
Downregulation of Apoptosis and Modulation of TGF-β1 by Sodium Selenate Prevents...
17
Downregulation of Apoptosis and Modulation of TGF-β1 by Sodium Selenate Prevents Streptozotocin-Induced Diabetic Rat Renal Impairment Souvik Roy & Sudheer Kumar Dontamalla & Anil Kumar Mondru & Santanu Sannigrahi & Prabhakar Reddy Veerareddy Received: 1 January 2010 / Accepted: 28 January 2010 / Published online: 20 February 2010 # Springer Science+Business Media, LLC 2010 Abstract To investigate whether sodium selenate treatment would impact on the onset of diabetic nephropathy, we examined blood glucose, serum biochemical components, and interrelationship between oxidative stress, TGF-β1, and apoptosis in streptozotocin (STZ) induced diabetic rats. Sixty male Wistar rats were divided into six groups. Group I (n =10), normal control; Group II (n =10), diabetic control; Group III (n =10), sodium selenate (16 μmoles/kg) + diabetic; Group IV (n =10), sodium selenate (32 μmoles/kg) + diabetic; Group V (n =10), sodium selenate (16 μmoles/kg) control; and Group VI (n =10), sodium selenate (32 μmoles/kg) control. Sodium selenate was administered via orogastric route for 10 weeks. In the diabetic group, diabetes was induced by single intraperitoneal injection of STZ (50 mg/kg). The levels of blood glucose were estimated and total cholesterol, high- density lipoprotein (HDL) cholesterol, triglycerides, creatinine, urea, and albumin were detected in serum. Antioxidant status was examined by measuring the superoxide dismutase (SOD), catalase, glutathione, and lipid peroxidation in kidney tissues. Histopathological studies were performed in the kidney tissue sections. The expression of TGF-β1 was estimated by the immunohistochemical analysis in kidneys. Apoptotic study in kidney was performed using the TdT-mediated dUTP nick end labeling technique. It was observed that blood glucose, serum, total cholesterol, HDL cholesterol, triglycerides, creatinine, urea, and albumin were significantly higher in diabetic control groups. Diabetic + sodium selenate (16 and 32 μmoles/kg) significantly reduced blood glucose, serum, total cholesterol, HDL cholesterol, triglycerides, creatinine, urea, and albumin levels. Selenium-treated groups significantly increased antioxidant enzyme activities (SOD, catalase, and glutathione) in kidneys of diabetic rats. All enzyme activities of selenium control groups did not differ compared with the normal control. Sodium selenate reduces significantly lipid peroxidation in diabetic rats. Cellular architecture of the diabetic rats was altered whereas sodium selenate administration rectifies the degenerative changes of the kidney. Profound immunopositivity of TGF-β1 was observed in the glomerular and tubulointerstitial cells of diabetic rat kidney. Immunopositivity of TGF-β1 was significantly reduced in both low and high dose of sodium- Biol Trace Elem Res (2011) 139:55–71 DOI 10.1007/s12011-010-8635-z S. Roy (*) : S. K. Dontamalla : A. K. Mondru : S. Sannigrahi : P. R. Veerareddy Department of Pharmacology, St. Peter’ s Institute of Pharmaceutical Sciences, Hanmakonda, Warangal, Andhra Pradesh 506001, India e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Downregulation of Apoptosis and Modulation of TGF-β1 by Sodium Selenate Prevents...

Downregulation of Apoptosis and Modulation of TGF-β1by Sodium Selenate Prevents Streptozotocin-InducedDiabetic Rat Renal Impairment
Souvik Roy & Sudheer Kumar Dontamalla &
Anil Kumar Mondru & Santanu Sannigrahi &Prabhakar Reddy Veerareddy
Received: 1 January 2010 /Accepted: 28 January 2010 /Published online: 20 February 2010# Springer Science+Business Media, LLC 2010
Abstract To investigate whether sodium selenate treatment would impact on the onset ofdiabetic nephropathy, we examined blood glucose, serum biochemical components, andinterrelationship between oxidative stress, TGF-β1, and apoptosis in streptozotocin (STZ)induced diabetic rats. Sixty male Wistar rats were divided into six groups. Group I (n=10),normal control; Group II (n=10), diabetic control; Group III (n=10), sodium selenate(16 μmoles/kg) + diabetic; Group IV (n=10), sodium selenate (32 μmoles/kg) + diabetic;Group V (n=10), sodium selenate (16 μmoles/kg) control; and Group VI (n=10), sodiumselenate (32 μmoles/kg) control. Sodium selenate was administered via orogastric route for10 weeks. In the diabetic group, diabetes was induced by single intraperitoneal injection ofSTZ (50 mg/kg). The levels of blood glucose were estimated and total cholesterol, high-density lipoprotein (HDL) cholesterol, triglycerides, creatinine, urea, and albumin weredetected in serum. Antioxidant status was examined by measuring the superoxide dismutase(SOD), catalase, glutathione, and lipid peroxidation in kidney tissues. Histopathologicalstudies were performed in the kidney tissue sections. The expression of TGF-β1 wasestimated by the immunohistochemical analysis in kidneys. Apoptotic study in kidney wasperformed using the TdT-mediated dUTP nick end labeling technique. It was observed thatblood glucose, serum, total cholesterol, HDL cholesterol, triglycerides, creatinine, urea, andalbumin were significantly higher in diabetic control groups. Diabetic + sodium selenate (16and 32 μmoles/kg) significantly reduced blood glucose, serum, total cholesterol, HDLcholesterol, triglycerides, creatinine, urea, and albumin levels. Selenium-treated groupssignificantly increased antioxidant enzyme activities (SOD, catalase, and glutathione) inkidneys of diabetic rats. All enzyme activities of selenium control groups did not differcompared with the normal control. Sodium selenate reduces significantly lipid peroxidation indiabetic rats. Cellular architecture of the diabetic rats was altered whereas sodium selenateadministration rectifies the degenerative changes of the kidney. Profound immunopositivity ofTGF-β1 was observed in the glomerular and tubulointerstitial cells of diabetic rat kidney.Immunopositivity of TGF-β1 was significantly reduced in both low and high dose of sodium-
Biol Trace Elem Res (2011) 139:55–71DOI 10.1007/s12011-010-8635-z
S. Roy (*) : S. K. Dontamalla : A. K. Mondru : S. Sannigrahi : P. R. VeerareddyDepartment of Pharmacology, St. Peter’s Institute of Pharmaceutical Sciences, Hanmakonda, Warangal,Andhra Pradesh 506001, Indiae-mail: [email protected]

