Culture Collection (Rockville, MD). ROS 17/2.8steoblast-like cells were a kind gift from Professor T. J. MartinMelbourne, Australia). All three cell lines were propagated in Dul-ecco’s modified Eagle’s medium, nonessential amino acids, 2.5g/ml gentamycin, 10% (v/v) FCS and 2.0 mM L-glutamine. Cell
ines were maintained at 37°C in a humidified atmosphere contain-ng 5% CO2. The effect of 1,25-(OH)2D3 was studied by the incubationf confluent cells in albumin-enriched, fetal calf serum-free mediumontaining varying concentrations of 1,25-(OH)2D3 for 24 h prior tossay. Each 1,25-(OH)2D3 dose was performed in triplicate and eachxperiment repeated at least twice.
Transfection of human VDR and RXR nuclear receptor expressionectors into COS-1 cells and preparation of nuclear extracts. Ex-ression plasmids pFLAG-CMV2 and pRc/CMV, for hVDR andRXRa, respectively, together with corresponding empty vectors,ere gifts from Professor J. Eisman (Garvan Institute, Sydney, Aus-
ralia) and were transfected into COS-1 cells by electroporationsing a Bio-Rad Gene Pulser apparatus at 450 V and 250 mF.
TABLE 1
Nucleotide Sequence of the Putative hPMCA1 VDRE andat Osteocalcin VDRE Oligonucleotides Used in Electro-horetic Mobility Shift Assay
hPMCA1 putative VDRE oligonucleotide sequence59-TTGC AGGCCA CCC GGTGCA GACGGGG-39
Rat osteocalcin VDRE sequence59-CTGCACT GGGTGA ATG AGGACA TTAC-39
Note. Three base 59 and 39 overhangs were constructed to enableabeling of each oligonucleotide using [a-32P]dCTP. The 59 and 39equences for the rat osteocalcin and putative hPMCA1 VDREs arehown in bold for ease of recognition.
723
cribed methodology (21).
Electrophoretic mobility shift assay. Oligonucleotides were man-factured by Geneworks (Adelaide, South Australia) and are shown
n Table 1. Approximately 3 pmol of double-stranded DNA probe tohe rat osteocalcin VDRE (nucleotide positions 2468 to 2447) orPMCA1 putative VDRE (nucleotide positions 1029 to 1054, Gen-ank Accession No. L14561) were end-labeled with [a-32P]dCTP
Geneworks, Adelaide, South Australia) using Klenow DNA poly-erase. DNA binding reactions (10 mM Hepes, 25 mM KCl, 2%icoll, 2% glycerol, 1 mg/ml poly(dI:dC), 0.4% bovine serum albumin)ere performed by incubation at room temperature for 30 min and
eparated on a 4% polyacrylamide gel in 13 TBE. Supershift exper-ments were performed by preincubating the nuclear proteins with 1g of monoclonal antibodies for 30 min on ice. Monoclonal VDRntibody F2A10 was a kind gift from Dr. H. DeLuca (University ofisconsin, Madison, WI) and the monoclonal RXR antibody 4RX-12, which cross-reacts with all three RXR isoforms, was kindly
upplied by Dr. P. Chambon (IGBMC, Illkirch, France). Competitionxperiments were performed by incubating fold excess of unlabelledompetitor DNA with the nuclear extracts for 10 min prior to theddition of labeled probe.
Transient transfection of promoter chimeric constructs. The en-ire 1.7-kb BamHI fragment (14) and various PCR-generated sub-ragments of the hPMCA1 promoter (Fig. 1) were cloned in front ofhe luciferase reporter gene in pGL2-Basic or pGL3-Basic vectorsPromega, Darra, Queensland). MDBK and ROS 17/2.8 cells wereransfected using FuGENE 6 transfection reagent (Roche Diagnos-ics, West Perth, WA) with 300–900 ng of the appropriate hPMCA1romoter-reporter gene vector and 100 ng of pcDNA3.1/CAT vectors control for transfection efficiency. Cleared cell lysates were pre-ared 48 h after transfection and 24 h after treatment with 1,25-OH)2D3.
