
Comparative study on antioxidant effects and vascular matrix metalloproteinase-2 downregulation by...
10
ORIGINAL ARTICLE Comparative study on antioxidant effects and vascular matrix metalloproteinase-2 downregulation by dihydropyridines in renovascular hypertension Diogo M. O. Marçal & Elen Rizzi & Alisson Martins-Oliveira & Carla S. Ceron & Danielle A. Guimaraes & Raquel F. Gerlach & Jose E. Tanus-Santos Received: 19 August 2010 / Accepted: 17 October 2010 / Published online: 31 October 2010 # Springer-Verlag 2010 Abstract The vascular remodeling associated with hyperten- sion involves oxidative stress and enhanced matrix metal- loproteinases (MMPs) expression/activity, especially MMP-2. While previous work showed that lercanidipine, a third- generation dihydropyridine calcium channel blocker (CCB), attenuated the oxidative stress and increased MMP-2 expres- sion/activity in two-kidney, one-clip (2K1C) hypertension, no previous study has examined whether first- or second- generation dihydropyridines produce similar effects. We compared the effects of nifedipine, nimodipine, and amlodipine on 2K1C hypertension-induced changes in systolic blood pressure (SBP), vascular remodeling, oxidative stress, and MMPs levels/activity. Sham-operated and 2K1C rats were treated with water, nifedipine 10 mg/kg/day, nimodipine 15 mg/kg/day, or amlodipine 10 mg/kg/day by gavage, starting 3 weeks after hypertension was induced. SBP was monitored weekly. After 6 weeks of treatment, quantitative morphometry of structural changes in the aortic wall was studied in hematoxylin/eosin-stained sections. Aortic and systemic reac- tive oxygen species levels were measured by using dihydroe- thidine and thiobarbituric acid-reactive substances (TBARs), respectively. Aortic MMP-2 levels and activity were deter- mined by gelatin zymography, in situ zymography, and immunofluorescence. Nifedipine, nimodipine, or amlodipine attenuated the increases in SBP in hypertensive rats by approximately 17% (P <0.05) and prevented vascular hyper- trophy (P <0.05). These CCBs blunted 2K1C-induced increases in vascular oxidative stress and plasma TBARs concentrations (P <0.05). All dihydropyridines attenuated the increases in aortic MMP-2 levels and activity associated with 2K1C hypertension. These findings suggest lack of superiority of one particular dihydropyridine, at least with respect to antioxidant effects, MMPs downregulation, and inhibition of vascular remodeling in hypertension. Keywords 2K1C hypertension . Calcium channel blockers . Dihydropyridines . Matrix metalloproteinases . Oxidative stress . Renovascular hypertension Introduction Hypertension clearly leads to structural and functional cardio- vascular modifications that can be prevented by pharmacolog- ical therapy. In this regard, drugs targeting relevant pathophysiological processes may reduce the incidence of cardiovascular events associated with hypertension (Cohuet and Struijker-Boudier 2006). Importantly, accumulating evi- dence implicates a group of zinc-dependent endopeptidases named matrix metalloproteinases (MMPs) in the vascular alterations associated with many cardiovascular conditions including hypertension (Galis and Khatri 2002). Upregulated MMPs promote excessive degradation of extracellular matrix and are involved in pathological vascular remodeling (Sluijter et al. 2006), which includes vascular smooth muscle cell migration and proliferation in the arterial wall (Newby 2006). D. M. O. Marçal : E. Rizzi : A. Martins-Oliveira : C. S. Ceron : D. A. Guimaraes : J. E. Tanus-Santos (*) Department of Pharmacology, Faculty of Medicine of Ribeirão Preto, University of São Paulo, Av. Bandeirantes, 3900, 14049-900 Ribeirão Preto, SP, Brazil e-mail: [email protected] J. E. Tanus-Santos e-mail: [email protected] R. F. Gerlach Department of Morphology, Estomatology and Physiology, Dental School of Ribeirão Preto, University of São Paulo, Av. Bandeirantes, 3900, Ribeirão Preto, SP, Brazil 14049-900 Naunyn-Schmied Arch Pharmacol (2011) 383:35–44 DOI 10.1007/s00210-010-0573-y
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Comparative study on antioxidant effects and vascular matrix metalloproteinase-2 downregulation by...

ORIGINAL ARTICLE
Comparative study on antioxidant effects and vascularmatrix metalloproteinase-2 downregulationby dihydropyridines in renovascular hypertension
Diogo M. O. Marçal & Elen Rizzi & Alisson Martins-Oliveira & Carla S. Ceron &
Danielle A. Guimaraes & Raquel F. Gerlach & Jose E. Tanus-Santos
Received: 19 August 2010 /Accepted: 17 October 2010 /Published online: 31 October 2010# Springer-Verlag 2010
Abstract The vascular remodeling associated with hyperten-sion involves oxidative stress and enhanced matrix metal-loproteinases (MMPs) expression/activity, especially MMP-2.While previous work showed that lercanidipine, a third-generation dihydropyridine calcium channel blocker (CCB),attenuated the oxidative stress and increased MMP-2 expres-sion/activity in two-kidney, one-clip (2K1C) hypertension, noprevious study has examined whether first- or second-generation dihydropyridines produce similar effects. Wecompared the effects of nifedipine, nimodipine, and amlodipineon 2K1C hypertension-induced changes in systolic bloodpressure (SBP), vascular remodeling, oxidative stress, andMMPs levels/activity. Sham-operated and 2K1C rats weretreated with water, nifedipine 10 mg/kg/day, nimodipine15 mg/kg/day, or amlodipine 10 mg/kg/day by gavage, starting3 weeks after hypertension was induced. SBP was monitoredweekly. After 6 weeks of treatment, quantitative morphometryof structural changes in the aortic wall was studied inhematoxylin/eosin-stained sections. Aortic and systemic reac-tive oxygen species levels were measured by using dihydroe-thidine and thiobarbituric acid-reactive substances (TBARs),
respectively. Aortic MMP-2 levels and activity were deter-mined by gelatin zymography, in situ zymography, andimmunofluorescence. Nifedipine, nimodipine, or amlodipineattenuated the increases in SBP in hypertensive rats byapproximately 17% (P<0.05) and prevented vascular hyper-trophy (P<0.05). These CCBs blunted 2K1C-inducedincreases in vascular oxidative stress and plasma TBARsconcentrations (P<0.05). All dihydropyridines attenuated theincreases in aortic MMP-2 levels and activity associated with2K1C hypertension. These findings suggest lack of superiorityof one particular dihydropyridine, at least with respect toantioxidant effects, MMPs downregulation, and inhibition ofvascular remodeling in hypertension.