selenate-treated rats (P<0.05, P<0.01). High numbers of apoptotic cells were observed indiabetic rats whereas sodium selenate in both doses significantly reduces the incidence ofapoptosis (P<0.05, P<0.01). We conclude herein that sodium selenate has the potential toplay a significant role in limiting the renal impairment by altering the apoptosis and TGF-β1in experimental diabetic rats.
Keywords Diabetes . Streptozotocin . Sodium selenate . TGF-β1 . Apoptosis .
Murine kidney
Introduction
Diabetes mellitus is one of best characterized chronic metabolic disorder, still a major concernin human health and shows increasing incidence [1]. According to WHO the estimated numberof patients with diabetes worldwide is around 170 million and projected to reach about 300million in 2025 [2]. It was reported that diabetes is a risk factor for microvascular(nephropathy, retinopathy, and neuropathy) and macrovascular complications, which are theleading cause of deaths in diabetic population [3, 4]. Hyperglycemia results in the generationof reactive oxygen species (ROS) and attenuates antioxidant defense systems, leading tooxidative stress in a variety of tissues [5]. Furthermore, antioxidative enzyme activities likesuperoxide dismutase (SOD), catalase, and glutathione might also be reduced due to glycationor increased lipid peroxidation products [6]. Hyperlipidemia, which is a common feature ofdiabetes, is also suggested to be involved in the increased lipid peroxidation [7]. Severalinvestigators suggested that treatment of hyperglycemia and hyperlipidemia may be beneficialin diminishing oxidative stress and related complications in diabetes mellitus [8]. There wasan evidence that hyperglycemia is associated with upregulation of renal TGF-β1 mRNAexpression [9]. It became clear that ameliorating oxidative stress through treatment withantioxidants might be an effective strategy for reducing diabetic complications.
Renal cell apoptosis is important in both physiological conditions such as normal renaldevelopment and pathological processes affecting the tubulointerstitial compartment of thekidney. Apoptosis may be particularly prominent during acute kidney injury, and it alsooccurs in progressive chronic kidney disease including chronic changes associated withnormal renal aging and age-related diseases such as hypertension and type 2 diabetes [10, 11].Apoptosis contribute to the late structural abnormalities in the diabetic kidney [12].
Selenium (Se) is a dietary essential trace element required for many cellular processes.Organic forms of selenium include selenomethionine, selenocysteine foundmainly in plants andanimals respectively whereas inorganic forms include selenate, selenite (mainly supplements)[13]. In diet, Se mostly present in organic forms, whereas inorganic Se compound exists muchless frequently and at very low amounts. Both organic and inorganic forms of Se appear to beutilized with similar efficacy in the body to produce selenoproteins [14]. Inorganic forms ofselenium are reduced by glutathione and number of intermediate metabolic steps lead to thegeneration of hydrogen selenide (H2Se), which either serves as a precursor for the synthesis ofselenoproteins or undergo stepwise methylation to generate the mono-, di-, and trimethylatedforms of selenium [15]. In rats, selenium dose close to LD50 values, i.e., 3.5 [16] and 4.5 mg/kgbody weight, [17] were applied to attenuate the diabetic status.
Selenium is a functional component of several enzymes with physiological antioxidantproperties, including glutathione peroxidase and thioredoxine [18]. For the first time the insulin-like effect of selenium was shown in in vitro studies [19]. In later studies it was reported thatfrequent intraperitoneal administration of selenate to streptozotocin (STZ)-induced diabetic rats
56 Roy et al.

for 7 weeks caused sustained decrease in their hyperglycemia [16]. It was reported that oralselenate induced a sustained improvement of glucose homeostasis in diabetic rats by insulin-like action, involving partial correction of altered pretranslational regulatory mechanisms inliver metabolism [20]. Sodium selenate corrected glucose tolerance and heart function indiabetic rats. An insulin mimetic role for selenate was also shown in type II db/db mice [21].
Streptozotocin is an antibiotic derived from streptomyces achromogenes, structurally anitrosourea derivative. It is one of most commonly used substance to induce diabetes inexperimental animals [22]. Streptozotocin causes alkylation or breakage of DNA strandsand a consequent increase in the activity of poly-ADP-ribose synthetase, an enzymedepleting the NAD+ in β cells leading to energy deprivation and finally necrosis of β cellsof pancreas [23]. Streptozotocin induces oxidative stress in both blood and tissuesparticularly in the liver and kidneys. Reduction in antioxidant states and an elevation inlipid peroxidation occur in kidneys of streptozotocin-treated rats [24].
This study was designed to investigate the effect of sodium selenate on the glucosehomeostasis, antioxidant defense systems, serum biochemical parameters, histological changes,cell damage biomarkers, and apoptosis in streptozotocin-induced diabetic rat kidneys.
Materials and Methods
Animals and Experimental Design
Male Wistar rats (200–230 g) were procured from National Institute of Nutrition,Hyderabad, India and were used for the experiment. Rats were maintained in an air-conditioned room (20±1°C) with a normal night and day cycle. Rats were feed with semipurified basal diet (Table 1) and demineralized drinking water ad libitum. The rats wereallowed to acclimatize to the laboratory environment for a week before the start of theexperiment. All experimental procedures were conducted in conformity with InstitutionalAnimal Ethics committee CPCSEA (Reg. No. 1126/bc/07 CPCSEA) for the care and use ofanimals and were strictly followed throughout the study.
After acclimatization, rats were randomly divided into six groups as follows: Group I—control rats (n=10), Group II—untreated diabetic rats (n=10), Group III—sodium selenate(16 μmoles/kg) treated diabetic rats (n=10), Group IV—high-dose sodium selenate
Table 1 Composition of Food Mixture
Ingredient Amount (g/kg diet)
Wheat flour (Triticum aestivum)a 500
Bengal gram (Cicer artetinum) 150
Maize dust (Zea mays) 150
Milk powder (spray dried) 150
Macromineral mixb 49.75
Micromineral mixc 0.25
aWheat flour, Bengal gram, maize, and spray dried milk contain sufficient amounts of vitamins and fattyacids and no extra vitamin mixtures were added to the foodbMacromineral mix provided (g/kg food): CaCO3, 16.7; NaHPO4, 13.7; NaCl, 6; KCl, 12; and MgO, 17.5cMicromineral mix provided (mg/kg food): FeSO4.7H2O, 216; CuSO4, 20; Ca(IO3)2, 1.3; MnO, 5;CoCl2.6H2O, 1.9; Na2SeO3, 0.30; CrCl3.8H2O, 4; and Na2Mo4.2H2O, 1.5
Sodium Selenate Prevents Streptozotocin-Induced Rat Renal Impairment 57

