
Preliminary Report on the Excavations at Jerusalem’s Southeastern Hill, Area D3
Upload
independentCategory
view
1download
0
ava i l ab l e a t www.sc i enced i r ec t . com
C l i n i ca l Immuno logy
www.e l sev i e r . com/ loca te /yc l im
Clinical Immunology (2011) 138, 212–221
1α,25-dihydroxyvitamin D3 (vitamin D3) catalyzessuppressive activity on human natural regulatory Tcells, uniquely modulates cell cycle progression, andaugments FOXP3Vanessa Morales-Tiradoa,1, David G. Wichlana, Thasia E. Leimig a,Shayna E.A. Street a,2, Kimberly A. Kasowb,3, Janice M. Riberdya,⁎
a Department of Immunology, St. Jude Children's Research Hospital, Memphis, TN 38105, USAb Division of Bone Marrow Transplantation & Cellular Therapy, Department of Oncology,St. Jude Children's Research Hospital, Memphis, TN 38105, USA
Received 15 July 2010; accepted with revision 6 November 2010Available online 16 December 2010
Abbreviations: nTregs, natural regreceptor; VDR, vitamin D receptor; IPtarget of rapamycin; HDAC, Histone de⁎ Corresponding author. Department
TN 38105, USA. Fax: +1 901 595 2176.E-mail address: Janice.riberdy@stj
1 Present Address: Division of Bone MaMemphis, TN 38105, USA.2 Present Address: Diamantina Institu
Buranda, QLD, 4102, Australia.3 Present Address: Bone Marrow and
1521-6616/$ – see front matter © 201doi:10.1016/j.clim.2010.11.003
KEYWORDSRegulatory T cells;FOXP3;Vitamin D3;Immune regulation;Immunotherapy;Tolerance
Abstract Human natural regulatory T cells (nTregs) show great promise for therapeuticallymodulating immune-mediated disease, but remain poorly understood. One explanation underintense scrutiny is how to induce suppressive function in non-nTregs and increase the size of theregulatory population. A second possibility would be to make existing nTregs more effective, likea catalyst raises the specific activity of an enzyme. The latter has been difficult to investigatedue to the lack of a robust short-term suppression assay. Using a microassay described herein wedemonstrate that nTregs in distinct phases of cell cycle progression exhibit graded degrees ofpotency. Moreover, we show that physiological concentrations of 1α,25-dihydroxyvitamin D3
(vitamin D3) boosts nTregs function. The enhanced suppressive capacity is likely due to vitaminD3's ability to uniquely modulate cell cycle progression and elevate FOXP3 expression. These datasuggest a role for vitamin D3 as a mechanism for catalyzing potency of nTregs.© 2010 Elsevier Inc. All rights reserved.ulatory T cells; vitamin D3, 1EX, immune dysregulation, polacetylases.of Hematology, Mail Stop 341, S
ude.org (J.M. Riberdy).rrow Transplantation & Cellular
te for Cancer Immunology & Me
Stem Cell Transplantation Pro
0 Elsevier Inc. All rights reser
α,25-dihydroxyvitamin D3; PFA, 1.5% paraformaldehyde; TCR, T cellyendocrinopathy, enteropathy, X-linked syndrome; mTOR, mammalian
t. Jude Children's Research Hospital, 262 Danny Thomas Place, Memphis,
Therapy, Department of Oncology, St. Jude Children's Research Hospital,
tabolic Medicine, University of Queensland, Princess Alexandra Hospital,
gram, University of North Carolina, Chapel Hill, NC, 27599, USA.
ved.

2131α,25-dihydroxyvitamin D3
1. Introduction
Immune-mediated disease causes substantial morbidity andmortality. Therapeutic modalities to reign in auto-aggressiveimmune reactions are limited and compromise a patient'sability to respond to life-threatening infections. Studies tounderstand the immune system's own rheostat mechanism forpreventing autoimmunity, i.e. natural regulatory T cells(nTregs), have shown promising efficacy in numerous animalmodels of disease. Many investigations have focused on theprocess of “infectious tolerance” or how non-regulatory T cellsacquire suppressive function. Fewer investigations regardingmechanisms to augment or “catalyze” regulatory activity fromexisting nTregs have been reported and parallel studies usinghuman nTregs have been particularly limited [1]. Given thechallenges associated with obtaining therapeutic quantities ofnTregs, elucidation of mechanisms which enhance suppressivepotency may provide insight to develop immunotherapieswhich achieve clinical efficacy with fewer cells.
To investigate the mechanisms controlling the potency ofhuman nTregs, we first considered how to best demonstratesuperior function when comparing samples from a heteroge-neous group of individuals. Sincemost experiments do not yieldsufficient primary nTregs for elegant in vivo models usingimmunocompromised mice [2–4], most studies have beenrelegated to in vitro proliferation assays. However, prolifera-tion assays can be cumbersome and are not always suitable forsmaller amounts of peripheral blood. Another caveat is thatstandard proliferation assays most commonly used to showsuppressive function (generally inhibition of CFSE dilution or3H-thymidine incorporation) take days to complete, therebyobscuring early molecular events. Thus, we developed a short-term suppression assay (6 hours) requiring yet fewer nTregs.Importantly this assay is amenable to awide titration of nTregs,allowing an objective measure of nTregs quality. Using thisassay, we demonstrate graded degrees of activity for primaryhuman nTregs in specific phases of cell cycle progression,directly linking this process with functional capacity. Theability to modulate cell cycle progression and known immuno-modulatory properties prompted us to evaluate vitamin D3 as aphysiologically important candidate for modulating nTregsfunction [5–8]. Utilizing the criteria described above,enhanced suppression across titrated doses of nTregs, weshow that vitamin D3 significantly augments suppressiveactivity. Moreover, vitamin D3 likely mediates its effectsthrough modulation of cell cycle progression and increasedFOXP3 expression. These data support the notion thatincreasing the quality of regulatory functionmaybe a promisingstrategy in settings where large quantities of therapeuticnTregs are not available.
2. Materials and methods
2.1. Peripheral blood samples
Peripheral blood was obtained from either healthy plateletdonors at St. Jude Children's Research Hospital Blood DonorCenter with permission from the Institutional Review Board(IRB) or purchased from Lifeblood Biological Services(Memphis, TN). Peripheral blood was obtained from a severelyill child diagnosed with IPEX at St. Jude Children's Research
Hospital with permission from the Institutional Review Board(IRB) and parental consent.
2.2. Purification of CD4+CD25− and CD4+CD25+
T lymphocytes
CD4+CD25− and CD4+CD25+ populations were isolated using anAutoMACS® cell sorter following manufacturer's instructions(CD4+CD25+ Regulatory T Cell Isolation Kit, Miltenyi BiotecInc., Auburn, CA). Purities (N95%) were assessed by flowcytometry.