Transcriptional activity of the hPMCA1 promoter was assessed byuciferase assay of cellular supernatant and quantitation by-counter. Following heat inactivation of endogenous deacetylasectivity, cell lysate was quantitated by b-counter for chlorampheni-ol acetyltransferase (CAT) activity. Using the absolute count valuesf the CAT assay for each sample, luciferase values were appropri-tely adjusted to give a luciferase/CAT ratio for each sample.
ESULTS
,25-(OH)2D3 Effects on hPMCA1 Promoter Activityin MDBK and ROS 17/2.8 Cells
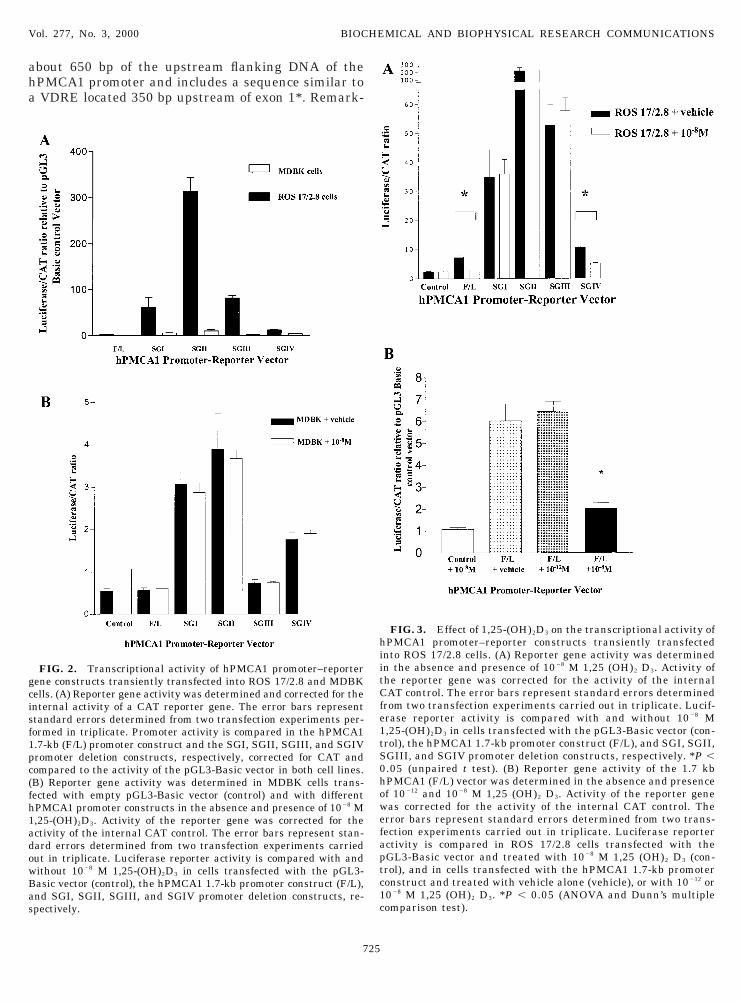
The hPMCA1 promoter segments shown in Fig. 1ere cloned in front of a luciferase reporter gene inGL2-Basic or pGL3-Basic vectors and transientlyransfected into ROS 17/2.8 and MDBK cells. In ROS7/2.8 cells, these constructs demonstrated basal pro-oter activity that was highly variable (Fig. 2A). The
romoter fragment SGII, corresponding to exon 1,howed the highest promoter activity, being up to 300-old more active than the pGL3-basic control vector.emarkably different results were obtained in MDBKells. Minimal promoter activity was demonstrableith all five constructs with luciferase values approx-
mately 2- to 3-fold higher for constructs SGI and SGIIhan values obtained with equivalent quantities of con-rol plasmid (Fig. 2A).
