
Ultrastructure of the Erythrocytic Stages of Plasmodium malariae1
8
J Pruru:oul., 34(3). 1987. pp. 267-274 6 1987 by the Society of Protoroologists Ultrastructure of the Erythrocytic Stages of Plasmodium malariael CARTER T. ATKINSON,* MASAMICHI AIKAWA,* EDWIN P. ROCK,** * KEVIN MARSH,*** PATRICIA M. ANDRYSIAK,**** GARY H. CAMPBELL,**** WILLIAM E. COLLINS,**** and RUSSELL J. HOWARD***** *Institute of Pathology, Case Western Reserve University, Cleveland, Ohio 44106, USA. **Howard Hughes Medical Institute, Bethesda, Maryland 20814, USA, ***Medical Research Council Laboratories, Fajara, The Gambia, ****Malaria Branch, Center for Infectious Diseases, Centers for Disease Control, Atlanta, Georgia 30333, USA, and *****Laboratory of Parasitic Diseases, National lnstitute of Allergy and Infectious Diseases, National Institutes of Health. Bethesda, Maryland 20205, USA ABSTRACT. This report describes the fine structure of the erythrocytic stages of Plasmodium malariae. Erythrocytic parasites from a naturally acquired human infection and an experimentally infected chimpanzee were morphologically indistinguishable and structurally similar to other primate malarias. New findings included observations of highly structured arrays of merozoite surface coat proteins in the cytoplasm of early schizonts and on the surface of budding merozoites and the presence of knobs in the membranes of Maurer's clefts. Morphological evidence is presented suggesting that proteins are transported between the erythrocyte surface and intracellular parasites via two routes: one associated with Maurer's clefts for transport of membrane-associated knob material and a second associated with caveolae in the host cell membrane for the import or export of host- or parasite-derived substances through the erythrocyte cytoplasm. F the four species of Plasmodium that cause human ma- 0 laria, P. malariae (Grassi & Feletti, 1890) is one of the most unusual in terms of its patchy geographic distribution and ability to persist at low intensities in infected individuals for long periods (1 0). In spite of its importance as a cause of ne- phropathy in endemic areas (1 5), P. malariae has not been thoroughly studied. This is largely because long-term in vitro culture of this species has not been successful and because sus- ceptible primate models rarely develop parasitemias in excess of 2-5% of the circulating erythrocytes. Consequently, only one brief study ofthe fine structure ofthis species has been published Discontinuous Percoll gradients with (6) or without (7) sor- bitol have been used recently for purifying P. falciparum-in- fected erythrocytes from infected humans and P. ovale-infected erythrocytes from the whole blood of Aotus monkeys and chim- panzees (7). These methods were used in this study, as well as a Plasmage13concentration technique (1 8), to purify P. malar- iae-infected erythrocytes from whole human and chimpanzee blood in order to allow observation of numerous parasitized cells by electron microscopy. In this report we describe new ultrastructural observations of both P. malariae and parasite- induced morphological changes of host erythrocytes. (20). MATERIALS AND METHODS An 18-year-old Gambian man attended the Medical Research Council outpatient clinic with a two-day history of fever, head- ache, and general body pain. On examination, he was afebrile ' We are grateful to the staff and patients of the Medical Research Council Laboratories, Fajara, for assistance in sample collection and for the excellent technical assistance of Ana Milosavljevic and Kiet Dan Luc. Yoshitsugu Matsumoto and John Rabbege provided helpful com- ments on the manuscript. This work was supported in part by grants from the U.S. Agency for International Development (DPE-0453-A- 00-4027), USPHS (AI- 10645), the U.S. Army Research and Develop- ment Command (DAMDI7-79C-9029),and the UNDP/World BanW WHO Special Programme for Research and Training in Tropical Dis- eases. This is contribution No. 1805 to the U.S. Army Drug Devel- opment Program. Edwin Rock is a research scholar at the Howard Hughes Medical Institute. Use of trade names is for identification only and does not imply endorsement by the Public Health Service or by the U.S. Department of Health and Human Services. and in good general health. His spleen was just palpable. No proteinurea was detected on dip-stick testing. Numerous P. ma- lariae trophozoites and gametocytes were seen on a thick blood film. The patient was treated with a complete course of chlo- roquine. He was well and without parasites on a repeat blood film prepared four weeks later. Plasmodium malariae-infected blood with a parasitemia of 0.5% was collected into heparin by venipuncture soon after the infection was diagnosed and washed twice with RPMI culture medium without serum. The parasites were predominantly pig- mented trophozoites at the time of collection. The blood was fractionated on a step gradient of Percoll/sorbitol (steps of 50, 60,70,80, and 90% Percoll containing 4% w/v sorbitol) in RPMI medium as described previously (6). A band collected at the 501 60% Percoll interface contained 95% mature trophozoite-in- fected erythrocytes. These cells were washed three times in RPMI medium before one portion of the sample was fixed with 2% (v/v) glutaraldehyde in 0.05 M phosphate buffer, pH 7.4, with 0.12 M sucrose for 1 h at room temperature. Fixed cells were subsequently washed with 0.1 M phosphate buffer, pH 7.4, post- fixed with 1% (w/v) osmium tetroxide in 0.1 M buffer, washed three times with 0.1 M buffer, rinsed with distilled water, and stained en bloc with 1% (w/v) aqueous uranyl acetate. The cells were then dehydrated with ethanol and propylene oxide and embedded in Epon 812. Ultrathin sections were mounted on nickel grids, stained with 2% (w/v) uranyl acetate in 50% ethanol and Reynold's lead citrate, carbon-coated, and examined with a JEOL lOOCX electron microscope. The Uganda I/CDC strain of P. malariae was also used to infect a splenectomized chimpanzee. This strain had been pas- saged in Aotus monkeys and chimpanzees before inoculation (1 1). Peripheral blood was obtained by venipuncture from the chimpanzee. Most leukocytes were removed by passage over CFl1 cellulose columns (Whatman, Inc., England) (9), and the parasite-infected erythrocytes were enriched to approximately 2% with Plasmagel (Cellular Products, Inc., Buffalo, New York) (1 8). Briefly, heparinized whole blood was washed with RPMI- 1640 medium and diluted to two times the original volume prior to passage over CFl 1 columns. The CF11 eluate was then incubated with Plasmagel for 30 min at 37°C. The cells in the resulting supernatant fractions were collected and washed ex- tensively. The parasite preparations were finally fixed with 2% (v/v) glutaraldehyde in 0.1 M cacodylate buffer, pH 7.4, with 0.12 M sucrose and 0.2 pM CaCI, for 1 h at room temperature. The fixed cells were washed twice with 0.1 M phosphate buffer and processed as described earlier. 267
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Ultrastructure of the Erythrocytic Stages of Plasmodium malariae1

J Pruru:oul., 34(3). 1987. pp. 267-274 6 1987 by the Society of Protoroologists
Ultrastructure of the Erythrocytic Stages of Plasmodium malariael CARTER T. ATKINSON,* MASAMICHI AIKAWA,* EDWIN P. ROCK,** * KEVIN MARSH,*** PATRICIA M. ANDRYSIAK,****
GARY H. CAMPBELL,**** WILLIAM E. COLLINS,**** and RUSSELL J. HOWARD***** *Institute of Pathology, Case Western Reserve University, Cleveland, Ohio 44106, USA.