Keywords 2K1C hypertension . Calcium channelblockers . Dihydropyridines .Matrix metalloproteinases .
Oxidative stress . Renovascular hypertension
Introduction
Hypertension clearly leads to structural and functional cardio-vascular modifications that can be prevented by pharmacolog-ical therapy. In this regard, drugs targeting relevantpathophysiological processes may reduce the incidence ofcardiovascular events associated with hypertension (Cohuetand Struijker-Boudier 2006). Importantly, accumulating evi-dence implicates a group of zinc-dependent endopeptidasesnamed matrix metalloproteinases (MMPs) in the vascularalterations associated with many cardiovascular conditionsincluding hypertension (Galis and Khatri 2002). UpregulatedMMPs promote excessive degradation of extracellular matrixand are involved in pathological vascular remodeling (Sluijteret al. 2006), which includes vascular smooth muscle cellmigration and proliferation in the arterial wall (Newby 2006).
D. M. O. Marçal : E. Rizzi :A. Martins-Oliveira : C. S. Ceron :D. A. Guimaraes : J. E. Tanus-Santos (*)Department of Pharmacology, Faculty of Medicine of RibeirãoPreto, University of São Paulo,Av. Bandeirantes, 3900,14049-900 Ribeirão Preto, SP, Brazile-mail: [email protected]
J. E. Tanus-Santose-mail: [email protected]
R. F. GerlachDepartment of Morphology, Estomatology and Physiology,Dental School of Ribeirão Preto, University of São Paulo,Av. Bandeirantes, 3900,Ribeirão Preto, SP, Brazil 14049-900
Naunyn-Schmied Arch Pharmacol (2011) 383:35–44DOI 10.1007/s00210-010-0573-y

Enhanced expression and activity of MMPs have beenconsistently implicated in vascular remodeling associated withhypertension in patients (Yasmin et al. 2005; Ahmed et al.2006) and animal models (Bouvet et al. 2005; Watts et al.2007; Lehoux et al. 2004; Flamant et al. 2007; Martinez et al.2008a; Castro et al. 2008, 2010). We have recently shown thatincreased vascular MMP-2 levels and activity may lead toimpaired endothelial-dependent vasorelaxation, arterial wallhypertrophy, and excessive collagen and elastin deposition intwo-kidney, one-clip (2K1C) hypertension, thereby contribut-ing to hypertensive vascular remodeling and dysfunction(Castro et al. 2008).
Although MMPs are regulated by their endogenous inhib-itors (tissue inhibitors of metalloproteinases (TIMPs)), in-creased formation of reactive oxygen species (ROS) is amajor factor activating MMPs expression/activity (Grote et al.2003; Nelson and Melendez 2004; Ra and Parks 2007), andincreased oxidative stress has been consistently shown inanimal models and in clinical hypertension (Griendling andFitzGerald 2003). Moreover, antioxidant drugs downregulatevascular MMP-2 expression and prevent the development ofstructural vascular changes associated with 2K1C hyperten-sion (Castro et al. 2009). Therefore, targeting MMPs (Raffettoand Khalil 2008), especially MMP-2 (Castro et al. 2008), maybe a promising therapeutic aim in the therapy or prevention ofthe alterations caused by hypertension.
In line with this suggestion, we have shown thatlercanidipine, a dihydropyridine calcium channel blocker(CCB) used in the therapy hypertension, produces pleiotropiceffects that may add cardiovascular protection to its directantihypertensive effects. Indeed, we found that lercanidipineexerts antioxidant effects, reduces MMPs activity/expression,and improves vascular function (Martinez et al. 2006, 2008a,b). However, it is not known whether other dihydropyridineCCBs can exert effects similar to those exerted bylercanidipine. In the present study, we compared the effectsof first-, second-, and third-generation dihydropyridineCCBs (nifedipine, nimodipine, and amlodipine, respectively)on 2K1C hypertension-induced changes in arterial bloodpressure, vascular remodeling, oxidative stress, and MMPslevels/activity.
Methods
Animals and treatments
This study complied with guidelines of the Faculty ofMedicine of Ribeirão Preto, University of São Paulo, andthe animals were handled according to the guidingprinciples published by the National Institutes of HealthGuide for the Care and Use of Laboratory Animals. MaleWistar rats (180–200 g) obtained from the colony at
University of São Paulo were maintained on 12-h light/dark cycle at 25°C with free access to rat chow and water.
2K1C hypertension was induced by clipping the left renalartery with a silver clip (0.2 mm). Sham-operated ratsunderwent the same surgical procedure (under generalanesthesia with ketamine 100 mg/kg and xylazine 10 mg/kgi.p.), except for the placement of the renal artery clip. Threeweeks after surgery, the animals were randomly assigned toone of eight groups: 2K1C and Sham groups that received tapwater; 2K1C and Sham groups that received nifedipine 10 mg/kg/day; 2K1C and Sham groups that received nimodipine15 mg/kg/day; and 2K1C and Sham groups that receivedamlodipine 10mg/kg/day. The animals were treatedwith wateror dihydropyridines by gavage for additional 6 weeks. Thedoses of CCBs used in the present study were based onprevious studies showing theses doses reduced systolic bloodpressure to similar extents in spontaneously hypertensive rats(approximately by 40 mmHg) (Tomassoni et al. 2003).
Body weight and tail systolic blood pressure were assessedweekly throughout the experiment period. The systolic bloodpressure was measured by tail-cuff plethysmography inconscious rats (Castro et al. 2004).