(32 μmoles/kg) treated diabetic rats (n=10), Group V—sodium selenate (16 μmoles/kg) treatedcontrol rats (n=10), and Group VI—high-dose sodium selenate (32 μmoles/kg) treated controlrats (n=10). Diabetes was induced in Groups II, III, and IV rats following overnight fasting byan i.p. injection of single dose of streptozotocin (50 mg/kg). Streptozotocin was dissolved in afreshly prepared 0.01 M citrate buffer at pH 4.5 while control rats were injected with bufferalone. Streptozotocin-injected animals were given 5% glucose for 24 h to prevent initialstreptozotocin-induced hypoglycemic mortality. Experiment was conducted up to 10 weeks.Sodium selenate was administered via orogastric route by dissolving in distilled water.
After 10 weeks of the experiment, blood samples were collected from tail vein for thebiochemical estimations, then rats were sacrificed by ether anesthesia and kidneys wereisolated for histopathological, immunohistochemical, and apoptotic studies.
Chemicals
Streptozotocin, sodium selenate (Na2SeO4), biotinylated rabbit anti-mouse IgG, streptavidinperoxidase, diaminobenzidine tetra hydrochloride (DAB) was obtained from Sigmachemicals (St. Louis, MO, USA). The rabbit anti-mouse TGF-β1 antibody was purchasedfrom ANASPEC Inc (San Jose, CA, USA). All other chemicals and reagents used were ofanalytical grade and purchased in purest form, available from local firms.
Biochemical Assays
Blood glucose level, serum cholesterol, high-density lipoprotein (HDL) cholesterol,triglycerides, creatinine, urea, and albumin were estimated by using commercially availablebiochemical estimation kits (Ensure Biotech Pvt. Ltd, Hyderabad, India).
Antioxidant Enzyme Activity and Lipid Peroxidation
The lipid peroxidation in the control and all treated groups of animals was measured by thequantification of thiobarbituric acid reactive substances (TBARS) determined by themethod of Buege and Aust [25]. The activity of superoxide dismutase in the kidneys ofcontrol and treated rats was measured by the xanthine–xanthine oxidasecytochrome Cmethod [26]. The activities of glutathione peroxidase and catalase were measured by themodified method of Pagila and Valentine [27] and Johansson and Hakan Borg [28] method.
Histological Examination
Ten weeks after the experiment, animals from each group were randomly selected. A portionof the kidneys were excised from ether anesthetized rat fixed in 10% formalin and processedfor histological studies. Tissues were dehydrated through 70%, 90%, and 100% alcohol andembedded in low melting point paraffin wax. Sections of 4 μm thickness were cut and placedserially on glass slide. The sections were deparaffinized in xylene and rehydrated through100%, 90%, and 70% alcohol. Three continuous sections were made from each kidney tissueand stained with hematoxylin and eosin for histological evaluation using light microscopy.
Immunohistochemical Analysis
Immunostaining of TGF-β1: immunohistochemical detection of TGF-β1 protein in cold-acetone-fixed, paraffin-embedded kidney section was performed by the avidin–biotin–
58 Roy et al.

peroxidase complex method [29]. Briefly, 5-μm thin sections of lysin-coated slides weredeparaffinized and rehydrated. For immunolabelling of TGF-β1 antigen retrievals werefacilitated by heating the sections in citrate buffer pH 6.0, for 20 min. Endogenous peroxidaseactivity was blocked with 1% H2O2 in 0.1 M Tris–Nacl, (pH 7.6) for 30 min. After incubationin 5% normal goat serum, sections were then separately incubated overnight at 4°C with theprimary rabbit anti-mouse TGF-β1 (AnaSpec, Inc, San Jose, CA, USA) polyclonal antibodyat 1:50 dilutions respectively in 1% bovine serum albumin. Sections were then incubated witha biotinylated secondary antibody, goat anti-rabbit IgG (Sigma) for 30 min at 37°C with 1:100dilution. This was followed by incubation with streptavidin peroxidase (1:100) for 1 h. Afterthat, 100–400 μl of DAB reagent was added to each section; as soon as sections turn brown,the slides were immersed in double-distilled water (ddH2O). Then slides were counter-stainedwith Harris hematoxylin for 2 min. The tissue sections washed in ddH2O for 5 min each. Thesections are dehydrated and mounted with DPX and served as a positive control. Thepercentage of immunopositive cells were counted under a light microscope.
Apoptosis by TUNEL Assay
Apoptotic cells in kidney tissue sections were determined by terminal deoxynucleotidyltransferase (TdT)-mediated dUTP nick end labeling (TUNEL) assay. The sections weredigested with proteinase K (20 μg/ml in PBS, Darmstadt, Germany) for 15 min at roomtemperature, and rinsed with double-distilled water. Slides were then quenched by 2% H2O2
for 5 min at room temperature, equilibrated with buffer. Slides were then incubated withTdT buffer (30 mM Trizma base, pH 7.2, 140 mM sodium cacodylate, 1 mM cobaltchloride), followed by TdT reaction solution containing TdT and dUTP for 90 min at 37°C,then washed with 2% standard saline citrate to stop the reaction for 10 min at roomtemperature. The slides were then washed with PBS for 5 min each and incubated with anti-digoxigenin peroxidase (Sigma) for 30 min (1:100) at room temperature. Color wasdeveloped using 0.05% 3,3′-dimethyl aminobenzene (Sigma, St. Louis, MO, USA) in0.01% H2O2 diluted with Tris–HCl (pH 7.5; DAB Kit Sigma, St. Louis, MO, USA), thenlightly counterstained with Leishman stain. Sections were then washed, dehydrated, andmounted. Apoptotic cells were identified by a brown stain over the nuclei.
Statistical Analysis
Data are expressed mean ± standard error of the mean (SEM) were calculated fromquantified data obtained from at least three replicative experimental conditions. Statisticalanalyses were performed using t test and one-way analysis of variance (ANOVA) followedby post Dunnet’s test as appropriate using graph pad prism. Differences are statisticallysignificant when P<0.01 and <0.05.
Results
Body Weight Changes
Figure 1 shows the body weight changes of different groups of rats during the experimentalperiod. The untreated diabetic rats gained weight at a much lower rate compared to controland sodium-selenate-treated groups. Administration of sodium selenate to diabetic (GroupsIII and IV) rats resulted in an increase in body weight compared to diabetic rats (Group II).
Sodium Selenate Prevents Streptozotocin-Induced Rat Renal Impairment 59