Interestingly, the 1.7-kb hPMCA1 promoter frag-ent demonstrated transrepression following 1,25-

(tdtf
tctt
E
cinptaacVbond
aw
ecttpRa
fi
mtu5tVcweoao
D
tt1p
resp
Vol. 277, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
OH)2D3 treatment in ROS 17/2.8 cells (Fig. 3A), andhis effect was dose-dependent (Fig. 3B). One promotereletion construct (SGIV), corresponding to exon 1* ofhe hPMCA1 gene, also demonstrated transrepressionollowing 1,25-(OH)2D3 treatment.
By contrast, 1,25-(OH)2D3 treatment of MDBK cells,ransiently transfected with all five hPMCA1 promoteronstructs in the pGL3-Basic plasmid, had no inhibi-ory effect on the already low basal promoter activity inhese cells (Fig. 2B).
MSA Analysis Using the Rat Osteocalcin andPutative hPMCA1 VDREs
Both hVDR and hRXRa nuclear receptors were suc-essfully overexpressed from their respective plasmidsn COS-1 cells which display no demonstrable endoge-ous expression of either VDR or RXR nuclear receptorrotein (data not shown). Using nuclear extracts fromransfected COS-1 cells and gel shift analysis, hVDRnd hRXRa were shown to complex as a heterodimernd bind to the osteocalcin VDRE (Fig. 4A). A mono-lonal antibody to RXR (4RX-D12) supershifted theDR/RXRa complex (Fig. 4A). The monoclonal anti-ody to VDR (F2A10) appeared to inhibit the formationf the hVDR/RXRa heterocomplex, whilst the monoclo-al antibody to estrogen receptor a (control) had noemonstrable effect (Fig. 4A).Studies using the putative hPMCA1 VDRE were un-
ble to demonstrate any difference in gel shift patternith nuclear extract from COS-1 cells transfected with
FIG. 1. Schematic representation of the hPMCA1 promoter andepresented diagrammatically on top and the 1.7-kb BamHI fragmenlarged below with the positions of the putative VDRE and of Sp1ubfragments SGI (nucleotides 1–639), SGII (640–1002), SG III (9GL2-Basic and pGL3-Basic luciferase reporter gene vectors are sho
724
mpty vector, compared to COS-1 cell nuclear extractsontaining hVDR and hRXRa (Fig. 4B). Varying pro-ein and salt concentrations, incubation period andemperature did not change the result. In addition,reincubation with antibodies to VDR (F2A10) andXR (4RX-D12) or 1028 M 1,25-(OH)2 D3 also did notlter the gel shift pattern (data not shown).Finally, competition assays were performed using a
xed amount of hVDR and hRXRa nuclear protein,32P-labeled rat osteocalcin VDRE and titration experi-
ents using increasing amounts of unlabeled rat os-eocalcin VDRE oligonucleotide (Fig. 4C, lanes 1–4) ornlabeled hPMCA1 oligonucleotide (Fig. 4C, lanes–8). Specificity of the observed protein-DNA interac-ion of hVDR and hRXRa with the rat osteocalcinDRE sequence was supported by results showingompetition of hVDR/RXRa heterocomplex formationhen unlabeled probe was used as competitor. Inter-stingly, and in contrast to the results with the cognatesteocalcin probe, unlabeled hPMCA1 oligonucleotidected to stabilize the hVDR/RXRa complex with thesteocalcin VDRE.
ISCUSSION
This study demonstrates that the 1.7 kb region andhe SGIV subfragment of the hPMCA1 promoter areransrepressed in ROS 17/2.8 osteoblast cells following,25-(OH)2 D3 treatment. The 1.7-kb fragment encom-asses exons 1 and 1* of the hPMCA1 gene as well as
omoter deletion constructs. The 59 portion of the hPMCA1 gene iscontaining the putative promoter and exon 1 and exon 1* is shown
CAAT sequences indicated symbolically. The 1.7-kb fragment and–1379), and SG IV (1370–1616) cloned separately in front of theschematically on the bottom.