Morphometric analysis of the vascular wall
After six treatments with tap water or dihydropyridines, the ratswere killed by decapitation. Their thoracic aorta wereharvested, connective tissue cleaned off, immediately fixed in4% phosphate-buffered paraformaldehyde, pH 7.4, and em-bedded in paraffin blocks. Four-micrometer-thick slices werestainedwith hematoxylin and eosin.Media cross-sectional area(CSA) was calculated by subtracting the lumen internal area(Ai) from the external area (Ae), which were measured intissue sections (×250; Castro et al. 2008). The externaldiameter (ED) and the internal diameter (ID) were calculatedas the square root of 4Ae/π and 4Ai/π, respectively. Mediathickness (M) was calculated as (ED−ID)/2. Finally, M tolumen diameter (M/L) was also calculated. The number ofvascular smooth muscle cells in the aortic wall was measuredby the tridimensional dissector method on two consecutivesections, as previously described (Castro et al. 2008). Thismethod is independent of nuclei orientation, form, and size.Stained sections were examined with light microscopy(DMLB; Leica, Bensheim, Germany), and the image wascaptured at ×400, as previously described (Castro et al. 2008).These structural analyses in the media were evaluated byusing ImageJ Program (National Institute of Health).
Assessment of vascular ROS and lipid peroxide levelsin plasma
Dihydroethidium (DHE), a sensitive superoxide probe, wasused to evaluate in situ production of ROS. Briefly, aortic
36 Naunyn-Schmied Arch Pharmacol (2011) 383:35–44

tissues were embedded vertically in Tissue-tek and thenfrozen and cut in serial 4-μm sections. Unfixed cryosec-tions were incubated at room temperature, in the dark, with10 μL of DHE (10 μmol/L) for 30 min (Castro et al. 2009).Sections were examined by fluorescence microscopy (LeicaImaging Systems Ltd., Cambridge, England), and the imagewas captured at ×400. Red fluorescence reflected superox-ide production and was evaluated by using the ImageJprogram (National Institutes of Health). We measured redfluorescence from 20 fields selected around the vesselcircumference, and the arithmetic mean of the fluorescencefrom the 20 fields was calculated for each slide (n=5–6;Castro et al. 2009).
Plasma lipid peroxide levels were determined bymeasuring thiobarbituric acid-reactive substances using afluorimetric method as previously described (Dias-Junior etal. 2008). This method requires excitation at 515 nm andemission at 553 nm and uses 1,1,3,3-tetramethoxypropaneas standard. The lipoperoxide levels were expressed interms of malondialdehyde (MDA; nmol/mL).
Measurement of aortic MMP-2 levels by gelatinzymography
Gelatin zymography was performed as previously described(Martinez et al. 2008a, b). Briefly, aortic extracts weresubjected to electrophoresis on 12% SDS-PAGE copoly-merized with gelatin (1%). Gelatinolytic activities weredetected as unstained bands against the background ofCoomassie-blue-stained gelatin, assayed densitometry usinga Kodak Electrophoresis Documentation and AnalysisSystem 290 (Kodak, Rochester, NY, USA).
Immunofluorescence for MMP-2
Aortic MMP-2 expression was examined by immunofluores-cence as previously described (Castro et al. 2010). Afteradding DQ gelatin, tissue sections were incubated withMMP-2 primary mouse anti-human monoclonal antibody(MAB3308; Chemicon, Temecula, CA, USA) for 1 h in darkhumidified chambers [MMP-2 (1:1,000)]. Red fluorescencewas visualized by adding rhodamine conjugated as secondaryantibody (1:200; AP160P; Chemicon) for 1 h. To confirm thespecificity of antibodies, the primary antibody was omittedand substituted by PBS +1% bovine serum albumin.Rhodamine did not bind nonspecifically to tissue sections.
In situ zymography
MMP activities in the media and intimae of frozen thoracicaortas were measured using DQ gelatin (E12055; MolecularProbes, Oregon, USA) as a fluorogenic substrate. Briefly,aortic tissues were embedded vertically in Tissue-tek and then
frozen and cut in serial 4 μm sections. Vessel sections wereincubated in dark humidified chambers for 1 h with 1.0 μg/mLDQ gelatin in Tris–CaCl2 buffer (50 mM Tris, 10 mM CaCl2,1 μM ZnCl2). The sections were examined with fluorescencemicroscopy (Leica Imaging Systems Ltd., Cambridge, Eng-land), and the image was captured at ×400. Proteolyticactivity was detected as bright green fluorescence, whichindicates substrate breakdown, and was evaluated by usingthe ImageJ program (National Institutes of Health, USA). Theassessment of gelatinolytic activity was made by quantifyingthe intensity of green fluorescence from 20 fields selectedaround the vessel circumference. The arithmetic mean of thefluorescence from the 20 fields was calculated for each slide(n=8–10). This number of fields per slide corresponds toapproximately 20–30% of the total aortic area being studiedand has led to interassay coefficients of variations of less than3%. Phenanthroline 1 mM and PMSF 1 mM were used toconfirm the activity of MMPs in thoracic aorta. Whereasphenanthroline fully inhibited MMP activity, PMSF producedno significant effects (data not shown).
Statistical analysis
Results are expressed as means±SEM. Between groups,comparisons were assessed by two- or one-way ANOVAfollowed by the Tukey test using GraphPad Prism software. Aprobability value <0.05 was considered significant.
Results
Effects of dihydropyridines on SBP levels in 2K1C rats
Baseline SBP and body weight (data not shown) were similarin the eight experimental groups (Fig. 1). Whereas nosignificant changes in SBP were seen in the sham and
0 1 2 3 4 5 6 7 8 9 1050
100
150
200
250
ShamSham+NifeSham+Nimo
Sham+Amlo
2K1C2K1C+Nife2K1C+Nimo
2K1C+Amlo*
** * * * * *
** ** ** **
Dihydropyridines or Vehicle
Time (weeks)
Sys
tolic
blo
od
pre
ssu
re (
mm
Hg
)
*
Fig. 1 Systolic blood pressure (mmHg) measured by tail-cuff methodin the eight experimental groups during the study period. Data areshown as mean±SEM; (n=10–13 per group). *P<0.01 for 2K1C and2K1C+dihydropyridines groups versus respective Sham groups. **P<0.05 versus 2K1C group
Naunyn-Schmied Arch Pharmacol (2011) 383:35–44 37

sham+dihydropyridines groups, SBP increased in the 2K1Cgroup (Fig. 1). Treatment with nifedipine, nimodipine, oramlodipine significantly attenuated the increases in SBPin hypertensive rats (SBP=174±4, 173±5, and 171±7 mmHg, respectively, versus 208±4 mmHg in hyper-tensive controls at week 9; all P<0.05; Fig. 1). Nosignificant differences in body weight were observedamong the experimental groups (data not shown).