Results suggested that sodium selenate treatment has positive effect on maintaining bodyweights in diabetic rats. A gradual increase in body weights of only sodium-selenate-treatedgroups (Groups V and VI) were similar to that of normal control rats.
Biochemical Changes
The effect of sodium selenate on blood glucose levels in diabetic and nondiabetic rats wasshown in (Fig. 2). After STZ injection a significant increase was observed in the bloodglucose levels of different groups of rats (data not shown). Sodium selenate (16 and32 μmoles/kg) treatment significantly decreased blood glucose levels in diabetic rats after10 weeks (P<0.01) as compared to diabetic animals. Blood glucose levels of only sodium-selenate-administered animals were similar to that of control rats (Group I).
Serum cholesterol levels were increased in diabetic group animals (Group II) comparedto control rats (Group I). Oral administration of sodium selenate at the doses of (16 and32 μmoles/kg) significantly decreased the serum cholesterol levels in diabetic rats (P<0.01;Fig. 3). Only sodium-selenate-treated rats (Groups V and VI) have serum cholesterol levelssimilar to control (Group I) rats. Figure 4 shows the concentrations of serum triglycerides indifferent groups of rats after 10 weeks of experiment. The triglyceride concentrations in theserum of diabetic rats increased compared to control rats. A significant difference in theserum triglycerides of sodium-selenate-treated diabetic rats (Groups III and IV) anduntreated diabetic rats were observed (P<0.01). No differences in the serum concentrationsof triglycerides in control and only sodium-selenate-treated rats were observed. Diabeticrats have decreased HDL cholesterol levels compared to control rats (P<0.01; Fig. 5). HDLcholesterol levels were increased significantly in sodium-selenate-administered diabetic rats
Fig. 1 Time course of the effects of sodium selenate on body weight within groups. Group I control rats,Group II diabetic rats, Group III sodium selenate (16 μmoles/kg) treated rats, Group IV sodium selenate(32 μmoles/kg) treated rats, Group V sodium selenate (16 μmoles/kg) treated control rats, and Group VIsodium selenate (32 μmoles/kg) treated control rats. Values are given as mean ± SEM for groups of sixanimals each
60 Roy et al.

(Groups III and IV) compared to untreated diabetic rats (P<0.01). The levels of HDLcholesterols in only sodium selenate administered rats were similar to control rats (Group I).
Serum levels of creatinine in different experimental groups were shown in (Fig. 6).Streptozotocin-induced diabetic rats showed increased levels of serum creatinine comparedto normal control (P<0.01). Significant differences between the sodium-selenate-treated(Groups III and IV) and diabetic rats (Group II) were observed (P<0.01). There was nosignificant difference between the sodium selenate control (Groups V and VI) groups andnormal control group (Group I). An increase in serum urea levels was observed in diabeticrats and these levels were decreased with both doses (16 and 32 μmoles/kg) of sodiumselenate administration (P<0.01). There was no significant difference between the sodiumselenate control (Groups V and VI) groups and normal control group (Group I; Fig. 7).
Fig. 3 Serum cholesterol levels within groups. Group I control rats, Group II diabetic rats, Group III sodiumselenate (16 μmoles/kg) treated rats, Group IV sodium selenate (32 μmoles/kg) treated rats, Group V sodiumselenate (16 μmoles/kg) treated control rats, and Group VI sodium selenate (32 μmoles/kg) treated controlrats. Values are given as mean ± SEM for groups of six animals each. *Significant difference from nontreateddiabetic rats (P<0.01)
Fig. 2 Blood glucose levels within groups after 10 weeks. Group I—control rats, Group II—diabetic rats,Group III—sodium selenate (16 μmoles/kg) treated rats, Group IV—sodium selenate (32 μmoles/kg) treatedrats, Group V—sodium selenate (16 μmoles/kg) treated control rats, and Group VI—sodium selenate(32 μmoles/kg) treated control rats. Values are given as mean ± SEM for groups of six animals each.*Significant difference from nontreated diabetic rats (P<0.01)
Sodium Selenate Prevents Streptozotocin-Induced Rat Renal Impairment 61

Serum albumin levels were higher in diabetic rats (Fig. 8). Significant difference betweensodium-selenate-administered rats (Groups III and IV) and diabetic rats (Group II) wasobserved (P<0.01). No differences in the serum concentrations of triglycerides in controland only sodium-selenate-treated rats were observed.
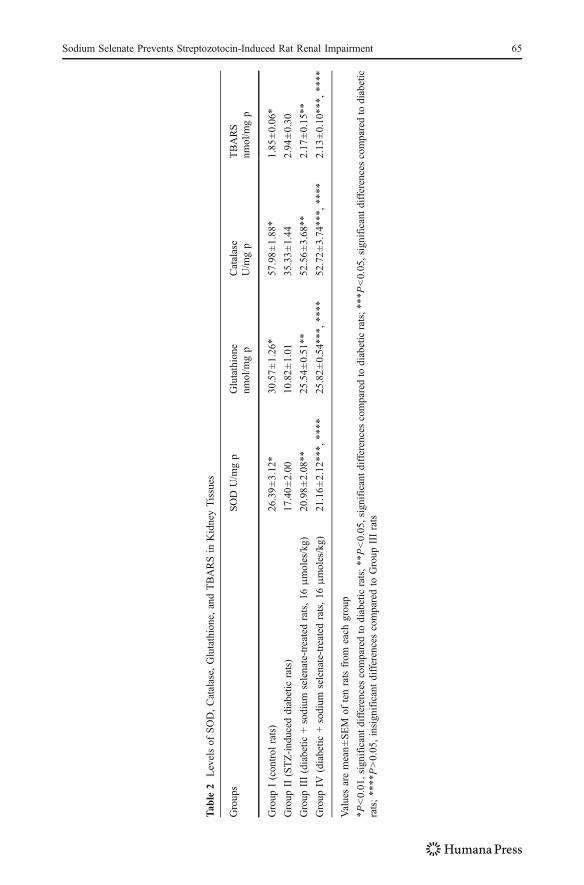
Antioxidant Enzyme Activity and Lipid Peroxidation
The activities of SOD, catalase, glutathione, and TBARS concentrations were given in(Table 2). In the kidneys of STZ-induced diabetic rats, SOD, catalase, and glutathione weresignificantly decreased (17.40±2.00, 35.33±1.44, and 10.82±0.80) compared to the controlrats (26.39±3.12, 57.98±1.88, and 20.18±2.47), while sodium-selenate-administereddiabetic rats (16 and 32 μmoles/kg) appeared to increase the activities of SOD, catalase,and glutathione significantly (P<0.05). No significant difference was observed in betweenGroups III (16 μmoles/kg) and IV (32 μmoles/kg) rats (P>0.05). Sodium-selenate-treatedcontrol rats (Groups V and VI) have SOD, catalase, and glutathione activities as normalcontrol rats (Group I; data not shown).
The lipid peroxidation (TBARS) in the diabetic rats were significantly elevated (2.94±0.30) compared to normal control rats (1.85±0.06). Administration of sodium selenate to
Fig. 5 Serum HDL cholesterol levels within groups. Group I control rats, Group II diabetic rats, Group IIIsodium selenate (16 μmoles/kg) treated rats, Group IV sodium selenate (32 μmoles/kg) treated rats, Group Vsodium selenate (16 μmoles/kg) treated control rats, and Group VI sodium selenate (32 μmoles/kg) treatedcontrol rats. Values are given as mean ± SEM for groups of six animals each. *Significant difference fromnontreated diabetic rats (P<0.01)
Fig. 4 Serum triglyceride levels within groups. Group I control rats, Group II diabetic rats, Group IIIsodium selenate (16 μmoles/kg) treated rats, Group IV sodium selenate (32 μmoles/kg) treated rats, Group Vsodium selenate (16 μmoles/kg) treated control rats, and Group VI sodium selenate (32 μmoles/kg) treatedcontrol rats. Values are given as mean ± SEM for groups of six animals each. *Significant difference fromnontreated diabetic rats (P<0.01)
62 Roy et al.

