
Proteomic Analysis of Glycated Proteins from Streptozotocin-Induced Diabetic Rat Kidney
11
RESEARCH Proteomic Analysis of Glycated Proteins from Streptozotocin-Induced Diabetic Rat Kidney Ashok D. Chougale • Shweta P. Bhat • Swapnil V. Bhujbal • Mandar R. Zambare • Shraddha Puntambekar • Rahul S. Somani • Ramanamurthy Boppana • Ashok P. Giri • Mahesh J. Kulkarni Published online: 23 April 2011 Ó Springer Science+Business Media, LLC 2011 Abstract Glycation of proteins leading to formation of advanced glycation end products (AGEs) has been con- sidered as one of the important causes of diabetic nephropathy. Therefore, in this study, glycated proteins were detected by anti-AGE antibodies from kidney of streptozotocin-induced diabetic rat showing nephropathic symptoms, by using two dimensional electrophoresis and western blot analysis. These glycated proteins were iden- tified and characterized by using combination of peptide mass finger printing and tandem mass spectrometric approaches. Glycated proteins identified included proteins from metabolic pathways, oxidative stress, cell signaling, and transport. Several of the proteins modified by glycation were involved in glucose metabolism. The extent of gly- cation was higher in diabetes compared to control, in the glycated proteins that were common to both control and diabetic kidney. Two dimensional electrophoresis proteins profiling of glycated proteins suggest that four of the gly- cated proteins were significantly up regulated in diabetes. Keywords Diabetes Nephropathy AGE Glucose toxicity Post-translation modification Introduction Diabetic nephropathy is one of the major complications caused by hyperglycemic condition. The condition is char- acterized by glomerular and tubular basement membrane thickening, microvascular damage, and mesangial extra- cellular matrix expansion with the accumulation of impaired proteins modified by advanced glycation end products (AGEs). Proteins from extracellular matrix, kidney, as well as proteins from circulation, get AGE modified and trapped in kidney [1]. In the diabetic kidney, both intracellular and extracellular AGEs have been observed. Glucose plays main role in extracellular AGEs formation, which activates the RAGEs leading to apoptosis and inflammation [2]. Whereas intracellular AGEs formation are mostly due to various dicarbonyls. Eventually, both types of the AGEs may be responsible for the kidney damage [3]. It was reported that accumulation of AGEs in the kidney was associated with increased accumulation of AGE peptides in plasma and urine in diabetic nephropathy [4]. Such AGE accumulation studied in diffuse and nodular diabetic nephropathy suggests that carboxy methyl lysine (CML) was the major AGE detected in diabetic mesangium, glomerular basement membranes, tubular basement membranes and vessel walls, while pent- osidine was the major AGE detected in interstitial collagen [4]. AGE modification in diabetes significantly increased quantities of methyl glyoxal, an intermediary compound of AGE reaction, accompanied with a superoxide induced oxidative damage of proteins [5]. Adaptive response to oxidative damage was observed in a proteomic analysis of glomeruli of db/db diabetic mice. The diabetic mice showed Electronic supplementary material The online version of this article (doi:10.1007/s12033-011-9409-3) contains supplementary material, which is available to authorized users. A. D. Chougale S. P. Bhat S. V. Bhujbal M. R. Zambare S. Puntambekar A. P. Giri M. J. Kulkarni (&) Chemical Proteomics Group, Biochemical Sciences Division, National Chemical Laboratory (CSIR), Pune 411008, India e-mail: [email protected] R. S. Somani Sinhgad College of Pharmacy, Vadgaon, Pune, India R. Boppana National Centre for Cell Science, Pune, India 123 Mol Biotechnol (2012) 50:28–38 DOI 10.1007/s12033-011-9409-3
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Proteomic Analysis of Glycated Proteins from Streptozotocin-Induced Diabetic Rat Kidney

RESEARCH
Proteomic Analysis of Glycated Proteinsfrom Streptozotocin-Induced Diabetic Rat Kidney
Ashok D. Chougale • Shweta P. Bhat • Swapnil V. Bhujbal •
Mandar R. Zambare • Shraddha Puntambekar • Rahul S. Somani •
Ramanamurthy Boppana • Ashok P. Giri • Mahesh J. Kulkarni
Published online: 23 April 2011
� Springer Science+Business Media, LLC 2011
Abstract Glycation of proteins leading to formation of
advanced glycation end products (AGEs) has been con-
sidered as one of the important causes of diabetic
nephropathy. Therefore, in this study, glycated proteins
were detected by anti-AGE antibodies from kidney of
streptozotocin-induced diabetic rat showing nephropathic
symptoms, by using two dimensional electrophoresis and
western blot analysis. These glycated proteins were iden-
tified and characterized by using combination of peptide
mass finger printing and tandem mass spectrometric
approaches. Glycated proteins identified included proteins
from metabolic pathways, oxidative stress, cell signaling,
and transport. Several of the proteins modified by glycation
were involved in glucose metabolism. The extent of gly-
cation was higher in diabetes compared to control, in the
glycated proteins that were common to both control and
diabetic kidney. Two dimensional electrophoresis proteins
profiling of glycated proteins suggest that four of the gly-
cated proteins were significantly up regulated in diabetes.
Keywords Diabetes � Nephropathy � AGE �Glucose toxicity � Post-translation modification
Introduction
Diabetic nephropathy is one of the major complications
caused by hyperglycemic condition. The condition is char-
acterized by glomerular and tubular basement membrane
thickening, microvascular damage, and mesangial extra-
cellular matrix expansion with the accumulation of impaired
proteins modified by advanced glycation end products
(AGEs). Proteins from extracellular matrix, kidney, as well
as proteins from circulation, get AGE modified and trapped
in kidney [1]. In the diabetic kidney, both intracellular and
extracellular AGEs have been observed. Glucose plays main
role in extracellular AGEs formation, which activates the
RAGEs leading to apoptosis and inflammation [2]. Whereas
intracellular AGEs formation are mostly due to various
dicarbonyls. Eventually, both types of the AGEs may be
responsible for the kidney damage [3]. It was reported that
accumulation of AGEs in the kidney was associated with
increased accumulation of AGE peptides in plasma and urine
in diabetic nephropathy [4]. Such AGE accumulation studied
in diffuse and nodular diabetic nephropathy suggests that
carboxy methyl lysine (CML) was the major AGE detected
in diabetic mesangium, glomerular basement membranes,
tubular basement membranes and vessel walls, while pent-
osidine was the major AGE detected in interstitial collagen
[4]. AGE modification in diabetes significantly increased
quantities of methyl glyoxal, an intermediary compound of
AGE reaction, accompanied with a superoxide induced
oxidative damage of proteins [5]. Adaptive response to
oxidative damage was observed in a proteomic analysis of
glomeruli of db/db diabetic mice. The diabetic mice showed
Electronic supplementary material The online version of thisarticle (doi:10.1007/s12033-011-9409-3) contains supplementarymaterial, which is available to authorized users.