Effects of dihydropyridines on vascular remodelingin 2K1C rats
Renovascular hypertension was associated with arterialwall hypertrophy, with significant increases in thenumber of VSMCs, increased aortic CSA, and increasedM/L ratio (P<0.05; Fig. 2a–d). Treatment of hypertensiverats with nifedipine, nimodipine, or amlodipine prevented
a
b
c
d
Sham+Nife
200µm 200µm
Sham+Nimo
200µm
Sham+Amlo
200µm
Sham
2K1C
200µm
2K1C+Nife
200µm
2K1C+Nimo
200µm
2K1C+Amlo
200µm
Sham
Sham
+Nife
Sham
+Nim
o
Sham
+Am
lo2K
1C
2K1C
+Nife
2K1C
+Nim
o
2K1C
+Am
lo0
50
100
150
*
Cel
ls/µ
m a
ort
a
Sham
Sham
+Nife
Sham
+Nim
o
Sham
+Am
lo2K
1C
2K1C
+Nife
2K1C
+Nim
o
2K1C
+Am
lo0
50000
100000
150000
200000
250000 *
CS
A (
µm2 )
Sham
Sham
+Nife
Sham
+Nim
o
Sham
+Am
lo2K
1C
2K1C
+Nife
2K1C
+Nim
o
2K1C
+Am
lo0
5
10
15
20 *
Med
ia/lu
men
(%
)
Fig. 2 Effects of dihydropyri-dines on aortic structural mod-ifications associated with 2K1Chypertension. a Representativephotographs of aortic samples(×400) stained by hematoxylinand eosin. b–d Values for vas-cular smooth muscle cell num-ber per length of aorta, medialCSA, and media to lumen ratio(M/L). Data are shown as mean±SEM (n=5–8 per group). *P<0.05 versus sham groups and2K1C+dihydropyridine groups
38 Naunyn-Schmied Arch Pharmacol (2011) 383:35–44

these histological changes associated with 2K1C hyper-tension (P<0.05; Fig. 2a–d).
Effects of dihydropyridines on vascular production of ROSand lipid peroxide levels in plasma from 2K1C rats
The vascular production of ROS was assessed by fluores-cence microscopy using DHE in isolated aortic segments.We found increased ROS levels in the media of thoracicaorta from 2K1C rats compared with the sham groups (P<0.05; Fig. 3a, b). Treatment of 2K1C rats with dihydropyr-idines blunted 2K1C-induced increases in oxidative stress(P<0.05; Fig. 3a, b).
Giving support to these findings, the plasma MDA levelswere significantly higher in the 2K1C group compared withthe sham groups (P<0.05; Fig. 3c). Treatment withnifedipine, nimodipine, or amlodipine blunted 2K1C-induced increases in oxidative stress, as suggested by lowerplasma lipid peroxide levels in the 2K1C+dihydropyridinesgroups (P<0.05; Fig. 3c).
Effects of dihydropyridines on aortic MMP-2 levelsin 2K1C rats
Gelatin zymography was used to assess MMP levels inaortic extracts. Figure 4a shows a representative zymogram
a
b
c
Sham+NimoSham
200µm
Sham+Nife
200µm 200µm
2K1C+Nife
200µm
2K1C+Nimo
200µm 200µm
2K1C
Sham+Amlo
200µm
2K1C+Amlo
200µm
Sham
Sham
+Nife
Sham
+Nim
o
Sham
+Am
lo2K
1C
2K1C
+Nife
2K1C
+Nim
o
2K1C
+Am
lo0
5
10
15*
Flu
ore
scen
t in
ten
sity
(Arb
itra
ry U
nit
s)
Sham
Sham
+Nife
Sham
+Nim
o
Sham
+Am
lo2K
1C
2K1C
+Nife
2K1C
+Nim
o
2K1C
+Am
lo0
5
10
15 *
Pla
sma
MD
A (n
mo
l/mL
)
Fig. 3 Effects of dihydropyri-dines on vascular ROS produc-tion. a Representativefluorescence photomicrographs(×400) of cryosections of DHE-aortic samples. Red fluorescencerepresents superoxide produc-tion. b The quantification ofaortic fluorescence (n=5–6 pergroup). c The effects of dihy-dropyridines on plasma ROSconcentrations. Thiobarbituricacid-reactive substances areexpressed in terms of MDAconcentrations in plasma sam-ples from the rats (n=8–10 pergroup). Data are shown as mean±SEM. *P<0.05 versus shamgroups and 2K1C+dihydropyri-dine groups
Naunyn-Schmied Arch Pharmacol (2011) 383:35–44 39

of aortic extracts with bands corresponding to molecularweights of the MMP-2 bands (75, 72, and 64 kDa; Fig. 4a).Aortas from 2K1C rats showed higher levels of the threeMMP-2 forms (75, 72, and 64 kDa) compared with the shamgroups (P<0.05; Fig. 4b–d). Treatment with nifedipine and
nimodipine, but not with amlodipine, significantly attenuated2K1C hypertension-induced increases in the 75- and 72-kDaMMP-2 forms (P<0.05; Fig. 4b–d). However, all dihydro-pyridines attenuated the increases in the 64-kDa MMP-2form associated with 2K1C hypertension (P<0.05; Fig. 4d).