prntand93wn
aha
gcisf1pc(fh1adowBas
hiitCfe1tS0howefaptc1c
Vol. 277, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
bout 650 bp of the upstream flanking DNA of thePMCA1 promoter and includes a sequence similar toVDRE located 350 bp upstream of exon 1*. Remark-
FIG. 2. Transcriptional activity of hPMCA1 promoter–reporterene constructs transiently transfected into ROS 17/2.8 and MDBKells. (A) Reporter gene activity was determined and corrected for thenternal activity of a CAT reporter gene. The error bars representtandard errors determined from two transfection experiments per-ormed in triplicate. Promoter activity is compared in the hPMCA1.7-kb (F/L) promoter construct and the SGI, SGII, SGIII, and SGIVromoter deletion constructs, respectively, corrected for CAT andompared to the activity of the pGL3-Basic vector in both cell lines.B) Reporter gene activity was determined in MDBK cells trans-ected with empty pGL3-Basic vector (control) and with differentPMCA1 promoter constructs in the absence and presence of 1028 M,25-(OH)2D3. Activity of the reporter gene was corrected for thectivity of the internal CAT control. The error bars represent stan-ard errors determined from two transfection experiments carriedut in triplicate. Luciferase reporter activity is compared with andithout 1028 M 1,25-(OH)2D3 in cells transfected with the pGL3-asic vector (control), the hPMCA1 1.7-kb promoter construct (F/L),nd SGI, SGII, SGIII, and SGIV promoter deletion constructs, re-pectively.
725
FIG. 3. Effect of 1,25-(OH)2D3 on the transcriptional activity ofPMCA1 promoter–reporter constructs transiently transfected
nto ROS 17/2.8 cells. (A) Reporter gene activity was determinedn the absence and presence of 1028 M 1,25 (OH)2 D3. Activity ofhe reporter gene was corrected for the activity of the internalAT control. The error bars represent standard errors determined
rom two transfection experiments carried out in triplicate. Lucif-rase reporter activity is compared with and without 1028 M,25-(OH)2D3 in cells transfected with the pGL3-Basic vector (con-rol), the hPMCA1 1.7-kb promoter construct (F/L), and SGI, SGII,GIII, and SGIV promoter deletion constructs, respectively. *P ,.05 (unpaired t test). (B) Reporter gene activity of the 1.7 kbPMCA1 (F/L) vector was determined in the absence and presencef 10212 and 1028 M 1,25 (OH)2 D3. Activity of the reporter geneas corrected for the activity of the internal CAT control. Therror bars represent standard errors determined from two trans-ection experiments carried out in triplicate. Luciferase reporterctivity is compared in ROS 17/2.8 cells transfected with theGL3-Basic vector and treated with 1028 M 1,25 (OH)2 D3 (con-rol), and in cells transfected with the hPMCA1 1.7-kb promoteronstruct and treated with vehicle alone (vehicle), or with 10212 or028 M 1,25 (OH)2 D3. *P , 0.05 (ANOVA and Dunn’s multipleomparison test).