A. D. Chougale � S. P. Bhat � S. V. Bhujbal �M. R. Zambare � S. Puntambekar � A. P. Giri �M. J. Kulkarni (&)
Chemical Proteomics Group, Biochemical Sciences Division,
National Chemical Laboratory (CSIR), Pune 411008, India
e-mail: [email protected]
R. S. Somani
Sinhgad College of Pharmacy, Vadgaon, Pune, India
R. Boppana
National Centre for Cell Science, Pune, India
123
Mol Biotechnol (2012) 50:28–38
DOI 10.1007/s12033-011-9409-3

up regulation of antioxidative enzymes peroxiredoxin 1 and
3, glutathione peroxidase 1, and super oxide dismutase 1 [6].
In another proteomic study with diabetes, influence of grape
seed proanthocyanidin extract (GSPE) on proteins involved
in oxidative stress, glycated hemoglobin, and AGE products
was determined. Differentially expressed proteins involved
in oxidative stress, glycosylation damage, and amino acids
metabolism were back regulated to normal levels after GSPE
therapy [7]. This differential expression of proteins could be
due to modification of these proteins by glycation and for-
mation of AGEs. It is important to identify AGE modifica-
tion of proteins and their expression to understand the
molecular mechanism of diabetic nephropathy.
Mass spectrometry has been extensively used to identify
and characterize proteins modified by glycation in both in
vitro and in vivo conditions [8]. In our previous study, we
have used MALDI-TOF-MS to study in vitro glycation of
insulin and albumin [9, 10]. In this study using Anti-AGE
antibodies, we have detected the proteins modified by
AGEs from the kidney of streptozotocin-induced diabetic
rat by using two-dimensional electrophoresis coupled with
western blot analysis. These proteins were characterized
for glycation modification by using peptide mass finger-
printing and tandem mass spectrometric approaches.
Experimental
Chemicals
All chemicals were purchased from Sigma-Aldrich unless
otherwise stated. Bradford protein estimation kit purchased
from the Bio-Rad chemicals. Anti-AGE antibodies obtained
from Millipore, whereas goat anti rabbit IgGs biotin conju-
gate, BCIP (5-Bromo-4-Chloro-30-Indolyphosphate p-Tolui-
dine Salt) and NBT (Nitro-Blue Tetrazolium Chloride)
substrate and streptavidin conjugated alkaline phosphatase
(ALP) were purchased from Bangalore Genei, microalbumin
detection kit was purchased from Chemelex, S.A
Induction of Diabetes and Kidney Sample Collection
All animal experiments were carried out at Sinhgad college
of Pharamacy approved by Institution Animal Ethics
Committee. Diabetes was induced by injecting 55 mg of
STZ/Kg body weight intra-peritoneally to the overnight
fasting Wistar rats. Animals surviving after diabetes
induction were maintained for 100–120 days. The diabetic
status of the rats was monitored by measuring their blood
glucose using glucometer (Entrust, Bayer India) and gly-
cated hemoglobin (HBA1c) using HBA1c analyzer (In2it,
Bio-Rad). Animals with higher HBA1c ([8%) were
selected for kidney collection (Table 1). Kidney tissue was
collected after killing the rats with CO2. The tissue was
washed with cold saline for three to four times and stored at
-80�C.
Microalbuminuria
Microalbumin from rat urine was measured by using the
antibody based kit from Chemelex S. A. according to the
procedure described by the manufacturers.
Histopathology of the Kidney Tissue
After killing the animals by exposing to CO2, kidneys were
removed and immediately placed in 10% formaldehyde.
The kidneys were embedded in paraffin and sectioned
using a microtome to get 5 lm thick sections for histo-
logical studies. Sections were stained with hematoxylin–
eosin stain and observed under microscope at 209
magnification.
Sample Preparation
Kidney was perfused with cold phosphate buffer saline to
remove blood stains prior to homogenization. The tissue
was homogenized to fine powder in liquid nitrogen and
washed two to three times with chilled acetone. Then, the
acetone powder was dissolved in buffer consisting of 7 M
Urea, 2 M Thiourea, 2% CHAPS, 1% DTT, 40 mM Tris,
and centrifuged at 14,0009g for 30 min at 4�C in the
HERMLE centrifuge. The supernatant was collected and
stored in aliquots at -80�C. Protein concentration was
estimated by Bradford method.
Electrophoresis and Western Blotting
Proteins were initially separated by isoelectric focusing, and
second dimension SDS-PAGE of the proteins was done
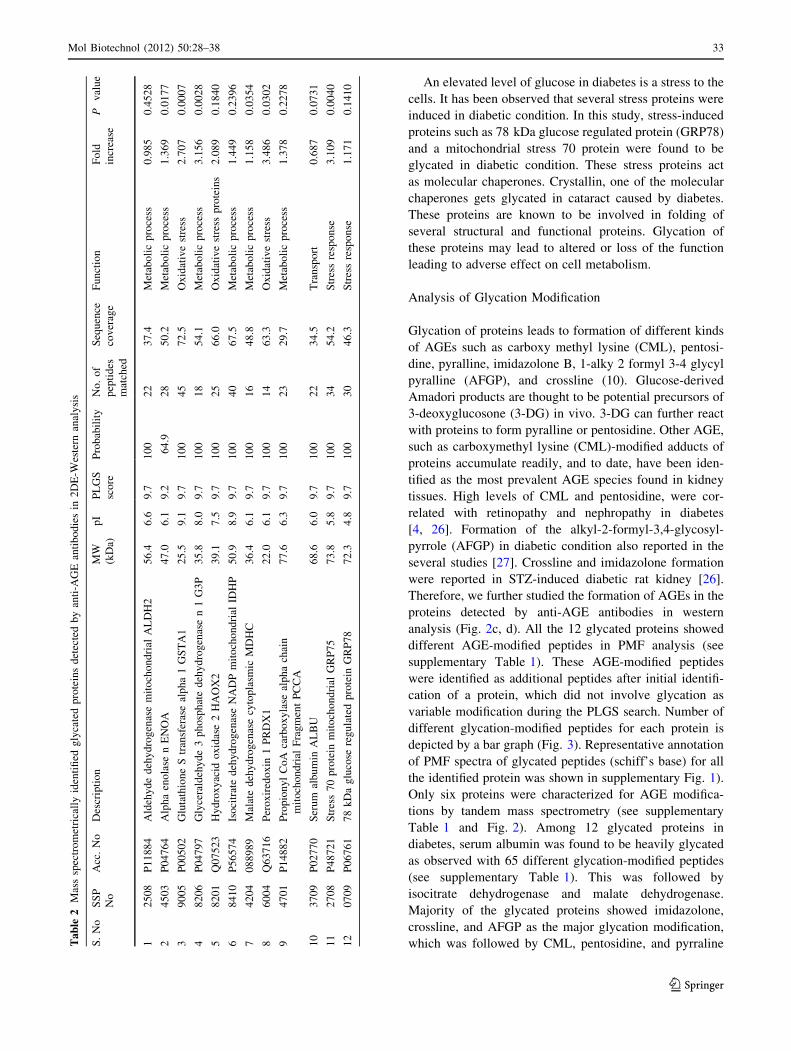
Table 1 Blood glucose, HbA1C level, and albumin excretion rate of diabetic and control rats
Animal model Body weight (g) Glucose (mg/dl) HbA1 (%) Albumin excretion
rate (lg/24 h)
Control (6) 225 ± 15 60.00 ± 10 4.5 ± 1.8 241.80 ± 71
Diabetic (6) 200 ± 10 371.47 ± 20* 8.5 ± 2.0* 1561.00 ± 37*
Value in parenthesis indicates the total number of rats in the group and (±) indicates the standard deviations. * P \ 0.005
Mol Biotechnol (2012) 50:28–38 29
123

essentially according to Laemmli [11]. Proteins on the gel
were visualized by coomassie brilliant blue (CBB) R-250
staining (Sigma). For protein identification, isoelectric
focusing was carried out by rehydrating 11 cm (pH 3–10) IPG
dry strips (BioRad) with 200 lg of proteins dissolved in
200 ll rehydration buffer (7 M Urea, 2 M thiourea, 40 mM
Tris 1% DTT, and 2% CHAPS) and focused at 8,000 V for
35,000 Vh using Protean IEF Cell (BioRad). Whereas for 2-D
western blot, 7 cm (pH 3–10) IPG dry strips (BioRad) were
