
10017363.pdf - UCL Discovery
269
DISTRIBUTION OF NTTRERGIC NERVES IN THE RAT ADRENAL GLAND: PLASTICITY IN AGING AND DISEASE Thesis Submitted to the University of London for the Degree of Doctor of Philosophy in the Faculty of Science by Mekbeb Afework, B.Sc., M. Phil. Department of Anatomy and Developmental Biology University College London June 1995
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of 10017363.pdf - UCL Discovery

DISTRIBUTION OF NTTRERGIC NERVES IN THE RAT ADRENAL GLAND:
PLASTICITY IN AGING AND DISEASE
Thesis Submitted to the University of London for
the Degree of Doctor of Philosophy in
the Faculty of Science
by
Mekbeb Afework, B.Sc., M. Phil.
Department of Anatomy and Developmental Biology
University College London
June 1995

ProQuest Number: 10017363
All rights reserved
INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10017363
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.This work is protected against unauthorized copying under Title 17, United States Code.
Microform Edition © ProQuest LLC.
ProQuest LLC 789 East Eisenhower Parkway
P.O. Box 1346 Ann Arbor, Ml 48106-1346

ABSTRACT
This thesis presents the localization and characterization of the
recently discovered nitric oxide (NO)-synthesizing nerves in the
adrenal glands of rats, and their changes in various conditions which
cause alteration in adrenal nerve and/or glandular activities. The
study was conducted mainly by qualitative analysis of the NO-
synthesizing enzyme, nitric oxide synthase (NOS) immunohistochemically
and/or by the use of reduced nicotinamide adenine dinucleotide
phosphate (NADPH)-diaphorase histochemistry which also marks the site
of neuronal NOS immunoreactivity. In parts of the study, where
quantification of the levels of NOS was required, computer assisted
image analysis and biochemical assay for measuring the activity of NOS
were also used.
In the adult rat adrenal glands NOS immunoreactivity occurred in a population
of nerve cell bodies and fibres, where NADPH-diaphorase staining also colocalized. In
the adrenal cortical cells NADPH-diaphorase staining alone was observed without
NOS immunoreactivity. As found from surgical extrinsic and intrinsic adrenal
denervation studies, the NOS-immunoreactive nerve fibres in the gland have both
extrinsic and intrinsic origins. The extrinsic nitrergic nerve fibres innervate adrenal
chromaffin and ganglion cells. The intrinsic nitrergic nerve fibres which arise from the
local neurons innervate adrenal blood vessels and the zona glomerulosa of cortex.
Colocalization studies revealed that a population of NADPH-diaphorase stained NO-
synthesizing adrenal intrinsic neuronal cell bodies contained vasoactive intestinal
peptide and neuropeptide Y, but not calcitonin gene-related peptide, substance P,
tyrosine hydroxylase or calretinin.
Developmental studies showed that NOS-immunoreactive nerve fibres were

present in adrenal glands of rats at all ages examined from the 16^ ̂day of gestation up
to 2 years of age. A considerable degree of variation in the distribution of the
immunoreactive nerve fibres in both medulla and the zona glomerulosa of cortex was
observed at different ages. The NOS-containing neuronal cell bodies within the
adrenal gland were found from the 2 0 ^ ̂ day of gestation onwards, and increased in
their number to reach to that of adult levels by the 4^ ̂ day after birth. In the glands
from the aging rats their number was increased by 28.6 % above the adult levels . NOS
immunoreactivity in the chromaffin cells was observed only in the glands of the aging
rats.
Guanethidine, 6 -hydroxydopamine or capsaicin treatments did not
cause any change in the adrenal NOS immunoreactivity and NADPH-
diaphorase staining. In contrast, reserpine treatment for 7 days
caused a significant decrease in the NOS immunoreactivity in the nerve
fibres that innervate the chromaffin and ganglion cells.
Hypophysectomy did not cause any change in the NOS immunoreactivity,
although it eliminated most of the NADPH-diaphorase staining adrenal
cortical cells. Streptozotocin-induced diabetes of 8 weeks duration
caused an increase in the NOS-immunoreactive nerve fibres and induced
NOS immunoreactivity in a small number of adrenal cortical cells. It
also caused an increase in the intensity of NADPH-diaphorase staining
in the adrenal cortical cells. A significant degree of prevention of
such diabetic-induced increase in both NOS and NADPH-diaphorase was
found with ganglioside treatment. At a later stage (12 weeks of
diabetes duration) the effect of streptozotocin-induced diabetes was a
decrease in the NOS-immunoreactive nerve fibres but an increase in the
occurrence of NOS-immunoreactive adrenal cortical cells. The intensity
of NADPH-diaphorase staining in the cortical cells was still increased.

CONTENTS
ABSTRACT......................................................................................................................2
CONTENTS......................................................................................................................4
ACKNOWLEDGEMENTS...........................................................................................9
LIST OF TABLES AND FIGURES............................................................................ 10
PUBLICATIONS ARISING FROM THE WORK PRESENTED
IN THIS THESIS............................................................................................................ 12
LIST OF ABBREVIATIONS....................................................................................... 13
PREFACE........................................................................................................................15
SECTION 1: INTRODUCTION.................................................................................. 19
1:1. Historical background to adrenal gland research.............................................. 20
1:2. Morphology of adrenal gland.................................................................................21
1:3. Development and aging of adrenal medulla.........................................................24
1:4. Development and aging of adrenal cortex............................................................25
1:5. Biosynthesis and secretion of adrenal gland hormones................................... 26
1:5:1. Catecholamines........................................................................................... 26
1:5:2. Steroids........................................................................................................ 28
1:6. Innervation of adrenal gland................................................................................ 29
1:6:1. Extrinsic adrenal innervation.................................................................... 29
1:6:2. Intrinsic adrenal innervation..................................................................... 32
1:6:3. Course and distribution o f nerve fibres and
terminals within the adrenal cortex ..........................................................32
1:6:4. Course and distribution o f nerve fibres and
terminals within the adrenal medulla .......................................................32

1:7. Neuromediators in adrenal gland..........................................................................33
1:7:1. Acetylcholine................................................................................................33
1:7:2. Biogenic amines...........................................................................................34
1:7:3. Adenosine 5'-triphosphate.........................................................................35
1:7:4. Neuropeptides..............................................................................................36
1:7:4:1. Enkephalins..................................................................................36
1:7:4:2. Vasoactive intestinal peptide ......................................................37
1:7:4:3. Somatostatin................................................................................39
1:7:4:4. Neurotensin.................................................................................. 39
1:7:4:5. Calcitonin gene-relatedpeptide ............................................... 40
1:7:4:6. Neuropeptide Y ............................................................................41
1:7:4:7. Substance P ..................................................................................42
1:7:4:8. Bombesin...................................................................................... 43
1:7:4:9. Galanin........................................................................................ 44
1:8. Nitric oxide................................................................................................................45
1:8:1. Biosynthesis o f nitric oxide: nitric oxide synthase ................................... 47
1:8:2. Functional significance o f nitric oxide ......................................................49
1:8:2:1. Nitric oxide in the cardiovascular system ................................. 50
1:8:2:2. Nitric oxide in immunity and inflammation..............................51
1:8:2:3. Nitric oxide in neural transmission............................................51
1:9. Plasticity in adrenal gland innervation and expression of
neuromediators........................................................................................................ 53
1:9:1. During development and aging .................................................................. 53
1:9:2. During stress ................................................................................................55
1:9:3. After selective surgical denervations.........................................................56
1:9:3:1. Extrinsic adrenal denervation............................................ 56
1:9:3:2. Intrinsic adrenal denervation.....................................................57
1:9:3:3. Hypophysectomy..........................................................................57

1:9:4 During chronic exposure to drugs ..............................................................58
1:9:4:1. Reserpine..................................................................................... 58
1:9:4:2. 6-hydroxydopamine................................................................... 58
1:9:4:3. Guanethidine................................................................................59
1:9:4:4. Capsaicin..................................................................................... 59
1:9:5. In diabetes mellitus..................................................................................... 59
SECTION 2: MATERIALS AND METHODS...........................................................62
2:1. Animals used in this study...................................................................................... 63
2:2. Maintenance of the animals.................................................................................... 63
2:3. Tissue processing...................................................................................................... 63
2:3:1. Preparation o f adrenal sections................................................................. 63
2:3:2. Immunohistochemistry................................................................................64
2:3:3. NADPH-diaphorase histochemistry.........................................................66
2:4. Microscopy and computer assisted image analysis............................................. 67
2:5. Biochemical assay of nitric oxide synthase activity............................................ 68
2:6. Surgical procedures.................................................................................................70
2:6:1. Extrinsic adrenal denervation.................................................................... 70
2:6:2. Intrinsic adrenal denervation..................................................................... 71
2:6:3. Hypophysectomy......................................................................................... 72
2:7. Drug treatment......................................................................................................... 73
2:7:1. Reserpine.....................................................................................................73
2:7:2. Guanethidine and 6-hydroxydopamine.................................................... 73
2:7:3. Capsaicin..................................................................................................... 74
2:8. Induction of diabetes................................................................................................75
2:9. Source of Antibodies, Chemicals and Reagents.................................................. 77

SECTION 3: RESULTS..................................................................................................80
3:1. Distribution and characteristics of nitric oxide-synthesizing
nerves in the rat adrenal gland..............................................................................81
3:1:1. Colocalization o f nitric oxide synthase and
NADPH-diaphorase in rat adrenal g land .............................................. 82
3:1:2. Distribution and colocalization o f nitric oxide synthase
and NADPH-diaphorase in adrenal gland o f developing, adult
and aging Sprague-Dawley ra ts ..............................................................93
3:1:3. The intra-adrenal distribution o f intrinsic
and extrinsic nitrergic nerve fibres in the r a t ........................................110
3:1:4. Colocalization o f neuropeptides and NADPH-diaphorase
in the intra-adrenal neuronal cell bodies and
fibres o f the r a t .........................................................................................121
3:1:5. Calretinin immunoreactivity in adrenal gland o f
developing, adult and aging Sprague-Dawley ra ts ............................. 133
3:2. Changes in the expression of nitric oxide synthase and
NADPH-diaphorase in the adrenal gland of rats Iffdevelopment,-
aging and altered adrenal gland acti vity .̂....................................................... 146
3:2:1. Effect o f reserpine treatment and hypophysectomy on
the nitric oxide synthase immunoreactivity and
NADPH-diaphorase staining in the rat adrenal g land ........................ 147
3:2:2. Increase in nitric oxide synthase and NADPH-diaphorase
in the adrenal gland o f streptozotocin-diabetic Wistar
rats and its prevention by ganglioside....................................................157
SECTION 4: GENERAL DISCUSSION....................................................................181
4:1. Adrenal nitric oxide-synthesizing nerves........................................................ 182
4:2. Nitric oxide in the regulation of adrenal medullary secretion........................184
7

4:3. Nitric oxide in the regulation of adrenal cortical secretion............................188
4:4. Nitric oxide in the regulation of adrenal blood flow ....................................... 190
4:5. Plasticity in nitrergic innervation........................................................................193
REFERENCES............................................................................................................. 198

ACKNOWLEDGEMENTS
I am extremely grateful to my supervisor Professor Geoffrey Bumstock for giving me the opportunity to pursue my education under his supervision by arranging the cost of the study. I thank him for his enthusiastic and invaluable guidance of the research work and his helpful constructive comments during the preparation of the manuscript.
I thank Dr Annette Tomlinson for the helpful discussions throughout the course of the study and the constructive comments in the preparation of the manuscript. I would also like to thank Dr Abebech Belai for teaching me the technique of immunohistochemistry, and the collaborative work in the injections of the animals for the studies reported in Section 3:2:2 and the helpful discussions and encouragements throughout the course of this study.
My sincere gratitude goes to my other collaborators Dr Vera Ralevic who injected the animals for the studies reported in Section 3:1:3 and Dr Jill Lincoln who performed the biochemical assay presented in Section 3:2:2.1 thank Jane Pendjiky and Christopher Sym for their help in the photographic work. I would also like to thank Dr Peter Abrahams, Dr Christopher Dean and the late Professor John Pegington for their initiation of my study in this department and their encouragements; Dr Barbara Pittam for administrative support; Annie Evans for her encouragements and support; Tim Robson for keeping up with the material supplies; Dr John Rogers and Dr Ulrich Forstermann for providing the calretinin and some of the NOS antisera, respectively; all members of the Bumstock's research group and everybody whose contribution was helpful in the course of the study.
This study was supported in part by the grants from Overseas Research Students award, University College London Dean's Scholarship and Department of Anatomy and Developmental Biology's Research Studentship.

LIST OF FIGURES AND TABLES
Pages
Section 3:1:1.
Fig. 1 .......................................................... 8 8
Fig. 2 ............................................................................................ 89
Section 3:1:2.
Fig. 3 .......................................................................................... 101
Fig. 4 .......................................................................................... 102
Fig. 5 .......................................................................................... 103
Fig. 6 .......................................................................................... 104
Table 1 .......................................................................................105
Section 3:1:3
Fig. 7 .......................................................................................... 116
Fig. 8 .......................................................................................... 117
Fig. 9 .......................................................................................... 118
Section 3:1:4
Fig. 1 0 ........................................................................................ 127
Fig. 1 1 ........................................................................................ 128
Fig. 1 2 ........................................................................................ 129
Section 3:1:5
Fig. 1 3 ........................................................................................ 140
Fig. 1 4 ........................................................................................ 141
Fig. 1 5 ........................................................................................ 142
Section 3:2:1
Fig. 1 6 ........................................................................................ 152
Fig. 1 7 ........................................................................................ 153
Table 2 .......................................................................................154
10

Section 3:2:2
Fig. 1 8 ...........................................................................................168
Fig. 1 9 .......................................................................................... 169
Fig. 2 0 .......................................................................................... 170
Fig. 2 1 .......................................................................................... 171
Fig. 2 2 .......................................................................................... 172
Table 3 ......................................................................................... 173
Table 4 ......................................................................................... 174
Table 5 ......................................................................................... 175
11

PUBLICATIONS ARISING FROM THE WORK PRESENTED IN THIS
THESIS
Afework, M., Tomlinson, A., Belai, A. and Bumstock, G. (1992) Colocalization of nitric
oxide synthase and NADPH-diaphorase in rat adrenal gland. NeuroReport 3, 893-
896.
Afework, M., Tomlinson, A. and Bumstock, G. (1994) Distribution and colocalization
of nitric oxide synthase and NADPH-diaphorase in adrenal gland of developing,
adult and aging Sprague-Dawley rats. Cell Tissue Res. 276,133-141.
Afework, M., Ralevic, V. and Bumstock, G. (1995) The intra-adrenal distribution of
intrinsic and extrinsic nitrergic nerve fibres in the rat. Neurosci Lett 190,109-112.
Afework, M. and Bumstock, G. (1995) Colocalization of neuropeptides and NADPH-
diaphorase in the intra-adrenal neuronal cell bodies and fibres of the rat. Cell Tissue
Res in press.
Afework, M. and Bumstock, G. (1995) Calretinin immunoreactivity in adrenal glands
of developing, adult and aging Sprague-Dawley rats. Int J Dev Neurosci in press.
Afework, M. and Bumstock, G. (1995) Effect of reserpine treatment and
hypophysectomy on the nitric oxide synthase immunoreactivity and NADPH-
diaphorase staining in the rat adrenal gland. Manuscript.
Afework, M., Lincoln, J., Belai, A. and Bumstock, G. (1995) Increase in nitric oxide
synthase and NADPH-diaphorase in the adrenal gland of streptozotocin-diabetic
Wistar rats and its prevention by ganglioside. Manuscript.
12

LIST OF ABBREVIATIONS
A cells .................... adrenaline-storing and secreting cells
A ChE......................acetylcholinesterase
ACTH .....................adrenocorticotrophic hormone
A T P ........................ adenosine 5 -triphosphate
BH4 ........................ tctrahydrobiopterine
cA M P..................... cyclic adenosine monophosphate
cGM P..................... cyclic guanosine monophosphate
CGRP...................... calcitonin gene-related peptide
ChAT......................choline acetvl transferasecp m ........................ count per minuteD BH ....................... dopamine p-nyaroxyiase
DOPA....................dihydroxyphenylalanine
dpm .........................disintegration per minute
E ..............................embryonic day
ED RF..................... endothelium-derived relaxing factor
FA D .......................flavin adenine dinucleotide
FG F........................fibroblast growth factor
F ig ...........................figure
Figs........................ figures
FM N...................... flavin mononucleotide
G TP....................... guanosine triphosphate
h ............................. hour
..........................tritiated hydrogen
IDDM ................... insulin-dependent diabetes mellitus
EPS ....................... lipopolysaccharides
13

L T P .......................long term potentiation
m in ........................minute
N A ........................ noradrenaline-storing and secreting cells
NADPH................reduced nicotinamide adenine dinucleotide phosphate
L-N A M E.............N^-nitro-L-arginine methyl ester
N G F......................nerve growth factor
NIDDM ................non-insulin-dependent diabetes mellitus
L-N M M A........... N^-monomethyl-L-arginine
L -N N A ................N^-nitro-L-arginine
N O ....................... nitric oxide
N O S..................... nitric oxide synthase
N PY ..................... neuropeptide Y
6 -O H D A .............. 6 -hydroxydopamine
PB S...................... phosphate-buffered saline
PG P...................... protein gene product
PNM T.................. phenylethanolamine-N-methyltransferase
SGC cells .............small granule containing cells
S P ........................ substance P
T H ........................ tyrosine hydroxylase
V IP .......................vasoactive intestinal peptide
14

PREFACE
15

The paired adrenals are small pyramid-shaped organs located at the superior
pole of the kidneys. Each adrenal is composed of an inner part, the adrenal medulla and
an outer part, the adrenal cortex that is surrounded by a capsule. The general function
of the adrenal gland is to enable the individual to cope with stress conditions. The
catecholamine secretion by the adrenal medulla has a widespread effect in the body
system in mobilizing glucose and fatty acids for energy and preparing the heart, lungs,
and muscles for action during acute stress and injury. During chronic stress, the steroid
hormones of the cortex protect against overreaction from the body's responses to stress.
They are also important in immune defence and inflammation. In addition, particularly
during stress conditions due to prolonged deprivation of food and fluid, the adrenal
steroids stimulate gluconeogenesis to maintain the supply of glucose and increase
sodium reabsorption to maintain body fluid content.
Classically, secretion of catecholamines from the adrenal medulla was
considered to be under the sole control of the cholinergic preganglionic sympathetic
innervation (Elliott 1913; Hoshi 1926; Feldberg, Minz and Tisudzimura 1934). It was
also thought that humoral factors were exclusively responsible for the control of
adrenal cortical function (Hechter 1949; Hayenes, Savard and Dorfman 1952).
However, recent discoveries on the occurrence of a growing number of neuroactive
substances, in both adrenal medullary nerves and chromaffin cells (Kondo 1985;
Pelto-Huikko 1989), have suggested that several neuromediators are involved in
regulating the process of synthesis and secretion of various neuroactive substances by
the adrenal chromaffin cells. In the adrenal cortex, morphological studies provided
evidence for the presence of nerve fibres and endings containing various mediators (see
Charlton 1990). Functional studies have shown the involvement of these nerves in the
regulation of adrenal cortical activities (see Vinson, Hinson and Toth 1994). Secretion
from the adrenal gland is also found to be influenced by adrenal blood flow (Vinson
and Hinson 1992).
The major neuroactive substances found in the adrenal neuronal components
16

and/or chromaffin cells include acetylcholine, biogenic amines, adenosine triphosphate
(ATP) and neuropeptides. In addition, a substance with possible functional role(s),
whose synthesis in the adrenal gland was recently recognized is nitric oxide (NO). It is
also a recent discovery that NO which is a noxious unstable gas has been found to be
secreted by mammalian cells with widespread functions in the cardiovascular system,
immune system, inflammation and neurotransmission. Furthermore, changes in its
levels of production has been associated with various pathophysiological
complications.
NO is synthesized by a family of enzymes called nitric oxide synthases which
catalyze the conversion of L-arginine and molecular oxygen to NO and L-citrulline
(see Forstermann, Schmidt, Pollock, Sheng, Mitchell, Warner, Nakane and Murad
1991). Immunohistochemically, nitric oxide synthase (NOS) has been localized in a
wide range of cells that synthesize NO, and has been colocalized with the
histochemical staining for reduced nicotinamide adenine dinuceleotide phosphate
(NADPH)-diaphorase. In the adrenal gland, the presence of NOS was originally
reported from a biochemical study which demonstrated the activity of the enzyme in
rat and bovine adrenal glands (Palacios, Knowles, Palmer and Moncada 1989).
Recently, immunohistochemical and enzyme histochemical studies have shown its
occurrence in rat adrenal medullary nerves (Bredt, Hwang and Snyder 1990; Dawson,
Bredt, Fotuchi, Hwang and Snyder 1991). These studies, however, were brief and apart
from the demonstration of the existence of NOS in the gland, they did not characterize
the nature and extent of its distribution. The aim of the study presented in this thesis
was therefore to fully investigate and characterize cellular elements containing the
neuronal NOS isoform in the adrenal glands of rats in health and during various
conditions causing altered adrenal nerve and/or glandular activities.
In this thesis, a literature survey on the historical background and current
knowledge of the adrenal gland and NO is given in the Introduction (Section 1). The
17

Materials and Methods section (Section 2) describes the general background of the
techniques and the protocols employed in the present study. These include
immunohistochemistry, NADPH-diaphorase histochemistry and an enzyme
biochemical method for assay of NOS activity. The various experimental approaches
employed in the present study to cause alteration in the adrenal innervation and/or
glandular secretions are also described in this section.
The results are presented in two main parts. The first part deals with studies on
the demonstration and characterization of the NO-synthesizing nerves in the adrenal
glands of rats. These include: a detailed account of the distribution of NOS
immunoreactivity and its colocalization with NADPH-diaphorase staining in the adult
rat adrenal gland (Section 3:1:1); an account of the changes in the distribution of NOS
that occur during development and aging (Section 3:1:2); intra-adrenal distribution
and characterization of the extrinsic and intrinsic NOS-containing nerve fibres as
determined by surgical denervations of extrinsic and intrinsic adrenal nerves, and by
chemical sympathectomy and sensory denervation (Section 3:1:3); studies on
colocalization of NOS with a number of neuropeptides and the rate limiting enzyme in
the catecholamine synthesis, tyrosine hydroxylase (TH) (Section 3:1:4) and with the
recently discovered calcium binding protein, calretinin Section 3:1:5. As this study is
the first to describe the distribution of calretinin in the adrenal gland, a detailed account
of its occurrence and changes in development and aging has also been examined in
Section 3:1:5.
In part II of the Results section, changes occurring in the adrenal NOS and
NADPH-diaphorase following alteration of the state of the adrenal innervation and/or
glandular secretions by reserpine treatment and hypophysectomy (Section 3:2:1) and
by streptozotocin-induced diabetes (Section 3:2:2) are examined.
The separate experimental work is discussed individually in each study. In
addition, a discussion of broader issues is given in the General Discussion section
(Section 4).
18

SECTION 1
INTRODUCTION
19

1:1. Historical background to adrenal gland research
In 1563 Bartholomeus Eustachius gave the first description of the adrenal glands
under the title "glandulae Renibus Incumbentes". The anatomy and functions of the
adrenals, however, remained a puzzle for nearly three centuries while they were
described with various names and supposed functions. Casper Bartholinus (1611) gave
an account of the adrenals and illustrated them in a woodcut as hollow capsule-like
organs filled with black bile. This led to the belief that the adrenal glands have ducts
which connect them to the kidneys and/or the gonads (see Rolleston 1936).
Later, Cuvier (1805) discovered the adrenal glands as solid organs, each formed
of two distinct tissues, rather than as previously thought of, a capsule with a cavity.
The anatomy of the gland was correctly identified as having a smaller central part, the
adrenal medulla, and a larger outer part, the adrenal cortex, surrounded by a fibrous
connective tissue capsule.
In 1855, Thomas Addison described disease of the adrenal glands and pointed
out their indispensability for life. This initiated a functional approach towards the study
of the adrenal glands and the following year, Brown-Sequard (1856) verified their
importance with the demonstration that adrenalectomy in various animals would result
in death.
Henlé (1865) found brown colouration of the adrenal medullary tissue upon
treatment with chromate salts and introduced the term chromaffin reaction.
Subsequently, a pressor substance was demonstrated to be contained in adrenal
medullary extract (Oliver and Schafer 1894) and was implicated for the histological
reactivity of the tissue with chromate salts (Moore 1895). The search for the
identification of this pressor substance in the medullary extract was carried out and in
1901 Takamine and Aldrich, working independently, isolated a crystalline substance
and named it adrenaline and epinephrine, respectively. It was, however, later in the mid
20

1900s that a second pressor substance found in the adrenal medulla was identified and
termed noradrenaline or norepinephrine (Holtz, Credener and Kroneberg 1947;
Bergstrorm, von Euler and Hamberg 1949; von Euler and Hamberg 1949).
The observations that removal of the interrenal body, in some fishes, was fatal
indicated that the indispensability of the adrenal glands for life is related to the cortex
but not the medulla (Biedle 1913). The essential substance found in the adrenal cortex
was then identified to be lipid extracts which maintained adrenalectomized cats
indefinitely (Hartman and Brownell 1930; Swingle and Pfiffner 1930). Later,
numerous steroids were isolated from the cortical extracts and the adrenal cortex was
discovered to have three main groups of steroids each of which consisted of multiple
corticosteroids (Pfiffiier 1942; Mason 1964; Gaunt 1975). Mineralocorticoids, mainly
secreted by cells of the zona glomerulosa are one of such steroids (Sarason 1943;
Deane and Creep 1946; Deane, Shaw and Creep 1948; Creep and Deane 1949).
Aldosterone is the primary mineralocorticoid. Glucocorticoids, secreted by cells of
zona fasciculata and zona reticularis are the second group of adrenal corticosteroids
(Sarason 1943; Deane and Creep 1946; Deane et al. 1948). In humans and most
mammals the primary glucocorticoid is cortisol, while in some rodents it is
corticosterone (Bush 1953, Sandor, Fazekas and Robinson 1976). The third group of
corticosteroids secreted from adrenal cortex are the adrenal androgens. They are
believed to be secreted from the zona reticularis of the adrenal cortex. Testosterone is
the physiologically potent androgen (Bondy 1985).
1:2. Morphology of adrenal gland
It is only in mammals that the adrenal cortex and medulla are found intimately
associated. In the other vertebrate classes, the two tissues may be completely separate
or intermingled to a varying degree (Bourne 1961; Coupland 1965). When separate, the
homologue of the adrenal medulla is simply considered as part of the chromaffin
21

tissue, while that of the adrenal cortex is called interrenal tissue.
The adrenal cortex forms the higher proportion of the organ. In rat, the cortex
accounts for about 90 % of the weight, whereas the adrenal medulla forms the
remaining 10 %. However, calculated medullary weight and volume have been shown
to vary by as much as 30 % with different methods of fixation and embedding
(Tomlinson, Durbin and Coupland 1987). The parenchyma of the adrenal cortex is
formed from steroid synthesizing and secreting epithelial cells, arranged in cords or
cylinders with varying architecture to form the three adrenal cortical zones. These are
an outer zona glomerulosa, an intermediate zona fasciculata and an inner zona
reticularis. In many mammalian species, cells of the juxtamedullary zone of the cortex
are sometimes found interdigitated with the medullary chromaffin cells and in
histological sections may appear as islands of cortical cells dispersed in the medulla
(Coupland 1965).
The adrenal medulla is mainly composed of the chromaffin cells arranged in an
anastomosing network in close relation to capillaries. It represents the largest aggregate
of the chromaffin tissue, which is embryologically derived from the neuroectoderm,
and is usually innervated by the preganglionic sympathetic nerve fibres and contains a
high concentration of catecholamines which usually results in the chromaffin reaction
(Coupland 1989). The chromaffin cells contain a large number of vesicles, which are
called chromaffin vesicles or granules. Schwann cells, vascular and connective tissue
elements, and species variable numbers of ganglion cells and islets or cords of cortical
cells are additional cellular components found in the adrenal medulla (Coupland 1965).
In most mammals, there are two types of principal adrenal medullary secretory
cells: the adrenaline-storing and secreting (A) cells and the noradrenaline-storing and
secreting (NA) cells (Eranko 1952; Hillarp and Hokfelt 1953; Coupland 1965). In
adrenals fixed with glutaraldehyde and postfixed with osmium tetroxide, the A cells
contain vesicles filled with an evenly distributed medium electron dense granules,
while the NA cells contain electron dense granules which in rodents are usually located
22

asymmetrically within the vesicle (Coupland 1965). The relative proportions of A to NA
cells vary with species and age of the animal. In most postnatal mammals A cells
predominate, although a variable but smaller amount of NA cells are also always
present (Coupland 1984).
Another group of chromaffin cells called the small granule-containing (SGC)
cells are also found in the adrenal medulla of many animal species (Unsicker 1973a;
Gorgas and Bock 1976; Coupland, Kobayashi and Tomlinson 1977; Unsicker, Kabura-
Fliih and Zwarg 1978). The SGC cells contain membrane bound granules of variably
moderate to low electron density, with few high electron dense cores. They are of the
same size as typical chromaffin cells but have smaller vesicles, a higher nucleus to
cytoplasm ratio and a great capacity to form processes. In rat, they account for not
more than 1 % of the chromaffin cell's population (Tomlinson and Coupland 1990).
The SGC cells are considered to be cells of chromaffin cell lineage, morphologically
intermediate between typical chromaffin cells and neurons (Coupland 1984).
The blood supply to the adrenal gland is by multiple small arteries which arise
from the abdominal aorta or one of its major branches between the point where it enters
abdomen and the renal arteries (Lever 1952; Coupland 1975; Coupland and Selby
1976). Several small arteries approach the adrenals where they divide and redivide to
form a capsular plexus. Many cortical arteries arise from this plexus, penetrate the
capsule and form another plexus in the subcapsular region and zona glomerulosa, from
which several capillaries branch and run through the zonae fasciculata and reticularis.
These capillaries join and rejoin progressively to form larger channels in the medulla
which eventually open into the central vein. Additionally, there are a few cortical
arteries which directly traverse the capsule and outer region of the cortex to join the
capillary plexus in the zona fasciculata, or take a looped course back towards the
surface before joining the cortical capillary plexus (Flint 1900; Lever 1952).
Although there are some investigators who favour the existence of a vascular
23

corticomedullary portal circulation (Pohrecky and Wurtman 1971; Henderon and Daniel
1978), morphological studies did not reveal any intra-adrenal portal system (Flint
1900; Bennett and Kilham 1940; Gersh and Grollman 1941, Dempster 1974; Coupland
1975; Coupland and Selby 1976; Sparrow and Coupland 1987; Murakami, Oukouchi,
Uno, Ohtsuka and Taguchi 1989b). Instead, another group of arteries, penetrate the
capsule and pass straight through the cortex without branching and reach to the
medulla. Here they form a medullary capillary plexus which ultimately joins the
peripheral radicles of the central vein. The central vein also receives other minor veins
of periadrenal origin to form the single adrenal vein, which drains into the posterior
vena cava, the left renal vein or one of its branches, or into the adrenolumbar vein
(Johnstone 1957; Coupland 1965).
1:3. Development and aging of adrenal medulla
Adrenal chromaffin cells develop from the neural crest cells (Le Douarin 1980).
This was demonstrated by transplanting the neural crest and tube from quail embryo
into the corresponding regions of chick embryo and tracing the fate of the cells with
the help of their distinct nucleolus. It was found that the migratory behaviour of the
neural crest cells depended on the pathway available while the phenotype of the cells
was determined by environmental factors. Glucocorticoids secreted from the adrenal
cortex, favour differentiation of the sympathoadrenal precursors of the neural crest
cells migrating to the adrenal gland into chromaffin cells. On the other hand, fibroblast
growth factor (FGF) and nerve growth factor (NGF) determine the neural phenotype in
the neural crest cells migrating to other regions (Le Douarin 1980; Anderson 1993).
Early in development, the migrating adrenal medullary precursor cells invade
the dorsomedial aspect of the adrenal cortical anlage and pass through three
developmental stages (Coupland 1952). The first stage is one of "primitive
sympathetic" cells with a round basophilic nucleus surrounded by a thin rim of
cytoplasm. The second stage comprises cells called "parasympathetic" or
24

phaeochromoblasts of elongated cells with an elongated nucleus. The third stage is the
chromaffin cells or phaeochromocytes, which are round to polyhedral cells displaying
the chromaffin reaction and have a rounded nucleus. Such later cell types showing the
chromaffin reaction may be found as early as 12-15 weeks in the human foetus, 20
days in foetal rabbit and 12 days in foetal rat (Coupland 1980).
During aging the adrenal medulla shows proliferative lesions (Fleischmann
1965; Haley 1976; Hollander and Snell 1976; Cheng 1980). In the rat, lesions occur as
nodular proliferations on a diffuse hyperplastic background (Tischeler, DeLellis,
Perlman, Allen, Costopoulos, Lee, Nunnemacher, Wolfe and Bloom 1985). Such
lesions are particularly observed after 1 year (Gillman, Gilbert and Spence 1953), and
their occurrence is influenced by several factors such as genetic, humoral, nourishment
and gender (Cheng 1980). In a lesioned adrenal medulla, there is occurrence of both
adrenaline and noradrenaline granules within the same chromaffin cell (Chalfe and
Perlman 1976). Moreover, the appearance of synaptic type vesicles with electron dense
contents in a number of NA cells of medulla of the aging rat suggested the
differentiation of adrenal chromaffin cells towards the neural phenotype in aging
(Coupland and Tomlinson 1989).
1:4. Development and aging of adrenal cortex
Adrenocortical primordia organize early in foetal life from the local mesoderm.
The process of growth, differentiation and cell renewal of the adrenal cortex has been a
matter of controversy. There were two principal hypotheses: the cell migration
hypothesis originally proposed by Gottschau (1883), and the zonal hypothesis
proposed by Chester Jones (1948) where each zone is thought to replenish its cells
independently. However, the observations from the regenerating capacity of the
adrenocortical cells in the enucleated adrenals (Ingle and Higgins 1938; Greep and
Deane 1949) supported the cell migration hypothesis. This was further proved with
25

studies from injection of radiolabelled thymidine which showed a continual centripetal
migration of subcapsular cells (Bertholet 1980; Zajicek, Ariel and Arber 1986). During
the migration, the cells are transformed progressively into the different cell types of the
zonae glomerulosa, fasciculata and reticularis. The migration ends in the zona
reticularis where the cells ultimately degenerate and die.
In aging rat, there is hypertrophy of the zonae fasciculata and reticularis
(Rebuffat, Belloni, Rocco, Andreis, Neri, Malendowicz, Gottardo, Mazzocchi and
Nussdorfer 1992). There is also an increase in the occurrence of nodular hyperplasia
(Attia 1985). In addition, an increase in the levels of lipid droplets contained in cells of
all the three zones and lipofÆin contained in cells of the zonae rticularis and
fasciculata is documented (Carsia and Malamed 1989; Belloni, Rebuffat,
Malendowicz, Mazzochi, Rocco and Nussdorfer 1992; Rebuffat et aL 1992).
1:5. Biosynthesis and secretion of adrenal gland hormones
1:5:1. Catecholamines
Adrenaline is synthesised from tyrosine in four enzyme catalyzed sequential
steps of dihydroxyphenylalanine (DOPA), dopamine, noradrenaline and adrenaline
(Blaschko 1939; Demis, Blaschko and Welch 1956; Masuoko, Schott, Akawie and
Clark 1956; Undenfriend and Wyngaarden 1956; Goodall and Kirshner 1957; Kirshner
1975). The conversion of tyrosine into DOPA is catalyzed by tyrosine hydroxylase
(TH). This is the rate limiting step in the pathway. DOPA is transformed into dopamine
by the enzyme DOPA decarboxylase. Both TH and DOPA decarboxylase have been
shown to C c t f j r in the cytosol, and therefore, the formations of DOPA and
dopamine take place in the cell cytoplasm. Dopamine is taken into the granules and
converted into noradrenaline by the enzyme dopamine 6̂ -hydroxylase (DBH) only
found within the chromaffin granules. In A cells noradrenaline leaks out of the
chromaffin granules and is converted into adrenaline in the cell cytoplasm by the
enzyme phenylethanolamine-N-methyltransferase (PNMT). From the cell cytoplasm,
26

adrenaline is taken up and stored in the chromaffin granules. The chromaffin vesicles
also contain nucleotides, chromogranins, lipids, glycoproteins, calcium,
mucopolysaccharides, ascorbic acid, and a number of neuroactive peptides (Winkler
1976, Winkler, Apps and Fischer-Colbrie 1986).
The secretion of catecholamines from the adrenal chromaffin cells is effected
mainly by acetylcholine released from the sympathetic preganglionic nerves.
Acetylcholine is believed to initiate opening of the calcium channels located on the
plasma membrane and brings an inward movement of calcium from extracellular fluid
to raise intracellular calcium levels which leads to the movement of chromaffin
granules towards the plasma membrane for subsequent fusion and exocytosis (Baker
and Knight 1978,1981; Knight and Baker 1982; Burgoyne 1991).
Acetylcholine receptors on the chromaffin cells are both nicotinic and
muscarinic (Feldberg et aL 1934). There is, however, species variation in the relative
distribution of this two subtypes of cholinergic receptors (Malmejac 1964; Rubin and
Miele 1968; Liang and Perlman 1979; Knight and Baker 1986). In the cat, selective
muscarinic agonists have been shown to release principally adrenaline, while nicotinic
agonists cause release of mainly noradrenaline (Douglas and Poisner 1965; Rubin and
Miele 1968). However, the absence of such preferential secretion with activation of
either of the receptor in many other species has led to the conclusion that dual control
may be peculiar to the cat (Unger and Phillips 1983). Both nicotinic and muscarinic
acetylcholine receptors contribute to activation of secretion of catecholamines
(Kirpekar, Prat and Shiavone 1982; Role and Perlman 1983; Wakade and Wakade
1983). The extent to which each receptor is involved depends on the concentration of
acetylcholine and the animal species, and it appears that it is mainly the nicotinic
receptor that is engaged in active secretion of catecholamines. The muscarinic
receptors are activated by a relatively low concentration of acetylcholine to cause an
efflux of calcium from the cell and an increase in intracellular cyclic guanosine
27

monophosphate (cGMP) levels that is inhibitory to nicotine-mediated release of
catecholamines (Schneider, Cline and Lemaire 1979; Derome, Tseng, Mercier,
Lemaire and Lemaire 1981). At relatively higher concentration of acetylcholine the
inhibition of release is overcome by activation of the nicotinic receptors. In addition to
acetylcholine, several other neuromediators (see section 1:7) and glucocorticoids have
also been shown to influence the biosynthesis and secretion of adrenal medullary
hormones.
1:5:2. Steroids
The starting material for the synthesis of adrenal steroid hormones is cholesterol
(Hechter, Solomon, Zaffaroni and Pincus 1953; Werbin and Chaikoff 1961; Krum,
Morris and Bennett 1964). Adrenal cortical cells synthesize cholesterol, or obtain it
from blood plasma, and mainly store it as cholesterol ester in the lipid droplets. When
required, the synthesis of steroid hormones occurs by a series of reactions involving
oxidation-reduction and hydroxylation, which are catalyzed respectively, by a
hydroxysteroid dehydrogenase system and a series of mixed function oxidases
(hydroxylases), utilising O2 , NADPH, adrenodoxin and cytochrome P-450 (Hyano,
Saba, Dorfman and Hechter 1956; Gower 1975; Samuels and Nelson 1975; Fraser
1992). Since the enzymes are located either in the mitochondria or the endoplasmic
reticulum, the metabolic intermediates move back and forth between the two organelles
during the reaction process. Cholesterol is released from the lipid droplets by the action
of cholesterol ester hydrolase and passes into the mitochondria, where it is converted
into a common intermediate precursor called pregnenolone. The pregnenolone passes
out of the mitochondria and enters into the endoplasmic reticulum and is converted into
androstenedione, 11-deoxycortisol and 11-deoxycorticosterone. Androstenedione is
further converted into the androgens, while 11-deoxycortisol and 11-
dexycorticosterone are transferred back into the mitochondria and yield cortisol and
corticosterone, respectively, as well as aldosterone.
28

Adrenocorticotrophic hormone (ACTH) regulates the synthesis of cortisol and
corticosterone (Hechter 1949; Haynes et al. 1952; Saffran, Grad and Bayliss 1952).
Binding of ACTH into receptors on adrenocortical cells activate adenylate cyclase
which increases intracellular levels of cyclic adenosine monophosphate (cAMP)
(Schuster 1974). The elevated levels of cAMP lead to phosphorylation of cholesterol
ester hydrolase and subsequent activation of this enzyme, to cause cleavage of the fatty
acid residues from the cholesterol ester (Jefcoate, DiBartolomeis, Williams and
McNamara, 1987; Simpson and Waterman 1988). In addition, cAMP triggered by
chronic response to ACTH regulates the expression of genes encoding steroidogenic
enzymes for a longer term maintenance of optimal levels of steroid hydroxylase
enzymes (Simpson and Waterman 1988,1992).
The renin-angiotensin system, extracellular ions and ACTH are the major
factors regulating the synthesis of aldosterone (Quinn and Williams 1988, 1992).
ACTH and prolactin are thought to control the production of adrenal androgens (Bondy
1985). In addition, the splanchnic nerves and several neuromediators have also been
demonstrated to have modulatory effects on adrenal steroidogenesis (see section 1:7).
1:6. Innervation of adrenal gland
1:6:1. Extrinsic adrenal innervation
The observation of a similar response of the gut upon stimulation of either the
greater splanchnic nerve or directly the adrenal gland led Jakobj (1892) to suggest that
the greater splanchnic nerve may innervate the gland. Dreyer (1899) provided more
direct evidence by demonstrating an increased pressor activity of the blood in the
adrenal vein of dog following stimulation of the greater splanchnic nerve. The
contribution of the lesser splanchnic nerve and branches arising from the lumbar
sympathetic chain in the innervation of the gland was later shown by Elliott (1912).
Several investigators have demonstrated that the splanchnic innervation to the gland is
29

ipsilateral (Elliott 1912; Cannon, Lewis and Britton 1926; Hollinshead 1936; Maycock
and Heslop 1939). However, a minor contralateral splanchnic innervation has also been
reported (Kahn 1911).
The observation that the secretory response of the adrenal medulla to electrical
stimulation of the splanchnic nerve could be abolished by nicotine, which also
produces sympathetic ganglionic block, indicated that the primary innervation to the
gland was preganglionic (Elliott 1913). A comparable degree of degeneration of nerve
fibres in the adrenal gland following section of either the greater splanchnic nerve or
branches from the coeliac ganglion to the gland, indicated that the adrenal nerve fibres
pass through the ganglion without synaptic interruption and further supported the
preganglionic nature of the major innervation to the gland (Hoshi 1926).
Studies from tracing of degenerating nerve fibres following nerve fibre
transection showed that the adrenal preganglionic nerves leave the spinal cord via the
anterior roots of third thoracic to second lumbar spinal nerves (Elliott 1913; Hoshi
1926; Swinyard 1937; Young 1939; MacFarland and Davenport 1941). Furthermore,
evidence from chromatolitic cell changes upon adrenal medullectomy, demonstrated
that the adrenal preganglionic nerve fibres originate from neurons located in the
intermediolateral cell column of the spinal cord (Cummings 1969). This was further
proved by studies involving injection of nerve tracing substances into the adrenal
medulla and locating the retrogradely labelled preganglionic cell bodies in the
intermediolateral cell column of thoracic and upper lumbar spinal cord segments, with
the majority of the neurons concentrated in the mid and lower thoracic regions (Ellison
and Clark 1975; Schramm, Adair, Stribling and Grey 1975; Hasse, Contestabile and
Flumerfelt 1982; Holets and Elde 1982; Bacon and Smith 1988; Kesse, Parker and
Coupland 1988; Strack, Sawyer, Marubio and Loewy 1988; Parker, Mohamed and
Coupland 1990b). The majority of the studies demonstrated an ipsilateral location of
the preganglionic neurons in the spinal cord. However, a bilateral location with a small
number of neurons found contralaterally has also been reported (Ross, Smolen and
30

Cherry 1980; Holets and Elde 1982).
Studies with nerve fibre tracer injection into the gland, have also indicated that a
small number of neurons located in the prcvcrtebral sympathetic chain in rat, guinea
pig and marmoset as well as the parasympathetic dorsal motor nuclei of the vagus in
rat and guinea pig have their terminals in the gland (Kesse et al. 1988; Coupland,
Parker, Kesse and Mohamed 1989; Parker et at. 1990b).
Earlier findings of a small number of degenerating nerve endings following
removal of the thoracic dorsal root ganglia suggested the presence of sensory nerve
fibres in the gland (Pines and Narowtschatowa 1931; Kiss 1951). More direct
morphological evidence was, however, obtained recently with the help of injection of
nerve fibre tracer substance into the gland and locating the retrogradely labelled
neurons in the ipsilateral dorsal root ganglia and the bilateral sensory vagal ganglia in
rat, guinea pig and marmoset (Afework 1988; Mohamed, Parker and Coupland 1988;
Coupland et al. 1989; Parker, Afework and Coupland 1990a; Parker, Kesse, Mohamed
and Afework 1993). The spinal sensory nerve fibres originate from the upper thoracic
to mid sacral dorsal root ganglia, and the majority of the neurons are located in the
lower thoracic ganglia.
Higher centres located in the brain stem and hypothalamus are implicated in the
regulation of adrenal medullary secretion. Electrical stimulation of various parts of the
hypothalamus and the midbrain causes catecholamine secretion in cat (Folkow and von
Euler 1954; Robinson, Culberson and Carmichael 1983; Stoddard-Apter, Siegel and
Levin 1983; Goadsby 1985) and rat (Gauthier and Reader 1982; Matsui 1984).
Stimulation of medulla oblongata has also been shown to result in catecholamine
secretion in rat (Matsui 1979). Moreover, transneuronal cell body labelling with
pseudorabies virus has provided morphological evidence for five centres within the
hypothalamus and brain stem that have connections with adrenal medullary nerve
fibres (Strack, Sawyer, Platt and Loewy 1989; Wesselingh, Li and Blessing 1989).
31

1:6:2. Intrinsic adrenal innervation
A small number of intrinsic neurons are found in the adrenal gland and are
believed to be involved with local adrenal innervation. Coupland (1965) described the
relative number of these neurons in comparison to the chromaffin cells in the adrenal
medulla of different mammalian species to be in descending order of guinea pig, man,
cat, dog, rat, mouse, sheep, ox and rabbit. The precise functional role of these neurons
is, however, not totally understood.
1:6:3. Course and distribution o f nerve fibres and terminals within adrenal cortex
Although it has long been shown that nerve fibres traverse the adrenal cortex,
there were a number of studies which failed to show local innervation (Hollinshead
1936; Swinyard 1937; MacFarland and Davenport 1941). Nevertheless, a number of
studies on axonal degeneration following section of adrenal nerve fibres have revealed
the existence of a subcapsular plexus in several species including rat, guinea pig, cat
dog and man (Alpert 1931; Willard 1936; Kiss 1951; Lever 1953; Mikhail and Mahran
1965). From the subcapsular plexus, nerve fibres have been described to innervate
cortical cells of the zonae glomerulosa (Alpert 1931; Willard 1936; Lever 1953) and
reticularis (Mikhail 1961; Mikhail and Mahran 1965). Evidence for the innervation of
adrenal cortex was further provided by ultrastructural studies which demonstrated
nerve terminals containing synaptic vesicles adjacent to the cortical cells in mouse, rat,
guinea pig, golden hamster, pig, sheep, monkey and man (Unsicker 1969, 1971;
Robinson, Perry, Hardy, Coghlan and Scoggins 1977; Uno 1977; Unsicker, Kabura-
Flüh and Zwarg 1978; Migally 1979; Dorovini-Zis and Zis 1991).
1:6:4. Course and distribution o f nerve fibres and terminals within adrenal medulla
Unlike that of adrenal cortex, the innervation of adrenal medulla is rich and has
been described by several investigators (MacFarland and Davenport 1941; Coupland
32

and Holmes 1958; Coupland 1965; Tomlinson and Coupland 1990). Nerve fibres branch
from the subcapsular plexus, and together with nerve bundles which directly cross the
capsule, pass through the cortex alongside the arteries and capillaries inward to the
medulla. Here, nerve fibres with associated Schwann cell cytoplasm branch and
rebranch to form a three dimensional network of nerve fibres around the chromaffin
and ganglion cells. The basement membrane of the Schwann cell fuses with that of the
chromaffin cell and the nerve fibres end as synaptic junctions.
1:7. Neuromediators in adrenal gland
A growing number of neuroactive substances have been demonstrated in the
adrenal chromaffin cells and/or neuronal components. These neuroactive substances
include acetylcholine, several biogenic amines, adenosine 5'-triphosphate (ATP) and
neuropeptides, as reviewed below.
1:7:1. Acetylcholine
The cholinergic nature of the great majority of the nerve fibres innervating the
adrenal medulla and which cause its secretion, was originally shown from functional
studies (Feldberg et al. 1934). Later, a diffuse network of cholinesterase-positive nerve
fibres were demonstrated in the adrenal medulla of cat, rat and rabbit (K ^le 1950;
Coupland and Holmes 1958). Electron microscopic studies have also revealed the
majority of adrenal medullary nerve terminals contain small synaptic electronlucent
vesicles, with positive cholinesterase activity in rat and bird (Lewis and Shute 1969;
Unsicker 1973b).
While the majority of the cholinergic nerve fibres traverse the cortex on their
way towards the medulla, some terminate in the cortex. Nerve fibres displaying
positive cholinesterase activity have been found in the adrenal cortex of rat, cat, rabbit,
sheep and goat (Coupland and Holmes 1958; Robinson et al. 1977; Purwar 1978;
33

Watanabe, Hiramtsu, Ohmori and Paik 1990). Furthermore, muscarinic receptors have
been demonstrated on bovine adrenocortical cells (Hadjian, Ventre and Chambaz
1981). Acetylcholine stimulates secretion of glucocorticoids and mineralocorticoids
from isolated adrenal gland of calf (Rosenfeld 1955) and frog (Benyamina,
Leboulenger, Lirhmann, Delarue, Fcullioley and Vaudry 1987). It also causes such
secretion from calf in vivo (Edwards and Jones 1990) and isolated adrenocortical cells
in cow (Hadjian, Guidicelli and Chambaz 1982).
1:7:2. Biogenic amines
The principal biogenic amines contained in the adrenal gland are adrenaline and
noradrenaline, which are synthesized and released by the adrenal chromaffin cells.
Substantial numbers of intra-adrenal nerves also contain catecholamines. Synaptic
vesicles typical of adrenergic profiles with predominantly small and some large
granular vesicles have been seen in association with adrenal chromaffin cells of toad
(Piezzi 1966). Electron microscopy combined with histochemistry, have shown a close
apposition of catecholaminergic nerve fibres to the adrenal chromaffin cells of bird
(Unsicker 1973c) and cat (Prentice and Wood 1975). Moreover, Omori, Okuno,
Fujisaw and Ono (1991) using TH immunoelectron microscopy have reported the
presence of catecholaminergic nerve fibres in close proximity to the rat adrenal
chromaffin cells. In contrast to such small number of studies which so far managed to
find catecholaminergic nerve fibres in association with the adrenal chromaffin cells,
there is an overwhelming evidence which showed the catecholamine-containing nerve
fibres innervate the capsule, cortex and blood vessels of the gland. Such evidence
comes from studies conducted with catecholamine histofluorescence combined with
electron microscopy in rat, Syrian hamster and sheep (Unsicker 1969; Robinson et al
1977; Kleitman and Holzwarth 1985); TH immunohistochemistry combined with
electron microscopy in rat (Oomori, Okuno, Fujisawa, Ishikawa, Satoh, Matsuda,
Yamano and Ono 1989; Oomori et al. 1991); the chromaffin reaction combined with
34

electron microscopy in rat (Vizi, Toth, Szalay, Windisch, Orso, Szabo and Vinson 1992)
and DBH immunohistochemistry in man (Gilchrist, Leake and Charlton 1993). The
catecholaminergic nerve fibres that are found in the cortex mainly innervate cells of the
zona glomerulosa, although a minor supply to those of the zona fasciculata has also
been found.
The catecholaminergic innervation to the gland is believed to be principally
noradrenergic. There is also evidence which suggested local noradrenergic axon
terminals may take up dopamine from the circulation and release it or convert it to
noradrenaline (Vizi, Toth, Orso, Szalay, Sz, Szabo, Baranyi and Vinson 1993).
Functionally, dopamine has been shown to inhibit aldosterone secretion from the zona
glomerulosa of rat (Pratt, Turner, Bowsher and Henry 1987). However, secretion of
aldosterone by direct /3-adrenergic stimulation has also been shown in cultures of
bovine adrenal cortical cells (De Lean, Racz, McNicoll and Desrosiers 1984).
The other biogenic amines immunohistochemically demonstrated in the adrenal
gland are histamine and serotonin. Histamine has been localized in NA cells of rat
adrenal gland and cultured bovine adrenal chromaffin cells (Hàppôlà, Soinila,
Paivarinta, Joh and Panula 1985; Tuominen, Karhunen, Panula and Yamatodani 1993).
It stimulates the release of catecholamines from isolated bovine adrenal chromaffin
cells (Pender and Burgoyne 1992) and perfused rat adrenals (Borges 1994). Histamine
also causes corticosteroid secretion from the adrenal gland of rat and isolated dog
adrenocortical cells (Mikolajczyk 1965; Hirose, Matsumoto and Aikawa 1978).
Serotonin has been demonstrated in A cells of adrenal gland of rat (Holzwarth,
Sawetawan and Brownfield 1984; Verhofstad and Jonsson 1983). It stimulates
corticosteroid secretion in the rat (Mikolajczyk 1965).
1:7:3. Adenosine 5'-triphosphate
In addition to its role as intracellular energy source, ATP has also extracellular
35

signalling functions and acts as a neurotransmitter and/or modulator (Bumstock 1972,
1975). In the chromaffin cells, ATP is costored and released with the catecholamines
(Douglas, Poisner and Rubin 1965; Stevens, Robinson, Van Dike and Stizel 1975;
Winkler and Westhead 1980). The released ATP has been suggested to exert a
modulatory role on catecholamine secretion, since ATP has been shown to inhibit
(Chem, Herrera, Kao and Westhead 1987) or facilitate (Chem, Kim, Slakey and
Westhead 1988; Kim and Westhead 1989) catecholamine release from isolated bovine
chromaffin cells. More recently, ATP has been shown to induce catecholamine
secretion from perfused bovine adrenal gland (Lin, Bott, Kao and Westhead 1995).
1:7:4. Neuropeptides
1:7:4:1. Enkephalins
Met-enkephalin and leu-enkephalin were the first two opioid peptides
originally isolated from the brain and described as 5 amino acid polypeptides (Hughes,
Smith, Kosterlitz, Forthergill, Morgan and Morris 1975). Subsequently, a number of
other peptides with opiate activity were discovered (Weber, Evans and Barchas 1983).
Enkephalins were also the first peptides to be identified immunohistochemically in the
adrenal chromaffin cells (Schultzberg, Lundberg, Hokfelt, Temius, Brandt, Elde and
Goldstein 1978). They have been demonstrated in the adrenal chromaffin cells of
several species including rat, guinea pig, hamster, cat, rabbit, dog, horse and man
(Schultzberg et at. 1978; Lundberg, Hamberger, Schultzberg, Hokfelt, Granberg,
Efendic, Terenius, Goldstein and Luft 1979; Linnoila, Diaugustine, Hervonen and
Miller 1980; Livett and Dean 1980; Livett, Day, Elde and Howe 1982; Vamdell,
Topia, Demey, Rush, Bloom and Polak 1982; Kobayashi, Ohashi, Fujita, Nakao,
Yoshimasa, Imura, Mochizuki, Yanaihara, Yanaihara and Verhofstad 1983; Kobayashi,
Ohashi, Uchida, Nakao, Imura, Yanaihara and Verhofstad 1984; Kondo, Kuramoto and
Iwanaga 1984; Heym 1985; Kondo 1985; Heym and Kummer 1988; Pelto-Huikko
1989). However, the localization of enkephalins to either A or NA cells is equivocal.
36

Schultzberg et al. (1978) reported as met-enkephalin immunoreactivity is located in the
NA cells of rat, with no such preference being found in the cat. Kobayashi et at. (1983,
1984) and Kondo et at. (1984) reported that met-enkephalin-arg-gly-leu and m et-
enkephalin-arg-phe have no preference of localization to either A or NA cells in rat,
cat, hamster and dog. Pelto-Huikko (1989), in contrast, described that
immunoreactivity to both met-enkephalin and leu-enkephalin was mainly found in A
cells in rat, cat and hamster, and only in NA cells in mouse.
Nerve fibres and terminals immunoreactive for Met-enkephalin have been
found in rat, mouse, guinea pig and cat adrenal medulla (Schltzberg et al. 1978;
Kobayashi et al. 1983; Pelto-Huikko 1989). Such nerve fibres innervate A cells and
some medullary neurons of the gland (Pelto-Huikko 1989).
Enkephalins cause a non-specific inhibition of the nicotinic-evoked, but an
enhancement of the basal, release of catecholamines from cultured bovine adrenal
chromaffin cells (Lemaire, Lemaire, Dean and Livett 1980; Dean, Lemaire and Livett
1982; Saiani and Guidotti 1982; Livett, Boksa, Dean, Mizobe and Lindenbaum 1983).
They reduce the release of acetylcholine and substance P, and thereby inhibit synaptic
transmission (Konishi, Tsunoo and Otsuka 1981). Both met- and leu-enkephalins
stimulate aldosterone secretion from bovine adrenal glomerulosa cells (Bruzzone and
Marusic 1988). They also stimulate aldosterone and corticosterone secretion in
perfused rat adrenal preparation (Hinson, Cameron, Purbick and Kapas 1994b; Hinson,
Purbick, Cameron and Kapas 1994c).
1:7:4:2. Vasoactive intestinal peptide
Vasoactive intestinal peptide (VIP) is a 28 amino acid polypeptide originally
isolated from pig small intestine (Said and Mutt 1970). It is part of a pre-prohormone
that is cleaved into VIP and peptide histidine-isoleucine and is structurally related to
secretin, glucagon and gastric inhibitory peptide (Itoh, Obata, Yanaihara and Okamoto
37