abiiptnp
d1stoi
c(iciD2
tmtqtai
oh3mterchpcccm
Vol. 277, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
bly, the 1.7-kb promoter segment showed a very lowasal activity in reporter gene assays. By contrast,ndividual subfragments, especially that correspond-ng to exon 1 (SGII), showed dramatically increasedromoter activity. These findings suggest a multipar-ite nature of the hPMCA1 promoter where strongegative regulatory elements are interspersed withositive control regions.Our studies also indicate a strong cell-type depen-
ence of the hPMCA1 promoter. In contrast to the ROS7/2.8 osteoblast cell line, MDBK cells were unable toustain the high reporter gene activity observed withhe SGII hPMCA1 promoter fragment. Cotransfectionf the constitutively active CAT reporter plasmid ver-fied high efficiency of transfection. Since the MDBK
FIG. 4. Effect of ectopically expressed hVDR and hRXRa on thsteocalcin VDRE was end-labeled with [a-32P]dCTP and incubated wVDR and hRXRa. Nuclear extracts were preincubated without anti), and VDR (lane 4) for 30 minutes on ice. The DNA binding reactixtures were resolved on a 4% polyacrylamide gel in 13 TBE buffer
he free probe are indicated by arrows on the right. (B) The rat osteocnd-labeled with [a-32P]dCTP and incubated with COS-1 nuclear extecombinant nuclear receptors (lane 2). The incubation mixtures womplexes were noted with the putative hPMCA1 VDRE sequence aPMCA1 VDRE sequence did not change when COS-1 nuclear extraattern produced with COS-1 nuclear extracts containing transientlomplex formation on [a-32P]dCTP end-labeled rat osteocalcin VDRompeting unlabeled rat osteocalcin VDRE oligonucleotides (lanes 1oncentrations of 0-, 25-, 50-, and 100-fold excess of each unlabelled sixtures were resolved on a 4% polyacrylamide gel in 13 TBE buff
726
ell line is capable of responding appropriately to 1,25-OH)2D3 using other reporter genes (22), the relativenactivity of the hPMCA1 promoter constructs in thisell line may be surprising. It is notable however, thatn the study of the promoter for the mouse calbindin28K gene, basal expression was highest in ROS 17/.8 cells compared to two other cell lines (20).The significance and mechanism of the observed
ransrepression of the 1.7-kb (and SGIV) hPMCA1 pro-oter fragments remains to be determined. This
ransrepression effect appears to be mediated by se-uence elements within the SG IV/exon 1* region, al-hough no candidate binding sites for putative trans-cting negative regulatory factors have so far beendentified. Nuclear run-on experiments may be helpful
t osteocalcin and hPMCA1 putative VDRE sequences. (A) The rat1 mg of nuclear extract from COS-1 cells transiently transfected withy (lane 1) or with monoclonal antibodies to ERa (lane 2), RXR (lanewas left at room temperature for 30 min and then the incubatione shifted hVDR/RXRa heterocomplex, the supershifted complex andin VDRE (lane 1) and putative hPMCA1 VDRE (lanes 2 and 3) werets overexpressing hVDR and hRXRa (lanes 1 and 3) or without any
resolved on a 4% polyacrylamide gel in 13 TBE buffer. Severalare designated by arrows. The mobility shift pattern of the putativecontaining overexpressed receptors (lane 3) was compared with theansfected pRC/CMV vector alone (lane 2). (C) hVDR/RXRa hetero-sequence was analyzed in the presence of increasing amounts of) or of unlabeled putative hPMCA1 VDRE (lanes 5–8). Increasingence were used, respectively, in lanes 1–4 and 5– 8. The incubation
Arrows indicate the VDR/RXRa heterocomplex and the free probe.
e raithbodion. Thalcracere
ndctsy trE–4equer.

iysr(Mcr
fqpfHfttibttYVtsrpret(wudmobmk
tt(oiD(f(is1igtt
ttap
A
hAacDN(vSGat
R
1
1
Vol. 277, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
n validating our findings; however, this form of anal-sis is insensitive to detecting small changes in tran-cription (23). Since PMCA1 represents a relativelyare transcript and the magnitude of the effect of 1,25-OH)2D3 on steady-state mRNA in ROS 17/2.8 and
DBK cells is modest (16), such studies may be incon-lusive as demonstrated in the recent study of PMCA1egulation in rat vascular smooth muscle cells (24).