rehydrated with 100 lg of protein dissolved in 100 ll rehy-
dration buffer (7 M Urea, 2 M thiourea, 40 mM Tris 1% DTT,
and 2% CHAPS) and focused at 4,000 V for 10,000 Vh using
Protean IEF Cell (BioRad). After isoelectric focusing, the IPG
strips were reduced and alkylated by using equilibration
buffers containing dithiothreitol and iodoacetamide, respec-
tively. Separation in the second dimension was performed on a
12% SDS-PAGE. After electrophoresis, 11 cm gels were used
for CBB staining and 7 cm gels were used for western blot-
ting. Proteins were transferred to PVDF membrane and
incubated overnight at 4�C in blocking buffer containing 2%
milk powder dissolved in 20 mM Tris–Cl (pH 7.5), 0.15 M
NaCl (TBS). The membranes were washed with TBS and
incubated for 1 h at 37�C with diluted (1:5,000) anti-AGE
antibodies (Millipore). After three washes with washing
buffer (TBS containing 0.05% Tween 20), the membranes
were incubated with goat anti rabbit IgGs biotin conjugate
(Bangalore Genei) at a dilution of 1:1,000 for 30 min at room
temperature. Again after three washes with washing buffer,
membranes were incubated with streptavidin-conjugated
alkaline phosphatase at dilution 1:2,000 for 30 min at RT.
Immunodetection was done by adding BCIP/NBT substrate of
ALP in dark.
Image Analysis and In-Gel Digestion
The Coomassie-stained gels (3–10 pH range) were scanned
using a densitometer GS-710 (Bio-Rad Laboratories)
and analyzed using advanced PD-Quest Software version
(Bio-Rad Laboratories). Gel patterns from each independent
analysis were matched and the relative abundance of each
spot was compared between control and diabetic gel. This
analysis was done for three biological replicates. Differen-
tially expressed protein spots were destained with 100 ll of
destaining solution (50 mM ammonium bicarbonate/aceto-
nitrile mixed 1:1 v/v). After thorough rinsing with 100 mM
ammonium bicarbonate, the gel pieces were dehydrated in
100% ACN, which was removed by air drying. The proteins
were in-gel digested overnight at 37�C with 15–20 ll of
20 ng/ll trypsin (Sigma) in 25 mM ammonium bicarbonate
buffer pH 8.4. Peptides were recovered by extraction with
50% ACN5% TFA. Tryptic protein digests were reconsti-
tuted in 50% ACN with 0.1% TFA solvent before subjecting
to mass spectrometric analysis.
Mass Spectrometric Analysis
1 ll of the reconstituted protein digest was premixed with
equal volume of CHCA matrix, vortexed well before
spotting on 96-well MALDI plate. Glu-Fibrinopeptide was
used as an external calibrant (lock mass). Mass spectro-
metric analysis was done on MALDI-Q-TOF (Synapt-
HDMS, M/S Waters). All mass spectra were acquired
200 Hz solid state UV laser in V mode by MassLynx 4.1.
The quadrupole profile was set to 500 m/z for 5% of the
scan time and then ramped to 1,500 m/z for remaining
period of the scan. The instrument mass calibration was
carried with Polyethylene glycol (PEG) MALDI mixture
provided by M/s Waters. Proteins were identified by
MALDI survey method, which uses combination of PMF
and tandem mass spectrometric (MS/MS) approaches for
identification of proteins. In the survey or MS mode, spectra
were recorded in the mass range of 800 to 4,000 m/z for
60 s. MS/MS analysis was done in a data dependent
manner for the top 10 peptides with higher relative inten-
sity for 30 s each. The product ion mass range was set to
100 to 1,500 m/z.
Protein Identification
The resulting MS, MS/MS data were analyzed by Protein
Lynx Global Survey (PLGS) software (Waters, India). Four
spectra were acquired for each protein. The data were
searched against Rattus norvegicus species database
(downloaded from UNIPROT) using PLGS search engine
with a mass tolerance set to 100 ppm and 0.03 Da for MS
and MS/MS, respectively. Fixed modification was set to
carbamidomethylation and variable modification was set to
methionine oxidation. Only those proteins were considered
identified where a same protein was identified by PLGS
with the top score in all four replications. The protein IDs
were examined for sequence coverage, number of peptides
matched agreement between theoretical and experimental
gel MW, and pI values and matching of major peaks of
PMF with the peptides identified in the protein.
PTM Analysis
After the protein identification by PLGS, the spectra were
again subjected to the PLGS search by including glycation
modifications in the search criteria. Modifications were
defined for Schiff’s base or Amodori product (?162.02),
pentosidine (?58.03) and carboxy methyl lysine (?58.02),
pyralline (?108.02), imidazolone (?142.03), crossline
(?252.11), and 1-Alkyl-2-formyl-3,4-glycosyl-pyrrole
(AFGP) (?270.07) as described [12, 13]. Only those pro-
teins were considered identified where a same protein was
identified by PLGS with the top score in all different
30 Mol Biotechnol (2012) 50:28–38
123

searches with different glycation modifications. Further-
more, each PMF spectrum was manually annotated for the
presence of a peak with increased mass caused by glyca-
tion. And the MS/MS spectrum of glycated peptide
(Amodori product or Schiffs base) was annotated for the
neutral loss or any/all of the following ion (M ? H - 18),
(M ? H - 36), (M ? H - 120), (M ? H - 162) accord-
ing to the procedure described by Brancia et al. [14]. The
same approach was also used to annotate the other glyca-
tion modifications.