Groups III (2.17±0.15) and IV rats (2.13±0.10) for 10 weeks decreased the TBARS levelssignificantly (P<0.05). No significant difference in between Groups III and IV (16 and32μmoles/kg) rats (P>0.05) were observed. Sodium-selenate-administered groups (Vand VI)did not show any marked changes in the TBARS levels.
Histopathological Changes
Figure 9 depicted the histological examinations of kidney tissue sections of normal control (a),as well as diabetic control (b) and sodium-selenate-treated rats (c and d). Figure 9a revealednormal control kidney tissues like glomeruli, mesangial cells, glomerular basementmembrane, capsular space, proximal convoluted tubules, and distal convoluted tubules.Normal cellular architecture of kidneys was preserved in control rats. The cellular architecturewas altered in 10-week diabetic rat kidneys (Fig. 9b). Distinctive changes were observed indiabetic rat kidneys like occasional ruptures and a shorting at the brush border accompaniedby the appearance of cytoplasmic debris and desquamated nuclei in the widened lumens of
Fig. 7 Serum urea levels within groups. Group I control rats, Group II diabetic rats, Group III sodiumselenate (16 μmoles/kg) treated rats, Group IV sodium selenate (32 μmoles/kg) treated rats, Group V sodiumselenate (16 μmoles/kg) treated control rats, and Group VI sodium selenate (32 μmoles/kg) treated controlrats. Values are given as mean ± SEM for groups of six animals each. *Significant difference from nontreateddiabetic rats (P<0.01)
Fig. 6 Serum creatinine levels within groups. Group I control rats, Group II diabetic rats, Group III sodiumselenate (16 μmoles/kg) treated rats, Group IV sodium selenate (32 μmoles/kg) treated rats, Group V sodiumselenate (16 μmoles/kg) treated control rats, and Group VI sodium selenate (32 μmoles/kg) treated controlrats. Values are given as mean ± SEM for groups of six animals each. *Significant difference from nontreateddiabetic rats (P<0.01)
Sodium Selenate Prevents Streptozotocin-Induced Rat Renal Impairment 63

the tubules. In addition, vacuolization, picnotic nuclei, and edema in the cells of tubules andhyalinization of arterioles with thickened basement membranes of proximal and distalconvoluted tubules were noted (Fig. 9b). It was observed from histological examinations thattreatment of diabetic rats by sodium selenate treatment (16 μmoles/kg) somehow corrects thechanges which were observed in diabetic rats like occasional ruptures, picnotic nucleus,cytoplasmic debris, vacuolization, and thickened basement membranes (Fig. 9c). There wereno signs of appearance of cytoplasmic debris, desquamated nuclei, edematous of tubules, andhyalinization of the cells observed in sodium-selenate-treated (32 μmoles/kg) rat kidneys andthey look like normal. Selenate control (Groups V and VI) showed no alterations of the basicstructure of the kidneys and look like same as normal control (figure not shown).
Immunohistochemical Analysis
The expression of TGF-β1 in kidney tissue was examined immunohistochemically(Table 3). We could not detect TGF-β1 expression in the normal control animals (Group I).Positive staining of this protein was observed in the glomerulus, tubulointerstitium of kidneysof STZ-treated diabetic animals (Fig. 10b). Sodium-selenate-treated kidney sections(Group III, 16 μmoles/kg) significantly decreased (17.5±3.3; P<0.05) the immunopo-sitivity as compared to STZ control animals (24.4±2.59). There was also very significantreduction (12.4±2.2; P<0.01) of the immunopositity of TGF-β1 expression in sodiumselenate (Group IV, 16 μmoles/kg) kidney sections was observed compared to diabeticcontrol animals (Group II). A significant reduction (P<0.05) of the immunopositivity ofTGF-β1 expression was observed in between the sodium-selenate-treated (16 μmoles/kg,32 μmoles/kg) kidney sections.
Apoptosis by TUNEL Assay
To further analyze the chemopreventive action of sodium selenate on diabetic nephropathy,we examined its effect on apoptosis on diabetic kidney sections. In kidneys from diabeticrats, TUNEL staining revealed a significant degree of apoptosis in proximal tubules, distaltubules, and interstitial cells. No apoptosis was observed in cells within the glomeruli ofdiabetic rats. Treatment with sodium selenate (16 and 32 μmoles/kg) decreased the numberof TUNEL positive apoptotic cells.
Fig. 8 Serum albumin levels within groups. Group I control rats, Group II diabetic rats, Group III sodiumselenate (16 μmoles/kg) treated rats, Group IV sodium selenate (32 μmoles/kg) treated rats, Group V sodiumselenate (16 μmoles/kg) treated control rats, and Group VI sodium selenate (32 μmoles/kg) treated controlrats. Values are given as mean ± SEM for groups of six animals each. *Significant difference from nontreateddiabetic rats (P<0.01)
64 Roy et al.
Tab
le2
Levelsof
SOD,Catalase,
Glutathione,andTBARSin
KidneyTissues
Groups
SOD
U/m
gp
Glutathione
nmol/m
gp
Catalase
U/m
gp
TBARS
nmol/m
gp
Group
I(control
rats)
26.39±3.12
*30
.57±1.26*
57.98±1.88
*1.85
±0.06*
Group
II(STZ-ind
uced
diabetic
rats)
17.40±2.00
10.82±1.01
35.33±1.44
2.94
±0.30
Group
III(diabetic
+sodium
selenate-treated
rats,16
μmoles/kg)
20.98±2.08**
25.54±0.51**
52.56±3.68**
2.17
±0.15**
Group
IV(diabetic
+sodium
selenate-treated
rats,16
μmoles/kg)
21.16±2.12
***,
****
25.82±0.54**
*,**
**52
.72±3.74
***,
****
2.13
±0.10**
*,**
**
Valuesaremean±SEM
oftenratsfrom
each
group
*P<0.01,significant
differencescomparedto
diabeticrats;**P<0.05,significant
differencescomparedto
diabeticrats;***P
<0.05,significant
differencescomparedto
diabetic
rats;**
**P>0.05,insignificantdifferencescomparedto
Group
IIIrats
Sodium Selenate Prevents Streptozotocin-Induced Rat Renal Impairment 65