1983). Immunoreactivity for VIP occurs in a small number of A cells of rat and human
adrenal medulla and cultured bovine chromaffin cells (Bryant, Bloom, Polak,
Albuquerque, Modlin and Pearse 1976; Kondo 1985; Siegel, Biden and Pruss 1985).
Immunoreactivity for VIP also occurs in a number of intra-adrenal neurons of rat
(Hokfelt, Lundberg, Schultzberg and Fahrenkrug 1981; Holthwarth 1984; Kondo 1985;
Dagerlind, Goldstein and Hokfelt 1990; Oomori, Okuno, Fujisawa, luchi, Ishikawa,
Satoh and Ono 1994).
VIP-immunoreactive nerve fibres forming a plexus with varicosities and nerve
terminals are found in the zona glomerulosa and subcapsular region of the cortex of
adrenal gland of several species, including rat, guinea pig, cat, dog, pig, hamster and
man (Linnoila et al. 1980; Hokfelt et al 1981; Holzwarth 1984; Kondo 1985; Heym
and Kummer 1988). A moderate distribution of VIP-immunoreactive nerve fibres also
occurs in the adrenal medulla.
VIP stimulates catecholamine release from adrenal medullary cells of foetal bird
(Cheung and Holzwarth 1986) and perfused rat adrenal gland (Guo and Wakade 1994).
A rise in the concentration of blood aldosterone and corticosterone levels occurs
following administration of VIP in rat (Nussdorfer and Mazzocchi 1987; Pralong,
Corder and Gaillard 1991). Prolonged (7 days) treatment of rat with VIP stimulates the
growth and steroidogenic capacity of the zona glomerulosa of adrenal cortex
(Mazzocchi, Robba, Malendowicz and Nussdorfer 1987). In perfused adrenal gland of
pig, administration of VIP causes an increase in both aldosterone and cortisol secretion
(Ehrhart-Bomstein, Bomstein, Scherbaum, Pfeiffer and Holst 1991). Similarly, VIP
causes an increase in secretion of both aldosterone and corticosterone in perfused rat
adrenal gland preparation (Hinson, Kapas, Orford and Vinson 1992; Hinson et al
1994b, c). Reports on the effect of VIP on isolated adrenocortical cells are, however,
not coherent. Some studies have found as VIP could stimulate steroidogenesis (Kowal,
Horst, Pensky and Alfonzo 1977; Morera, Cathiard, Lurthe and Saez 1979; Li, Queen
and LaBella 1990), while some others found it with no effect (Hinson et al 1992;
38

Enyedi, Szabo and Spat 1983).
1:7:4:3. Somatostatin
Somatostatin is a 14 amino acid polypeptide originally isolated from bovine
hypothalamus (Brazeau, Vale, Burgus, Ling, Butcher, Rivier and Guillemin 1973).
Singly dispersed somatostatin-immunoreactive chromaffin cells occur in human
adrenal gland (Lundberg et al. 1979). In addition, nerve fibres immunoreactive for
somatostatin are found in the adrenal medulla of guinea pig and cat (Heym and
Kummer 1988).
Somatostatin causes a noncompetitive inhibition of catecholamine secretion
from isolated bovine and pig chromaffin cells (Mizobe, Kozousek, Dean and Livett
1979; Role, Leeman and Perlman 1981; Livett et al. 1983). It also inhibits the growth
of the zona glomerulosa and its steroidogenic capacity in rat (Rebuffat, Robba,
Mazzocchi and Nussdorfer 1984).
1:7:4:4. Neurotensin
Neurotensin was originally isolated from bovine hypothalamus and
characterized as a 13 amino acid polypeptide (Carraway and Leeman 1973, 1975). In
the cat, immunoreactivity for neurotensin occurs in the majority of the NA cells
(Lundberg, Rokaeus, Hokfelt, Rosell, Brown and Goldstein 1982; Terenghi, Polak,
Vamdell, Lee, Wharton and Bloom 1983). However, it is found only in a few adrenal
chromaffin cells in guinea pig, rat and dog (Heym 1985; Reinecke 1985). Neurotensin-
immunoreactive nerve terminals innervate NA cells and intra-adrenal neurons in
hamster and cat (Lundberg et al. 1982; Pelto-Huikko, Salminen, Partnen, Toivanen
and Hervonen 1985).
Administration of neurotensin in the rat causes hypertrophy of the zona
glomerulosa and elevates plasma aldosterone levels (Malendowicz, Nussdorfer,
39

Majchrzak, Nowak and Lesniewska 1992). It has also been shown that infusion of
neurotensin into isolated rat adrenal gland causes an increase in both aldosterone and
corticosterone secretion (Hinson et al. 1994b, c). Moreover, the finding of high levels
of specific binding sites for neurotensin on the cells of zona reticularis of rat adrenal
gland has suggested that circulating neurotensin may also influence steroid secretion
from this region of the gland as well (Goedert, Mantyh, Hunt and Emson 1984).
1:7:4:5. Calcitonin gene-relatedpeptide
Calcitonin gene-related peptide (CGRP) was first isolated from rat adrenal
medullary cells as a 37 amino acid polypeptide which is expressed by the same gene as
that of calcitonin by alternative processing of the mRNA (Rosenfeld, Mermod, Amara,
Swanson, Sawchenko, Rivier, Vale and Evans 1983). Subsequent studies have shown
its localization in a number of A cells of adrenal medulla in mouse, rat, cat, hamster
and man (Kondo 1985; Kuramoto, Kondo and Fujita 1987; Pelto-Huikko 1989).
Several CGRP-immunoreactive nerve fibres are found in the adrenal gland of
rat, cat, mouse, hamster, rabbit and pig (Rosenfeld et al. 1983; Kuramoto et al. 1987;
Heym and Kummer 1988; Pelto-Huikko 1989). Such immunoreactive fibres are
associated with chromaffin cells and ganglion cells in the medulla and some blood
vessels in the cortex.
CGRP has inhibitory effect on aldosterone secretion when administered to dog
and rat in vivo (Murakami, Hiromichi, Nakajima, Nakamoto, Kageyama and Saruta
1989a; Mazzochi, Malendowicz, Meneghelli and Nussdorfer 1992). It also causes
inhibition of steroid biosynthesis in isolated rabbit adrenal glomerulosa cells
(Murakami et al. 1989a). However, in isolated perfused rat adrenal gland, CGRP has
caused vasodilatation accompanied with an increased secretion of aldosterone (Hinson
and Vinson 1990). It has also been shown that CGRP causes cortisol secretion from
adrenal gland of calf in vivo (Bloom, Edwards and Jones 1989).
40

1:7:4:6. Neuropeptide Y
Neuropeptide Y (NPY) is a 36 amino acid polypeptide and shares a considerable
sequence homology with various pancreatic polypeptides and peptide YY (Tatemoto,
Carlquist and Mutt 1982). Immunohistochemically, NPY has been demonstrated in
adrenal chromaffin cells in mouse, rat, guinea pig, hamster, cat, pig, dog, horse, cow
and man (Vamdell, Polak, Allen, Terengchi and Bloom 1984; Kuramoto, Kondo and
Fujita 1986; Lundberg, Hokfelt, Hcnscn, Theodorsson-Norheim, Pemow, Hamberger
and Goldstein 1986; Schalling, Seroogy, Hokfelt, Chai, Hallman, Persson, Larhammar,
Ericsson, Terenius, Graffi, Massoulie and Goldstein 1988b; Pelto-Huikko 1989). The
localization of NPY immunoreactivity in A and NA cells is not coherent, with
conflicting reports in the literature. NPY immunoreactivity was originally described in
the NA cells of adrenal medulla of mouse, rat, guinea pig, cat, horse and cow (Vamdell
et al. 1984; Majane, Alho, Kataoka, Lee and Yang 1985). However, subsequent
investigators have reported NPY immunoreactivity is instead located in the A cells of
mouse, rat, hamster, guinea pig, cat, bovine and man adrenal medulla (De Quidt and
Emson 1986; Kuramoto et al. 1986; Lundberg et al. 1986; Pelto-Huikko 1989).
Furthermore, Schalling et al. (1988b) described NPY immunoreactivity in all A and a
small number of NA cells of rat adrenal medulla.
Immunoreactivity for NPY also occurs in the ganglion cells of the adrenal gland
in rat, guinea pig and hamster (Pelto-Huikko 1989; Dagerlind et al. 1990; Oomori et
al. 1994). Several nerve fibres and terminals immunoreactive for NPY are found in the
adrenal gland of rat, mouse, guinea pig, hamster, dog, horse and cow (Vamdell et al.
1984; Majane et al. 1985; De Quidt and Emson 1986; Kuramoto et al. 1986; Pelto-
Huikko 1989; Maubert, Tramu, Croix, Beauvillain and Dupouy 1990). Such
immunoreactive nerve fibres form a plexus in the zona glomemlosa, and make synaptic
contacts with cortical cells, chromaffin and ganglion cells of medulla and some blood
vessels in the gland.
41

Specific binding sites for NPY have been localized on the glomerulosa cells of
bovine adrenal cortex (Torda, Cruciani and Saavedra 1988). In the rat, NPY is shown
to cause aldosterone secretion from adrenal gland in vivo (Mazzocchi and Nussdorfer
1987) and perfused adrenal gland preparation (Hinson et al. 1994b). In addition,
hypertrophy of the rat zona glomerulosa, accompanied with a rise in plasma levels of
aldosterone, were observed following prolonged treatment with NPY (Rebuffat,
Malendowicz, Belloni, Mazzocchi and Nussdorfer 1988a). In contrast, prolonged
treatment of a smaller dose of NPY has resulted in inhibition of aldosterone and
corticosterone levels accompanied with a decrease in the volume of the zona
fasciculata cells of adrenal gland in rat (Lesniewska, Nowak, Miskowak, Nussdorfer
and Malendowicz 1990). NPY has also been found to inhibit corticosterone secretion
from dispersed rat zonae fasciculata and reticularis cells (Malendowicz, Lesniewska
and Miskowiak 1990).
1:7:4:7. Substance P
It was von Euler and Gaddum (1931) who first extracted substance P (SP) from
horse brain and intestine, and observed that it could cause hypotension and tachycardia
when administered intravenously. Later, SP was identified as an 11 amino acid
polypeptide (Chang, Leeman and Niall 1971). Immunoreactivity for SP has been
observed in a number of adrenal chromaffin cells in rat, dog, guinea pig and rabbit
(Heym 1985; Kuramoto, Kondo and Fujita 1985; Heym and Kummer 1988).
SP-immunoreactive nerve fibres have been localized in the capsule and zona
glomerulosa of adrenal cortex in rat, guinea pig and man (Linnoila et al. 1980;
Kuramoto et al. 1985). SP-immunoreactive nerve fibres with varicosities have also
been demonstrated in apposition to chromaffin cells of rat, guinea pig, hamster, dog,
pig, cat and man adrenal medulla (Linnoila et al. 1980; Kuramoto et al. 1985; Heym
and Kummer 1988).
At a relatively high concentration (> 10” ^ mol/1), SP inhibits the nicotinic
42

acetylcholine-mediated catecholamine release from adrenal chromaffin cells (Livett,
Kozousek, Mizobe and Dean 1979; Mizobe, Kozouzek, Dean and Livett 1979; Livett
et aL 1983). At a lower concentration (< 10” ^ mol/1), SP prevents the nicotinic
receptor desensitization and thereby protects and prolongs catecholamine release from
adrenal chromaffin cells (Livett et al. 1983; Boksa and Livett 1984). It has also been
shown that SP facilitates acetylcholine release from the splanchnic nerve terminals
presynaptically (Zhou and Livett 1990). By itself SP has no effect on secretion, but
requires the presence of acetylcholine or a nicotinic agonist for effect (Livett, Marley,
Wan and Xin-Fu 1990). It has therefore been suggested that SP by acting as a
modulator maintains a prolonged output of catecholamines during stress condition
(Livett et at. 1983; Livett et al. 1990).
SP enhances aldosterone secretion from rat adrenal gland in vivo (Nussdorfer,
Malendowicz, Belloni, Mazzocchi and Rebuffat 1988). Prolonged infusion of SP
results in the hypertrophy of zona glomerulosa cells associated with a rise in
aldosterone levels (Nussdorfer et al. 1988). In addition, SP increases both aldosterone
and corticosterone secretion from isolated perfused rat adrenal gland (Hinson et al.
1994b, c).
1:7:4:8. Bombesin
Bombesin was originally isolated from frog skin extracts and characterized as a
14 amino acid polypeptide (Anastasi, Erspamer and Bucci 1971). Later, it was found in
the mammalian central and peripheral nervous system (Panula 1986).
Immunohistochemically, bombesin has been demonstrated in bovine adrenal medullary
cells (Lemaire, Chouinard, Mercier and Day 1986) and guinea pig adrenal medullary
nerve fibres (Heym and Kummer 1988). Bombesin has a direct stimulating action on
catecholamine secretion from rat adrenal gland in vivo (Okubo, Kaku, Kaneko and
Yanaihra 1985). In contrast, it inhibits basal corticosterone secretion from isolated cells
43

of the zonae fasciculata and reticularis of rat adrenal cortex (Malendowicz, Lesniewska,
Baranowska, Nowak and Majchrzak 1991).
1:7:4:9. Galanin
The 29 amino acid polypeptide, galanin was first isolated from pig intestine
(Tatemoto, Rokaeus, Jomvall, McDonald and Mutt 1983). Immunoreactivity for
galanin has been shown in adrenal chromaffin cells of mouse, hamster and cat (Pelto-
Huikko 1989). A few nerve fibres showing galanin immunoreactivity have been found
near intra-adrenal blood vessels in rat, mouse, cat and hamster (Pelto-Huikko 1989).
Galanin stimulates secretion of corticosterone and aldosterone in rat in vivo
(Mazzocchi, Malendowicz, Rebuffat and Nussdorfer 1992). It also causes release of
both corticosterone and aldosterone from isolated perfused porcine adrenal gland
(Holst, Ehrhart-Bomstein, Messell, Poulsen and Harling 1991).
While the above described neuromediators are the most established ones with
functional evidence in the adrenal glandular activities, there are also other additional
neuroactive substances demonstrated in the gland. These include: atrial natriuretic
polypeptide (Inagaki, Kubota, Kito, Kangawa and Matsuo 1986; Heym and Kummer
1988); corticotrophin-releasing hormone (Bruhn, Engeland, Anthony, Gann and
Jackson 1987; Rundle, Benedict, Robinson and Funder, 1988); vasopressin and
oxytocin (Hawthorn, Nussey, Henderson and Jenkins 1987); delta sleep-inducing
peptide (Ekman, Bjartell, Eklblad and Sundler 1987); growth-associated protein
(Grant, Konig, Deloume, Aunis and Langley 1992; Dorsey and Schmidt 1993) and
gamma-aminobutyric acid (Kataoka, Fujmoto, Alho, Guidotti, Geffard, Kelly and
Hambauer 1986; Oomori, luchi, Nakaya, Tanaka, Ishikawa, Satoh and Ono 1993). In
addition, by the time the work presented in this thesis was commenced a few studies
have indicated the presence of the NO-synthesizing enzyme, NOS in bovine and rat
adrenal glands (Palacios et al. 1989; Bredt et al. 1990; Dawson et al. 1991). While this
work was in progress, additional reports on localization of NOS in rat and human
44

adrenal glands have also appeared (Aim, Larsson, Ekblad, Sundler and Andersson 1993;
Dun, Dun, Wu and Forstermann 1993; Heym, Colombo-Benckmann and Mayer 1994).
As the present study is concerned with investigation of the NO-synthesizing cellular
elements in the adrenal gland of rat, relevant literatures regarding NO is reviewed
below.
1:8. Nitric oxide
NO is a colourless gas with an extremely simple chemical structure. It is a free
radical, labile molecule whose activity, when superfused over a tissue, decays by 50 %
in about 4 seconds (Moncada, Palmer and Higgs 1989). It reacts with oxygen to form
nitrogen dioxide, ultimately breaking down into the stable end products nitrite and
nitrate. Recently, it has come apparent from observations by investigators in different
unrelated research areas that NO is a major secretory product of mammalian cells in
host defence and homeostasis.
Mitchell, Shonle and Grindley (1916) first suggested that mammals produced
oxides of nitrogen. This was, however, largely ignored or thought to be due to
intestinal nitrifying bacteria (White 1975; Tannenbaum, Fett, Young, Land and Bruce
1978). Later, Green, Tannenbaum and Goldman (1981), while analyzing the metabolic
balance of nitrate levels observed that germ-free and ordinary rats excreted
comparably more nitrate than they ingested. Therefore, nitrate synthesis in mammals
was suggested to be a normal process, than is due to intestinal nitrifying bacteria.
Furthermore, the observation of an enhanced nitrate excretion in a human with an
episode of fever and diarrhoea instigated investigations and the discovery that rats
injected with Escherichia coli lipopolysaccharides (LPS) could result in an increase in
nitrate excretion (Wagner and Tannenbaum 1982; Wagner, Young and Tannenbaum
1983). Subsequently, Stuehr and Marietta (1985) with studies on LPS-induced nitrate
biosynthesis in LPS-sensitive versus LPS-resistant mice combined with experiments
45

on cells in culture showed that macrophages can account at least, in part, for the increase
in nitrate excretion. In such nitrate biosynthesis pathway of macrophages, L-arginine
was found to be the substrate while L-citrulline was a coproduct (Hibbs, Taintor and
Vavrin 1987a; Iyengar, Stuehr and Marietta 1987). It was subsequently found that NO
was an intermediate coproduct along with L-citrulline (Marietta, Yoon, Iyengar, Leaf
and Wishnok 1988).
In another laboratory, Furchgott and Zawadzki (1980) observed the relaxation of
isolated preparations of rabbit thoracic aorta by acetylcholine, required an intact
endothelium. This led to the postulation that a labile substance, which was termed
endothelium derived relaxing factor (EDRF), was released from stimulated
endothelium to bring about vasodilatation. Subsequently, the effect of EDRF in the
smooth muscle of blood vessels was shown to be acting by increasing the levels of
cGMP (Rapoport and Murad 1983). This indicated the similarity of the mode of action
of EDRF to that of nitrovasodilators (Katsuki and Murad 1977; Katsuki, Arnold and
Murad 1977), which do not require endothelium for their effect (Furchgott and
Zwadzki 1980; Rapoport and Murad 1983). As nitrovasodilators had already been
speculated to exert their effect via NO (Arnold, Mittal, Katsuki and Murad 1977;
Gruetter, Barry, McNamara, Gruetter, Kadowitz and Ignarro 1979) EDRF was
postulated to be NO. Eventually, based on the indistinguishability of EDRF from NO
in terms of biological activity, stability and susceptibility to an inhibitor and to a
potentiator, EDRF was identified to be identical to NO (Ignarro, Buga, Wood, By ms
and Chudhuri 1987; Palmer, Ferrige and Moncada 1987).
Another interesting finding was made by Garthwaite, Charles and Chess-
Williams (1988). Upon stimulation of cerebellar slices with excitatory amino acids,
they observed that there was release of a labile substance which had pharmacological
properties similar to those of NO, including the ability to raise cGMP and to relax
vascular smooth muscle. In retrospect, NO was then considered to be responsible for
the earlier unexplained observations of increased cGMP levels in certain brain regions
46

upon stimulation by neurotransmitters, particularly amino acids (Drumond 1983). In
fact, there had been previous reports where NO was shown to stimulate guanylyl
cyclase activity in the brain (Arnold et al. 1977; Miki, Kawabe and Kuriyama 1977),
although this was considered to be simply a pharmacological phenomenon.
1:8:1. Biosynthesis o f nitric oxide: nitric oxide synthase
In mammalian cells, NO is synthesized from the guanidino nitrogen of L -
arginine by the enzyme NOS (EC 1.14.13.39) through a process that consumes five
electrons and results in the formation of L-citrulline. There are three isoforms of NOS
which have been identified and their cDNAs are isolated. These are: NOS isoform I, in
brain; NOS isoform II, in cytokine and endotoxin-induced cells such as macrophages,
hepatocytes and vascular smooth muscles; and NOS isoform III, in endothelial cells
(see Forstermann et al. 1991; Nathan 1992; Forstermann, Close, Pollock, Nakane,
Schwartz, Gath and Kleinert 1994). In the literature, there are varying nomenclatures
assigned to the three NOS isoforms. Isoform I is also referred to as brain, neuronal or
constitutive NOS. Isoform II is also called inducible or macrophage NOS, while
isoform III is referred to as endothelial or constitutive NOS (Forstermann et al 1994;
Nathan and Xie 1994a).
Each of the NOS isoforms is produced by distinct genes with the deduced NOS
amino acid sequences from the three different cDNAs being approximately 50-60 %
identical with one another. In humans, the genes for NOS isoform I are located on
chromosome 12 (Nakane, Schmidt, Pollock, Forstermann and Murad 1993), whereas
those of isoforms II and III are located on chromosomes 17 and 7, respectively
(Marsden, Heng, Schere, Stewart, Hall, Shi, Tssi and Schappert 1993; Xu, Charles,
Moncada, Gorman, Sheer, Liu and Emson 1994). The molecular mass of the denatured
proteins of NOS isoforms I, II and III are 150-160 kDa, 133-135 kDa and 125-135
kDa, respectively (Bredt and Snyder 1990; Revel, White and Marietta 1991; Schmidt,
47

Pollock, Nakane, Gorsky, Forstermann and Murad 1991; Stuehr, Cho, Kwon, Weise and
Nathan 1991; Lamas, Marsden, Li, Tempt and Michel 1992; Sessa, Harrison, Barber,
Zeng, Durieux, D'Angelo, Lynch and Peach 1992).
All the three isoforms of NOS use L-arginine and molecular oxygen as
substrate and require the cofactors NADPH, tetrahydrobiopterine (BH4 ), flavin adenine
dinucleotide (FAD) and flavin mononucleotide (FMN). They all bind calmodulin and
contain heme (Marietta et al. 1988; Mayer, John, Heinzel, Werner, Wachter, Schultz
and Bohme 1991; Forstermann et at. 1994). The C-terminal half of NOS has similarity
to the cytochrome P-450 reductase where there are binding sites for FMN, FAD and
NADPH (Sessa 1994).
The NOS isoforms I and III are generally constitutively expressed by cells
which contain them, and require calcium and calmodulin for their activity (Knowles,
Palacios, Palmer and Moncada 1989; Bredt and Snyder 1990; Malinski and Taha
1992). Following receptor or physiologically induced increase in the intracellular
calcium level, calcium binds to calmodulin and the calcium-calmodulin complex
activates NOS. This facilitates the formation of NO in small amounts (picomoles) for a
short period of up to several minutes (Malinski and Taha 1992). Such intermittent
productions of NO in small amounts transmit signals. Although, originally NOS
isoform I is thought to be exclusively a cytosolic enzyme (Schmidt et al. 1991), recent
findings show that up to 60 % of its total activity in rat and rabbit cerebella is found in
the particulate fraction (Hecker, Miilsch and Busse 1994). About 95 % of the NOS
isoform III is found as particulate, while the remaining 5 % is cytosolic (Pollock,
Forstermaim, Mitchell, Warner, Schmidt, Nakane and Murad 1991).
The NOS isoform II is normally absent from the cell until it is activated by
cytokines, endotoxins or exotoxins (Moncada, Palmer and Higgs 1991; Nathan 1992).
Upon its activation, the enzyme facilitates the release of large amount of NO
(nanomoles), for a period of up to 5 days, which acts as a cytotoxic molecule against
invading microorganisms and tumour cells (Vodovotz, Kwon, Pospischil, Manning,
48

Paik and Nathan 1994). The NOS isoform II is found as a soluble fraction. Like the
isoforms I and III which are calcium dependent, the NOS isoform II has also binding
site for calmodulin despite the calcium independence of its activity (Cho, Xie,
Calaycay, Mumford, Swiderek, Lee and Nathan 1992; Lowenstein, Glatt, Bredt and
Snyder 1992; Schini and Vanhoutte 1992). However, in NOS isoform II, calmodulin is
very tightly bound and is unaffected by calcium.
NOS isoform I has been localized immunohistochemically in neurons found in
both central and peripheral nervous tissue; astrocytes; epithelial cells of lung, uterus
and stomach; kidney macula densa cells; and pancreatic islet cells (see Stuehr and
Griffith 1992; Henry, Lepoivre, Drapier, Ducrocqu, Boucher and Guissani 1993;
Forstermann et aL 1994). Induced NOS isoform II has been localized
immunohistochemically in a variety of cells which include macrophages; lymphocytes;
neutrophils and eosinophils in red pulp of spleen; kupffer cells; endothelial cells and
hepatocytes of liver; alveolar macrophages in lung; macrophages and endothelial cells
in adrenal gland; eosinophils mast cells and endothelial cells in colon; nerves
supplying mesenteric blood vessels; and epithelium of large air ways (Bandaletova,
Brouet, Bartsch, Sugimura, Esumi and Ohshima 1993; Buttery, Springall, Carpenter,
Riveros-Moreno, Moncada, Cohen and Polak 1993; Kobzik, Bredt, Lowenstein,
Drazen, Gaston, Sugarbaker and Stamler 1993). NOS isoform III has been localized in
endothelial cells of a wide variety of arterial and venous blood vessels (Pollock,
Nakane, Buttery, Martinez, Springall, Polak, Forstermann and Murad 1993).
1:8:2. Functional significance o f nitric oxide
The best characterized molecular target of NO in the cell is iron contained in
certain proteins as a heme group or as an iron-sulphur complex (Henry et al. 1993).
Binding of NO to such iron-containing enzymes leads to either activation or
inactivation of the enzyme. One enzyme which is activated by NO is guanylatc
49

cyclase. Binding of NO to the iron located in the heme group of guanylate cyclase
causes the production of cGMP, which in turn activates a cascade of other cellular
processes leading to activities such as dilatation of blood vessels (Moncada et al. 1991)
and neuronal transmission (Garthwaite 1991; Schuman and Madison 1994). Important
enzymes that are inhibited by the binding of NO to the iron contained in their catalytic
centre include: ribonucleotide reductase, which is rate-limiting in DNA replication;
iron-sulphur cluster-dependent enzymes (complex I and II) involved in mitochondrial
electron transport; and *'cis”-aconitase in the citric acid cycle (Kwon, Stuehr and
Nathan 1991; Lepoivre, Fieschi, Coves, Thelander and Fontecave 1991; Nathan and
Hibbs 1991; Beredeaux 1993; Stamler 1994). Such diversity of the targets of NO
coupled with its production by wide variety of cells make NO to be involved in
multiple functions. So far it has been implicated in the cardiovascular system,
immunity and inflammation and neuronal transmission.
1:8:2:1. Nitric oxide in the cardiovascular system
Small amounts of NO continuously released from the vascular endothelial cells
brings about a basal level of vascular smooth muscle relaxation (Moncada et al. 1991).
Furthermore, NO released from the endothelial cells and also possibly from the
platelets causes inhibition of platelet aggregation and adhesion, and thereby it prevents
blood clot and facilitates blood flow (Radomski and Moncada 1991). Release of NO
along with other possible factors from endocardium have been suggested to be
involved in the regulation of cardiac function (Smith, Shah and Lewis 1991; Smith,
Shah, Fort and Lewis 1992). NO released form perivascular nerves of certain blood
vessels may also contribute to the regulation of blood flow and pressure (Rand 1992).
By such means, NO regulates the systemic blood pressure as well as local blood flow
in specific vascular beds in brain, kidney, lung, heart and gastrointestinal tract
(Moncada and Higgs 1993; Lowenstein, Dinerman and Snyder 1994). Decrease in the
systemic NO production could therefore lead to vasospasm or hypertension, as for
50

example, decreased endothelium-dependent relaxation is found in patients with
essential hypertension (Panza, Quyyami, Brush and Epstein 1990). In contrast,
overproduction of NO is implicated in hypotension of sepsis, and may also be
associated with rare causes of abnormally low blood pressure, such as idiopathic
orthostatic hypotension (Lowenstein et al. 1994).
1:8:2:2. Nitric oxide in immunity and inflammation
Locally released NO in large quantities from activated cells, such as
macrophages and neutrophils, as well as destroying invading pathogenic organisms
may damage normal near by cells. By such a mechanism, NO kills or inhibits the
growth of many pathogens including bacteria, fungi, parasites and tumour cells, and
also normal host cells in autoimmune diseases (Lowenstein et al. 1994). Inhibitors of
NOS have been shown to reduce acute inflammation in rat (lalenti, Ignaro, Moncada
and Di Rosa 1992). In addition, glucocorticoids, in a potency which correlates to that
of their antiinflammatory action have been found to inhibit induction of the calcium-
independent NOS, but not the activity of the induced enzyme (Moncada and Palmer
1991). It has, therefore, been suggested that NO may be involved in the pathogenesis
of inflammation (Moncada and Palmer 1991; lalenti et al. 1992).
1:8:2:3. Nitric oxide in neuronal transmission
In the central nervous system, NO has been suggested to participate in
modulating neuronal transmission. NO released postsynaptically is suggested to diffuse
in retrograde fashion and affect the presynaptic neurons resulting in a stable increase in
synaptic transmission, as a phenomenon of long-term potentiation (LTP) (Bohme,
Bon, Stutzmann, Doble and Blanchard 1991; Schuman and Madison 1994). Such LTP
is thought to be linked to memory function. Indeed, NOS inhibitors have been found to
impair learning and can produce amnesia in chick and rat (Chapman, Atkins, Allen,
51

Haley and Steinmetz 1992; Hoischer and Rose 1992; Bohme, Bon, Lemaire, Reibaud,
Plot, Sutzmann, Doble and Blanchard 1993), The NO released from the postsynaptic
neurons during LTP is, however, found to be generated by the NOS isoform III rather
than the NOS isoform I (O’Dell, Huang, Dawson, Dinerman, Snyder, Kandel and
Fishman 1994).
Another activity where NO has been implicated in the central nervous system
concerns neurotoxicity. Excessive calcium entry during overstimulation with excitatory
amino acids has been associated with high levels of NO production in the central
nervous system leading to convulsion and neurotoxicity (Garthwaite 1991; Moncada
and Higgs 1993; Triggle 1994). In such manner, NO has been implicated in neuronal
damage during a variety of neuropathological conditions including stroke, Parkinson's
disease, multiple sclerosis and the dementia associated with Alzheimer's disease and
acquired immunodeficiency syndrome (Moncada and Higgs 1993; Triggle 1994).
In the peripheral autonomic nervous system, NO is implicated in the non-
adrenergic non-cholinergic inhibitory neurotransmission. NO has been found to
mediate some forms of gastrointestinal relaxation, including dilatation of the stomach
to adapt to increase in intra-gastric pressure (Rand 1992). NO released from pelvic
plexus neurons has been shown to be responsible for penile erection (Ignarro 1990;
Burnett, Lowenstein, Bredt, Chang and Snyder 1992). Additionally, NO released from
nerve fibres innervating adventitia of certain blood vessels may participate in the
regulation of blood flow (Rand 1992). NO has also been found involved in the
relaxation of tracheal muscle (Belvis, Stretton, Yacoub and Barnes 1992) and bladder
(Persson, Igawa, Mattiasson and Andersson 1992). In the adrenal gland, functional
studies have suggested the involvement of NO during secretion of catecholamines
(Dohi, Morita and Tsujimoto 1983; O'Sullivan and Bourgoyne 1990; Uchiyama,
Morita, Kitayama, Suemitsu, Minami, Miyasako and Dohi 1994) and corticosterone
(Adams, Nock, Truong and Cicero 1992; Cameron and Hinson 1993).
NO is, however, atypical transmitter in many ways and therefore differ from
52

hitherto known neurotransmitters. Unlike the classical mediators and the neuropeptides
which are frequently stored in vesicles and released specifically (see Bumstock 1986;
Kupfermann 1991), NO is not stored but diffuses freely from its site of formation
without any specialized release machinery (see Garthwaite 1991; Schuman and
Madison 1994). Being a free radical molecule it permeates membrane and pass normal
signal transduction routes without interaction with synaptic membrane receptors and
forms covalent bonds fairly easily with its target molecules (Stamler et al. 1992;
Stamler 1994), in contrast to the classical transmitters and the neuropeptides which
have complex structures and depend for their action on a complimentary fit to a
specific receptor. The action of conventional neurotransmitter is terminated by
presynaptic reuptake or enzymatic degradation, whereas the action of NO is
presumably terminated by diffusion away from its targets as well as by forming
covalent linkage to the superoxide anion or scavenger proteins (Stamler 1994). NO
acting as a covalently reactive redox type mediator may not be the only one of its type,
and may be the first example of a completely new signalling molecule. It has already
been suggested that CO, another activator of guanylate cyclase could be a neuronal
messenger (Verma, Hirsch, Glatt, Ronnett and Snyder 1993).
1:9. Plasticity in adrenal gland innervation and expression of neuromediators
1:9:1. During development and aging
As shown in the rat, the developmental expression of various mediators found in
the adrenal gland is not synchronous. In the developing adrenal chromaffin cells,
noradrenergic expression, immunoreactivity for TH and DBH, is evident by embryonic
day (E)12.5, while adrenergic expression, immunoreactivity for TH, DBH and PNMT,
is not observed prior to E17 (Bohn, Goldstein and Black 1981). During late foetal life,
all developing adrenal medullary chromaffin cells of the rat achieve adrenergic
expression, and solely noradrenaline expressing cells start reappearing by 2-3 days
53

postnatally (Verhofstad, Coupland, Parker and Goldstein 1985). The relative
concentration of normal adult proportion of about 80 % adrenaline and 2 0 %
noradrenaline is reached by the 4^ ̂ day of postnatal life (Verhofstad et al. 1985).
Immunoreactivity for NPY and Leu-enkephalin in the developing adrenal medullary
cells of rat begin to appear at E15 and E16, respectively (Henion and Landis 1990).
The proportions of adrenal chromaffin cells expressing met-enkephalin, CORF and
neurotensin during the first week are higher than those found in the adult (Holgert,
Dagerlind, Hokfelt and Lagercrantz 1994). NPY and galanin are expressed in
equivalent proportions of chromaffin cells in the glands from developing and adult rats
(Holgert et at. 1994).
The development of adrenal innervation begins during foetal life, although on
birth, functional innervation depends on the state of maturity of the neonate (Slotkin
1986). Ultrastracturally, synaptic profiles associated with the adrenal medullary
chromaffin cells of foetal rats are observed as early as 15.5 days of gestation (Daikoku,
Kinutani and Sato 1977). However, as found during the first postnatal week, most of
the synaptic profiles on the chromaffin cells are immature, although, they account for
about 50 % of those found in the adult rat (Tomlinson and Coupland 1990).
Pharmacological studies have also revealed that the innervation of adrenal medulla of
rat is non-functional until about the end of the first week, and becomes mature by 1 0
days after birth (Slotkin, Smith, Lau and Bareis 1980; Slotkin, Chantry and Bartolomé
1982). However, a neuronal connection between the adrenal medulla and the spinal
cord is already established by 2 days after birth (Parker, Kesse, Tomlinson and
Coupland 1988).
As shown with immunohistochemical studies, bundles of nerve fibres labelled
with neurofilament 160 kDa are visible in the embryonic adrenal medulla as early as 15
days of gestation (Henion and Landis 1990). Immunoreactive nerve terminals
associated with the chromaffin cells are, however, found only postnatally by the 2 ^^
day for neurofilament 160 kDa and choline acetyl transferase (ChAT), and by the 5^^
54

day for leu-enkephalin (Henion and Landis 1990). Intra-adrenal nerve fibres
immunoreactive for CGRP and galanin are found fully developed by day 2 after birth
(Holgert et al. 1994). However, intra-adrenal nerve fibres immunoreactive for ChAT,
leu-enkephalin and acetylcholinesterase (AChE) increase during the early postnatal
period and reach to that of adult levels by days 11-12, 14 and 16, respectively (Henion
and Landis 1990; Holgert et al. 1994). Met-enkephalin-immunoreactive nerve fibres
were found only by the 1̂ ̂ (Holgert et al. 1994) or the 2”^ week (Kent and Coupland
1989) after birth and increased afterwards throughout the developmental period
(Holgert et al. 1994).
There are a few studies carried out on the state of innervation and expression of
neuromediators in the adrenal gland during aging. An increased sympathoadrenal
activity associated with elevated plasma catecholamine levels was reported by Ito, Sato
and Suzuki (1986). Reduced numbers of AChE-positive nerve fibres were described in
the nodular hyperplastic regions that occur in aging Long-Evans rat (Tischler et al.
1985). However, studies by Tomlinson and Coupland (1990) did not reveal any change
in the frequency of synaptic profiles associated with the chromaffin cells of aging
Wistar rat from that observed in young adult rat.
1:9:2. During stress
Varieties of stress stimuli such as immobilization, cold, swimming, electric
shock and insulin administration activate secretion of glucocorticoids and
catecholamines from the adrenal gland. They also produce changes in the activities of
the catecholamine biosynthetic enzymes (Kvetnansky, Weise and Kopin 1970;
Kvetinansky, Sun, Lake, Thoa, Torda and Kopin 1978; Axelrod and Reisine 1984;
Stachwiak, Fluharty, Strieker, Zigmond and Kaplan 1986; Sietzen, Schober, Fischer-
Colbrie, Scherman, Sperk and Winkler 1987). Biosynthesis and secretion of peptides
from the gland have also been found influenced by stress (Sietzen et al. 1987; Fisher-
55

Colbrie, lacangelo and Eiden 1988).
1:9:3. After selective surgical denervations
1:9:3:1. Extrinsic adrenal denervation
As the synthesis and secretion of adrenal medullary hormones are mainly
regulated transynaptically via the splanchnic nerves (Thoenen, Muller and Axelrod
1969; Axelrod 1971; Thoenen 1975; Zigmond 1985), transection of these nerves
causes derangements in the biosynthetic and secretory activities of the gland.
Splanchnic nerve section has been found to cause small but significant decrease in the
levels of mRNAs for TH, PNMT and NPY (Schalling, Franco-Cereceda, Hemsen,
Dagerlind, Seroogy, Persson, Hokfelt and Lundberg 1991). It also causes a decrease in
the catecholamine content of the adrenal medulla in cat and rat (Vogt 1952; Fleminger,
Lahm and Udenfriend 1984). In contrast, splanchnic nerve transection increases the
number of immunoreactive chromaffin cells for enkephalin, CGRP and neurotensin in
the adrenal medulla of rat (Schultzburg et al. 1978; Lewis, Stem, Kilpatrik, Gerber,
Rossier, Stein and Udenfriend 1981; Flemingr et al. 1984; Kilpatrick, Howells,
Fleminger and Udenfriend 1984; Pelto-Huikko 1989; Dagerlind, Pelto-Huikko,
Lundberg, Ubink, Verhofstad, Brimijoin and Hokfelt 1994).
Since the majority of adrenal medullary nerve fibres degenerate upon splanchnic
nerve section (Elliott 1913; Hoshi 1926; Hollinshead 1936; Swinyard 1937; McFarland
and Davenport 1941), the occurrence of changes in the intra-adrenal nerve fibres
immunoreactive for various mediators is also expected. Indeed, a marked loss of nerve
fibres immunoreactive for AChE, and total loss of those immunoreactive for
enkephalin, CGRP and SP from rat adrenal gland have been found after splanchnic
nerve section (Pelto-Huikko 1989; Dagerlind et al. 1994). In contrast, ligation of the
splanchnic nerve in rat, results in an increased intra-adrenal VIP-immunoreactive
nerve fibres and cell bodies (Holzwarth 1984). Splanchnic nerve transection has also
been shown to increase the number of galanin-immunoreactive ganglion cells found in
56

the rat adrenal medulla (Dagerlind et al. 1994).
1:9:3:2. Intrinsic adrenal denervation
Following adrenal demedullation which causes intrinsic denervation, a marked
loss of VIP-immunoreactive nerve fibres occurs from the zona glomerulosa of rat
adrenal cortex (Holzwarth 1984). Reduction in the NPY-immunoreactive nerve fibres
forming a plexus in the capsule and zona glomerulosa of rat adrenal gland also follows
after adrenal demedullation (Maubcrt, Dopouy and Bemet 1993).
1:9:3:3. Hypophysectomy
Hypophysectomy results in a major atrophy of adrenal cortical cells of zonae
fasciculata and reticularis accompanied with diminished glucocorticoids secretion
(Deane and Creep 1946; Sayers 1950). The activities of the catecholamine synthesizing
enzymes TH, DBH and PNMT are also reduced (Wurtman and Axelrod 1966; Mueller,
Thoenen and Axelrod 1970; Weinshilboum and Axelrod 1970; Sietzen et al. 1987).
However, indicating that an intact pituitary-adrenocortical axis is not a prerequisite for
the synthesis of these enzymes, the neurally mediated induction of the enzymes is still
possible in hypophysectomized animals (Thoenen et al. 1969; Ciaranello, Wooten and
Axelrod 1976).
No change occurs in the peptide levels of enkephalins and NPY as well as their
mRNAs after hypophysectomy (Sietzen et al. 1987; Yobum, Franklin, Calvano and
Inturrisi 1987; Fisher-Colbrie et al. 1988). However, it has been shown that the
denervation-induced increase in adrenal enkephalin synthesis is glucocorticoid-
dependent, since the increase in enkephalin is markedly reduced or even totally
abolished by hypophysectomy and can be restored by corticosterone and
dexamethasone treatment (Yobum et al. 1987).
57

1:9:4. During chronic exposure to drugs
1:9:4:1. Reserpine
An enhanced adrenal catecholamine synthesis and secretion occurs after
administration of reserpine (Thoenen et al. 1969, Molinoff, Brimijoin, Weinshiloum
and Axelrod 1970). It increases mRNAs for TH and NPY, but decreases that of PNMT
in the rat adrenal chromaffin cells (Schalling, Dagerlind, Brené, Hallman, Djurfeldt,
Persson, Terenius, Goldstein, Schlesinger and Hokfelt 1988a). Reserpine also causes
an increase in the peptide levels of met-enkephalin, CGRP and NPY in the rat adrenal
medullary cells (Wilson, Abou-Donia, Chang and Viveros 1981; Bohn, Kessler,
Golightly and Black 1983; Seitzen et at. 1987; Hofle, Weiler, Fisher-Colbrie, Humpel,
Laslop, Wohlfarter, Hogue-Angeletti, Saria, Flemong and Winkler 1991).
1:9:4:2. 6 -Hydroxydopamine
Administration of 6 -Hydroxydopamine (6 -OHDA) causes an increased
synthesis and turnover of adrenal catecholamines (Mueller, Thoenen and Axelrod
1969). It causes transynaptic induction of TH and DBH (Muller et at. 1969; Molinoff
et at. 1970). Injection of 6 -OHDA in rat during early foetal and neonatal period has
been shown to cause a substantial reduction in the number of intra-adrenal ganglion
cells, although, the chromaffin cells remained unaffected (Aloe and Levi-Montalcini
1980).
6 -OHDA administered to adult rat causes an increase in the number of CGRP-
immunoreactive adrenal chromaffin cells, but decrease in the staining intensity of those
immunoreactive for NPY (Pelto-Huikko 1989). Furthermore, treatment of neonatal
rats with 6 -OHDA results in elimination of the catecholaminergic nerve fibres from
the capsule and the zona glomerulosa of adrenal gland (Kleitman and Holzwarth 1985).
In contrast, 6 -OHDA does not cause any change in the rat adrenal nerve fibres
immunoreactive for enkephalin, NPY or CGRP (Bohn et at. 1983; Pelto-Huikko and
Salminen 1987; Pelto-Huikko 1989).
58

1:9:4:3. Guanethidine
A marker reduction in the number of adrenal chromaffin and ganglion ceils in
the adrenal medulla of 14 day-old rats which were exposed to guanethidine during
foetal and neonatal period have been reported by Aloe and Levi-Montalcini (1980).
However, no such effect was observed in the adrenal medulla of adult rats treated with
guanethidine as neonate or adult (Angeletti and Levi-Montalchini 1972; Johnson and
O'Brien 1976). Guanethidine administered to neonatal rats has been shown to eliminate
the catecholaminergic nerve fibres from the capsule and the zona glomerulosa of
adrenal gland (Kleitman and Holzwarth 1985).
1:9:4:4. Capsaicin
When capsaicin is administered to both neonatal and adult rat and hamster, the
number of CGRP-immunoreactive chromaffin cells show an early increase followed
by a gradual decrease to the normal levels (Pelto-Huikko 1989). It also causes a
decrease in the staining intensity of enkephalin and NPY immunoreactivity in the
chromaffin cells of rat adrenal medulla (Pelto-Huikko 1989). In addition,
administration of capsaicin into neonatal rat eliminates the few galanin-
immunoreactive chromaffin cells (Pelto-Huikko 1989). Substantial reduction in the rat
adrenal nerve fibres immunoreactive for CGRP occurs after capsaicin treatment to both
neonatal and adult rats (Pelto-Huikko 1989). However, capsaicin does not have any
effect on nerve fibres immunoreactive for neurotensin, NPY and enkephalins (Bohn et
al. 1983; Pelto-Huikko 1989).
1:9:5. In diabetes mellitus
Diabetes mellitus is a disease complication caused by a relative or complete lack
of insulin secretion by the ^6-cells of the pancreas or of defects of the insulin receptors.
59

There are two major forms of diabetes which are known as insulin-dependent diabetes
mellitus (IDDM or type I diabetes) and non-insulin-dependent diabetes mellitus
(NIDDM or type II diabetes) (Keen and Ng Tang Fui 1982). The former is
characterized by an absolute deficiency of insulin and massive ^5-cell lesions and
necrosis. NIDDM is characterized by significant insulin production, varying from less
than normal to above normal but always in quantities insufficient to maintain glucose
homeostasis, and by target-organ resistance to insulin. One of the complications
associated with diabetes mellitus is diabetic neuropathy.
The adrenal gland in diabetic rat is enlarged (Bennett and Knoeff 1946;
Rebuffat, Belloni, Malendowicz, Mazzocchi, Gottardo and Nussdorfer 1988b;
Rebuffat, Belloni, Malendowicz, Mazzocchi, Meneghelli and Nussdorfer 1988c). This
is mainly associated with hypertrophy of the zona fasciculata, although, in contrast,
there is also atrophy of the zona glomerulosa of cortex (Rebuffat et al. 1988b, c).
Furthermore, there is occurrence of adrenal medullary fibrosis in long standing human
patients with IDDM (Brown, Zuckerman, Longway and Rabinowe 1989; Brown,
Smith, Longway and Rabinowe 1990). Reduction in the number of adrenal medullary
preganglionic nerve fibres in diabetic rats has also been reported (Wilke, Riley, Lelkes
and Hillard 1993).
Derangements in the secretion from the adrenal gland occurs in diabetes. There
is an increased levels of plasma cortisol in diabetic man (Lentle and Thomas 1964;
Cameron, Thomas, Tiongco, Hariharan and Greden 1987; Tsigos, Young and White
1993). Similarly, increased adrenal and plasma levels of corticosterone occurs in
diabetic rat (De Nicola, Fridman, Del Castillo and Foglia 1976; Torenello, Coirini and
De Nicola 1981; Rebuffat etal. 1988b). In contrast, adrenaline secretion in response to
insulin-induced hypoglycemia is diminished in diabetic man (Hilsted 1982; Hoeldtke,
Boden, Shuman and Owen 1982; Cryer 1989). Decrease in adrenaline secretion also
occurs in diabetic rat (Patel 1983; Wilke et at. 1993).
Other additional conditions in which a number of reports have indicated the
60

occurrence of changes in the adrenal glandular activity include hypertension (McCarty,
Chiueh and Kopin 1978; Kvetnasky, McCarty, Thoa, Lake and Kopin 1979; Sato, Sato
and Suzuki 1987) and pregnancy (Watanabe, Meeker, Gray, Sims and Solomon 1968;
Rosenthal, Slaunwhite and Sandberg 1969; Ehrlich, Nolten, Oparil and Lindheimer
1976; Natrajan, McGarrigle, Lawrence and Lachelin 1982). It is also apparent that the
adrenal secretory activity is affected during diseases such as Phaeochromocytoma and
Addison's disease, as well as Cushing's, Conn's and adrenogenital syndromes which
directly affect the adrenal gland.
61

SECTION 2
MATERIALS AND METHODS
62

2:1. Animals used in this study
For studies involving the distribution of NOS immunoreactivity in the adult rats
reported in Section 3:1:1 and that of diabetic experiments reported in Section 3:2:2
Wistar rats were used, while all remaining studies were carried out on Sprague-
Dawley rats. All studies on postnatal rats were carried out on male rats, while either
sex of the prenatal rats were used. Sprgue-Dawley rats for the studies on the effects of
reserpine treatment and hypophysectomy, and all Wistar rats were obtained from
Charles River UK Ltd (Morgate, UK). All remaining Sprgue-Dawley rats were
supplied by the animal breeding unit of the Biological Services at the University
College London (London, UK).
2:2. Maintenance of the animals
Animals were maintained in the Animal House Unit of the Biological Services
at the University College London. The animal house had normal conditions of 21 °C
temperature, 12 h light and 12 h darkness. The animals were fed standard rat chow, and
had access to water ad labitum.
2:3. Tissue processing
2:3:1. Preparation o f adrenal sections
Animals were killed by asphyxiation with carbon dioxide or ether. Adrenals
were quickly dissected out, and in order to facilitate fixation, the adrenal glands
(except those from the early developing rats) were bisected. The adrenals from the
early developing rats, which were small in size, were fixed intact. Fixation was done
by immersion of the adrenals in 4 % paraformaldehyde (TAAB, Reading, UK) in 0.1
M phosphate buffered saline (PBS), pH 7.4, for 2 h at room temperature. After fixation,
the adrenal glands were cryoprotected by leaving them in 15 % sucrose in PBS
63

overnight, at 4°C.
The adrenals were then embedded in OCT, Tissue-Tek compound (Miles Inc,
Elhart, USA) and frozen onto specimen blocks by gradually immersing in iso-pentane
precooled at -150°C. Frozen tissue blocks were stored in liquid nitrogen at -190°C
until used. Sections were cut at 14 fjm in a cryostat (Reichert-Jung, Germany) at
cutting temperature of -24°C, and thaw-mounted onto 0.1 % poly L-lysine (sigma,
Poole, UK) coated glass slides, and then stored in the dark at 4°C until processed
within 1 week.
2:3:2. Immunohistochemistry
Introduction
Immunohistochemistry, introduced by Coons and his colleges (Coons, Creech
and Jones 1941; Coons and Kaplan, 1950) is a method by which cell and tissue
constituents are made visible in situ by the application of antigen-antibody reaction.
The tissue constituent acts as an antigen and its site is visualized by the application of
its antibody conjugated to various labels such as fluorescent compounds, enzymes or
ferritin or colloidal gold particles (see Polak and Van Noorden 1987; Beltz and Burd
1989). The original approach was the direct or one step method, where the label is
conjugated straight to the antibody directed against the antigen (Coons et al. 1941).
Subsequently, the indirect method was introduced by the inclusion of additional step(s)
(Coons and Kaplan 1950). When a secondary antibody directed against unlabelled
primary antibody is conjugated with a label and added as a second layer, the method is
called two step method. A three step method involves the addition of a tertiary labelled
antibody as a third layer directed against unlabelled secondary antibody.
The indirect method has been modified to incorporate avidin and biotin into the
system in order to obtain the advantage of the very high affinity of avidin for biotin.
Avidin is a large glycoprotein from egg white and some bacteria. It has four binding
sites per molecule for biotin which is a small vitamin molecule found in egg yolk. In
64

this method, biotin is conjugated to a secondary antibody, directed against the
immunoglobulin G of the host in which the primary antibody was raised. This is
followed by incubation with avidin coupled with a label. Strcptavidin, which is derived
from Streptococcus avidini and behaves in a similar way to avidin could also be used
as a substitute for avidin (Coggi, Dell'Orto and Viale 1986). For its high sensitivity and
efficiency, the biotin-streptavidin method was used in the present study.
Experimental protocols
Sections were rinsed in PBS containing 0.1 % Triton X-100 for 5 min to
facilitate antibody penetration, and arranged in a humidified incubating chamber. In
each chamber sections from experimental and their respective controls were processed
in parallel. Sections were incubated by covering them with the respective primary
antisera at the optimum dilution in PBS (pH 7.4) containing 0.1 % Triton X-100, 0.1
% lysine, 0.01 % bovine serum albumin and 0.1 % sodium azide. After 18 h of
incubation at room temperature, sections were washed 3 times for 5 min each with PBS
and incubated with biotinylated donkey anti-rabbit antibody (Amersham Ltd,
Amersham, UK) at a dilution of 1:250 for 1 h. Sections were washed ( 3 x 5 min) with
PBS and incubated with streptavidin fluorescein (Amersham Ltd) at a dilution of 1:100
for 1 h, washed in PBS and then mounted with citifluor mountant (City University,
London, UK).
Replacements of the primary antibodies with preimmune serum were used as
negative controls to establish the specificity of each of the antiserum. Furthermore,
replacements of the primary antibodies with non-immune serum and/or omissions of
the primary antibodies were served as a routine negative controls each time the
immunohistochemistry was carried out.
65

2:3:3: NADPH-diaphorase histochemistry
Introduction
NADPH-diaphorase histochemistry is an enzyme histochemicai method which
employs the principle of dehydrogenase histochemistry (Farber, Stemberger and
Dunlap 1956; Nachlas, Walker and Seligman 1958; Pearse 1972). Dehydrogenases are
enzyme systems which bring oxidation of organic compounds by transferring the
hydrogen atoms to another organic molecules, generally thought of as the coenzymes
(cofactors) in the system. In dehydrogenase histochemistry the reduced cofactors are
used to reduce in turn a dye which is visible as an indication for the presence of the
enzyme either directly or via a second enzyme known as a diaphorase. NADPH-
diaphorase is one of such enzymes that utilizes the cofactor NADPH. Recently, studies
have found that neuronal NADPH-diaphorase is NOS (Dawson et al. 1991; Hope,
Michael, Knigge and Vincent 1991).
In NADPH-diaphorase histochemistry, as in dehydrogenase histochemistry,
various tétrazolium salts whose redox potentials are suitable to be easily reduced by
certain enzymes may be used as final substrate (i.e., dye) for localization of the site of
the enzyme. In their oxidized form tétrazolium salts are colourless, while upon
reduction they precipitate to give coloured water insoluble compounds called
formazans. Nitroblue tétrazolium chloride is the most commonly used tétrazolium salt
employing the light microscopic study (Tsou, Cheng, Nachlas and Seligman 1956). It
readily accepts electrons to be reduced and give formazan of good pigment quality as
amorphous red-blue coloured dye. The formazan is insoluble in aqueous and common
organic solvents. In addition, it is resistant to oxidation by molecular oxygen back to
tétrazolium. It is therefore, this tétrazolium salt that has been utilized as a marker for
the present NADPH-diaphorase histochemicai study.
Experimental protocols
Cryostat sections, were rinsed in 0.1 M TRIS HCl buffer, pH 8 for 5 min and
66

incubated in a freshly prepared incubating medium for 30 min at 37°C, in a humidified
chamber. The incubating medium contained: 1 mg/ml ^-NADPH (Sigma), 0.2 mg/ml
nitroblue tétrazolium (Sigma), 2.7 mg/ml L-malic acid (Sigma) and 0.1 % Triton X -
100 in 0.1 M TRIS HCL buffer, pH 8 . The reaction was stopped by rinsing in 0.1 M
TRIS HCl buffer followed by several washes with distilled water. Sections were then
mounted with citifluor mountant.
In control experiments, in order to test for endogenous reduction in the tissue
which may convert the nitroblue tétrazolium salt to the formazan product, one set of
section was incubated in media devoid of ^-NADPH. Another set of section was
incubated with 0.1 M TRIS HCl buffer heated at 75°C for 10 min before incubation in
order to test the heat stability of the NADPH-diaphorase activity. Such procedures
should give negative reaction (Hope and Vincent 1989).
2:4. Microscopy and computer assisted image analysis
Immunoprocessed and NADPH-diaphorase stained tissue sections were
analyzed with a Zeiss photomicroscope (Carl Zeiss, Inc., Thomwood, New York)
equipped with both epifluorescence and appropriate filters as well as ordinary light
sources. Immunolabelled cells and nerve fibres were studied and photographed with
Kodak, T-max black-and-white film (ASA 3200; Kodak, Rochester, New York).
Similarly sections processed for NADPH-diaphorase histochemistry were studied and
labelled cellular elements were photographed with Kodak T-max black-and-white
film (ASA 100; Kodak, England).
When required, measuring of the labelled cell bodies was achieved by aligning
the long and short axes of the labelled cell bodies with clear nuclear outlines
perpendicular to each other to calibration bars fitted to an eye-piece. From the long (1)
and short (s) axes a mean diameter (d) of the labelled cells was calculated as d = VI x s
(see Neuhber, Sandoz and Fryscak 1986). Indication to the shape of the labelled cell
67

bodies were obtained from the ratio of the long to the short axes.
Quantitative analysis of the immunoreactive nerve fibres for the studies reported
in Section 3:2:1 was made using a computer image analyzer (Seescan Imaging Ltd.,
Cambridge, UK) connected to a Zeiss photomicroscope via a video camera (Sony,
Japan). With a x l 6 objective regions of adrenal gland in every 10^^ consecutive
sections were randomly selected and analyzed in both the medulla as well as the
subcapsular region and zona glomerulosa of the cortex. The frame area of measured
region in the medulla was always kept constant to the maximum possible view on the
screen monitor, and the analyzed regions had comparable medullary tissue and were
free of labelled neuron. In the adrenal cortex, a smaller standard measuring area was
set to include only the subcapsular region and zona glomerulosa, and no region of zona
fasciculata or reticularis was included.
The levels of sensitivity and light intensity were adjusted until optimum at the
beginning of each set of the analysis and then kept constant throughout the subsequent
measurements. Values have been expressed as mean ± standard deviation of the
percentage of the ratio of positive to negative immunofluorescence for each
experiment.
2:5. Biochemical assay of nitric oxide synthase activity
Introduction
A radiometric assay has been developed for measuring the activity of NOS in
the neurons of both central and peripheral neural tissue (Bredt and Snyder 1990;
Schmidt et al. 1991; Lincoln and Messersmith 1995). The assay is based on the
conversion of [^H]-labelled arginine to [^H]-labelled citrulline, in the presence of
NADPH, BH4 , FAD, CaClo and calmodulin at optimum concentrations and
conditions. In order to permit optimal and reproducible enzyme activity the
composition of the buffer utilized for homoginization and/or incubation includes
EDTA, dithiothreitol, leupeptin, soybean trypsin inhibitor and aprotonin. EDTA binds
68

divalent cations which may inhibit the enzyme activity. Dithiothreitol protects against
oxidative damage of the enzyme. Leupeptin, soybean trypsin inhibitor and aprotonin
inhibit proteinase activity, thereby protect the enzyme.
The reaction is terminated by the addition of Dowex ion exchange resin which
binds any residual arginine leaving the newly formed citrulline in the supernatant
which is transferred into liquid scintilants for counting the radioactivity and then
corrected to dpm. In order to remove any activity due to artifacts, blanks such as the
assay of the homogenizing medium alone are also carried out in identical process. The
radioactivity of the blanks converted into dpm is subtracted from that of the samples
and the residual dpm is then converted into enzyme activity using the specific activity
of the L-arginine used.
Experimental protocols
The biochemical assay of NOS activity was used for the studies reported in
Section 3:2:2. Adrenal glands were dissected and stored in liquid nitrogen until assay.
When required, the adrenal glands were weighed and homogenized in 10 mM HEPES
buffer (pH 7.4) containing 0.32 M sucrose, 0.1 mM EDTA, 1 mM dithiothreitol, 10
pg/ml leupeptin, 10 ^g/ml soybean trypsin inhibitor and 2 jug/m\ aprotonin to give a
tissue concentration of 50 mg/ml. The homogenate was centrifuged at 12,000 g for 10
min and NOS activity was measured in the supernatant.
The control incubating medium was 50 mM HEPES (pH 7.4) containing 1 mM
EDTA, 1 mM dithiothreitol, 1 mM NADPH, 1.25 mM CaClo, 10 yt/g/ml calmodulin, 10
pM FAD and 100 pM BH4 . The [^H]-arginine solution was prepared by diluting
[^H]-arginine with non-radioactive arginine to give a final arginine concentration of
10 pM with a specific activity of 4.05 Ci/mmol. 25 p\ of each supernatant was
equilibrated with 100 p[ of the incubating medium at 37®C. The reaction was started by
the addition of 25 p\ of [^H]-arginine solution. After 10 min at 37®C, the reaction was
69