Quantification of Glycation
Protein glycation was quantified by comparing the actual
number of protein spots detected by anti-AGE antibody in
the western blot analysis. Wherever a same protein was
detected to be glycated in both control and diabetes, the
extent of glycation was quantified by comparing
(1) number of protein specific glycated peptides in
control and diabetes,
(2) the intensity of protein specific glycated peptide in the
PMF spectrum from control and diabetes,
(3) the densities of glycated spots in control and diabetes
in the western analysis.
Statistical Analysis
Statistical analysis was done for following parameters
(a) Intensity of glycation modified peptides:
Heat map analysis was done to study the extent of gly-
cation modification specific glycated peptides in control and
diabetes. The average ion count of individual modified
peptide from three replication was converted to per cent
relative intensity for generation of heat map analysis. Heat
map analysis was done using MeV Multiexperiment Viewer
Version 4.6 according to Saeed et al. [15]. The relative
intensities were color coded from light green to dark red for
lower intensity to higher intensity, respectively.
(b) For protein expression analysis
The CBB stained 2D gels were analyzed by PDQUEST
image analysis software. The density of each protein was
expressed as mean ± SD for three experiments. The sta-
tistical significance was established by Student’s t-test.
Differences were considered significant if P B 0.05.
Results and Discussion
Identification of Glycated Proteins
Glycation of kidney proteins was studied in STZ-induced
diabetic rats having nephropathic symptoms. The histopa-
thology of glomeruli shows the basement membrane
thickening and mesangial proliferation without nodule
formation in diabetic rat kidney (Fig. 1). The HBA1c was
8.5% and albumin excretion rate (AER) was about
1,561 lg/24 h (Table 1) in diabetic rats, whereas in control
rats the HBA1c was 4.5% and AER was 241 lg/24 h.
These tissues were used to identify and characterize gly-
cated proteins by using combination of approaches
involving 2DE, western analysis with anti-AGE antibodies
and mass spectrometry. The 2DE-western analysis led to
detection of 38 well-resolved glycated protein spots in
diabetes and 17 glycated spots in control sample (Fig. 2).
Out of these glycated proteins detected, 12 proteins were
identified in diabetes, and 3 proteins were identified from
control. (Other glycated proteins were not identified as we
did get the same first hit in all the four replications using
PLGS. Henceforth only 12 proteins identified were
Fig. 1 Transverse section of
the kidney glomerulus of
a control rat and b diabetic rat,
indicating basement thickening
Mol Biotechnol (2012) 50:28–38 31
123

considered for this study.) These 12 glycated proteins
identified belong to proteins involved in metabolic path-
ways, oxidative stress, stress proteins, and gene regulation
(Table 2). Glyceraldehyde-3-phosphate dehydrogenase
(GAPDH), malate dehydrogenase (MDH) a-enolase, iso-
citrate dehydogenase, aldehyde dehydrogenase, and pro-
pionyl carboxylase constituted important glycated proteins
involved in metabolic pathways. It was interesting to find
several proteins involved in glucose metabolism were the
targets of glycation. An earlier study has shown that gly-
cation of GAPDH decreases its activity in rabbit muscle
and human erythrocyte [16, 17]. Additionally, it was also
demonstrated that the glycation of GAPDH contributes to
the development of diabetic vascular complications by
activating generation of intracellular advanced glycation
end products [18, 19]. Glycation of isocitrate dehydroge-
nase at its active site lysine residue (K212) reduces the
activity of the enzyme and disturbs the antioxidant system
as it is NADPH dependent enzyme [20]. Similarly, glyca-
tion of mitochondrial isocitrate dehydrogenase was
observed in the kidneys of diabetic rats and in the lenses of
diabetic patients suffering from cataracts, accompanied
with a decrease in the enzyme activity. The glycation-
mediated damage to isocitrate dehydrogenase may result in
the perturbation of cellular antioxidant defense mecha-
nisms and subsequently may contribute to development of
various pathologies associated with long-term complica-
tions of diabetes. The glycation of propionyl CoA car-
boxylase was not known earlier.
Glycation and oxidative stress act synergistically
in developing vascular complications in hyperglycemic
condition [21–23]. In this study, it was observed that the
oxidative enzymes like glutathione S transferase, perrox-
iredoxin and hydroxyacid oxidase were glycated. Glutathi-
one S transferase is an antioxidant protein, whose activity
gets reduced in long-term hyperglycemia. Perhaps, the
decrease in the activity of glutathione S transferase could be
due to glycation [24, 25]. The activity of perroxiredoxin has
been shown to increase in diabetic condition, in our study the
protein was found to be glycated [6]. Glycation of hydroxy
acid oxidase in kidney was also observed in the present study.
Glycation of such proteins largely may affect the oxidative
status of the tissue and leads to the oxidative stress which is
responsible for the damage of tissue.
A B
C D
Fig. 2 Two dimensional
electrophoresis of rat kidney
proteins either stained with
coomassie brilliant blue (a) and
(b), or immunoblotted with
anti-AGE antibodies (c) and
(d) of control and diabetes,
respectively
32 Mol Biotechnol (2012) 50:28–38
123
An elevated level of glucose in diabetes is a stress to the
cells. It has been observed that several stress proteins were
induced in diabetic condition. In this study, stress-induced
proteins such as 78 kDa glucose regulated protein (GRP78)
and a mitochondrial stress 70 protein were found to be
glycated in diabetic condition. These stress proteins act
as molecular chaperones. Crystallin, one of the molecular
chaperones gets glycated in cataract caused by diabetes.
These proteins are known to be involved in folding of
several structural and functional proteins. Glycation of
these proteins may lead to altered or loss of the function
leading to adverse effect on cell metabolism.
Analysis of Glycation Modification
Glycation of proteins leads to formation of different kinds
of AGEs such as carboxy methyl lysine (CML), pentosi-
dine, pyralline, imidazolone B, 1-alky 2 formyl 3-4 glycyl
pyralline (AFGP), and crossline (10). Glucose-derived
Amadori products are thought to be potential precursors of
3-deoxyglucosone (3-DG) in vivo. 3-DG can further react
with proteins to form pyralline or pentosidine. Other AGE,
such as carboxymethyl lysine (CML)-modified adducts of
proteins accumulate readily, and to date, have been iden-
tified as the most prevalent AGE species found in kidney
tissues. High levels of CML and pentosidine, were cor-
related with retinopathy and nephropathy in diabetes
[4, 26]. Formation of the alkyl-2-formyl-3,4-glycosyl-
pyrrole (AFGP) in diabetic condition also reported in the
several studies [27]. Crossline and imidazolone formation
were reported in STZ-induced diabetic rat kidney [26].