2.3. Cell culture
CD4+CD25+ Tregs in culture medium (X-Vivo supplementedwith 2 mM L-glutamine, 15% Human Serum, and 10U/mlrecombinant human IL2) were activated with anti-CD2/CD3/CD28 antibody-coated beads (MACS® T Cell Activation/Expansion Kit—Miltenyi Biotec Inc.) following manufacturer'sconditions for the indicated times.
Vitamin D3 was added at 20 nM where indicated.
2.4. Suppression microassay
10,000 CD4+CD25− T cells were activated as described abovein the presence or absence of nTregs for 6 hours, unlessotherwise indicated. Control conditions included nTregs cellcultures and CD4+CD25− T cell cultures alone.
2.5. nTregs identification by labeling withfluorescent probes
Isolated nTregs were labeled with 1 μM CMRA Cell Tracker™(Molecular Probes, Invitrogen, CA) using serum-free media.Cells were incubated for 45 minutes at 37 °C, followed bytwo washes using serum-free media prior to resuspension inculture media.
2.6. Propidium iodide staining
Cell samples were washed once with PBS followed by labelingwith Propidium Iodide solution (Propidium Iodide, sodiumcitrate and Triton X-100, Sigma, St. Louis, MO). Sampleswere treated with RNAse (Calbiochem, USA) for 30 minutesat room temperature prior to analysis.
2.7. Hoescht 33342
Cells were labeled with 10 μM Hoescht 33342 in mediacontaining human serum (Sigma, St. Louis, MO) for 40minutes at 37 °C, followed by two washes using PBS/2%human serum. Hoescht 33342 was added at a finalconcentration of 1 μM to avoid efflux of the dye.
2.8. mRNA analysis
Total RNA was extracted using the QIAGEN RNeasy® Micro Kit(QIAGEN, Valencia, CA) and reverse transcribed using theTaqMan® Reverse Transcription Kit (Applied Biosystems,

214 V. Morales-Tirado et al.
Foster City, CA). Human IL2 mRNA was tested using ABL1 asthe endogenous control. nTregs mRNA was used as acalibrator sample and mRNA from CD4+CD25− T cells as apositive control. Human VDR and FOXP3 mRNA were testedusing the TaqMan® Gene Expression Assays. Gene Assays: IL2:Hs00174114_m1, ABL1: Hs00245445_m1, FOXP3:Hs00203958_m1, VDR: Hs00172113_m1.
2.9. Confocal microscopy analysis
nTregs were activated as described above in the presence orabsence of 20nM vitamin D3 in 35 mm Poly-D-lysine dishes with10 mm coverslips for 5 days. Cells were fixed with 1.5%paraformaldehyde (PFA) and permeabilized with 0.3% TritonX-100 followed by anti- FOXP3 Alexa 488 (clone 259D—BioLegend, San Diego, CA) staining. ProLong® Gold anti fadereagentwithDAPIwas usedasmounting solutionprior to study.Analysis was performed using Nikon C1Si confocal/ TE2000microscope and NIS Elements software (Nikon Inc., Melville,NY).
2.10. Flow cytometry
Cellular surfacemarkers were assessed by flow cytometry usingthe following antibodies: anti-CD4 APC (clone SK3, BectonDickinson, CA) anti-CD25 PE (clone 4E3, Miltenyi Biotec); anti-CD127 FITC (clone hIL-7R-M21, Becton Dickinson). Sampleswere run on a FACS Calibur using Cell Quest software (BDBiosciences) and analyzed using FlowJo 8.8.2 software(Ashland, OR).
2.11. Intracellular staining
Cells were fixed in 1.5% PFA and permeabilized with cold 100%methanol prior to staining (clone 259D BioLegend, San Diego,CA; clone PCH101 eBioscience, San Diego, CA; and the isotypecontrols recommended by the respective manufacturers).FOXP3 protein in nTregs from the patient with IPEX wasdetected using the anti-human FOXP3 Flow kit (BioLegend);Alexa-Fluor 488 anti-FOXP3 (clone 259D), Alexa-Fluor 488mouse IgG1 k isotype control and anti-human CD4-PE-Cy5/CD25-PE cocktail (clones RPA-T4/BC96) as per manufacturer'sinstructions. Plots show the comparison between the anti-FOXP3 staining or respective IgG antibody isotype control usingFlowJo 8.8.2 software.
2.12. Protein analysis by Western blot
Cell extracts were lysed and fractionated (cytoplasmic andnuclear extracts) using the NE-PER® Nuclear and CytoplasmicExtraction Reagents (Thermo Scientific, Rockford, IL) as permanufacturer's conditions. Quantitation was done using theBCA™ Protein Assay Kit (Thermo Scientific). 10 μg of proteinwas loadedper lane, separatedby SDS-PAGE and transferred toan Immobilon™ PVDF Transfer Membrane (Millipore, Bedford,MA), followed by overnight blocking. Primary antibodies wereincubated for 2 hours at RT, followed by several washes andincubation with secondary antibodies for 1 hour at RT.Membranes were developed using the SuperSignal® WestPico Chemiluminescent Substrate as per manufacturer's
instructions (Thermo Scientific). FOXP3 detection: cloneH190 rabbit polyclonal IgG (Santa Cruz Biotechnology, Inc.,Santa Cruz, CA).
2.13. Analysis and statistics
Where possible, samples were set up in triplicate and eachrespective cDNA was analyzed for IL2 in triplicate. Standarddeviations (SD) were determined by paired t test usingGraphPad Prism software (San Diego, CA). If culture sampleswere set up in duplicate followed by analyzing the cDNA for IL2content in triplicate, standard errors (SE) were determined byExcel software (Henderson, NV).
3. Results
Initial attempts from our laboratory to identify physiologicalparameters, which catalyze human nTregs activity, suggestedthat control of cell cycle progression may be linked tosuppressive potency (Riberdy et al.,manuscript in preparation).However, to more carefully explore the relationship betweenregulatory potency and cell cycle progression, an assay meetingseveral criteriawas needed. First, to interpret events that occurduring a given roundof division itwas critical to reduce the assayduration from days (as required for standard proliferationassays) to one completed within hours. Secondly, a functionalassay sufficiently robust to measure suppression across aheterogeneous range of responses, yet adequate to detectthose approaching the lower end of the spectrum wasimportant. Thirdly, since human nTregs investigations arehampered by the limited availability of cells, particularly inclinical studies of various immune-mediated diseases,suppression assays necessitating fewer cells would be highlydesirable. The crucial role of IL2 production and features such asrapid mRNA up-regulation [9,10], combined with sensitivemethods for detection, suggested that such a readout wouldbe superior to the standardproliferation assay [9,11–14]. Fig. 1Aillustrates the dynamic spectrum of IL2 mRNA signal fromisolated CD4+CD25− T cells at various times after activation,with 7 of 9 samples peaking between 6 and 12 hours afteractivation before declining. Since the priority was to develop ashort-term assay, subsequent experiments were done using a 6hour activation period. Of note, we have empirically observedthat despite the 6- to 12-hour range for peak IL2 mRNAproduction among individuals, themajority of CD4+CD25− T cellstested at 6 hours post-activation yielded sufficient signal toreliably assess suppression (Fig. 1B and data not shown).Similarly prioritizing the need to reduce the number of cells ina short-term suppression assay, titrated doses of CD4+CD25− Tcells from different individuals identified 10,000 cells as theminimumnumber of cellsmost reliably yielding signal above thenegative controls (negative control CT values ranged 35–40;Fig. 1B and data not shown). It is interesting to note that aminority of samples remain below the limits of detection at thehighest dose of cells, perhaps reflecting the lower end of the IL2response range or the donors’ health status (Fig. 1B).