Figure 11a–d shows representative apoptotic pictures of kidney tissue sections of rats fromnormal control (Group I), diabetic rat (Group II), sodium-selenate-treated (16 μmoles/kg;Group III), and sodium-selenate-treated (32 μmoles/kg; Group IV), respectively. Thechromogen-generated brown stain is an indication of apoptotic cells. It should be noted thatin almost every case, the brown stain overlaps the condensed chromatin of apoptotic bodies,thus confirming that the TUNEL assay correlates well with the morphological appearance ofapoptosis. The rate of apoptosis is generally very low in normal control group (Group I).Typically, from diabetic control group slides, we found an average of 30–35 apoptotic cells in a
Fig. 9 a Normal histological appearance of kidney tissues of control rat (H&E), ×200. b Histologicalappearance of kidney tissues of a diabetic rat. Occasional ruptures ( ), shorting at the brush border
( ), cytoplasmic debris ( ), desquamated nuclei ( ), vacuolization ( ), picnotic nuclei ( ) in thecells of tubules, and thickening of glomerular basement membrane ( ; H&E); ×200. c Histologicalappearance of kidney tissues of sodium selenate (16 μmoles/kg) treated diabetic rat (H&E); ×200. dHistological appearance of kidney tissues of sodium selenate (32 μmoles/kg) treated diabetic rat (H&E); ×200
Table 3 TGF-β1 Immunopositive Cells in Kidney Tissues
Groups TGF-β1 immunopositive cells (%)
Group II—diabetic control rats 24.4±2.59a
Group III—diabetic + sodium selenate-treated rats (16 μmoles/kg) 17.5±3.3*
Group IV—diabetic + sodium selenate-treated rats (32 μmoles/kg) 12.4±2.2**, ***
Approximately 200 cells were counted per field, five fields were examined per slide and five slides wereexamined per group. Results are analyzed by t test and one-way ANOVA. The expression of TGF-β1 proteinwas not detectable in kidneys of normal group (Group I) animals
*P<0.05, significant difference compared to diabetic control rats; **P<0.01, significant difference comparedto diabetic control rats; ***P<0.05, significant difference compared to Group III ratsa Values are presented mean ± SEM
66 Roy et al.

field of about 200 cells (Fig. 11b). This number decreased to an average of seven to eightstained cells in the sodium selenate (16 μmoles/kg; Fig. 11c) and three to four stained cells inthe sodium selenate (32 μmoles/kg) treated group (Fig. 11d). As seen in Table 4, apoptoticindexwas calculated. The percentage of TUNEL positive apoptotic cells was denoted as apoptoticindex. Quantitative analysis revealed a marked increase in apoptotic cells in kidneys (17.5±0.02)from streptozotocin-induced diabetic rats at 10 weeks. An effect that was completely reversed by(16 μmoles/kg) sodium selenate and the apoptotic index (6.2±0.01) which was statisticallysignificant (P<0.05) when compared to diabetic control using one-way ANOVA. Further thereduction of apoptosis (1.5±0.05) in high dose of sodium selenate (32 μmoles/kg) was morestatistically significant (P<0.01) when compared to diabetic control group. In contrast, featuresof cell necrosis were not observed in streptozotocin-induced diabetic rat kidney.
Thus, it appears that the changes in renal cellular architecture in diabetes were associatedwith the progressive cell loss, which was further restored by the sodium selenate intervention.
Discussion
Diabetes mellitus is a pathologic condition, resulting in severe metabolic imbalances andnonphysiologic changes in many tissues, where oxidative stress plays an important role inthe etiology of this disease [30]. STZ causes diabetes by rapid depletion of beta cells whichleads to reduction in insulin release and hyperglycemia [31]. Hyperglycemia causes oxidativedamage by generation of ROS and development of diabetic complications [32]. STZ-induceddiabetes is characterized by severe loss of body weight due to increased muscle wasting in
Fig. 10 a Expression of TGF-β1 in glomeruli, tubulointerstitium of control rat kidney (H&E); ×450. bExpression of TGF-β1 in diabetic rat kidney (H&E); ×450. c Expression of TGF-β1 in sodium selenate(16 μmoles/kg) treated diabetic rat kidney (H&E); ×450. d Expression of TGF-β1 in sodium selenate(32 μmoles/kg) treated diabetic rat kidney (H&E); ×450. Arrow represents the expression of TGF-β1 inglomeruli, tubulointerstitium of kidney
Sodium Selenate Prevents Streptozotocin-Induced Rat Renal Impairment 67

diabetes [33]. Sodium selenate administered orally to diabetic rats showed modest increase inthe average body weights compared to the untreated diabetic rats (Fig. 1).
Diabetic rats exhibited increased blood glucose levels; administration of sodium selenate todiabetic rats reduced blood glucose levels significantly (P<0.01; Fig. 2). These effects may beattributed to insulin-like actions of selenate include stimulation of glucose uptake andregulation of metabolic processes such as glycolysis, gluconeogenesis, and fatty acidsynthesis. The mechanism by which selenate is capable of mimicking insulin is not clear [34].
The degree of hypercholesterolemia is directly proportional to severity in diabetes [35].In the present study, we have observed higher levels of serum cholesterol levels in diabetic
Fig. 11 Representative immunohistochemistry pictures (×450) of apoptotic cells in kidney tissue. Brown-stained cells are undergoing apoptosis. Arrowheads represents the expression of TUNEL positive apoptoticcells. a Represents normal control. b Represents diabetic control. c Represents sodium selenate (16 μmoles/kg)treated rat. d Represents sodium selenate (32 μmoles/kg) treated rat
Table 4 Apoptotic Value of Kidney Tissue Sections
Groups Apoptotic indexa (AI)
Group I—normal control rats 0.5±0.01*
Group II—diabetic control rats 17.5±0.02
Group III—diabetic + sodium selenate-treated rats (16 μmoles/kg) 6.2±0.01**
Group IV—diabetic + sodium selenate-treated rats (32 μmoles/kg) 1.5±0.05***
Approximately 200 cells were counted per field, five fields were examined per slide and five slides wereexamined per group. The percentage of TUNEL positive apoptotic cells was denoted as apoptotic index.Results are analyzed by t test and one-way ANOVA
*P<0.01, significant difference compared to diabetic control rats; **P<0.05, significant difference comparedto diabetic control rats; ***P<0.01, significant difference compared to diabetic control ratsa Values are presented mean ± SEM
68 Roy et al.