Liquid scintilation counting was carried out using a Beckman LS-600IC liquid
scintiiation counter. This equipment is fitted with automatic quench calibration using
the H number method. The percentage efficiency of counting was therefore measured
for each sample individually and the cpm corrected to dpm.

stopped by the addition of 2 ml of Dowex ion-exchange resin in water (1:1) followed
by vortex mixing. The tubes were stored on ice and the resin allowed to settle. The
supernatants were transferred to scintillation vials containing 5 ml Optiphase "HiSafe”
3 for liquid scintillation counting. ; :. ̂ Blanks
consisted of the homogenizing medium, which was taken through an identical process.
NOS activity was further characterized by carrying out the assay in the presence of the
following inhibitors: L-NAME (1 mM); BGTA (2 mM). All samples were assayed in
triplicate and corrected for the blanks. NOS activity was calculated as nmol citrulline
formed/h/gland and expressed as total activity, L-NAME-sensitive activity, and
EGTA-sensitive activity.
2:6. Surgical procedures
2:6:1. Extrinsic adrenal denervation
Introduction
In the rat, the extrinsic nerve fibres to the adrenal gland come through the
greater and lesser splanchnic nerves which join at the suprarenal ganglion, found as a
lateral extension of the coeliac plexus (Baljet and Drukker 1979). Several nerve trunks
branch from the suprarenal ganglion and enter the adrenal gland. Denervation of the
extrinsic nerve fibres to the gland is therefore, best achieved by impairing the nerve at
the suprarenal ganglion or proximal to it before it gives off the small adrenal branches.
Experimental protocols
Rats were anaesthetized by intramuscular injection of fentanyl-fluanisone
(Hypnorm; Janssen, Oxford, UK; 3 mg/kg body wt) and subcutaneous injection of
diazepam (Valium; Phoenix Pharmaceutical Ltd, Gloucester, UK; 2 mg/kg body wt).
The loss of the stretch reflex in the hind limb was taken as indicative of the level of
anaesthesia.
Anaesthetized rats were placed in the supine position over an operating board.
70

The skin over the left flank was shaved and swabbed with 70 % ethanol. An L-shaped
incision was then made through the skin and muscle separately and reflected laterally
and held with artery forceps. The stomach, intestine and spleen were gently moved
laterally onto warm saline soaked cotton gauze placed over the reflected muscle flap,
and covered with similar saline soaked cotton gauze. Under an operating binocular
microscope the left splanchnic nerve along with the suprarenal ganglion was transected
using a pair of microscissors and removed. The stomach, intestine and spleen were
carefully placed back to their original positions, and the muscle and the skin incisions
were separately sutured with continuous suture using 3/0 chromic catgut, 16 mm
cutting (Ethicon Ltd, Edinburgh, UK). Animals were then resuscitated and kept in
cages bedded with shredded tissue papers.
For sham control studies, in age-matched rats the left splanchnic nerve along
with the suprarenal ganglion were exposed in a similar manner as above, but no nerve
was cut.
2:6:2. Intrinsic adrenal denervation
Introduction
As the intrinsic adrenal neurons are found intermingled with the other cells of
the gland, intrinsic surgical denervation of the gland is difficult and is impossible to
perform without damaging the adrenal medulla as well. A method utilized for such
denervation takes advantages of the fact that if the adrenal gland is bilaterally
enucleated, the cortex will regenerate while the medulla and the intrinsic neurons
remain unregenerated (Ingle and Higgins 1938; Creep and Deane 1949). Such method
of intrinsic adrenal denervation in association with studies from extrinsic adrenal
denervation is a valuable approach to determine source of the nerve fibres found in the
gland with regard to their extra-adrenal or intra-adrenal origin (see for example,
Holzwarth 1984).
71

Experimental protocols
Rats were anaesthetized as described in Section 2:6:1, and placed in the prone
position over an operating board. The skin of the back around the midline at the region
of the lower throracic and upper lumbar vertebrae was shaved and swabbed with 70 %
ethanol. A midline incision was made through the skin from the level of the eighth
thoracic to the fifth lumbar vertebrae. The left skin flap was reflected laterally and held
with artery forceps. A small vertical parasagittal incision was made on the muscle
below the last rib and retracted laterally with an artery forceps. With the help of a
cotton bud, the left adrenal gland was gently manipulated from the surrounding adipose
tissue found in the retroperitoneal space and gently grasped with small forceps. The
capsule at the posterior aspect of the gland was slit with a pair of microscissors, and by
a gentle squeeze with small curved forceps the content of the gland was extruded
intact. The capsule of the enucleated gland, with the pedicle of the gland untouched
was then restored to position. The muscle incision was closed by a continuous suture
using 3/0 chromic catgut, 16 mm cutting.
A similar procedure was repeated on the right side to expose and enucleate the
right adrenal gland. Finally, the skin incision was sutured using 3/0 chromic cutgut, 16
mm cutting. Throughout the operation the exposed tissue was kept moist by warm
physiological saline solution. Animals were resuscitated and kept in a cage bedded
with shredded paper.
For sham operation, the adrenal glands in age-matched control rats were
exposed and manipulated in a similar manner as above, except that the capsules were
not slit nor the adrenals were enucleated.
2:6:3. Hypophysectomy
Hypophysectomized, sham-hypophysectomized and age-matched non
operated rats were obtained from Charles River UK Ltd. Hypophysectomized rats
72

gained less body weight as compared to their age-matched unoperated and sham
operated rats.
2:7. Drug treatment
2:7:1. Reserpine
Introduction
Administration of reserpine into animals causes depletion of catecholamines
from neurons and adrenal chromaffin cells (Brodie, Pletscher and Shore 1955; Carlsson
1965; Thoenen et al. 1969). It also activates the splanchnic nerves (Haeusler 1974;
Pemow, Thorén, Millberg and Lundberg 1988). Reserpine can therefore be used as an
agent in studies involving depletion of adrenal catecholamines and/or splanchnic nerve
stimulation (see fore example, Sietzen et al. 1987; Schalling et al. 1991).
Experimental protocols
Rats were subcutaneously injected with 2.5 mg/kg body wt of reserpine (Sigma;
10 mg dissolved in 30 p\ glacial acetic acid, 0.25 ml propylene glycol, 0.25 ml ethanol,
1.47 ml HoO [Sietzen et al. 1987]) on day 1, 3, 5 and 7, and killed on day 8 . Age-
matched control rats were injected with the solvent alone.
2:7:2. Guanethidine and 6 -hydroxydopamine
Introduction
Prolonged administration of guanethidine both to newborn and adult rats
produces an irreversible destruction of the cell bodies of sympathetic neurons (Jensen-
Holm and Juul 1970; Bumstock, Evans, Gannon, Heath and James 1971; Eranko and
Eranko 1971; Angeletti and Levi-Montalcini 1972; Johnson and O'Brien 1976).
Similarly, injection of 6 -hydroxydopamine (6 -OHDA) into the newborn animals
results in selective and permanent destruction of cell bodies in sympathetic ganglia
73

(Angeletti and Levi-Montalcini 1970). However, administration of 6 -OHDA into
adult animals results i n destruction of adrenergic nerve terminals while cell bodies in
the sympathetic ganglia are resistant, and therefore regeneration of the adrenergic
terminals follows after sometime (Thoenen and Tranzer 1968; Tranzer and Thoenen
1968; Bennett, Bumstock, Cobb and Malmfors 1970; Furness, Campbell, Gillard,
Malmfors, Cobb and Bumstock 1970; De Champlain 1971; Jonsson and Sachs 1972).
For these reasons, guanethidine is usually utilized for studies involving chronic
sympathetic denervation, while 6 -OHDA is employed for studies involving acute
denervation of the sympathetic nerve terminals in adult animals.
Experimental protocols
Litter rats were injected subcutaneously with 50 mg/kg body wt of guanethidine
sulphate (Ismelin; Ciba Laboratories, Horsham, UK) from day 8 after birth for 3 weeks,
5 days per week, and killed at 12-14 weeks of age (Aberdeen, Corr, Milner, Lincoln
and Bumstock 1990). Age-matched control rats were injected with saline alone.
Another rats which were 13 weeks old were subcutaneously injected with 6 -OHDA
(Sigma, Poole, UK) dissolved in sterile saline containing 1 mg/ml ascorbic acid, at
doses of 100 mg/kg body wt on day 1 and 250 mg/kg body wt on day 2, and killed on
day 8 (Aberdeen et al. 1990). Age-matched control rats were injected with the solvent
alone.
2:7:3. Capsaicin
Introduction
Administration of capsaicin into adult animals causes desensitization for
noxious chemical stimuli which lasts for weeks or months (Jancso 1968). In newbom
rats, administration of capsaicin brings about destmction of the jS-type peptide
containing primary afferent neurons with loss of specific sensory functions (Jancso,
Kiraly and Jancso-Gabor 1977; Gamse, Holzer and Lembeck 1980; Nagy, Hunt,
74

Iversen and Emson 1981; Jancso, Hokfelt, Lundberg, Kiraly, Halasz, Nilsson,
Terenius, Rehfeld, Steinbush, Veihofstad, Hide, Said and Brown 1981; Lawson 1987).
Neonatal treatment of capsaicin is therefore utilized for studies aimed at denervation of
such sensory nerves.
Experimental protocols
Neonatal rats were subcutaneously treated with 50 mg/kg body wt of capsaicin
(Sigma, Poole, UK) dissolved in saline containing 10 % Tween 80 and 10 % ethanol at
1 , 2, 3, 5, 7 and 14 days of age, and killed at 14 weeks (Mione, Cavanagh, Kirkpatrik
and Bumstock 19992). For control study litter-matched neonates were injected with
same volume of vehicle. Ice anaesthesia was used for injections of the 1 and 2 days old
rats.
2:8. Induction of diabetes
Introduction
There are various experimental as well as spontaneous animal diabetic models
which are utilized to study diabetes (Rerup 1970; Grodsky, Anderson, Coleman,
Craighead, Gerritsen, Hansen, Herberg, Howard, Lemmark, Matschinsky, Rayfield,
Riley and Rossini 1982; Mordes and Rossini 1985). In the experimental animal models
the most successful method of provoking diabetes is by use of chemical agents, such as
alloxan and streptozotocin which exert toxicity on the ^-cells of pancreas. In rat, the
specificity of streptozotocin on ^-cells of pancreas is superior to that of alloxan
(Junod, Lambert, Orci, Pictet, Gonet and Renold 1967), where a single injection of 25-
100 mg/kg body wt streptozotocin induces diabetes (Rakieten, Rakieten and Nadkami
1963; Junod, Lambert, Stauffacher and Renold 1969). Such diabetic rats show rapid
body weight loss, hyperglycemia, intermittent to continuous diarrhoea, polydipsia,
polyuria and glycosuria (Junod et al. 1969; Ganda, Rossini and Like 1976).
75

Although diabetic neuropathy is known as a major clinical problem, there has
not been a satisfactory treatment so far available. One of the various therapeutic
modalities investigated to prevent and/or cure diabetic neuropathy involves
ganglioside. Ganglioside is a diverse group of sialic acid-containing glycosphingolipid
that is present in the plasma membrane of mammalian cells, being particularly
abundant in neurons and associated glial cells (Wiegandt 1971; Ledeen 1978; Ledeen
and Yu 1982; Ando 1983; Stults, Sweeley and Macher 1989).
A ganglioside molecule is composed of a hydrophilic portion called syalosyl-
oligosaccharide and a hydrophobic part called ceramide. The ceramide is formed of
sphingosine and fatty acid and is inserted into the outer leaflet of the lipid bilayer of
the plasma membrane, whereas the negatively hydrophilic sugar moiety, the syalosyl-
oligosaccharide, protrudes towards the extracellular environment (see Cuello 1990).
The asymmetrical structure of ganglioside and its location in the plasma membrane
makes it best placed to interact with other constituents of the plasma membrane of the
same or neighbouring cells and other exogenous molecules coming in contact with the
cell. Treatment of ganglioside, both in vivo and in vitro stimulates nurite outgrowth,
axonal sprouting, synapse formation and regeneration. Furthermore, ganglioside has
been shown to prevent or facilitate recovery from diabetic-induced neuropathic
changes (see Cuello 1990). In the present study, if ganglioside treatment has any effect
on results obtained in the streptozotocin-induced diabetic rats was also investigated.
Experimental protocols
Diabetes was induced by a single intraperitoneal injection of streptozotocin (65
mg/kg body wt; Upjohn, Kalamazoo, USA) in saline. Onset of diabetes was established
by rapid body wt loss, polyuria and glycosuria. To investigate the effect of ganglioside
upon changes occurring in diabetic rats, starting one week after induction of diabetes, a
group of diabetic rats were given intraperitoneal injections of 30 mg/kg body wt bovine
brain ganglioside (AGFj^; Fidia, Abano Terme, Italy) twice a week for 7 weeks.
76

2:9. Sources of Antibodies, Chemicals and Reagents
Antibodies
Calretinin..........................Gift from Dr J. Rogers, Cambridge, UK
CGRP................................Cambridge Research Biochemicals, UK
N O S..................................Gift from Dr U. Forstermann, Abbot Laboratories, USA
(For studies reported in Sections 3:1:1-2 and 3:2:2.
....................................... Euro-Diagnostica, Maimo, Sweden
(For studies reported in Sections 3:1:3 and 3:2.1.
N PY .................................. UCB, Belgium
PGP 9 .5 ............................ Ultraclon, UK
S P ......................................Cambridge Research Biochemicals
T H .....................................Affinity, UK
V IP ....................................Incstar, UK
Biotinylated donkey anti-rabbit antibody.....................Amersham Ltd, Amersham, UK
Biotinylated sheep anti-mouse antibody...................... Amersham
Rhodamine conjugated goat anti-rabbit antibody Nordic Immunological
Laboratories, Maidenhead Berks, UK
Streptavidin fluorescein..................................................Amersham
Chemicals and Reagents
Aprotonin...........................Sigma, Poole, UK
L-arginine........................ Dupont/New Research Products, Stevenage, UK
BH4 ................................... Research Biochemicals Inc, St Albans, UK
Bovine serum albumin Sigma
CaClo.................................. Sigma
Calmodulin..........................Sigma
77

Capsaicin......................... Sigma
Citifluor........................... City University, London, UK
Colchicine........................ Sigma
Diastix..............................Ames, Slough, UK
Diazepam (Valium) Phonix Pharmaceutical Ltd, Gloster, UK
Dithiothreitol...................Sigma
Dowex 50X8-200 (H form) ion-exchange resin Aldrich Chemical Co Ltd,
Gillingham, UK
EDTA............................... Sigma
EGTA............................... Sigma
FA D .................................. Sigma
Fentanyl-fluanisone (Hypnorm)........... Janssen, Oxford, UK
Glacial acetic ac id .................................. BDH, Poole, UK
Ganglioside.............................................Fidia, Abano Terme, Italy
Guanethidine sulphate (Ismelin)...........Ciba Laboratories, Horsham, UK
H C l........................................................... BDH
Iso-pentane..............................................BDH
Leupeptin..................................................Sigma
Lysine.......................................................Sigma
L-malic ac id ............................................Sigma
^-N A D PH ...............................................Sigma
L-N A M E................................................Sigma
N aC l........................................................ DBH
NaOH...................................................... Sigma
Nitroblue tétrazolium.............................Sigma
OCT (Tissue-tek).................................. Miles Inc., Elhart, USA
6 -O H D A ............................................... Sigma
78

Optiphase "HiSafe" 3 .......................... Fisons, Loughbonigh, UK
Paraformaldehyde................................TAAB, Reading, UK
Phosphate-buffered saline..................BDH
Physiological saline..............................Antigen Pharmaceuticals Ltd., Roscrea, Ireland
Poly L -lysine .......................................Sigma
Propylene glycol..................................Sigma
Reserpine.............................................Sigma
Sodium azide...................................... Sigma
Soybean trypsin inhibitor...................Sigma
Streptozotocin.....................................Upjohn, Kalamazoo, USA
Sucrose................................................BDH
T R IS .................................................... Sigma
Triton X -1 0 0 ......................................BDH
Tween 8 0 ............................................ Sigma
79

SECTION 3
RESULTS
80

SECTION 3:1
DISTRIBUTION AND CHARACTERISTICS OF NITRIC OXIDE-
SYNTHESIZING NERVES IN THE RAT ADRENAL GLAND
81

SECTION 3:1:1
COLOCALIZATION OF NITRIC OXIDE SYNTHASE AND
NADPH-DIAPHORASE IN RAT ADRENAL GLAND
82

Abstract
The distribution and colocalization of nitric oxide synthase and NADPH-
diaphorase was studied in the adrenal gland of rat. Ganglion cells and many nerve
fibres in the gland showed both nitric oxide synthase immunoreactivity and NADPH-
diaphorase staining. The adrenal cortical cells showed NADPH-diaphorase staining
but were not immunoreactive for nitric oxide synthase.
Positive labelling for both NADPH-diaphorase and nitric oxide synthase was
found in bundles and in single fibres with varicosities, preferentially located around the
noradrenaline-storing cells. Adrenaline-storing cells and ganglion cells in the medulla,
along with the cortical cells and blood vessels in the zona glomerulosa received
relatively fewer positive fibres.
83

Introduction
There are recent reports that refer briefly to the occurrence of NOS-
immunoreactive and NADPH-diaphorase-reactive ganglion cells and nerve fibres in
the adrenal gland of the rat (Bredt et al. 1990; Dawson et al. 1991). The ganglion cells
of the adrenal gland are poorly understood with regard to their transmitters and
function, although there is some evidence for the existence of both cholinergic
(Coupland and Holmes 1958) and noradrenergic (Dagerlind et al. 1990) neurons in the
gland.
The present study, utilizing NOS immunohistochemistry and NADPH-
diaphorase histochemistry, was carried out in order to give a fuller understanding of the
nature and distribution of NOS-containing cells and nerve fibres in the adrenal gland
of rat. This study demonstrates for the first time the differential distribution of NOS-
containing fibres within different regions of the gland and characterizes the NOS-
containing neurons.
84

Materials and methods
Five adult male Wistar rats were killed by an overdose inhalation of carbon
dioxide. The adrenal glands were then removed, bisected and fixed in 4 %
paraformaldehyde in 0.1 M PBS and cryostat sections were prepared as described in
Section 2:3:1. Fixation with 4 % paraformaldehyde enables NA cells to be
distinguished from A cells by fluorescence microscopy (Eranko 1952, 1955).
Therefore, prior to processing for NOS immunohistochemistry, the adrenal sections
were mounted in PBS and examined under fluorescence microscope and photographed.
The objective of this process was to examine if there is any correlation between the
distribution of NOS-immunoreactive elements and either NA or A cells. Coverslips
were then removed and adrenal sections were rinsed in PBS and
immunohistochemistry for NOS was carried out as described in Section 2:3:2. The
NOS antibody used was raised in rabbit against purified soluble NOS extracted from
rat cerebellum (Schmidt et al. 1991), and was used at a dilution of 1:2500.
Immunoprocessed adrenal sections were examined under a fluorescence microscope
and labelled cellular elements were studied and photographed. Coverslips were
removed and adrenal sections were rinsed in PBS.
NADPH-diaphorase histochemistry as described in Section 2:3:3 was carried
out on adrenal gland sections previously immunoreacted for NOS and their non-
immunoreacted consecutive sections. NADPH-diaphorase stained adrenal sections- t u e .
were then inspected under ordinary light illumination using^same microscope as for
NOS immunoreacted ones.
85

Results
Control experiments in which the NOS antiserum was omitted or substituted
with non-immune serum and preimmune serum failed to show labelling. Similarly, no
NADPH-diaphorase staining was seen in the control sections incubated in media
devoid of )3-NADPH or in those sections pretreated with TRIS HCl heated at 75°C.
Examination of the sections immunostained with NOS antisera revealed that
several ganglion cells and many nerve fibres were positively immunoreactive, while
chromaffin and cortical cells were non-reactive. The majority of the immunoreactive
ganglion cells were large with an average diameter of 32 fim and were round in shape
with a circular nucleus devoid of reaction product (Fig. la). A small number of
immunoreactive neurons appeared to contain two nuclei (Fig. Ic). The NOS-positive
neurons generally occurred as small clusters of cells ranging up to 6 in number,
although occasionally they were found singly or in pairs. Apart from a few
immunoreactive ganglion cells which appeared in the juxtamedullary region of the
zona reticularis, no reactive cell was observed in the cortex. The majority of the
immunoreactive neurons showed similar bright, homogeneous fluorescence. A small
number of ganglion cells showed very light fluorescence.
Occasional NOS-immunoreactive nerve fibres were rarely observed in the
capsule of the gland. However, NOS-positive fibres with varicosities formed a plexus
in the subcapsular region and zona glomerulosa of the cortex (Fig. 2a). Single nerve
fibres with varicosities were seen to arise from this plexus, branch and run among the
cortical cells and blood vessels in the zona glomerulosa.
In the zonae fasciculata and reticularis occasional NOS-immunoreactive nerve
bundles without varicosities were encountered running among blood vessels and cords
of cortical cells (Fig. 2b).
In the adrenal medulla several NOS-immunoreactive nerve bundles were
observed, in addition to many single nerve fibres with varicosities. The single nerve
86

fibres appeared to form a closely associated network particularly around the NA cells
(Figs. 2c, d). Relatively less positive nerve fibres were observed around the A cells and
immunoreactive ganglion cells.
Examination of sections processed for the NADPH-diaphorase histochemistry
showed that all cortical cells forming the parenchyma of the cortex and those
occasionally found dispersed in the medulla as cortical islands, along with several
ganglion cells and nerve fibres, were positively stained (Fig. lb). The reactive ganglion
cells were similar in morphology to the NOS-immunoreactive cells described above. A
number of the NADPH-diaphorase stained neurons displayed a cytoplasmic process
extending from one pole of the cell.
When consecutive sections were compared, almost all NOS-immunoreactive
ganglion cells were found to be stained for NADPH-diaphorase. In double labelling
studies all NOS-immunopositive neurons were also found to be stained for NADPH-
diaphorase (Figs. la, b). As for NOS immunoreactivity, a small number of ganglion
cells showed very light staining. These neurons accounted for not more than 5 % as
compared to the strongly positive reactive neurons counted, which ranged in number
from 87 to 114, in 5 animals. The intensity of NADPH-diaphorase staining in the
double labelling experiment was reduced as compared to the NADPH-diaphorase
staining alone in consecutive sections. This was particularly noticeable for cortical
cells.
NADPH-diaphorase stained nerve fibres in both adrenal medulla (Fig. lb) and
cortex showed a similar pattern of distribution to that observed and described with the
NOS immunoreaction. However, the intensity of the staining and the frequency of the
NADPH-diaphorase stained fibres appeared relatively less in comparison with those
immunopositive for NOS, and no NADPH-diaphorase-positive fibre was seen in the
zona glomerulosa of the adrenal glands of 2 of the 5 animals.
87

Fig. 1. Micrographs of rat adrenal gland sections demonstrating:
a, b. double labelled ganglion cells and nerve fibres with NOS
immunoreactivity (a) and NADPH-diaphorase staining (b); arrowheads point to
labelled nerve fibres. Note in b the cortical cells (Co) are stained for NADPH-
diaphorase. M: Medulla. Bars = 35 ywm.
c. NOS-immunoreactive binucleated ganglion cell {arrowhead)^ as well as
mononucleated ganglion cell {open arrow)\ arrow points to NOS-
immunoreactive nerve bundle. Co: Cortex; M: Medulla. Bar = 35 /rni.
88
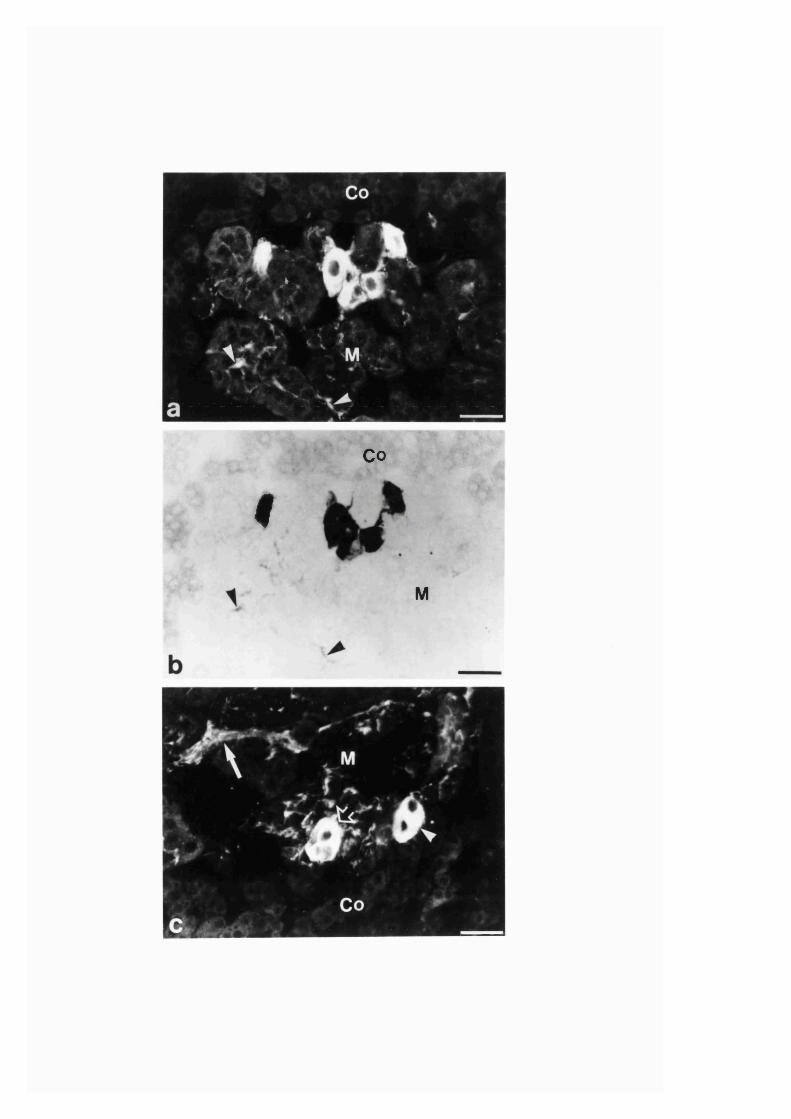
- -
- - . f ' .1
f
Co
IM

Fig. 2. Fluorescence micrographs of rat adrenal gland demonstrating:
a,b. NOS-immunoreactive nerve fibres in the subcapsular region and zona
glomerulosa (ZG; a), and zonae fasciculata (ZF) and reticularis (ZR; b); Ca:
capsule. Bars = 60 fjm.
c,d. catecholamine fluorescence (c) and the same section immunostained for
NOS (d). Note that the noradrenaline-storing cells (NA) are more densely
innervated than the adrenaline-storing cells (A). Bars = 6 0 ^ .
89
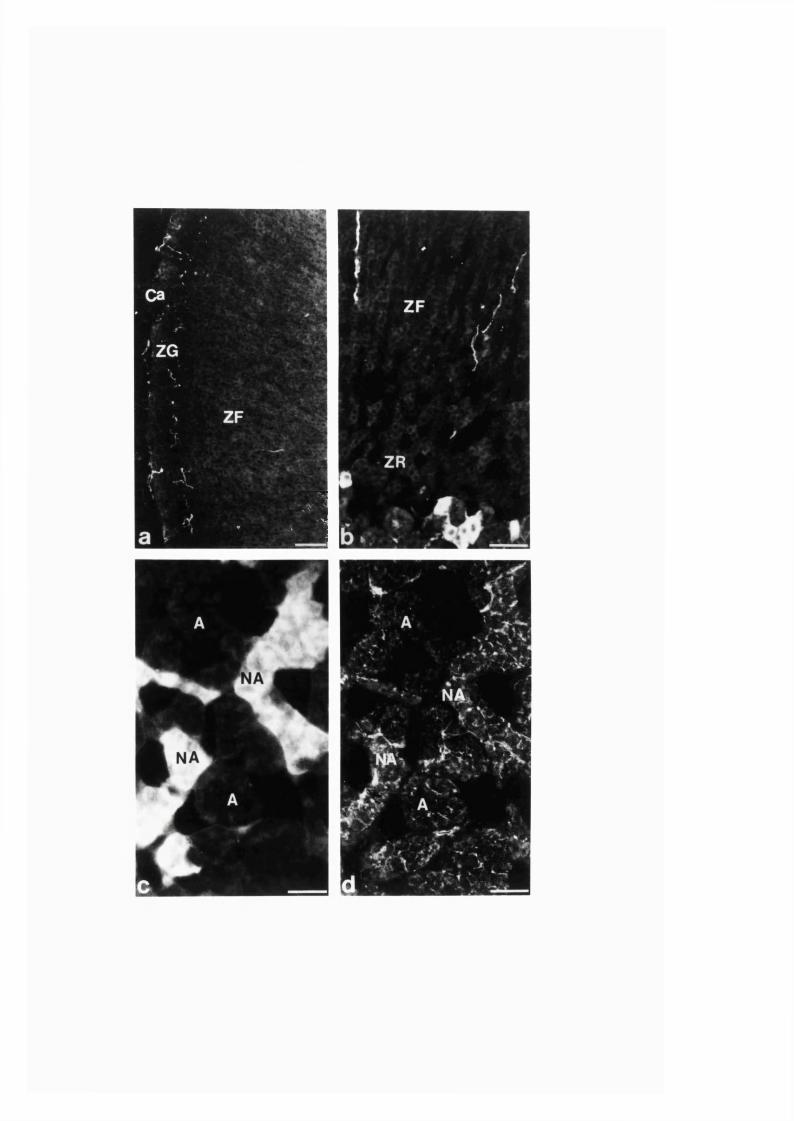
^Bl

Discussion
The failure of immunoiabelling in the control experiments in which the NOS
antiserum was omitted or substituted with preimmune serum and non-immune serum
showed that the NOS-immunoreactivity observed with the NOS antibody was specific.
In addition, the NOS antisera used was tested on other tissues, such as cultured
myenteric plexus of the guinea-pig (Saffrey, Hassall, Hoyle, Belai, Moss, Schmidt,
Forstermann, Murad and Bumstock 1992) and whole mount preparations of rat
myenteric plexus (Belai, Schmidt, Hoyle, Hassall, Saffrey, Moss, Forstermann, Murad
and Bumstock 1992), and its specificity has also been established. Therefore, the NOS
immunoreaction observed in this study was due to NOS present in the tissue.
Adrenal cortical cells are involved in steroid synthesis and contain NADPH-
requiring steroidogenic enzymes (Sweat and Lipscomb 1955). It may be because of
these enzymes other than NOS that the cortical cells gave staining for NADPH-
diaphorase but failed to immunoreact for NOS antisera. Altematively, as NOS is
known to exist in different isoforms (see Forstermann et al. 1991), the cortical
NADPH-diaphorase may be related to another isoform, which could not be recognized
by the neuronal NOS antibody used in this study. This possibility cannot be ruled out,
and awaits further investigation.
The labelling of the ganglion cells and the nerve fibres of the adrenal gland with
both NOS and NADPH-diaphorase confirms earlier observations by other workers
(Bredt et al. 1990; Dawson et al. 1991) and supports the suggestion that NOS and
neuronal NADPH-diaphorase label similar sites (Dawson et al. 1991; Hope et al.
1991). The reduction in the intensity of the NADPH-diaphorase staining in neurons
and the lack of staining of some of the nerve fibres during double labelling may be due
to the weakening of the reactivity as a result of one or more of the prior
immunoreaction procedures.
The occasional immunoreactive fibres observed in the capsule suggest that some
90

NOS fibres in the gland may have an extrinsic origin rather than originating from
intrinsic reactive neurons. The sympathoadrenal preganglionic neurons located in the
intermediolateral cell column of the spinal cord (Kesse et al. 1988) show NADPH-
diaphorase staining (Blottner and Baumgarten 1992). In addition, neurons in the dorsal
root ganglia and the vagal ganglia from where the adrenal gland gets its additional
nerve supply (Parker et al. 1990a; Coupland et al. 1989) also show NADPH-
diaphorase activity (Aimi, Fujimura, Vincent and Kimura 1991). Denervation
experiments are carried out in order to explore the possible existence of an extrinsic
innervation and its contribution in comparison to the intrinsic one (see Section 3:1:3).
NO from both pre- and postsynaptic neural sources has been implicated with
various functions (see Garthwaite 1991). Administration of L-nitroarginine
methylester, one of the NOS inhibitor drugs, has been shown to increase mean arterial
blood pressure and decrease the adrenal blood flow from the adrenal gland of dog
(Breslow, Tobin, Bredt, Ferris, Snyder and Traystman 1992). Similarly, the NOS-
immunoreactive fibres in the adrenal gland of rat may modulate vasodilation and
increase the adrenal blood flow during the secretory activities of the gland.
The close association of a plexus of NOS-positive varicose fibres with the
cellular elements of the gland suggests that NO may have a role concerning cellular
function. Catecholamine secretion from the adrenal medulla of rat is mediated by
cholinergic and non-cholinergic mechanisms (Malhotra and Wakade 1987a). While
the cholinergic component comes from the preganglionic nerve fibres in the spinal cord
(Kesse et al. 1988), the non-cholinergic component is not well understood in terms of
the type and origin of transmitters, although some peptidergic nerve fibres found in the
gland have been suggested for their possible involvement (see Livett et al. 1990). The
morphological observations of the NOS-positive fibres presented here provide support
for the view that NO may be an additional transmitter involved in the control of
catecholamine secretion from the gland. As reported here the NOS-positive fibres are
91

more densely associated with NA than A cells. The functional implication of these
observations needs resolution.
While the adrenal cortex has traditionally been considered to be principally
controlled by ACTH, there is now increasing morphological and functional evidence
for neural control of such functions as steroidogenesis (Holzwarth, Cunningham and
Kleitman 1987), compensatory adrenal growth (Dallman, Engeland and Shinsako
1976) and adrenal blood flow (Engeland, Lilly and Gann 1985). The NOS-positive
fibres found in a varicose plexus in the zona glomerulosa may therefore imply that NO
in this region of the cortex exerts its effect in either one or a combination of the above
activities.
In conclusion, the present immunohistochemical and histochemical study shows
that NOS and NADPH-diaphorase are colocalized in the adrenal ganglion cells. This
further confirms that neuronal NADPH-diaphorase is identical to NOS in the nervous
system and provides morphological evidence for the synthesis of NO in nerves
supplying the adrenal gland. Several NOS-positive fibres arising from the intrinsic
neurons and possibly also from another extrinsic source, innervate the adrenal medulla
and zona glomerulosa of the adrenal cortex. In the adrenal medulla the NA cells
receive greater proportion of the NOS-positive nerve fibres than the A cells or the
ganglion cells. It seems that NO may have a regulatory and/or modulatory role in the
cellular function of the adrenal medulla and zona glomerulosa of the adrenal cortex
and/or blood flow of the gland.
92

SECTION 3:1:2
DISTRIBUTION AND COLOCALIZATION OF NITRIC OXIDE SYNTHASE
AND NADPH-DIAPHORASE IN ADRENAL GLAND OF DEVELOPING,
ADULT AND AGING SPRAGUE-DAWLEY RATS
93

Abstract
The distribution and colocalization of nitric oxide synthase and NADPH-
diaphorase was investigated in the adrenal gland of developing, adult and aging rats
with the use of immunohistochemical and histochemical techniques. Nitric oxide
synthase-immunoreactive neurons within the adrenal gland were found from the 20^^
day of gestation onwards. During early development the neurons were found as small
clusters of smaller-size cells compared to those observed in the adult gland. Their
number reached that of adult level by the 4^ ̂ day after birth, and in the glands from
aging rats a 28.6 % increase was observed.
Whilst no immunofluorescence was seen in chromaffin cells during early
development, some cells from glands of aging rats showed nitric oxide synthase-
immunoreactivity with varying intensity. The immunoreactive neurons from postnatal
rat adrenals were also positive for NADPH-diaphorase, whilst those in prenatal rats
were negative or lightly stained.
Nitric oxide synthase-immunoreactive nerve fibres were present in all adrenal
glands examined from the 16^ ̂ day of gestation onwards. A considerable degree of
variation in the distribution of immunoreactive fibres both in medulla and outer region
of cortex at the different age groups was observed and described. Most, but not all,
nitric oxide synthase-immunoreactive nerve fibres also showed NADPH-diaphorase
staining.
94

Introduction
Innervation of the adrenal gland of the rat shows developmental plasticity.
Although synaptic elements can be identified as early as 15.5 days of gestation
(Daikoku et al. 1977), functional splanchnic innervation of the medulla develops
during the first week of postnatal life reaching maturity by day 10 (Slotkin et al. 1980,
1982). During development, there is also evidence of transitional changes in various
messenger molecules such as acetylcholine and catecholamines (Tomlinson and
Coupland 1990), and neuropeptides (Henion and Landis 1990) contained in the gland.
In aging, the adrenal medullary cells show evidence of proliferative lesions and
structural changes (Tischler et al. 1985; Tischler and DeLellis 1988; Coupland and
Tomlinson 1989). An increase in adrenal sympathetic neural activity associated with an
increase in catecholamine secretion is another phenomenon observed during aging (Ito
et al. 1986). Therefore, the distribution and colocalization of NOS and NADPH-
diaphorase in the developing, adult and aging rat adrenal gland warrants examination.
Using immunohistochemical and histochemical techniques, this study demonstrates
evidence of plasticity in the distribution of NOS in the adrenal gland of the rat during
development and aging.
95

Materials and methods
The study was conducted on male (postnatal) and either sex (prenatal) of
Sprague-Dawley rats. The age of the animals was determined by counting from the
appearance of a cervical mucous plug as day 0 of embryonic life (EO) and the day of
delivery as day 0 of postnatal life. Adrenal glands from 5 rats of each of the following
age were studied: developing rats of E16, E18, E20, 1, 4, 7, 14 and 21 days of age;
adult rats of 16 weeks of age and aging rats of 24 months.
Pregnant rats were with carbon dioxide and foetuses were
removed and their adrenals quickly dissected out and immersion fixed in 4 %
paraformaldehyde in 0.1 M PBS for 2 h at room temperature. Similarly, all other rats
were sacrificed by carbon dioxide asphyxiation, their adrenals removed and immersion
fixed for 2 h. Cryostat sections were prepared as described in Section 2:3:1 and
immunohistochemistry for NOS antibody was carried out as described in Section 2:3:2.
The NOS antibody was raised in rabbit against soluble NOS extracted from rat
cerebellum (Schmidt et al. 1991), and used at a dilution of 1:2500. Immunoprocessed
adrenal sections were examined under fluorescence microscope and labelled cellular
elements were studied and photographed. Coverslips were removed and sections were
rinsed in PBS.
Adrenal gland sections that were previously immunostained with NOS
antiserum, their consecutive sections and several other consecutive sections were
processed for NADPH-diaphorase histochemistry as described in Section 2:3:3.
Adrenal sections processed for NADPH-diaphorase histochemistry were inspected
using same microscope under ordinary light illuminations.
All labelled neurons were counted and the diameter of representative neurons
were measured. To determine whether significant differences existed among the mean
number of labelled neurons at the different age groups, statistical tests with one way
analysis of variance were performed using Fisher's least significant difference
procedure.
96

Results
Adult rats
The NOS immunoreactivity and NADPH-diaphorase staining in the adrenal
glands of the adult Sprague-Dawley rats were comparable to that of Wistar rats
described in Section 3:1:1. NOS-immunoreactivity was found in several ganglion cells
of the adrenal gland (Fig. 3a). A small number of these neurons were located in the
zona reticularis, whereas the remainder was found in the adrenal medulla. Within the
medulla the majority of the NOS-immunoreactive neurons appeared to be peripherally
localized in comparison to a variable number of larger sized, closely packed non-
immunoreactive ganglion cells seen mainly towards the centre of the medulla.
Several NOS-immunoreactive nerve fibres, running in bundles and as single
nerve fibres with varicosities, were found in both adrenal medulla and cortex. In the
medulla, the single nerve fibres were closely associated with chromaffin cells (Fig. 3c).
As reported in Section 3:1:1, paraformaldehyde-induced fluorescence showed that NA
cells were more densely innervated than the A cells. Ganglion cells in the gland were
also innervated, with the large non-reactive neurons being more densely innervated
than those immunoreactive for NOS.
In the adrenal cortex the NOS-immunoreactive nerve fibres were observed in
the subcapsular region and zona glomerulosa (Fig. 6g). In the inner two zonae of the
gland, immunoreactive nerve bundles were seen occasionally running along blood
vessels and cords of cortical cells. In addition, in the zona reticularis and also the
medulla, a small number of blood vessels was seen closely surrounded by
immunoreactive nerve fibres. Some of the fibres were seen projecting from nearby
reactive neurons (Fig. 3d).
NADPH-diaphorase histochemistry revealed that all cortical cells, occasional
cellular elements dispersed in the medulla and several ganglion cells as well as nerve
97

fibres were positively stained (Figs. 3b, e, f). The majority of the dispersed positively
stained cells found in the medulla were morphologically similar to cortical cells.
However, a smaller number of the cells, some of which were elongated with a tapering
end observed between groups of chromaffin cells (Fig. 3f) and apposed to non-reactive
neurons, appeared to be morphologically different. These cells were also obvious due
to their intensity of staining, which was similar to other cells at the corticomedullary
junction (Fig. 3f).
In double-labelling experiments, all neurons and most of the nerve fibres that
were immunoreactive for NOS were also found to be positive for NADPH-diaphorase.
However, reduction in staining intensity of the cortical cells was noticeable on NOS-
preimmunolabelled sections as compared to sections stained for NADPH-diaphorase
alone.
Developing rats
While a weak fluorescence was observed in most cells of the adrenal cortical
primordium of prenatal rats, no clearly labelled NOS-immunoreactive cell was
observed in the adrenal glands at E l6 and E l8. However, in the extra-adrenal
medullary blastema, accumulated at the medial aspect of the developing adrenal glands
of E18 rats, a small number of small-sized immunoreactive neural cells (10-13 pm in
diameter) and nerve fibres were observed (Fig. 4a).
Intra-adrenal NOS-immunoreactive neurons were found in glands from rats of
E20 and older. At E2Ü the immunoreaction in the majority of neurons and nerve fibres
was of lower intensity than that observed in postnatal rats. The neurons, in the
developing rat adrenals, were small in diameter (15-20 pm) and found as small
compact clusters located in the cortex at E20 and day 1 (Figs. 4d, g) and in the outer
regions of the medulla in all other age groups of developing rats (Fig. 4i). Cytoplasmic
processes were seen arising from some of the neurons found in the developing rat
adrenals. In addition to the NOS-immunofluorescence, the cytoplasmic processes
98

helped to ascertain the neural identity of the labelled cells in glands from early stages of
development where NADPH-diaphorase staining intensity of the labelled neurons was
relatively indistinguishable from the surrounding cortical ceils.
NOS-immunoreactive nerve fibres with varying intensity and distribution were
seen in all age groups of developing rat adrenals studied. At E l 6 and E l8 a few fibres
were found as faintly labelled nerve bundles running through the cortical primordium.
In the medulla of E20 and older rats, an increase in frequency of the immunoreactive
fibres was found with increasing age of the animal (Figs. 4c, f, i), reaching at 21 days
the level seen in the adult rats. In the outer cortex, the immunoreactive fibres
progressively increased in frequency up to 7 days, followed by a progressive decrease
in 14 and 21 day-old rats (Figs. 6a-f).
NADPH-diaphorase histochemistry of adrenal sections from prenatal rats
showed staining in a few nerve fibres and most of the cortical cells with varying
intensity. In double-labelling experiments, nerve fibres and neurons that were
immunoreactive for NOS antiserum were negative or stained weakly for NADPH-
diaphorase (Figs. 4b, e). In postnatal developing adrenals, NADPH-diaphorase
staining was found in the same cellular elements, and also was colocalized with NOS-
immunoreactivity in a similar pattern as described for the adult gland (Figs. 4h, j).
Aging rats
Neurons comparable in morphology and size (20-35 /^m) to those seen in the
adult gland, found occasionally in small groups but mainly occurring singly in the
medulla, exhibited bright NOS-immunofluorescence (Fig. 5a). Occasionally, regions
of adrenal medullary chromaffin cells showed a weak heterogeneous
immunofluorescence (Fig 5c). Furthermore, in two specimens, an occasional group of
chromaffin cells displayed bright homogeneous fluorescence (Fig. 5d).
In the regions of adrenal medulla showing weak heterogeneous
99

immunofluorescence, an increase in NOS-immunoreactive nerve fibres was observed,
A dense innervation, similar to that of adult rats, was also seen in the region of the
non-reactive large ganglion cells (Fig. 5c). Alternatively, the immunoreactive nerve
fibres in the remaining regions of the adrenal medulla (Fig. 5a) and cortex (Fig. 6h)
were reduced in comparison with those in the adult rats. The NADPH-diaphorase
staining was localized in the same cellular elements as described in the adult gland and
was colocalized with NOS-immunoreactivity in the same pattern (Figs. 5a, b), except
for the immunoreactive chromaffin cells which failed to stain for NADPH-diaphorase
(Figs. 5d, e).
100

Fig. 3. Photomicrographs of adrenal gland sections demonstrating:
a, b. NOS-immunoreacted (a) and NADPH-diaphorase double-stained (b)
ganglion cells in adrenal medulla of 16 week-old rat. Arrows in b point to
NADPH-diaphorase stained cortical cells located in medulla. Bars = 33 /am.
c. NOS-immunoreactive nerve fibres in adrenal medulla of 16 week-old rat
associated to medullary cells and also surrounding the outlines of large-size
non-immunoreactive neurons (arrows). Bar = 33 /am.
d,e. NOS-immunoreactive (d) and NADPH-diaphorase-stained (e) nerve
processes projecting from nearby reactive neurons to encircle a blood vessel in
zona reticularis of cortex (Co) of 16 week-old rat. Co: cortex; M: medulla. Bars
= 33 /am.
f. NADPH-diaphorase-staining in 16 week-old rat. Arrows point to NADPH-
diaphorase-positive elongated cells located between groups of medullary cells.
Note at corticomedullary junction a group of cells (arrowhead) displaying a
similar intensity of staining as those found scattered in the medulla, but heavier
than cortical cells. Co: cortex; M: medulla. Bar = 33 /cm.
101
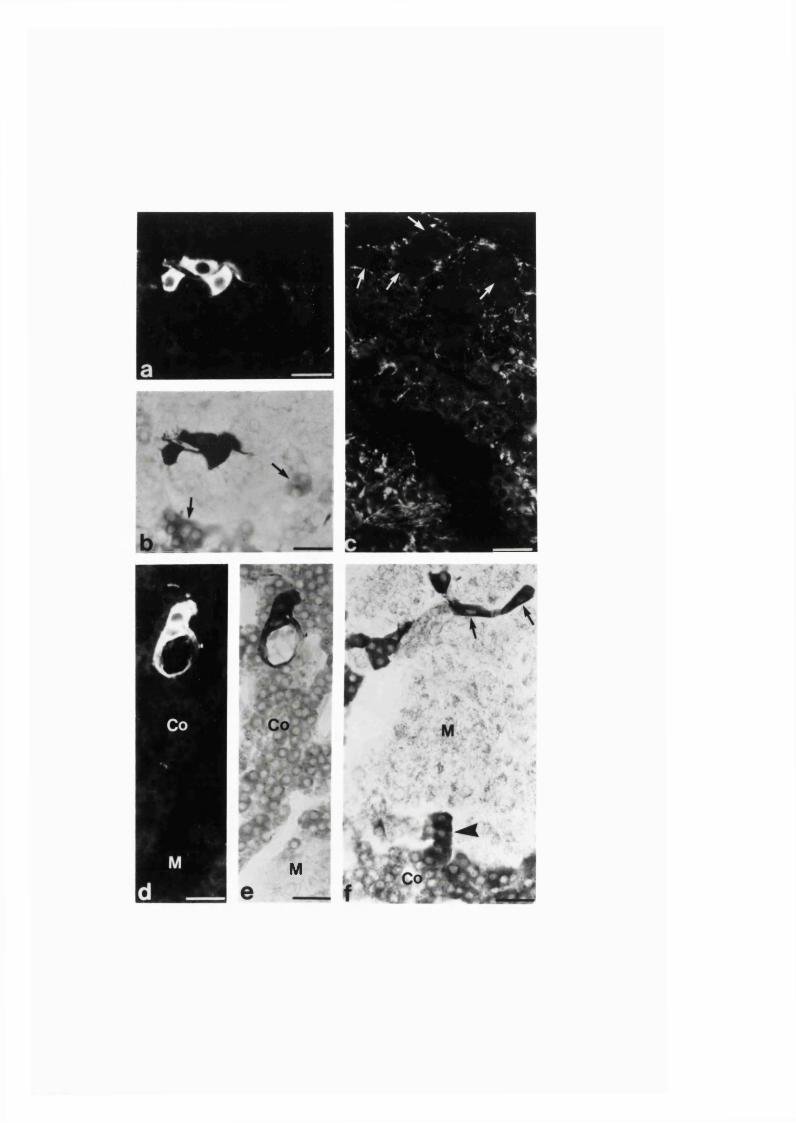
M

Fig. 4. Photomicrographs of:
a, b. NOS-immunoreactive neurons and nerve fibres in the extra-adrenal
medullary blastema of E18 rat (a), and same section double-stained for
NADPH-diaphorase (b). Note NOS-immunoreactive neurons and nerve fibres
are negative or weakly stained for NADPH-diaphorase. Bars = 33 f/m.
c-e. Sections of adrenal gland from E20 rat showing NOS-immunoreactive
nerve fibres (c) and neurons (d), and same section as Figure d double-stained
for NADPH-diaphorase (e). Note that the NOS-immunoreactive neurons are
weakly stained for NADPH-diaphorase. Arrows in Figure c point to
immunoreactive nerve fibres. Ca: capsule; Co: cortex; M: medulla. Bars = 33
jum,
f-h. Sections of adrenal gland firom 1 day-old rat showing NOS-
immunoreactive nerve fibres (f) and neurons (g), and same section as g double
labelled with NADPH-diaphorase (h). Co: cortex; M: medulla. Bars = 33 jum.
i, j. NOS-immunoreactive (i) and NADPH-diaphorase double-stained (j)
neurons and nerve fibres in sections of adrenal medulla from 7 day-old rat. Bars
= 33 jum.
102
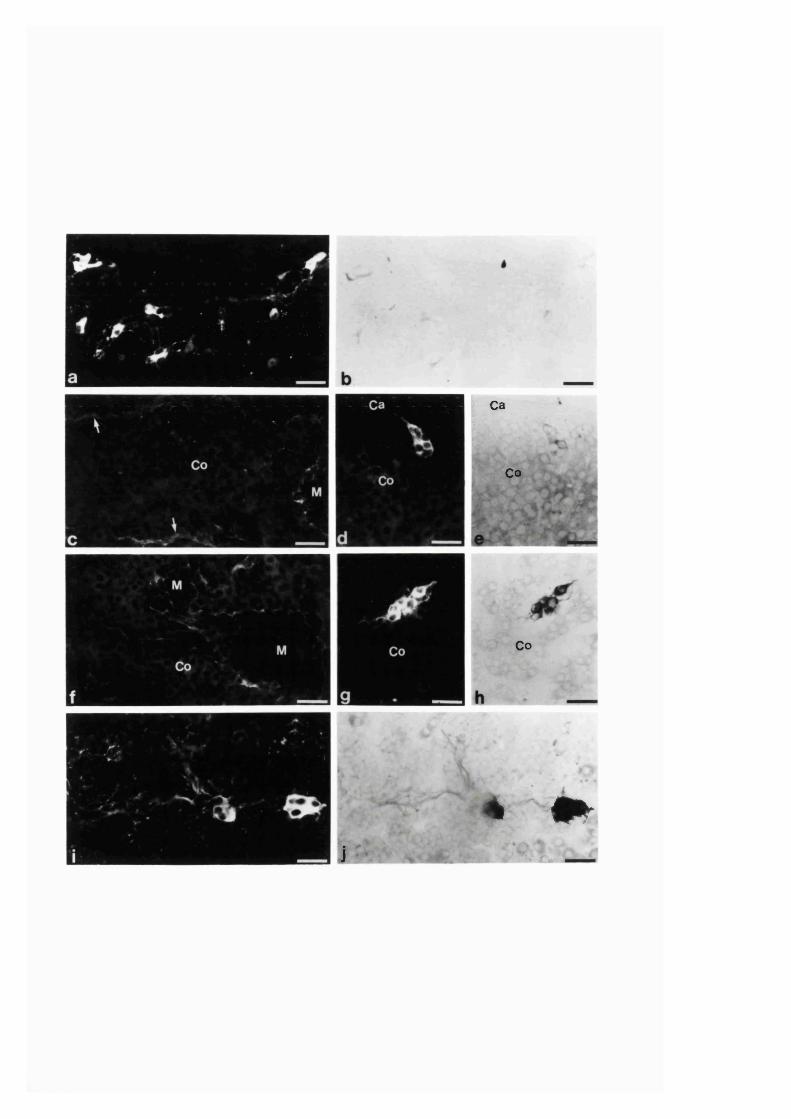
' ' 'A
•
j
ca
Co
Co
* - m

Fig. 5. Photomicrographs of:
a, b. Section of adrenal gland from 24 month-old rat showing labelled neurons
and nerve fibres for NOS-immunoreactivity (a) and the same section double
stained for NADPH-diaphorase (b). Note occasional reactive neuron adjacent to
sinusoid (S). Co: cortex. Bars = 33 fim.
c. NOS-immunoreactive neurons and nerve fibres in section of adrenal medulla
from 24 month-old rat. Note dispersed immunoreactive granules in chromaffin
cells. point to large-sized non-immunoreactive neurons. Bar = 5 2 ^ .
d, e. NOS-immunoreactive chromaffin cells of adrenal medulla (d) and same
section double processed for NADPH-diaphorase (e) in 24 month-old rat. Note
the NOS-positive chromaffin cells are negative for NADPH-diaphorase. Bars =
33ywm.
103
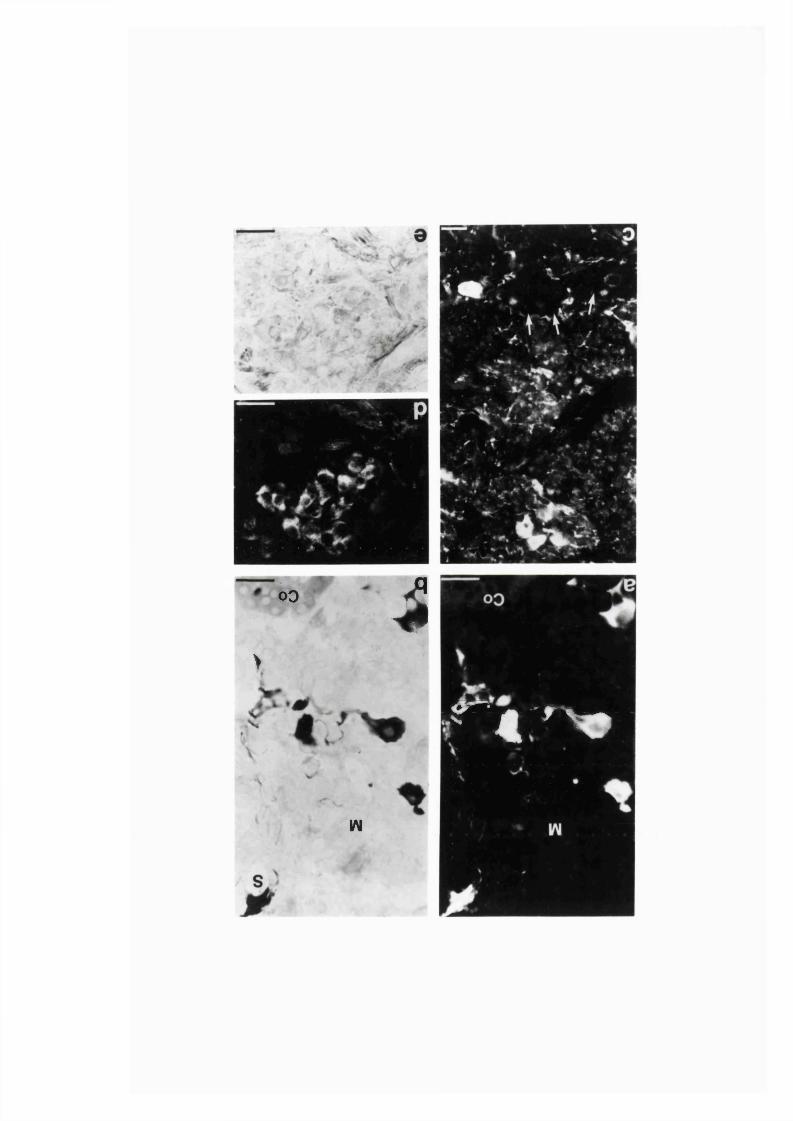
0 3
i
V
r 4 "\#l/\l