Having reduced the time for a CD4+CD25− T cell functionalreadout assay from days to 6 hours, we were poised toinvestigate whether cell cycle progression was linked to nTregspotency. However, three technical issues needed to beaddressed. First, since an exclusive marker for human nTregs

Figure 1 Cell cycle progression modulates nTregs function. (A) Optimal time for IL2 mRNA detection was determined by activating10,000 CD4+CD25− T cells in vitro for the indicated times, followed by qRT-PCR analysis for cytokine [n=9]. (B) The minimum numberof cells for reliable IL2mRNA detection was determined by assaying titrated doses of in vitro activated CD4+CD25− T cells for cytokinevia qRT-PCR [1 k n=8, 5 k n=12, 10 k n=18, 25 k n=21, 50 k n=18]. (C) nTregs were activated in vitro for 5 days, labeled with Hoescht33342, sorted into the indicated fractions and assayed for DNA content at time 0 and 6 hrs under suppression assay conditions. This isrepresentative of 2 experiments. (D) 10,000 nTregs in the indicated cell cycle phases were sorted and incubated with 10,000CD4+CD25− T cells under the described suppression microassay conditions for 6 hrs prior to IL2mRNA analysis. Error bars represent SDand this is representative of 3 experiments. (E) Purified nTregs were activated and fractionated as in 1D, prior to assaying forsuppressive activity. Error bars represent SE and this is representative of 4 experiments.
2151α,25-dihydroxyvitamin D3
is lacking, we purified anergic cells expressing aCD4+CD25brightCD127loFOXP3+ phenotype (Supplemental Fig. 1and data not shown). These characteristics comprise a broadconsensus phenotype for human nTregs, at least until adefinitive lineage specification marker is elucidated [15,16]Secondly, conventional cell cycle analysis evaluates DNAcontent by propidium iodide staining of fixed cells [17],precluding functional analysis of specific subsets. Thirdly,although nTregs show evidence of rapid proliferation in vivo[18,19], freshly isolated cells are largely anergic unlessprovided additional signals [20–23]. To circumvent the lattertwo hurdles, primary human nTregs were activated and labeledfor DNAcontentwithHoechst 33342. Fivedays of activationwasthe shortest activation periodwhere sufficient numbers of cellswere found in the S and G2/M phases (Supplemental Fig. 2).Next, viable nTregs in indicated phases of cell cycle progressionwere sorted by flow cytometry and immediately assayed forregulatory function. As an additional control, DNA content ofthe sorted cells was evaluated at time 0 and 6 hours to verifythat the nTregs remained within the desired phases for theduration of the assay (Fig. 1C). Surprisingly, nTregs in eitherS/G2/M combined, or S and G2/M fractionated, exhibitedsignificantly more potent suppressor activity than those in theG0/G1 pool, further substantiating the notion that cell cycleprogression is directly linked to nTregs potency (Fig. 1D and E).
If modulating cell cycle progression is a physiologicallyrelevant mechanism for controlling nTregs function, then onewould predict that important biologicalmolecules participate inthis process. Since vitamin D3 has been shown to alter cell cycleprogression [24] and is emerging as a key immunomodulator[8,25,26], we investigated whether this molecule couldpotentiate nTregs function. Fig. 2A shows that nTregs activatedin thepresence of physiological concentrations of vitaminD3aresignificantly more effective at suppression than mock treatednTregs. Importantly, this enhancement is maintained when afixed number of nTregs interacts with an increasing numberof CD4+CD25− T cells, providing an objective demonstrationof amplified potency. We were curious whether vitamin D3mediated enhancement of suppressor function could also beobserved at the level of IL2 protein production. However, thediffering turnover time for IL2 mRNA versus IL2 protein makesdirectly adapting the above mRNA suppression assay tointracellular cytokine staining challenging and additional kineticstudies will be required for optimization. Albeit modest, anincrease in suppression of IL2 protein production was observedwhen vitamin D3-treated nTreg were assayed, suggesting thatregulatory activity can be detected at both the protein andmRNA levels (Fig. 2B). Heightened function is not due to carryover of vitamin D3 acting on the responding cells themselves, asaddition of vitamin D3 directly to CD4+CD25− T cells does not

Figure 2 Vitamin D3 modulates human nTregs potency at both the mRNA and protein levels. (A) nTregs were either mock treated ortreated with vitamin D3 for 5 days as described in Materials and methods, assayed for suppressive potency with 10,000 CD4+CD25− Tcells at the indicated ratios, and analyzed for IL2 mRNA. Data represents 2 of 7 experiments. Sufficient nTregs were available fromDonor 2 to set up each condition in triplicate, while the yield of nTregs from Donor 1 only permitted a single well for each condition.Error bars in the panel from Donor 2 represent SD. (B) Mock treated or vitamin D3 treated nTregs were assayed for the ability to inhibitIL2 protein production by CD4+CD25− T cells. nTregs were labeled with CMRA Cell Tracker™ to distinguish them from the CD4+CD25− Tcells, mock treated or vitamin D3 treated, and assayed for suppressive function with the following changes to the microassay. 10,000CD4+CD25− T cells were incubated with an equivalent number of nTregs under the described microassay conditions for 10 hrs.Brefeldin A was added during the last 5 hrs to inhibit cytokine secretion prior to intracellular IL2 staining. (C) Exposure to vitamin D3 atdifferent stages of nTregs activation differentially alters potency. 20nM vitamin D3 was added to the nTregs activation cultures at0 hrs (Early); 96 hrs (Late); or 0, 48, and 96 hrs (Continuous). nTregs were assayed for suppressive function during the described 6 hrsuppression microassay (n=5 for the different nTregs groups).
216 V. Morales-Tirado et al.
diminish the level of detectable IL2 mRNA or protein(Supplemental Fig. 2). All nTregss populations we have testedthus far have exhibited increased regulatory activity whenexposed to vitamin D3, but the kinetics of this boost variesamong individuals (data not shown). This heterogeneity likelyreflects genetic modifiers, placing the delineation of a precisetime course beyond the scope of this study. Thus, to investigatewhether the timing of vitamin D3 exposure was important in amore generalized manner, the potency of nTregs receiving asingle dose either at the beginning or end of T cell receptor(TCR) stimulationwas compared to those receiving a continuallyreplenished supply. Fig. 2C shows graded degrees of suppressionamong the groups, with those getting vitamin D3 throughout theactivation program being most effective.