a
Std Vehicle Nife Nimo Amlo Vehicle Nife Nimo Amlo
Sham 2K1C
b
c
d
75 kDa72 kDa64 kDa
Sham
Sham
+Nife
Sham
+Nim
o
Sham
+Am
lo2K
1C
2K1C
+Nife
2K1C
+Nim
o
2K1C
+Am
lo0
1
2
3
4
* #
75 K
Da
MM
P-2
(Arb
itrar
y U
nits
)
Sham
Sham
+Nife
Sham
+Nim
o
Sham
+Am
lo2K
1C
2K1C
+Nife
2K1C
+Nim
o
2K1C
+Am
lo0
5
10
15
*
72 K
Da
MM
P-2
(Arb
itrar
y U
nits
)
Sham
Sham
+Nife
Sham
+Nim
o
Sham
+Am
lo2K
1C
2K1C
+Nife
2K1C
+Nim
o
2K1C
+Am
lo0
1
2
3
4
5
*
64 K
Da
MM
P-2
(Arb
itra
ry U
nit
s)
Fig. 4 a A representative SDS-PAGE gelatin zymogram ofaortic samples. Molecularweights of MMP-2 bands wereidentified after electrophoresis.Std internal standard. b–d Thevalues for each molecularweight form of MMP-2 (75, 72,and 64 kDa, respectively) in theaortas. Data are shown as mean±SEM (n=8–14 per group). *P<0.05 versus all the other ex-perimental groups, but not ver-sus the 2K1C+Amlo group in band c. #P<0.05 versus Sham+Amlo group
40 Naunyn-Schmied Arch Pharmacol (2011) 383:35–44
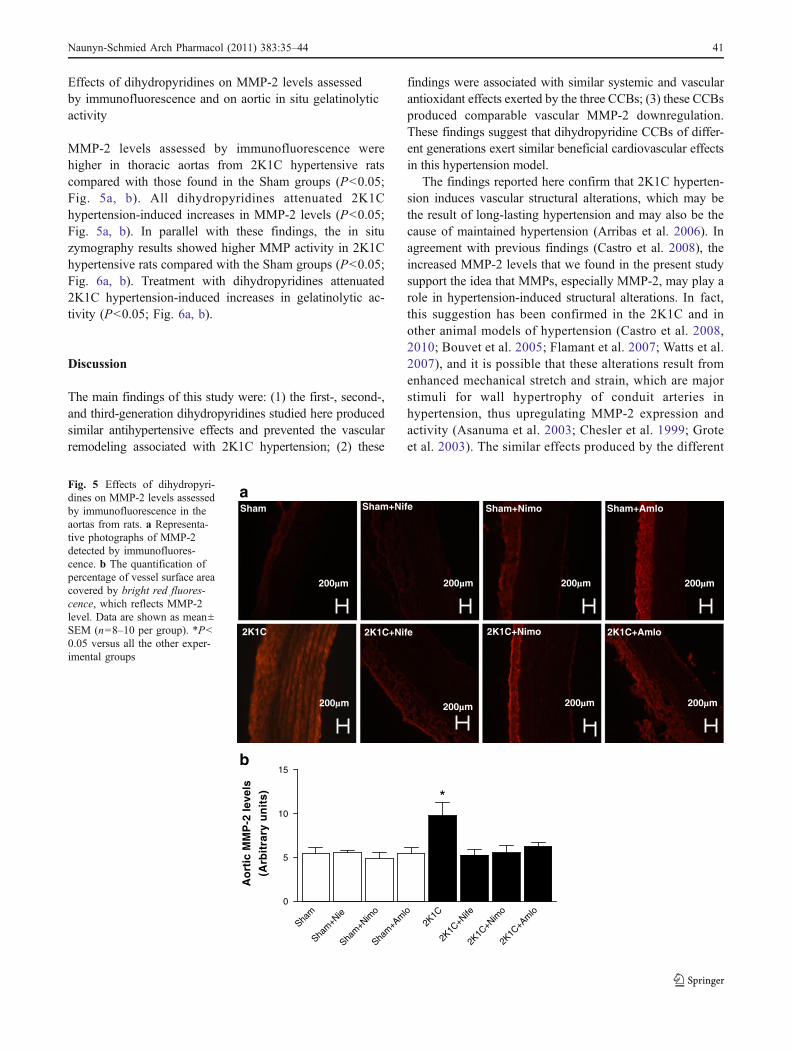
Effects of dihydropyridines on MMP-2 levels assessedby immunofluorescence and on aortic in situ gelatinolyticactivity
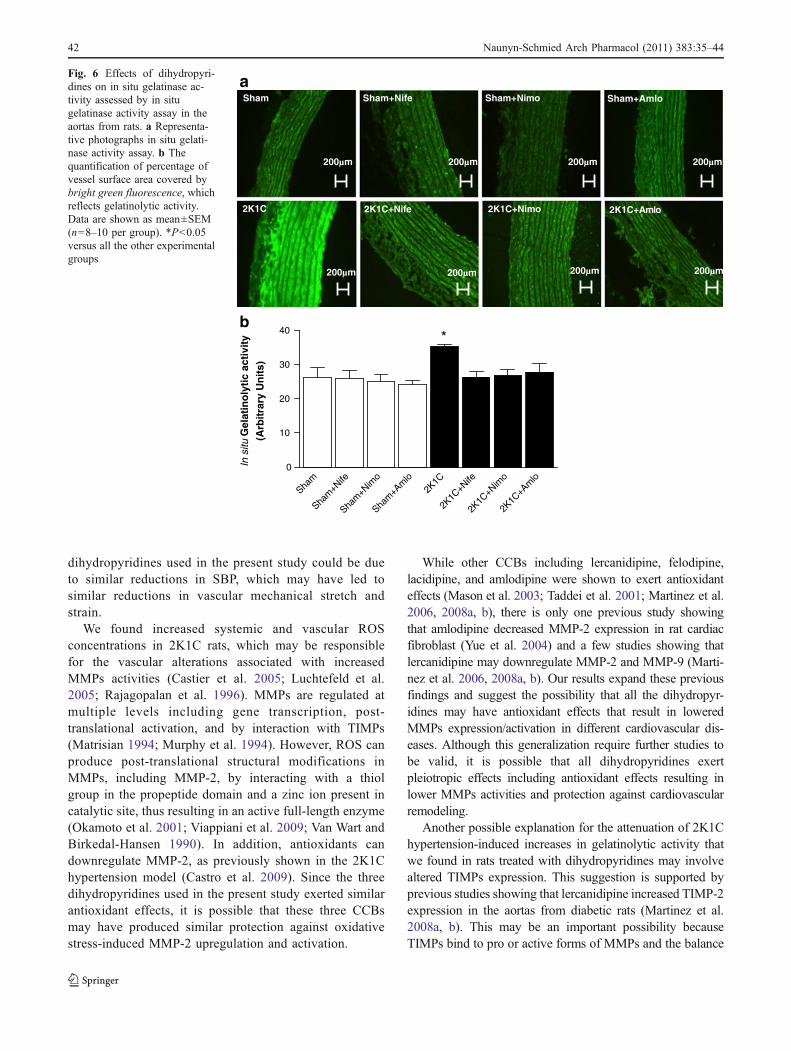
MMP-2 levels assessed by immunofluorescence werehigher in thoracic aortas from 2K1C hypertensive ratscompared with those found in the Sham groups (P<0.05;Fig. 5a, b). All dihydropyridines attenuated 2K1Chypertension-induced increases in MMP-2 levels (P<0.05;Fig. 5a, b). In parallel with these findings, the in situzymography results showed higher MMP activity in 2K1Chypertensive rats compared with the Sham groups (P<0.05;Fig. 6a, b). Treatment with dihydropyridines attenuated2K1C hypertension-induced increases in gelatinolytic ac-tivity (P<0.05; Fig. 6a, b).