Therefore, we further studied the formation of AGEs in the
proteins detected by anti-AGE antibodies in western
analysis (Fig. 2c, d). All the 12 glycated proteins showed
different AGE-modified peptides in PMF analysis (see
supplementary Table 1). These AGE-modified peptides
were identified as additional peptides after initial identifi-
cation of a protein, which did not involve glycation as
variable modification during the PLGS search. Number of
different glycation-modified peptides for each protein is
depicted by a bar graph (Fig. 3). Representative annotation
of PMF spectra of glycated peptides (schiff’s base) for all
the identified protein was shown in supplementary Fig. 1).
Only six proteins were characterized for AGE modifica-
tions by tandem mass spectrometry (see supplementary
Table 1 and Fig. 2). Among 12 glycated proteins in
diabetes, serum albumin was found to be heavily glycated
as observed with 65 different glycation-modified peptides
(see supplementary Table 1). This was followed by
isocitrate dehydrogenase and malate dehydrogenase.
Majority of the glycated proteins showed imidazolone,
crossline, and AFGP as the major glycation modification,
which was followed by CML, pentosidine, and pyrralineTa
ble
2M
ass
spec
tro
met
rica
lly
iden
tifi
edg
lyca
ted
pro
tein
sd
etec
ted
by
anti
-AG
Ean
tib
od
ies
in2
DE
-Wes
tern
anal
ysi
s
S.
No
SS
P
No
Acc
.N
oD
escr
ipti
on
MW
(kD
a)
pI
PL
GS
sco
re
Pro
bab
ilit
yN
o.
of
pep
tid
es
mat
ched
Seq
uen
ce
cov
erag
e
Fu
nct
ion
Fo
ld
incr
ease
Pv
alu
e
12
50
8P
11
88
4A
ldeh
yd
ed
ehy
dro
gen
ase
mit
och
on
dri
alA
LD
H2
56
.46
.69
.71
00
22
37
.4M
etab
oli
cp
roce
ss0
.98
50
.45
28
24
50
3P
04
76
4A
lph
aen
ola
sen
EN
OA
47
.06
.19
.26
4.9
28
50
.2M
etab
oli
cp
roce
ss1
.36
90
.01
77
39
00
5P
00
50
2G
luta
thio
ne
Str
ansf
eras
eal
ph
a1
GS
TA
12
5.5
9.1
9.7
10
04
57
2.5
Ox
idat
ive
stre
ss2
.70
70
.00
07
48
20
6P
04
79
7G
lyce
rald
ehy
de
3p
ho
sph
ate
deh
yd
rog
enas
en
1G
3P
35
.88
.09
.71
00
18
54
.1M
etab
oli
cp
roce
ss3
.15
60
.00
28
58
20
1Q
07
52
3H
yd
rox
yac
ido
xid
ase
2H
AO
X2
39
.17
.59
.71
00
25
66
.0O
xid
ativ
est
ress
pro
tein
s2
.08
90
.18
40
68
41
0P
56
57
4Is
oci
trat
ed
ehy
dro
gen
ase
NA
DP
mit
och
on
dri
alID
HP
50
.98
.99
.71
00
40
67
.5M
etab
oli
cp
roce
ss1
.44
90
.23
96
74
20
40
88
98
9M
alat
ed
ehy
dro
gen
ase
cyto
pla
smic
MD
HC
36
.46
.19
.71
00
16
48
.8M
etab
oli
cp
roce
ss1
.15
80
.03
54
86
00
4Q
63
71
6P
ero
xir
edo
xin
1P
RD
X1
22
.06
.19
.71
00
14
63
.3O
xid
ativ
est
ress
3.4
86
0.0
30
2
94
70
1P
14
88
2P
rop
ion
yl
Co
Aca
rbo
xy
lase
alp
ha
chai
n
mit
och
on
dri
alF
rag
men
tP
CC
A
77
.66
.39
.71
00
23
29
.7M
etab
oli
cp
roce
ss1
.37
80
.22
78
10
37
09
P0
27
70
Ser
um
alb
um
inA
LB
U6
8.6
6.0
9.7
10
02
23
4.5
Tra
nsp
ort
0.6
87
0.0
73
1
11
27
08
P4
87
21
Str
ess
70
pro
tein
mit
och
on
dri
alG
RP
75
73
.85
.89
.71
00
34
54
.2S
tres
sre
spo
nse
3.1
09
0.0
04
0
12
07
09
P0
67
61
78
kD
ag
luco
sere
gu
late
dp
rote
inG
RP
78
72
.34
.89
.71
00
30
46
.3S
tres
sre
spo
nse
1.1
71
0.1
41
0
Mol Biotechnol (2012) 50:28–38 33
123

modifications (Fig. 3). In diabetes, human serum albumin
(HSA) was one of the heavily glycated proteins in the
plasma [28]. The biological impact of glycated albumin on
cell physiology and its role as a biological marker of
diabetes was recently reviewed in great detail [29].
Quantification of Glycation
It was natural to observe more number of glycated proteins
in the kidney of STZ-induced diabetic rat. However, in
addition to those specifically glycated proteins in diabetes,
few glycated proteins were also detected to control by
western blot analysis using anti-AGE antibody. These
glycated proteins were GAPDH, serum albumin, and MDH
(Fig. 4). Therefore, in these glycated proteins a comparison
between control and diabetes was made to establish the
extent of glycation. The extent of glycation was measured
by, (a) density of glycated spot in western blot, (b) number
of glycation modified peptides, and (c) intensity of glyca-
tion modified peptide. In the western blot analysis using
anti-AGE antibodies, it was observed that the density of
glycated forms of serum albumin, GAPDH, and MDH was
higher in diabetes than that of control (Fig. 4a). In all these
three glycated proteins, the number of glycated peptides
were found to be more in diabetes than that of its corre-
sponding control protein (Fig. 4b). These glycated pep-
tides included peptides modified with schiff’s base, CML,
pyralline, pentosidine, imidazolone, crossline, and AFGP.
Furthermore, the intensities of different glycation-modified
peptides were compared between control and diabetes. A
representative mass spectra of PMF analysis of GAPDH
control and diabetes was shown in Fig. 5a. The intensities
of encircled glycated peptide (2,547 m/z) clearly suggest
that extent of glycation was higher in diabetes than control.