To better understand the molecular mechanism by whichvitamin D3 acts on nTregs, we examined the effects of TCRstimulation on vitamin D receptor (VDR) expression. PurifiednTregs were activated ex vivo, either in the presence orabsence of vitamin D3 and assayed at the indicated times forVDR mRNA. Expression of VDR peaks around 12 hours afteractivation, remains partially elevated for at least 14 days,and is not differentially regulated in the presence of ligand(Fig. 3A and data not shown). Thus, the profile of VDRregulation in nTregs is similar to that described forconventional T cells (i.e. VDR increases upon TCR stimulationin a ligand-independent fashion) [27,28].
It has been reported for multiple cell types that vitaminD3 blocks the transition from G0/G1 to S [29,30]. Consistentwith such a block, vitamin D3 treated CD4+CD25− T cellsexhibited fewer cells in the S and G2/M phases at both 48 and120 hours, compared to mock treated cells (Fig. 3B). At 48hours, nTregs similarly showed decreased numbers of cells inS and G2/M phases. However, the percentage of cells in S orG2/M phases unexpectedly increased in the vitamin D3treated nTregs after 120 hours (approximately 3- and10-fold, respectively), suggesting a release or less stringentblock in cell cycle progression at this time (Fig. 3C). Next, wedetermined whether this pattern was unique to a specificindividual or reflective of a more generalized trend, byperforming a similar time course of cell cycle analysis onnTregs from 6 additional donors. To normalize for variationsamong different individuals, we combined the percentage ofcells in S with those in G2/M as an overall measurement of“proliferating” cells within the total population (referred toas S/G2/M). Table 1 illustrates that after 120 hours ofactivation in the presence of vitamin D3, 5 of 6 individualsincrease the number of cells in S/G2/M, indicating thatnTregs exhibit a late release from the G0/G1 block.
FOXP3 is critical for nTregs function [31–36] and in silicostudies suggested that the promoter for this gene has twopotential VDR binding elements (Riberdy, unpublished).Hypothesizing that vitamin D3 may be mediating its activity
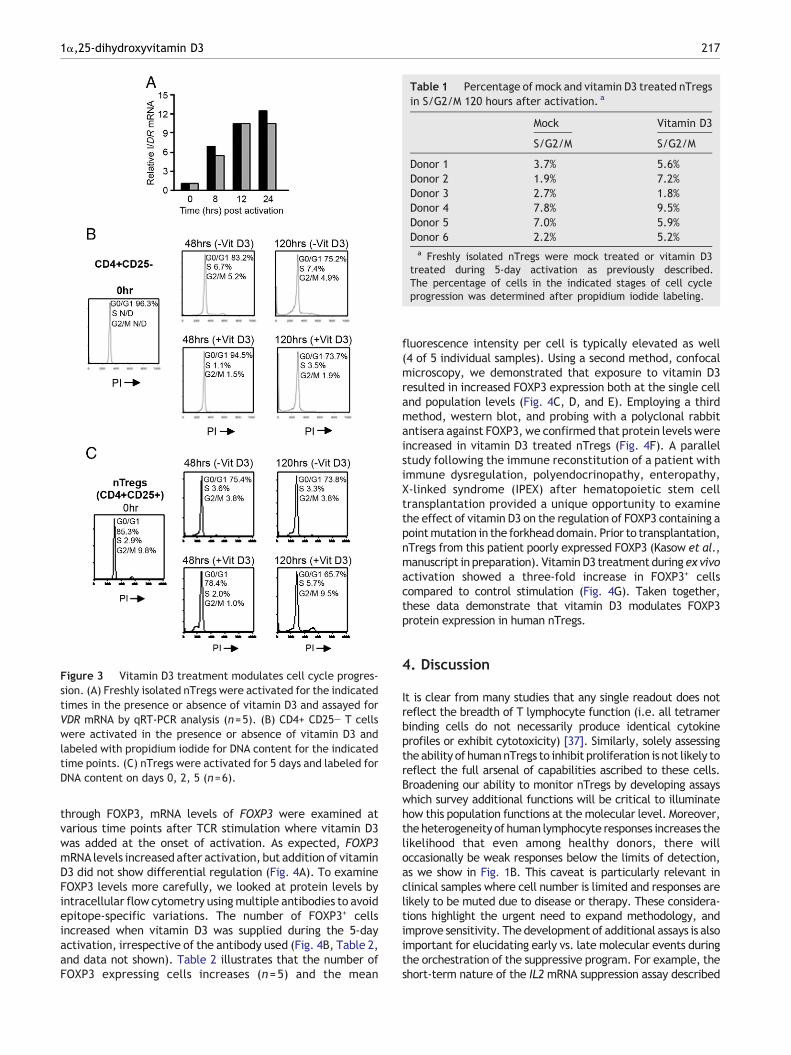
Figure 3 Vitamin D3 treatment modulates cell cycle progres-sion. (A) Freshly isolated nTregs were activated for the indicatedtimes in the presence or absence of vitamin D3 and assayed forVDR mRNA by qRT-PCR analysis (n=5). (B) CD4+ CD25− T cellswere activated in the presence or absence of vitamin D3 andlabeled with propidium iodide for DNA content for the indicatedtime points. (C) nTregs were activated for 5 days and labeled forDNA content on days 0, 2, 5 (n=6).
Table 1 Percentage of mock and vitamin D3 treated nTregsin S/G2/M 120 hours after activation. a
Mock Vitamin D3
S/G2/M S/G2/M
Donor 1 3.7% 5.6%Donor 2 1.9% 7.2%Donor 3 2.7% 1.8%Donor 4 7.8% 9.5%Donor 5 7.0% 5.9%Donor 6 2.2% 5.2%a Freshly isolated nTregs were mock treated or vitamin D3
treated during 5-day activation as previously described.The percentage of cells in the indicated stages of cell cycleprogression was determined after propidium iodide labeling.