**Howard Hughes Medical Institute, Bethesda, Maryland 20814, USA, ***Medical Research Council Laboratories, Fajara, The Gambia,
****Malaria Branch, Center for Infectious Diseases, Centers for Disease Control, Atlanta, Georgia 30333, USA, and *****Laboratory of Parasitic Diseases, National lnstitute of Allergy and Infectious Diseases,
National Institutes of Health. Bethesda, Maryland 20205, USA
ABSTRACT. This report describes the fine structure of the erythrocytic stages of Plasmodium malariae. Erythrocytic parasites from a naturally acquired human infection and an experimentally infected chimpanzee were morphologically indistinguishable and structurally similar to other primate malarias. New findings included observations of highly structured arrays of merozoite surface coat proteins in the cytoplasm of early schizonts and on the surface of budding merozoites and the presence of knobs in the membranes of Maurer's clefts. Morphological evidence is presented suggesting that proteins are transported between the erythrocyte surface and intracellular parasites via two routes: one associated with Maurer's clefts for transport of membrane-associated knob material and a second associated with caveolae in the host cell membrane for the import or export of host- or parasite-derived substances through the erythrocyte cytoplasm.
F the four species of Plasmodium that cause human ma- 0 laria, P. malariae (Grassi & Feletti, 1890) is one of the most unusual in terms of its patchy geographic distribution and ability to persist at low intensities in infected individuals for long periods (1 0). In spite of its importance as a cause of ne- phropathy in endemic areas ( 1 5), P. malariae has not been thoroughly studied. This is largely because long-term in vitro culture of this species has not been successful and because sus- ceptible primate models rarely develop parasitemias in excess of 2-5% of the circulating erythrocytes. Consequently, only one brief study ofthe fine structure ofthis species has been published
Discontinuous Percoll gradients with (6) or without (7) sor- bitol have been used recently for purifying P. falciparum-in- fected erythrocytes from infected humans and P. ovale-infected erythrocytes from the whole blood of Aotus monkeys and chim- panzees (7). These methods were used in this study, as well as a Plasmage13 concentration technique (1 8), to purify P. malar- iae-infected erythrocytes from whole human and chimpanzee blood in order to allow observation of numerous parasitized cells by electron microscopy. In this report we describe new ultrastructural observations of both P. malariae and parasite- induced morphological changes of host erythrocytes.
(20).
MATERIALS AND METHODS An 18-year-old Gambian man attended the Medical Research
Council outpatient clinic with a two-day history of fever, head- ache, and general body pain. On examination, he was afebrile
' We are grateful to the staff and patients of the Medical Research Council Laboratories, Fajara, for assistance in sample collection and for the excellent technical assistance of Ana Milosavljevic and Kiet Dan Luc. Yoshitsugu Matsumoto and John Rabbege provided helpful com- ments on the manuscript. This work was supported in part by grants from the U.S. Agency for International Development (DPE-0453-A- 00-4027), USPHS (AI- 10645), the U.S. Army Research and Develop- ment Command (DAMDI 7-79C-9029), and the UNDP/World BanW WHO Special Programme for Research and Training in Tropical Dis- eases. This is contribution No. 1805 to the U.S. Army Drug Devel- opment Program.
Edwin Rock is a research scholar at the Howard Hughes Medical Institute.
Use of trade names is for identification only and does not imply endorsement by the Public Health Service or by the U.S. Department of Health and Human Services.
and in good general health. His spleen was just palpable. No proteinurea was detected on dip-stick testing. Numerous P. ma- lariae trophozoites and gametocytes were seen on a thick blood film. The patient was treated with a complete course of chlo- roquine. He was well and without parasites on a repeat blood film prepared four weeks later.
Plasmodium malariae-infected blood with a parasitemia of 0.5% was collected into heparin by venipuncture soon after the infection was diagnosed and washed twice with RPMI culture medium without serum. The parasites were predominantly pig- mented trophozoites at the time of collection. The blood was fractionated on a step gradient of Percoll/sorbitol (steps of 50, 60,70,80, and 90% Percoll containing 4% w/v sorbitol) in RPMI medium as described previously (6). A band collected at the 501 60% Percoll interface contained 95% mature trophozoite-in- fected erythrocytes. These cells were washed three times in RPMI medium before one portion of the sample was fixed with 2% (v/v) glutaraldehyde in 0.05 M phosphate buffer, pH 7.4, with 0.12 M sucrose for 1 h at room temperature. Fixed cells were subsequently washed with 0.1 M phosphate buffer, pH 7.4, post- fixed with 1% (w/v) osmium tetroxide in 0.1 M buffer, washed three times with 0.1 M buffer, rinsed with distilled water, and stained en bloc with 1% (w/v) aqueous uranyl acetate. The cells were then dehydrated with ethanol and propylene oxide and embedded in Epon 812. Ultrathin sections were mounted on nickel grids, stained with 2% (w/v) uranyl acetate in 50% ethanol and Reynold's lead citrate, carbon-coated, and examined with a JEOL lOOCX electron microscope.