Fig. 6a-h. Photomicrographs of NOS-immunoreactive nerve fibres in the
subcapsular and outer cortical regions of adrenal sections from E20 (a), 1 day-
(b), 4 day- (c), 7 day- (d), 2 week- (e), 3 week- (f), 16 week- (g) and 24
month- (h) old rats. Note that the frequency of NOS-immunoreactive nerve
fibres increased with increasing age up to 7 days (d) and showed reduction in 14
days (e) and 21 days (f), to increase again in the adult (g), which then slightly
reduced during aging (h). Ca: capsule; Co: cortex; PC: periadrenal connective
tissue containing NOS-immunoreactive nerve fibres approaching the adrenal
gland of 4 day-old rat. Bars = 52 fim.
104
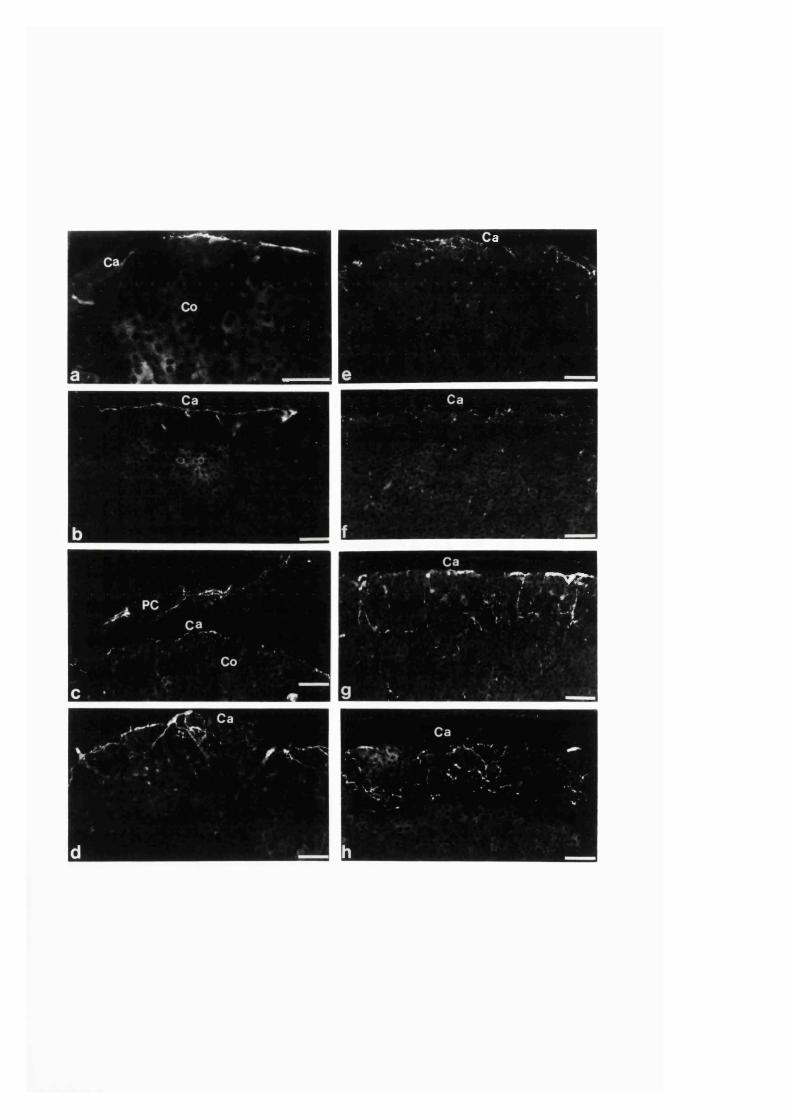
-s®
r ■'~yzr

Table 1. Number of NOS-containing neurons per adrenal gland of the different age
groups, as counted from NOS-immunoreactive and/or NADPH-diaphorase-stained,
14 ̂ m consecutive sections. Values are mean ± standard deviation, n = 5.
Age E20 1 day 4 days 7 days 14 days 21 days 16 weeks 24 months
No o fNOS-\-ve 6±3 20±8.7 108±16.8 92±6.5 104±19.6 107±9.2 112±10.7 144±27.8neurons
105

Discussion
Chromaffin cells and neurons of the adrenal medulla develop from neural crest
cells, which migrate and invade the medial aspect of the cortical analage during foetal
life (Le Douarin 1980; Landis and Patterson 1981; Coupland 1989). Before entering
the adrenal cortical primordium, the migrating cells accumulate at the medial aspect of
the gland and are identifiable as extra-adrenal medullary blastema (Verhofstad et al.
1985). The NOS-immunoreactive neural cells observed in the present study at E18,
near the medial aspect of the adrenals, most likely therefore represent NOS-containing
neurons amongst migrating cells. With increasing age, the progressive inward
localization of the immunoreactive neurons towards the medulla, away from the
periphery of the cortex, followed the developmental pathway taken by the cells.
During early development, the NOS-containing neurons in the adrenal glands
were found to be fewer in number and occurred as clusters of smaller-size cells as
compared to those found in the other age groups. Statistical analysis revealed that with
a level of probability of 0.05 or less the number of neurons found in the gland showed
significant increase at two age levels (see Table 1). The first was between the 1̂ ̂ and
4^ ̂ day after birth where the mean number of neurons increased by 5.4 fold and
reached a number not significantly different from that found in the adult. The second
was in aging rats where a 28.6 % increase above that of adult was observed. An
increase in the number of neurons during postnatal development in the rat is
documented in both the central (Altman and Das 1966) and peripheral (Gabella 1971)
nervous system. In addition, increase in average neural size and reduction in package
density is also a characteristic of developing visceral neurons (Gabella 1971). The
increase in number of neurons may be due to cell division, migration and
transformation (Altman and Das 1966) or subsequent development of expression of a
neuronal marker used in previously negative neurons. It is not known at present which
one or combinations of these factors may be responsible for the observed increase in
106

NOS-positive neurons during development and aging.
The increase in size of NOS-positive neurons during development may be
attributed to an increase in the neuroplasm associated with an increase in size of the
gland. Such a correlated increase of the size of neurons in relation to the size of the
innervated target tissue has been reported in the myenteric ganglia (Gabella and Trigg
1984). In addition, elimination of the small-size neurons as a result of naturally
occurring neural death during postnatal development (see Oppenheim 1991) and their
substitution by larger-size, newly appearing cells may also be possible.
The colocalization of NADPH-diaphorase staining in all NOS-immunoreactive
neurons in postnatal rats is consistent with previous observations in adult rats that
neuronal NADPH-diaphorase is identical to NOS labelling (Dawson et al. 1991; Hope
et al. 1991; Bêlai et al. 1992; Hassall, Saffrey, Bêlai, Hoyle, Moules, Moss, Schmidt,
Murad, Fôrstermann and Bumstock 1992; Saffrey et al. 1992; also Section 3:1:1).
However, the dramatic reduction or failure of the NOS-reactive cells to stain for
NADPH-diaphorase in prenatal rats was interesting. This may indicate that although
NOS is present in a number of neural cells before birth its activity may not be fully
developed to the extent of detection by NADPH-diaphorase histochemistry.
Alternatively, the amount of the NOS present in the prenatal adrenal gland may be
smaller than in glands from postnatal rats and may be below the level of detection with
NADPH-diaphorase histochemistry. The weak immunofluorescence in most NOS-
positive intra-adrenal neurons and nerve fibres before birth may be similarly
accounted by the smaller quantity of the available enzyme. Nevertheless, the lack of
correlation between NADPH-diaphorase activity and NOS immunoreaction in the
adrenal neurons during early development may be an example of the non-functional
nature of adrenal innervation during this period (Slotkin et al. 1980,1982).
There is increasing frequency of spontaneous change in adrenal chromaffin cells
during aging. This change occurs as proliferative lesions of the adrenal medullary cells
107

from hyperplasia, phaeochromocytoma and/or hypertrophy of chromaffin ceils (Tischler
et al. 1985; Tischler and DeLellis 1988; Coupland and Tomlinson 1989). A number of
noradrenaline-storing cells also shows synaptic type vesicles with electron-dense
content indicating differentiation towards the neural phenotype (Coupland and
Tomlinson 1989). It is debatable whether such chromaffin cells displaying
morphological plasticity with age actually present NOS immunoreaction.
The NOS-immunoreactive nerve fibres seen in the subcapsular region and zona
glomerulosa are of interest. Those found as single nerve fibres with varicosities may
indicate a local role for NO in the region, in addition to the possible passage of
extrinsic fibres destined for the medulla. Innervation of this region of the adrenal
cortex with nerve fibres containing several different neurotransmitters with possible
functional implication has been described (see Holzwarth et al. 1987; Charlton 1990;
Hinson 1990). Neural mechanism appears to modulate steroidogenesis in adrenal
cortical cells, as electrical stimulation of the splanchnic nerve has been shown to
enhance the sensitivity of adrenal cortex to ACTH in hypophysectomized animal
(Edwards and Jones 1987). It is a matter to be clarified if the NOS-containing nerve
fibres observed in the zona glomerulosa may innervate cortical cells in the region and
modulate steroidogensis. Furthermore, noradrenaline and adrenaline have been
demonstrated to stimulate release of cortisol and aldosterone from isolated perfused pig
adrenal preparation (Bomstein, Ehrhart-Bomstein, Scherbaum, Pfeiffer and Holst
1990). Since there is evidence for the presence of islets of chromaffin cells in the zona
glomerulosa of rat adrenals (Palacios and Lafarga 1975; Bomstein, Ehrhart-Bomstein,
Usadel, Bockmann and Scherbaum, 1991) influence of locally released catecholamines
from the chromaffin cells and local adrenergic nerve terminals on steroidogenesis has
been suggested (Bomstein et al. 1990; Vizi et al. 1992). Further study is needed to
investigate if NOS-containing nerve fibres innervate chromaffin cells in outer cortex,
similar to those found in adrenal medulla, and play a secondary role in steroidogenesis.
Steroid secretion from adrenal cortex follows an increase in the rate of adrenal blood
108

flow (see Vinson and Hinson 1992). It may therefore be possible for NO, released from
the NOS-containing nerve fibres found in the subcapsular region, to facilitate
steroidogenesis by regulating blood flow to the gland. Nevertheless, such speculation,
on the possible functional implication of the distribution of NOS-containing fibres in
relation to the observed developmental plasticity needs further resolution.
109

SECTION 3:1:3
THE INTRA-ADRENAL DISTRIBUTION OF INTRINSIC AND EXTRINSIC
NITRERGIC NERVE FIBRES IN THE RAT
110

Abstract
The intra-adrenal distribution of nitric oxide synthase-immunoreactive nerve
fibres was studied in rats subjected to various denervations. Splanchnic nerve section
eliminated the nitric oxide synthase-immunoreactive nerve fibres which innervate
adrenal chromaffin and neuronal cells. It did not affect those innervating blood vessels
and zona glomerulosa, which instead were affected by adrenal demedullation.
Guanethidine, 6-hydroxydopamine and capsaicin treatments, however, did not produce
any change.
These results suggest that nitrergic nerves which innervate adrenal medullary
cells are extrinsic (largely preganglionic sympathetic), whilst those innervating zona
glomerulosa and the majority of adrenal blood vessels are intrinsic, and that they do
not belong to nerves sensitive to the sympathetic nerve neurotoxins, guanethidine and
6-hydroxydopamine, or the sensory neurotoxin, capsaicin.
I l l

Introduction
In the rat, NOS-immunoreactive nerve fibres are closely associated with adrenal
chromaffin and ganglion cells and cortical cells of zona glomerulosa as well as with
intra-adrenal blood vessels (Bredt et al. 1990; Sections 3:1:1-2). Such ample
distribution of the nitrergic nerve fibres in the gland suggests the possible presence of
an extrinsic input, since the number of local NOS-immunoreactive neurons, which
would account for intrinsic innervation, is relatively small (Sections 3 :l:l-2 ). In
addition, NOS-positive preganglionic neurons found in the intermediolateral cell
column of the spinal cord have been shown to have their terminals in the gland
(Blottner and Baumgarten 1992). The intra-adrenal distribution of extrinsic with
respect to intrinsic NOS-immunoreactive nerve fibres is, however, not known. Thus,
this study examined the intra-adrenal distribution of NOS-immunoreactive nerve
fibres following either section of the splanchnic nerve or demedullation. In addition, to
investigate the nature of NOS-immunoreactive adrenal nerves, the distribution of
NOS-immunoreactive nerves in the gland was also studied following chemical
denervation of sympathetic nerves with guanethidine or 6-OHDA and denervation of
primary sensory neurons with capsaicin.
112

Materials and Methods
Studies were carried out on male Sprague-Dawley rats. In each of six 15 week-
old rats, the left splanchnic nerve along with the suprarenal ganglion was removed as
described in Section 2:6:1. The rats were allowed to survive for 5 days postoperation.
In each of another six rats, which were 10 week-old, bilateral adrenal enucleations
were performed as described in Section 2:6:2. These rats were given physiological
saline solution instead of water for 10 days in order to maintain their electrolyte
balance in the absence of normal aldosterone secretion (Holzwarth 1984). The rats
were allowed to survive for 6 weeks postoperation.
Six other rats for each of guanethidine, 6-OHDA and capsaicin treatments were
also included in the study. Guanethidine and 6-OHDA treatments were carried out as
described in Section 2:7:2, while capsaicin treatment was done as described in Section
2:7:3. Guanethidine and capsaicin were treated neonatal, whereas 6-OHDA was
administered into adult rats.
As controls, adrenal glands from the following rats were studied: 6 normal non-
denervated rats; 3 sham-operated rats for each of the two surgical adrenal
denervations; and 6 rats of 3 groups, each group treated with either one of the solvents
in which guanethidine, 6-OHDA or capsaicin was dissolved. The adrenal glands from
the contralateral side of the splanchnic nerve section also served as controls.
Animals were sacrificed by ether asphyxiation and the adrenals were quickly
dissected out, bisected and immersion fixed in 4 % paraformaldehyde in 0.1 M PBS,
and cryostat sections were prepared as described in Section 2:3:1,
Immunohistochemistry for rabbit primary antibody against neuronal NOS (Euro-
Diagnostica, Malmo, Sweden) at a dilution of 1:2500 was carried out as described in
Section 2:3:2. On consecutive adrenal sections of the NOS-immunoreacted ones,
NADPH-diaphorase histochemistry was carried out as described in Section 2:3:3.
113

Results
In the adrenal glands from all groups of control rats NOS immunoreactivity was
observed in several intrinsic neurons located in the medulla and nerve fibres found in
both medulla and cortex, as described in Sections 3 :l: l-2 . In the medulla, the nerve
fibres were closely associated with chromaffin and neuronal cells and blood vessels
(Fig. 7a). However, in the adrenal medulla of the glands from the contralateral side of
the splanchnic nerve section, the NOS-immunoreactive nerve fibres associated with
the large NOS-immunonegative ganglion cells were found to be increased (Fig. 7b). In
the adrenal cortex, immunoreactive nerve fibres formed a plexus in the subcapsular
region and zona glomerulosa (Fig. 7c).
In the adrenal glands ipsilateral to the splanchnic nerve section side, only a few
immunoreactive nerve fibres were found near the chromaffin cells (Fig. 8a). The vast
majority of the nerve fibresjseen innervating the chromaffin and neuronal cells in the
medulla of control rat^ were absent. There were, however, occasional NOS-
immunoreactive nerve fibres found in association with the blood vessels (Fig. 8a).
Some of such nerve fibres were seen projecting from nearby reactive neurons, which
did not show any apparent change from that of the controls. Occasional
immunoreactive nerve fibres and moderate size nerve bundles were seen running
radially across the cortex (Fig. 8b). The immunoreactive nerve fibre plexus in the
subcapsular region and the zona glomerulosa of cortex was similar to that in controls
(Fig. 8c).
In sections of the demedullated adrenal glands a small number of
immunoreactive nerve fibres were seen in the connective tissue which replaced the
region of the enucleated medulla of the gland (Fig. 9a). Some of these nerve fibres
were closely associated with a few large blood vessels found in the region. A few large
immunoreactive nerve bundles were also seen running radially in the superficial region
of the cortex, some crossing the capsule (Fig. 9b). However, the type of the nerve fibre
114

plexus found in the subcapsular region and zona glomerulosa of the control rat adrenals
was not present.
No apparent change was observed in NOS immunoreactivity in adrenal gland
sections from rats treated with guanethidine, 6-OHDA or capsaicin with respect to the
controls.
NADPH-diaphorase-staining of the adrenal gland sections from all groups of
rats, both experimental and controls, closely matched their respective NOS
immunoreactivity.
115

Fig. 7. Fluorescence photomicrographs of NOS immunoreactivity in control
sections from: (a) adrenal medulla of normal non-denervated rat; (b) adrenal
medulla of the contralateral side of the splanchnic nerve section; and (c) adrenal
cortex of normal non-denervated rats. Note in b the high density of NOS-
immunoreactive nerve fibres around large non-immunoreactive ganglion cells
(arrows). In a and b arrowheads and arrows point to NOS-immunopositive and
immunonegative neuronal cell bodies, respectively. V: blood vessel; ZG: zona
glomerulosa; ZF: zona fasciculata. Bars = 52 fim.
116


Fig. 8. Fluorescence photomicrographs of NOS immunoreactivity in sections of
adrenal gland ipsilateral to the splanchnic nerve section side showing NOS-
immunoreactive nerve fibres are: (a) absent from medulla (M), but present in
relation to a blood vessel (V); (b) traversing radially the adrenal cortex; (c)
present as plexus in the zona glomerulosa (ZG) of cortex. Small arrows in a and
b point to immunoreactive nerve fibres, while arrowheads and large arrow in a
indicate NOS-immunopositive and immunonegative neurons, respectively. Co:
cortex; ZF: zona fasciculata; ZR: zona reticularis. Bars = 52//m.
117
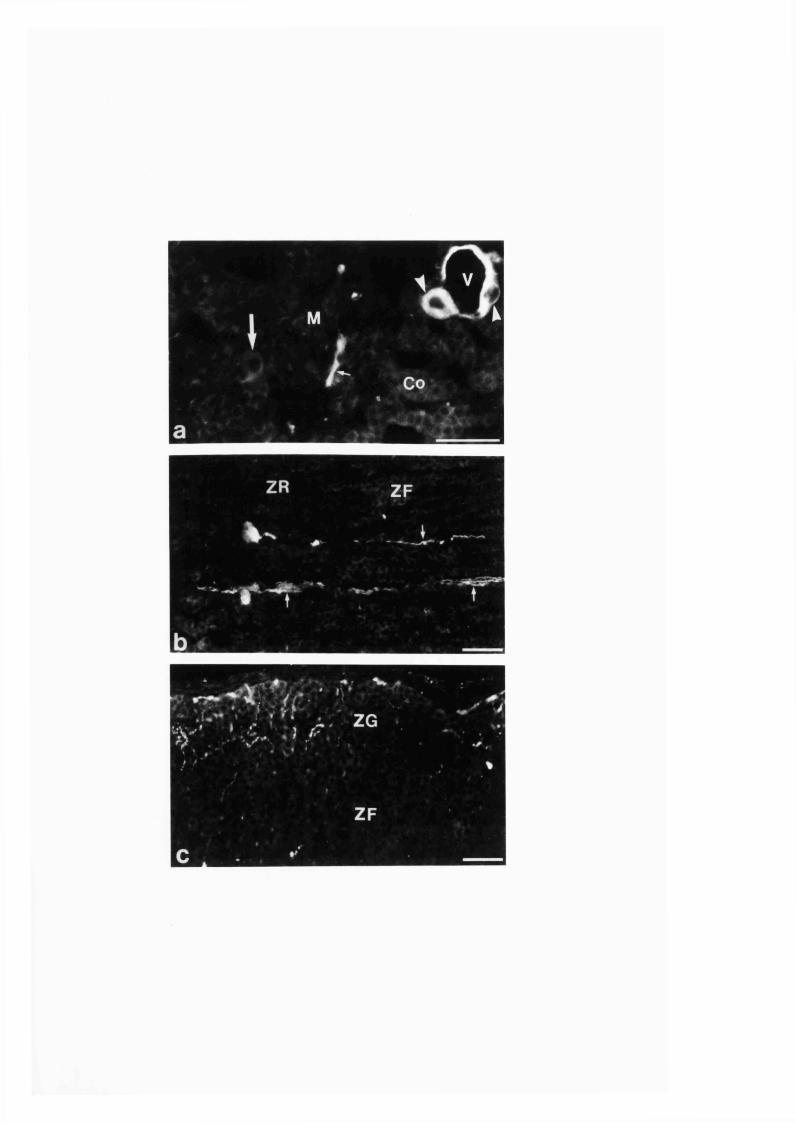

Fig. 9. Fluorescence photomicrographs of NOS immunoreactivity in sections of
demedullated adrenal gland, (a) NOS-immunoreactive nerve fibres (small
arrows) are seen in the connective tissue occupying the region of enucleated
medulla, and some of such fibres are associated with large blood vessels (V). (b)
NOS-immunoreactive nerve bundles (small arrows) are seen in the periadrenal
connective tissue, and one of such bundle is seen crossing the adrenal capsule
(Ca). Note the absence of immunoreactive nerve plexus from the subcapsular
region and the zona glomerulosa (ZG) of cortex. Open arrows in a point to
adrenal cortical cells. ZF: zona fasciculata. Bars = 52 pm.
118

4 '
4..' .. '.f

Discussion
The increase in NOS-immunoreactive nerve fibres observed innervating the
large ganglion cells in the adrenal gland from the contralateral side of section of the
splanchnic nerve is intriguing. It may be related to increased secretory activity of the
gland occurring to compensate for a reduced secretion from the denervated gland.
However, it is not clear at present as to the significance of such nitrergic innervation of
the ganglion cells in the glandular activity.
Both intrinsic as well as extrinsic origins for the NOS-immunoreactive intra
adrenal nerve fibres could be inferred from the present study. Those that innervate
chromaffin and neuronal cells appear to be extrinsic as shown by the disappearance of
these fibres following section of the splanchnic nerve. In contrast, the NOS-
immunoreactive nerve fibres found as plexus in the subcapsular region and the zona
glomerulosa of the cortex seem to arise from the intrinsic neurons. This is implied
since these fibres were unaffected by splanchnic nerve section, but disappeared upon
demedullation.
The nitrergic innervation of the adrenal vasculature also appears to originate
largely from the intrinsic NOS-immunoreactive neurons; indeed some immunoreactive
neurons have been seen to project to nearby vessels. This is also in agreement with the
finding that no apparent change in the vascular nitrergic innervation occurred
following section of the splanchnic nerve. In contrast, the existence of a small number
of immunoreactive nerve fibres associated with the vasculature in the fibrous
connective tissue occupying the central region of the demedullated gland suggests that
there may be an additional minor extrinsic innervation as well. It is possible that the
loss of such minor extrinsic nerve fibres may not be detected by splanchnic nerve
section alone. However, it is also possible that these fibres may not pass via the
splanchnic nerve. Instead, they may use an alternative route, probably running along
with the adrenal blood vessels, and therefore may have been missed during splanchnic
119

nerve section. Alternatively, such innervations of the blood vessels observed in the
demedullated gland may be a compensatory development for the loss of innervation
following removal of the intrinsic nerves.
The absence of any detectable change in the distribution of NOS
immunoreactivity in the adrenal gland of rats treated with any of guanethidine, 6-
OHDA or capsaicin indicate that the NOS-immunoreactive nerves in the gland are not
components of the sympathetic or primary sensory nerves which are affected by the
above drugs (Fumuss et al. 1970; Bumstock et al. 1971; Jancsô et al. 1977). It is also
evident from the present study that the loss of nerves sensitive to these drugs and found
within or outside the gland do not appear to have any secondary effect on the
distribution of NO-synthesizing nerves in the gland. The majority of the extrinsic
NOS-immunoreactive nerve fibres may be sympathetic preganglionic fibres as shown
previously with tracer injection into the gland and colocalizing NOS in the retrogradely
labelled neurons located in the intermediolateral cell column of the spinal cord
(Blottner and Baumgarten 1992). Further similar studies which also include the other
neural projection sites such as the sympathetic chain ganglia, dorsal root ganglia and
vagal ganglia (Parker et al. 1993) may be needed to fully elucidate the origin and
relative proportions of the extrinsic nitrergic nerve fibres to the gland.
120

SECTION 3:1:4
COLOCALIZATION OF NEUROPEPTIDES AND NADPH-DIAPHORASE
IN THE INTRA-ADRENAL NEURONAL CELL BODIES AND FIBRES OF
THE RAT
121

Abstract
Colocalization of vasoactive intestinal peptide, neuropeptide Y, calcitonin
gene-related peptide, substance P, and tyrosine hydroxylase, respectively, with
NADPH-diaphorase staining in rat adrenal gland was investigated using the double
labelling technique.
All vasoactive intestinal peptide- and some neuropeptide Y-immunoreactive
intrinsic neuronal cell bodies seen in the gland were double stained with NADPH-
diaphorase. Double labelling also occurred in some nerve fibres immunoreactive to
vasoactive intestinal peptide and neuropeptide Y in the medulla and cortex.
No colocalization of calcitonin gene-related peptide, substance P or tyrosine
hydroxylase immunoreactivity with NADPH-diaphorase staining was observed.
However, nerve fibres with varicosities immunoreactive for all the neuropeptides
examined were closely associated with some of the NADPH-diaphorase-stained
neuronal cell bodies. Thus, in rat adrenal gland, nitric oxide is synthesized in all
ganglion cells containing vasoactive intestinal peptide and in some containing
neuropeptide Y, but not in those containing calcitonin gene-related peptide, substance
P or tyrosine hydroxylase.
122

Introduction
In rat adrenal gland, NOS immunoreactivity is found in some intrinsic ganglion
cell bodies and nerve fibres. Immunoreactivity to TH, NPY and VIP has also been
demonstrated in the intra-adrenal ganglion cell bodies and nerve fibres of the rat
(Kondo 1985; Pelto-Huikko 1989; Dagerlind et al. 1990; Oomori et al. 1994).
However, it is not known whether NOS is coexpressed with either of these neuroactive
substances or whether it is found in other distinct population of nerves. Elsewhere,
neural NOS has been found colocalized with VIP, NPY, SP or enkephalin (Costa,
Furness, Pompolo, Brookes, Bomstein, Bredt and Snyder 1992; Kummer, Fisher,
Mundel, Mayer, Hoba, Philippin and Preissler 1992; Yamamoto, Bredt, Snyder and
Stone 1993; Heym et al. 1994; Tay, Moules and Bumstock 1994).
To investigate the situation in the rat adrenal gland, colocalization of neuronal
NADPH-diaphorase, which marks the site of NOS (Dawson et al. 1991; Hope et al.
1991; Bêlai et al. 1992; Hassall et al. 1992; Saffrey et al. 1992; also Sections 3 :l:l-2 ),
with the neuropeptides VIP, NPY, CGRP and SP, and the rate limiting enzyme in the
catecholamine synthesis, TH were carried out by a double labelling technique.
123

Materials and methods
A total of 40 adult Sprague-Dawley rats were used; 15 rats were
intraperitoneally injected with colchicine (Sigma, Poole, UK; Img/lOOg body wt, in
saline) whereas 10 other rats were injected with saline alone, 6 h prior to sacrifice. Rats
were killed t'f carbon dioxide and the adrenals were quickly dissected
out, bisected and immersion-fixed in 4 % paraformaldehyde in 0.1 M PBS, and
cryostat sections were prepared as described in Section 2:3:1. Immunohistochemistry
on every other consecutive adrenal section for rabbit polyclonal primary antibodies
against VIP (1:2000; Incstar, UK), NPY (1:3000; UCB, Belgium), CGRP (1:2000;
Cambridge Research Biochemicals, UK), SP (1:2000; Cambridge Research
Biochemicals) and TH (1:1000; Affinity, UK) was carried out as described in Section
2:3:2. Adrenal gland sections from 3 untreated, 3 colchicine-treated and 2 saline-
treated rats were studied for each of the antibody. Immunoprocessed adrenal sections
were inspected under fluorescence microscope and labelled cellular elements were
studied and photographed. Coverslips were removed and sections were rinsed in PBS.
All immunoreacted sections together with their respective fresh consecutive
sections were then processed for NADPH-diaphorase histochemistry as described in
Section 2:3:3.
124

Results
Examination of the immunoprocessed sections of the gland showed labelling in
cellular elements with the respective antibodies as described by previous investigators
(Kondo 1985; Pelto-Huikko 1989; Dagerlind et al. 1990; Oomori et al. 1994), whereas
control experiments in which the primary antibodies were omitted or substituted with
preimmune and non-immune sera resulted in no labelling. Immunoreactivity was
enhanced in the adrenal gland sections of rats pretreated with colchicine compared with
that of untreated glands.
Application of NADPH-diaphorase histochemistry to the VIP-immunoreacted
sections revealed that all VIP-immunoreactive intrinsic neuronal cell bodies observed
in the gland were also labelled positively for NADPH-diaphorase (Figs. lOa-b, d-e).
Such neuronal cell bodies were only occasionally observed and formed a minority of
the NADPH-diaphorase-stained neuronal cell bodies in the gland. Double labelling
was also observed in some VIP-immunoreactive nerve fibres seen in medulla and
cortex (Figs. 10b, e) and a few of those forming a plexus in the zona glomerulosa and
subcapsular region. However, the VIP-immunoreactive nerve fibres observed around a
small number of double labelled neuronal cell bodies did not stain for NADPH-
diaphorase (Figs. 10b, e). In addition, VIP-immunoreactive chromaffin cells were
negative for NADPH-diaphorase (Figs. 10c, f). Alternatively, some nerve fibres in the
medulla and cortex were negative for the VIP immunoreaction but positive for
NADPH-diaphorase staining (Figs. 10c, f).
Double labelling with NADPH-diaphorase was also observed in some NPY-
immunoreactive neuronal perikarya (Figs. 11a, d). The intensity of the NADPH-
diaphorase staining in these neurons was moderate compared with the intense staining
seen in most of the other NADPH-diaphorase-positive neurons. In addition, NADPH-
diaphorase staining and NPY immunoreactivity were seen in some nerve bundles found
radially traversing the cortex (Figs. 11b, e). However, colocalization occurred only in
125

occasional nerve fibres and varicosities within the bundles. A few nerve fibres in the
zona glomerulosa and subcapsular region were also double labelled for NPY and
NADPH-diaphorase. Conversely, no NADPH-diaphorase staining was found in NPY-
immunoreactive chromaffin cells (Figs. l la -f ) , or the majority of NPY-
immunoreactive ganglion cells (Figs. 11c, f) and nerve fibres. The NPY and NADPH-
diaphorase double labelled neuronal cell bodies appeared to be smaller than the NPY-
immunoreactive non-NADPH-diaphorase-stained neuronal cell bodies. Moreover,
NPY immunoreactivity in the former neuronal cell bodies was seen as homogeneous
immunofluorescence throughout the cytoplasm except the nucleus (Fig. 11a), in
contrast to the granular immunofluorescence mainly located at the perinuclear region
in the latter neurons (Fig. 11c). A small number of NPY-immunoreactive nerve fibres
were seen around some NADPH-diaphorase-stained neuronal cell bodies. The
occurrence of NADPH-diaphorase labelling in some NPY-immunonegative neuronal
cell bodies and nerve fibres was also observed.
Immunoreactivity to CGRP, SP or TH was not seen together with staining for
NADPH-diaphorase (Figs. 12a-b, c-d, e-f, respectively). However, nerve fibres with
varicosities immunoreactive for either CGRP or SP were found closely associated with
some NADPH-diaphorase-stained neuronal cell bodies (Figs. 12a-b, c-d,
respectively). The relative density of such innervation and the number of NADPH-
diaphorase-labelled neuronal cell bodies being innervated appeared to be higher for
CGRP than SP.
Similar observations to the above described results were also made by
comparison of the observations from studies carried out on consecutive sections
processed separately for NADPH-diaphorase and neuropeptides or TH.
126

Fig. 10. Photomicrographs of adrenal gland sections processed for VIP
immunoreaction (a-c) and NADPH-diaphorase double staining (d-f,
respectively). VIP-immunoreactive neuronal cell bodies (large arrowheads in
a, b) are double stained for NADPH-diaphorase (large arrowheads in d, e,
respectively), whereas an immunoreactive chromaffin cell (large arrow in c) is
NADPH-diaphorase-negative (large arrow in f). Some VIP-immunoreactive
nerve fibres seen in the cortex (small arrowheads in b) are double labelled with
NADPH-diaphorase (small arrowheads in e) but some others found around VIP
and NADPH-diaphorase double positive neuronal cell bodies (small arrow in b)
are not stained for NADPH-diaphorase (small arrow in e). On the other hand,
some nerve fibres that are VIP-immunonegative (small arrowheads in c) are
NADPH-diaphorase-positive (small arrowheads in f). Co: cortex; M: medulla.
Bar = 33 fjm.
127
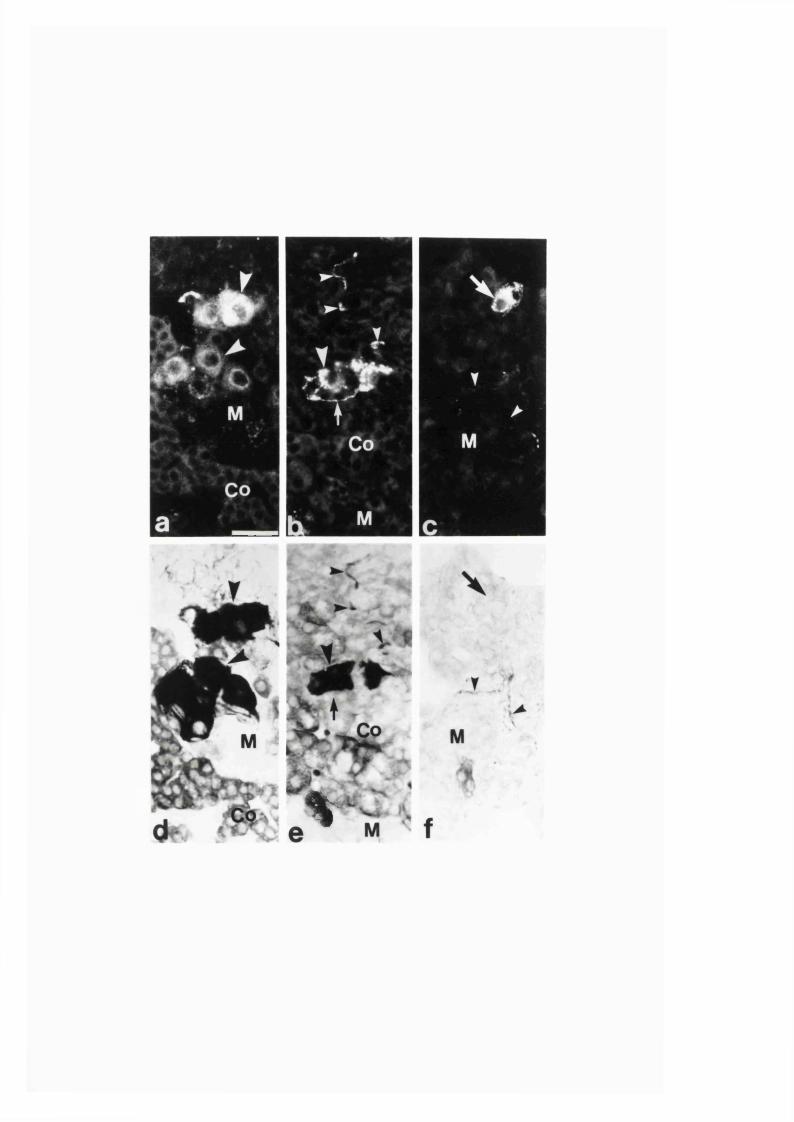
%

Fig. 11. Photomicrographs of adrenal gland sections immunoreacted for NPY
(a-c) and double stained for NADPH-diaphorase (d-f, respectively). NPY-
immunoreactive neuronal cell bodies (large arrowheads in a) and nerve bundles
(small arrowheads in b) are NADPH-diaphorase double positive (large
arrowheads in d and small arrowheads in e, respectively). Other NPY-
immunoreactive neuronal cell bodies (large arrows in c) are NADPH-
diaphorase-negative (large arrows in f). Note that NPY-immunoreacted
chromaffin cells found in medulla (a-c) are NADPH-diaphorase-negative (d-f,
respectively), whereas NADPH-diaphorase-positive cortical cells (e) are NPY-
immunonegative (b). Co: cortex; M: medulla. Bar = 33 ^m.
128
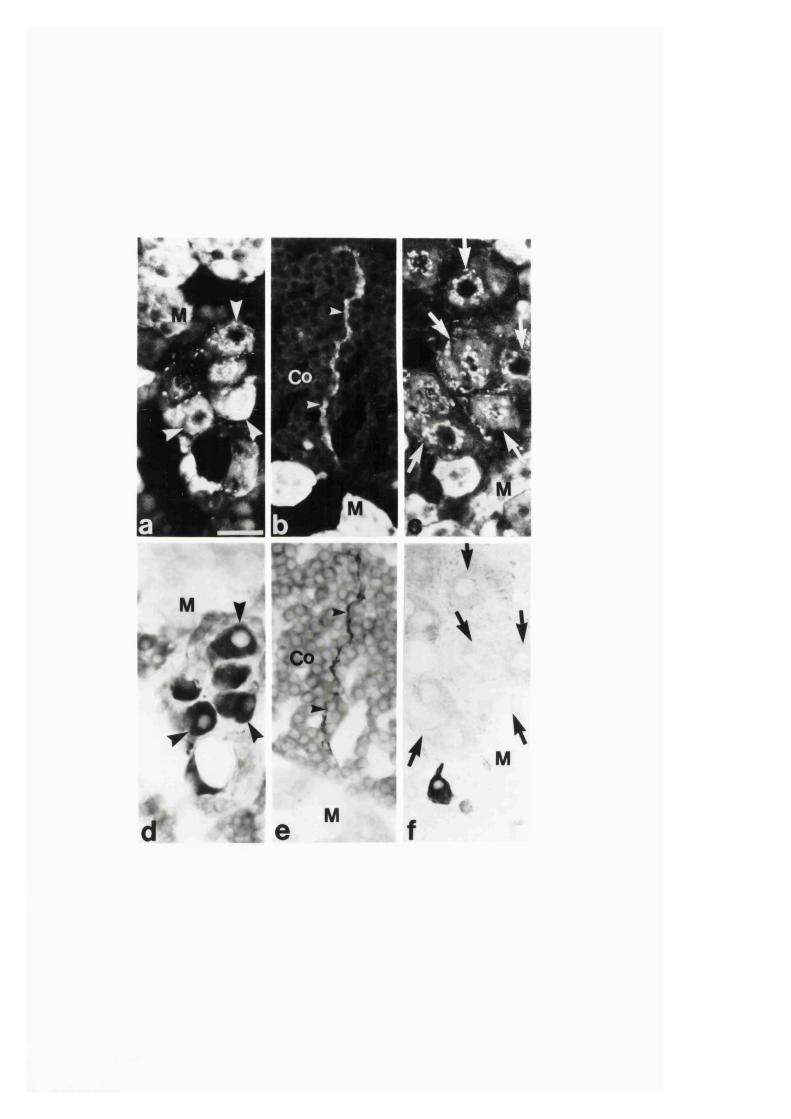

Fig. 12. Photomicrographs of:
a, b. Adrenal gland section immunoreacted for CGRP (a) and double stained for
NADPH-diaphorase (b) showing CGRP-immunoreactive but NADPH-
diaphorase-negative nerve fibres with varicosities {small arrowheads in a, b,
respectively) are associated with neuronal cell bodies that are CGRP-
immunonegative but NADPH-diaphorase-positive (large arrowheads in a, b,
respectively). A CGRP-immunoreactive chromaffin cell (large arrow in a) is
NADPH-diaphorase-negative (large arrow in b). M: medulla. Bar = 33 ywm.
c, d. SP immunoreactivity (c) and NADPH-diaphorase double staining (d) of an
adrenal gland section showing an SP-immunoreactive but
NADPH-diaphorase-negative nerve fibre with varicosities (small arrowheads
in c, d, respectively). This fibre approaches neuronal cell bodies that are
negative for SP but positive for NADPH-diaphorase (large arrowheads in c, d,
respectively). M: medulla. Bar = 33 /mi.
e, f. TH immunoreactivity (e) and NADPH-diaphorase double staining (f) of an
adrenal gland section. Non-TH-immunoreactive neuronal cell bodies (large
arrowheads in e) and nerve fibres (small arrowheads in e) are NADPH-
diaphorase-positive (large and small arrowheads in f, respectively). Note that
TH-immunoreactive chromaffin cells in the medulla are NADPH-diaphorase-
negative. The moderately NADPH-diaphorase-positive islands of cells seen
near the intensely NADPH-diaphorase-positive neuronal cells and fibres in f
are cortical cells, and note that they are TH-immunonegative in e. M: medulla.
Bar = 33 /mi.
129

1
%

Discussion
Neuronal NADPH-diaphorase staining and NOS immunoreactivity have been
shown to be identical and mark similar sites in various regions of central and peripheral
nervous tissue, including those found in the adrenal gland (Dawson et al. 1991; Hope et
al. 1991; Bêlai et al. 1992; Hassall et al. 1992; Saffrey et al. 1992; also Sections 3:1:1-
2). It is therefore evident from the present study that some intrinsic adrenal neurons
that synthesize NO are VIP-immunoreactive. A small number of these neurons are
innervated by VIP-immunoreactive NADPH-diaphorase-negative nerve fibres. This
suggests that VIP-containing fibres, most probably arising from non-NO-synthesizing
extra-adrenal neurons, influence some of the NO-producing neurons in the gland. In
this connection, stimulation of NO production by VIP has been shown during
relaxation of the gut (Grider 1993) and lung (Lilly, Stamles, Gaston, Meckel, Loscalzo
and Drazen 1993).
The other neuropeptide found to be colocalized in NO-synthesizing neurons of
the gland is NPY. There is no evidence from previous investigations that NPY and VIP
are colocalized in ganglion cells of the gland (Pelto-Huikko 1989; Maubert et al. 1990;
Oomori et al. 1994); therefore, it appears that these neuropeptides are contained in
separate subpopulations of NO-producing intrinsic neurons. At least some of the nerve
fibres double labelled for NPY and NADPH-diaphorase, and those double labelled for
VIP and NADPH-diaphorase, may arise from intrinsic double labelled neuronal cell
bodies with the respective markers. However, an extrinsic input cannot be ruled out.
Unlike in the intra-adrenal neuronal cell bodies, NPY and VIP have been found to be
colocalized in some nerve fibres that are regarded as extrinsic and that are mainly
found in the superficial region of the cortex (Maubert et al. 1990; Oomori et al. 1994).
Some of these fibres also show TH immunoreactivity (Oomori et al. 1994). Although it
is possible that NADPH-diaphorase may also coexist in some of the nerve fibres
containing VIP and NPY, the lack of NADPH-diaphorase staining in the TH-
130

immunoreactive nerve fibres implies that it is not found in those fibres that also contain
TH.
The absence of CGRP or SP immunoreactivity in the rat adrenal ganglion cells
suggests that intra-adrenal nerve fibres immunoreactive to either of these
neuropeptides are extrinsic. As there are a large number of CGRP-immunoreactive and
SP-immunoreactive neurons in the dorsal root ganglia (Gibson, Polak, Bloom, Sabate,
Mulderry, Ghatei, McGregor, Morrison, Kelly, Evans and Rosefeld 1984), and because
most of the intra-adrenal immunoreactive nerve fibres for these neuropeptides
disappear following capsaicin treatment (Pelto-Huikko 1989), nerve fibres containing
CGRP and SP in the gland are likely to be sensory fibres. It has recently been
demonstrated that the adrenal gland of rat receives a substantial sensory innervation
from dorsal root and sensory vagal ganglia (Parker et al. 1993); some of the neurons in
the former ganglia contain SP (Zhou, Oldfield and Livett 1991). The observation of the
CGRP-containing or SP-containing nerve fibres in association with the NO-
synthesizing neurons may therefore indicate their sensory innervation.
The absence of colocalization of TH immunoreactivity and NADPH-diaphorase
staining suggests that NO is not synthesized by noradrenaline-containing neurons in
the gland. Furthermore, the large-sized ganglion cells that have been shown to be
noradrenergic (Dagerlind et al. 1990) are negative for NADPH-diaphorase staining.
Thus, NO and noradrenaline appear to be produced in different intra-adrenal neurons.
The colocalization of NADPH-diaphorase with VIP and NPY immunoreactivity
in the intra-adrenal nerves of the rat demonstrated in the present study is different from
that in the human gland (Heym et al. 1994) where immunoreactivity to SP in the
nerves, and VIP and enkephalin in the chromaffin cells are coexpressed with NOS. The
distribution of NOS and its colocalization with other neuroactive substances in adrenal
gland thus show species variation.
In conclusion, NO-synthesizing intra-adrenal nerves of the rat appear to
131

contain VIP or NPY, but not SP or CGRP, and are not noradrenergic. The evidence that
only subpopulations of the NADPH-diaphorase-stained nerves immunoreact for NPY
or VIP, or receive distinct peptidergic innervation indicates the diversity of the NO-
synthesizing nerves found in the gland.
132

SECTION 3:1:5
CALRETININ IMMUNOREACTIVITY IN ADRENAL GLAND OF
DEVELOPING, ADULT AND AGING SPRAGUE-DAWLEY RATS
133

Abstract
Localization of cairetinin immunoreactivity in the rat adrenal gland was studied
using immunohistochemical methods. Calretinin-immunoreactive adrenal chromaffin
cells and nerve fibres were found at all of the ages examined from embryonic day 16
up to 2 year-old. Immunoreactive chromaffin cells showed a decrease in number with
increasing age, and in the adult and aging rats they were found among the adrenaline-
storing chromaffin cells. Cairetinin immunofluorescence was also observed in a few
ganglion cells of the adult and aging rats, and in most of the cortical cells at embryonic
day 16. The density of immunoreactive nerve fibres in the gland showed a progressive
increase with age to a peak by day 4 after birth and then a gradual decrease afterwards;
they became associated mainly with some of the ganglion cells in the adult rat. As
revealed by double labelling of cairetinin immunoreactivity in adrenal sections from
the adult rat with NADPH-diaphorase histochemistry, no colocalization existed
between cairetinin and NADPH-diaphorase; however, a small number of neuronal cell
bodies which contained nitric oxide synthase were found heavily surrounded with
calretinin-immunoreactive nerve fibres. In aging rats, an increased density of
calretinin-immunoreactive nerve fibres was seen associated largely with the
chromaffin cells.
It is concluded that cairetinin is found in the rat adrenal gland; the high degree
of variability in its expression with age suggests that cairetinin may be concerned with
age-related activities in the gland.
134

Introduction
Cairetinin is a 29 kDa, more recently discovered member of a family of calcium
binding proteins (Rogers 1987; Winsky, Nakata, Martin and Jacobowitz 1989). It is
mainly found in the nervous tissue and structurally related to the 28 kDa calbindin,
with 58 % amino acid sequence homology (Rogers 1987). Immunohistochemically,
cairetinin has been localized in neurons with different features found in several regions
of both central (Rogers 1987, 1989; Winsky et al. 1989; Résibois and Rogers 1992;
Ren, Ruda and Jacobowitz 1993) and peripheral nervous tissue (Rogers 1989; Brookes,
Steele and Costa 1991; Siou, Belai and Bumstock 1992). The precise function of
cairetinin has not yet been recognized, although it is thought that it may have
additional cellular function apart from just buffering intracellular calcium
concentration (Rogers 1991; Anderssen, Bliimcke and Celio 1993).
Identification of its occurrence in further tissues and examining changes in its
distribution in development and aging may give a clue to its function. Accordingly, this
study investigated immunohistochemically its localization in adrenal glands of adult
rats, and examined the changes that occur during development and aging. Furthermore,
by applying NADPH-diaphorase histochemistry, which demonstrates the NO-
synthesizing neurons (Dawson et al. 1991; Hope et al. 1991; Belai et al. 1992; Hassall
et al. 1992; Saffrey et al. 1992; also Sections 3 :l:l-2 ) , onto the cairetinin
immunoreacted sections, this study also has examined if there is any structural
relationship between calretinin-immunoreactive elements and the NOS-containing
nerves recently described in the rat adrenal gland (Bredt et al. 1990; Dawson et al.
1991; also Sections 3 :l:l-2 ).
135

Materials and methods
Male (postnatal) and either sex (prenatal) of Sprague-Dawley rats were used.
Five rats from each of the following ages were studied: developing rats of E l 6, E18,
E20, 1, 4, 7, 14, and 21 days; adult rats of 16 weeks and aging rats of 24 months. The
age of the developing rats was determined by counting the appearance of the cervical
mucous plug as EO and the day of delivery as day 0 of postnatal life. Animals were
sacrificed by carbon dioxide asphyxiation. Foetuses from the pregnant rats were
removed and their adrenals along with those from rats at the other ages were dissected
out and immersion fixed in 4 % paraformaldehyde in 0.1 M PBS and cryostat sections
were cut as described in Section 2:3:1. Immunohistochemistry for rabbit primary
antibody against cairetinin (gift from J. Rogers, Cambridge, UK) at a dilution of
1:5000 was carried out as described in Section 2:3:2.
In the adrenal gland sections at E l6, where the chromaffin and cortical cells are
found intermingled with each other, in order to identify between these cell types some
sections were also incubated as above in a mixture of the antibody against cairetinin
and a mouse monoclonal antibody against protein gene product 9.5 (PGP 9.5; 1:2000;
Ultraclone, UK). PGP 9.5 is a cytosolic neuronal and neuroendocrine ubiquitin and
used as a general marker for neuronal and neuroendocrine cells (Thompson, Doran,
Jackson, Dhillon and Rode 1983; Wilson, Barber, Hamid, Power, Dhillon, Rode, Day,
Thompson and Polak 1988; Wilkinson, Lee, Deshpande, Duerksen-Hughes, Boss and
Pohl 1989). Previously, PGP 9.5 has been used for a similar study in the adrenal gland
of rat where it has been found labelling the chromaffin but not the cortical cells (Kent
and Coupland 1989). In the latter preparations the sites of antibody-antigen reaction
were revealed by incubation with biotinylated sheep anti-mouse antibody (1:250;
Amersham) followed by streptavidin fluorescein (1:100) for PGP 9.5 and rhodamine
conjugated goat anti-rabbit antibody (1:40; Nordic Immunological Laboratories, UK)
for cairetinin.
136

Immunoreacted sections were inspected using a fluorescence Zeiss microscope.
In order to examine whether calretinin-immunoreactive chromaffin cells have any
correlation with either A or NA cells, regions of adrenal medulla with labelled
chromaffin cells were examined with a filter for paraformaldehyde-induced
catecholamine fluorescence.
To investigate any structural relationship that exists between calretinin-
immunoreactive cellular elements and the NO-synthesizing nerves in the gland,
coverslips were removed from immunoprocessed sections and NADPH-diaphorase
histochemistry was applied onto the calretinin-immunoreacted sections from the adult
rats as described in Section 2:3:3.
137

Results
No labelling was found in control experiments in which the primary antibodies
were omitted.
Developing rat
In adrenal glands from rats at E l 6 most of the chromaffin and cortical cells gave
positive calretinin immunoreactivity (Fig. 13a, b). The fluorescence intensity among
the labelled cells varied, with a smaller proportion being intensely stained than the rest
of the cells. As revealed by the colocalization studies with the PGP 9.5, the intensely
labelled cells were cortical cells (Figs. 13a, b). In the intensely labelled cells the
immunofluorescence was homogeneously located throughout the cell cytoplasm,
including the nucleus. In the adrenal gland sections from rats at the remaining ages of
development the occurrence of immunoreactive chromaffin cells progressively
declined with increasing age and no labelled cortical cell was found.
A small number of weakly reactive nerve fibres was already present at E16. The
frequency of labelled nerve fibres progressively increased with age reaching a peak by
the 4^ ̂day after birth (Fig. 13c) and then decreasing with age afterwards. The reactive
fibres were seen running in bundles and single nerve fibres within the gland, and a few
were found associated with chromaffin and ganglion cells.
Adult rat
Calretinin-immunoreactive chromaffin cells were also found in sections of
adrenal glands from the adult rats (Fig. 14a, c, g). In most of the adrenal medullary
sections up to 5 labelled chromaffin cells were found singly dispersed or in a small
group. As evidenced from paraformaldehyde-induced catecholamine fluorescence, the
immunoreactive cells were always found among the A cells (Figs. 14a, b). A few
ganglion cells positively labelled with calretinin immunoreactivity were also observed
138

(Fig. 14c). However, unlike in the reactive chromaffin cells, the intensity of the
immunofluorescence in the neuronal perikarya was weak and the outline of the nucleus
was not labelled.
Occasional calretinin-immunoreactive nerve bundles were seen traversing
radially across the cortex towards the medulla (Fig. 14d). In the medulla, calretinin-
immunoreactive nerve fibres with varicosities and terminals of varying density were
seen accumulated around immunoreactive (Fig. 14c) and non-immunoreactive intra
adrenal ganglion cell bodies. A small number of the non-immunoreactive ganglion
cells which were densely surrounded by calretinin-immunoreactive nerve fibres were
found to be NADPH-diaphorase-positive (Figs. 14e, f), while the majority of the
ganglion cells which received a moderate degree of innervation were NADPH-
diaphorase-negative (Figs 14g, h). Calretinin-immunoreactive neuronal perikarya
were not NADPH-diaphorase reactive. Only a few calretinin-immunoreactive nerve
fibres were seen in relation to the chromaffin cells of the adrenal medulla of the adult
rats.
Aging rat
The occurrence of calretinin-immunoreactive chromaffin cells in the adrenals of
aged rats was reduced compared to that in the adult rat, and observed only
occasionally. However, similar to those in the adult rat, such labelled cells occurred
among the A cells, and a few ganglion cells were seen with faint immunofluorescence.
Conversely, an increased number of immunoreactive nerve fibres with varicosities and
endings were found in close association with some adrenal chromaffin cells (Fig. 15),
in addition to those found in relation to ganglion cells.
139

Fig. 13a-c. Photomicrographs of adrenal gland sections of developing rats at (a,
b) E l 6 and (C) 4 day-old. Figures a and c show calretinin immunoreactivity,
while Figure b shows immunoreactivity for PGP 9.5 on same section as a. Note
the occurrence of calretinin immunoreactivity in cells of both developing
adrenal medulla (M) and cortex (Co), and that some cortical cells (arrows) are
intensely calretinin-immunoreactive (a, b). Note also in c the calretinin-
immunoreactive nerve fibres run singly as well as in bundles (arrows). Bar = 33
fjm.
140
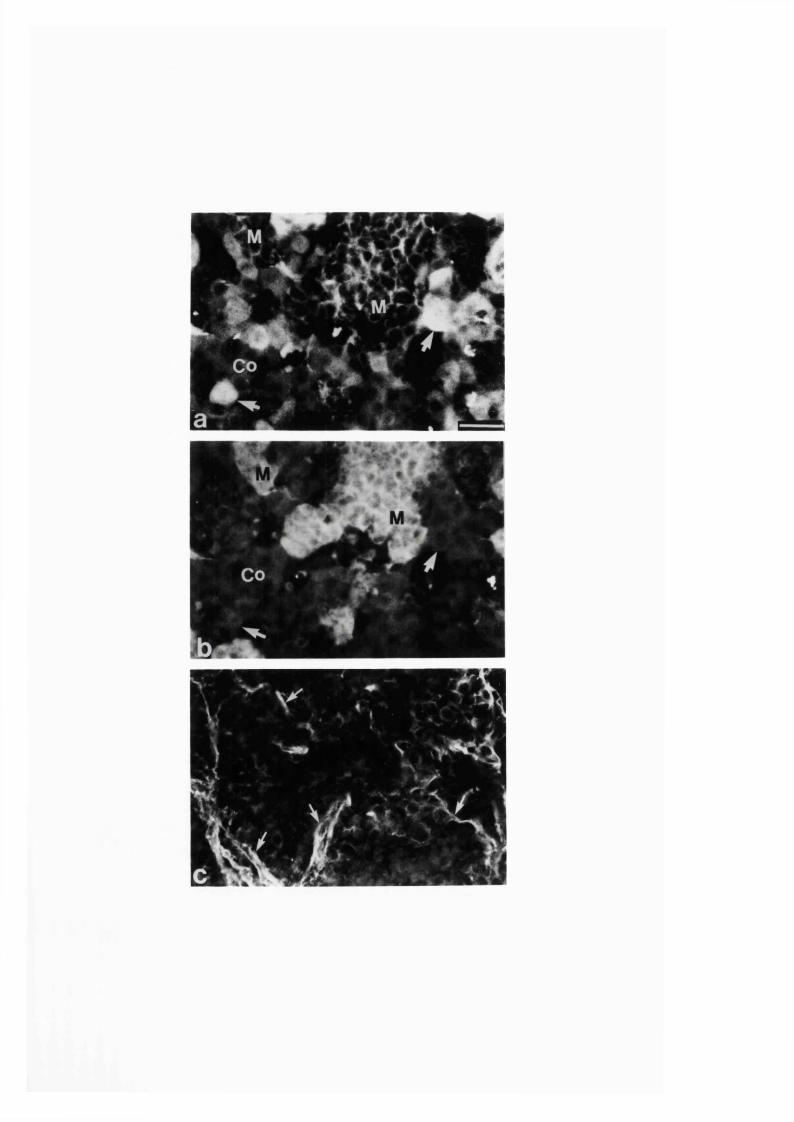

Fig. 13a-c. Photomicrographs of adrenal gland sections of developing rats at (a,
b) E l6 and (C) 4 day-old. Figures a and c show calretinin immunoreactivity,
while Figure b shows immunoreactivity for PGP 9.5 on same section as a. Note
the occurrence of calretinin immunoreactivity in cells of both developing
adrenal medulla (M) and cortex (Co), and that some cortical cells (arrows) are
intensely calretinin-immunoreactive (a, b). Note also in c the calretinin-
immunoreactive nerve fibres run singly as well as in bundles (arrows). Bar = 33
140
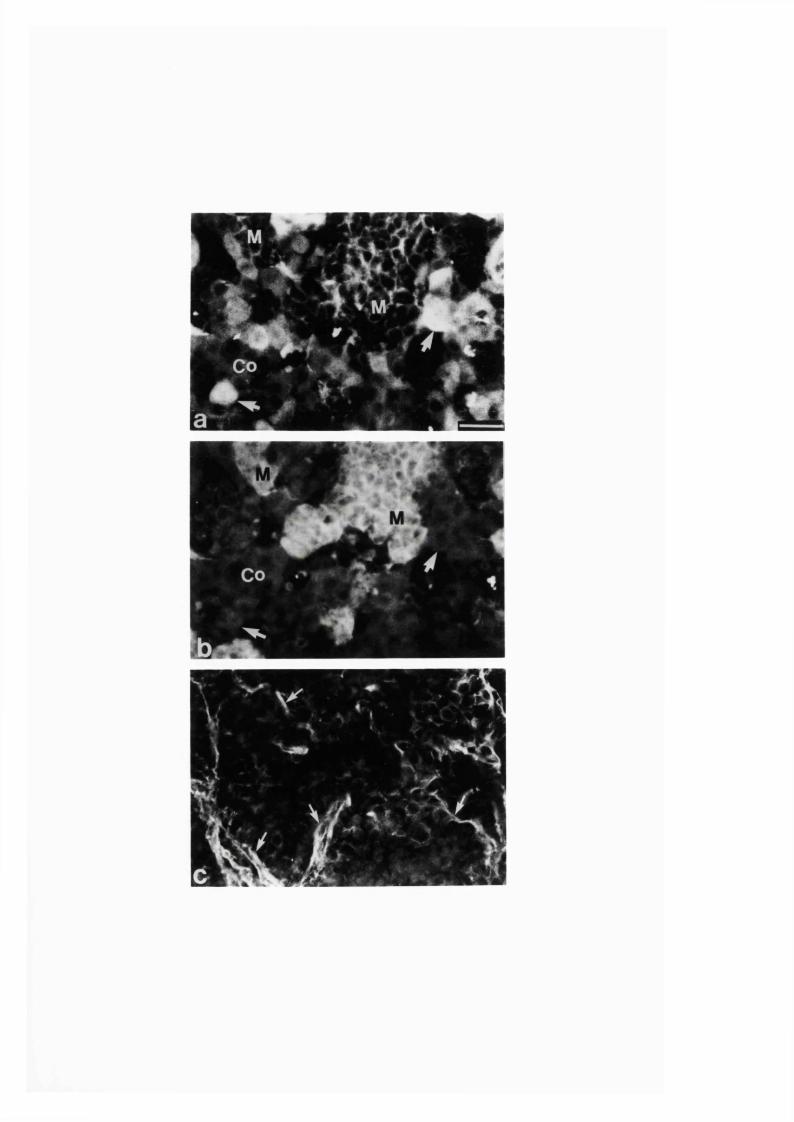

Fig. 14a-h. Photomicrographs of sections of adrenal medulla (a-c, e-h) and
cortex (d) from 16 week-old rats. A group of calretinin-immunoreactive
chromaffin cells (arrow in a) are seen among the adrenaline-storing chromaffin
cells (A in b) as shown with paraformaldehyde-induced catecholamine
fluorescence on the same section as a. An immunoreactive ganglion cell
(arrowhead in c) is shown which is approached by immunoreactive nerve fibres
(small arrows in c). Calretinin-immunoreactive nerve bundles (small arrows in
d) are seen radially crossing the outer region of the adrenal cortex. Figures e-h
show calretinin immunoreactivity (e, g) and the same sections double stained for
NADPH-diaphorase (f, h, respectively). Note the calretinin-negative cells
(arrowheads in e) surrounded by dense calretinin-immunoreactive nerve fibres
(small arrows in e) are NADPH-diaphorase-positive neurons (arrowheads in f).
However, the calretinin-negative cells (arrowheads in g) moderately
surrounded by calretinin-immunoreactive nerve fibres (small arrows in g) are
NADPH-diaphorase-negative neurons (arrowheads in h). Note also that
calretinin-immunoreactive chromaffin cell (large arrow in g) is not stained for
NADPH-diaphorase (large arrow in h). Large and small arrows show
immunoreactive chromaffin cells and nerve fibres, respectively, while
arrowheads point to ganglion cells. A: adrenaline-storing cells; N:
noradrenaline-storing cells; Ca: capsule; Co: cortex. Bar = 33 fjm.
I4l
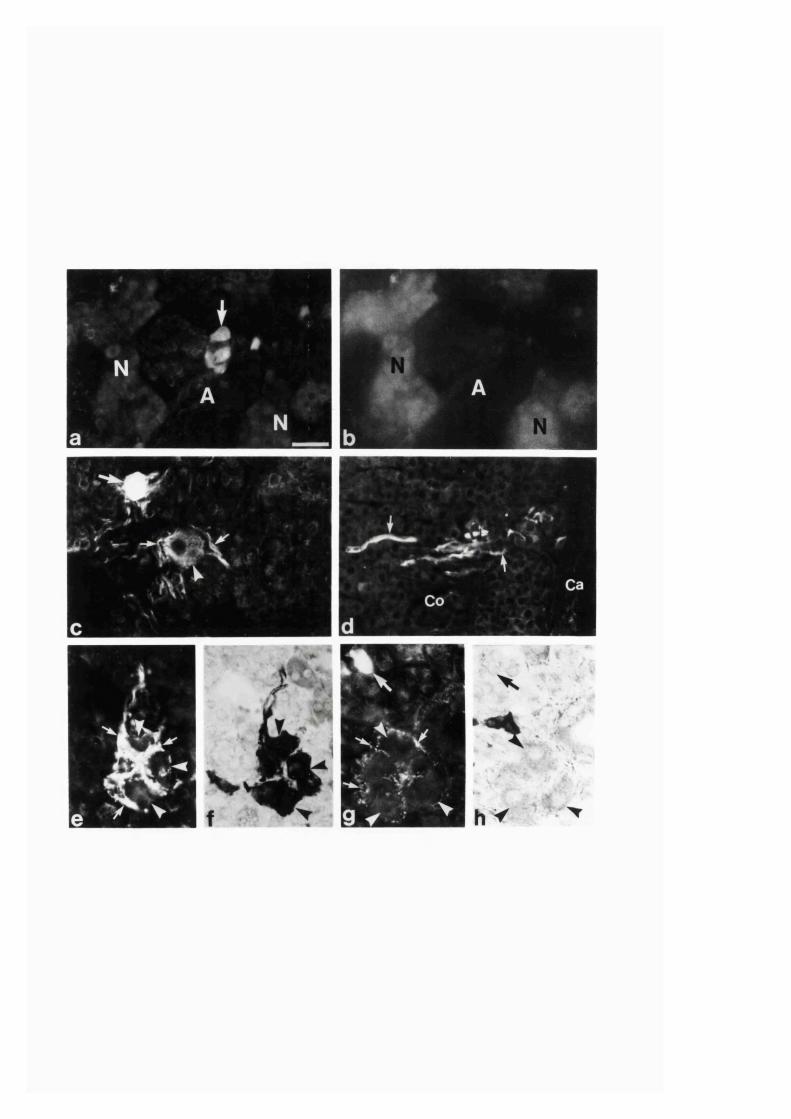
- V '
T é.