2171α,25-dihydroxyvitamin D3
through FOXP3, mRNA levels of FOXP3 were examined atvarious time points after TCR stimulation where vitamin D3was added at the onset of activation. As expected, FOXP3mRNA levels increased after activation, but addition of vitaminD3 did not show differential regulation (Fig. 4A). To examineFOXP3 levels more carefully, we looked at protein levels byintracellular flow cytometry usingmultiple antibodies to avoidepitope-specific variations. The number of FOXP3+ cellsincreased when vitamin D3 was supplied during the 5-dayactivation, irrespective of the antibody used (Fig. 4B, Table 2,and data not shown). Table 2 illustrates that the number ofFOXP3 expressing cells increases (n=5) and the mean
fluorescence intensity per cell is typically elevated as well(4 of 5 individual samples). Using a second method, confocalmicroscopy, we demonstrated that exposure to vitamin D3resulted in increased FOXP3 expression both at the single celland population levels (Fig. 4C, D, and E). Employing a thirdmethod, western blot, and probing with a polyclonal rabbitantisera against FOXP3, we confirmed that protein levels wereincreased in vitamin D3 treated nTregs (Fig. 4F). A parallelstudy following the immune reconstitution of a patient withimmune dysregulation, polyendocrinopathy, enteropathy,X-linked syndrome (IPEX) after hematopoietic stem celltransplantation provided a unique opportunity to examinethe effect of vitamin D3 on the regulation of FOXP3 containing apointmutation in the forkheaddomain. Prior to transplantation,nTregs from this patient poorly expressed FOXP3 (Kasow et al.,manuscript in preparation). VitaminD3 treatment during ex vivoactivation showed a three-fold increase in FOXP3+ cellscompared to control stimulation (Fig. 4G). Taken together,these data demonstrate that vitamin D3 modulates FOXP3protein expression in human nTregs.
4. Discussion
It is clear from many studies that any single readout does notreflect the breadth of T lymphocyte function (i.e. all tetramerbinding cells do not necessarily produce identical cytokineprofiles or exhibit cytotoxicity) [37]. Similarly, solely assessingtheability of humannTregs to inhibit proliferation is not likely toreflect the full arsenal of capabilities ascribed to these cells.Broadening our ability to monitor nTregs by developing assayswhich survey additional functions will be critical to illuminatehow this population functions at the molecular level. Moreover,the heterogeneity of human lymphocyte responses increases thelikelihood that even among healthy donors, there willoccasionally be weak responses below the limits of detection,as we show in Fig. 1B. This caveat is particularly relevant inclinical samples where cell number is limited and responses arelikely to be muted due to disease or therapy. These considera-tions highlight the urgent need to expand methodology, andimprove sensitivity. The development of additional assays is alsoimportant for elucidating early vs. late molecular events duringthe orchestration of the suppressive program. For example, theshort-term nature of the IL2mRNA suppression assay described

Figure 4 FOXP3 protein expression is augmented upon Vitamin D3 treatment. (A) Freshly isolated nTregs were activated andassayed for FOXP3 mRNA by qRT-PCR analysis (n=5). (B) Freshly isolated nTregs were activated for 5 days as previously described intext. Gating strategy: CD4+ T cells (99%) were selected and analyzed for CD25 vs. FOXP3. IgG isotype controls and unlabelled cellswere used to define gates. The percentage of CD4+ T cells expressing both CD25 and FOXP3 as depicted by the oval gate, are indicated.(C–E) Confocal microscopy analysis of nTregs. (C) Activated cells were fixed and permeabilized, followed by anti-FOXP3 staining.Controls included unlabeled cells and isotype controls (data not shown). Anti-fade reagent with DAPI was used as mounting solutionand nuclear marker. Mean intensity of anti-FOXP3 was analyzed both at the single cell (D) and population (E) levels (representing 1 of 3experiments) using NIS Elements Software. (F) Western blot analysis for FOXP3 expression using 10 mg of protein per lane as describedin Materials and methods. (G) nTregs purified from a patient with IPEX were either mock treated or vitamin D3 treated during ex vivoactivation and stained for FOXP3 expression. Gating strategy: CD4+ T cells (99%) were selected and analyzed for CD25 vs. FOXP3. IgGisotype controls and unlabeled cells were used to define gates. The percentage of CD4+ T cells expressing both CD25 and FOXP3, asdepicted by the gate are indicated.
Table 2 FOXP3 expression in mock and vitamin D3 treatednTregs during activation. a
Vitamin D3 %CD4+CD25+FOXP3+ MFI FOXP3
Donor 1 − 29 195+ 38.6 304
Donor 2 − 29.5 28+ 48 35
Donor 3 − 75.9 7.5+ 83.7 8
Donor 4 − 19.7 19+ 35.9 12.3
Donor 5 − 25.8 170+ 38.6 295
a Freshly isolated nTregs were activated for 5 days in thepresence or absence of vitamin D3 followed by flow cytometryanalysis of FOXP3 expression. The same gating strategydescribed in Fig. 4 was used.
218 V. Morales-Tirado et al.
in this work was critical for the cell cycle progression studies,which could not have been done using a proliferation-basedmethod. Additionally, the vitamin D3-mediated enhancementof nTregs potency would have been missed without a 6-hourassay. It is tempting to speculate that because an immuneresponse grows exponentially, a small enhancement of IL2suppression during the first 6 hours will likely result in a largeimpact on the overall magnitude.
Initial attempts to examine the effect of vitamin D3 usingproliferation assays were inconclusive and prompted us toexamine whether augmented potency may be transient.Indeed altering the dosing of vitamin D3 during nTregsactivation suggests that this molecule make act temporally(Fig. 2C). It is intriguing to consider that the transient effect ofvitamin D3 may provide an additional fine-tune adjustmentsuch that autoimmunity is ablated in one anatomical locale,yet at another site the body remains poised to respond topathogens. It has been suggested that vitamin D3 can facilitatethe recruitment of induced-Tregs into the modulation process

2191α,25-dihydroxyvitamin D3
[8,38], while our data demonstrate a novel role for thismolecule, augmenting function from existing nTregs. Such ascenario would support the concept that vitamin D3 rapidlyacts on nearby nTregs to blunt immune genesis. However, inphysical environments requiring a larger number of regulatoryT cells such as the gut, induced-Tregs could be enrolled toassist.
Given the lack of a single cell suppression assay, it remainsunknown whether all the nTregs subjected to in vitro analysisactually participate in the modulatory process or whetheronly a subpopulation mediates the bulk of activity. This isperhaps most relevant in the cell cycle studies. Given theobservation that nTregs in S or G2/M are more potent, it isreasonable to speculate that the increase in S/G2/M cellsafter vitamin D3 exposure results in the enhanced suppressivefunction. To test this directly and compare treated vs. non-treated nTregs in S/G2/M, additional methods requiring evenfewer cells need to be developed because sufficient cellscannot readily be obtained from peripheral blood. Nonethe-less, it is important to point out the hypothetical scenariodescribed above does not necessarily imply that the nTregs inS/G2/M are the subset most efficient at mediating othersuppressive functions such as inhibiting proliferation,modulating dendritic cell function, or arresting cytotoxicity.Rather, it is likely that population of cells characterized by aCD4+CD25brightCD127loFOXP3+ phenotype will be furtherpartitioned into functional subsets, perhaps by markerssuch as CD45RA or HLA-DR, which are already known tocorrelate with specific functional capabilities [39,40].