Fig. 3 Bar graph representing
number of different glycation
modified peptides in each of the
glycated proteins from kidney
of diabetic rat
34 Mol Biotechnol (2012) 50:28–38
123
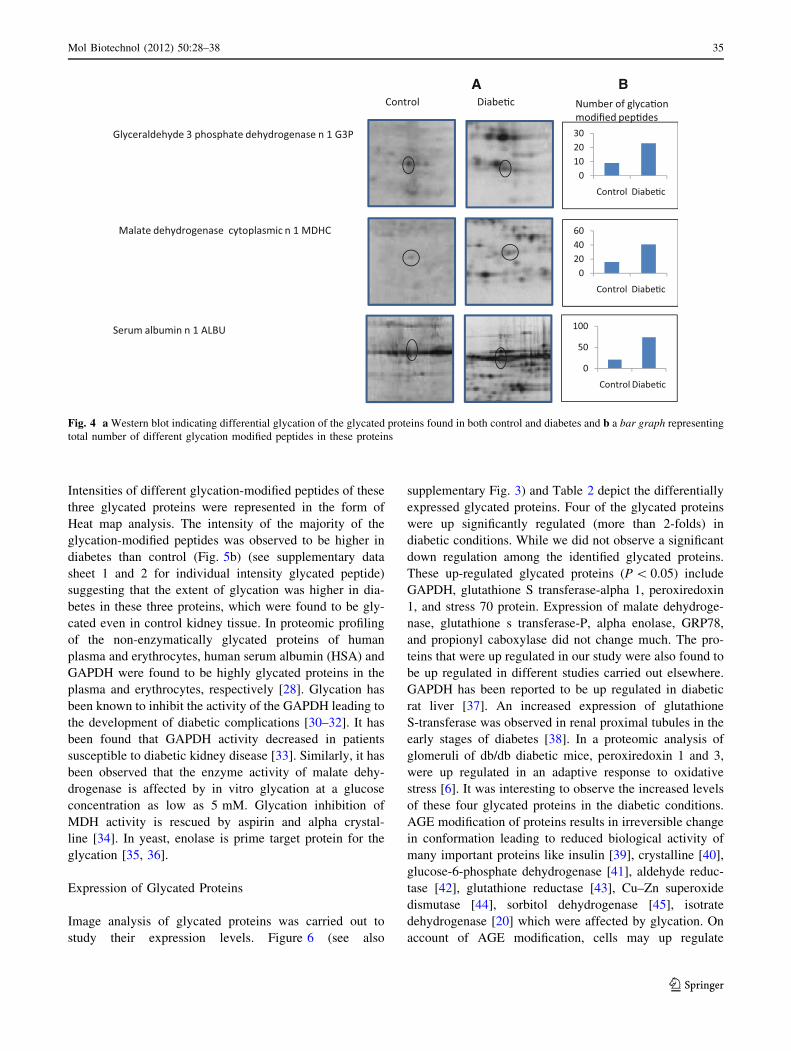
Intensities of different glycation-modified peptides of these
three glycated proteins were represented in the form of
Heat map analysis. The intensity of the majority of the
glycation-modified peptides was observed to be higher in
diabetes than control (Fig. 5b) (see supplementary data
sheet 1 and 2 for individual intensity glycated peptide)
suggesting that the extent of glycation was higher in dia-
betes in these three proteins, which were found to be gly-
cated even in control kidney tissue. In proteomic profiling
of the non-enzymatically glycated proteins of human
plasma and erythrocytes, human serum albumin (HSA) and
GAPDH were found to be highly glycated proteins in the
plasma and erythrocytes, respectively [28]. Glycation has
been known to inhibit the activity of the GAPDH leading to
the development of diabetic complications [30–32]. It has
been found that GAPDH activity decreased in patients
susceptible to diabetic kidney disease [33]. Similarly, it has
been observed that the enzyme activity of malate dehy-
drogenase is affected by in vitro glycation at a glucose
concentration as low as 5 mM. Glycation inhibition of
MDH activity is rescued by aspirin and alpha crystal-
line [34]. In yeast, enolase is prime target protein for the
glycation [35, 36].
Expression of Glycated Proteins
Image analysis of glycated proteins was carried out to
study their expression levels. Figure 6 (see also
supplementary Fig. 3) and Table 2 depict the differentially
expressed glycated proteins. Four of the glycated proteins
were up significantly regulated (more than 2-folds) in
diabetic conditions. While we did not observe a significant
down regulation among the identified glycated proteins.
These up-regulated glycated proteins (P \ 0.05) include
GAPDH, glutathione S transferase-alpha 1, peroxiredoxin
1, and stress 70 protein. Expression of malate dehydroge-
nase, glutathione s transferase-P, alpha enolase, GRP78,
and propionyl caboxylase did not change much. The pro-
teins that were up regulated in our study were also found to
be up regulated in different studies carried out elsewhere.
GAPDH has been reported to be up regulated in diabetic
rat liver [37]. An increased expression of glutathione
S-transferase was observed in renal proximal tubules in the
early stages of diabetes [38]. In a proteomic analysis of
glomeruli of db/db diabetic mice, peroxiredoxin 1 and 3,
were up regulated in an adaptive response to oxidative
stress [6]. It was interesting to observe the increased levels
of these four glycated proteins in the diabetic conditions.
AGE modification of proteins results in irreversible change
in conformation leading to reduced biological activity of
many important proteins like insulin [39], crystalline [40],
glucose-6-phosphate dehydrogenase [41], aldehyde reduc-
tase [42], glutathione reductase [43], Cu–Zn superoxide
dismutase [44], sorbitol dehydrogenase [45], isotrate
dehydrogenase [20] which were affected by glycation. On
account of AGE modification, cells may up regulate
A B
Fig. 4 a Western blot indicating differential glycation of the glycated proteins found in both control and diabetes and b a bar graph representing
total number of different glycation modified peptides in these proteins
Mol Biotechnol (2012) 50:28–38 35
123

Fig. 5 a Representative peptide
mass finger printing analysis of
glyceraldehyde 3 phosphate
dehydrogenase from control
(i) and diabetes (ii). The
encircled peaks correspond to
mass of glycated peptides.
b Heat map analysis of
intensities of different
glycation-modified peptides of
glycated proteins from control
and diabetes
36 Mol Biotechnol (2012) 50:28–38
123

expression of those glycated proteins to compensate for the
loss of biological activity caused by glycation. In fact,
glycation-induced inactivation of sorbitol dehydrogenase
was accompanied with increased levels of mRNA and
protein in liver of STZ-induced diabetic rats [45]. Alter-
natively, the increased level of such proteins could be due
to formation of protease resistant protein aggregates caused
by AGE modification as well as by decreased proteolytic
activity. It has been observed that the glomerular protein-
ase activity was decreased in the streptozotocin-induced
diabetic rats [46]. Additionally, methyl glyoxal covalently
modifies the 20S proteasome, decreasing its activity in the
diabetic kidney [47]. All these factors may contribute
toward elevated levels of glycated proteins.
In conclusion, we have comprehensively analyzed the
glycation of kidney proteins from STZ-induced diabetic
and control rats. The number of glycated proteins detected
in the diabetic kidney were more than that of control.