rats. Administration of sodium selenate to diabetic rats (Groups III and IV) normalizescholesterol levels significantly (Fig. 3).
Diabetic rats had marked elevated levels of triglycerides, but decreased levels of HDLcholesterol [36]. Administration of sodium selenate to the diabetic rats decreased the serumlevels of triglycerides and increased the HDL cholesterol levels. The efficacy of sodiumselenate on serum triglycerides level and serum HDL cholesterol level may be attributed toincreased lipoprotein lipase activity [37]. Profound changes in protein metabolism occur indiabetes, enhanced catabolism of muscle proteins in diabetes elevates the serum creatininelevels [38]. Our findings (Fig. 6) concluded that sodium selenate ameliorates the renalfunctions in diabetic rats by decreasing the creatinine levels. The diabetic hyperglycemiainduces the elevation of serum levels of urea and albumin. In our study, STZ-induced diabeticrats increased serum urea, albumin concentrations compared to control group. Administrationof sodium selenate decreased the levels of serum urea and albumin (Figs. 7 and 8).
An untreated diabetic state results in exaggerated synthesis of free radicals and defectivescavenging systems which in turn cause lipid peroxidation [39]. Several studies have shownincreased lipid peroxidation in experimental diabetes [40]. STZ has been shown to produceoxygen free radicals and lipid peroxide mediate tissue damages. Previous studies havereported that lipid peroxidation in kidneys of diabetic rats was increased [41]. In agreementwith our result, the concentrations of renal TBARS was high in STZ-induced diabetic rats,where as these levels in sodium-selenate-treated rats showed decreased the concentrationsof TBARS which indicate the decreased rate of lipid peroxidation (Table 2).
Few studies have shown that selenium supplementation increase antioxidant enzymeactivities, decrease lipid peroxidation levels, and upregulated mRNA expression forantioxidant enzymes in patients and animal models of diabetes [42]. Interestingly, weobserved in diabetic rat that the decreased activities of renal SOD, catalase, and glutathione,which play an important role in scavenging free radicals. In line with this, our results alsoshowed that oral administration of sodium selenate to diabetic rats for 10 weeks increasedthe activities of renal SOD, catalase, and glutathione (Table 2). Thus, sodium selenateprovides a protective effect by alteration in antioxidant status.
The kidney exhibits a characteristic pattern of changes during diabetes [43]. The histologicalevidence suggests that structural alterations occur around 6 weeks after induction of diabetes,but oxidative stress starts quite early in diabetes. Increase in oxygen free radical could causelipid peroxidation and cellular damage in the glomeruli and tubular interstitium [44]. In thisstudy an increase in occasional ruptures and a shorting at the brush border, widened lumens ofthe tubules, and increase in dark cells containing a large number of mitochondria may supportthe increased oxygen consumption which is associated with free radical generation and tissuedamage in diabetic rats (Fig. 9b), sodium selenate treatment rectifies the degenerative changesof cellular architecture satisfactorily (Fig. 9c, d).
TGF-β1 induces oxidative stress in vitro in the kidney of both normal and diabetic ratsand this oxidative stress is abrogated by the antibody to TGF-β1 [45]. TGF-β1 has beenshown to decrease catalase and glutathione peroxidase activities in pancreatic beta cells andvascular smooth muscle cells [46]. Neutralizing anti-TGF-β1 antibodies in vivo suppresseddiabetes-related kidney growth [47]. Expression of TGF-β1 gradually increases with theprogression of diabetes. These findings is in agreement with recent study which showedthat positive staining for TGF-β1 mainly occurred within the glomeruli, especially themesangial area, while being weak in the tubules. This increase in positive staining for TGF-β1 was observed in the diabetic rat kidneys (Fig. 10b). In contrast, sodium selenatetreatment dramatically reduced the expression of TGF-β1 protein in the glomeruli(Fig. 10c, d) compared to diabetic rats.
Sodium Selenate Prevents Streptozotocin-Induced Rat Renal Impairment 69

Apoptosis is an ongoing process in many experimental kidney diseases [48, 49]. In ratswith renal ischemia/reperfusion injury, pre-treatment with the anti-apoptotic agent IGF-1 orZVAD-fmk (a caspase inactivator) prevent the onset of tubular cell apoptosis and reducedreperfusion-induced inflammation, associated with improved renal function [50]. The majorfindings of the present study are that apoptosis occurs in tubular and interstitial cells of thekidney at 10 weeks in streptozotocin-induced diabetes, associated with increased TGF-β1expression in glomerulus and tubulointerstitium and with no evidence of necrosis. Sodiumselenate treatment can relieve the symptoms of diabetes through suppression of TGF-β1and apoptotic signals in diabetic rats.
Taken together our result show that the sodium selenate lowered blood glucose andbiochemical profiles associated with kidney damage and alters the antioxidant status, TGF-β1, and apoptotic response in diabetic rats indicating that sodium selenate can relieve orprevent the incidence of diabetic renal impairment.
References
1. American Diabetes Association (2005) Diagnosis and classification of diabetes mellitus. Diabetes Care28:37–42
2. Wild S, Roglic G, Green A, Sicree R, King H (2004) Global prevalence of diabetes estimates for the year2000 and projections for 2030. Diabetes Care 27:1047–1053
3. Pyorala K, Laakso M, Unsitupa M (1987) Diabetes and atherosclerosis a epidemiologic view. DiabetesMetab Rev 3(2):463–524
4. The Diabetes Control and Complications Trial Research Group (1993) The effect of intensive treatmentof diabetes on the development and progression of long term complications in insulin dependent diabetesmellitus. N Engl J Med 329(14):977–986
5. Brownlee M (2001) Biochemistry and molecular cell biology of diabetic complications. Nature 414:813–8206. Sozmen EY, Sozmen B, Delen Y, Onat T (2001) Catalase/superoxide dismutase (SOD) and catalase/
paroxonase (PON) ratios may implicate poor glycemic control. Arch Med Res 32:283–2877. Davi G, Falco A, Patrono C (2005) Lipid peroxidation in diabetesmellitus. Antioxid Redox Signal 7:256–2688. Godin DV, Wohaieb SA, Garnett ME, Goumeniouk AD (1988) Antioxidant enzyme alterations in
experimental and clinical diabetes. Mol Cell Biochem 84:223–2319. Rocco MV, Chen Y, Goldfard S, Ziyadeh FN (1992) Elevated glucose stimulates TGF- β gene
expression and bioactivity in proximal tubule. Kidney Int 41:107–11410. Kerr JF, Gobe GC, Winterford CM, Harmon BV (1995) Anatomical methods in cell death. Methods Cell
Biol 46:1–2711. Andrade L, Vieira JM, Safirstein R (2000) How cells die counts. Am J Kidney Dis 36:662–66812. Nagata S (1997) Apoptosis by death factor. Cell 88:355–36513. Pennington JA, Young BE (1991) Total diet study nutritional elements. J Am Diet Assoc 91:179–18314. Shiobara Y, Yoshida T, Suzuki KT (1998) Effect of dietary selenium species on selenium concentrations
in hair, blood and urine. Toxicol Appl Pharmacol 152:309–31415. Letavayova L, Vlckova V, Brozmanova J (2006) Selenium, from cancer prevention to DNA damage.
Toxicology 227:1–1416. McNeill JH, Delgatty HL, Battell ML (1991) Insulin like effects of sodium selenate in streptozotocin
induced diabetic rats. Diabetes 40:1675–167817. Battell ML, Delgatty HL, McNeill JH (1998) Sodium selenate corrects glucose tolerance and heart
function in STZ diabetic rats. Mol Cell Biochem 179:27–3418. Xia L, Nordman T, Olsson JM, Damdimopoulos A, Bjorkhem-Bergman L, Nalvarte T, Eriksson LL,
Arner ESJ, Spyrou G, Bjornstedt M (2002) The mammalian selenoenzyme thioredoxin reductase reducesubiquinone. A novel mechanism for defense against oxidative stress. J Biol Chem 278:2141–2146
19. Ezaki O (1990) The insulin-like effects of selenate in rat adipocytes. J Biol Chem 265:1124–113020. Becker DJ, Reul B, Ozcelikay AT, Buchet JP, Henquin JC, Brichard SM (1996) Oral selenate improves
glucose homeostasis and partly reverses abnormal expression of liver glycogenic and gluconeogenicenzymes in diabetic rats. Diabetologia 39:3–11
21. Mueller AS, Pallauf J, Rafael J (2003) The chemical form of selenium affects insulinomimetic propertiesof the trace element investigaiton in type II diabetic dbdb mice. J Nutr Biochem 14:637–647
70 Roy et al.