The ability of vitamin D3 to enhance nTregs function,possibly via increasing FOXP3, emphasizes the need toconsider both quality and quantity when considering thedynamics of immunomodulation. This issue is highlighted bythe demonstration that vitamin D3 increases the number ofnTregs expressing FOXP3, as well as the amount of protein percell (Fig. 4 and Table 2). However, it is unclear whether thenTregs expressing the most FOXP3 are actually more potent.Additional studies are necessary to elucidate how quantity(amount of FOXP3) and quality (perhaps as a post-transla-tional modification) relate to suppressive function. Theunique regulation of FOXP3 in human T cells poses additionalconstraints on interpretation of quality and quantity. Forexample, FOXP3 is restricted to murine T cells withregulatory function while all human T lymphocytes transient-ly express FOXP3 upon TCR stimulation, without necessarilyacquiring suppressive function [41]. Thus, simple enumera-tion studies using FOXP3 alone may not reflect bona fidenTregs. An additional qualitative point is demonstrated byectopic expression of FOXP3 in CD4+CD25− T cells to induceregulatory function. In these studies ectopic expression ofFOXP3 in murine CD4+CD25− T cells is sufficient to inducepotent regulatory function [4,31,42], while similar experi-ments with human T lymphocytes have resulted in onlymodest levels of activity [32,43–45]. However, potentsuppressive activity can be obtained if high levels of ectopicFOXP3 are sustained for several days, perhaps reflecting arequirement for epigenetic changes [46]. The need forepigenetic alterations is further supported by the observationthat bona fide regulatory cells fully demethylate a CpG islandin the promoter of FOXP3. In contrast, those exhibiting apartial demethylation status show only transient expressionand lack functional activity [47–49].
The ability to overcome the cell cycle block mediated byvitamin D3 is reminiscent of the observation that nTregsselectively expand in the presence of rapamycin, an anti-proliferative agent known to inhibit the activity of themammalian target of rapamycin (mTOR) [50,51]. Histonedeacetylases (HDAC) are key molecules in regulating cellcycle progression not only by virtue of their activity onhistones and therefore control of transcription, but also ontranscription factors themselves. Interestingly, HDAC inhibi-tors increase acetylation of FOXP3 itself and this modifica-tion is associated with enhanced nTregs function [52]. Thecombined observations that nTregs potency is enhanced byvitamin D3, rapamycin, and HDAC inhibitors, strongly suggestthat cell cycle progression is uniquely regulated in nTregs.Understanding the molecular events controlling cell cycleprogression in human nTregs may reveal novel targets forenhancing their function and will be critical for translatingthese observations into strategies which harness the fulltherapeutic potential of this elusive population.
Supplementary materials related to this article can befound online at doi:10.1016/j.clim.2010.11.003.
Acknowledgments
The authors would like to thank Laura McLaughlin, LauraSpaulding, Victoria Fabry and Jaime Beine, for technicalassistance in developing methods, which could be applied tothis study. We also thank Jim Houston and Drs. Anne MarieHamilton-Easton and Richard Ashmun for invaluable assis-tance with flow cytometry, Jennifer Peters and Sam Connellfor confocal microscopy, and the St. Jude Blood DonorCenter for help in obtaining samples. This work wassupported by the Assisi Foundation of Memphis, Program ofOncology Training Grant NCI 5R25CA023944, Cancer CenterSupport Grant 2P30CA021765 and the American Lebaneseand Syrian Associated Charities (ALSAC).
References
[1] J.L. Riley, C.H. June, B.R. Blazar, Human T regulatory celltherapy: take a billion or so and call me in the morning,Immunity 30 (2009) 656–665.
[2] F. Powrie, M.W. Leach, S. Mauze, L.B. Caddle, R.L. Coffman,Phenotypically distinct subsets of CD4+ T cells induce orprotect from chronic intestinal inflammation in C. B-17 scidmice, Int. Immunol. 5 (1993) 1461–1471.
[3] T. Mutis, R.S. van Rijn, E.R. Simonetti, T. Aarts-Riemens, M.E.Emmelot, L. van Bloois, A. Martens, L.F. Verdonck, S.B.Ebeling, Human regulatory T cells control xenogeneic graft-versus-host disease induced by autologous T cells in RAG2-/-gammac-/- immunodeficient mice, Clin. Cancer Res. 12 (2006)5520–5525.
[4] S. Hori, T. Nomura, S. Sakaguchi, Control of regulatory T celldevelopment by the transcription factor Foxp3, Science 299(2003) 1057–1061.
[5] J.S. Adams, M. Hewison, Unexpected actions of vitamin D: newperspectives on the regulation of innate and adaptiveimmunity, Nat. Clin. Pract. Endocrinol. Metab. 4 (2008) 80–90.
[6] A.K. Bhalla, E.P. Amento, T.L. Clemens, M.F. Holick, S.M. Krane,Specific high-affinity receptors for 1, 25-dihydroxyvitamin D3 inhuman peripheral blood mononuclear cells: presence in mono-cytes and induction inT lymphocytes following activation, J.Clin.Endocrinol. Metab. 57 (1983) 1308–1310.

220 V. Morales-Tirado et al.
[7] W.F. Rigby, S. Denome, M.W. Fanger, Regulation of lymphokineproduction and human T lymphocyte activation by 1, 25-dihydroxyvitamin D3. Specific inhibition at the level ofmessengerRNA, J. Clin. Invest. 79 (1987) 1659–1664.
[8] A. Boonstra, F.J. Barrat, C. Crain, V.L. Heath, H.F. Savelkoul,A. O'Garra, 1alpha, 25-Dihydroxyvitamin d3 has a direct effecton naive CD4(+) T cells to enhance the development of Th2cells, J. Immunol. 167 (2001) 4974–4980.
[9] N. Oberle, N. Eberhardt, C.S. Falk, P.H. Krammer, E. Suri-Payer,Rapid suppression of cytokine transcription in human CD4+CD25T cells by CD4+Foxp3+ regulatory T cells: independence of IL-2consumption, TGF-beta, and various inhibitors of TCR signaling,J. Immunol. 179 (2007) 3578–3587.
[10] J. Shaw, K. Meerovitch, R.C. Bleackley, V. Paetkau, Mechanismsregulating the level of IL-2 mRNA in T lymphocytes, J. Immunol.140 (1988) 2243–2248.
[11] M.K. Levings, R. Sangregorio, M.G. Roncarolo, Human cd25(+)cd4(+) t regulatory cells suppress naive and memory T cellproliferation and can be expanded in vitro without loss offunction, J. Exp. Med. 193 (2001) 1295–1302.
[12] KruisbeekA.M. , ShevachE. , ThorntonA.M. , Proliferative assaysfor T cell function, Curr. Protoc. Immunol. (2004)8 Chapter 3,Unit 3 12.