Discussion
The main findings of this study were: (1) the first-, second-,and third-generation dihydropyridines studied here producedsimilar antihypertensive effects and prevented the vascularremodeling associated with 2K1C hypertension; (2) these
findings were associated with similar systemic and vascularantioxidant effects exerted by the three CCBs; (3) these CCBsproduced comparable vascular MMP-2 downregulation.These findings suggest that dihydropyridine CCBs of differ-ent generations exert similar beneficial cardiovascular effectsin this hypertension model.
The findings reported here confirm that 2K1C hyperten-sion induces vascular structural alterations, which may bethe result of long-lasting hypertension and may also be thecause of maintained hypertension (Arribas et al. 2006). Inagreement with previous findings (Castro et al. 2008), theincreased MMP-2 levels that we found in the present studysupport the idea that MMPs, especially MMP-2, may play arole in hypertension-induced structural alterations. In fact,this suggestion has been confirmed in the 2K1C and inother animal models of hypertension (Castro et al. 2008,2010; Bouvet et al. 2005; Flamant et al. 2007; Watts et al.2007), and it is possible that these alterations result fromenhanced mechanical stretch and strain, which are majorstimuli for wall hypertrophy of conduit arteries inhypertension, thus upregulating MMP-2 expression andactivity (Asanuma et al. 2003; Chesler et al. 1999; Groteet al. 2003). The similar effects produced by the different
a
b
2K1C
200µm
2K1C+Nimo
200µm
2K1C+Amlo
200µm
200µm
Sham
200µm
Sham+Nife
200µm
Sham+Nimo
200µm
Sham+Amlo
2K1C+Nife
200µm
Sham
Sham
+Nie
Sham
+Nim
o
Sham
+Am
lo2K
1C
2K1C
+Nife
2K1C
+Nim
o
2K1C
+Am
lo0
5
10
15
*
Ao
rtic
MM
P-2
leve
ls
(Arb
itra
ry u
nit
s)
Fig. 5 Effects of dihydropyri-dines on MMP-2 levels assessedby immunofluorescence in theaortas from rats. a Representa-tive photographs of MMP-2detected by immunofluores-cence. b The quantification ofpercentage of vessel surface areacovered by bright red fluores-cence, which reflects MMP-2level. Data are shown as mean±SEM (n=8–10 per group). *P<0.05 versus all the other exper-imental groups
Naunyn-Schmied Arch Pharmacol (2011) 383:35–44 41
dihydropyridines used in the present study could be dueto similar reductions in SBP, which may have led tosimilar reductions in vascular mechanical stretch andstrain.
We found increased systemic and vascular ROSconcentrations in 2K1C rats, which may be responsiblefor the vascular alterations associated with increasedMMPs activities (Castier et al. 2005; Luchtefeld et al.2005; Rajagopalan et al. 1996). MMPs are regulated atmultiple levels including gene transcription, post-translational activation, and by interaction with TIMPs(Matrisian 1994; Murphy et al. 1994). However, ROS canproduce post-translational structural modifications inMMPs, including MMP-2, by interacting with a thiolgroup in the propeptide domain and a zinc ion present incatalytic site, thus resulting in an active full-length enzyme(Okamoto et al. 2001; Viappiani et al. 2009; Van Wart andBirkedal-Hansen 1990). In addition, antioxidants candownregulate MMP-2, as previously shown in the 2K1Chypertension model (Castro et al. 2009). Since the threedihydropyridines used in the present study exerted similarantioxidant effects, it is possible that these three CCBsmay have produced similar protection against oxidativestress-induced MMP-2 upregulation and activation.
While other CCBs including lercanidipine, felodipine,lacidipine, and amlodipine were shown to exert antioxidanteffects (Mason et al. 2003; Taddei et al. 2001; Martinez et al.2006, 2008a, b), there is only one previous study showingthat amlodipine decreased MMP-2 expression in rat cardiacfibroblast (Yue et al. 2004) and a few studies showing thatlercanidipine may downregulate MMP-2 and MMP-9 (Marti-nez et al. 2006, 2008a, b). Our results expand these previousfindings and suggest the possibility that all the dihydropyr-idines may have antioxidant effects that result in loweredMMPs expression/activation in different cardiovascular dis-eases. Although this generalization require further studies tobe valid, it is possible that all dihydropyridines exertpleiotropic effects including antioxidant effects resulting inlower MMPs activities and protection against cardiovascularremodeling.
Another possible explanation for the attenuation of 2K1Chypertension-induced increases in gelatinolytic activity thatwe found in rats treated with dihydropyridines may involvealtered TIMPs expression. This suggestion is supported byprevious studies showing that lercanidipine increased TIMP-2expression in the aortas from diabetic rats (Martinez et al.2008a, b). This may be an important possibility becauseTIMPs bind to pro or active forms of MMPs and the balance
a
b
Sham
200µm
Sham+Nimo
200µm
Sham+Amlo
200µm
Sham+Nife
200µm
2K1C
200µm
2K1C+Nife
200µm
2K1C+Nimo
200µm
2K1C+Amlo
200µm
Sham
Sham
+Nife
Sham
+Nim
o
Sham
+Am
lo2K
1C
2K1C
+Nife
2K1C
+Nim
o
2K1C
+Am
lo0
10
20
30
40 *
In s
itu G
elat
ino
lyti
c ac
tivi
ty
(Arb
itra
ry U
nit
s)
Fig. 6 Effects of dihydropyri-dines on in situ gelatinase ac-tivity assessed by in situgelatinase activity assay in theaortas from rats. a Representa-tive photographs in situ gelati-nase activity assay. b Thequantification of percentage ofvessel surface area covered bybright green fluorescence, whichreflects gelatinolytic activity.Data are shown as mean±SEM(n=8–10 per group). *P<0.05versus all the other experimentalgroups
42 Naunyn-Schmied Arch Pharmacol (2011) 383:35–44

between MMPs and TIMPs determines net proteolyticactivity.