Fig. 15. Photomicrograph of calretinin immunoreactivity in section of adrenal
medulla from 24 month-old rat showing immunofluorescent nerve endings and
varicosities (arrows) associated with chromaffin cells. Bar = 33 yum.
142


Discussion
The present study has demonstrated that calretinin-immunoreactive cell bodies
and nerve fibres are present in the adrenal gland of the rat. Fixation with 4 %
paraformaldehyde allows, under the appropriate filter, the identification of fluorescing
NA cells from non-fluorescing A cells (Eranko 1992, 1955). This procedure enabled
us to note that the calretinin-immunoreactive chromaffin cells in the adrenal glands of
the adult and aging rats occurred among the A cells. However, since the
paraformaldehyde-induced fluorescence was so weak in glands from the developing
rats, such a correlation of the labelled cells to either type of the chromaffin cell was not
possible. It is interesting that the occurrence of calretinin in a subpopulation of A cells
in the adult and aging rats is different fi"om its closely related calcium binding protein,
calbindin, which exclusively occurs in the NA cells of rat adrenal gland (Buffa, Mare,
Salvadore, Solcia, Furness and Lawson 1989).
As bundles of calretinin-immunoreactive nerve fibres were seen running across
the cortex towards the medulla, and since only a small number of neurons showed
weak calretinin immunoreactivity, it seems likely that intra-adrenal reactive nerve
fibres are mainly extrinsic in origin. Although the adrenal medulla receives its major
innervation from sympathetic preganglionic neurons in the intermediolateral cell
column (Kesse et al, 1988), there are additional nerve fibres projecting to the gland
from postganglionic sympathetic, as well as parasympathetic and sensory, ganglia
located at various sites (Parker et al. 1993). The intra-adrenal calretinin-
immunoreactive nerve fibres may thus originate from either, or combinations of, these
regions. In fact, calretinin immunoreactivity has been demonstrated in neurons found
in the lateral cell column of the spinal cord as well as in dorsal root ganglia of the rat
(Ren et al. 1993). Retrograde fibre tracing combined with colocalization studies may
be required to determine the source and the relative proportion of the projections of the
calretinin-immunoreactive fibres in the gland.
143

Although at the early period of development calretinin-immunoreactive nerve
fibres in the gland were also associated with chromaffin cells, at later stages of
development and in the adult they were mainly confined to some ganglion cells. A
smaller number of such ganglion cells receiving a higher level of innervation are NO-
synthesizing neurons as compared to the moderately innervated ones which do not
synthesize NO. The functional implication of such specificity in the innervation of the
intrinsic neurons of the adrenal gland is not known at present and is a matter of future
investigation. On the other hand, in the aged rat adrenals in addition to the ganglion
cells, some chromaffin cells also received immunoreactive nerve fibres. The adrenal
medulla of aging rats shows signs of proliferative lesions and differentiation of the
chromaffin cells towards a neuronal phenotype (Tischler et al. 1985; Coupland and
Tomlinson 1989) as well as changes in its pattern of cholinergic (Tischler et al. 1985)
and nitrergic (Section 3:1:2) innervation. It is interesting that such changes in
innervation also occur with calretinin-immunoreactive nerve fibres. However, the
cause and effect of the change in the innervation with calretinin-containing nerves, and
whether such a change of innervation is associated with those chromaffin cells affected
in aging remains to be established.
The function of calretinin in the adrenal gland, as in other regions, is not yet
known. Nevertheless, several possible roles in cellular activities may be considered.
One suggestion is that calretinin is involved in activities such as short term intracellular
calcium buffering. Changes in intracellular levels of calcium are known to modulate
several important processes including cell metabolism, cell division, cell movement,
nerve impulse conduction and exocytosis (Campbell 1983; Carafoli and Penniston
1985; Miller 1988). However, in excess, calcium is toxic and causes cell death
(Schanne, Kane, Young and Farber 1979). In a protective role against such damage,
calretinin may be one of the calcium binding proteins involved in calcium buffering.
Indeed, recently it has been shown that calretinin-immunoreactive cultured rat brain
1#

cortical neurons are more resistant to calcium overload and excitotoxicity as compared
to those which are not calretinin-immunoreactive (Lukas and Jones 1994). The
increase in calretinin-immunoreactive adrenal cells and nerve fibres during early
development as observed in this study may be related to an increase in calcium levels
as required by the cells for developmental activities such as cell division and cell
migration occurring during this stage. The observation of calretinin immunoreactivity
in the adrenal cortical cells at E l6 may also indicate involvement of calretinin in early
developmental activities. During development the temporary occurrence of calretinin
has also been described in several neurons as well as in non-neuronal cells including
photoreceptors and ependymal cells of the brain (Ellis, Richards and Rogers 1991;
Anderssen et al. 1993; Bastianelli and Pochet 1993). However, as calretinin
immunoreactivity is found in distinct subpopulations of cells and nerve fibres,
calretinin may alternatively have additional unknown functions. In such cases, the
variability in the occurrence of calretinin immunoreactivity in cell bodies and nerve
fibres of the adrenal gland during different ages may be related to the appearance and
disappearance of cells due to cell division and cell death, respectively.
In conclusion, the present study reveals that calretinin-containing cell bodies
and nerve fibres show dramatic changes of expression in development and aging in the
adrenal gland of the rat. The plasticity in the distribution of calretinin with age may be
indicative of age-related functions in the gland.
145

SECTION 3:2
CHANGES IN THE EXPRESSION OF NITRIC OXIDE SYNTHASE
AND NADPH-DIAPHORASE IN THE ADRENAL GLAND OF RATS IN
DEVELOPMENT, AGING AND ALTERED ADRENAL GLAND ACTIVITY
146

SECTION 3:2:1
EFFECT OF RESERPINE TREATMENT AND HYPOPHYSECTOMY ON THE
NITRIC OXIDE SYNTHASE IMMUNOREACTIVITY AND NADPH-
DIAPHORASE STAINING IN THE RAT ADRENAL GLAND
147

Abstract
Nitric oxide synthase immunoreactivity and NADPH-diaphorase staining in the
adrenal gland of rats subjected to either reserpine treatment or hypophysectomy as
compared with the controls were studied. Reserpine treatment caused a significant
decrease in immunoreactivity to the neuronal isoform of Nitric oxide synthase in the
nerve fibres that innervate adrenal medullary cells. Hypophysectomy did not cause any
change in the Nitric oxide synthase immunoreactivity, although it eliminated most of
the NADPH-diaphorase staining non-NOS-immunoreactive cortical cells. The
reduction in Nitric oxide synthase, and hence decreased production of Nitric oxide in
the adrenal gland of reserpine-treated rats may be related to the increased biosynthesis
and secretion of catecholamines following treatment of the drug.
148

Introduction
The distribution and age related changes of the NO-synthesizing nerves in the
adrenal gland of rat has been described (Sections 3:1:2). Changes in the expression of
the NOS isoform has also been detected in sensory ganglia and central nervous system
neurons under various conditions (Verge, Xu, Xu, Weisenfeld-Hallin and Hokfelt
1992; Wu 1993; Bredt and Snyder 1994; Sessa 1994). It is not known if the nitrergic
innervation of the adrenal gland is also changed in different conditions that cause
changes in the activity of its innervation and glandular secretion. In the study reported
here the state of neuronal NOS immunoreactivity in the adrenal gland of rats was
assessed following reserpine treatment or hypophysectomy. Reserpine causes depletion
of adrenal catecholamine and increases its biosynthesis via a transynaptic mechanism
(Thoenen et al. 1969). Hypophysectomy is known to cause a major atrophy of adrenal
cortical cells, and thereby diminishes the secretion of glucocorticoids (Sayers 1950)
which have a regulatory influence on adrenal medullary catecholamines and peptide
synthesis and secretion (Wurtman and Axelrod 1966; Youbum et at. 1987).
149

Materials and Methods
15 week-old Sprague-Dawley rats were injected subcutaneousiy with 2.5
mg/kg reserpine as described in Section 2:7:1 on day 1, 3, 5, and 7 and killed on day 8.
14 week-old hypophysectomized, sham-hypophysectomized and non-operated
control Sprague-Dawley rats were obtained from Charles River Ltd, and allowed to
survive for two weeks. Six rats were studied for each of the experiments.
Animals were sacrificed by ether asphyxiation and the adrenal glands were
quickly dissected out, bisected and immersion fixed in 4 % paraformaldehyde in 0.1 M
PBS for 2 h and cryostat sections were prepared as described in Section 2:3:1.
NOS immunohistochemistry against the neuronal NOS (1:2500; Euro-
Diagnostica, Sweden) and NADPH-diaphorase histochemistry were carried out as
described in Sections 2:3:2 and 2:3:3, respectively. Quantitative analysis of the NOS-
immunoreactive nerve fibres in every 10^ ̂ consecutive sections of the adrenal glands,
in both the medulla and the zona glomerulosa and subcapsular region of the cortex,
was made using a computer image analyzer as described in Section 2:4. To determine
whether significant differences existed among the means from each groups, statistical
analysis was performed using one way analysis of variance with Fisher's least
significant difference procedure.
150

Results
In the adrenal glands from all groups of control rats NOS immunoreactivity was
observed in several intrinsic neuronal cell bodies located in medulla and in nerve fibres
found in both medulla and cortex, as described in Sections 3 :l: l-2 . In the medulla, the
nerve fibres were closely associated with chromaffin cells (Fig. 16a), neuronal cells
and blood vessels. In addition, the immunoreactive nerve fibres formed a plexus in the
subcapsular region and the zona glomerulosa of cortex.
In sections of the adrenal gland of the reserpine-treated rats, NOS-
immunoreactive nerve fibres that innervate the adrenal medullary chromaffin cells and
also those associated with the neuronal cell bodies were significantly reduced (Fig.
16b; Table 2) as compared with that of the controls (Fig. 16a). However, no change
was found in the NOS-immunoreactive nerve fibres that form a plexus in the
subcapsular region and zona glomerulosa (Table 2). The NOS-immunoreactive nerve
fibres found in association with blood vessels and the NOS-positive intrinsic neuronal
cell bodies were also unaffected in the reserpine-treated rats.
The weight of adrenal glands from hypophysectomized rats was reduced by
about 76 % as compared with the controls. However, there was no significant change
in the NOS immunoreactivity in the adrenal gland of the hypophysectomized rats
(Table 2).
NADPH-diaphorase-staining of the neuronal elements in the sections of
adrenal gland from all groups of rats both experimental and controls, closely matched
the NOS immunoreactivity. The adrenal cortical cells were also positively stained.
NADPH-diaphorase stained and non-NOS-immunoreactive cells that are found as
islands or singly dispersed in the medulla of the glands from the control rats were not
present in the medulla of the hypophysectomized rats (Figs. 17a, b). There was also a
substantial loss of the cortical cells of the zonae fasciculata and reticularis of the
adrenal cortex of the hypophysectomized rats.
151

Fig. 16. Fluorescence photomicrographs of NOS immunoreactivity in sections
of adrenal medulla of (a) control and (b) reserpine-treated rats. Arrows and
arrowheads indicate NOS-immunoreactive neuronal cell bodies and nerve
fibres, respectively. Note the absence of the large majority of NOS-
immunoreactive nerve fibres in the medulla of the reserpine-treated rat (b) as
compared with that of the control rat (a). Bar: 33 fm .
152
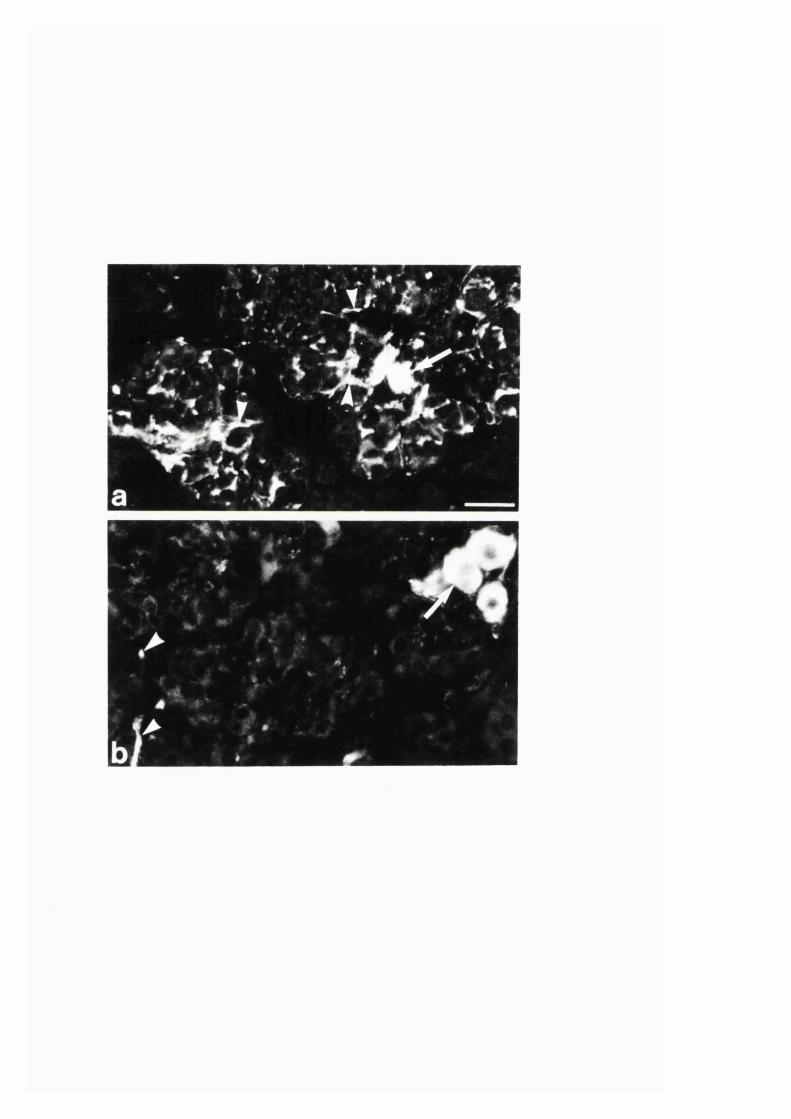

Fig. 17. Photomicrographs of NADPH-diaphorase stained adrenal gland
sections of (a) control and (b) hypophysectomized rats. Note that NADPH-
diaphorase stained cells found dispersed in the medulla of control ratot*
(arrowheads in a) are absent^^reduced from that of the hypophysectomized rat
(b). Co, cortex; M, medulla. Bar: 75 fjm.
153
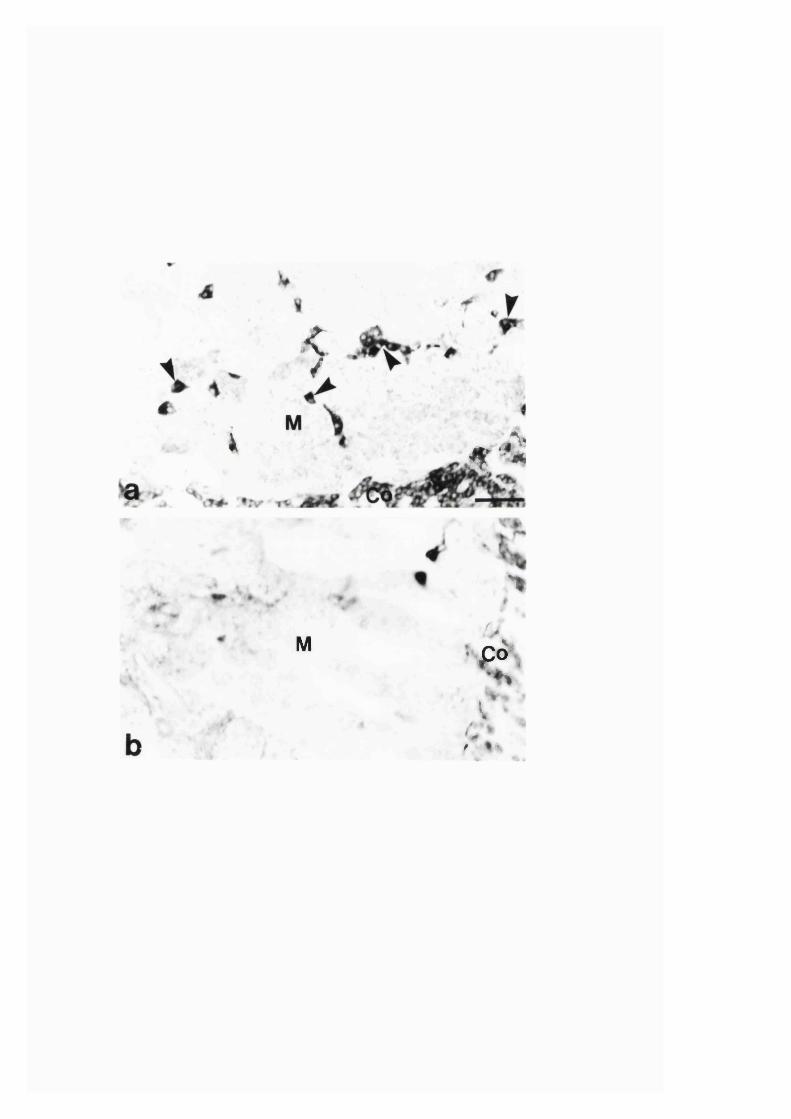
i
^ *
5 - ' i
>
M />
• Po% ♦>c
f " »>
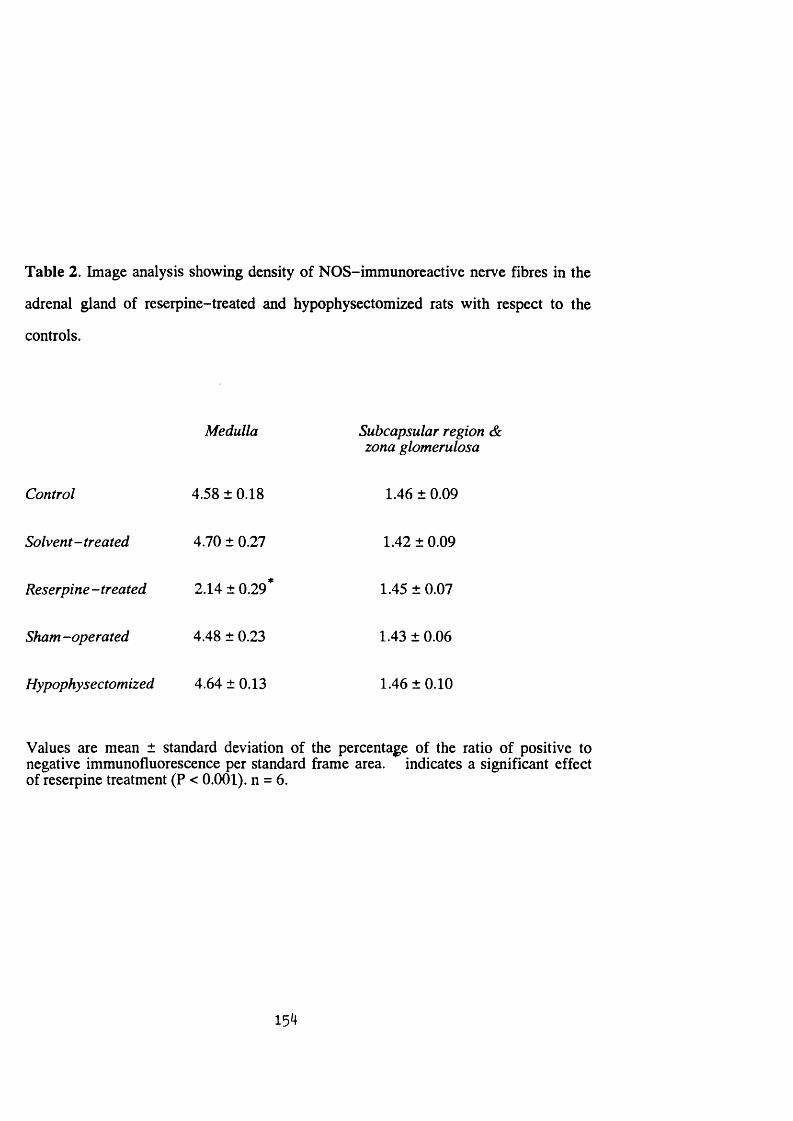
Table 2. Image analysis showing density of NOS-immunoreactive nerve fibres in the
adrenal gland of reserpine-treated and hypophysectomized rats with respect to the
controls.
Medulla Subcapsular region &zona glomerulosa
Control 4.58 ± 0.18 1.46 ± 0.09
Solvent-treated 4.70 ± 0.27 1.42 ± 0.09
Reserpine - treated 2.14 ± 0.29* 1.45 ± 0.07
Sham -operated 4.48 ± 0.23 1.43 ± 0.06
Hypophysectomized 4.64 ± 0.13 1.46 ± 0.10
Values are mean ± standard deviation of the percentage of the ratio of positive to negative immunofluorescence per standard frame area, indicates a significant effect of reserpine treatment (P < 0.001). n = 6.
154

Discussion
Neuronal and endothelial NOS isoforms are thought to be only expressed
constitutively and to be regulated by calcium and calmodulin. However, recently a
number of conditions have been found to cause changes in the expression and/or
activity of these isoforms (Verge et al. 1992; Wu 1993; Bredt and Snyder 1994; Sessa
1994; also Section 3:1:2). In the present study, the expression of NOS in the adrenal
gland of rats histochemically detected by an antibody against the neuronal isoform also
showed a significant change after reserpine treatment. The reduction in the NOS
immunoreactivity was found in the nerve fibres that innervate adrenal medullary cells.
As found from studies reported in Section 3:1:3, these nerve fibres are extrinsic and
that they appear to be preganglionic cholinergic nerve terminals (Blottner and
Baumgarten 1992). The mechanism(s) involved in the changes caused by reserpine in
these nerves is not clear at present and is a matter of speculation. Reserpine is known
to cause depletion of catecholamines and coexisting transmitters, such as NPY, from
adrenergic nerves (Thoenen et al. 1969; Lundberg, Saria, Franco-Cerceda and
Theodorsson-Northeim 1985). However, since reserpine has not been claimed to affect
the preganglionic cholinergic nerves directly, the change we observed in the present
study may have not occurred as a direct consequence of reserpine treatment. Instead,
indirect effects of the drug may be the possible cause of the change. Such an indirect
effect may be related to a retrograde feedback mechanism from the gland associated
with the increased adrenal catecholamine biosynthesis and secretion that are caused by
reserpine treatment (Thoenen et al. 1969).
NO appears to be a non-cholinergic transmitter and/or modulator in adrenal
medullary catecholamine secretory activity. However, since NO has been implicated in
both inhibition (Torres, Ceballos and Rubio 1994) and potentiation (Oset-Gasque,
Parramon, Hortelano, Bosca and Gonzalez 1994) of catecholamine secretion from
adrenal chromaffin cells, it also appears that its involvement is complex. In view of
155

this, and the enhanced biosynthesis and secretion of catecholamines from the adrenal
chromaffin cells occurring after reserpine treatment, the reduction in the level of NOS
observed in this study may be related to suppression of the production of NO from the
preganglionic nerve fibres which consequently reduces any possible inhibitory effect
that NO may otherwise have in the biosynthesis and secretion of catecholamines in the
gland. Nevertheless, although further functional study is needed to clarify such
speculation, the reduction of the levels of NOS in the cholinergic preganglionic nerve
fibres at the time when their activity is supposedly increased (Thoenen et al. 1969)
may indicate that the production of NO is regulated differently from that of
acetylcholine in these nerves.
Although the adrenal medullary secretion is affected by adrenal glucocorticoids,
and hypophysectomy has been shown to affect adrenal medullary catecholamine and
peptide biosynthesis (Wurtman and Axelrod 1966; Yobum et at. 1987),
hypophysectomy did not cause any change in the NOS immunoreactivity in the gland
in the present study. However, the disappearance of the NADPH-diaphorase stained
cells, which are found dispersed in the medulla of control rats, further supports the
suggestion that these cells are cortical cells. It is a matter for further investigation if
such cortical cells located in the medulla could be, at least partly, the source for local
glucocorticoids needed in the regulation of adrenal medullary secretory activity in the
gland.
156

SECTION 3:2:2
INCREASE IN NITRIC OXIDE SYNTHASE AND NADPH-DIAPHORASE IN
THE ADRENAL GLAND OF STREPTOZOTOCIN-DIABETIC WISTAR RATS
AND ITS PREVENTION BY GANGLIOSIDE
157

A bstract
Levels of nitric oxide synthase and NADPH-diaphorase in adrenal glands of
streptozotocin-diabetic rats of 8 and 12 weeks duration compared with control rats
were assessed with histochemical and biochemical techniques. Adrenal glands from
streptozotocin-diabetic rats of 8 weeks duration treated with ganglioside were also
examined. In the adrenal medulla of 8 weeks and 12 weeks diabetic rats nitric oxide
synthase-immunoreactive nerve fibres were increased and decreased, respectively;
additional nitric oxide synthase-immunoreactive and NADPH-diaphorase stained
cells, which appeared to be cortical cells, were located in medulla and cortex as
compared with controls. Increased intensity in NADPH-diaphorase staining of the
cortical cells of diabetic rats was also observed. Ganglioside treatment of the 8 weeks
diabetic rats prevented the diabetic-induced increase in nitric oxide synthase-
immunoreactive nerve fibres. It also reduced most of the diabetic-induced increase in
the nitric oxide synthase-immunoreactive and NADPH-diaphorase stained cells and
the intensity of NADPH-diaphorase staining of cortical cells. With biochemical assay
a significant increase in nitric oxide synthase activity was found in the adrenal glands
from 8 weeks diabetic rats, and this increase was reduced by ganglioside treatment in
4 out of 6 diabetic rats.
In summary, streptozotocin-induced diabetes causes an initial increase in the
levels of nitric oxide synthase and NADPH-diaphorase in adrenal gland of rat, which
was prevented by ganglioside treatment.
158

Introduction
Chromaffin cells of adrenal medulla synthesize and secrete adrenaline which is
involved in glucose counteyegulation next to glucagon (Cryer 1981). In long standing
diabetes in humans, the glucagon response to insulin-induced hypoglycemia is reduced
(Gerich, Langlois, Noacco, Karam and Forsham 1973) and is absent in diabetes with
autonomic neuropathy (Maher, Tanenberg, Greenberg, Hoffman, Doe and Goetz
1977). Reduction in glucagon response to insulin-induced hypoglycemia has also been
observed in alloxan-diabetic (Lykkelund and Lund 1979), streptozotocin-diabetic
(Patel 1983) and B B -W istar diabetic (Wilke et al. 1993) rats. In the absence of
glucagon, maintenance of glucose homeostasis depends on adrenaline secreted from
adrenal medulla (Cryer 1981; Bolli, De Feo, Campagnucci, Cartechini, Angeletti,
Santeucanio and Brunett 1982), the other glucose counte^egulatory hormones: cortisol
and growth hormone not being critical. However, an eventual reduction of adrenaline
release in response to insulin-induced hypoglycemia has also been found in some
patients with IDDM (Hoeldtkeer at. 1982) as well as in streptozotocin-diabetic (Patel
1983) and B B -W istar diabetic (Wilke et al. 1993) rats. The cause for such
complications in diabetes is not known conclusively, although a correlation with
autonomic neuropathy has been found in studies of clinical (Hoeldtk et al. 1982; Cryer
1989) and experimental diabetes in rat (Wilke et al. 1993). However, other clinical
studies did not report such a correlation (Ryder, Owens Hayes, Ghatei and Bloom
1990).
In this study, the distribution of NOS and NA DPH-diaphorase in the adrenal
glands of streptozotocin-diabetic rats was investigated. Ganglioside is a sialic ac id -
containing glycolipid that is found in the plasma membrane of mammalian cells, being
especially abundant in neurons and associated glial cells (Ledeen and Yu 1982; Ando
1983). It has been claimed to increase neural survival in a wide variety of
neuropathological conditions (Cuello 1990; Rodden, W iegant and Bauer 1991). In this
159

laboratory, ganglioside treatment has been shown to prevent diabetes-induced changes
in the innervation of the pyloric sphincter of rat (Soediono, Belai and Bumstock 1993).
Thus, in this study, if ganglioside can affect changes in the level of NOS and NADPH-
diaphorase in adrenal glands of streptozotocin-diabetic rats was also investigated.
160

Materials and methods
Male Wistar rats with initial body weight of 400-450 g were used. Diabetes was
induced in 27 rats by a single intraperitoneal injection of streptozotocin (65 mg/kg
body wt; Upjohn, Kalamazoo, USA) in saline. Onset of diabetes was established by
rapid body weight loss, polyuria and glycosuria (Diastix; Ames, Slough, UK).
Starting one week after induction of diabetes, 11 diabetic rats were given
intraperitoneal injections of mixtures of bovine brain ganglioside (AGF^, 30 mg/kg
body wt; Fidia, Abano Terme, Italy) twice a week for 7 weeks. AGF^ is a mixture of
ganglioside whose carboxyl group of the sialic acids of GMj ,̂ GD^^, GD^y and GT^y
are esterified with the hydroxyl groups of the neighbouring saccharides.
Two groups of age-matched controls, which consisted of 16 untreated non
diabetic and 11 ganglioside-treated non-diabetic rats similar to ganglioside-treated
diabetic rats were used. Most of the rats were killed 8 weeks after injection with
streptozotocin, while 5 untreated diabetic and 5 untreated non-diabetic rats were killed
12 weeks after induction of diabetes. Animals were killed with an overdose inhalation
of ether and blood samples (3-5 ml) were quickly withdrawn from the posterior vena
cava into heparinized syringes for plasma-glucose level determination by the
hexokinase method, using Sigma diagnostics kit (Sigma, Poole, UK). The adrenal
glands were then dissected out.
Histochemistry
Adrenal glands were immersion fixed in 4 % paraformaldehyde in 0.1 M PBS
and cryostat sections were prepared as described in Section 2:3:1. Every other
consecutive sections were processed for NOS immunohistochemistry as described in
Section 2:3:2. The NOS antibody used was raised in rabbit against purified soluble
NOS extracted from rat cerebellum (Schmidt et al. 1991), and was at a dilution of
1:2500.
161

After studying the immunoprocessed sections under fluorescence microscope,
coverslips were removed and sections were rinsed in PBS. NADPH-diaphorase
histochemistry, as described in Section 2:3:3, was carried out on adrenal sections
which were previously immunoreacted with NOS antiserum and other consecutive
sections.
Biochemical Assay o f NOS Activity
More than one type of NADPH-diaphorase exists in the adrenal gland. In
addition, NADPH-diaphorase activity measured biochemically does not correlate with
NOS activity (Tracey, Nakane, Pollock and Forstermann 1993). Therefore, the
biochemical study was restricted to the measurement of NOS activity.
Adrenal glands from diabetic and ganglioside-treated diabetic rats of 8 weeks
duration and that of ganglioside-untreated and treated non-diabetic controls were
assayed for NOS enzyme activity as described in Section 2:5. Statistical analysis of the
biochemical data was carried out by two way analysis of variance with diabetes and
ganglioside treatment as factors by the general linear model for unbalanced data using
the Minitab 10 programme. Such statistical analysis also revealed any outlying values
within each group that were significantly outside normal distribution. In such cases the
data was calculated and analyzed both with and without the outlying values as
suggested by Stevens (1990).
162

RESULTS
Upon death both diabetic and ganglioside-treated diabetic rats had lower body
weight, increased adrenal gland weight and higher blood glucose level as compared
with ganglioside-untreated and treated non-diabetic control rats (Table 3).
NOS immunoreactivity in medulla o f control rats
Control experiments in which the NOS antibody was omitted did not give any
staining. A comparable distribution of NOS immunoreactivity was observed in the
adrenal medulla from the untreated non-diabetic (Fig. 18a) and ganglioside-treated
non-diabetic control rats, and was as reported previously (Sections 3:1:1-2). Briefly,
NOS immunoreactivity was localized in several intrinsic neuronal cell bodies, and
nerve fibres which were associated with chromaffin cells, ganglion cells and blood
vessels.
NOS immunoreactivity in medulla o f 8 weeks diabetic rats
No apparent change was observed in the NOS immunoreactivity of the intrinsic
neuronal cell bodies. However, increased NOS-immunoreactive nerve fibres were seen
in the adrenal medulla from the diabetic rats of 8 weeks duration (Fig. 18b). The
increased nerve fibres were found accumulated in some regions of medulla near
chromaffin cells and large non-immunoreactive ganglion cells. Furthermore, several
additional cells occasionally found as columns of cells in between groups of
chromaffin cells and non-immunoreactive ganglion cells displayed homogeneous NOS
immunofluorescence (Fig. 18b). Most of these cells were small in diameter (6-10 ytrni)
and had a small central nucleus.
NOS immunoreactivity in medulla o f ganglioside-treated 8 weeks diabetic rats
In the ganglioside-treated diabetic rat adrenal medulla the diabetic-induced
163

increase in NOS-immunoreactive nerve fibres was not observed, and the pattern of
innervation was similar to that of the non-diabetic controls (Fig. 18c). In addition, the
majority of NOS-positive cells that appeared in the medulla of diabetic rats were not
observed with ganglioside treatment. However, several cells found dispersed in the
outer region of medulla persistently gave NOS immunoreactivity.
NOS immunoreactivity in cortex o f control rats
In the adrenal cortex of ganglioside-untreated and treated non-diabetic control
rats, there were NOS-immunoreactive nerve fibres in the zona glomerulosa and
subcapsular region (Fig. 19a), and around some blood vessels.
NOS immunoreactivity in cortex o f 8 weeks diabetic and ganglioside-treated diabetic
rats
Immunoreactive nerve fibres similar to that of controls were present in the
adrenal cortex of both diabetic and ganglioside-treated diabetic rats of 8 weeks
duration. However, a small number of cortical cells located in some regions of the
zonae glomerulosa and fasciculata of the diabetic rats gave positive NOS
immunoreactivity (Fig. 19b). Such immunoreactive cortical cells were not found in the
glands from ganglioside-treated diabetic rats (Fig. 19c).
NOS immunoreactivity in medulla and cortex o f 12 weeks diabetic rats
In the diabetic rats which were left for 12 weeks after induction of diabetes, the
occurrence of NOS-immunoreactive nerve fibres in medulla was reduced as compared
with age-matched non-diabetic controls (Figs. 20a, b). In addition, a weaker
immunofluorescence was seen in the chromaffin cells of some regions of the medulla.
However, similar to the 8 weeks diabetic rat gland, NOS immunoreactivity in
occasional small sized cells was found in the medulla (Fig. 20b). Furthermore, in the
164

12 weeks diabetic rats, the NOS-immunoreactive nerve fibres found in the outer cortex
were reduced as compared with controls, while the occurrence of NOS-
immunoreactive cortical cells in the region was increased (Figs. 21a, b).
NADPH-diaphorase staining in control rats
No NADPH-diaphorase staining was seen in the control sections incubated in
media devoid of ^-NADPH or in those sections pretreated with TRIS HCl heated at
75°C. A similar pattern and intensity of NADPH-diaphorase staining was seen in the
adrenal glands from ganglioside-untreated and treated non-diabetic control rats.
NOS-immunoreactive cellular elements were double positive for NADPH-diaphorase.
Cortical cells forming the parenchyma of adrenal cortex and those found as occasional
dispersed cells in medulla were also positively stained for NADPH-diaphorase (Fig.
22a). The intensity of NADPH-diaphorase staining in the cortical cells varied between
regions of the gland, being weaker in those found in zona glomerulosa than those
found in the zonae fasciculata and reticularis. Small groups of cells located at the inner
aspect of zona reticularis and at the corticomedullary junction gave a heavier NADPH-
diaphorase staining than the remaining regions. The staining intensity of these cells
matched those of cortical cells occasionally found dispersed in medulla.
NADPH-diaphorase staining in diabetic rats
NOS-immunoreactive cellular elements in the glands from all groups of the
diabetic rats were also NADPH-diaphorase double positive. However, cortical cells of
adrenal glands from diabetic rats of both duration gave a heavier intensity of NADPH-
diaphorase staining compared with that of non-diabetic control rats (Figs. 22a, b). The
increase in staining was more significant in cells of zonae fasciculata and reticularis,
and those cells found dispersed in medulla. In glands of ganglioside-treated 8 weeks
diabetic rats, the intensity of the NADPH-diaphorase staining of the cortical cells (Fig.
22c) was lower than that of diabetic rats, but was higher than that found in the non
165

diabetic controls.
Biochemical assay o f NOS activity
In the assay for NOS activity in the soluble fraction from adrenal glands of
untreated non-diabetic control rats, the production of citrulline was totally inhibited
(99.6 ± 0.3 %, mean ± standard error of mean, n = 6) by the arginine analogue, (N ^ -
nitro-L-arginine methyl ester (L-NAME). L-NAME inhibits both constitutive and
inducible NOS activity. The percentage inhibition by EGTA, a calcium chelator, was
89.7 ± 3.1 % (n = 6), indicating that virtually all the NOS activity measured was
calcium-dependent and thus of the constitutive isoform of the enzyme. Neither
diabetes nor ganglioside treatment had any significant effect on the degree of inhibition
by L-NAME or EGTA. The levels of NOS activity in the adrenal glands from the four
groups of animals are given in Table 4.
Total NOS activity/gland was significantly increased (p < 0.001) in the adrenal
glands from diabetic rats. Ganglioside treatment did not have any significant effect on
NOS activity in control rats. However, it was apparent that ganglioside treatment did
result in increased variability in the levels of NOS activity measured in diabetic rats.
Two outlying values of NOS activity, were revealed in the ganglioside-treated diabetic
group during two way analysis of variance. When these values were removed
ganglioside treatment did have a significant effect on NOS activity in diabetes (p =
0.025). Thus there was a significant interaction (P < 0.05) between the effect of
diabetes and the effect of ganglioside treatment. A similar pattern of change was
observed when results were expressed as activity inhibited by L-NAME and EGTA
(Table 4) indicating that the increase in NOS activity in diabetes was due to an increase
in constitutive NOS activity. The increase in NOS activity/gland in diabetes was not
simply due to the increased weight of adrenal glands induced by diabetes. When NOS
activity was expressed per gram tissue there was still a significant increase in diabetic
166

tissue (Total NOS activity, nmoi/h/g tissue; Controls = 73.14 ± 4.42, n = 6; Diabetics =
141.16 ± 8.51, n = 5; P < 0.001). Furthermore, the decrease in NOS activity/gland in 4
out of the 6 ganglioside-treated diabetics was observed despite the fact that
ganglioside treatment did not significantly affect the diabetic-induced increase in
adrenal gland weight (Table 3).
167

Fig. 18a-c. Photomicrographs of NOS immunoreactivity in sections of adrenal
medulla of (a) non-diabetic control, (b) 8 weeks diabetic and (c) ganglioside-
treated 8 weeks diabetic rats. Arrowheads in a indicate NOS-immunoreactive
neurons, while small arrows show NOS-immunoreactive nerve fibres in this
and subsequent figures. Inset in b shows a group of small size immunoreactive
cells (large arrow) found in between groups of chromaffin cells. Note the
increased immunoreactive nerve fibres that occurred in the medulla of the
diabetic rat (b). Co: cortex. Bar = 33 fnn.
168
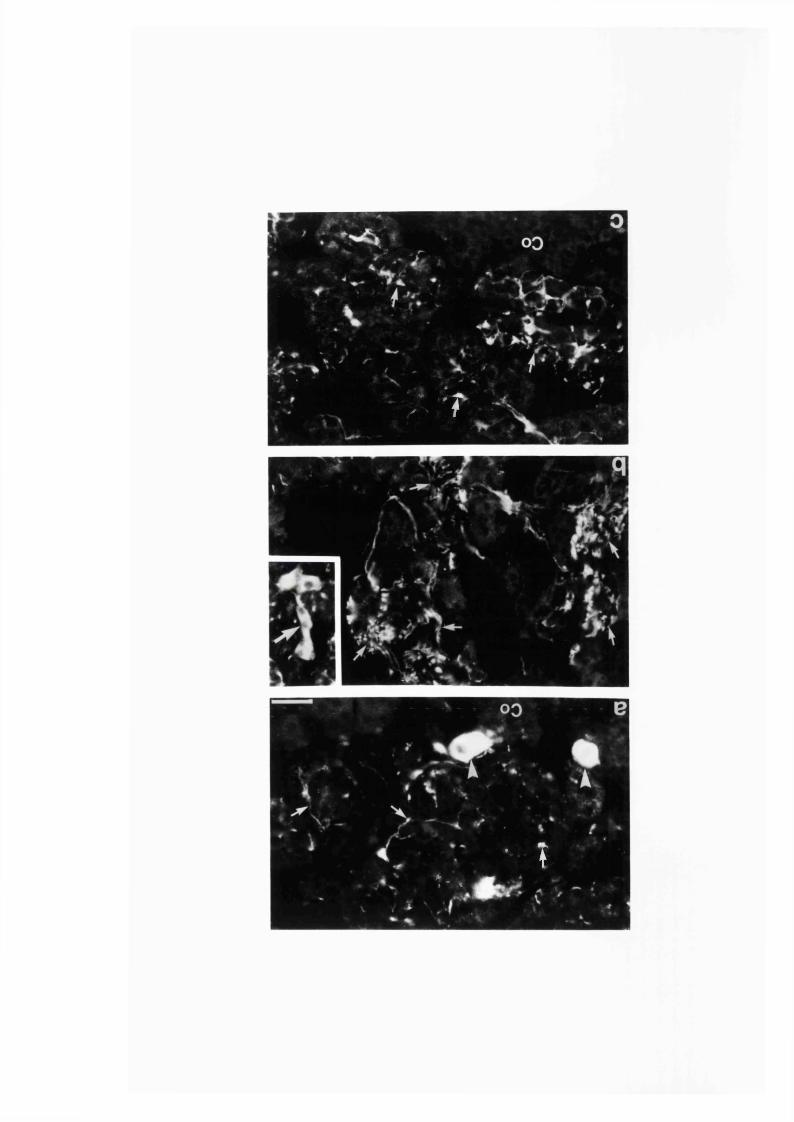
; ’ Î
• Ît - .

Fig. 19a-c. Photomicrograph of NOS immunoreactivity in sections of adrenal
cortex of (a) non-diabetic control, (b) 8 weeks diabetic and (c) ganglioside-
treated 8 weeks diabetic rats. Note the presence of immunoreactive cells {arrows
in b) in a region of cortex of the adrenal gland from diabetic rat. ZG: zona
glomerulosa; ZF: zona fasciculata. Bar = 33 ̂ m.
169
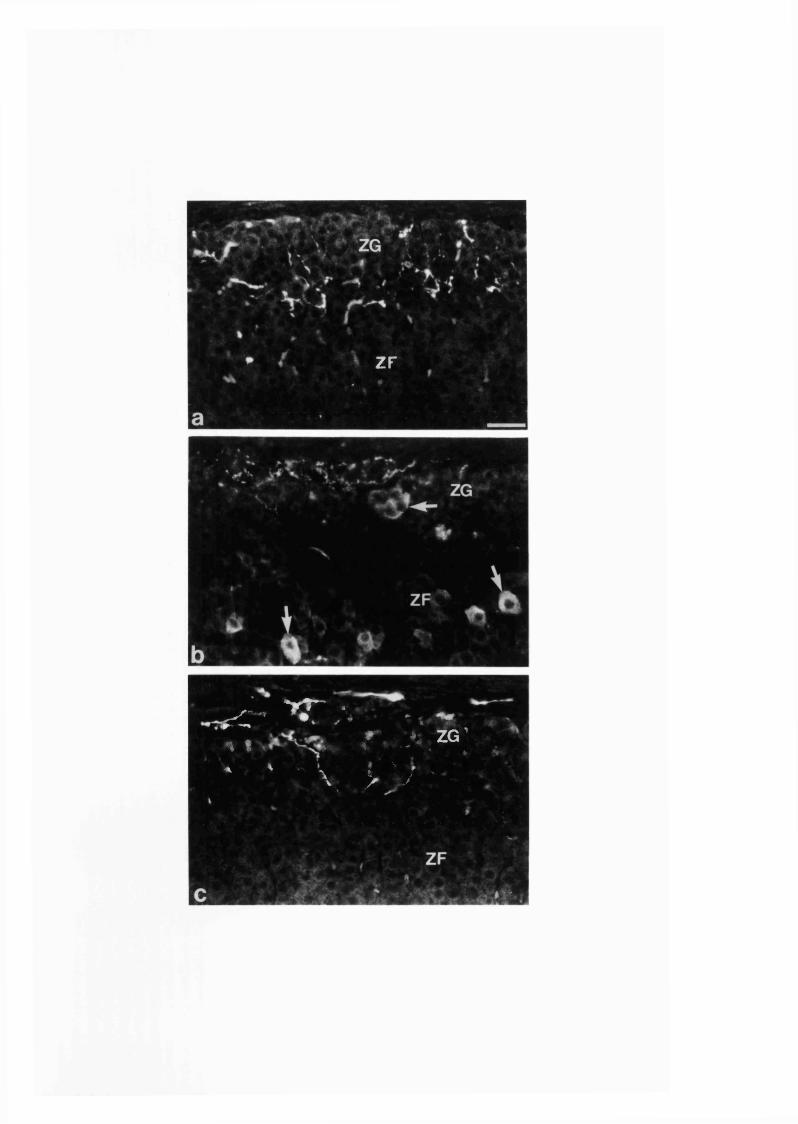
' ZF
-■ * * r t ^ .rC ' • . * • “̂ ,. , T / ' ZG
' ' \ > \ ; -

Fig. 20a, b Photomicrographs of NOS immunoreactivity in sections of adrenal
medulla from (a) non-diabetic control rat and (b) diabetic rat of 12 weeks
duration. Inset in b shows a group of small size NOS-immunoreactive cells
{large arrow) found in between groups of chromaffin cells. Note the absence of
the large majority of immunoreactive nerve fibres in the gland from diabetic rat
(b) as compared with that of the non-diabetic controls (a). Small arrows in a
show immunoreactive nerve fibres. Arrowhead in b points to an
immunoreactive neuron. Bar = 33 ^m.
170


Fig. 21a, b Photomicrographs of NOS immunoreactivity in sections of adrenal
cortex from (a) non-diabetic control and (b) 12 weeks diabetic rats. Note in b
the presence of several immunoreactive cells {arrows) and the absence of
immunoreactive nerve fibres. ZG: zona glomerulosa, ZF: zona fasciculata. Bar =
33y«m.
171
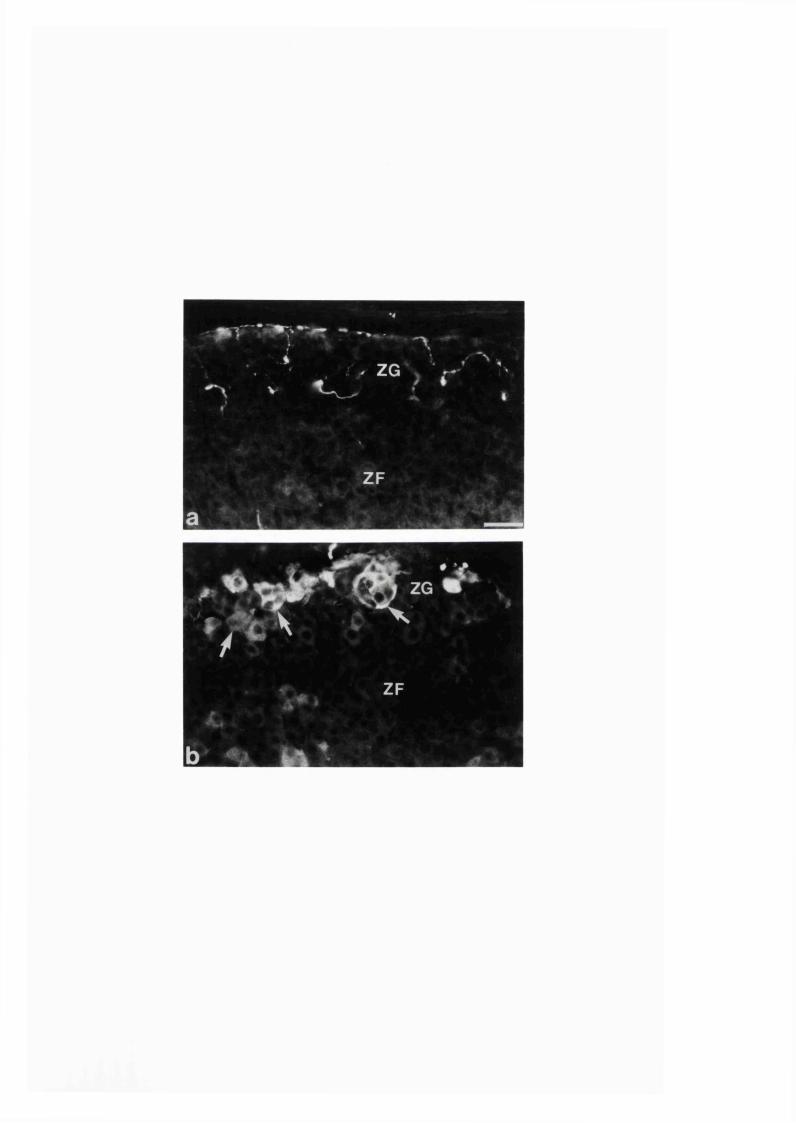

Fig. 22a-c. Photomicrographs of sections of adrenal cortex stained black for
NADPH-diaphorase of (a) non-diabetic control, (b) 8 weeks diabetic and (c)
ganglioside-treated 8 weeks diabetic rats. Note the high intensity of NADPH-
diaphorase staining in adrenal cortex of the diabetic rat in b which is reduced by
ganglioside treatment (c), although not to the level of that of the non-diabetic
control rat (a). Black arrowheads in a show groups of intensely stained cells
occasionally found in some regions of zona reticularis and at corticomedullary
junction. ZG; zona glomerulosa; ZF: zona fasciculata; ZR: zona reticularis. Bar
= 64 /Mm.
172

m

Table 3. Comparison of body weight, adrenal gland weight and blood-glucose level of
diabetic and ganglioside-treated diabetic rats of 8 weeks duration with respect to the
non-diabetic controls. Values represent mean ± standard error of mean. ̂ indicates p <
0.001, and ^ indicates p < 0.05 with respect to both ganglioside-untreated and treated
non-diabetic controls.
Controls Ganglioside'treated
Diabetics Ganglioside- treated
Bodywt(g)(n = 11)
609±14.48 594±9.91 382±16.16^ 399±9.35^
Adrenal wt (mg) (n = 6)
25±0.89 23.8±0.92 32.6±2.73^ 29.811.18'’
B lood- glucose (mM) (n = 11)
10±0.55 11.5±0.47 44.5±2.27^ 45±1.60^
173

Table 4. Effects of diabetes and ganglioside treatment on NOS activity (nmol/h/gland)
in the adrenal gland of diabetic rats of 8 weeks duration.
Total L -N A M E - EGTA-sensitive sensitive
Controls (n = 6) 1.95±0.20 1.94±0.19 1.73±0.14
Ganglioside-treated 1.80±0.33 1.80±0.33 1.64±0.33controls (n = 6)
Diabetics (n = 5) 4.61±0.52* 4.59+0.51* 4.05+0.40*
Ganglioside-treateddiabeticsa) Whole group (n = 6) 3.94±0.81 3.92±0.81 3.45±0.69
b) Outliers removed 2.74±0.47'*^ 2.74±0.47‘*'̂ 2.49±0.45'*‘̂(n = 4)
NOS activity was measured by the conversion of arginine to citrulline. Total activity represents activity corrected for a reagent blank. L-NAME-sensitive and EGTA- sensitive activity represent the difference in NOS activity measured in the presence and absence of L-NAME and EGTA, respectively. Analysis of the data was carried out using two way analysis of variance. Where such analysis revealed the presence of values in individual groups significantly outside the normal limits of distribution the data has been calculated both with and without the outliers. Results have been given as the mean ± standard error of mean. * indicates a significant effect of diabetes (p < 0.001), + indicates a significant effect of ganglioside treatment (p < 0.025), # indicates a significant interaction between the effects of diabetes and of ganglioside treatment (p < 0.05).
174
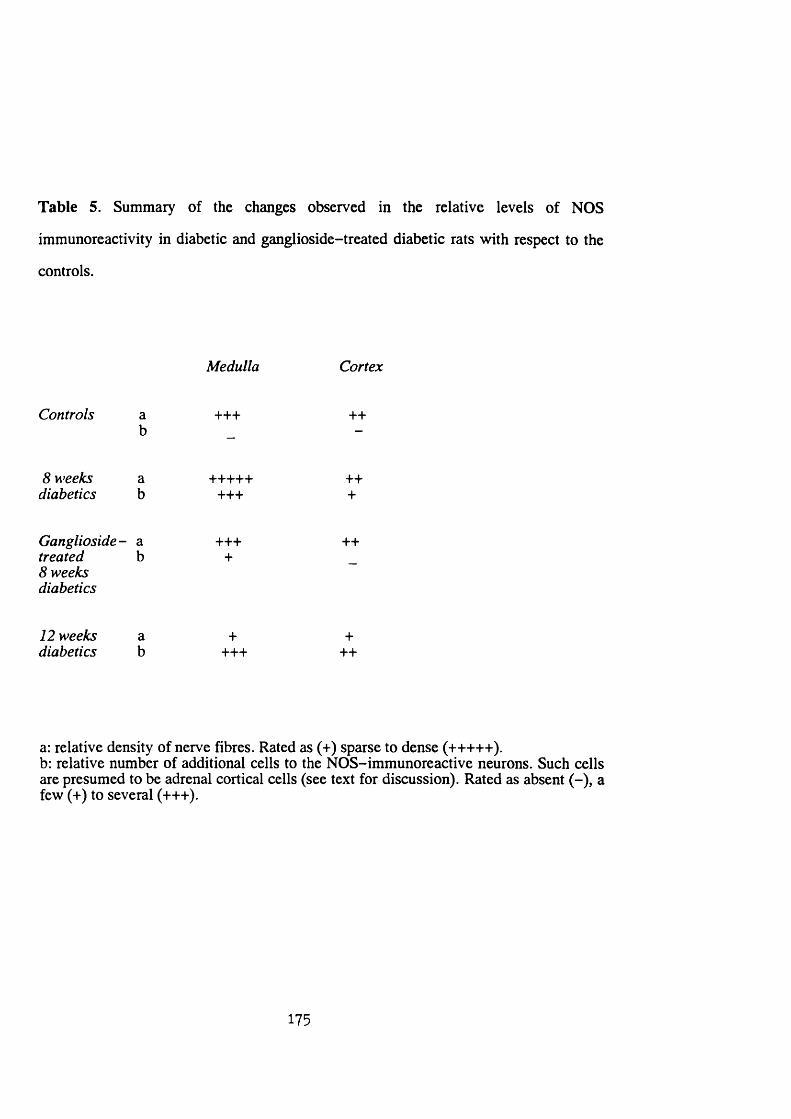
Table 5. Summary of the changes observed in the relative levels of NOS
immunoreactivity in diabetic and ganglioside-treated diabetic rats with respect to the
controls.
Medulla Cortex
Controls +++ ++
8 weeks diabetics
++++++++
+++
Ganglioside- a treated b8 weeks diabetics
++++
++
12 weeks diabetics
++++
+++
a: relative density of nerve fibres. Rated as (+) sparse to dense (+++++). b: relative number of additional cells to the NOS-immunoreactive neurons. Such cells are presumed to be adrenal cortical cells (see text for discussion). Rated as absent (-), a few (+) to several (+++).
175