[13] A.M. Thornton, E.E. Donovan, C.A. Piccirillo, E.M. Shevach,Cutting edge: IL-2 is critically required for the in vitroactivation of CD4+CD25+ T cell suppressor function, J.Immunol. 172 (2004) 6519–6523.
[14] A.M. Thornton, E.M. Shevach, CD4+CD25+ immunoregulatoryT cells suppress polyclonal T cell activation in vitro by inhibitinginterleukin 2 production, J. Exp. Med. 188 (1998) 287–296.
[15] W. Liu, A.L. Putnam, Z. Xu-Yu, G.L. Szot, M.R. Lee, S. Zhu, P.A.Gottlieb, P. Kapranov, T.R. Gingeras, B. Fazekas de St Groth, C.Clayberger, D.M. Soper, S.F. Ziegler, J.A. Bluestone, CD127expression inversely correlates with FoxP3 and suppressivefunction of human CD4+ T reg cells, J. Exp. Med. 203 (2006)1701–1711.
[16] N. Seddiki, B. Santner-Nanan, J. Martinson, J. Zaunders, S.Sasson, A. Landay, M. Solomon, W. Selby, S.I. Alexander, R.Nanan, A. Kelleher, B. Fazekas de St Groth, Expression ofinterleukin (IL)-2 and IL-7 receptors discriminates betweenhuman regulatory and activated T cells, J. Exp. Med. 203 (2006)1693–1700.
[17] A. Krishan, Rapid flow cytofluorometric analysis of mammaliancell cycle by propidium iodide staining, J. Cell Biol. 66 (1975)188–193.
[18] M.A. Gavin, S.R. Clarke, E. Negrou, A. Gallegos, A. Rudensky,Homeostasis and anergy of CD4(+)CD25(+) suppressor T cells invivo, Nat. Immunol. 3 (2002) 33–41.
[19] M. Vukmanovic-Stejic, Y. Zhang, J.E. Cook, J.M. Fletcher, A.McQuaid, J.E. Masters, M.H. Rustin, L.S. Taams, P.C.Beverley, D.C. Macallan, A.N. Akbar, Human CD4+ CD25hiFoxp3+ regulatory T cells are derived by rapid turnover ofmemory populations in vivo, J. Clin. Invest. 116 (2006)2423–2433.
[20] M. Itoh, T. Takahashi, N. Sakaguchi, Y. Kuniyasu, J. Shimizu, F.Otsuka, S. Sakaguchi, Thymus and autoimmunity: production ofCD25+CD4+ naturally anergic and suppressive T cells as a keyfunction of the thymus inmaintaining immunologic self-tolerance,J. Immunol. 162 (1999) 5317–5326.
[21] L.S. Walker, A. Chodos, M. Eggena, H. Dooms, A.K. Abbas,Antigen-dependent proliferation of CD4+ CD25+ regulatoryT cells in vivo, J. Exp. Med. 198 (2003) 249–258.
[22] L. Klein, K. Khazaie, H. von Boehmer, In vivo dynamics ofantigen-specific regulatory T cells not predicted from behaviorin vitro, Proc. Natl Acad. Sci. USA 100 (2003) 8886–8891.
[23] Y. Kuniyasu, T. Takahashi, M. Itoh, J. Shimizu, G. Toda, S.Sakaguchi, Naturally anergic and suppressive CD25(+)CD4(+) T cells as a functionally and phenotypically distinct
immunoregulatory T cell subpopulation, Int. Immunol. 12(2000) 1145–1155.
[24] P. Banerjee, M. Chatterjee, Antiproliferative role of vitamin Dand its analogs—a brief overview, Mol. Cell. Biochem. 253(2003) 247–254.
[25] P.T. Liu, S. Stenger, H. Li, L. Wenzel, B.H. Tan, S.R. Krutzik,M.T. Ochoa, J. Schauber, K. Wu, C. Meinken, D.L. Kamen, M.Wagner, R. Bals, A. Steinmeyer, U. Zugel, R.L. Gallo, D.Eisenberg, M. Hewison, B.W. Hollis, J.S. Adams, B.R. Bloom,R.L. Modlin, Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response, Science 311 (2006)1770–1773.
[26] J. Rosenblatt, A. Bissonnette, R. Ahmad, Z. Wu, B. Vasir, K.Stevenson, C. Zarwan, W. Keefe, B. Glotzbecker, H. Mills, R.Joyce, J.D. Levine, D. Tzachanis, V. Boussiotis, D. Kufe, D.Avigan, Immunomodulatory effects of vitamin D: implications forGVHD, Bone Marrow Transplant 45 (2010) 1463–1468.
[27] H.F. Deluca, M.T. Cantorna, Vitamin D: its role and uses inimmunology, FASEB J. 15 (2001) 2579–2585.
[28] C.M. Veldman, M.T. Cantorna, H.F. DeLuca, Expression of 1,25-dihydroxyvitamin D(3) receptor in the immune system,Arch. Biochem. Biophys. 374 (2000) 334–338.
[29] C.S. Johnson, J.R. Muindi, P.A. Hershberger, D.L. Trump, Theantitumor efficacy of calcitriol: preclinical studies, AnticancerRes. 26 (2006) 2543–2549.
[30] D.L. Trump, P.A. Hershberger, R.J. Bernardi, S. Ahmed, J.Muindi, M. Fakih, W.D. Yu, C.S. Johnson, Anti-tumor activity ofcalcitriol: pre-clinical and clinical studies, J. Steroid Biochem.Mol. Biol. 89–90 (2004) 519–526.
[31] J.D. Fontenot, M.A. Gavin, A.Y. Rudensky, Foxp3 programs thedevelopment and function of CD4+CD25+ regulatory T cells,Nat. Immunol. 4 (2003) 330–336.
[32] H. Yagi, T. Nomura, K. Nakamura, S. Yamazaki, T. Kitawaki, S.Hori, M. Maeda, M. Onodera, T. Uchiyama, S. Fujii, S.Sakaguchi, Crucial role of FOXP3 in the development andfunction of human CD25+CD4+ regulatory T cells, Int. Immunol.16 (2004) 1643–1656.
[33] M.E. Brunkow, E.W. Jeffery, K.A. Hjerrild, B. Paeper, L.B.Clark, S.A. Yasayko, J.E. Wilkinson, D. Galas, S.F. Ziegler, F.Ramsdell, Disruption of a new forkhead/winged-helix protein,scurfin, results in the fatal lymphoproliferative disorder of thescurfy mouse, Nat. Genet. 27 (2001) 68–73.
[34] C.L. Bennett, J. Christie, F. Ramsdell, M.E. Brunkow, P.J.Ferguson, L. Whitesell, T.E. Kelly, F.T. Saulsbury, P.F. Chance,H.D. Ochs, The immune dysregulation, polyendocrinopathy,enteropathy, X-linked syndrome (IPEX) is caused by mutationsof FOXP3, Nat. Genet. 27 (2001) 20–21.