CCBs are a heterogeneous group of drugs with differentpharmacologic effects and are classified as phenylalkyl-amines, benzothiazepines, and dihydropyridines. Whilenifedipine was the first dihydropyridine CCB available,newer generations of dihydropyridines became available,and now, CCBs are an important group of drugs used totreat clinical hypertension. All dihydropyridine CCBs areknown to reduce calcium entry through voltage-dependentchannels and produce vasodilation, and the dihydropyri-dines (including nifedipine, nimodipine, and amlodipine)are more potent than the other subclasses of CCBs (Sica2006). However, their pleiotropic effects may differsignificantly, as previously suggested (Mason et al. 2003).Importantly, while dihydropyridines present pharmacologicsimilarities, they may have different sites and modes ofactions that are not well understood. In fact, differencesbetween CCBs with respect to their effects on endothelin-1production in diseased vessels, on nitric oxide bioavailabil-ity, and maybe on their antioxidant effects may affect theirtherapeutic efficacy, as reviewed elsewhere (Godfraind2005). In fact, drugs producing antioxidant effects andimproving nitric oxide bioavailability showed relevantantihypertensive effects in the hypertension model used inthe present study (Montenegro et al. 2010). Our findingssuggest that dihydropyridines probably are antihypertensivedrugs that abrogate key pathophysiological mechanisms,including oxidative stress and MMPs activation, as shownfor other antihypertensive drugs (Ceron et al. 2010), andthus may prove to be superior than other drugs that simplylower blood pressure.
In conclusion, based on our findings, we suggest lack ofsuperiority of one particular dihydropyridine CCB at leastwith respect to antioxidant effects, MMPs downregulation,and inhibition of vascular remodeling in hypertension.
Acknowledgements This study was funded by Fundação de Amparo aPesquisa do Estado de São Paulo (FAPESP-Brazil) and ConselhoNacional de Desenvolvimento Científico e Tecnológico (CNPq-Brazil).
Conflicts of interest The authors declare that they have no conflictof interest.
References
Ahmed SH, Clark LL, Pennington WR, Webb CS, Bonnema DD,Leonardi AH, McClure CD, Spinale FG, Zile MR (2006) Matrixmetalloproteinases/tissue inhibitors of metalloproteinases: relation-ship between changes in proteolytic determinants of matrixcomposition and structural, functional, and clinical manifestationsof hypertensive heart disease. Circulation 113(17):2089–2096
Arribas SM, Hinek A, Gonzalez MC (2006) Elastic fibres and vascularstructure in hypertension. Pharmacol Ther 111(3):771–791
Asanuma K, Magid R, Johnson C, Nerem RM, Galis ZS, O’CallaghanCJ, Williams B (2003) Uniaxial strain upregulates matrix-degrading enzymes produced by human vascular smooth musclecells. Am J Physiol Heart Circ Physiol 284(5):H1778–H1784
Bouvet C, Gilbert LA, Girardot D, deBlois D, Moreau P (2005)Different involvement of extracellular matrix components insmall and large arteries during chronic no synthase inhibition.Hypertension 45(3):432–437
Castier Y, Brandes RP, Leseche G, Tedgui A, Lehoux S (2005)P47phox-dependent NADPH oxidase regulates flow-inducedvascular remodeling. Circ Res 97:533–540
Castro MM, Rizzi E, Rascado RR, Nagassaki S, Bendhack LM,Tanus-Santos JE (2004) Atorvastatin enhances sildenafil-inducedvasodilation through nitric oxide-mediated mechanisms. Eur JPharmacol 498(1–3):189–194
Castro MM, Rizzi E, Figueiredo-Lopes L, Fernandes K, BendhackLM, Pitol DL, Gerlach RF, Tanus-Santos JE (2008) Metal-loproteinase inhibition ameliorates hypertension and preventsvascular dysfunction and remodeling in renovascular hyperten-sive rats. Atherosclerosis 198(2):320–331
Castro MM, Rizzi E, Rodrigues GJ, Ceron CS, Bendhack LM,Gerlach RF, Tanus-Santos JE (2009) Antioxidant treatmentreduces matrix metalloproteinase-2-induced vascular changes inrenovascular hypertension. Free Radic Biol Med 46(9):1298–1307
Castro MM, Rizzi E, Prado CM, Rossi MA, Tanus-Santos JE, GerlachRF (2010) Imbalance between matrix metalloproteinases andtissue inhibitor of metalloproteinases in hypertensive vascularremodeling. Matrix Biol 29(3):194–201
Ceron CS, Castro MM, Rizzi E, Montenegro MF, Fontana V, SalgadoMC, Gerlach RF, Tanus-Santos JE (2010) Spironolactone andhydrochlorothiazide exert antioxidant effects and reduce vascularmatrix metalloproteinase-2 activity and expression in a model ofrenovascular hypertension. Br J Pharmacol 160(1):77–87
Chesler NC, Ku DN, Galis ZS (1999) Transmural pressure inducesmatrix-degrading activity in porcine arteries ex vivo. Am JPhysiol 277(5 Pt 2):H2002–H2009
Cohuet G, Struijker-Boudier H (2006) Mechanisms of target organdamage caused by hypertension: therapeutic potential. PharmacolTher 111(1):81–98
Dias-Junior CA, Montenegro MF, Florencio BC, Tanus-Santos JE(2008) Sildenafil improves the beneficial haemodynamic effectsof intravenous nitrite infusion during acute pulmonary embolism.Basic Clin Pharmacol Toxicol 103(4):374–379
Flamant M, Placier S, Dubroca C, Esposito B, Lopes I, Chatziantoniou C,Tedgui A, Dussaule JC, Lehoux S (2007) Role of matrix metal-loproteinases in early hypertensive vascular remodeling. Hyperten-sion 50(1):212–218
Galis ZS, Khatri JJ (2002) Matrix metalloproteinases in vascularremodeling and atherogenesis: the good, the bad, and the ugly.Circ Res 90(3):251–262