Discussion
Effect o f diabetes
The increase in the weight of the adrenal glands of diabetic rats observed in the
present study is in agreement with earlier reports that the adrenal gland of diabetic rat
is enlarged (Bennett and Koneff 1946; Rebuffat et al. 1988b, c). As has been
summarized in Table 5, the changes on the NOS immunoreactivity caused by diabetes
and the effect of ganglioside treatment on such diabetic-induced changes are marked.
In the adrenal glands from the diabetic rats of 8 weeks duration, the occurrence of NOS
immunoreactivity in additional cells and nerve fibres when compared with the glands
from the non-diabetic control rats was observed immunohistochemically. Biochemical
assay also demonstrated a significant increase in the NOS activity in the adrenal glands
from the 8 weeks diabetic rats with respect to the non-diabetic controls, confirming the
immunohistochemical results.
The NOS activity measured biochemically by the conversion of arginine to
citrulline was calcium-dependent, indicating that it was of the constitutive isoform of
the enzyme. Both endothelial and neuronal NOS are of the constitutive isoform;
however, more than 90 % of the endothelial NOS is membrane bound whereas
neuronal NOS is mainly found free in the cytoplasm (Forstermann et at. 1994). Since
no detergent was added to the homogenizing medium and only the supernatant
(cytoplasmic fraction) was analyzed for NOS activity, it is unlikely that endothelial
NOS contributed to the results reported here. The conclusion that NOS activity in the
cytoplasmic fraction reflects neuronal rather than endothelial NOS activity has been
reported previously (Bush, Gonzalez and Ignarro 1992). Since the increased NOS
activity in the adrenal glands from the diabetic rat was also calcium-dependent the
increase was due to constitutive NOS, as found immunohistochemically using antibody
against the constitutive NOS type I isoform (Schmidt et at. 1991), rather than induction
of the inducible isoform of the enzyme.
176

As shown histochcmically, in the diabetic rat adrenals there were NOS-
immunoreactive and NADPH-diaphorase stained small sized cells found in between
the chromaffin cells and ganglion cells of the medulla. As found in Sections 3 :l:l-2 ,
there are some cells found scattered in the adrenal medulla in between groups of
chromaffin cells as well as ganglion cells that display NADPH-diaphorase staining,
but not NOS immunoreaction. These cells are negative for the chromaffin cell marker
TH (Section 3:1:4) and the general neuronal marker PGP 9.5 (unpublished
observation), and are thought to be cortical cells that are found dispersed as islands in
the medulla of rat (Coupland 1965). The NOS-immunoreactive cells observed in
diabetic rats may be of these cortical cell type where diabetes has increased their
number and induced the expression of NOS. This is further indicated by the finding of
similar NOS-immunoreactive cells in the adrenal cortex of the diabetic rats. However,
it is not known as what could be the cause and effect of expression of NOS in such
cortical cells during diabetes. It has been shown that in streptozotocin-induced diabetic
rats there is atrophy and hypertrophy of zona glomerulosa and fasciculata cells,
respectively (Rebuffat et al. 1988b, c). It may be either of such affected cortical cells
that expressed NOS in diabetes.
This study has also demonstrated an increase in the density of NOS-
immunoreactive nerve fibres found in the adrenal medulla of diabetic rats of 8 weeks
duration and the dramatic disappearance of such nerve fibres with increasing duration
of diabetes at 12 weeks. Previous studies have investigated changes in neurotransmitter
content of autonomic nerves in diabetes (Lincoln, Bokor, Crowe, Griffith, Haven and
Bumstock 1984; Belai, Lincoln, Milner, Crowe, Loesch and Bumstock 1985; Belai,
Lincoln, Milner and Bumstock 1988; Bumstock 1990; Soediono et al. 1993). In a
similar pattem to the present observation on NOS-immunoreactive nerve fibres in
adrenals of the diabetic rats, in rat proximal colon an initial increase in tissue level and
immunoreactivity of VIP and noradrenaline at 8 weeks has been found to be followed
177

by a subsequent decrease at 16 and 25 weeks after induction of diabetes with
streptozotocin (Belai et al. 1988). In addition, an early increase of VIP and PGP 9.5
immunoreactivity in the skin of human IDDM patient has been found to precede a
subsequent depletion (Properzi, Francavilla, Poccia, Aloisi, Gu, Terenghi and Polak
1993). Increased activities of NOS and NADPH-diaphorase have also been found in
myenteric nerves of 8 weeks diabetic rat ileum (Lincoln, Messersmith, Belai and
Bumstock 1993). NO has been implicated in neural death in cerebral infarction
(Nowicki, Duval, Poignet and Scatton 1991) and in activated microglia in culture (Boje
and Arora 1992). It is possible that the increase in the NOS immunoreactivity in the
nerve fibres and the appearance of NOS-containing cellular elements in the glands of 8
weeks diabetic rats may be indicative of enhanced production of NO as an event
leading to neuropathy and cellular degeneration. In this connection, reduction in the
amount of preganglionic nerve terminals innervating the chromaffin cells in addition to
changes in adrenal cellular structure has been found in the diabetic rats of long duration
(Wilke et al. 1993). It has also been reported that there is adrenal medullary fibrosis in
human IDDM patients of long duration (Brown et al. 1989,1990).
Although NOS is generally colocalized with NADPH-diaphorase, the reverse is
not always true, since the latter can exist independently of the former in some tissues.
In adrenal cortex, where NADPH is utilized by several steroidogenic enzymes (Sweat
and Lipscomb 1955), NADPH-diaphorase labelling of the cells with a characteristic
zonal intensity of staining distribution occurs. Comparison of such staining of adrenal
glands from experimental animals with that of controls has been utilized in the past to
assess the state of adrenal cortical activities. Accordingly, the increase in NADPH-
diaphorase staining that we observed in the adrenal cortex of the diabetic rats over that
of the non-diabetic controls may indicate increased enzymatic adrenocortical activities
with increased corticosteroid production. In this respect, in streptozotocin-treated rats
a rise in the intra-adrenal (Tomello et al. 1981) and plasma (De Nicola et al. 1976;
Rebuffat et al. 1988c) corticosterone level have been reported. As indicated by the
178

increased plasma level of ACTH, increased activity of the hypothalamic-pituitary-
adrenal axis has been implicated in the increase in corticosterone level during diabetes
(Rebuffat et al. 1988b, c). In clinical studies the cause for such an increase in
hypothalamic-pituitary-adrenal axis has been suggested to be diabetic neuropathy
(Tsigos et at. 1993). It is therefore a matter to be investigated if the increased
NADPH-diaphorase staining in the cortical cells of the diabetic rats reflects diabetic
neuropathy somewhere along the hypothalamic-pituitary-adrenal axis.
Effect o f ganglioside treatment
The immunohistochemical results indicate that ganglioside treatment tends to
prevent the diabetic-induced increase in NOS and NADPH-diaphorase reactivity in
the 8 weeks diabetic rat adrenal glands. The biochemical assay results for the effect of
ganglioside treatment on NOS activity are somewhat equivocal due to the fact that
ganglioside treatment per se appeared to result in increased variability in the data.
According to the results from assay of NOS activity with ganglioside treatment, there
was a significant reduction in the diabetes-induced increase in NOS activity in the
adrenal glands from four out of the six diabetic animals studied, although such
reduction did not occur in the remaining two rats. The reason for the inconsistency
between the biochemical and histochemical approaches is unknown at present. The
persistency of some reactive cells in the ganglioside -treated diabetic rats, as observed
histochemically, may in part be accounted for by the observed variability in the NOS
activity.
Ganglioside has been shown to prevent diabetic-induced neuropathic changes
(Soediono et al. 1993) and facilitate recovery from diabetic-induced neural
dysfunction (Norido, Canella, Zanoni and Gorio 1984; Triban, Guidolin, Fabris,
Marini, Schiauinato, Dona, Bortolami, Giamberardino and Fiori 1989; Figliomeni,
Bacci, Panozzo, Fogarolo, Triban and Fiori 1992). It is therefore possible that
179

ganglioside treatment, by preventing an increase in NO production in the adrenal glands
from diabetic rats may protect against neuropathy. Furthermore, it remains to be
established if the observed inhibition of the increase in adrenal cortical NADPH-
diaphorase staining intensity in the diabetic rats by ganglioside is secondary to possible
protection from neuropathy. However, the possibility that ganglioside may have a
direct parallel influence on the cortical cells similar to that on neural tissue can not be
ruled out and awaits investigation.
180

SECTION 4
GENERAL DISCUSSION
181

Each of the separate studies presented in this thesis has been discussed
individually in detail in Section 3. In this Section of the thesis a general discussion with
regard to the feature of adrenal NOS-synthesizing nerves and their possible functional
involvement in the adrenal gland activity is presented. Plasticity in the expression of
the nitrergic nerves and its possible cause and mechanism has also been discussed here.
4:1. Adrenal nitric oxide-synthesizing nerves
The NOS immunoreactivity in the adrenal gland from the adult rats was found
only in the neuronal elements. This is also in agreement with the results obtained by
others (Bredt et al. 1990; Dawson et al. 1991; Aim et al. 1993), except one group (Dun
et al. 1993) who claimed the occurrence of the immunoreactivity in all of the
chromaffin cells as well. The lack of NADPH-diaphorase staining in the chromaffin
cells also further confirmed the absence of NOS from the adrenal chromaffin cells of
the adult rats. However, indicating possible species difference, NOS immunoreactivity
has been reported in a small number of chromaffin cells in human adrenal gland (Heym
et al 1994).
The adrenal nitrergic nerves are heterogeneous, since they costore different
neuromediators (Section 3:1:4) and the nerve fibres originate from extrinsic as well as
intrinsic adrenal neuronal sources (Section 3:1:3). The source for the extrinsic NOS-
containing nerve fibres that innervate adrenal medullary cells has not been totally
demonstrated, although sympathetic preganglionic cell bodies in the intermediolateral
cell column of the spinal cord have been shown to have their terminals in the gland
(Blottner and Baumgarten 1992). However, the nitrergic nerve fibres found as a plexus
in the subcapsular region and the zona glomerulosa of the adrenal cortex, as well as
those associated with some intra-adrenal blood vessels, come from the NOS-
containing intrinsic adrenal neurons (Section 3:1:3).
The nature and function of the adrenal intrinsic neurons have been a mystery for
182

many years. Two principal types of intra-adrenal neurons have been described (Unsicker
et al. 1978). One group of neurons are large multipolar, which are typical of ganglion
cells. These neurons express TH, NA and NPY, and do not have mRNA for choline
acetyltransferase and arc therefore regarded to be sympathetic postganglionic neurons
producing noradrenaline (Dagerlind et al. 1990; Oomori et al. 1994). They are
surrounded by Schwann cells and are contacted by dense nerve terminals. In the guinea
pig, such terminals contain both small clear and large dense cored vesicles, and as
neither of them show uptake of 5-OHDA are considered to be cholinergic (Unsicker et
al. 1978). These type of neurons do not contain NOS, but are heavily innervated with
nerve fibres containing NOS and calretinin (Sections 3:l:4-5). The second group of
intrinsic neurons are smaller in size (Unsicker et al. 1978). From their morphology and
the colocalization studies, it could be inferred that it is this second type of neurons
which demonstrate the NOS immunoreaction and NADPH-diaphorase staining
(Section 3:1:4). Some of these neurons also contain VIP and NPY, and are distinctly
innervated by nerve fibres containing SP, CGRP, VIP and NOS. A few of these
neurons which contain NOS are densely surrounded by calretinin-containing nerve
fibres (Section 3:1:5).
Because of the poor accessibility of the adrenal intrinsic neurons no evidence is
available which shows their functional role. However, based on the frequent
observations of their processes contacting nearby blood vessels (Kondo 1985; Oomori
et al. 1994; also present study) and the present selective surgical denervation studies
(Section 3:1:3), their possible involvement in the innervation of intra-adrenal blood
vessels could be suggested. Furthermore, previous studies which used extrinsic and/or
intrinsic surgical adrenal denervations have shown VIP and NPY-containing adrenal
intrinsic neurons innervate the outer region of adrenal cortex (Hokfelt et al. 1981;
Holzwarth 1984; Maubert et al. 1993). The present study for the NOS-containing
nerves has also supported the suggestion that adrenal intrinsic neurons innervate
183

adrenal cortex (Section 3:1:3).
4:2. Nitric oxide in the regulation of adrenal medullary secretion
The adrenal chromaffin cells continuously secrete catecholamines to maintain
the homeostatic balance of the body. There are also occasions of extreme activation of
these cells with increased secretion of catecholamines during stress conditions. The
chromaffin cells of the adrenal medulla also synthesize several other substances, such
as the neuropeptides. In order to achieve such secretory activities in a harmonious
manner, the adrenal chromaffin cells may be regulated by a variety of
neurotransmitters and hormones. Although acetylcholine released from the
preganglionic nerve terminals is the main regulator of these cells, several studies have
shown evidence for a non-cholinergic transmission as well. Stimulation of adrenal
medulla with subcutaneous injection of nicotine in the rat produced induction of TH
expression which could only be partially blocked with the nicotine receptor
antagonists, hexaméthonium and mecamylamine (Fossom, Carlson and Tank 1991a).
In addition, in the denervated adrenal gland, the induction of TH by nicotine could be
prevented by the above nicotinic receptor antagonists (Fossom, Sterling and Tank
1991b). These observations led to the suggestion that nicotine activates the adrenal
chromaffin cell nicotinic receptors directly as well as activating the central nervous
system pathway resulting in the release of substances from the splanchnic nerves which
interact with adrenal chromaffin cell receptors other than the cholinergic nicotinic
receptors (Fossom et al. 1991a, b). It has also been found that denervation, but not the
nicotinic receptor antagonist chlorisondamine, could completely block the reserpine-
induced increase in adrenal TH levels (Mueller et at. 1970; Schalling et al. 1991). In
addition, the cold-induced increase in the level of PNMT mRNA could not be
inhibited by chlorisondamine or atropine (Baruchin, Vollmer, Miner, Sell, Strieker and
Kapllan 1993), suggesting possible presence of non-cholinergic neuronal transmission
in the regulation of PNMT expression. Studies in isolated perfused rat and bovine
184

adrenal glands have also provided evidence for a significant contribution of non-
cholinergic, secretomotor nerve fibres in the catecholamine secretion elicited by field
stimulation of adrenal nerve fibres (Malhotra and Wakade 1987a; Wakade 1988;
Wakade, Blank, Malhotra, Pourcho and Wakade 1991; Marley, Thomson and
Smardencas 1993).
Several neuromediators that are contained in the splanchnic nerves, intrinsic
neurons and/or the chromaffin cells themselves may be involved in the non-
cholinergic mechanism of regulation of adrenal medullary hormone biosynthesis and
secretion. In rat, VIP has been shown to be involved in non-cholinergic transmission
(Malhotra and Wakade 1987a, b; Wakade 1991; Wakade et al. 1991; Guo and Wakade
1994). Pituitary adenylate cyclase-activating polypeptide which is structurally related
to VIP has also been shown to elicit catecholamine secretion fi’om rat adrenal
chromaffin cells (Wakade, Guo, Strong, Arimura and Haycock 1992).
The occurrence of NO in the preganglionic nerve terminals that innervate
adrenal chromaffin cells (Bredt et al. 1990; Blottner and Baumgarten 1992; also
present study) suggests that NO as a non-cholinergic transmitter may anterogradely
regulate adrenal medullary hormone biosynthesis and secretion. Being an easily
diffusible molecule, NO could also translocate from the preganglionic nerve terminals
to adjacent nerve terminals for a collateral modulation. In addition, the occurrence of
NO in the intrinsic adrenal neurons (Dawson et al. 1991; Aim et al. 1993; also presents
study) suggest that NO may also serve in the gland as a signal arising from
postganglionic neurons for possible modulation of adrenal medullary secretion.
As shown in this study, the nitrergic nerve fibres are found closely associated
with the chromaffin cells, and the NA cells are innervated with a higher proportion
than the A cells (Section 3:1:1). There are other studies that also show that adrenal NA
cells generally receive more nerve endings than the A cells (Allen, ErankO and Hunter
1958; Eranko 1959; Grynszpan-Winograd 1974). Such differential innervation may
185

suggest distinct anatomical pathways controlling A and NA cells. In fact, selective
secretion of adrenaline or noradrenaline can be achieved by stimulating specific
regions in the central nervous system in rat and cat (Folkow and von Euler 1954;
Matsui 1979, 1984). However, the proportion of catecholamines released may also
vary with different conditions such as stress and the state of stimulation. In stress-
induced catecholamine secretion, for example, the proportion of adrenaline and
noradrenaline secreted depends upon the type of the stress experienced (Feuerstcin and
Gutman 1971; Khalil, Livett and Marley 1986). Differential secretion of adrenaline or
noradrenaline can also be achieved by varying the pattem of electrical stimulation of
the splanchnic nerve alone (Mirkin 1961; Edwards 1982).
The involvement of NO in adrenal catecholamine secretory activity has been
suggested by several authors, although their findings are not coherent with regard to
the precise role of NO in causing secretion. Effluents from the endothelial cells and the
chromaffin cells were found to inhibit potassium or the nicotinic agonist
l,l-dimethyl-4-phenylpiperazinium-induced catecholamine secretion from cultured
bovine chromaffin cells (Torres et al, 1994). Since the NOS inhibitors L-NAME or
N^-monoethyl-L-arginine (L-NMMA) and the guanylate cyclase inhibitor
methylene blue could prevent such inhibition of catecholamine secretion caused by the
effluents, but augment the catecholamine secretion induced by the secretagogues, it
was suggested that NO is involved in inhibition of catecholamine secretion via
activation of guanylate cyclase (Torres et al. 1994). In fact, cGMP which is produced
by activated guanylate cyclase and whose cellular level is known to be regulated by
NO (see Garthwaite 1991) has previously been found to inhibit catecholamine
secretion from bovine chromaffin cells (Schneider et al. 1979; Derome et al. 1981) and
PC 12 cells (Fiscus, Robles, Waldman and Murad 1987; Drewett, Marchand, Ziegler
and Trachte 1988). However, a biphasic catecholamine secretion has also been caused
by 8 -bromo-cGMP from cultured bovine chromaffin cells (O'Sullivan and Burgoyne
1990). At low doses, 8 -bromo-cGMP caused potentiation, while at supramaximal
186

doses it caused inhibition of catecholamine release (O'Sullivan and Burgoyne 1990).
Moreover, NO and other NO generating agents have been shown to stimulate
catecholamine release accompanied with increased output of cGMP from perfused dog
adrenal gland (Dohi et al, 1983). In addition, NO has produced calcium and
concentration-dependent stimulation of basal catecholamine secretion from bovine
chromaffin cells (Oset-Gasque et at. 1994). It has also been found that NO stimulates
the catecholamine secretion evoked by small doses of nicotine, but inhibits at higher
doses of nicotine (Oset-Gasque et al. 1994). Furthermore, N ^-nitro-L-arginine (L-
NNA), an inhibitor of NOS, has been shown to reduce the acetylcholine-induced
increase in intracellular calcium level and catecholamine release which could be
restored by L-arginine (Uchiyama et al. 1994).
There are other studies which do not implicate NO and/or cGMP in
catecholamine secretion. The splanchnic nerve stimulation-induced catecholamine
secretion in pentobaribitol-anaesthetized dog was not affected by L-NAME (Breslow
et al. 1992). Certain stimuli that evoke catecholamine secretion, such as electrical
stimulation of splanchnic nerves, methacholine, dimethyl-4-phenylpiperazinium
iodide, high levels of potassium and alamethicin have caused a rise in cGMP from
perfused cat adrenal medulla (Moro, Michelena, Sanchez-Garcia, Palmer, Moncada
and Garcia 1993). However, although, L-NAME could abolish such rise in cGMP, it
did not cause any change in the catecholamine release (Moro et al. 1993). It is not
known at present as to what could be the reason for the discrepancies on the effect of
NO in the catecholamine secretion as inferred from the above mentioned studies. It is,
however, possible that differences in the methods and preparations used to elicit
catecholamine release, in the type of secretagogues used, and in their concentration and
duration of the stimuli as suggested by Torres et al. (1994), may partly account for the
discrepancies.
NO in the adrenal medulla may also be involved in the synthesis of other
187

substances such as the neuropeptides whose biosynthesis may not directly correlate with
that of the catecholamines. Although, adrenal peptides are costored and coreleased
with the catecholamines, their biosynthesis may be differently regulated, as is found for
enkephalins (see Naranjo, Mocchetti, Schwartz and Costa 1987). Elsewhere, NO has
already been found to modulate the release of corticotrophin-rcleasing hormone from
rat hypothalamus in vitro (Costa, Trainer, Besser and Grossman 1993) and inhibit
vasopressin and oxytocin secretion from isolated neural lobe of the rat pituitary gland
(Lutz-Bucher and Koch 1994). There is also evidence to suggest that NO modulates
the transcriptional induction of a number of genes, since it has been found to activate
the immediate early genes c-fos and jun-B in the PC 12 cells (Peunova and
Enikolopov 1993; Haby, Lisovoski, Aunis and Zwiller 1994).
4:3. Nitric oxide in the regulation of adrenal cortical secretion
It is apparent that adrenal cortical activity is influenced by neuronal
mechanisms, in addition to hormonal regulation. Electrical stimulation of the
splanchnic nerves in calf and dog causes an increase in the rate of ACTH-induced
cortisol secretion (Edwards and Jones 1987; Engeland and Gann 1989). Splanchnic
nerve stimulation has also been found to enhance the activity of 1 1 ^-hydroxylase,
which in turn increases the synthesis of cortisol from 11 deoxycortisol (Engeland and
Gann 1989). In contrast, transection of the splanchnic nerve in lamb and calf causes a
decrease in adrenocortical sensitivity to ACTH (Edwards and Jones 1987; Edwards,
Jones and Bloom 1986). Moreover, several mediators, including NO, which exist in the
intra-adrenal nerves and/or the chromaffin cells have been shown to influence
corticosteroid secretions from the gland (see Section 1:7).
Treatment of rats with the NOS inhibitor L-NAME causes a dose-dependent
increase in serum corticosterone levels, suggesting endogenous NO may negatively
regulate steroidogenesis (Adams et al. 1992). Perfusion of isolated rat adrenal gland
preparation with a medium which contains L-arginine at a higher dose above 500
188

fimo\/\ results in a decrease in adrenal corticosterone secretion (Cameron and Hinson
1993). However, at a lower dose of L-arginine upto 500 /^mol/1 an increase in the
corticosterone secretion occurs in a dose dependent manner (Cameron and Hinson
1993). It therefore appears that NO could influence adrenal cortical secretory activity.
With the majority of NOS-containing nerve fibres located in the subcapsular region
and the zona glomerulosa of the cortex, NO liberated from these nerve fibres may
reach the cortical cells in the other region by diffusion. Since the NOS-containing
intrinsic neurons are the source for the NO-containing nerve fibres which innervate the
cortex (Section 3:1:3), adrenal medullary neurons which synthesize NO therefore
appear to regulate the activity of adrenal cortical cells. Such local regulation has also
been suggested for adrenal neurons that contain the neuropeptides VIP and NPY
(Hokfelt et al. 1981; Holzwarth 1984; Maubert et al. 1993). Furthermore, there are a
population of chromaffin cells located in the cortex which are thought to exert a
paracrine influence on adrenal cortical activity (Palacios and Lafarga 1975; Bomstein
etal. 1990,1991; Vizi etal. 1992).
The adrenal cortex in turn is known to influence the secretion of adrenal
medullary chromaffin cells. Following hypophysectomy roughly 100 times more
glucocorticoids are required to restore the activity of adrenal PNMT than required for
those of extra-adrenal enzymes regulated by glucocorticoids (Pohorecky and Wurtman
1971; Wurtman, Pohorecky and Baliga 1972). Adrenaline synthesis by the adrenal
chromaffin cells has therefore been suggested to be regulated by very high
concentrations of glucocorticoids compared to the level found in the circulation
(Pohorecky and Wurtman 1971; Wurtman et al. 1972). Glucocorticoids also regulate
the expression of other adrenal chromaffin cell products, such as chromogranin A
(Fischer-Colbrie et al. 1988) and Leu-enkephalin (Henion and Landis 1992).
Furthermore, in the rat, the initial expression of PNMT and Leu-enkephalin occurs
after the precursor cells have arrived in the adrenal gland, and is temporarily correlated
189

with the large increase in the production of glucocorticoids by adrenal cortical cells at
E16 (Seidl and Unsicker 1989; Henion and Landis 1992). It therefore appears that there
is inter-regulation between medulla and cortex of the gland. However, although the
influence of adrenal medulla over cortex appears to be accomplished by the projections
of adrenal intrinsic neurons located in the medulla and the paracrine action of adrenal
chromaffin cells located in the cortex, it has not been unequivocally resolved as to how
the secretion from the adrenal cortex could reach the medulla in the high concentration
needed. The hypothesis of a portal circulation to facilitate such influence is unlikely,
since it is not supported by anatomical evidence (Coupland and Selby 1976; Murakami
et al. 1989a). Hamaji, Miyata and Kawashima (1985) have suggested that
glucocorticoids may reach the medullary cells either via the interstitial fluid of the
cortex or by free diffusion firom the adrenal cortical veins into the medullary interstitial
fluid. Moreover, there are islands of cortical cells found in the medulla of the gland
(Coupland 1965). As found in the present study, significant numbers of NADPH-
diaphorase positive but NOS-negative cells and which appear to be cortical cells are
seen in groups or singly dispersed throughout the medulla. These cells are negative for
the chromaffin cell marker TH (Section 3:1:4) and the general neuronal marker PGP
9.5 (unpublished observation). In addition, the disappearance of these cells following
hypophysectomy further suggested their cortical cell identity (Section 3:2:1). There is
therefore a possibility that such cortical cells found in the medulla could, at least partly,
be the source of local glucocorticoids needed by the chromaffin cells, and that further
work needs to be performed.
4:4. Nitric oxide in the regulation of adrenal blood flow
The mammalian adrenal gland is a highly vascular organ and receives high
levels of blood flow compared with other organs (see Vinson and Hinson 1992). The
blood flow through the gland is actively regulated and does not simply follow changes
in systemic blood pressure. However, a number of physiological conditions which
190

cause secretion from either medulla or cortex also result in increased adrenal blood
flow.
As found in the dog, blood flow between medulla and cortex may be regulated
independently. Splanchnic nerve stimulation is shown to cause an increase in the rate
of adrenal medullary blood flow without altering that of the adrenal cortex (Breslow,
Jordan, Thellman and Traystman 1987). The increase in adrenal medullary blood flow
in response to haemorrhagic hypotension is only accompanied by a transient decrease
in cortical blood flow (Breslow, Mennen, Koehler and Traystman 1986). However, in
the rat, indicating possible species difference, the proportion of blood flow to medulla
and cortex is conserved following haemorrhagic hypotension (Sparrow and Coupland
1987).
Although adrenal blood flow accompanies adrenal catecholamine secretion, it
appears that these two phenomena are regulated independently. As found from analysis
of the levels of catecholamine secretions with respect to the changes in the rate of
delivery of perfusion medium in isolated perfused rat adrenal gland, increase in adrenal
flow rate by itself has no effect on catecholamine secretion (Bouloux, Perrett, Sopwith
and Besser 1986). Furthermore, in spite of a 95 % abolition of the splanchnic nerve-
induced increase in catecholamine secretion that could be caused by administration of a
nicotinic antagonist hexaméthonium, the increase in medullary vasodilatation could
only be abolished by a combination of nicotinic and muscarinic antagonists but not by
either one alone (Kennedy, Breslow, Tobin and Traystman 1991). In contrast, adrenal
steroid secretion appears to be closely regulated with adrenal blood flow. At
submaximal concentration of ACTH, enhancement of adrenal steroid secretion in rat
and dog could be achieved by mechanically increasing the rate of blood flow in the
gland (Porter and Klaiber 1965; Urquhart 1965). In the calf, changes in the adrenal
flow rate is shown to cause changes in the rate of presentation of ACTH which in turn
correlates with the rate of adrenal steroid secretion (Jones, Edwards and Bloom 1990).
191

The regulation of blood flow in the gland may be effected through various
agents. ACTH causes a large increase in the rate of blood flow in the adrenal gland of
rat and dog (Hartman, Brownell and Liu 1955; Holzbauer and Vogt 1957). Certain
neuropeptides, which are known to exist in the splanchnic nerves, chromaffin cells
and/or adrenal intrinsic neurons, also appear to regulate blood flow as shown in
isolated perfused rat adrenal gland preparations (Hinson, Cameron, Purbic and Kapas
1994a). VIP was found to have the greatest effect causing 136 % increase in adrenal
flow rate. Met-enkephalin caused a 50 % increase, while leu-enkephalin, neurotensin
and substance P had up to 35 % increase in perfusion medium flow rate. In contrast,
NPY caused a 30 % decrease in the perfusion medium flow rate. Since NO is a
powerful vasodilator in several regions of the body (see Moncada et al. 1991;
Lowenstein et al. 1994), it may also be involved in the regulation of blood flow in the
adrenal gland. Indeed, as found in the isolated perfused rat adrenal gland preparation,
infusion with medium containing L-arginine increases adrenal flow rate in a direct
dose dependent manner, and such an increase is inhibited by L-NAME (Cameron and
Hinson 1993). In the anaesthetized dog, a substantial reduction in adrenal cortical
blood flow occurs following administration of L-NAME (Breslow, Tobin, Bredt,
Ferris, Snyder and Traysman 1993). Moreover, the increase in the adrenal blood flow
which accompanies the catecholamine secretion induced by splanchnic nerve
stimulation is associated with an increase in NOS activity, and that inhibition of NOS
by L-NAME abolishes the vasodilation response to splanchnic nerve stimulation
(Breslow et al. 1992,1993).
The NOS-positive nerve fibres found closely associated with adrenal blood
vessels and in the subcapsular region, may be the source of NO involved in the
regulation of blood flow in the gland. NADPH-diaphorase staining has been shown to
mark the presence of NOS in the endothelial cells of some blood vessels (Loesch, Belai
and Bumstock 1993, 1994; Tomimoto, Nishimura, Suenaga, Nakamura, Akiguchi,
Wakita, Kimura and Mayer 1994). However, the absence of NADPH-diaphorase
192

staining in the endothelial cells within the adrenal gland may suggest that the adrenal
nerves are the only source of the NO needed in normal physiological condition in the
gland. Nevertheless, further studies with specific antibody which recognizes
endothelial NOS may be needed to fully investigate the possibility of the presence of
NO-synthesizing endothelial cells which may have not been detected by NADPH-
diaphorase histochemistry in the gland.
4:5. Plasticity in nitrergic innervation
The production of NO is thought to be regulated in at least two ways. The NO
produced by the NOS isoforms I and III is mainly regulated by calcium and
calmodulin, where increase in the intracellular calcium concentration binds to
calmodulin and activates the enzyme (Knowles et al. 1989; Bredt and Snyder 1990;
Malinski and Taha 1992). The NO produced by the NOS isoform II, on the other hand,
is caused by an induction of the synthesis of the enzyme after challenge with a variety
of immunologic or inflammatory stimuli (Hibbs et al. 1987b; Vodovotz et al. 1994).
However, as found in the present study, the level of NOS immunohistochemically
recognized with antibody specific to isoform I, shows changes during development and
aging (Section 3:1:2), after reserpine treatment (Section 3:2:1) and in streptozotocin-
induced diabetes (Section 3:2:2). This suggests that the NOS isoform I may also be
induced. Other recent investigations have also shown the protein levels and/or
activities of the NOS isoforms I and III can be affected under various conditions such
as during development, pregnancy, neuronal lesions, neuronal activity, mechanical
stimuli and exercise, and by several agents such as NGF, estrogens, testosterone and
cytokines (Verge et a/.1992; Fiallos-Estrada, Kummer, Mayer, Bravo, Zimmermann
and Herdegen 1993; Hirsch, Steiner, Dawson, Mammen, Hayek and Snyder 1993;
Matsumoto, Pollock, Nakane and Forstermann 1993; Weiner, Lizasoan, Bayliz,
Knowles, Charles and Moncada 1993; Wu 1993; Wu and Li 1993; Bredt and Snyder
193

1994; Roskams, Bredt, Dawson and Ronnett 1994; Sessa et al. 1994).
The changes in the expression of NOS isoform I during development and after
neuronal lesions has thrown light onto possible additional functions, in addition to
neurotransmission. During early development, several regions of the brain c«idc the
cortical plate, sensory ganglia and olfactory epithelium express NOS (Bredt and
Snyder 1994; Roskams et at. 1994). The expression of NOS in these regions appears
when the cells have completed cell division and begun to extend processes. In some of
the regions NOS is expressed transiently and lost later in development (Bredt and
Snyder 1994). Furthermore, corticothalamic fibres express NOS when cortical neurons
cease dividing and extend processes (Bredt and Snyder 1994). Treatment of PC 12
cells with NGF, which also causes neurite extension (Greene and Tischler 1976), elicits
expression of the NOS isoform I (Hirsch et al. 1993). Based on such observations a
role for NO in modification of synaptic efficacy in the developing nervous system has
been suggested (Bredt and Snyder 1994; Roskams et al. 1994).
Several studies have shown the occurrence of changes in NOS expression
immediately following neuronal lesions. In adult rat, after olfactory bulboctomy the
newly developing neuronal cells become NOS positive as they extend processes to
replace those lost (Roskams et al. 1994). The number of NOS-positive neurons in
sensory ganglia increases after peripheral axotomy (Verge et al. 1992; Fiallos-Estrada
et al. 1993). Ventral root avulsion has resulted in expression of NOS staining in motor
neurons which are negative in unlesioned animals (Wu 1993). However, the increase in
expression of NOS, and thus enhanced production of NO following neuronal lesions is
not known as to whether it is related to trophic action or involved in the degeneration
process of the lesioned nerves. There is evidence where lesions of medial forebrain
bundle and mammilothalamic tract causes NOS staining in cell bodies of these
bundles, which persists for up to 150 days, implying a possible neuroprotective action
for NO (Herdegen, Brecht, Mayer, Leah, Kummer, Bravo and Zimmermann 1993). In
contrast, the NOS positive motor neurons expressed in the spinal cord following
194

ventral root avulsion have been found to die ultimately, whereas treatment of such
animals with the NOS inhibitor nitroarginine prior to ventral root avulsion protects the
cells from death (Wu and Li 1993). Furthermore, NO-generating agents have been
shown to cause a collapse of neuronal growth cones in regenerating dorsal root
ganglion cells (Hess, Patterson, Smith and Skene 1993).
Various factors and mechanisms may be responsible for the changes in the
expression of NOS. Increase in NOS expression in a tissue may be due to an increase
in number of cells that express the enzyme as a result of induction of its synthesis in
cells which previously did not contain the enzyme. It could also be due to proliferation
of cells with the capacity of expressing the enzyme. In contrast, decrease in the levels
of expression of the enzyme in a tissue may reflect a cessation of its synthesis from
cells, or may be due to a disappearance of the cells containing the enzyme as a result of
degeneration or cell death.
The induction and disappearance of transmitters and their synthesizing enzymes,
such as the NOS, in intact nerves may be responsible for causing changes in neuronal
phenotype and hence plasticity that occur in response to different stimuli. The basis for
the changes in the levels of transmitters and their synthesizing enzymes appears to be
effected mainly at the gene level, although up or down regulation of the activities of
preexisting enzymes may also be possible. It appears that all neurons carry the genes
responsible for the neurotransmitters and their synthesizing enzymes. Some of the
genes are silenced in certain subpopulations of neurons, while some other genes that
are appropriate to a given differentiated neurons are expressed by an active control
mechanism. Such processes enables neurotransmitter choice that is neither
precommitted nor stable, but is continuously influenced by environmental conditions.
Several factors such as the state of membrane electrical activity, and the levels of
neurotrophic factors, neurotransmitters and hormones could influence the expression of
the genes.
195

The mechanisms of change for the expression of NOS are not fully understood
at present, although most likely are mediated through alterations in NOS gene
expression, mRNA stability or protein biosynthesis (see Morris and Billiar 1994;
Nathan and Xie 1994b; Sessa 1994). In many instances the changes in the activity of
NOS isoform II have been correlated with similar changes in its mRNA abundance
which suggested the major part of the regulation of this enzyme is at a pretranslational
step such as transcription or mRNA stability (Xie, Cho, Calaycay, Mumford, Swiderck,
Lie, Ding, Troso and Nathan 1992; Nunokawa, Ishida and Tanaka 1993). Such
correlation of enzyme levels and/or activities with mRNA levels have also been
observed in studies involving the NOS isoforms I and III (Yoshizumi, Perrella, Burnett
and Lee 1993; Roskams et aL 1994; Sessa, Pritchard, Seyedi, Wang and Hintze 1994).
The levels of substrates, cofactors and products may also influence the state of
enzyme activity and could play an important role in determining rates of cellular NO
production. GTP-cyclohydrolase I, the first and rate-limiting enzyme in the synthesis
of BH4 from GTP, has been shown to be responsible for tumour necrosis factor-a,
interferon-r or LPS-induced increases in endothelial cell NOS activity (Wemer-
Felmayer, Werner, Fuchs, Hausen, Reibnegger, Schmidt, Weis and Wachter 1993).
Moreover, it has been shown that NO can inhibit the production of NO in cerebellum,
macrophages and central nervous system glial cells (Rogers and Ignarro 1992;
Assreuy, Cunha, Liew and Moncada 1993; Griscavage, Rogers, Shermann and Ignarro
1993; Park, Lin and Morphy 1994). Such inhibition of the production of NO by NO
itself is effected through the regulation of NOS catalytic activity by interaction with the
heme moiety of the enzyme (Rogers and Ignarro 1992; Griscavage et al. 1993) and
through the regulation of NOS expression by limiting transcriptional induction of the
gene (Park et al. 1994).
The expression of NOS and/or production of NO therefore appear to be complex
and is subject to regulation at various cellular levels. It is likely that some of the
changes in the expression of NOS in the adrenal gland which were observed in the
196

present study are correlated with changes at the molecular levels, such as gene
expression, mRNA stability and/or derangements in the enzyme biosynthesis or
activity. They may also be correlated with cellular and nerve fibre degeneration or
proliferation. Such changes and their effect may have not been fully revealed by the
present histochemical studies at the light microscopic level. Future investigations
which employ electron microscopy and techniques such as molecular biology and
biochemical methods may reveal more changes and throw light onto the mechanisms
of the plasticity.
197

REFERENCE
198

Aberdeen, J., Con, L., Milner, P., Lincoln, J. and Bumstock, G. (1990) Marked increase in calcitonin gene-related peptide-containing nerves in the developing rat following long-term sympathectomy with guanethidine. Neuroscience 35, 175- 184.
Adams, M.L., Nock, B., Truong, R. and Ckero, T J. (1992) Nitric oxide control of steroidogenesis: endocrine effects of N ^-nitro-L-arginine and comparisons to alcohol. Life Sci 50, PL-35-PL-40.
Addison, T. (1855) On the constitutional and local effects o f disease o f suprarenal capsule. S. Highley: London..
Afework, M. (1988) The Sensory Innervation o f the Rat Adrenal Gland. M. Phil Thesis, University of Nottingham.
Aimi, Y., Fujimura, M., Vincent, S.R. and Kimura, H. (1991) Localization of NADPH-diaphorase-containing neurons in sensory ganglia of the rat. J Comp Neurol 306, 382-392.
Aldrich, T.B. (1901) A preliminary report on the active principle of the suprarenal Am J Physiol 5, 457-461.
Allen, J.M., Eranko, O. and Hunter, R.L. (1958) A histochemical study of the esterases of the adrenal medulla of the rat. J Anat 102, 93-116.
Aim, P., Larsson, B., Ekblad, E., Sundler, F. and Andersson, K.-E. (1993) Immunohistochemical localization of peripheral nitric oxide synthase-containing nerves using antibodies raised against synthesized C - and N-terminal fragments of a cloned enzyme from rat bidAn.Acta Physiol Scand 148, 421-429.
Aloe, L. and Levi-Montalcini, R. (1980) Comparative studies on the effect elicited by pre- and postnatal injections of anti-NGF, guanethidine, and 6 -hydroxydopamine in chromaffin and ganglion cells of the adrenal medulla and carotid body in infant rats. Adv Biochem Psychopharmacol 25, 222-226.
Alpert, L.K. (1931) The innervation of the suprarenal gldûtids. Anat Rec 50, 221-233.
Altman, J. and Das, G.D. (1966) Autoradiographic and histological studies of postanatal neurogenesis. I. A longitudinal investigation of the kinetics, migration and transformation of cells incorporating tritiated thymidine in neonate rats, with special reference to postnatal neurogenesis in some brain regions. J Comp Neurol 126, 337-390.
Anastasi, A., Erspamer, V. and Bucci, M. (1971) Isolation and structure of bombesin and alytesin, two analogous active peptides from the skin of the European amphibians and Alytes. Experimentia 27,166-167.
Anderson, D.J. (1993) Molecular control of cell fate in the neural crest: the sympathoadrenal lineage, Rev Neurosci 16,129-158.
Anderssen C., Blümcke, I. and Celio, M R. (1993) Calcium-binding proteins: selective markers of nerve cells. Cell Tissue Res 271,181-208.
Ando, S. (1983) Gangliosides in the nervous system. Neurochem Int 5, 507-537.
199

Angeletti, P.U. and Levi-Montalcini, R. (1970) Sympathetic nerve cell destruction in newborn mammals by 6 -hydroxydopamine. Proc Natl Acad Sci USA 65,112-121.
Angeletti, P.U. and Levi-Montalcini, R. (1972) Growth inhibition of sympathetic cells by some adrenergic blocking agents. Proc Natl Acad Sci USA 69, 8 6 - 8 8 .
Arnold, W.P., Mittal, C.K., Katsuki, S. and Murad, F. (1977) Nitric oxide activates guanilate cyclase and increases guanosine 3':5-cyclic monophosphate levels in various tissue preparations. Proc Natl Acad Sci USA 74, 3203-3207.
Assreuy, J., Cunha, F.Q., Liew, F.Y. and Moncada, S. (1993) Feedback inhibition of nitric oxide synthase activity by nitric oxide. B rJ Pharmacol 108, 833-837.
Attia, M.A. (1985) Neoplastic and noneoplastic lesions in aging female rats with special reference to the functional morphology of the hyperplastic and neoplastic changes in the pituitary gland. /trcA Toxicol 57, 77-83.
Axelrod, J. (1971) Noradrenaline: fate and control of its biosynthesis. Science 173, 598-606.
Axelrod, J. and Reisine, T.D. (1984) Stress hormones: their interaction and regulation. Science 224, 452-459.
Bacon, S.J. and Smith, A.D. (1988) Preganglionic sympathetic neurones innervating the rat adrenal medulla: immunocytochemical evidence of synaptic input from nerve terminals containing substance P, GABA or 5-hydroxytryptamine. J Auton Nerv Syst 24, 97-122.
Baker, P.F. and Kniÿit, D.E. (1978) Calcium dependent exocytosis in bovine adrenal medullary cells with leaky plasma membranes. Nature 276, 620-622.
Baker, P.F. and Knight, D.E. (1981) Calcium control of exocytosis and endocytosis in bovine adrenal medullary cells. Phil Trans Roy Soc B96, 83-103.
Baljet, B. and Drukker, J. (1979) The extrinsic innervation of the abdominal organs in the female rat. Acta Anat 104, 243-267.
Bandaletova, T., Brouet, I., Bartsch, H., Sugimura, T., Esumi, H. and Ohshima, H. (1993) Immunohistochemical localization of an inducible form of nitric oxide synthase in various organs of rats treated with propionibacterium-acnes and lipopolysaccharide. APM/5 101, 330-336.
Bartholinus, C. (1611) Anatomicae Institutiones Corporis Rumania, Wittenberg. Quoted By Rolleston (1936).
Baruchin, A., Vollmer, R.R., Miner, L.L., Sell, S.L., Strieker, E.M. and Kapllan, B.B. (1993) Cold-induced increases in phenylethanolamine N-methyltransferase (PNMT) mRNA are mediated by non-cholinergic mechanisms in the rat adrenal gland. Neurochem Res 18, 759-766.
Bastianelli, E. and Pochet, R. (1993) Transient expression of calretinin during development of chick cerebellum. Comparison with calbindin - D28K. Neurosci Res 17, 53-61.
200

Bêlai, A., Lincoln, J., Milner, P. and Bumstock, G. (1988) Progressive changes in adrenergic, serotonergic, and peptidergic nerves in proximal colon of streptozotocin-diabetic rats. Gastroenterology 95,1234-1241.
Belai, A., Lincoln, J., Milner, P., Crowe, R., Locsch, A. and Bumstock, G. (1985) Enteric nerves in diabetic rats: increase in vasoactive intestinal polypeptide but not substance P. Gastroenterology 89, 967-976.
Belai, A., Schmidt, H.H.H.V., Hoyle, C.H.V., Hassall, C.J.S., Saffrey, M.S., Moss, J., Forstermann, U., Murad, F. and Bumstock, G. (1992) Colocalization of nitric oxide synthase and NADPH-diaphorase in the myenteric plexus of the rat gut. Neurosci Lett 143, 60-64.
Belloni, A S., Rebuffat, P., Malendowicz, L.K., Mazzocchi, G., Rocco, S. Nussdorfer,G.G. (1992) Age-related changes in the morphology and function of the zona glomemlosa of the rat adrenal cortex. Tissue Cell 24, 835-842.
Beltz, B.S. and Burd, G.D. (1989) Immunocytochemical Techniques: Principles and Practice, Blackwell: Cambridge.
Belvis, M.G., Stretton, C.D., Yacoub, M. and Bames, P.J. (1992) Nitric oxide is the endogenous neurotransmitter of bronchodilator nerves in humans. Eur J Pharmacol 210, 221- 222.
Bennett, H.S. and Kilham, L. (1940) The blood vessels of the adrenal gland of the adult CdX.Anat Rec 77, 447-471.
Bennett, L.L. and Koneff, A.A. (1946) Atrophy of the thyroid and hypertrophy of the adrenal in rats with alloxan diabetes, Rec 96,1-12.
Bennett, T., Bumstock, G., Cobb, J.L.S. and Malmfors, T. (1970) An ultrastmctural and histochemical study of the short-term effects of 6 -hydroxydopamine on adrenergic nerves in the domestic fowl. Br J Pharmacol 38, 802-809.
Bordeaux, A. (1993) Nitric oxide: an ubiquitos messenger. Fundam Clin Pharmacol 7, 401-411.
Benyamina, M., Leboulenger, F., Lirhmann, I., Delarue, C., Feullioley M. and Vaudry,H. (1987) Acetylcholine stimulates steroidogenesis in isolated frog adrenal gland through muscarinic receptors: evidence for a desensitization mechanism. J Endocrinol 113, 339-348.
Bergstrom, S., von Euler, U.S. and Hamberg, U. (1949) Isolation of noradrenaline from the adrenal gland. Acta Chem Scand 3, 305.
Bertholet, J.Y. (1980) Proliferative activity and cell migration in the adrenal cortex of fetal and neonatal rats: an autoradiographic study. J Endocrinol 87 ,1-9 .
Biedl, A. (1913) The Internal Secretory Organs. Their Physiology and Pathology. W.M. Woods: New York.
Blaschko, H. (1939) The specific action of L-DOPA decarboxilase. J Physiol 96, 50- 51P.
Bloom, S R., Edwards, A.V. and Jones, C.T. (1989) Adrenal responses to calcitonin
201

gene-related peptide in conscious hypopysectomised CdX\^s.J Physiol 409, 29-41.
Blottner, D. and Baumgarten, H.G. (1992) Nitric oxide synthase (NOS)-containing sympathoadrenal cholinergic neurons of the rat IML-cell column: evidence from histochemistry, immunohistochemistry, and retrograde labelling. J Comp Neurol 316, 45-55.
Bohme, G.A., Bon, C., Stutzmann, J.-M., Doble, A. and Blanchard, J.-C. (1991) Possible involvement of nitric oxide in long-term potentiation. Eur J Pharmacol 199, 379-381.
Bohme, G A., Bon, C., Lemaire, M., Reibaud, M., Piot, O., Stutzmann, J.-M., Doble, A. and Blanchard, J.-C. (1993) Altered synaptic plasticity and memory formation in nitric oxide inhibitor treated rats. Proc Natl Acad Sci USA 90, 9191-9194.
Bohn, M.C., Goldstein, M. and Black, I B. (1981) Role of glucocorticoids in expression of the adrenergic phenotype in rat embryonic adrenal gland. Dev Biol 82, 1- 10.
Bohn, M., Kessler, L., Golightly, L. and Black, I. (1983) Appearance of enkephalin- immunoreactivity in rat adrenal medulla following treatment with nicotinic antagonist or reserpine. Cell Tissue Res 231, 469-479.
Boje, K.M. and Arora, P.K. (1992) Microglial-produced nitric oxide and reactive nitrogen oxides mediate neuronal cell death. Brain Res 587, 250-256.
Boksa, P. and Livett, B.G. (1984) Substance P protects against desensitization of the nicotinic response in isolated adrenal chromaffin ccWs.J Neurochem 42, 618-627.
Bolli, G., De Feo, P., Campagnucci, P., Cartechini, M.G., Angeletti, G., Santeusanio, F. and Brunetti, P. (1982) Important role of adrenergic mechanisms in acute glucose counterregulation following insulin-induced hypoglycemia in type I diabetes: evidence for an effect mediated by beta-adrenocepotors. Diabetes 31, 641-647.
Bondy, P.K. (1985) Disorders of the adrenal cortex. In: Textbook o f Endocrinology, 7^ ̂edn. (eds. J.D. Wilson and D.W. Foster), pp. 816-890. W.B. Sounders: Philadelphia.
Borges, R. (1994) Histamine HI receptor activation mediates the preferential release of adrenaline in the adrenal gland. Life Sci 54, 631-640.
Bomstein, S R., Ehrhart-Bomstein, M., Scherbaum, W.A., Pfeiffer, E.F., and Holst, J.J. (1990) Effects of splanchnic nerve stimulation on the adrenal cortex may be mediated by chromaffin cells in a paracrine manner. Endocrinology 127, 900-906.
Bomstein, S R., Ehrhart-Bomstein, M., Usadel, H., Bockmann, M. and Scherbaum, W.A. (1991) Morphological evidence for a close interaction of chromaffin cells with cortical cells within the adrenal gland. Cell Tissue Res 256,1-9.
Bouloux, P.-M.G., Perrett, D., Sopwith, M. and Besser, G.M. (1986) An in situ isolated rat adrenal perfusion system for study of neuronally mediated catecholamine secretion: effects of morphine, a met-enkephalin analogue, and naloxane on catecholamine scciction. J Endocrinol 111, 7-15.
Boume, G.H. (1961) Comparative anatomy and embryology of the adrenal gland. In:
202

The Adrenal Cortex (ed. H.D. Moon), pp. 1-22. Paul B Hoeber Inc: USA.
Brazeau, P., Vale, W., Burgus, K., Ling, N., Butcher, M., Rivier, J. and Guillemin, R. (1973) Hypothalamic polypeptide that inhibits the secretion of immunoreactive pituitary growth hormone. Science 179, 77-79.
Bredt, D.S. and Snyder, S.H. (1990) Isolation of nitric oxide synthase, a calmoduline- requiring enzyme. Proc Natl Acad Sci USA 87, 682-685.
Bredt, D.S. and Snyder, S.H. (1994) Transient nitric oxide synthase neurons in embryonic cerebral cortical plate, sensory ganglia, and olfactory epithelium. Neuron 13, 301-313.
Bredt, D.S., Hwang, P.M. and Snyder, S.H. (1990) Localization of nitric oxide synthase indicating a neuronal role for nitric oxide. Nature 347, 768-770.
Breslow, M.J., Jordan, D.A., Thellman, S T. and Traystman, R.J. (1987) Neural control of adrenal medullary and cortical blood flow during hemorrhage. Am J Physiol 252, H521-H528.
Breslow, M.J., Mennen, A., Koehler, R.C. and Traystman, R.J. (1986) Adrenal medullary and cortical blood flow during hemorrhage. Am J Physiol 250, H954- H960.
Breslow, M.J., Tobin, J R., Bredt, O.S., Ferris, C D., Snyder, S.H. and Traystman, R.J.(1992) Role of nitric oxide in adrenal medullary vasodilatation during catecholamine secretion. Eur J Pharmacol 210,105-106.
Breslow, M.J., Tobin, J R., Bredt, D.S., Ferris, C D., Snyder, S.H. and Traystman, R.J.(1993) Nitric oxide as a regulator of adrenal blood flow. Am J Physiol 33, H464- H469.
Brodie, B.B., Pletscher, A. and Shore, P.A. (1955) Evidence that serotonin has a role in brain function. Science 122, 968.
Brookes, S.J.H., Steele, P.A. and Costa, M. (1991) Calretinin immunoreactivity in cholinergic motor neurones, intemeurones and vasomotor neurones in the guinea- pig small intestine. Cell Tissue Res 263, 471-481.
Brown, F.M., Smith, A.M., Longway, S. and Rabinowe, S.L. (1990) adrenal medullitis in type I diabetes. J Clin Endocrinol Metab 71,1491-1495.
Brown, F.M., Zuckerman, M., Longway, S. and Rabinowe, S.L. (1989) Adrenal medullary fibrosis in IDDM of long duration. Diabetes Care 12, 494-497.
Brown-Sequard, C.E. (1856) Recherches expérimentales sur la physiologie et la pathologie des capsules surrénales. Compt Red 43, 422-425. Quoted by Gaunt (1975).
Bnihn, T.O., Engeland, W.C., Anthony, E.L., Gann, D.S. and Jackson, I.M. (1987) Corticotropin-releasing factor in the dog adrenal medulla is secreted in response to hemorrhage. Endocrinology 120, 25-33.
Bruzzone, M E. and Marusic, E.T. (1988) Effects of opioid peptides on aldosterone production: stimulatory effect of Leu-enkephalin. Endocrinology 122, 402-406.
203

Bryant, M.G., Bloom, S.R., Polak, J.M., Albuquerque, R.H., Modlin, I. and Pearse, A.G. (1976) Possible dual role for vasoactive intestinal peptide as gastro-intestinal hormone and neurotransmitter substance. Lancet 1, 991-993.
Buffa, R., Mare, P., Salvadore, M., Solcia, E., Furness, J.B. and Lawson, D E M. (1989) Calbindin 28 kDa in endocrine cells of known or putative calcium- regulating function. Thyro-parathyroid C cells, gastric ECL cells, intestinal secretin and enteroglucagon cells, pancreatic glucagon, insulin and pp cells, adrenal medullary NA cells and some pituitary (TSH?) cells. Histochemistry 91,107-113.
Burgoyne, R.D. (1991) Control of exocytosis in adrenal chromaffin cells. Biochem Biophys Acta 1071,174-202.
Burnett, A.L., Lowenstein, C.J., Bredt, D.S., Chang, T.S. and Snyder, S.H. (1992) Nitric oxide: a physiologic mediator of penile erection. Science 257, 401-403.
Bumstock, G. (1972) Purinergic nerves. Pharmacol Rev 24, 509-581.
Bumstock, G. (1975) Purinergic transmission. In: Handbook o f Psychopharmacology, vol. 5 (eds. L.L. Iversen, S.D. Iversen and S.H. Snyder), pp. 131-194. Plenum Press: New York.
Bumstock, G. (1986) Autonomic neuromuscular junctions: current developments and future directions. 146,1-30.
Bumstock, G. (1990) Changes in expression of autonomic nerves in aging and disease. J Auton Nerv Syst 30, S25-S34.
Bumstock, G., Evans, B., Gannon, B.J., Heath, J.W. and James V. (1971) A new method of destroying adrenergic nerves in adult animals using guanethidine. Br J Pharmacol 43, 295-301.
Bush, LG. (1953) Species differences in adrenocortical secretion. J Endocrinol 9, 95- 100.
Bush, P.A., Gonzalez, N.E. and Ignarro, L.J. (1992) Biosynthesis of nitric oxide and citmlline from L-arginine by constitutive nitric oxide synthase present in rabbit corpus cavemosum. Biochem Biophys Res Commun 186, 308-314.
Buttery, L.D.K., Springall, D R., Carpenter, A., Riveros-Moreo, V., Moncada, S., Cohen, J. and Polak, J.M. (1993) Distinct localization of inducible nitric oxide synthase in endotoxin-treated rats. Endothelium [Suppl] 1, S27.
Cameron, L.A. and Hinson, J.P. (1993) The role of nitric oxide derived from L- arginine in the control of steroidogenesis, and perfusion medium flow rate in the isolated perfused rat adrenal gland. 7 Endocrinol 139, 415-423.
Cameron, O.G., Thomas, B., Tiongco, D., Hariharan, M. and Greden, J.F. (1987) Hypercortisolism in diabetes mellitus. Diabetes Care 10, 662-664.
Campbell, A.K. (1983) Intracellular calcium: its universal role as regulator. In: Monographs in Molecular Biophysics and Biochemistry (ed. H. Gutfreund), pp. 1- 556. John Wiley: New York.
204

Cannon, W.B., Lewis, J.T. and Britton, S.W. (1926) Studies on the conditions of activity in endocrine glands. XVII. A lasting preparation of the denervated heart for detecting internal secretion, with evidence for accessory accelerator fibres from the thoracic sympathetic chain. Am J Physiol 77, 326-352.
Carafoli, E. and Penniston, J.T. (1985) The calcium signal. Sci Am 253, 70-78.
Carlsson, A. (1965) Drugs that block storage of 5-hydroxytryptamine and related amines. In: Handbook o f Experimental Pharmacology^ vol. 19 (ed. V. Erspamer), pp. 529-592. Springer: Berlin.
Carmichael, S.W. (1983) The adrenal chromaffin vesicle: an histochemical perspective. J Auton Nerv Syst 7, 7-12.
Carraway, R. and Leeman, S.E. (1975) The amino acid sequence of a hypothalamic peptide, neurotensin. / Bio/ Chem 250,1907-1911.
Carraway, R. and Leeman, S. (1973) The isolation of a new hypotensive peptide, neurotensin, from bovine hypothalami. J Bio/ Chem 248, 6854-6861.
Carsia, R.V. and Malamed, S. (1989) The adrenals. In: Development, Maturation, and Senescence o f Neuroendocrine System: A Comparative Approach (eds. M.P. Schreibman and C.G. Scanes), pp. 353-380. Academic Press: New York.
Chalfe, M. and Perlman, R.L. (1976) Studies of transplantable rat phaeochromocytoma: biochemical characterization and catecholamine secretion. J Pharmacol Exp Ther 197, 615-622.
Chang, M.M., Leeman, S.E. and Niall, H.D. (1971) Amino-acid sequence of substance P. Nature 232, 86-87.
Chapman, P.P., Atkin, C.M., Allen, M.T., Haley, I.E. and Steinmetz, I.E. (1992) Inhibition of nitric oxide synthesis impairs two different forms of learning. NeuroReport 3, 567-570.
Charlton, B.G. (1990) Adrenal cortical innervation and glucocorticoid secretion. J Endocrinol 126, 5-8.
Cheng, L. (1980) Phaeochromocytoma in rats: Incidence, etiology, morphology and functional activity. 7 Bnviro/i Pathol Toxicol 4, 219-228.
Chem, Y.-J., Herrera, M., Kao, L.S. and Westhead, E.W. (1987) Inhibition of catecholamine secretion from bovine chromaffin cells by adenine nucleotides and adenosine. J Neurochem 48,1573-1576.
Chem, Y.-J., Kim, K.T., Slakey, L.L. and Westhead, E. (1988) Adenosine receptors activate adenylate cyclase and enhance secretion firom bovine adrenal chromaffin cells in the presence of forskoWn. J Neurochem 50,1484-1493.
Chester Jones, I. (1948) Variation in the mouse adrenal cortex with special reference to the zona reticularis and to brown degeneration, together with a discussion of the "cell migration" theory. Quart J Microsc Sci 89, 53-74.
Cheung, C.Y. and Holzwarth, M.A. (1986) Fetal adrenal VIP: distribution and effect on medullary catecholamine secretion. Peptides 7, 413-418.
205