The Uganda I/CDC strain of P. malariae was also used to infect a splenectomized chimpanzee. This strain had been pas- saged in Aotus monkeys and chimpanzees before inoculation (1 1). Peripheral blood was obtained by venipuncture from the chimpanzee. Most leukocytes were removed by passage over CFl1 cellulose columns (Whatman, Inc., England) (9 ) , and the parasite-infected erythrocytes were enriched to approximately 2% with Plasmagel (Cellular Products, Inc., Buffalo, New York) (1 8). Briefly, heparinized whole blood was washed with RPMI- 1640 medium and diluted to two times the original volume prior to passage over CFl 1 columns. The CF11 eluate was then incubated with Plasmagel for 30 min at 37°C. The cells in the resulting supernatant fractions were collected and washed ex- tensively. The parasite preparations were finally fixed with 2% (v/v) glutaraldehyde in 0.1 M cacodylate buffer, pH 7.4, with 0.12 M sucrose and 0.2 pM CaCI, for 1 h at room temperature. The fixed cells were washed twice with 0.1 M phosphate buffer and processed as described earlier.
267
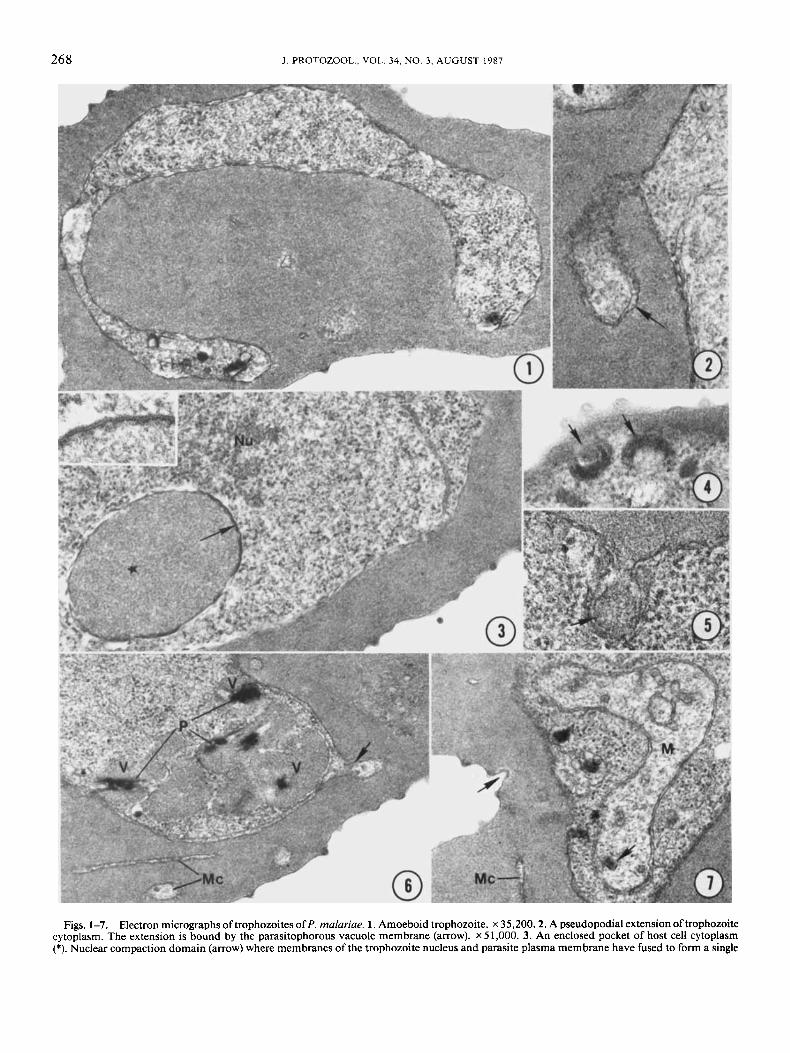
268 J. PROTOZOOL., VOL. 34, NO. 3, AUGUST 1987
Figs. 1-7. Electron micrographs oftrophozoites of P. malariae. 1. Amoeboid trophozoite. x 35,200.2. A pseudopodial extension oftrophozoite cytoplasm. The extension is bound by the parasitophorous vacuole membrane (arrow). x 51,000. 3. An enclosed pocket of host cell cytoplasm (*). Nuclear compaction domain (arrow) where membranes of the trophozoite nucleus and parasite plasma membrane have fused to form a single

ATKINSON ET AL.-FINE STRUCTURE O F PLASMODIUM MAL.4RIAE 269
RESULTS Trophozoites. Erythrocytic forms of P. malariae from the nat-
ural human infection and from the experimentally infected chimpanzee were morphologically indistinguishable. The par- asites resided within a membrane-bounded parasitophorous vacuole and were surrounded by a single plasma membrane. Young trophozoites were amoeboid (Fig. 1). Pseudopodial extensions of the trophozoite cytoplasm often extended through- out the cytoplasm of infected erythrocytes (Fig. 2). The pseu- dopods were surrounded by a parasitophorous vacuole mem- brane that often appeared to loop and branch beyond the parasite cytoplasm as tube-like extensions or Maurer’s clefts in the eryth- rocyte cytoplasm. Some pseudopodial extensions occasionally formed loops and pockets that enclosed indentations of host cell cytoplasm (Fig. 3 ) . Enclosed erythrocyte cytoplasm was less electron dense and more “foamy” in appearance than the red blood cell stroma outside the pockets.