Additionally, the extent of glycation was higher in few
glycated proteins that were present in both control and
diabetic kidney. Moreover, some of these glycated proteins
were found to be up regulated in diabetes. Functional
characterization of such glycated proteins may help better
in understanding their role in development of nephropathy.
Acknowledgments This work was carried out as a part of network
project NWP-4 supported by Council of Scientific and Industrial
Research, India. Authors thank Dr. Vidya Gupta, Chair, Biochemical
Sciences, Dr. S. Sivaram and Dr. Sourav Pal, Director, NCL for their
support and encouragement. We also thank Dr. Chetan Gadgil and
Mr. Avinash Ghanate for their inputs in Heatmap analysis. ADC also
thanks Department of Biotechnology, India for the post-doctoral
fellowship.
References
1. Verbeke, P., Perichon, M., Borot–Laloi, C., Schaeverbeke, J., &
Bakala, H. (1997). Accumulation of advanced glycation end
products in the rat nephron: Link with circulating AGEs during
aging. Journal of Histochemistry and Cytochemistry, 45, 1059–
1068.
2. Figueroa-Romero, C., Sadidi, M., & Feldman, E. L. (2008).
Feldman mechanisms of disease: The oxidative stress theory of
diabetic neuropathy. Reviews in Endocrine & Metabolic Disor-ders, 9, 301–314.
3. Ahmad, W., Li, L., & Deng, Y. (2008). Identification of AGE-
precursors and AGE formation in glycation-induced BSA
peptides. BMB Reports, 41, 516–522.
4. Tangi, N., Markowitz, G. S., Fu, C., Kislinger, T., Taguchi, A.,
Pischetsrieder, M., et al. (2000). Expression of advanced glyca-
tion end products and their cellular receptor RAGE in diabetic
nephropathy and nondiabetic renal disease. Journal of theAmerican Society of Nephrology, 11, 1656–1666.
5. Rosca, M. G., Mustata, T. G., Kinter, M. T., Ozdemir, A. M.,
Kern, T. S., Szweda, L., et al. (2005). Glycation of mitochondrial
proteins from diabetic rat kidney is associated with excess
superoxide formation. American Journal of Physiology: RenalPhysiology, 289, 420–430.
6. Barati, M. T., Merchant, M. L., Kain, A. B., Jevans, A. W.,
McLeish, K. R., & Klein, J. B. (2007). Proteomic analysis defines
altered cellular redox pathways and advanced glycation end-
product metabolism in glomeruli of db/db diabetic mice.
American Journal of Physiology: Renal Physiology, 293(4),
1157–1165.
7. Li, B., Cheng, M., Gao, H., Ma, Y. B., Xu, L., Li, X. H., et al.
(2008). Back-regulation of six oxidative stress proteins with
grape seed proanthocyanidin extracts in rat diabetic nephropathy.
Journal of Cellular Biochemistry, 104, 668–679.
8. Lapolla, A., Fedele, D., Seraglia, R., & Traldi, P. (2006). The role
of mass spectrometry in the study of non-enzymatic protein
glycation in diabetes: An update. Mass Spectrometry Reviews, 25,
775–797.
9. Bhonsle, H. S., Singh, S. K., Srivastava, G., Boppana, R., &
Kulkarni, M. J. (2008). Albumin competitively inhibits glycation
of less abundant proteins. Protein & Peptide Letters, 15,
663–667.
10. Golegaonkar, S. B., Bhonsle, H. S., Boppana, R., & Kulkarni, M.
J. (2010). Discovery of rifampicin as a new anti-glycating com-
pound by matrix-assisted laser desorption/ionization mass spec-
trometry-based insulin glycation assay. European Journal ofMass Spectrometry, 16, 221–226.
11. Laemmli, U. K. (1970). Cleavage of structural proteins during
the assembly of the head of bacteriophage T4. Nature, 227, 680–
685.
12. Zhang, Y., Cocklin, R. R., Bidasee, K. R., & Wang, M. (2003).
Rapid determination of advanced glycation end products of
proteins using MALDI-TOF-MS and PERL script peptide
searching algorithm. Journal of Biomolecular Techniques, 4,
224–230.
13. Wa, C., Cerny, R. L., Clarke, W. A., & Hage, D. S. (2007).
Characterisation of glycation adduction human serum albumin by
MALDI-TOF/MS. Clinica Chimica Acta, 385(1–2), 48–60.
14. Brancia, F. L., Bereszczak, J. Z., Lapolla, A., Fedel, D., Baccarin,
L., Seraglia, R., et al. (2006). Comprehensive analysis of glycated
human serum albumin tryptic peptides byoffline liquid chroma-
tography followed by MALDI analysis on time-of-flight/curved
field reflectron tandem mass spectrometer. Mass Spectrometry,41(9), 1179–1185.
Fig. 6 Representation of increased levels of glycated proteins from
control and diabetic rat kidney analyzed by PDQUEST (Bio-Rad)
having fold increase more than 2 and P \ 0.05
Mol Biotechnol (2012) 50:28–38 37
123

15. Saeed, A. L., Bhagabati, N. K., Bralsted, J. C., Liang, W., Sharov,
V., Howe, E. A., et al. (2006). TM4 microarray software suite.
Methods in Enzymology, 411, 134–193.
16. He, R. Q., Yang, M. D., Zheng, X., & Zhou, J. X. (1995). Iso-
lation and some properties of glycated D-glyceraldehyde-3-
phosphate dehydrogenase from rabbit muscle. BiochemicalJournal, 309, 133–139.
17. He, R. Q., Li, Y. G., Wu, X. Q., & Li, L. (1995). Inactivation and
conformation changes of the glycated and non-glycated D-gly-
ceraldehyde-3-phosphate dehydrogenase during guanidine-HCl
denaturation. Biochimica et Biophysica Acta, 1253, 47–56.
18. Kiss, L., & Szabo, C. (2005). The pathogenesis of diabetic
complications: the role of DNA injury and poly, ADP-ribose,
polymerase activation in peroxynitrite-mediated cytotoxicity.
Memorias do Instituto Oswaldo Cruz, 100, 29–37.
19. Szabo, C. (2009). Role of nitrosative stress in the pathogenesis of
diabetic vascular dysfunction. British Journal of Pharmacology,156, 13–27.
20. Kil, I. S., Lee, J. H., Shin, A. H., & Park, J. W. (2004). Glycation-
induced inactivation of NADP(?)-dependent isocitrate dehydro-
genase: implications for diabetes and aging. Free Radical Biologyand Medicine, 37, 1765–1778.
21. Lal, M. A., Brismar, H., Eklof, A., & Aperia, A. (2002). Role of
oxidative stress in advanced glycation end product-induced
mesangial cell activation. Kidney International, 61, 2006–2014.
22. Node, K., & Inoue, T. (2009). Postprandial hyperglycemia as an
etiological factor in vascular failure. Cardiovascular Diabetol-ogy, 8–23.