Cho, H.J., Xie, Q.-U., Calaycay, J., Mumford, RA., Swiderck, K.M., Lee, T.D., and Nathan, C. (1992) Calmodulin is a subunit of nitric oxide synthase from macrophages. J Exp Med 176, 599-604.
Ciaranello, R.D., Wooten, G.F. and Axelrod, J. (1976) Regulation of rat adrenal dopamine /5-hydroxilase. II. Receptor interaction in the regulation of enzyme synthesis and degradation. Brain Res 113, 349-362.
Coggi, E., DeirOrto, P. and Viale, G. (1986) Avidin-biodn methods. In: Immunocytochemistry, Modern Methods and Applications, 2'*^ edn. (eds. J.M. Polak and S. Van Noorden), pp. 54-70. John W ri^ t and Sons: Bristol.
Coons, A.H. and Kaplan, M.H. (1950) Localization of antigen in tissue cells. II. Improvements in a method for the detection of antigen by means of fluorescent antibody. J Exp Med 91,1-13.
Coons, A.H., Creech, H.J. and Jones, R.N. (1941) Immunological properties of an antibody containing a fluorescent group. Proc Soc Exp Biol Med 47, 200-202.
Costa, A., Trainer, P., Besser, M. and Grossman, A. (1993) Nitric oxide modulates the release of corticotropin-releasing hormone from the rat hypothalamus in vitro. Brain Res 605,187-192.
Costa, M., Furness, J.B., Pompolo, S., Brookes, S.J.H., Bomstein, J.C., Bredt, D.S. and Snyder, S.H. (1992) Projections and chemical coding of neurons with immunoreactivity for nitric oxide synthase in the guinea-pig small intestine. Neurosci Lett 148,121-125.
Coupland, R.B. (1952) The prenatal development of the abdominal para-aortic bodies in man. J Anat 8 6 , 357-372.
Coupland, R.E. (1965) The Natural History o f the Chromaffin Cells. Longmans: U)ndon.
Coupland, R.E. (1975) Blood supply of the adrenal gland. In: Handbook o f Physiology, Section 7, Endocrinology, Vol. 11, Adrenal Glar^ (eds. H. Blaschko, G. Sayers and A.D. Smith), pp. 283-294. American Physiological Society: Washington DC.
Coupland, R.E. (1980) The development and fate of catecholamine secreting endocrine cells. In: Biogenic Amines in Development (eds. H. Parvez and S. Parvez), pp. 3-28. Elsevier: Amsterdam.
Coupland, R.E. (1984) Ultrastmctural features of the mammalian adrenal medulla. In: Ultrastructure o f Endocrine Cells and Tissues, (ed. P.M. Motta), pp. 168-179. Martinus Nijhoff: Boston.
Coupland, R.E. (1989) The natural history of the chromaffin cell - Twenty five years on the beginning, Histol Cytol 52, [Suppl] 331-341.
Coupland, R.E. and Holmes, R.L. (1958) The distribution of cholinesterase in the adrenal glands of the rat, cat and labbit. J Physiol 141, 97-106.
Coupland, R.E. and Selby, J.E. (1976) The blood supply of the mammalian adrenal medulla: a comparative study. 7 122, 539-551.
206

Coupland, R.E. and Tomlinson, A. (1989) The development and maturation of adrenal medullary chromaffin cells of the rat in vivo: a descriptive and quantitative study. Int J Dev Neurosci 7, 419-438.
Coupland, R.E., Kobayashi, S. and Tomlinson, A. (1977) On the presence of small granule chromaffin (SGC) cells in the rodent adrenal medulla. J Anat 124, 488-489.
Coupland, R.E., Parker, T.L., Kesse, W.K. and Mohammed, A.A. (1989) The innervation of the adrenal gland. III. Vagal innervation. 163,173-181.
Cryer, P.E. (1981) Glucose countrregulation in man. Diabetes 30, 261-264.
Cryer, P.E. (1989) Decreased sympathochromaffin activity in IDDM. Diabetes 38, 405-409.
Cuello, A C. (1990) Glycosphingolipids that can regulate nerve growth and repair. Adv Pharmacol 21,1-50.
Cummings, J.F. (1969) Thoracolumbar preganglionic neurons and adrenal innervation in the dog. Acta Anat 73, 27-37.
Cuvier, G. (1805) Leçons d'anatomie Comparée, Paris, v 240. Quoted by Rolleston (1936).
Dagerlind, A., Goldstein, M. and Hokfelt, T. (1990) Most ganglion cells in the rat adrenal medulla are noradrenergic. NeuroReport 1,137-140.
Dagerlind, Â., Pelto-Huikko, M., Lundberg, J.M., Ubink, R., Vcrhofstad, A., Brimijoin, S and Hokfelt, T. (1994) Immunologically induced sympathectomy of preganglionic nerves by antibodies against acetylcholinesterase: increased levels of peptides and their mRNAs in rat adrenal chromaffin cells. Neuroscience 62, 217- 239.
Daikoku, S., Kinutani, M. and Sato, M. (1977) Development of the adrenal medullary cells in rats with reference to synaptogenesis. Cell Tissue Res 179, 77-86.
Dallman, M.F., Engeland, W.C. and Shinsako, J. (1976) Compensatory adrenal growth: a neurally mediated leüex. Am J Physiol 231, 408-414.
Dawson, T.M., Bredt, D.S., Fotuchi, M., Hwang, P.M. and Snyder, S.H. (1991) Nitric oxide synthase and neuronal NADPH diaphorase are identical in brain and peripheral tissues. Proc Natl Acad Sci USA 8 8 , 7797-7801.
Dean, D M., Lemaire, S. and Livett, B.G. (1982) Evidence that inhibition of nicotine- mediated catecholamine secretion from adrenal chromaffin cells by enkephalin, fi- endorphin, dynorphin (1-13), and opiates is not mediated via specific opiate leceptOTS. J Neurochem 38, 606-614.
Deane, H.M. and Greep, R.O. (1946) A morphological and histochemical study of the rat's adrenal cortex after hypophysectomy, with comments on the liver. Am J Anat 79,117-145.
Deane, H.M., Shaw, J.H. and Greep, R.O. (1948) The effect of altered sodium or potassium intake on the width and cytochemistry of the zona glomemlosa of the
207

rat's adrenal cortex. Endocrinology 43,133-153.
De Chaplain, J. (1971) Degeneration and regrowth of adrenergic nerve fibres in the rat peripheral tissues after 6 -hydroxydopamine. Canad J Physiol Pharmacol 49, 345- 355.
De Lean, A., Racz, K., McNicoll, N. and Desrosiers, M.L. (1984) Direct )5-adrenergic stimulation of aldosterone secretion in cultured bovine adrenal subcapsular cells. Endocrinology 115, 485-492.
Demis, D.J., Blaschko, H. and Welch, A.D. (1956) The conversion of dihydroxyphenylalanine-2-C (DOPA) to norepinephrine by bovine adrenal homogenates./P/iarmaco/117, 208-212.
Dempster, W.J. (1974) The nature of the venous systems in the adrenal gland. Tohoku J Exp Med 112, 63-77.
De Nicola, A.F., Fridman, O., Del Castillo, E.J. and Foglia, V.G. (1976) The influence of streptozotocin diabetes on adrenal function in male rats. Horm Metab Res 8 , 388-392.
De Quidt, M.E. and Emson, P.C. (1986) Neuropeptide Y in the adrenal gland: characterisation, distribution and drug effects. Neuroscience 19,1011-1022.
Derome, G., Tseng, R., Mercier, P., Lemaire, I. and Lemaire, S (1981) Possible muscarinic regulation of catecholamine secretion mediated by cyclic GMP in isolated bovine adrenal chromaffin cell. Biochem Pharmacol 30, 855-860.
Dohi, T., Morita, K. and Tsujimoto, A. (1983) Effect of sodium azide on catecholamine release from isolated adrenal gland and on guanylate cyclase. Eur J Pharmacol 94, 331-335.
Dorovini-Zis, K. and Zis, A.P. (1991) Innervation of the zona fasciculata of the adult human adrenal cortex: a light and microscopic study. JWewra/ Transm 84, 75-84.
Dorsey, D.A. and Schmidt, R.E. (1993) Correlation of GAP-43 immunoreactivity with subpopulations of chromaffin cells in rat adrenal medulla. Neurosci Lett 162, 29- 33.
Douglas, W.W. and Poisner, A.M. (1965) Preferential release of adrenaline from adrenal medulla by muscarine and pilocarpine. Nature 208,1102-1103.
Douglas, W.W., Poisner, A.M. and Rubin, R.P. (1965) Exflux of adenine nucleotides from perfused adrenal glands exposed to nicotine and other chromaffin cells stimulants. J Physiol 179,130-137.
Drewett, J., Marchand, G., Ziegler, R. and Trachte, G. (1988) Atrial natriuretic factor inhibits norepineplnine release in an adrenergic clonal cell line (PC12). Eur J Pharmacol 150,175-179.
Dreyer, G.P. (1899) On secretory nerves to suprarenal capsules. Am J Physiol 2, 203- 219.
Drumond, G.I. (1983) Cyclic nucleotides in the nervous system. Adv Cyclic Nucleotide Res 15, 373-494.
208

Dun, N.J., Dun, S.L., Wu., S.Y. and Forstermann, U. (1993) Nitric oxide synthase immunoreactivity in rat superior cervical ganglia and adrenal glands. Neurosci Lett 158, 51-54.
Edwards, A.V. (1982) Adrenal catecholamine output in response to stimulation of the splanchnic nerve in bursts in the conscious ceXi. J Physiol 327, 409-419.
Edwards, A.V. and Jones, C.T. (1987) The effect of splanchnic nerve stimulation on adrenocortical activity in conscious caXvts. J Physiol 382, 385-396.
Edwards, A.V. and Jones, C.T. (1990) Adrenal responses to intra-aortic infusions of acetylcholine in conscious, functionally hypophysectomised calves. J Physiol 423, 13p.
Edwards, A.V., Jones, C.T. and Bloom, S.R. (1986) Reduced adrenal cortical sensitivity to ACTH in lambs with cut splanchnic nerves. J Endocrinol 110, 81-85.
Ehrhart-Bomstein, M., Bomstein, S R., Scherbaum, W.A., Pfeiffer, E.F. and Holst, J. (1991) Role of the vasoactive intestinal peptide in a neuroendocrine regulation of the adrenal cortex. Neuroendocrinology 54, 628-633.
Ehrlich, E.N., Nolten, W.E., Oparil, S. and Lindheimer, M.D. (1976) Mineralocorticoids in normal pregnancy. In: Hypertension in Pregnancy (eds. M.D. Lindheimer, A.L. Katz and F.P. Zuspan), pp. 189-201. John Wiley and Sons: New York.
Ekman, R., Bjartell, A., Eklblad, E. and Sundler, F. (1987) Immunoreactive delta sleep-inducing peptide in pituitary adrcnocorticotrophic/alpha-melanotropin cells and adrenal medullary cells of the pig. Neuroendocrinology 45, 298-304.
Elliott, T.R. (1912) The control of the suprarenal glands by the splanchnic nerves. J Physiol 44, 374-409.
Elliott, T.R. (1913) The innervation of the adrenal ^3.nd. J Physiol 46, 285-290.
Ellis, J.H., Richards, D.E. and Rogers, J.H. (1991) Calretinin and calbindin in the retina of the developing chick. Cell Tissue Res 264,197-208.
Ellison, J.P. and Clark, G.M. (1975) Retrograde axonal transport of horseradish peroxidase in peripheral autonomic nerves. J Comp Neurol 161, 103-114.
Engeland, W.C. and Gann, D.S. (1989) Splanchnic nerve stimulation modulates steroid secretion in hypophysectomized dogs. Neuroendocrinology 50,124-131.
Engeland, W.C., Lilly, M.P. and Gann, D.S. (1985) Sympathetic adrenal denervation decrease adrenal blood flow without altering the cortisol response to hemorrhage. Endocrinology 117,1000-1010.
Enyedi, P., Szabo, B. and Spat, A. (1983) Failure of vasoactive intestinal peptide to stimulate aldosterone production, Physiol Hung 61, 77-79.
Eranko, O. (1952) On the histochemistry of the adrenal medulla of the rat, with special reference to acid phosphatase. Acta Anat [Basel, Suppl] 17,1-60.
209

Erânkô, O. (1955) Distribution of adrenaline and noradrenaline in the adrenal medulla. Nature 175, 88-89.
Eranko, O. (1959) Specific demonstration of acetylcholinesterase and nonspecific cholinesterase in the adrenal gland of the rat. Histochemie 1, 257-267.
Eranko, O. and Eranko, L. (1971) Histochemical evidence of chemical sympathectomy by guanethidine in newborn rats. Histochem J 3, 451-456.
Eustachius, B. (1563) Opuscula anatomica libellus de renibus vent. Quoted by Rolleston (1936).
Farber, E., Sternberg, W.H. and Dunlap, C.E. (1956) Histochemical localization of specific oxidative enzymes. 1. Tétrazolium stains for diphosphpyridine nucleotide diaphorase.///wtocAcm Cytochem 4, 254-265.
Feldberg, W., Minz, B and Tsudzimura, H. (1934) The mechanism of the nervous discharge of adrenaline. / P/iyszo/ 81, 286-304.
Feuerstein, G. and Gutman, Y. (1971) Preferential secretion of adrenaline or noradrenaline by the cat adrenal in vivo in response to different stimuli. Br J Pharmacol 43, 764-775.
Fiallos-Estrada, C.E., Kummer, W., Mayer, B., Bravo, R., Zimmermann, M. and Herdegen, T. (1993) Long-lasting increase of nitric oxide synthase immunoreactivity. NADPH-diaphorase reaction and cJUN co-expression in rat dorsal root ganglion neurons follows sciatic nerve transection. Neurosci Lett 150, 169-173.
Figliomeni, B., Bacci, B., Panozzo, C., Fogarolo, F., Triban, C. and Fiori, M.G. (1992) Experimental diabetic neuropathy. Effect of ganglioside treatment on axonal transport of cytoskeletal proteins. Diabetes 41, 866-871.
Fischer-Colbrie, R., lacangelo, A. and Eiden, L.E. (1988) Neural and humoral factors separately regulate neuropeptide Y, enkephalin, and chromogranin A and B mRNA levels in rat adrenal medulla. Proc Natl Acad Sci USA 85, 3240-3244.
Fiscus, R.R., Robles, B.T., Waldman, S.A. and Murad, F. (1987) Atrial natriuretic factors stimulate accumulation and efflux of cyclic GMP in C6-2B, rat glioma and PC12 rat phaeochromocytoma cell cultures. J Neurochem 48, 522-528.
Fleischmann, W. (1965) Spontaneous endocrine disease in laboratory animals. In: The Pathology o f Laboratory Animals (eds. W.E. Ribelin and J R. McCoy), pp. 60-92. Charles C Thomas: Illinois.
Fleminger, G., Lahm, H.-W., and Udenfriend, S. (1984) Changes in rat adrenal catecholamines and proenkephalin metabolism after denervation. Proc Natl Acad Sci USA 81, 3587-3590.
Flint, J.M. (1900) The blood vessels, angiogenesis, organogenesis, reticulum, and histology of the adrenal. Johns Hopkins Hos Rep 9,153-229.
Folkow, B. and von Euler, U.S. (1954) Selective activation of noradrenaline producing cells in the cat's adrenal gland by hypothalamic stimulation. Circ Res 2,191-195.
210

Fossom, L.H., Carlson, G.D. and Tank, A.W. (1991a) Stimulation of tyrosine hydroxylase gene transcription rate by nicotine in rat adrenal medulla. Mol Pharmacol 40,193-202.
Fossom, L.H., Sterling, C. and Tank, A.W. (1991b) Activation of tyrosine hydroxylase by nicotine in rat adrenal g^and. J Neurochem 57, 2070-2077.
Forstermann, U., Gloss, E.I., Pollock, J.S., Nakane, M., Schwartz, P., Gath, I. and Kleinert, H. (1994) Nitric oxide synthase isozymes. Characterization, purification, molecular cloning, and function. Hypertension 23,1121-1131.
Forstermann, U., Schmidt, Pollock, J.S., Sheng, H., Mitchell, J.A., Warner,T.D., Nakane, M. and Murad, F. (1991) Isoforms of nitric oxide synthase: characterization and purification from different cell types. Biochem Pharmacol 42, 1849-1857.
Fraser, R. (1992) Biosynthesis of adrenocortical steroids. In: The Adrenal Gland, 2”^ edn. (ed. V.H.T. James), pp. 117-130. Raven Press: New York.
Furchgott, R.F. and Zawadzki, J.V. (1980) The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 288, 373-376.
Furness, J.B., Campbell, G.R., Gillard, S.M., Malmfors, T., Cobb, J.L.S. and Bumstock, G. (1970) Cellular studies of sympathetic denervation produced by 6 - hydroxydopamine in the vas defeiens. J Pharmacol Exp Ther 174,111-122.
Gabella, G. (1971) Neuron size and number in the myenteric plexus of the newborn and adult rat, J Anat 109, 81-95.
Gabella, G. and Trigg, P. (1984) Size of neurons and glial cells in the enteric ganglia of mice, guinea-pigs, rabbits and sheep. J Neurocytoi 13, 49-71.
Gamse, R., Holzer, P. and Lembeck, F. (1980) Decrease of substance P in primary afferent neurons and impairment of neurogenic plasma extravasation by capsaicin. B rJ Pharmacol 6 8 , 207-213.
Ganda, O.P., Rossini, A.A. and Like, A.A. (1976) Studies on streptozotocin diabetes. Diabetes 25, 595-603.
Garthwaite, J. (1991) Glutamate, nitric oxide and cell-cell signalling in the nervous system. Trends Neurosci 14, 60-67.
Garthwaite, J., Charles, S.L. and Chess-Williams, R. (1988) Endothelium-derived relaxing factor release on activation of NMDA receptors suggests role as intercellular messenger in the brain. Nature 336, 383-388.
Gaunt, R. (1975) History of the adrenal Cortex. InHandbook o f Physiology Section 7, Endocrinology, Vol. VI, Adrenal Gland (eds. H. Blaschko, G. Sayers and A.D. Smith), pp. 1-12. American Physiological Society: Washington DC.
Gauthier, P. and Reader, T.A. (1982) Adrenomedullary secretory response to midbrain stimulation in rat: effects of depletion of brain catecholamines or serotonin. Can J Physiol Pharmacol 60,1464-1474.
Gerich, I.E., Langlois, M., Noacco, C., Karam, J.H. and Forsham, P. (1973) Lack of
211

glucagon response to hypoglycaemia in diabetes: evidence for an intrinsic pancreatic alpha cell defect. Science 182,171-173.
Gersh, I. and Grollman, A. (1941) The vascular pattern of the adrenal gland of the mouse and rat and its physioloical response to changes in glandular activity. Contrib Embryol 29,113-125.
Gibson, S.J., Polak, J.M., Bloom, S.R., Sabate, I.M., Mulderry, P.M., Ghatei, M.A., McGregor, G.P., Morrison, J.F.B., Kelly, J.S., Evans, R.M. and Rosefeld, M.G. (1984) Calcitonin gene-related peptide immunoreactivity in the spinal cord of man and eight other sçecies. J Neurosci 4, 3101-3111.
Gilchrist, A.B., Leake, A. and Charlton, B.G. (1993) Innervation of the human adrenal cortex: simultaneous visualisation using acetylcholinesterase histochemistry and dopamine ̂ -hydroxilase immunohistochemistry. 146, 31-35.
Gillman, J., Gilbert, C. and Spence, I. (1953) Phaeochromocytoma in the rat. Pathogenesis and collateral reactions and its relation to comparable tumours in man. Cancer 6 , 494-511.
Goadsby, P.J. (1985) Brainstem activation of the adrenal medulla in the cat. Brain Res 327, 241-248.
Goedert, M., Mantyh, P.W., Hunt, S.P. and Emson, P.C. (1984) Localization of specific neurotensin binding sites in the rat adrenal gland. Brain Res 299, 389-392.
Goodall, M.C.C. and Kirshner, N. (1957) Biosynthesis of adrenaline and noradrenaline in vitro. J Chem 226, 213-221.
Gottschau, M. (1883) Structur und embryonale entwickelung der nebennieren bei saugethieren. Archiv fur Anatomie und Entwicklungsgeschichte Anatomische Abteilung. Quoted by Bertolet (1980).
Gorgas, K. and Bock, P. (1976) Morphology and histochemistry of the adrenal medulla. I. Various types of primary catecholamine-storing cells in the mouse adrenal medulla. Histochemistry 50,17-31.
Gower, D.B. (1975) Biosynthesis of the corticosteroids. In: Biochemistry o f Steroid Hormones (ed. H.L.J. Makin), pp. 47-75. Blakwell Scientific Publications: Oxford.
Grant, N.J., Konig, F., Deloume, J.-C., Aunis, D. and Langley, K. (1992) Noradrenergic, but not adrenergic chromaffin cells in the adrenal gland express neuromoduline (GAP-43). Eur J Neurosci 4,1257-1263.
Green, L., Tannebaum, S.R. and Goldman, P. (1981) Nitrate synthesis in germfree and conventional rat. Science 212, 56-58.
Greene, L.A. and Tischeler, A S. (1976) Establishment of a nonadrenergic clonal line of rat adrenal phaeochromocytoma cells which respond to nerve growth factor. Proc Natl Acad Sci USA 73, 2424-2428.
Greep, R.O. and Deane, H.W. (1949) Histological, cytochemical and physiological observations on the regeneration of the rat's adrenal gland following enucleation. Endocrinology 45, 42-56.
212

Grider, J.R. (1993) Interplay of VIP and nitric oxide in regulation of the descending relaxation phase of peristalsis. Am J Physiol 264, G334-G340.
Griscavage, J.M., Rogers, N.E., Schcrman, M.P. and Ignarro, L.J. (1993) Inducible nitric oxide synthase from a rat alveolar macrophage cell line is inhibited by nitric o x ié t.J Immunol 151, 6329-6337.
Grodsky, G.M., Anderson, C.E., Coleman, D.L., Craighead, J.E., Gerritsen, G.C., Hansen, C.T., Herberg, L., Howard, C.F., Jr., Lemmark, A., Matschinsky, P.M., Rayfield, E., Riley, W.J. and Rossini, A.A. (1982) Metabolic and underlying causes of diabetes mellitus. Diabetes 31 [Suppl 1], 45-53.
Gruetter, CA., Barry, B.K., McNamara, D.B., Gruetter, D.Y., Kadowitz, P.J. and Ignarro, L.J. (1979) Relaxation of bovine coronary artery and activation of coronary arterial guanylate cyclase by nitric oxide, nitroprusside and a carcinogenic nitrosoamine. J Cyclic Nucleotide Res 5, 211-224.
Grynspan-Winograd, O. (1974) Adrenaline and noradrenaline cells in the adrenal medulla of the hamster. A morphological study of their innervation. J Neurocytol 3, 341-361.
Guo, X. and Wakade, A.R. (1994) Differential secretion of catecholamines in response to peptidergic and cholinergic transmitters in rat adrenals. J Physiol 475, 539-545.
Haby, C., Lisovoski, P., Aunis, D. and Zwiller, J. (1994) Stimulation of cyclic GMP pathway by NO induces expression of the immediate early genes c-fos and junB in PC 12 czWs.JNeurochem 62, 496-501.
Hadjian, A.J., Guidicelli, C. and Chambaz, E.M. (1982) Cholinergic muscarinic stimulation of steroidogenesis in bovine adrenal cortex fasciculata cell suspensions. Biochem Biophys Acta 714,157-163.
Hadjian, A.J., Ventre, R. and Chambaz, E.M. (1981) Cholinergic muscarinic receptors in bovine adrenal cortex. Biochem Biophys Res Commun 98, 892-900.
Haeusler, G. (1974) Clonidine-induced inhibition of sympathetic nerve activity: no induction for a central presynaptic or an indirect sympathetic mode of action. Naunyn-Schmiedeberg's Arch Pharmacol 286, 97-111.
Haley, T.J. (1976) Control data in animal populations: a retrospective study of various rat strains. Proc West Pharmacol Soc 19, 23-27.
Hamaji, M., Miyata, M. and Kawashima, Y. (1985) A study of the vascular arrangement in the rat adrenal gland using non-radioactive microspheres. Cell Tissue Res 240, 277-280.
Hâppôlâ, O., Soinila, S., Paivarinta, H., Joh, T.H. and Panula, P. (1985) Histamine- immunoreactive endocrine cells in the adrenal medulla of the rat. Brain Res 339, 393-396.
Hartman, P.A. and Brownell, K.A. (1930) The hormone of the adrenal cortex. Science 72, 76.
Hartman, P.A., Brownell, K.A. and Liu, T.Y. (1955) Blood flow through the dog adrenal. Am J Physiol 180, 375-377.
213

Hassall, C.J., Saffrey, M.J., Belai, A., Hoyle, C.H.V., Moules, E.W., Moss, J., Schmidt,H.H.H.W., Murad, F., Forstermann, U. and Bumstock, G. (1992) Nitric oxide synthase immunoreactivity and NADPH-diaphorase activity in a subpopulation of intrinsic neurones of the guinea-pig heart. Neuroscience Lett 143, 65-68.
Hasse, P., Contestabile, A. and Flumerfelt, B.A. (1982) Preganglionic innervation of the adrenal gland of the rat using hoseradish peroxida^. Exp Neurol 78, 217-221.
Hawthorn, J., Nussey, S.S., Henderson, J.R. and Jenkins, J.S. (1987) Immunohistochemical localization of oxytocin and vasopressin in the guinea pig. Cell Tissue Res 250,1-6.
Haynes, R., Savard, K. and Dorfman, R.I. (1952) An action of ACTH on adrenal slices. Science 116, 690-691.
Hechter, O. (1949) Corticosteroid release from the isolated adrenal gland. Fed Proc 8, 70-71.
Hechter, O., Solomon, M.M., Zaffaroni, A. and Pincus, G. (1953) Transformation of cholesterol and acetate to adrenal cortical hormones. Arch Biochem Biophys 46, 2201-2214.
Hecker, M., Miilsch, A. and Busse, R. (1994) Subcellular localization and characterization of neuronal nitric oxide synthase. J Neurochem 62,1524-1529.
Henion, P.D. and Landis, S.C. (1990) Asynchronous appearance and topographic segregation of neuropeptide-containing cells in the developing rat adrenal medulla. J Neurosci 10, 2886-2896.
Henion, P.D. and Landis, S.C. (1992) Developmental regulation of leucine-enkephalin expression in adrenal chromaffin cells by glucocorticoids and innervation. J Neurosci 12, 3818-3827.
Henlé, J. (1865) Ueber das gewebe der nebenniere und der hypophyse. J Rationelle Med 24,143-152. Quoted by Carmichael (1983).
Henry, Y., Lepoivre, M., Drapier, J.-C., Ducrocqu, C., Boucher, J.-L. and Guissani, A. (1993) EPR characterization of molecular targets for NO in mammalian cells and organells. FASEB J 7,1124-1134.
Herdegen, T., Brecht, S., Mayer, B., Leah, J., Kummer, W., Bravo, R. and Zimmermann, M. (1993) Long-lasting expression of JUN and KROX transcription factors and nitric oxide synthase in intrinsic neurons of the rat brain following axotomy. J A^ewro^d 13, 4130-4145.
Hess, D.T., Patterson, S.L, Smith, D.S. and Skene, J.H. (1993) Neuronal growth cone collapse and inhibition of protein fatty acylation by nitric oxide. Nature 366, 562- 565.
Hevel, J.M., White, K.A. and Marietta, M.A. (1991) Purification of the inducible murine macrophage nitric oxide synthase. Identification as a flavoprotien. J Biol Chem 266, 22789-22791.
Heym, C. (1985) Neuropeptieds in paraganglia of various mammals. Fortschr Zool 30,
214

563-569.
Heym, C. and Kummer, W. (1988) Regulatory pcptieds in paraganglia. Prog Histochem Cytochem 18,1-95.
Heym, C., Colombo-Benckmann, M. and Mayer, B (1994) Immunohistochemical demonstration of the synthesis enzyme for nitric oxide and of comediators in neurons and chromaffin cells of the human adrenal medulla./1/m/I/iar 176,11-16.
Hibbs, J.B., Taintor, R.D. and Vavrin, Z. (1987a) Macrophage cytotoxicity: role for L- arginine deiminase and imino nitrogen oxidation to nitrate. Science 235, 473-476.
Hibbs, J.B., Vavrin, Z. and Taintor, R.R. (1987b) L-arginine is required for expression of the activated macrophage effecter mechanism causing selective metabolic inhibition in target cells. J Immunol 138,550-556.
Hillarp, N.A. and Hokfelt, B. (1953) Evidence of adrenaline and noradrenaline in separate adrenal medullary cells, Physiol Scand 30, 55-68.
Hilsted, J. (1982) Pathophysiology in diabetic autonomic neuropathy: cardiovascular, hormonal and metabolic studies. Diabetes 31, 730-737.
Hinson, J.P. (1990) Paracrine control of adrenocortical function: a new role for the meÔMWdi. J Endocrinol 124, 7-9.
Hinson, J.P. and Vinson, G.P. (1990) Calcitonin gene-related peptide stimulates adrenocortical function in the isolated perfused rat adrenal gland in situ. Neuropeptide 16,129-134.
Hinson, J.P., Cameron, L.A., Purbick, A. and Kapas, S. (1994a) The role ofneuropeptides in the regulation of adrenal vascular tone: effects of vasoactive intestinal polypeptide, substance P, neuropeptide Y, neurotensin, Met-enkephalin and Leu-enkephalin on perfusion medium flow rate in the intact perfused rat adrenal. Regul Pept 51, 55-61.
Hinson, J.P., Cameron, L.A., Purbick, A. and Kapas, S. (1994b) The role ofneuropeptides in the regulation of adrenal zona glomemlosa function: effects ofsubstance P, neuropeptide Y, neurotensin, Met-enkephalin, Leu-enkephalin and corticotrophin-releasing hormone on aldosterone secretion in the intact perfused rat dàitndX.J Endocrinol 140, 91-96.
Hinson, J.P., Kapas, S., Orford, C D. and Vinson, G.P. (1992) Vasoactive intestinal peptide stimulation of aldosterone secretion by the rat adrenal cortex may be mediated by the local release of c a t e c h o l a m i n e s . 133, 253-258.
Hinson, J.P., Purbrick, A., Cameron, L.A. and Kapas, S. (1994c) The role ofneuropeptides in the regulation of adrenal zona fasciculata/reticularis function: effects of vasoactive intestinal polypeptide, substance P, neuropeptide Y, met- enkephalin, leu-enkephalin neurotensin and corticosterone secretion in the intact perfused rat adrenal gland in situ. Neuropeptides 26, 391-397.
Hirose, T., Matsumoto, I. and Aikawa, T. (1978) Direct effect of histamine on cortisol and corticosterone production by isolated dog adrenal cell. J Endocrinol 76 371- 372.
215

Hirsch, D.B., Steiner, J.P., Dawson, T.M., Mammen, A., Hayek, E. and Snyder, S.H.(1993) Neurotransmitter release regulated by nitric oxide in PC-12 cells and brain synaptosomes. Curr Biol 3, 749-754.
Hoeldtke, R.D., Boden, G., Shuman, C.R. and Owen, O.E. (1982) Reduced epinephrine secretion and hypoglycemia unawareness in diabetic autonomic neuropathy. Ann Intern Med 96, 459-462.
Hofle, G., Weiler, R., Fisher-Colbrie, R., Humpel, C., Laslop, A., Wohlfarter, T., Hogue-Angeletti, R., Saria, A., Flemong, P.J. and Winkler, H. (1991) Stimulation of rat adrenal medulla can induce differential changes in the peptide and mRNA levels of chromogranins, neuropeptides and other constituents of chromaffin granules. Regul Pept 32,321-331.
Hokfelt, T., Lundberg, J.M., Schultzberg, M. and Fahrenkrug, J. (1981) Immunohistochemical evidence for a local VIP-ergic system in the adrenal gland of the rat. Physiol Scand 113, 575-576.
Holets, V. and Hide, R. (1982) The differential distribution and relationship of serotonergic and peptidergic fibers to sympathoadrenal nerves in the intermediolateral cell column of the rat: a combined retrograde axonal transport and immunofluorescence study. Neuroscience 7,1155-1174.
Holgert, H., Dagerlind, Â., Hokfelt, T. and Lagercrantz, H. (1994) Neuronal markers, peptides and enzymes in nerves and chromaffin cells in the rat adrenal medulla during postnatal development. Dev Brain Res 83, 35-52.
Hollander, G.F. and Snell, K.C. (1976) Tumors of the adrenal gland. lARC Sci Publ I (Part 2): 273-293.
Hollinshed, W.H. (1936) The innervation of the adrenal glands. J Comp Neurol 64, 449-467.
Holscher, C. and Ross, S.P.R. (1992) An inhibitor of nitric oxide synthesis prevent memory formation in the chick. Neurosci Lett 145,165-167.
Holst, J.J., Ehrhart-Bomstein, M., Messell, T., Poulsen, S.S. and Harling, H. (1991) Release of galanin from isolated perfused porcine adrenal glands: role of splanchnic nerves. Am J Physiol 261, E31-E40.
Holtz, P., Credener, K. and Kroneberg, G. (1947) Tiber das sympathicometische pressorische prinzip des ham (urosympathin). Naunyn Schmiedebergs Arch Exp Phth Pharmak 204, 228-243. Quoted by Coupland (1965).
Holzbauer, M. and Vogt, M. (1957) Functional changes produced in the adrenal cortex of the rat by administration or by release of corticotrophin. J Physiol 138, 449-459.
Holzwarth, M.A. (1984) The distribution of vasoactive intestinal peptide in the rat adrenal cortex and medulla. J Auton Nerv Syst 11, 269-283.
Holzwarth, M.A., Cunningham, L.A. and Kleitman, N. (1987) The role of adrenal nerves in the regulation of adrenaocortical functions. Ann N Y Acad Sci 512, 449- 464.
Holzwarth, M.A., Sawetawan, C. and Brownfield, M.S. (1984) Serotonin-
216

immunoreactivity in the adrenal medulla: distribution and response topharmacological manipulation. Brain Res Bull 13, 299-308.
Hope, B.T. and Vincent, S.R. (1989) Histochemical characterization of neuronal NADPH diaphorase.///z5toc/i/n Cytochem 37, 653-661.
Hope, B.T., Michael, G.J., Knigge, K.M. and Vincent, S.R. (1991) Neuronal NADPH diaphorase is a nitric oxide synthase. Proc Natl Acad Sci USA 88, 2811-2814.
Hoshi, T. (1926) Morphologisch experimentelle untersuchungen uber die innervation der nebeniere. MittAlg Path Sendai 3,328-342. Quoted by Coupland (1965).
Hughes, J., Smith, T.W., Kosterlitz, H.W., Forthergill, L.A., Morgan, B.A. and Morris, H R. (1975) Identification of two related pentapeptides from the brain with potent opiate activity. Nature 258, 577-579.
Hyano, M., Saba, N., Dorfman, R.I. and Hechter, O. (1956) Some aspects of the biogenesis of adrenal steroid hormones. Recent Prog Horm Res 12, 79-123.
lalenti. A., Iganaro, A., Moncada, S. and Di Rosa, S. (1992) Modulation of acute inflammation by endogenous nitric oxide. Eur J Pharmacol 211,177-182.
Ignarro L.J. (1990) Biosynthesis and metabolism of endothelium-derived nitric oxide. Annu Rev Pharmacol Toxicol 30, 535-560.
Ignarro, L.J., Buga, G.M., Wood, K.S., Byms, R.E. and Chudhuri, G. (1987) Endothelium-derived relaxing factor produced from artery and vein is nitric oxide. Proc Natl Acad Sci USA 84, 9265-9269.
Inagaki, S., Kubota, Y., Kito, S., Kangawa, K. and Matsuo, H. (1986) Immunoreactive atrial natriuretic polypeptides in the adrenal medulla and sympathetic ganglia. Reg Pept 15, 249-260.
Ingle, D.J. and Higgins, G.M. (1938) Regeneration of the adrenal gland following enucleation. /Im J Afed Sci 196, 232-239.
Ito, K., Sato, Y. and Suzuki, H. (1986) Increase in adrenal catecholamine secretion and sympathetic nerve unitary activities with aging in rats. Neurosci Lett 69, 263-268.
Itoh, N., Obata, K., Yanaihara, N. and Okamoto, H. (1983) Human prcprovasoactive intaestinal polypeptide contains a novel PHI-27-like peptide, PHM-27. Nature 304, 547-549.
Iyengar, R., Stuehr, DJ. and Marietta, M.A. (1987) Macrophage synthesis of nitrite, nitrate, and N-nitrosamines: precursors and role of the respiratory bust. Proc Natl Acad Sci USA 84, 6369-6373.
Jakobj, C. (1892) Beitrage zur physiologischcn und pharmakogischen kenntniss der darmbewegun gen mit besonderer berucksuhitigung der beziehung der nebenniere zu denslben. /IrcA Exp Path Pharmak 29,171-211. Quoted by Coupland (1965).
Jancso, G., Kiraly, E. and Jancsô-Gâbor, A. (1977) Pharmacologically induced selective degeneration of chemosensitive primary sensory neurons. Nature 270, 741-743.
217

Jancso, G., Hokfelt, T., Lundberg, J.M., Kiraly, E., Halasz, N., Nilsson, G., Terenius, L., Rehfeld, J., Steinbusch, H., Verhostad, A., Eide, R., Said, S. and Brown, M.(1981) Immunohistochemical studies on the effect of capsaicin on spinal and medullary peptide and monoamine neurons using antisera to substance P, gastrin CCK, somatostatin, VIP, enkephalin, neurotensin and 5-hydroxytryptamine. J Neurocytol 10, 963-980.
Jancso, N. (1968) Desensitization with capsaicine and related acylamides as a tool for studying the function of pain receptors. In: Pharmacology and Pain (ed. R.K.S. Lim), pp. 33-35. Pergamon Press: Oxford.
Jefcoate, C.R., DiBartolomeis, M.J., Williams, CA. and McNamara, B.C. (1987) ACTH regulation of cholesterol movement in isolated adrenal cells. J Steroid Biochem 27, 721-729.
Jensen-Holm, J. and Juul, R. (1970) The effects of guanethidine, pre- and postganglionic nerve division on the rat superior cervical ganglion: cholinesterase and catecholamines (histochemistry), and histology. Acta Pharmacol Toxicol 28, 283- 298.
Johnson, E. Jr. and O'Brien, F. (1976) Evaluation of the permanent sympathectomy produced by the administration of guanethidine to adult rats. J Pharmacol Exp Ther 196, 53-61.
Johnstone, F.R.C. (1957) The suprarenal veins, 94, 615-620.
Jones, C.T., Edwards, A.V. and Bloom, S.R. (1990) The effect of changes in adrenal blood flow on adrenal cortical responses to adrenocorticotrophin in conscious calves. J Physiol 429, 377-386.
Jonsson, G. and Sachs, C. (1972) Neurochemical properties of adrenergic nerves regenerated after 6-hydroxydopamine. JA^ewroc/iem 19, 2577-2585.
Junod, A., Lambert, A.E., Stauffacher, W. and Renold, A.E. (1969) Diabetogenic action of streptozotocin: relationship of dose to metabolic response. J Clin Invest 48,2129-2139.
Junod, A., Lambert, A.E., Orci, L., Pictet, R., Gonet, A.E. and Renold, A.E. (1967) Studies of the diabetogenic action of streptozotocin. Proc Soc Exp Biol Med 126, 201-205.
Kahn, R.H. (1911) Zuckerstich und nebennieren. F ^ g Arch Physiol 140, 209-255. Quoted by Coupland (1965).
Kataoka, Y., Fujimoto, M., Alho, H., Guidotti, A., Geffard, M., Kelly, G.D. and Hambauer, I. (1986) Intrinsic gamma aminobutric acid receptors modulate the release of catecholamine from canine adrenal gland in situ. J Pharmacol Exp Ther 239, 584-590.
Katsuki, S. and Murad, F. (1977) Regulation of adenosine cyclase 3',5'-monophospate and guanosine cyclase 3',5',-monophosphate levels and contractility in bovine tracheal smooth muscle. Mol Pharmacol 13, 330-341.
Katsuki, S., Arnold, W.P. and Murad, F. (1977) Effects of sodium nitroprusside, nitroglycerin, and sodium azide on levels of cyclic nucleotides and mechanical
218

activity of various tissues. J Cyclic Nucleotide Res 3, 239-247.
Keen, H. and Ng Tang Fui, S. (1982) The definition and classification of diabetes miilitus. In: Clinics in Endocrinology and Metabolism. New Aspects o f Diabetes, Vol. X I (eds. D.G. Johnston and K.G.M.M. Albesti), pp. 179-305. W.S. Saunders: London.
Kennedy, J.G., Breslow, M.J., Tobin, J.R. and Traystman, R.J. (1991) Cholinergic reulation of adrenal medullary blood flow. Am J Physiol 261, H1836-H1841.
Kent, C. and Coupland^ R E. (1989) Localisation of chromo^anin A and B, met- enkephalin-arg"-gly^-leu° and PGP 9.5-like immunoreactivity in the developing and adult rat adrenal medulla and extra-adrenal chromaffin tissue. J Anat 66, 213- 225.
Kesse, W.K., Parker, T.L. and Coupland, R.E. (1988) The innervation of the adrenal gland. I. The source of pre- and postganglionic nerve fibres to the rat adrenal gland. JAnat 157, 33-41.
Khalil, Z., Livett, B.G. and Marley, P.D. (1986) The role of sensory fibres in the rat splanchnic nerve in the regulation of adrenal medullary secretion during stress. J Physiol 370, 201-215.
Kilpatric, D.L., Howells, R.D., Fleminger, G. and Udenfriend, S. (1984) denervation of rat adrenal glands markedly increases pre-proenkephalin mRNA. Proc Natl Acad Sci USA 81, 7221-7223.
Kim, K.T. and Westhead, E. (1989) Cellular response to Ca"'*' firom extracellular and intracellular ^ources are different as shown by simultaneous measurements of cytosolic Ca '̂*' and secretion from bovine chromaffin cells. Proc Natl Acad Sci USA 86, 9881-9885.
Kirpekar, S.M., Prat, J.C. and Schiavone, M.T. (1982) Effect of muscarine on release of catecholamines from the perfused adrenal gland of the cat. Br J Pharmacol 77, 455-460.
Kirshner, N. (1975) Biosynthesis of the catecholamines. In: Handbook o f Physiology, Section 7, Endocrinology, vol. VI Adrenal Gland (eds. H. Blaschko, G. Sayers and A.D. Smith), pp. 341-355. American Physiological Society: Washington, D C.
Kiss, T. (1951) Experimentaell-morphologische analyse der nebennieren innervation. Acta Anat 13, 81-89.
Kleitman, N. and Holzwarth, M.A. (1985) Catecholaminergic innervation of the rat adrenal cortex. Cell Tissue Res 241,139-147.
Knight, D.E. and Baker, P.T. (1982) Calcium dependence of catecholamine release from bovine adrenal medullary cells after exposure to intense electric fields. J Metab Biol 68,107-140.
Knight, D.E. and Baker, P.T. (1986) Observations on the muscarinic activation of catecholamine secretion in the chick adrenal. Neuroscience 19, 357-366.
Knowles, R.G., Palcios, M., Palmer, R.M.J. and Moncada, S. (1989) Formation of nitric oxide from L-arginine in the central nervous system: a transduction
219

mechanism for stimulation of the soluble guanilate cyclase. Proc Natl Acad Sci USA 86, 5159-5162.
Kobayashi, S., Ohashi, T., Fujita, T., Nakao, K., Yoshimasa, T., Imura, H., Mochizuki, T., Yanaihara, C., Yanaihara, N. and Verhoftad, AA.J. (1983) ^immunohistochemical study on the costor^e and/or of met-enkephalin-Arg - Gly -Leu° and m et-enkephalin-Arg^-Phe'w ith adrenaline and/or noradrenaline in the adrenal chromaffin cells of rat, dog and cat. Blamed Res 4, 433-442.
Kobayashi, S., Ohashi, T., Uchida, T., Nakao, K., Imura, H., Yanaihara, N. and Verhoftad, A.A.J. (1984) C^storage of adrenaline and noradrenaline with met- enkephalin-Arg^-Gly'-Lcu° and met-enkephalin-Arg^-Phe' in chromaffin cells of hamster adrenal medulla. Blamed Res [Suppl] 4,151-158.
Kobzik, L., Bredt, D.S., Lowenstein, CJ., Drazen, J., Gaston, B., Sugarbaker, D. and Stamler, J.S. (1993) Nitric oxide synthase in human and rat lung: immunocytochemical and histochemical localization. Am J Resplr Cell Mai Blal 9, 371-377.
K&lle, G.B. (1950) The histochemical differentiation of types of cholinesterase and their localizations in tissues of the cat. J Pharmacol Exp Ther 100,158-179.
Kondo, H. (1985)) Immunohistochemical analysis of the localization of neuropeptides in the adrenal gland. Hlstal Jap 48, 453-481.
Kondo, H., Kuramoto, H., and Iwanaga, T. (1984) Immunohistochemical study on met-enkephalin-arg-gly-like immunoreactive nerve fibres in the rat adrenal medulla. Brain Res 310, 371-375.
Konishi, S., Tsunoo, A. and Otsuka, M. (1981) Enkephalin as a transmitter for presynaptic inhibition in sympathetic ganglia. Nature 294, 80-82.
Kowal, J., Horst, I., Pensky, J. and Alfonzo, M. (1977) A comparison of the effects of ACTH, VIP and cholera toxin on adrenal cAMP and steroid synthesis. Ann N Y Acad Sci 297, 314-328.
Krum, A.A., Morris, M.D. and Bennett, L.L. (1964) Role of cholesterol in the vivo biogenesis of adrenal steroids by the dog. Embryology 74, 543-547.
Kummer, W., Fisher, A., Mundel, P., Mayer, B., Hoba, B., Philippin, B. and Preissler, U. (1992) Nitric oxide synthase in VIP-containing vasodilator nerve fibres in the guinea-pig. NeuroReport 3, 653-655.
Kupfermann, I. (1991) Functional studies of cotransmission. Physiol Rev 71, 683-732.
Kuramoto, H., Kondo, H. and Fujita, T. (1985) Substance P-like immunoreactivity in adrenal chromaffin cells and intraadrenal nerve fibres of rats. Histochemistry 82, 507-512.
Kuramoto, H., Kondo, H. and Fujita, T. (1986) Neuropeptide tyrosine (NPY)-like immunoreactivity in adrenal chromaffin cells and intraadrenal nerve fibers of rats. Anat Rec 214, 321-328.
Kuramoto, H., Kondo, H. and Fujita, T. (1987) Calcitonin gene-related peptide (CGRP)-like immunoreactivity in scattered chromaffin cells and nerve fibres in the
220

adrenal gland of rats. Cell Tissue Res 247, 309-315.
Kvetnansky, R., Weise, V.K. and Kopin, LJ. (1970) Elevation of adrenal tyrosinehydroxylase and phenylethanolamine-N-methyltransferase by repeatedimmobilization of rats. Endocrinology 87, 744-749.
Kvetnansky, R., McCarty, R., Thoa, N.B., Lake, C.R. and Kopin, I.J. (1979)Sympatho-adrenal responses of spontaneously hypertensive rats to immobilazation stress. Am J Physiol 236, 4457-4462.
Kvetnansky, R., Sun, C., Lake, C.R., Thoa, N.B., Torda, T. and Kopin, I.J. (1978) Effect of handling and forced immobilization on rat plasma levels of epinephrine, norepinephrine and dopamine j8-hydroxylase. Endocrinology 103,1868-1874.
Kwon, N.S., Stueher, D.J. and Nathan, C.F. (1991) Inhibition of tumor cell ribonucleotide reductase by macrophage-derived nitric oxide. J Exp Med 174, 761- 768.
Lamas, S., Marsden, P.A., Li, G.K., Tempt, P. and Michel, T. (1992) Endothelial nitric oxide synthase: molecular cloning and characterization of a distinct constitutive enzyme isoform. Proc Natl Acad Sci USA 89, 6348-6352.
Landis, S.C. and Patterson, P.H. (1981) Neural crest cell lineage. Trends Neurosci 4, 172-175.
Lawson, S.W. (1987) The morphological consequences of neonatal treatment with capsaicin on primary afferent neurones in adult rats. Acta Physiol Hangarica 69, 315-321.
Ledeen, R.W. (1978) Ganglioside structures and distribution: are they localized at the nerve ending? J Supramol Strct 8 ,1-17.
Ledeen, R.W. and Yu, R.K. (1982) Gangliosides: structure, isolation, and analysis. Methods Enzymol 83,139-191.
Le Douarin, N.M. (1980) The ontogeny of the neural crest in avian embryo chimaeras. Nature 286, 663-669.
Lemaire, S., Chouinard, L., Mercier, P. and Day, R. (1986) Bombesin-like immunoreactivity in bovine adrenal medulla. Regul Pept 13,133-146.
Lemaire, S., Lemaire, I., Dean, D M. and Livett, B.G. (1980) Opiate receptors and adrenal medullary function. Nature 288, 303-304.
Lentle, B,C. and Thomas, J.P. (1964) Adrenal function and the complications of diabetes mellitus. Lancet 2, 544-549.
Lepoivre, M., Fieschi, F., Coves, J., Thelander, L. and Fontecave, M. (1991) Inactivation of ribonucleotide reductase by nitric oxide. Biochem Biophys Res Commun 179, 442-448.
Lesniewska, B., Nowak, M., Miskowak, B., Nussdorfer, G.G. and Malendowicz, L.K.(1990) Long-term effects of neuropeptide-Y on the adrenal cortex. Neuropeptides 16, 9-13.
221

Lever, J.D. (1952) Observation on the adrenal blood vessels in the rat. J Anat 86, 459- 467.
Lever, J.D. (1953) Nerve fibres in the adrenal cortex of the rat. Nature 171, 882-883.
Lewis, P.R. and Shute, C.C.D. (1969) An electron-microscopic study of cholinesterase distribution in the rat adrenal medulla. JAficro^c 89,181-193.
Lewis, R.V., Stem, AS., Kilpatrik, D.L., Gerber, L.D., Rossier, J., Stein, S. and Udenfriend, S. (1981) Marked increases in large enkephalin-containing polypeptides in the rat adrenal gland following denervation. J JVigwrojcf 1, 80-82.
Li, Z.-G., Queen, G. and LaBella, F.S. (1990) Adrenocorticotropin, vasoactive intestinal polypeptide, growth-hormone-releasing factor, and dynorphin compete for common receptors in brain and adrenal. Endocrinology 126,1327-1333.
Liang, B.T. and Perlman, R.L. (1979) catecholamine secretion by hamster adrenal ceWs. J Neurochem 32, 927-933.
Lilly, C M., Stamles, J.S., Gaston, B., Meckel, C., Loscalzo, J. and Drazen, J.M. (1993) Modulation of vasoactive intestinal peptide pulmonary relaxation by NO in tracheally superfused guinea pig lungs. Am J Physiol 265, L410-L415.
Lin, L.F., Bott, M.C., Kao, L.-S. and Westhead, E.W. (1995) ATP stimulated catecholamine secretion: response in perfused adrenal glands and a subpopulation of cultured chromaffin cells. Neurosci Lett 183,147-150.
Lincoln, J. and Messersmith, W.A. (1995) Conditions required for the measurement of nitric oxide synthase activity in a myenteric plexus / smooth muscle preparation from the rat ileum. J Neurosci Methods, in press
Lincoln, J., Messersmith, W.A., Belai, A. and Bumstock, G. (1993) The effects of streptozotocin-induced diabetes and ganglioside treatment on the activities of the enzymes involved in nitric oxide synthesis in the rat ileum myenteric plexus. Diahet Med 10 [Suppl 3], P71.
Lincoln, J., Bokor, J.T., Crowe, R., Griffith, S.G., Haven, A.J. and Bumstock, G. (1984) Myenteric plexus in streptozotocin-treated rats. Neurochemical and histochemical evidence for diabetic neuropathy in the gut. Gastroenterology 86, 654-661.
Linnoila, I., Diaugustine, R.P., Hervonen, A. and Miller, R. (1980) Distribution of (leu5) and (met5) enkephalin-, VIP- and SP-like immunopreactivity in human adrenal glands. Neuroscience 5, 2247-2259.
Livett, B.G. and Dean, M.D. (1980) Distribution of immunoreactive enkephalins in adrenal paraneurones: preferential localization in varicose processes and terminals. Neuropeptides 1, 3-13.
Livett, B.G., Day, R., Bide, R.P. and Howe, P.R.C. (1982) Co-storage of enkephalins and adrenaline in the bovine adrenal medulla. Neuroscience 7,1323-1332.
Livett, B.G., Kozousek, V., Mizobe, F. and Dean, D M. (1979) Substance P inhibits nicotinic activation of chromaffin cells. Nature 287, 256-257.
222

Livett, B.G., Marley, P.D., Wan, D.C.C. and Xin-Fu, Z. (1990) Peptide regulation of adrenal medullary function. JA^eura/ Transm [Suppl] 29, 77-89.
Livett, B.G., Boksa, P., Dean, D M., Mizobe, F. and Lindenbaum, M.H. (1983) Use of isolated chromaffin cells to study basic release mechanisms. J Auton Nerv Syst 7, 59-86.
Loesch, A., Belai, A. and Bumstock, G. (1993) Ultrastractural localization of NADPH-diaphorase and colocalization of nitric oxide synthase in endothelial cells of the rabbit aorta. Cell Tissue Res 274,539-545.
Loesch, A., Belai, A. and Bumstock, G. (1994) An ultrastractural study of NADPH- diaphorase and nitric oxide synthase in the perivascular nerves and vascular endothelium of the rat basilar artery. J Neurocytolol 23, 49-59.
Lowenstein, C.J., Dinerman, J.L. and Snyder, S.H. (1994) Nitric oxide - a physioloic messenger. Ann Intern Med 120, 227-237.
Lowenstein, C.J., Glatt, C.S., Bredt, D.S. and Snyder, S.H. (1992) Cloned and expressed macrophage nitric oxide synthase contrasts with the brain enzyme. Proc Natl Acad Sci USA 89, 6711-6715.
Lukas W. and Jones K. A. (1994) Cortical neurons containing calretinin are selectively resistant to calcium overload and excitotoxicity in vitro. Neuroscience 61, 307-316.
Lundberg, J.M., Saria, A., Franco-Cerceda, A. and Theodorsson-Northeim, E. (1985) Mechanisms underlying changes in the contents of neuropeptide Y in cardiovascular nerves and adrenal gland induced by sympatholytic drugs. Acta Physiol Scand 124, 603-611.
Lundberg, J.M., Rokaeus, A., Hokfelt, T., Rosell, S., Brown, M. and Goldstein, M.(1982) Neurtensin-like immunoreactivity in the preganglionic sympathetic nerves and in adrenal medulla of the cat. Acta Physiol Scand 114,153-155.
Lundberg, J.M., Hokfelt, T., Hensen, A., Theodorsson-Norheim, E., Pemow, J., Hamberger, B. and Goldstein, M., (1986) Neuropeptide Y-like immunoreactivity in adrenaline cells of adrenal medulla and in tumors and plasma of phaeochromocytoma patients. Regul Pept 13,169-182.
Lundberg, J.M., Hamberger, B., Schultzberg, M., Hokfelt, T., Granberg, P.O., Efendic, S., Terenius, L., Goldstein, M. and Luft, R. (1979) Enkephalin- and somatostatinlike immunoreactivities in human adrenal medulla and phaeochromocytoma. Proc Natl Acad Sci USA 76, 4079-4083.
Lutz-Bucher, B. and Koch, B. (1994) Evidence for inhibitory effect of nitric oxide synthase on neuropeptide secretion from isolated neural lobe of the pituitary gland. Neurosci Lett 165, 48-50.
Lykkelund, C. and Lund, E.D. (1979) Plasma glucagon responses to insulin-induced hypoglycemia and arginine in normal and alloxan diabetic rats. Scand J Clin Lab Invest 39,151-157.
MacFarland, W.E. and Davenport, H.A. (1941) Adrenal innervation. J Comp Neurol 75, 219-233.
223