Godfraind T (2005) Antioxidant effects and the therapeutic modeof action of calcium channel blockers in hypertension andatherosclerosis. Philos Trans R Soc Lond B Biol Sci 360(1464):2259–2272
Griendling KK, FitzGerald GA (2003) Oxidative stress and cardio-vascular injury: part II: animal and human studies. Circulation108(17):2034–2040
Grote K, Flach I, Luchtefeld M, Akin E, Holland SM, Drexler H,Schieffer B (2003) Mechanical stretch enhances mRNA expressionand proenzyme release of matrix metalloproteinase-2 (MMP-2) viaNAD(P)H oxidase-derived reactive oxygen species. Circ Res 92(11):e80–e86
Lehoux S, Lemarie CA, Esposito B, Lijnen HR, Tedgui A (2004)Pressure-induced matrix metalloproteinase-9 contributes to earlyhypertensive remodeling. Circulation 109(8):1041–1047
Naunyn-Schmied Arch Pharmacol (2011) 383:35–44 43

Luchtefeld M, Grote K, Grothusen C, Bley S, Bandlow N, Selle T,Struber M, Haverich A, Bavendiek U, Drexler H, Schieffer B(2005) Angiotensin II induces MMP-2 in a p47phox-dependentmanner. Biochem Biophys Res Commun 328:183–188
Martinez ML, Lopes LF, Coelho EB, Nobre F, Rocha JB, Gerlach RF,Tanus-Santos JE (2006) Lercanidipine reduces matrixmetalloproteinase-9 activity in patients with hypertension. JCardiovasc Pharmacol 47(1):117–122
Martinez ML, Castro MM, Rizzi E, Fernandes K, Demacq C,Bendhack LM, Gerlach RF, Tanus-Santos JE (2008a) Lercanidi-pine reduces matrix metalloproteinase-2 activity and reversesvascular dysfunction in renovascular hypertensive rats. Eur JPharmacol 591(1–3):224–230
Martinez ML, Rizzi E, Castro MM, Fernandes K, Bendhack LM,Gerlach RF, Tanus-Santos JE (2008b) Lercanidipine decreasesvascular matrix metalloproteinase-2 activity and protects againstvascular dysfunction in diabetic rats. Eur J Pharmacol 599(1–3):110–116
Mason RP, Marche P, Hintze TH (2003) Novel vascular biology ofthird-generation L-type calcium channel antagonists: ancillaryactions of amlodipine. Arterioscler Thromb Vasc Biol 23(12):2155–2163
Matrisian LM (1994) Matrix metalloproteinase gene expression. AnnNY Acad Sci 732:42–50
Montenegro MF, Neto-Neves EM, Dias-Junior CA, Ceron CS, CastroMM, Gomes VA, Kanashiro A, Tanus-Santos JE (2010) Quercetinrestores plasma nitrite and nitroso species levels in renovascularhypertension. Naunyn Schmiedebergs Arch Pharmacol 382(4):293–301
Murphy G, Willenbrock F, Crabbe T, O’Shea M, Ward R, AtkinsonS, O’Connell J, Docherty A (1994) Regulation of matrixmetalloproteinase activity. Ann NY Acad Sci 732:31–41
Nelson KK, Melendez JA (2004) Mitochondrial redox control ofmatrix metalloproteinases. Free Radic Biol Med 37(6):768–784
Newby AC (2006) Matrix metalloproteinases regulate migration,proliferation, and death of vascular smooth muscle cells bydegrading matrix and non-matrix substrates. Cardiovasc Res 69(3):614–624
Okamoto T, Akaike T, Sawa T, Miyamoto Y, van der Vliet A, Maeda H(2001) Activation of matrix metalloproteinases by peroxynitrite-induced protein S-glutathiolation via disulfide S-oxide formation. JBiol Chem 276(31):29596–29602
Ra HJ, Parks WC (2007) Control of matrix metalloproteinase catalyticactivity. Matrix Biol 26(8):587–596
Raffetto JD, Khalil RA (2008) Matrix metalloproteinases and theirinhibitors in vascular remodeling and vascular disease. BiochemPharmacol 75(2):346–359
Rajagopalan S, Meng XP, Ramasamy S, Harrison DG, Galis ZS (1996)Reactive oxygen species produced by macrophage-derived foamcells regulate the activity of vascular matrix metalloproteinases invitro. Implications for atherosclerotic plaque stability. J Clin Invest98:2572–2579
Sica DA (2006) Pharmacotherapy review: calcium channel blockers. JClin Hypertens (Greenwich) 8(1):53–56
Sluijter JP, de Kleijn DP, Pasterkamp G (2006) Vascular remodelingand protease inhibition—bench to bedside. Cardiovasc Res 69(3):595–603
Taddei S, Virdis A, Ghiadoni L, Magagna A, Pasini AF, Garbin U,Cominacini L, Salvetti A (2001) Effect of calcium antagonist or betablockade treatment on nitric oxide-dependent vasodilation andoxidative stress in essential hypertensive patients. J Hypertens 19(8):1379–1386
Tomassoni D, Sabbatini M, Amenta F (2003) Effect of differentdihydropyridine-type Ca2+ antagonists on left ventricle hypertrophyand coronary changes in spontaneously hypertensive rats. JCardiovasc Pharmacol 41(4):544–552
VanWart HE, Birkedal-Hansen H (1990) The cysteine switch: a principleof regulation of metalloproteinase activity with potential applica-bility to the entire matrix metalloproteinase gene family. Proc NatlAcad Sci USA 87:5578–5582
Viappiani S, Nicolescu AC, Holt A, Sawicki G, Crawford BD, LeonH, van Mulligen T, Schulz R (2009) Activation and modulationof 72 kDa matrix metalloproteinase-2 by peroxynitrite andglutathione. Biochem Pharmacol 77(5):826–834
Watts SW, Rondelli C, Thakali K, Li X, Uhal B, Pervaiz MH,Watson RE, Fink GD (2007) Morphological and biochemicalcharacterization of remodeling in aorta and vena cava ofDOCA-salt hypertensive rats. Am J Physiol Heart Circ Physiol292(5):H2438–H2448
Yasmin McEniery CM, Wallace S, Dakham Z, Pulsalkar P, Maki-PetajaK, Ashby MJ, Cockcroft JR, Wilkinson IB (2005) Matrixmetalloproteinase-9 (MMP-9), MMP-2, and serum elastase activityare associated with systolic hypertension and arterial stiffness.Arterioscler Thromb Vasc Biol 25(2):372
Yue H, Uzui H, Shimizu H, Nakano A, Mitsuke Y, Ueda T, Lee JD(2004) Different effects of calcium channel blockers on matrixmetalloproteinase-2 expression in cultured rat cardiac fibroblasts.J Cardiovasc Pharmacol 44(2):223–230
44 Naunyn-Schmied Arch Pharmacol (2011) 383:35–44




