We have been unable to verify the presence of aunctional VDRE in the best candidate hPMCA1 se-uence using EMSA. While we used ectopically ex-ressed hRXRa, it is conceivable that hVDR formsunctional complexes with one other isoform of hRXR.owever, as RXRa is the predominant isoform partner
or VDR in studies of the rat osteocalcin and rat os-eopontin VDREs (25) this seems unlikely. Competi-ion studies demonstrated that the hPMCA1 sequences capable of stabilizing or enhancing hVDR/RXRainding to its cognate DNA sequence, suggesting thathe hPMCA1 sequence might sequester factors inhibi-ory for receptor-DNA binding such as COUP-TF orY1 which can compete with VDR/RXR binding at DR3DREs (26, 27). Finally, although the sequence iden-
ified in the 1.7 kb hPMCA1 promoter fragment repre-ents the best fit to a minimal consensus VDRE in thisegion, and is highly conserved in the mouse PMCA1romoter (28), this sequence is neither a perfect directepeat nor may it be the only candidate VDRE in thentire hPMCA1 gene regulatory region. Thus, a func-ional VDRE conferring positive regulation by 1,25-OH)2D3 on the hPMCA1 gene may be present else-here in the gene, either at a position .650 bppstream of the transcription start sites or .1000 bpownstream in an intronic sequence. Such VDRE(s)ay be able to override any negative regulatory effects
f the 1.7 kb promoter fragment and thus be responsi-le, at least in part, for the previously observed vita-in D3-dependent upregulation of PMCA1 mRNA in
idney, intestine and bone-forming tissues (12, 16).On the other hand, whilst avian calbindin D28K is
ranscriptionally regulated by 1,25-(OH)2D3, post-ranscriptional regulation is probably more important29). Recent study of the rat calbindin D9K gene dem-nstrated that whilst a weak VDRE was demonstrablen vitro (30), this site was not responsible for vitamin
3 induction following 1,25-(OH)2D3 treatment in vivo31). Study of the human calbindin D9K gene promoterailed to demonstrate 1,25-(OH)2D3 responsiveness32) despite increased steady-state mRNA expressionn response to 1,25-(OH)2D3 (33). Our own recent workhowed that the increase in PMCA1 transcript levels in,25-(OH)2D3 treated MDBK cells may be due to anncreased mRNA half-life (16). The size of the hPMCA1ene is such that it may take in excess of 30 minutes toranscribe the entire .100 kb gene (14). Given thisime constraint, an effect of 1,25-(OH)2D3 on post-
727
ranscriptional processing of PMCA1 mRNA, ratherhan on transcriptional activity of the PMCA1 gene, is
plausible way to induce rapid changes in calciumump abundance.
CKNOWLEDGMENTS
The authors thank Professor J. Martin, Melbourne, Australia, foris kind donation of ROS 17/2.8 cells, Professor J. Eisman, Sydney,ustralia for his kind donation of the vectors encoding human VDRnd RXRa, Dr. H. DeLuca, Madison, Wisconsin, for the F2A10 mono-lonal antibody, and Dr. P. Chambon, Illkirch, France, for the 4RX-12 monoclonal antibody. Paul Glendenning was supported by theational Health and Medical Research Council of Australia
NHMRC), Medical Postgraduate Scholarship (987643), and the Uni-ersity of Western Australia, Athelstan and Amy Saw Foundationtipend. This project was supported by a grant from the Sir Charlesairdner Hospital Research Foundation, by the Mayo Foundation,nd by the philanthropy of the Esther V. Lind Endowment Fund andhe Ronald and Nancy Reagan Alzheimers Fund.
EFERENCES
1. Carafoli, E., and Stauffer, T. (1994) The plasma membrane cal-cium pump: Functional domains, regulation of the activity, andtissue specificity of isoform expression. J. Neurobiol. 25, 312–324.
2. Stauffer, T. P., Hilfiker, H., Carafoli, E., and Strehler, E. E.(1993) Quantitative analysis of alternative splicing options ofhuman plasma membrane calcium pump genes. J. Biol. Chem.268, 25993–26003.
3. Stauffer, T. P., Guerini, D., and Carafoli, E. (1995) Tissue dis-tribution of the four gene products of the plasma membrane Ca21
pump. A study using specific antibodies. J. Biol. Chem. 270,12184–12190.
4. Kumar, R., Haugen, J. D., and Penniston, J. T. (1993) Molecularcloning of a plasma membrane calcium pump from human os-teoblasts. J. Bone Miner. Res. 8, 505–513.
5. DeLuca, H. F. (1988) The vitamin D story: A collaborative effortof basic science and clinical medicine. FASEB J. 2, 224–236.