Maher, T.D., Tanenberg, R.J., Greenberg, B.Z., Hoffman, J.E., Doe, R.P. and Goetz,F.C. (1977) Lack of glucagon response to hypoglycemia in diabetic autonomic neuropathy. Diabetes 26,196-200.
Majane, E.A., Alho, H., Kataoka, Y., Lee, C.H. and Yang, H.-Y. (1985) Neuropeptide Y in bovine adrenal glands: distribution and characterization. Endocrinology 117, 1162-1168.
Malendowicz, L.K., Lesniewska, B. and Miskowiak, B. (1990) Neuropeptide Y inhibits corticosterone secretion by isolated rat adrenocortical cells. Experientia 46, 721-722.
Malendowicz, L.K., Lesniewska, B., Baranowska, B., Nowak., M. and Majchrzak, M.(1991) Effect of bombesin on the structure and function of the rat adrenal cortex. Res Exp Med 191,121-128.
Malendowicz, L.K., Nussdorfer, G.G., Majchizak, M., Nowak, M. and Lesniewska, B.,(1992) Neurotensin stimulates the growth and secretion of rat adrenal zona glomemlosa. In Vivo 6, 523-526.
Malhotra, R.K. and Wakade, A.R. (1987a) Non-cholinergic component of rat splanchnic nerve predominates at low neuronal activity and is eliminated by nalaxone. J Physiol 383, 639-652.
Malhotra, R.K. and Wakade, A.R. (1987b) Vasoactive intestinal polypeptide stimulates the secretion of catecholamines from the rat adrenal gland. J Physiol 388, 285-294.
Malinski, T. and Taha, Z. (1992) Nitric oxide release from a single cell measured in situ by a porphyrinic-based microsensor. Nature 358, 676-678.
Malmejac, J. (1964) Activity of the adrenal medulla and its regulation. Physiol Rev 44, 186-218.
Marietta, M.A., Yoon, P.S., Iyengar, R., Leaf, CD. and Wishnok, J.S. (1988) Macrophage oxidation of L-arginine to nitrate and nitrite: nitric oxide is an mtcnncdiatc. Biochemistry 27, 8706-8711.
Marley, P.D., Thomson, K.A. and Smardencas, A. (1993) Non-cholinergic nervous control of catecholamine secretion from perfused bovine adrenal gland. J Physiol 465, 489-500.
Marsden, P.A., Heng, H.H.Q., Schere, S.W., Stewart, R.J., Hall, A.V., Shi, X.-M., Tssi, L.-C. and Schappert, K.T. (1993) Stmcture and chromosomal localization of the human constitutive endothelial nitric oxide synthase gene. J Biol Chem 268, 17478-17488.
Mason, H.L. (1964) Only the best is good enough. J Clin Endocrinol Metab 24, 1214- 1218.
Masuoko, D.T., Schott, H.F., Akawie, F. and Clark, W.G. (1956) Conversion of C - arterenol to epinephrine in vivo. Proc Soc Exp Biol Med 93, 5.
Matsui, H. (1979) Adrenal medullary secretory responses to stimulation of the medulla oblongata in the rat. Neuroendocrinology 29, 385-390.
224

Matsui, H. (1984) Adrenal medullary secretion in response to diencephalic stimulation in the rat. Neuroendocrinology 38,164-168.
Matsumoto, T., Pollock, J.S., Nakane, M. and Forstermann, U. (1993) Developmental changes of cytosolic and particulate nitric oxide synthase in rat brain. Dev Brain Res 73,199-203.
Maubert, E., Dupouy, J.P. and Bemet, F. (1993) Effect of adrenal demedullation on neuropeptide Y content of the capsule/^omerulosa zone of the rat adrenal gland. Neurosci Lett 156, 5-8.
Maubert, E., Tramu, G., Croix, D., Beauvillain, J.C. and Dupouy, J.P. (1990) Colocalization of vasoactive intestinal polypeptide and neuropeptide Y immunoreactivities in the nerve fibres of the rat adrenal gland. Neurosci Lett 113, 121-126.
Maycock, W.D.A. and Heslop, T.S. (1939) An experimental investigation of the nerve supply of the adrenal medulla of the cat. J Anat 73, 551-558.
Mayer, B., John, M., Heinzel, B., Werner, E.R., Wachter, H., Schultz, G. and Bohme, E. (1991) Brain nitric oxide synthase is a biopterin- and flavin- containing multifunctional oxidoreductase. FEES Lett 288,187-191.
Mazzocchi, G. and Nussdorfer, G.G. (1987) Neuropeptide-Y acutely stimulates rat zona glomerulosa in vivo. Neuropeptides 9, 257-262.
Mazzocchi, G., Malendowicz, L.K., Meneghelli, V. and Nussdorfer, G.G. (1992) Calcitonin gene-related peptide depresses the growth and secretory activity of rat adrenal zona glomerulsa. Neuropeptides 21,157-161.
Mazzocchi, G., Malendowicz, L.K., Rebuffat, P. and Nussdorfer, G.G. (1992) Effect of galanin on the secretory activity of the rat adrenal cortex: in vivo and in vitro studies. Res Exp Med Berl 192, 373-381.
Mazzocchi, G., Robba, C., Malendowicz, L.K. and Nussdorfer, G.G. (1987) Stimulatory effect of vasoactive intestinal peptide (VIP) on the growth and steroidogenic capacity of the rat adrenal zona glomerulosa. Biomed Res 8,19-23.
McCarty, R., Chiueh, C.C. and Kopin, I.J. (1978) Spontaneously hypertensive rats: adrenergic hyperresponsivity to anticipation of electric shock. Behev Biol 23, 180- 188.
Migally, N. (1979) The innervation of the mouse adrenal cortex. Anat Rec 194, 105- 112.
Mikhail, Y. (1961) Innervation of the different zones of the adrenal cortex. J Comp Neurol 117, 365-369.
Mikhail, Y. and Mahran, Z. (1965) Innervation of the cortical medullary portion of the adrenal gland of the rat during postnatal life. Anat Rec 152, 431-437.
Miki, N., Kawabe, Y. and Kuriyama, K. (1977) Activation of cerebral guanylate cyclase by nitric oxide. Biochem Biophys Res Commun 75, 851-856.
Mikolajiczk, H. (1965) Urinary output of 17-ks and microscopic changes in the
225

adrenal cortex of the rats treated with heparin, histamine or serotonin. Endocrinol Polska 347, 358.
Miller, R.J. (1988) Introduction to special issue: calcium signalling in neurons. Trends Neurosci. 11, 415-419.
Mione, M.C., Cavanagh, T.F.R., Kirkpatrick, KA. and Bumstock, G. (1992) Plasticity in expression of calcitonin gene-related peptide and substance P immunoreactivity in ganglia and fibres following guanethidine and/or capsaicin denervation. Cell Tissue Res 168, 491-504.
Mirkin, B.L. (1961) Factors influencing the selective secretion of adrenal medullary hormones. J Pharmacol Exp Ther 132, 218-225.
Mitchell, H.H., Schonle, H.A. and Grindly, H.S. (1916) The origin of the nitrates in the urine. J Biol Chem 24, 416-490.
Mizobe, F., Kozousek, V., Dean, D M. and Livett, B.G. (1979) Pharmacological characterization of adrenal paranurons: substance P and somatostatin as inhibitory modulators of the nicotinic release. Brain Res 178, 555-566.
Mohamed, A.A., Parker, T.L. and Coupland, R.E. (1988) The innervation of the adrenal gland. II. The source of spinal afferent nerve fibres to the guinea pig adrenal
Anat 160, 51-58.
Molinoff, P.B., Brimijoin, S., Weinshilboum, R. and Axelrod, J. (1970) Neuronally- mediated increase in dopamine ^-hydroxylase activity. Proc Natl Acad Sci USA 66, 453-458.
Moncada, S. and Higgs, A. (1993) The L-arginine-nitric oxide pathway. New E J Med 329, 2002-2012.
Moncada, S.H. and Palmer, R.M.J. (1991) Inhibition of the induction of nitric oxide synthase by glucocorticoids: yet another explanation for their anti-inflammatory effects? Trends Pharmacol Sci 12,130-131.
Moncada, S., Palmer, R.M.J. and Higgs, E.A. (1989) Biosynthesis of nitric oxide from L-arginine - a pathway for the regulation of cell function and communication. Biochem Pharmacol 38,1709-1715.
Moncada, S., Palmer, R.M.J. and Higgs, E.A. (1991) Nitric oxide: Physiology, pathophysiology and pharmacology. Pharmacol Rev 43,109-142.
Moore, B. (1895) On the chemical nature of a physiologically active substance occurring in the suprarenal J Physiol 17,14-17.
Mordes, J.P. and Rossini, A.A. (1985) Animal models of diabetes mellitus. In: Jouslin's Diabetes Mellitus, 12^" edn. (eds. A. Marble, L.P. Krall, R.F. Bradley, A.R. Christieb, J.S. Soeldner), pp. 110-137. Lea and Febiger: Philadelphia.
Morera, A.M., Cathiard, A.M., Lurthe, M. and Saez, J.M. (1979) Interaction of vasoactive intestinal peptide (VIP) with a mouse adrenal cell line (Y-1): specific binding and biological effects. Biochem Biophys Res Commun 90, 78-85.
Moro, M.A., Michelena, P., Sanchez-Garcia, P., Palmer, R., Moncada, S. and Garcia,
226

A.G. (1993) Activation of adrenal medullary L-arginine: nitric oxide pathway by stimuli which induce the release of catecholamines. Eur J Pharmacol 246, 213- 218.
Morris, S.M., Jr. and Billiar, T.R. (1994) New insights into the regulation of inducible nitric oxide synthesis. Am J Physiol 266, E829-E839.
Mueller, R.A., Thoenen, H. and Axelrod, J. (1969) Adrenal tyrosine hydroxylase: compensatory increase in activity after chemical sympathectomy. Science 163, 468- 469.
Mueller, R.A., Thoenen, H and Axelrod, J. (1970) Effect of pituitary and ACTH on the maintenance of basal tyrosine hydroxylase activity in the rat adrenal gland. Endocrinology 86, 751-755.
Markham, M., Hiromichi, S., Nakajima, S., Nakamoto, H., Kageyama, Y. and Saruta, T. (1989a) Calcitonin gene-related peptide is an inhibitor of aldostrone secretion. Endocrinology 125, 2227-2229.
Murakami, T., Oukouchi, H., Uno, Y., Ohtsuka, A. and Taguchi, T. (1989b) Blood vascular beds of rat adrenal and accessory adrenal glands, with special reference to the corticomedullary portal system: a further scanning electron microscopic study of corrosion casts and tissue specimens. Arch Histol Cytol 52, 461-476.
Nachlas, M.M., Walker, D.G. and Seligman, A.M. (1958) A histochemical method for the demonstration of diphosphopyridine nucleotide diaphorase. J Biophys Biochem Cytol 4, 29-37.
Nagy, J.I., Hunt, S.P., Iversen, L.L. and Emson, P. (1981) Biochemical and anatomical observations on the degeneration of peptide-containing primary afferent neurons after neonatal capsaicin. Neuroscience 6,1923-1934.
Nakane, M., Schmidt, H.H.H.W., Pollock, J.S., Forstermann, U. and Murad, F. (1993) Cloned human brain nitric oxide synthase is highly expressed in skeletal muscle. FEBS Lett 31 6 ,175-180.
Naranjo, J.R., Mocchetti, I., Schwartz, J.P. and Costa, E. (1986) Permissive effect of dexamethasone on the increase of proenkephalin mRNA induced by depolarization of chromaffin cells. Proc Natl Acad Sci USA 83,1513-1517.
Nathan, C. (1992) Nitric oxide as a secretory product of mammalian cells. FASEB J 6, 3051-3064.
Nathan, C.F. and Hibbs, J.B. (1991) Role of nitric oxide synthesis in macrophage antimicrobial activity. Cur Opin Immunol 3, 65-70.
Nathan, C. and Xie, Q.-W. (1994b) Regulation of biosynthesis of nitric oxide. J Biol Chem 269, 13725-13728.
Nathan, C. and Xie, Q.-W. (1994a) Nitric oxide synthase: roles, tolls, and controls. Cell 78, 915-918.
Natrajan, P.G., McGarrigle, H.H.G., Lawrence, D.M. and Lachelin, G.C.L. (1982) Plasma noradrenaline and adrenaline levels in normal pregnancy and in pregnancy- induced hypertension. B rJO bst Gynaecol 89,1041-1045.
227

Neuhber, W.L., Sandoz, P.A. and Fryscak, T. (1986) The central projections of primary afferent neurons of greater splanchnic and intercostal nerves in the rat. A horseradish peroxidase sXuùy. Anat Embryol 174,123-144.
Norido, P., Canella, R., Zanoni, R. and Gorio, A. (1984) Development of diabetic neuropathy in the C57BL/KS (db/db) mouse and its treatment with gangliosides. Exp Neurol 83, 221-232.
Nowicki, J.P., Duval, D., Poignet, H. and Scatton, B. (1991) Nitric oxide mediates neuronal death after focal cerebral ischemia in the mouse. Eur J Pharmacol 204, 339-340.
Nunokawa, Y., Ishida, N. and Tanaka, S. (1993) Cloning of inducible nitric oxide synthase in rat vascular smooth muscle. Biochem Biophys Res Commun 191, 89-94.
Nussdorfer, G.G. and Mazzocchi, G. (1987) Vasoactive intestinal peptide (VIP) stimulates aldosterone secretion by rat adrenal glands in vivo. J Steroid Biochem 26, 203-205.
Nussdorfer, G.G., Malendowicz, L.K., Belloni, A.S., Mazzocchi, G. and Rebuffat, P. (1988) Effects of substance P on the rat adrenal zona glomerulosa in vivo. Peptides 9,1145-1149.
O’Dell, T.J., Huang, P.L., Dawson, T.M., Dinerman, J.L., Snyder, S.H., Kandel, E.R. Fishman, M.C. (1994) Endothelial NOS and the blockade of LTP by NOS inhibitors in mice lacking neuronal NOS. Science 265, 542-546.
Okubo, M., Kaku, K., Kaneko, T. and Yanaihara, N. (1985) Effects of bombesin and gastrin releasing peptide on catecholamine secretion from rat adrenal gland in vitro. EndocrinolJpn 32, 21-27.
Oliver, G. and Schafer, F.A. (1894) On the physiological action of extract of the suprarenal capsules. 7 P/iy5i<9/16,1-4.
Oomori, Y., Okuno, S., Fujisawa, H. and Ono, K. (1991) Immunoelectron microscopic study of tyrosine hydroxylase immunoreactive nerve fibres and ganglion cells in the rat adrenal Anat Rec 229, 407-414.
Oomori, Y., luchi, H., Nakaya, K., Tanaka, H., Ishikawa, K., Satoh, Y. and Ono, K.(1993) Gamma-aminobutric acid (GABA) immunoreactivity in the mouse adrenal gland. Histochemistry 100, 203-213.
Oomori, Y., Okuno, S., Fujisawa, H., luchi, H., Ishikawa, K., Satoh, Y. and Ono, K.(1994) Ganglion cells immunoreactive for catecholamine-synthesising enzymes, neuropeptide Y and vasoactive intestinal polypeptide in the rat adrenal gland. Cell Tissue Res 275, 201-213.
Oomori, Y., Okuno, S., Fujisawa, H., Ishikawa, K., Satoh, Y., Matsuda, M., Yamano, M. and Ono, K. (1989) Tyrosine-hydroxylase-immunoreactive nerve fibres in the separated capsule of the rat adrenal gland. Acta Anat 136, 49-54.
Oppenheim, R.W. (1991) Cell death during development of the nervous system. Annu Rev Neurosci 14, 453-501.
228

Oset-Gasque, M.J., Parramôn, M., Hortclano, S., Boscâ, L. and Gonzalez, M.P. (1994) Nitric oxide implication in the control of neurosecretion by chromaffin cells. J Neurochem 63,1693-1700.
O'Sullivan, A.J. and Bourgoyne, R.D. (1990) Cyclic GMP regulates nicotine-induced secretion from cultured bovine adrenal chromaffin cells: effects of 8-bromo-cyclic GMP, atrial natriuretic peptide, and nitroprusside (nitric oxide). J Neurochem 54, 1805-1808.
Palacios, G. and Lafarga, M. (1975) Chromaffin cells in the glomerulosa zone of adult rat adrenal cortex. Cell Tissue Res 164, 275-278.
Palacios, M., Knowles, R.G., Palmer, R.M.J. and Moncada, S. (1989) Nitric oxide from L-arginine stimulates soluble guanilate cyclase in adrenal gland. Biochem Biophys Res Comms 165, 802-809.
Palmer, R., Ferrige, A.G. and Moncada, S. (1987) Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature 327, 524-526.
Panula, P. (1986) Histochemistry and function of bombesin-like peptides. Med Biol 64,177-192.
Panza, J.A., Quyyami, A.A., Brush, I.E., Jr. and Epstein, S.E. (1990) Abnormal endothelium-dependent vascular relaxation in patients with essential hypertension. NEngl J Med 323, 22-27.
Park, S.K., Lin, H.L. and Morphy, S. (1994) Nitric oxide limits transcriptional induction of nitric oxide synthase in CNS glial cells. Biochem Biophys Res Commun 201, 762-768.
Parker, T.L., Afework, M. and Coupland, R.E. (1990a) Sensory innervation of the rat adrenal gland. Neurosci Lett 38, S62.
Parker, T.L., Mohamed, A.A. and Coupland, R.E. (1990b) The innervation of the adrenal gland. IV. The source of pre- and postganglionic nerve fibres to the guinea-pig adrenal gland. 172,17-24.
Parker, T.L., Kesse, W.K., Mohamed, A.A. and Afework, M. (1993) The innervation of the mammalian adrenal gland. J Anat 183, 265-276.
Parker, T.L., Kesse, W.K., Tomlinson, A. and Coupland, R.E. (1988) Ontogenesis of preganglionic sympathetic innervation of the rat adrenal chromaffin cells. In: Progress in Catecholamine Research, Part A. Basic Aspects and Peripheraal Mechanisms (eds. A. Dhalstrom, R.H. Belmaker and M. Sandler), pp. 227-232. Alan R Liss: New York.
Patel, D.G. (1983) Lack of glucagon response to hypoglycemia in long-term experimental diabetic rats. Diabetes 32, 55-60.
Pearse, A G E. (1972) Histochemistry. Theoretical and Applied. Vol. II, 3̂ *̂ edn., pp. 761-1518. Churchill Livingstone: Edinburgh.
Pelto-Huikko, M. (1989) Immunocytochemical localization of neuropeptides in the adrenal medulla. J Electron Microsc Tech 12, 364-379.
229

Pelto-Huikko, M. and Saiminen, T. (1987) Localization of calcitonin gene-related peptide (CGRP) in chromaffin cells and nerve terminals in adrenal medulla. Exp Brain Res Series 16, 73-77.
Pelto-Huikko, M., Saiminen, T., Partnen, M., Toivanen, M. and Hervonen, A. (1985) Immunohistochemicai localization of neurotensin in hamster adrenal medulla. The Anatomical Record 211, 458-464.
Pender, N. and Burgoyne, R.D. (1992) Histamine stimulates exocytosis in a subpopulation of bovine adrenal chromaffin cells. Neurosci Lett 144, 207-210.
Peunova, N. and Enikolopov, G. (1993) Amplification of calcium-induced gene transcription by nitric oxide in neuronal cells. Nature 364,450-453.
Pemow, J., Thorén, P., Millberg, B.-I. and Lundberg, J.M. (1988) Renal sympathetic nerve activation in relation to neuropeptide Y in the kidney of the rat. Acta Physiol Scand 134, 53-59.
Persson, K., Igawa, Y., Mattiasson, A. and Andersson, K.-E. (1992) Effects of inhibition of the L-arginine/nitric oxide pathway in the rat lower urinary tract in vivo and in vitro. B rJ Pharmacol 107,178-184.
Pfiffner, J.J. (1942) The adrenal cortical hormones. Advan Enzymol 2, 325-356.
Piezzi, R.S. (1966) Two types of synapses in the chromaffin tissue of the toad's adrenal. /Iera Physiol Latinoam 16, 282-285.
Pines, L. and Narowtschatowa, K. (1931) Uber die innervation der nebennieren. Z Micr Anat Forsch 25, 518-538. Quoted by Coupland (1965).
Pohorecky, L A. and Wurtman, R.J. (1971) Adrenocortcal control of epinephrine synthesis. Pharmacol Rev 23,1-35.
Polak, J.M. and Van Noorden, S. (1987) An introduction to immunocytochemistry: current techniques and problems. Microscopy Handbooks, Revised edn. Oxford University Press: Oxford.
Pollock, J.S., Forstermann, U., Mitchell, J.A., Warner, T.D., Schmidt, H.H.H.W., Nakane, M. and Murad, F. (1991) Purification and characterization of particulate endothelium-derived relaxing factor synthase from cultured and native bovine aortic endothelial cells. Proc Natl Acad Sci USA 88,10480-10484.
Pollock, J.S., Nakane, M., Buttery, L.D., Martinez, A., Springall, D., Polak, J.M., Forstermann, U., and Murad, F. (1993) Characterization and localization of endothelial nitric oxide synthase using specific monoclonal antibodies. Am J Physiol 265, C1379-C1387.
Porter, J.C. and Klaiber, M.S. (1965) Corticosterone secretion in rats as a function of ACTH input and adrenal blood Üov/.J Physiol 209, 811-814.
Pralong, F.P., Corder, R. and Gaillard, R.C. (1991) Responses of the rat pituitary- adrenal axis to hypotensive infusions of corticotropin-releasing factor, vasoactive intestinal peptide and other depressor agents. Regul Pept 32, 217-226.
Pratt, J.H., Turner, D.A., Bowsher, R.R. and Henry, D P. (1987) Dopamine in rat
230

adrenal glomerulosa. Life Sci 40, 811-816.
Prentice, F.D. and Wood, J.G. (1975) Adrenrgic innervation of cat adrenal medulla. Anat Rec 181, 689-704.
Properzi, G., Francavilla, S., Poccia, G., Aloisi, P., Gu, X., Terenghi, G. and Polak, J.M. (1993) Early increase precedes a depletion of VIP and PGP-9.5 in the skin of insulin-dependent diabetics - correlation between quantitative immunohistochemistry and clinical assessment of peripheral neuropathy. J Pathol 169, 269-277.
Purwar, R.S. (1978) Neuro-histochemical study of the adrenal gland of the Rattus rattus rufescens (Indian black rat) as revealed by cholinesterase technique. Acta Anat 102, 29-32.
Quinn, S.J. and Williams, G.H. (1988) Regulation of aldosterone secretion. Annu Rev Physiol 50, 409-426.
Quinn, S.J. and Williams, G.H. (1992) Regulation of aldosterone secretion. In: The Adrenal Gland, 2^^ edn. (ed. V.H.T. James), pp. 159-189. Raven Press: New York.
Radomski, M.W. and Moncada, S. (1991) Biological role of nitric oxide in platelet function. In: Clinical Relevance o f Nitric Oxide in the Cardiovascular System (eds. S. Moncada, E.A. Higgs and J R. Berrazueta), pp. 45-56. EDICOMPLET: Madrid.
Rakieten, N., Rakieten, M.L. and Nadkami, M.V. (1963) Studies on the diabetogenic action of streptozotocin (NSC-37917). Cancer Chemother Rep 29, 91-98.
Rand, M.J. (1992) Nitrergic transmission: nitric oxide as a mediator of non-adrenergic, non-cholinergic neuro-effector transmission. Clin Exp Pharmacol Physiol 19, 147-169.
Rapoport, R.M. and Murad, F. (1983) Agonist-induced endothelium-dependent relaxation in rat thoracic aorta may be mediated through cGMP. Circ Res 52, 352- 357.
Rebuffat, P., Robba, C., Mazzocchi, G. and Nussdorfer, G.G. (1984) Inhibitory effect of somatostatin on the growth and steroidogenic capacity of rat adrenal zona g^omcuilosa. J Steroid Biochem 21, 387-390.
Rebuffat, P., Malendowicz, L.K., Belloni, A S., Mazzocchi, G. and Nussdorfer, G.G. (1988a) Long-term stimulatory effect of neuropeptide-Y on the growth and steroidogenic capacity of rat adrenal zona glomerulosa. Neuropeptides 11,133-136.
Rebuffat, P., Belloni, A S., Malendowicz, L.K., Mazzocchi, G., Gottardo, G. and Nussdorfer, G.G. (1988b) Zona glomerulosa morphology and function in streptozotocin-induced diabetic rats. Endocrinology 123, 949-955.
Rebuffat, P., Belloni, A S., Malendowicz, L.K., Mazzocchi, G., Meneghelli, U. and Nussdorfer, G.G. (1988c) Effect of streptozotocin-induced experimental diabetes on the morphology and function of the zona fasciculata of rat adrenal cortex. Virchows Archiv B Cell Pathol 1,13-19.
Rebuffat, P., Belloni, A S., Rocco, S., Andreis, P.G., Neri, G., Malendowicz, L.K., Gottardo, G., Mazzocchi, G. and Nussdorfer, G.G. (1992) The effect of aging on the
231

morphology and function of the zonae fasciculata and reticularis of the rat adrenal cortex. Cell Tissue Res 270, 265-272.
Reinecke, M. (1985) Neurotensin. Immunohistochemicai localization in central and peripheral nervous system and in endocrine cells and its functional role as neurotransmitter and endocrine hormone. Prog Histochem Cytochem 16,1-175.
Ren, K., Ruda, M.A. and Jacobowitz, D.M. (1993) Immunohistochemicai localization of calretinin in the dorsal root ganglia and spinal cord of the rat. Brain Res Bull 31, 13-22.
Rerup, C.C. (1970) Drugs producing diabetes through damage of the insulin secrting cells. Pharmacol Rev 22, 485-518^.
Résibois, A. and Rogers, J.H. (1992) Calretinin in rat brain: an immunohistochemicai study. Neuroscience 46,101-134.
Robinson, P.M., Perry, R.A., Hardy, K.J., Coghlan, J.P. and Scoggins, B.A. (1977) The innervation of the adrenal cortex in the sheep, Ovis ovis.J Anat 124,117-129.
Robinson, R.L., Culberson, J.L. and Carmichael, S.W. (1983) Influence of hypothalamic stimulation on the secretion of adrenal medullary catecholamines. J Auton Nerv Syst 8, 89-96.
Rodden, P., Wiegandt, H. and Bauer, B. (1991) Gangliosides: the relevance of current research to neurosurgery. 74, 606-619.
Rogers, J.H. (1987) Calretinin: a gene for a novel calcium-binding protein expressed principally in neurons. J Cell Biol 105,1343-1353.
Rogers, J.H. (1989) Immunoreactivity for calretinin and other calcium-binding proteins in cerebellum. Neuroscience 31, 711-721.
Rogers, J.H. (1991) Calretinin: In: Novel Calcium-Binding Proteins. Fundamentals and Clinical Implications (ed. C.W. Heizmann), pp. 251-276. Springer-Verlag: London.
Rogers, N.E. and Ignarro, L.J. (1992) Constitutive nitric oxide synthase from cerebellum is reversibly inhibited by nitric oxide formed from L-arginine. Biochem Biophys Res Commun 189, 242-249.
Role, L.W. and Perlman, R.L. (1983) Both nicotinic and muscarinic receptors mediate catecholamine secretion by isolated guinea-pig chromaffin cells. Neuroscience 10, 979-985.
Role, L.W., Leeman, S.E. and Perlman, R.L. (1981) Somatostatin and substance P inhibit catecholamine secretion from isolated cells of guinea-pig adrenal medulla. Neuroscience 6,1813-1821.
Rolleston, H.D. (1936) The Endocrine Organs in Health and Disease. Oxford University Press: London.
Rosenfeld, G. (1955) Stimulative effect of acetylcholine on the adrenocortical function of isolated perfused calf adicmls. Am J Physiol 183, 272-278.
232

Rosenfeld, M.G., Mermod, J.J., Amara, S.G., Swanson, L.W., Sawchenko, J.R., Rivier, J., Vale, W.W. and Evans, R.M. (1983) Production of a novel neuropeptied encoded by the calcitonin gene via tissue-specific RNA processing. Nature 304,129-135.
Rosenthal, HE., Slaunwhite, W.R., Jr., and Sandberg, A A . (1969) Transcortin: a corticosteroid binding protein of the plasma. X. Cortisol and progesterone interplay and the unbound levels of these steroids in pregnancy. J Clin Endocrinol Metab 29, 352-367.
Roskams, A.J., Bredt, D., Dawson, T.M. and Ronnett, G.V. (1994) Nitric oxide mediates the formation of synaptic connections in developing and regenerating olfactory receptor neurons. Neuron 13, 289-299.
Ross, L.L., Smolen, A. and Cherry, J. (1980) Supraspinal transection interferes with the normal development of sympathetic preganglionic neurons. Soc Neurosci Abst 6, 385.
Rubin, R.F. and Miele, E. (1968) A study of the differential secretion of epinephrine and norepinephrine from the perfused cat adrenal gland. J Pharmacol Exp Ther 164, 115-121.
Rundle, S.E., Benedict, J.C., Robinson, P.M. and Funder, J.W. (1988) Innervation of the sheep adrenal cortex: an immunohistochemicai study with rat corticotropin- releasing factor antiserum. Neuroendocrinology 48, 8-15.
Ryder, R.E.J., Owens, D R., Hayes, T.M., Ghatei, M.A. and Bloom, S.R. (1990) Unawareness of hypoglycaemia and in adequate hypoglycaemia counterregulation: no causal relation with diabetic autonomic neuropathy. Br Med J 301, 783-787.
Saffran, M., Grad, B. and Bayliss, M.J. (1952)) Production of corticosteroids by rat adrenals in vitro. Endocrinology 50, 639-643.
Saffrey, M.J., Hassall, C.J.S., Hoyle, C.H.V., Belai, A., Moss, J., Schmidt, H.H.H.W., Forstermann, U., Murad, F. and Bumstock, G. (1992) Colocalization of nitric oxide synthase and NADPH-diaphorase in cultured myenteric neurones. NeuroReport 3, 333-336.
Saiani, L. and Guidotti, A. (1982) Opiate receptor-mediated inhibition of catecholamine release in primary cultures of bovine adrenal chromaffin cells. J Neurochem 39,1669-1676.
Said, S.I. and Mutt, V. (1970) Polypeptide with broad biological activity: isolation from small intestine. Science 169,1217-1218.
Samuels, L.T. and Nelson, D.H. (1975) Biosynthesis of corticosteroids. In: Handbook o f Physiology, Section 7, Endocrinology, Vol. VI, Adrenal Gland (eds. H. Blaschko,G. Sayers and A.D. Smith), pp. 41-53. American Physiological Society: Washington DC.
Sandor, T., Fazekas, A.G. and Robinson, B.H. (1976) The biosynthesis of corticosteroids throughout the vertebrates. In: General, Comparative and Clinical Endocrinology o f the Adrenal Cortex, Vol. I (eds. I. Chester Jones and I.W. Henderson), pp. 25-142. Academic Press: London.
Sarason, E.L. (1943) Morphologic changes in the rats's adrenal cortex under various
233

experimental conditions. Arch Pathol 35, 373-390.
Sato, A., Sato, H. and Suzuki, H. (1987) Changes in sympatho-adrenal medullary functions during aging. In: Organization o f the Autonomic Nervous System: Central and Peripheral Mechanisms (eds. J Ciriello, F.R. Calareso, L.P. Renaud and C. Polosa), pp. 27-36. Alan R Liss: New York.
Sayers, G. (1950) The adrenal cortex and homeostasis. Physiol Rev 30, 241-320.
Schalling, M., Franco-Cereceda, A., Hemsen, A., Dagerlind, Â., Seroogy, K., Persson, H., Hokfelt, T. and Lundberg, J. M. (1991) NPY and catecholamines in rat sympathetic neurons and adrenal gland: studies on expression, synthesis and axonal transport after pharmacological and experimental manipulations using hybridization techniques and radioimmunoassay. Neuroscience 41, 753-766.
Schalling, M., Dagerlind, Â., Berne, S., Hallman, H., Djurfeldt, M., Persson, H., Terenius, L., Goldstein, M., Schlesinger, D. and Hokfelt, T.(1988a) Coexistence and gene expression of phenylethanolamine N-methyletransferase, tyrosine hydroxylase and neuropeptide tyrosine in rat and bovine adrenal gland: effect of reserpine. Proc Natl Acad Sci USA 85, 8306-8310.
Schalling, M., Seroogy, K., Hokfelt, T., Chai, S.Y., Hallman, H., Persson, H., Larhammar, D., Ericsson, A., Terenius, L., Graffi, J., Massoulié, J. and Goldstein, M. (1988b) Neuropeptide tyrosine in the rat adrenal ^and - immunohistochemicai and in situ hybridization study. Neuroscience 24, 337-349.
Schanne, F.A.X., Kane, A.B., Young, B.E. and Farber, J.L. (1979) Calcium dependence of toxic cell death: a final common pathway. Science 206, 700-702.
Schini, V.B. and Vanhoutte, P.M. (1992) Inhibitors of calmodulin impair the constitutive but not the inducible nitric oxide synthase activity in the rat aorta. J Pharmacol Exp Ther 261, 553-559.
Schmidt, H.H.H.W., Pollock, J.S., Nakane, M., Gorsky, L.D., Forstermann, U. and Murad, F. (1991) Purification of a soluble isoform of guanylyl cyclase-activating- factor synthase. Proc Natl Acad Sci USA 88, 365-369.
Schneider, AS., Cline, H.T. and Lemaire, S. (1979) Rapid rise in cyclic GMP accompanies catecholamine secretion in suspensions of isolated adrenal chromaffin cells. Li/e Science 24,1389-1394.
Schramm, L.P., Adair, J R., Stribling, J.M. and Grey, L.P. (1975) Preganglionic innervation of the adrenal gland of the rat: a study using horsradish peroxidase. Exp Neurol 49, 504-553.
Schultzberg, M., Lundberg, J.M., Hokfelt, T., Temius, L., Brandt, J., Elde, R. and Goldstein, M. (1978) Enkephalin-like immunoreactivity in gland cells and nerve terminals of the adrenal medulla. Neuroscience 3,1169-1186.
Schuman, E.M. and Madison, D.V. (1994) Nitric oxide and synaptic function. Ann Rev Neurosci 17,153-183.
Schuster, D. (1974) Adrenocorticotrophic hormone and the control of corticosteroidogencsis.^dv Steroid Biochem Pharmacol 4, 233-294.
234

Seidl, K. and Unsicker, K. (1989) The determination of the adrenal medullary cell fate during embryogenesis. Dev Biol 136, 481-490.
Sessa, W.C. (1994) The nitric oxide synthase family of proteins. J Vase Res 31, 131- 143.
Sessa, W.C., Pritchard, K., Seyedi, N., Wang, J. and Hintze, T.H. (1994) Chronic exercise in dogs increases coronary vascular nitric oxide production and endothelial cell nitric oxide synthase gene expression. Circ Res 74, 349-353.
Sessa, W.C., Harrison, J.K., Barber, C.M., Zeng, D., Durieux, M.E., D'Angelo, D.D., Lynch, K.R. and Peach, M.J. (1 ^ 2 ) Molecular cloning and expression of a cDNA encoding endothelial cell nitric oxide synthase. J Bio/ Chem 267,15274-15276.
Siegel, R.E., Eiden, L.E. and Pruss, R.M. (1985) Multiple populations of neuropeptide-containing cells in cultures of the bovine adrenal medulla. Dev Brain Res 17, 267-270.
Sietzen, M., Schober, M., Fischer-Colbriem, R., Scherman, D., Sperk, G. and Winkler,H. (1987) Rat adrenal medulla: levels of chromogranins, enkephalins, dopamine ^ - hydroxylase and of the amine transporter are changed by nervous activity and hypophysectomy. Neuroscience 22,131-139.
Simpson, E.R. and Waterman, M R. (1988) Regulation of the synthesis of steroidogenic enzymes in adrenal cortical cells by ACTH. Annu Rev Physiol 50, 422-440.
Simpson, E.R. and Waterman, M R. (1 ^ 2 ) Regulation of expression of adrenocortical enzymes. \n:The Adrenal Glands, 2^^ edn. (ed. V.H.T. James), pp. 191-207. Raven Press: New York.
Siou, P.G.S., Belai, A. and Bumstock, G. (1992) Age-related changes in galanin- and calretinin-immunoreactive nerves of guinea-pig gallbladder. NeuroReport 3, 990- 992.
Slotkin, T.A. (1986) Development of the sympathoadrenal axis. In: Developmental Neurobiology o f the Autonomic Nervous System (ed. P.M. Gootman), pp. 69-96. Humana Press: Clifton, New Jersey.
Slotkin, T.A., Chantry, C.J. and Bartolomé, J. (1982) Development of central control of adrenal catecholamine biosynthesis and release. Biosci 36, 95-102.
Slotkin, T.A., Smith, P.G., Lau, C. and Bareis, D.L. (1980) Functional aspects of development of catecholamine biosynthesis and release in the sympathetic nervous system. In: Biogenic Amines in Development (eds. H. Parvez and S. Parvez), pp. 29-48. Elsevier: North Holand, Amsterdam.
Smith, J.A., Shah, A.M. and Lewis, M.J. (1991) Factors released from endocardium of the ferret and pig modulate myocardial contraction. 7 P/iy5zo/ 439,1-14.
Smith, J.A., Shah, A.M., Fort, S.I and Lewis, M.J. (1992) The influence of endocardial endothelium on myocardial contraction. Trends Pharmacol Sci 13,113-116.
Soediono, P., Belai, A. and Bumstock, G. (1993) Prevention of neuropathy in the pyloric sphincter of streptozotocin-diabetic rats by gangliosides. Gastroenterology
235

104,1072-1082.
Sparrow, R.A. and Coupland, R.E. (1987) Blood flow to the adrenal gland of the rat: its distribution between the cortex and the medulla before and after haemorrhage. J Anat 155, 51-61.
Stachwiak, M.K., Fluharty, S.J., Strieker, E.M., Zigmond, M.J. and Kaplan, B.B. (1986) Molecular adaptations in catecholamine biosynthesis induced by cold stress and sympathectomy. J Neurosci Res 16,13-24.
Stamler, J.S. (1994) Redox signalling: nitrosylation and related target interaction of nitric oxide. Cell 78, 931-936.
Stamler, J.S., Singel, D.J. and Loscalzo, J. (1992) Biochemistry of nitric oxide and its redox-activated forms. Science 258,1898-1902.
Stevens, J (1990) Intermediate Statistics: A Modern Approach. Lawrence Erlbaum: London.
Stevens, P., Robinson, R.L., Van Dike, K. and Stizel, R. (1975) Synthesis, storage and drug induced release of ATP-8--^H in the perfused bovine adrenal gland. Pharmacology 13, 40-55.
Stoddard-Apter, S.L., Siegel, A. and Levin, B.E. (1983) Plasma Catecholamine and cardiovascular responses following hypothalamic stimulation in the awake cat. J Auton Nerv Syst 8, 343-360.
Strack, A.M., Sawyer, W.B., Marubio, L.M. and Loewy, A.D. (1988) Spinal origin of sympathetic preganglionic neurons in the rat. Brain Res 455,187-191.
Strack, A.M., Sawyer, W.B., Platt, K.B. and Loewy, A.D. (1989) CNS cell groups regulating the sympathetic outflow to adrenal gland as revealed by transneuronal cell body labelling with pseudorabies virus. Brain Res 491, 274-296.
Stuehr, D.J. and Griffith, O.W. (1992) Mammalian nitric oxide synthases. Adv Enzymol 65, 287-346.
Stuehr, D.J. and Marietta, M.A. (1985) Mammalian nitrate biosynthesis: mouse macrophages produce nitrite and nitrate in response to Escherichia coli lipopolysaccharide. Proc Natl Acad Sci USA 82, 7738-7742.
Steuher, D.J., Cho, H.J., Know, N.S., Weise, M.F. and Nathan, C.F. (1991) Purification and characterization of the cytokine-induced macrophage nitric oxide synthase: an FAD- and FMN-containing flavoprotein. Proc Natl Acad Sci USA 88, 7773-7777.
Stults, C.L.M., Sweeley, C.C. and Macher, B. (1989) Glycosphingolipids: structure, biological source, and properties. Methods Enzymol 179,167-214.
Sweat, M.L. and Lipscomb, M.D. (1955) A transhydrogenase and reduced triphosphopyridinenucleotide involved in the oxidation of desoxycorticosteme to corticosterone by adrenal t i s s u e . Chem Soc 77, 5185-5187.
Swingle, W.W. and Pfiffner, J.J. (1930) An aqueous extract of the suprarenal cortex which maintains the life of bilaterally adrenalectomized cats. Science 71, 321-322.
236

Swinyard, C.A. (1937) The innervation of the suprarenal gland. Anat Rec 68, 417-429.
Symington, T. (1969) Functional Pathology o f the Suprarenal Gland. E. and S. Livingston: London.
Takamine, J. (1901) Adrenaline, the active principle of the suprarenal glands and its mode of picparation. Am J Pharmacol 73, 523-531.
Tannenbaum, S.R., Fett, D., Young, V.R., Land, P.D. and Bruce, W.R. (1978) Nitrite and nitrate are formed by endogenous synthesis in the human intestine. Science 200, 1487-1489.
Tatemoto, K., Carlquist, M. and Mutt, V. (1982) Neuropeptide Y - a novel brain peptide with structural similarities to peptide YY and pancreatic polypeptide. Nature 296, 659-660.
Tatemoto, K., Rokacus, Â., Jomvall, H., McDonald, T.J. and Mutt, V. (1983) Galanin - a novel biologically active peptide from porcine intestine. FEES Lett 164, 124- 128.
Tay, S.S.W., Moules, E.W. and Bumstock, G. (1994) Colocalization of NADPH- diaphorase with nitric oxide synthase and vasoactive intestinal polypeptide in newborn pancriatic neurons. J Anat 184, 545-552.
Terenghi, G., Polak, J.M., Vamdell, I.M., Lee, Y.C., Wharton, J. Bloom, S.R. (1983) Neurotensin-like immunoreactivity in a subpopulation of noradrenaline-containing cells of the cat adrenal gland. Endocrinology 12, 226-233.
Thoenen, H. (1975) Transynaptic regulation of neuronal enzyme synthesis. In: Handbook o f Psychopharmacology, vol. I ll (eds. L.L. Iversen, S.D. Iversen and S. Snyder), pp. 443-475. Plenum Press: New York.
Thoenen, H. and Tranzer, J.P. (1968) Chemical sympathectomy by selective destruction of adrenergic nerve endings with 6-hydroxy dopamine. Naunyn- Schmiedebergs Arch Pharmakol 261, 271-288.
Thoenen, H., Mueller, R.A. and Axelrod, J. (1969) Trans-synaptic induction of adrenal tyrosine hydroxylase. J P/iarmaco/ Exp Ther 169, 249-254.
Thomas, P.K. (1991) Diabetic neuropathy: epidemiology and pathgenesis. In: Peripheral Neuropathy Vol. II (eds. J. Pickup and G. Williams), pp. 613-622. Blackwell Scientific Publications: Oxford.
Thompson, R.J., Doran, J.F., Jackson, P., Dhillon, A.P. and Rode, J. (1983) PGP 9.5 - a new marker for vertebrate neurons and neuroendocrine cells. Brain Res 278, 224- 228.
Tischler, A S. and DeLellis, R.A. (1988) The rat adrenal medulla. I. The normal adrenal. 7 Coll Toxicol 7 ,1-21.
Tischler, A S., DeLellis, R.A., Perlman, R.L., Allen, J.M., Costopoulos, D., Lee, Y.C., Nunnemacher, G., Wolfe, H.J. and Bloom, S.R. (1985) Spontaneous proliferative lesions of the adrenal medulla in aging Long-evans rats. Lab Invest 51, 486-498.
Tomimoto, H., Nishimura, M., Suenaga, T., Nakamura, S., Akiguchi, I., Wakita, H.,
237

Kimura, J. and Mayer, B. (1994) Distribution of nitric oxide synthase in the human cerebral blood vessels and brain tissues. J Cereb Blood Flow Metab 14, 930-938.
Tomlinson, A. and Coupland, R.E. (1990) The innervation of the adrenal gland. IV. Innervation of the rat adrenal medulla from birth to old age. A descriptive and quantitative morphometric and biochemical study of the innervation of chromaffin cells and adrenal medullary neurons in Wistar rats. J Anat 169, 209-236.
Tomlinson, A., Durbin, J. and Coupland, R.E. (1987) A quantitative analysis of rat adrenal chromaffin tissue: morphometric analysis at tissue and cellular level correlated with catecholamine content. Neuroscience 20, 895-904.
Torda, T., Cruciani, R.A. and Saavedra, J.M. (1988) Localization of neuropeptide Y binding sites in the zona glomerulosa of the bovine adrenal gland. Neuroendocrinology 48, 207-210.
Tomello, S., Coirini, H. and De Nicola, A.F. (1981) Effects of experimental diabetes on the concentration of corticosterone in central nervous system, serum and adrenal gland. J Steroid Biochm 14,1279-1284.
Torres, M., Ceballos, G. and Rubio, R. (1994) Possible role of nitric oxide in catecholamine secretion by chromaffin cells in the presence and absence of cultured endothelial ctWs.J Neurochem 63, 988-996.
Tracey, W.R., Nakane, M., Pollock, J.S. and Forstermann, U. (1993) Nitric oxide synthase in neuronal cells, macrophages and endothelium are NADPH diaphorase, but represent only a firaction of total cellular NADPH diaphorase activity. Biochem Biophys Res Commun 195,1035-1040.
Tranzer, J.P. and Thoeonen, H. (1968) An electron microscopic study of selective, acute degeneration of sympathetic nerve terminals after administration of 6 - hydroxydopamine. Experientia 24,155-156.
Triban, C., Guidolin, D., Fabris, M., Marini, P., Schiauinato, A., Dona, M., Bortolami, M.C., Giamberardino, L.D. and Fiori, M.G. (1989) Ganglioside treatment and improved axonal regeneration capacity in experimental diabetic neuropathy. Diabetes 38,1012-1022.
Triggle, C.R. (1994) Is NO the answer? (or NO news is good news). Pharmaceutical News 1, 9-14.
Tsigos, C., Young, R.J. and White, A. (1993) Diabetic neuropathy is associated with increased activity of the hypothalamic-pituitary-adrenal axis. J Clin Endocrinol Metab 76, 554-558.
Tsou, K-C., Cheng, C-S, Nachlas, M.M. and Seligman, A.M. (1956) Synthesis of some /?-nitrophenyl substituted tétrazolium salts as electron acceptors dehydrogenases. Chem Soc 78, 6139-6144.
Tuominen, R.K., Karhunen, T., Panula, P. and Yamatodani, A. (1993) Endogenous histamine in cultured bovine adrenal chromaffin cells. Eur J Neurosci 5, 1436- 1441.
Uchiyama, Y., Morita, K., Kitayama, S., Suemitsu, T., Minami, N., Miyasako, T. and Dohi, T. (1994) Possible involvement of nitric oxide in acetylcholin-induced
238

increase of intracellular Ca^" ̂ concentration and catecholamine release in bovine adrenal chromaffin cells. Jpn J Pharmacol 65, 73-77.
Undenfriend, S. and Wyngaarden, J.B. (1956) Precursors of adrenal epinephrine and norepinephrine in vivo. Biochem Biophys Acta 20, 48-52.
Unger, A. and Phillips, J.H. (1983) Regulation of the adrenal medulla. Physiol Rev 63, 787-843.
Uno, H. (1977) Catecholaminergic terminals in the pcrisinusoidal spaces of the hepatic acini and adrenal cortex of macaques, Rec 187, 735.
Unsicker, K. (1969) Zur innervation der nebennierenride vom goldhamster. Z Zellforsch 95, 608-619.
Unsicker, K. (1971) On the innervation of the rat and pig adrenal cortex. Z Zellforsch 116,151-156.
Unsicker, K. (1973a) Fine structure and innervation of the avian adrenal gland. I. Fine structure of adrenal chromaffin cells and ganglion cells. Z Zellforsch 145, 389-416.
Unsicker, K. (1973b) Fine structure and innervation of the avian adrenal gland. II. Cholinergic innervation of adrenal chromaffin cells. Z Zellforsch 145, 417-442.
Unsicker, K. (1973c) Fine structure and innervation of the avian adrenal gland. III. Non-cholinergic fibres. Z Zellforsch 145, 557-575.
Unsicker, K., Kabura-Flüh, O. and Zwarg, U. (1978) Different types of small granule- containing cells and neurons in the guinea pig adrenal medulla. Cell Tissue Res 189, 109-130.
Urquhart, J. (1965) Adrenal blood flow and the adrenocortical response to corticotropin, J 209,1162-1168.
Vamdell, I.M., Polak, J.M., Allen, J.M., Terengehi, G. and Bloom, S.R. (1984) Neuropeptide tyrosine (NPY) immunoreactivity in norepinephrine containing cells and nerves of the mammalian adrenal gland. Endocrinology 114,1460-1462.
Vamdell, I.M., Topia, F.J., Demey, J., Rush, R.A., Bloom, S.R. and Polak, J.M. (1982) Electron immunocytochemical localization of enkephalin-like material in catecholamine-containing cells of the carotid body, the adrenal medulla and phaeochromocytomas of man and other mammals. J Histochem Cytochem 30 682- 690.
Verge, V.M.K., Xu, Z., Xu, X.-J., Wiesenfeld-Hallin, Z. and Hokfelt, T. (1992) Marked increase in nitric oxide synthase mRNA in rat dorsal root ganglia after peripheral axotomy: in situ hybridization and functional studies. Proc Natl Acad Sci USA 89,11617-11621.
Verhofstad, A.A.J. and Jonsson, G. (1983) Immunohistochemicai and neurochemical evidence for the presence of serotonin in the adrenal medulla of rat. Neuroscience 10,1443-1453.
Verhofstad, A.A.J., Coupland, R.E., Parker, T.R. and Goldstein, M. (1985) Immunohistochemicai and biochemical study on the development of the
239

noradrenaline- and adrenalin-storing ceils of the adrenal medulla of the rat. Cell Tissue Res 242, 233-243.
Verma, A., Hirsch, D.J., Glatt, C.E., Ronnett, G.V. and Snyder, S.H. (1993) Carbon monoxide: a putative neural messenger. Science 259, 381-384.
Vinson, G.P. and Hinson, J.P. (199^31ood flow and hormone secretion in the adrenal gland. In: The Adrenal Gland, ^ edn. (ed. V.H.T. James), pp. 71-86. New York: Raven Press.
Vinson, G.P., Hinson, J.P. and Tôth, I.E. (1994) The neuroendocrinology of the adrenal coitcx. J Neuroendocrinol 6, 235-246.
Vizi, E.S., Toth, I.E., Ors6, E., Szaiay, Sz., Szabé, D., Baranyi, M. and Vinson, G.P.(1993) Dopamine is taken up from the circulation by, and released from, local noradrenergic varicose axon terminals in zona glomerulosa of the rat: a neurochemical and immunocytochemical study. J Endocrinol 139, 213-226.
Vizi, E.S., Tôth, I.E., Szaiay, K. S., Windisch, K., Ors6, E., Szabô, D. and Vinson, G.P.(1992) Catecholamines released from local adrenergic axon terminals are possibly involved in fine tuning of steroid secretion from zona glomerulosa cells: functional and morphological e v i d e n c e . 135, 551-561.
Vodovotz, Y., Kwon, N.S., Pospischil, M., Manning, J., Paik, J. and Nathan, C. (1994) Inactivation of nitric oxide synthase after prolonged incubation of mouse macrophages with IF N -r and bacterial lipopolysaccharide. J Immunol 152, 4110- 4118.
Vogt, M. (1952) The secretion of the denervated medulla of the cat. Br J Pharmacol 7, 325-330.
von Euler, U S and Gaddum, J.H. (1931) An unidentified depressor substance in certain tissue extidiCXs. J Physiol 72, 74-87.
von Euler, U.S and Hamberg, U. (1949) L-noradrenaline in the suprarenal medulla. Nature 163, 642-643.
Wagner, D A. and Tannenbaum, S.R. (1982) Enhancement of nitrate biosynthesis by Echerichia coli lipopolysaccharide. In: Nitrosamines and Human Cancer (ed. P.N. Magee), pp. 437-441. Cold Spring Harbor Laboratory: USA.
W a^er, D.A., Young, V.R. and Tannenbaum, S.R. (1983) Mammalian nitrate biosynthesis: incorporation of ^^NHo into nitrate is enhanced by nitrate treatment. Proc Natl Acad Sci USA 80, 4518-4521.
Wakade, A.R. (1988) Noncholinergic transmitter(s) maintains secretion of catecholamines from rat adrenal medulla for several hours of continuous stimulation of splanchnic neuions. J Neurochem 50,1302-1308.
Wakade, A.R. (1991) Contribution of acetylcholine and vasoactive intestinal polypeptide in controlling catecholamine secretion. J Auton Nerv Syst 50, 1302- 1308.
Wakade, A.R. and Wakade, T.D. (1983) Contribution of nicotinic and muscarinic receptors in the secretion of catecholamines evoked by endogenous and exogenous
240

acetylcholine. Neuroscience 10, 973-978.
Wakade, A.T., Guo, X., Strong, R., Arimura, A. and Haycock, J. (1992) Pituitary adenylate cyclase-activating polypeptide (PACAP) as a neurotransmitter in rat adrenal medulla. Regul Pept 37, 331.
Wakade, T.D., Blank, M.A., Malhotra, R.K., Pourcho, R. and Wakade, A.R. (1991) The peptide VIP is a neurotransmitter in rat adrenal medulla: physiological role in controlling catecholamine secretion. J Physiol 444,349-362.
Watanabe, I., Hiramtsu, K., Ohmori, Y. and Paik, Y.K. (1990) Histo- and cytochemical studies on the distribution of acetylcholinesterase-positive nerve fibres in the goat adrenal gland. Anat Histol Embryol 19, 245-254.
Watanabe, M., Meeker, C.I., Gray, M.J., Sims, E.A.H. and Solomon, S. (1968) Secretion rate of aldosterone in normal pregnancy. J Clin Invest 42,1619-1631.
Weber, E., Evans, C.J. and Barchas, J.D. (1983) Multiple endogenous ligands for opioid receptors. Trends Neurosci 6, 333-336.
Weiner, C.P., Lizasoain, I., Baylis, S.A., Knowles, R.G., Charles, I.G. Moncada, S.(1994) Induction of calcium-dependent nitric oxide syntheses by sex hormones. Proc Natl Acad Sci USA 91, 5212-5216.
Weinshilboum, R. and Axelrod, J. (1970) Dopamine ^-hydroxylase activity in the rat after hypophysectomy. Endocrinology 87, 894-899.
Werbin, H. and Chaikoff, I.L. (1961) Utilization of adrenal gland cholesterol for synthesis of cortisol by the intact normal and the ACTH-treated guinea pig. Arch Biochem Biophys 93, 476-482.
Wemer-Felmayer, G., Werner, E.R., Fuchs, D., Hausen, A., Reibnegger, G., Schmidt, K., Weiss, G. and Wachter, H. (1993) Pteridine biosynthesis in human endothelial cells. Impact on nitric oxide-mediated formation of cyclic GMP. J Biol Chem 268, 1842-1846.
Wesselingh, S.L., Li, Y.W. and Blessing, W.W. (1989) PNMT-containing neurones in the rostral medulla oblongata (Cl, C3 groups) are transneuronally labelled afterinjection of herps simplex virus type 1 into the adrenal gland. Neurosci Lett 106,99-104.
White, J.W. Jr. (1975) Relative significance of dietary sources of nitrate and nitrite. J Agric Food Chem 23, 886-891.
Wiegandt, H. (1971) Glycosphingolipids. Adv Lipid Res 9, 249-289.
Wilke, R.A., Riley, D.A., Lelkes, P.I. and Hillard, C.J. (1993) Decreasedcatecholamine secretion from the adrenal medullae of chronically diabetic BB- Wistar rats. Diabetes 42, 862-868.
Wilkinson, K.D., Lee, K., Deshpande, S., Duerksen-Hughes, P., Boss, J.M. and Pohl, J. (1989) The neuron-specific protein PGP 9.5 is a ubiquitin carboxy-terminal hydrolyse. Science 246, 670-672.
Willard, D.M. (1936) The innervation of the adrenal glands of mammals: a
241

contribution to the study of nerve endings. Q J Microscop 78, 475-486.
Wilson, P.O.G., Barber, P.C., Hamid, QA., Power, B.F., Dhillon, A.P., Rode, J., Day,I.N.M., Thompson, R.J. and Polak, J.M. (1988) The immunolocalization of protein gene product 9.5 using rabbit polyclonal and mouse monoclonal antibodies. Br J Exp Pathol 69, 91-104.
Wilson, S.P., Abou-Donia, M.M., Chang, K.-J. and Viveros, O.H. (1981) Reserpine increases opiate-like peptide content and tyrosine hydroxylase activity in adrenal medullary chromaffin cells in culture. Neuroscience 6,71-79.
Winkler, H. (1976) The composition of adrenal chromaffin granules: an assessment of controversial results. Neuroscience 1, 65-80.
Winkler, H. and Westhead, E. (1980) The molecular organization of adrenal chromaffin granules. Neuroscience 5,1803-1823.
Winkler, H., Apps, D.K. and Fischer-Colbrie, R. (1986) The molecular function of adrenal chromaffin granules: established facts and unresolved topics. Neuroscience 18, 261-290.
Winsky, L., Nakata, H., Martin, B.M. and Jacobowitz, D.M. (1989) Isolation, partial amino acid sequence, and immunohistochemicai localization of a brain-specific calcium-binding protein. Proc Natl Acad Sci USA 86,10139-10143.
Wu, W. (1993) Expression of nitric oxide synthase (NOS) in injured CNS neurons as shown by NADPH diaphorase histochemistry. Exp Neurol 120,153-159.
Wu, W. and Li, L. (1993) Inhibition of nitric oxide synthase reduces motoneuron death due to spinal root avulsion. Neurosci Lett 153,121-124.
Wurtman, R.J. and Axelrod, J. (1966) Control of enzymatic synthesis of adrenaline in the adrenal medulla by adrenal cortical steroid. / B/o/ Chem 241, 2301-2305.
Wurtman, R.J., Pohorecky, L.A. and Baliga, B.S. (1972) Adrenocortical control of the biosynthesis of epinephrine and proteins in the adrenal medulla. Pharmacol Rev 24, 411-426.
Yamamoto, R., Bredt, D.S., Snyder, S.H. and Stone, R.A. (1993) The localization of nitric oxide synthase in the rat eye and related cranial ganglia. Neurosci 54, 189- 200.
Yobum, B.C., Franklin, S.O., Calvano, S.E. and Inturrisi, C.E. (1987) Regulation of rat adrenal medullary enkephalins by glucocorticoids. Life Sci 40, 2495-2503.
Yoshizumi, M., Perrella, M.A., Burnett, J.C. and Lee, M. (1993) Tumor necrosis factor downregulated an endothelial nitric oxide synthase mRNA by shortening its half- life. Circ Res 73, 205-209.
Young, J.Z. (1939) Partial degeneration of the nerve supply of the adrenal. A study in autonomic i n n e r v a t i o n . 73, 504-550.
Xie, Q-w., Cho, H.J., Calaycay, J., Mumford, R.A., Swiderek, K.M., Lee, T.D., Ding, A., Troso, T. and Nathan, C. (1992) Cloning and characterization of inducible nitric oxide synthase from mouse macrophages. Science 256, 225-228.
242

Xu, W., Charles, LG., Moncada, S., Goman, P., Sheer, D., Liu, L. and Emson, P.(1994) Mapping of the genes encoding human inducible and endothelial nitric oxide synthase (NOS2 and NOS3) to the pericentric region of the chromosome 17 and to chromosome 7 respectively. Genomics 21,419-422.
Zajicek, G., Ariel, I. and Arber, N. (1986) The streaming adrenal cortex: direct evidence of centripetal migration of adrenocytes by estimation of cell turnover rate. J Endocrinol 111, 477-482.
Zhou, X.-F. and Livett, B.G. (1990) Substance P increases catecholamine secretion from perfused rat adrenal glands evoked by prolonged field stimulation. J Physiol 425, 321-334.
Zhou, X.-F., Oldfield, B.J. and Livett, B.G. (1991) Substance P-containing sensory neurons in the rat dorsal root ganglia innervate the adrenal medulla. J Auton Nerv Syst 33, 247-254.
Zigmond, R.E. (1985) Biochemical consequences of synaptic stimulation: the regulation of tyrosine hydroxylase activity by multiple transmitters. Trends Neurosci 8, 63-69.
243




