23. Coughlan, M. T., Mibus, A. L., & Forbes, J. M. (2008). Oxidative
stress and advanced glycation in diabetic nephropathy. Annals ofthe New York Academy of Sciences, 1126, 190–193.
24. Petlevski, R., Hadzija, M., Slijepcevic, M., & Petrik, D. (2003).
Glutathione S-transferases and malondialdehyde in the liver of
NOD mice on short-term treatment with plant mixture extract
P-9801091. Journal of Phytotherapy Research, 17(4), 311–314.
25. Lapshina, E. A., Sudnikovich, E. J., Maksimchik, J. Z.,
Zabrodskaya, S. V., Zavodnik, L. B., Kubyshin, V. L., et al.
(2006). Antioxidative enzyme and glutathione S-transferase
activities in diabetic rats exposed to long-term ASA treatment.
Life Sciences, 79(19), 1804–1811.
26. Lapolla, A., Traldi, P., & Fedele, D. (2005). Importance of
measuring products of non-enzymatic glycation of proteins.
Clinical Biochemistry, 38, 103–115.
27. Bidasee, K. R., Zhang, Y., Shao, C. H., Wang, M., Patel, K. P.,
Dincer, D., et al. (2004). Diabetes increases formation of
advanced glycation end products on sarco(endo)plasmic reticu-
lum Ca2?-ATPase. Diabetes, 53, 463–473.
28. Zhang, Q., Tang, N., Schepmoes, A. A., Phillips, L. S., Smith, R.
D., & Metz, T. O. (2008). Proteomic profiling of nonenzymati-
cally glycated proteins in human plasma and erythrocyte mem-
branes. Journal of Proteome Research, 7(5), 2025–2032.
29. Rondeau, P., & Bourdon, E. (2011). The glycation of albumin:
Structural and functional impacts. Biochimie. doi:10.1016/j.biochi.
2010.12.003.
30. Morgan, P. E., Dean, R. T., & Davies, M. J. (2002). Inactivation
of cellular enzymes by carbonyls and protein-bound glycation/
glycoxidation products. Archives of Biochemistry and Biophysics,403, 259–269.
31. Ray, M., Basu, N., & Ray, S. (1997). Inactivation of glycer-
aldehyde-3-phosphate dehydrogenase of human malignant cells
by methylglyoxal. Molecular and Cellular Biochemistry, 177,
21–26.
32. Hook, D. W., & Harding, J. J. (1997). Inactivation of glyceral-
dehyde 3-phosphate dehydrogenase by sugars, prednisolone-21-
hemisuccinate, cyanate and other small molecules. Biochimica etBiophysica Acta, 1362, 232–242.
33. Lee, H. J., Howell, S. K., Sanford, R. J., & Beisswenger, P. J.
(2005). Methylglyoxal can modify GAPDH activity and struc-
ture. Annals of the New York Academy of Sciences, 1043,
135–145.
34. Heath, M. M., Rixon, K. C., & Harding, J. J. (1996). Glycation-
induced inactivation of malate dehydrogenase protection by
aspirin and a lens molecular chaperone, alpha-crystallin. Bio-chimica et Biophysica Acta, 12, 176–184.
35. Pietkiewicz, J., Gamian, A., Staniszewska, M., & Danielewicz, R.
(2009). Inhibition of human muscle-specific enolase by methyl-
glyoxal and irreversible formation of advanced glycation end
products. Journal of Enzyme Inhibition and Medicinal Chemistry,24, 356–364.
36. Gomes, R. A., Vicente, M. H., Silva, M. S., Graca, G., Coelho, A.
V., Ferreira, A. E., et al. (2006). Yeast protein glycation in vivo
by methylglyoxal. FEBS Journal, 273, 5273–5287.
37. Johnson, T. D., Harris, R. A., French, S., Aponte, A., & Balaban,
R. S. (2009). Proteomic changes associated with diabetes in the
BB-DP rat. American Journal of Physiology: Endocrinology andMetabolism, 296, 422–432.
38. Fujitaa, H., Haseyamaa, T., Kayo, T., Nozaki, J., Wada, Y., Ito,
S., et al. (2001). Increased expression of glutathione S-transferase
in renal proximal tubules in the early stages of diabetes: A study
of type-2 diabetes in the Akita mouse model. ExperimentalNephrology, 9, 380–386.
39. Hunter, S. J., Boyd, A. C., O’Harte, F. P. M., McKillop, A. M.,
Wiggam, M. I., Mooney, M. H., et al. (2003). Demonstration of
glycated insulin in human diabetic plasma and decreased bio-
logical activity assessed by euglycemic-hyperinsulinemic clamp
technique in humans. Diabetes, 52, 492–498.
40. Abraham, E. C., Huaqian, J., Aziz, A., Kumarasamy, A., & Datta,
P. (2008). Role of the specifically targeted lysine residues in the
glycation dependent loss of chaperone activity of alpha A- and
alpha B-crystallins. Molecular and Cellular Biochemistry, 310,
235–239.
41. Ganea, E., & Harding, J. J. (1994). Inactivation of glucose-
6-phosphate dehydrogenase by glycation. Biochemical SocietyTransactions, 5S, 22–44.
42. Takahashi, M., Lu, Y., Myint, T., Fujii, J., Wada, Y., & Tanig-
uchi, N. (1995). In vivo glycation of aldehyde reductase, a major
3-deoxyglucosone reducing enzyme: Identification of glycation
sites. Journal of Biochemistry, 34, 1433–1438.
43. Blakytny, R., & Harding, J. J. (1992). Glycation (non-enzymic
glycosylation) inactivates glutathione reductase. Biochemistry,288, 303–307.
44. Arai, K., Maguchi, S., Fuji, S., Ishibashi, H., Oikawa, K., &
Taniguchi, N. (1987). Glycation and inactivation of human Cu–
Zn superoxidase dismutase. Journal of Biological Chemistry,262, 16969–16972.
45. Hoshi, A., Takahashi, M., Fujii, J., Myint, T., Kaneto, H., Suzuki,
K., et al. (1996). Glycation and inactivation of sorbitol dehy-
drogenase in normal and diabetic rats. Biochemical Journal, 318,
119–123.
46. Stauber, W. T., & Fritz, V. K. (1985). Decreased lysosomal
protease content of skeletal muscles from streptozotocin-induced
diabetic rats: A biochemical and histochemical study. The His-tochemical Journal, 17, 613–622.
47. Queisser, M. A., Yao, D., Geisler, S., Hammes, H. P., Lochnit,
G., Schleicher, E. D., et al. (2010). Hyperglycemia impairs pro-
teasome function by methylglyoxal. Diabetes, 59, 670–678.
38 Mol Biotechnol (2012) 50:28–38
123




