6. Wasserman, R. H., and Fullmer, C. S. (1989) On the molecularmechanism of intestinal calcium transport. Adv. Exp. Med. Biol.249, 45–65.
7. Zelinski, J. M., Sykes, D. E., and Weiser, M. M. (1991) The effectof vitamin D on rat intestinal plasma membrane CA-pumpmRNA. Biochem. Biophys. Res. Commun. 179, 749–755.
8. Wasserman, R. H., Smith, C. A., Brindak, M. E., De Talamoni,N., Fullmer, C. S., Penniston, J. T., and Kumar, R. (1992) Vita-min D and mineral deficiencies increase the plasma membranecalcium pump of chicken intestine. Gastroenterology 102, 886–894.
9. Cai, Q., Chandler, J. S., Wasserman, R. H., Kumar, R., andPenniston, J. T. (1993) Vitamin D and adaptation to dietarycalcium and phosphate deficiencies increase intestinal plasmamembrane calcium pump gene expression. Proc. Natl. Acad. Sci.USA 90, 1345–1349.
0. Armbrecht, H. J., Boltz, M. A., and Kumar, V. B. (1999) Intesti-nal plasma membrane calcium pump protein and its inductionby 1,25(OH) (2)D(3) decrease with age. Am. J. Physiol. 277,G41–G47.
1. Kumar, R., Strehler, E. E., and Cai, Q. (1997) Vitamin D and theplasma membrane calcium pump. In Molecular Nephrology: Kid-

ney Function in Health and Disease (Bonventre, J. V., and
1
1
1
1
1
1
1
1
2
2
2
2
interleukin-5 gene transcription by a novel nuclear run on
2
2
2
2
2
2
3
3
3
3
Vol. 277, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Schlondorff, D., Eds.), pp. 253–267. Dekker, New York.2. Pannabecker, T. L., Chandler, J. S., and Wasserman, R. H.
(1995) Vitamin-D-dependent transcriptional regulation of theintestinal plasma membrane calcium pump. Biochem. Biophys.Res. Commun. 213, 499–505.
3. Haussler, M. R., Mangelsdorf, D. J., Komm, B. S., Terpening,C. M., Yamaoka, K., Allegretto, E. A., Baker, A. R., Shine, J.,McDonnell, D. P., Hughes, M., et al. (1988) Molecular biology ofthe vitamin D hormone. Rec. Prog. Horm. Res. 44, 263–305.
4. Hilfiker, H., Strehler-Page, M. A., Stauffer, T. P., Carafoli, E.,and Strehler, E. E. (1993) Structure of the gene encoding thehuman plasma membrane calcium pump isoform 1. J. Biol.Chem. 268, 19717–19725.
5. Colnot, S., Lambert, M., Blin, C., Thomasset, M., and Perret, C.(1995) Identification of DNA sequences that bind retinoid Xreceptor–1,25(OH)2D3-receptor heterodimers with high affinity.Mol. Cell Endocrinol. 113, 89–98.
6. Glendenning, P., Ratajczak, T., Dick, I. M., and Prince, R. L.(2000) Calcitriol upregulates expression and activity of the 1bisoform of the plasma membrane calcium pump in immortaliseddistal kidney tubular cells. Arch. Biochem. Biophys. 380, 126–132, doi:10.1006/abbi.2000.1908.
7. Williams, G. R., Bland, R., and Sheppard, M. C. (1994) Charac-terization of thyroid hormone (T3) receptors in three osteosar-coma cell lines of distinct osteoblast phenotype: Interactionsamong T3, vitamin D3, and retinoid signaling. Endocrinology135, 2375–2385.
8. Meszaros, J. G., and Karin, N. J. (1993) Osteoblasts express thePMCA1b isoform of the plasma membrane Ca(21)- ATPase. J,Bone Miner, Res. 8, 1235–1240.
9. Abramowitz, J., Gonzalez, J. M., Rouse, D., and Suki, W. N.(1995) Differential expression of plasma membrane calciumpump mRNA isoforms in rat osteoblast-like cells. Miner. Elec-trolyte Metab. 21, 367–374.