22. Szkudelski T (2001) The mechanism of alloxan and streptozotocin in action in beta cells of rat pancreas.Physiol Res 50:536–546
23. Mythili MD, Vyas R, Akila G, Gunasekaran S (2004) Effect of streptozotocin on the ultra structure of ratpancreatic islets. Microsc Res Tech 63:274–281
24. Kedziora K, Szram S, Komotowski T, Szadujkis-Szaduriki L, Bartosz G (2002) The effect of verapamilon the antioxidant defense system in diabetic kidney. Clin Chim Acta 322:105–112
25. Buege JA, Aust SD (1984) Microsomal lipid peroxidation. Meth Enzymol 105:302–31026. Reddi AS, Bollineni JS (1997) Renal cortical expression of mRNAs for antioxidant enzymes in normal
and diabetic rats. Biochem Biophys Res Commun 235:598–60127. Pagila DE, Valentine WN (1967) Studies on the quantitative and qualitative characterization of
erythrocyte glutathione peroxidase. J Lab Clin Med 70:158–16928. Johansson LH, Hakan Borg LA (1988) A spectrophotometric method for determination of catalase
activity in small tissue samples. Anal Biochem 174:331–33629. Jin R, Chow VT, Tan PH, Dheen ST, Duan W, Bay BH (2002) Metallothionein 2A expression is
associated with cell proliferation in breast cancer. Carcinogenesis 23:81–8630. Baynes JW, Thorpe SR (1996) The role of oxidative stress in diabetic complications. Curr Opin
Endocrinol 3:277–28431. Gupta S, Kataria M, Gupta PK, Murganandam RC, Yashroy RC (2004) Protective role of extracts of
neem seeds in diabetes caused by Streptozotocin in rats. J Ethnopharmacol 90:185–18932. Baynes JW, Thorpe SR (1999) Role of oxidative stress in diabetic complications. A new perspective on
an old paradigm. Diabetes 48:1–933. Ravi K, Ramachandran B, Subramanian S (2004) Protective effect of Eugenia jambolana seed kernel on
tissue antioxidants in STZ induced diabetic rats. Biol Pharm Bull 27:1212–121734. Stapleton SR (2005) Selenium: an insulin mimetic. Cell Mol Life Sci 57:1874–187935. Al-Shamaony L, Al-Khazaraji SM, Twaiji IA (1994) Hypoglycemic effect of Arteminia herba alba II
effect of a valuable extract on some blood parameters in diabetic animals. J Ethnopharmacol 43:167–17136. Naziroglu M, Dilsiz N, Cay M (1999) Protective role of intraperitoneally administered vitamin C and E
and Selenium on the levels of lipid peroxidation in the lens of rats made diabetic with streptozotocin.Biol Trace Elem Res 70:223–232
37. Bayraktitan U (2002) Free radical, diabetes and endothelial function. Diabetes Obes Metab 4:224–23838. Almdal TP, Vilstrup H (1988) Strict insulin treatment normalizes the organic nitrogen contents and the
capacity of urea-N synthesis in experimental diabetes in rats. Diabetologica 31:114–11839. Giugliano D, Ceriello A (1996) Oxidative stress and diabetic vascular complications. Diabetes Care
19:257–26740. Kakkar R, Kalra J, Mantha SV, Prasad K (1995) Lipid peroxidation and activity of antioxidant enzymes
in diabetic rats. Mol Cell Biochem 151:113–11941. Venkateswaran S, Pari L (2002) Antioxidant effect of Phaseolus vulgaris in streptozotocin induced
diabetic rats. Asia Pacific J Clin Nutr 11:206–20942. Naziroglu M, Cay M (2001) Protective role of intraperitoneally administered vitamin E and selenium on
the antioxidative defense mechanisms in rats with diabetes induced by streptozotocin. Biol Trace ElemRes 79:149–159
43. Sharma KP, Mc Cue SR, Dunn (2003) Diabetic kidney disease in the db/db mouse. Am J Physiol 28:F1138–F1144
44. Parinandi NL, Thompson EW, Schmid HHO (1990) Diabetic heart and kidney exhibit increasedresistance to lipid peroxidation. Biochim Biophys Acta 1047:63–69
45. Reddi AS, Camerini, Davalos RA (1990) Diabetes nephropathy an update. Arch Intern Med 150:31–4346. Nishio E, Watanabe Y (1997) Transforming growth factor β is a modulator of platelet derived growth
factor action in vascular smooth muscle cells: a possible role for catalase activity and glutathioneperoxidase activity. Biochem Biophys Res Commun 232:1–4
47. Sharma K, Jin Y, Guo J, Ziyadeh FN (1996) Neutralization of TGF beta by anti TGF beta antibodyattenuates kidney hypertrophy and the enhanced extracellular matrix gene expression in STZ induceddiabetic mice. Diabetes 45:522–530
48. Schelling JR, Cleveland RP (1999) Involvement of Fas-dependent apoptosis in renal tubular epithelialcell deletion in chronic renal failure. Kidney Int 56:1313–1316
49. Sugiyama H, Kashihara N, Makino H, Yamasaki Y, Ota A (1996) Apoptosis in glomerular sclerosis.Kidney Int 49:103–111
50. Daemen MA, Van t Veer C, Denecker G, Heemskerk VH, Wolfs TG, Clauss M, Vandenabeele P,Buurman WA (1999) Inhibition of apoptosis induced by ischaemia reperfusion prevents inflammation. JClin Invest 104:541–549
Sodium Selenate Prevents Streptozotocin-Induced Rat Renal Impairment 71




