[35] L.A. Schubert, E. Jeffery, Y. Zhang, F. Ramsdell, S.F. Ziegler,Scurfin (FOXP3) acts as a repressor of transcription andregulates T cell activation, J. Biol. Chem. 276 (2001)37672–37679.
[36] M.R. Walker, D.J. Kasprowicz, V.H. Gersuk, A. Benard, M. VanLandeghen, J.H. Buckner, S.F. Ziegler, Induction of FoxP3 andacquisition of T regulatory activity by stimulated human CD4+CD25- T cells, J. Clin. Invest. 112 (2003) 1437–1443.
[37] J.E. Snyder, W.J. Bowers, A.M. Livingstone, F.E. Lee, H.J.Federoff, T.R. Mosmann, Measuring the frequency of mouseand human cytotoxic T cells by the Lysispot assay: independentregulation of cytokine secretion and short-term killing, Nat.Med. 9 (2003) 231–235.
[38] Z. Urry, E. Xystrakis, D.F. Richards, J. McDonald, Z. Sattar, D.J.Cousins, C.J. Corrigan, E. Hickman, Z. Brown, C.M. Hawrylowicz,Ligation of TLR9 induced on human IL-10-secreting Tregs by1alpha, 25-dihydroxyvitamin D3 abrogates regulatory function,J. Clin. Invest. 119 (2009) 387–398.
[39] C. Baecher-Allan, E. Wolf, D.A. Hafler, MHC class II expressionidentifies functionally distinct human regulatory T cells,J. Immunol. 176 (2006) 4622–4631.

2211α,25-dihydroxyvitamin D3
[40] P. Hoffmann, R. Eder, T.J. Boeld, K. Doser, B. Piseshka, R.Andreesen, M. Edinger, Only the CD45RA+subpopulation ofCD4+CD25high T cells gives rise to homogeneous regulatoryT-cell lines upon in vitro expansion, Blood 108 (2006)4260–4267.
[41] S.E. Allan, S.Q. Crome, N.K. Crellin, L. Passerini, T.S. Steiner,R. Bacchetta, M.G. Roncarolo, M.K. Levings, Activation-inducedFOXP3 in human T effector cells does not suppress proliferationor cytokine production, Int. Immunol. 19 (2007) 345–354.
[42] R. Khattri, T. Cox, S.A. Yasayko, F. Ramsdell, An essential rolefor Scurfin in CD4+CD25+ T regulatory cells, Nat. Immunol. 4(2003) 337–342.
[43] F. Ocklenburg, D. Moharregh-Khiabani, R. Geffers, V. Janke, S.Pfoertner, H. Garritsen, L. Groebe, J. Klempnauer, K.E.Dittmar, S. Weiss, J. Buer, M. Probst-Kepper, UBD, adownstream element of FOXP3, allows the identification ofLGALS3, a new marker of human regulatory T cells, Lab.Investig. 86 (2006) 724–737.
[44] S.F. Ziegler, FOXP3: of mice and men, Annu. Rev. Immunol. 24(2006) 209–226.
[45] S.E. Allan, L. Passerini, R. Bacchetta, N. Crellin, M. Dai, P.C.Orban, S.F. Ziegler, M.G. Roncarolo, M.K. Levings, The role of 2FOXP3 isoforms in the generation of human CD4+ Tregs, J. Clin.Invest. 115 (2005) 3276–3284.
[46] S.E. Allan, A.N. Alstad, N. Merindol, N.K. Crellin, M. Amendola,R. Bacchetta, L. Naldini, M.G. Roncarolo, H. Soudeyns, M.K.Levings, Generation of potent and stable human CD4+ T
regulatory cells by activation-independent expression ofFOXP3, Mol. Ther. 16 (2008) 194–202.
[47] G. Lal, N. Zhang, W. van der Touw, Y. Ding, W. Ju, E.P.Bottinger, S.P. Reid, D.E. Levy, J.S. Bromberg, Epigeneticregulation of Foxp3 expression in regulatory T cells by DNAmethylation, J. Immunol. 182 (2009) 259–273.
[48] U. Baron, S. Floess, G. Wieczorek, K. Baumann, A. Grutzkau, J.Dong, A. Thiel, T.J. Boeld, P. Hoffmann,M. Edinger, I. Turbachova,A. Hamann, S. Olek, J. Huehn, DNA demethylation in the humanFOXP3 locus discriminates regulatory T cells from activated FOXP3(+) conventional T cells, Eur. J. Immunol. 37 (2007) 2378–2389.
[49] S. Floess, J. Freyer, C. Siewert, U. Baron, S. Olek, J. Polansky,K. Schlawe, H.D. Chang, T. Bopp, E. Schmitt, S. Klein-Hessling,E. Serfling, A. Hamann, J. Huehn, Epigenetic control of thefoxp3 locus in regulatory T cells, PLoS Biol. 5 (2007) e38.
[50] J.J. Coenen, H.J. Koenen, E. van Rijssen, L.B. Hilbrands, I.Joosten, Rapamycin, and not cyclosporin A, preserves the highlysuppressive CD27+ subset of human CD4+CD25+ regulatoryT cells, Blood 107 (2006) 1018–1023.
[51] M. Battaglia, A. Stabilini, M.G. Roncarolo, Rapamycin selectivelyexpands CD4+CD25+FoxP3+ regulatory T cells, Blood 105 (2005)4743–4748.
[52] R. Tao, E.F. de Zoeten, E. Ozkaynak, C. Chen, L. Wang, P.M.Porrett, B. Li, L.A. Turka, E.N. Olson, M.I. Greene, A.D. Wells,W.W. Hancock, Deacetylase inhibition promotes the generationand function of regulatory T cells, Nat. Med. 13 (2007)1299–1307.
Copyright © 2022 FDOKUMEN



![D3: e`ad T`ca`cReV ]`R_ hRZgVc ]Zde - SPLessons](https://static.fdokumen.com/doc/165x107/6322423f28c445989105b166/d3-ead-tcacrev-r-hrzgvc-zde-splessons.jpg)



![D2/D3 dopamine receptor binding with [F-18]fallypride in thalamus and cortex of patients with schizophrenia](https://static.fdokumen.com/doc/165x107/633625b864d291d2a302c45f/d2d3-dopamine-receptor-binding-with-f-18fallypride-in-thalamus-and-cortex-of.jpg)





![[3H]4-(dimethylamino)-N-(4-(4-(2-methoxyphenyl)piperazin-1-yl) butyl)benzamide: A selective radioligand for dopamine D3 receptors. II. Quantitative analysis of dopamine D3 and D2 receptor](https://static.fdokumen.com/doc/165x107/6335c3b902a8c1a4ec01e90f/3h4-dimethylamino-n-4-4-2-methoxyphenylpiperazin-1-yl-butylbenzamide.jpg)