0. Gill, R. K., and Christakos, S. (1993) Identification of sequenceelements in mouse calbindin-D28k gene that confer 1,25-dihydroxyvitamin D3- and butyrate-inducible responses. Proc.Natl. Acad. Sci. USA 90, 2984–2988.
1. Andrews, N. C., and Faller, D. V. (1991) A rapid microprepara-tion technique for extraction of DNA-binding proteins from lim-iting numbers of mammalian cells. Nucleic Acids Res. 19, 2499.
2. Sugawara, A., Sanno, N., Takahashi, N., Osamura, R. Y., andAbe, K. (1997) Retinoid X receptors in the kidney: Their proteinexpression and functional significance. Endocrinology 138,3175–3180.
3. Rolfe, F. G., and Sewell, W. A. (1997) Analysis of human
728
method based on the polymerase chain reaction. J. Immunol.Methods 202, 143–151.
4. Afroze, T., and Husain, M. (2000) c-Myb-binding sites mediateG(1)/S-associated repression of the plasma membrane Ca(21)-ATPase-1 promoter. J. Biol. Chem. 275, 9062–9069.
5. Staal, A., van Wijnen, A. J., Birkenhager, J. C., Pols, H. A.,Prahl, J., DeLuca, H., Gaub, M. P., Lian, J. B., Stein, G. S., vanLeeuwen, J. P., and Stein, J. L. (1996) Distinct conformations ofvitamin D receptor/retinoid X receptor-alpha heterodimers arespecified by dinucleotide differences in the vitamin D-responsiveelements of the osteocalcin and osteopontin genes. Mol. Endo-crinol. 10, 1444–1456.
6. Cooney, A. J., Leng, X., Tsai, S. Y., O’Malley, B. W., and Tsai,M. J. (1993) Multiple mechanisms of chicken ovalbumin up-stream promoter transcription factor-dependent repression oftransactivation by the vitamin D, thyroid hormone, and retinoicacid receptors. J. Biol. Chem. 268, 4152–4160.
7. Guo, B., Aslam, F., van Wijnen, A. J., Roberts, S. G., Frenkel, B.,Green, M. R., DeLuca, H., Lian, J. B., Stein, G. S., and Stein,J. L. (1997) YY1 regulates vitamin D receptor/retinoid X receptormediated transactivation of the vitamin D responsive osteocalcingene. Proc. Natl. Acad. Sci. USA 94, 121–126.
8. Du, Y., Carlock, L., and Kuo, T. H. (1995) The mouse plasmamembrane Ca21 pump isoform 1 promoter: Cloning and charac-terization. Arch. Biochem. Biophys. 316, 302–10.
9. Enomoto, H., Hendy, G. N., Andrews, G. K., and Clemens, T. L.(1992) Regulation of avian calbindin-D28K gene expression inprimary chick kidney cells: Importance of posttranscriptionalmechanisms and calcium ion concentration. Endocrinology 130,3467–3474.
0. Darwish, H. M., and DeLuca, H. F. (1992) Identification of a1,25-dihydroxyvitamin D3-response element in the 59-flankingregion of the rat calbindin D-9k gene. Proc. Natl. Acad. Sci. USA89, 603–607.
1. Colnot, S., Ovejero, C., Romagnolo, B., Porteu, A., Lacourte, P.,Thomasset, M., and Perret, C. (2000) Transgenic analysis of theresponse of the rat calbindin-D 9k gene to vitamin D. Endocri-nology 141, 2301–2308.
2. Barley, N. F., Prathalingam, S. R., Zhi, P., Legon, S., Howard, A.,and Walters, J. R. (1999) Factors involved in the duodenal ex-pression of the human calbindin-D9k gene. Biochem. J. 341,491–500.
3. Walters, J. R., Howard, A., Lowery, L. J., Mawer, E. B., andLegon, S. (1999) Expression of genes involved in calcium absorp-tion in human duodenum. Eur. J. Clin. Invest. 29, 214–219.




