Trophozoites possessed a variety of organelles, including a small cytostome composed of two concentric, electron-dense rings with an internal diameter of 50-60 nm (Figs. 4, 5); pe- ripherally arranged food vacuoles that contained red blood cell cytoplasm and angular crystals of malarial pigment (Fig. 6); one or more mitochondria with tubular cristae and occasional elec- tron-dense bodies (Fig. 7); and a nucleus with a prominent nu- cleolus (Fig. 3) . The trophozoite cytoplasm was packed with numerous ribosomes.
The nucleus of older trophozoites was often located at the margin of the parasite cytoplasm and closely appressed to the plasma membrane. Nuclear compaction domains were occa- sionally observed along these areas (Fig. 3) . The domains were composed of regions where the parasite plasma membrane and the two nuclear membranes were fused into a single, electron- dense, thickened membrane (Fig. 3, inset).
Schizonts. As trophozoites continued their growth, they be- came larger and rounder in shape. The earliest schizogonic stages were identified by the appearance of electron-dense, centriolar plaques in the nuclear membrane. When nuclear division began, scattered electron-dense thickenings developed under the plas- ma membrane. These thickenings formed the apical ends of merozoites as they differentiated and budded from a centrally located residual body.
The cytoplasm of early schizonts occasionally contained free masses of cuboidal or rhomboidal striated material between nuclei and the parasite plasma membrane (Fig. 8). The striated arrays had a regular period of - 12 nm. Similar material was observed in the parasitophorous vacuole space and on the sur- face of budding merozoites (Figs. 8, 9). Material on the surface of budding merozoites was composed of regular denticulations spaced about 12 nm apart that resembled a “zipper.” The zip- per-like material graded gradually into a granular electron-dense matrix in the parasitophorous vacuole space that appeared to be composed of subunits approximately the same size and shape as individual striations that formed the elongate “zippers” (Fig. 9).
Mature segmenters contained free merozoites and a residual body that were embedded in the electron-dense matrix described above and surrounded by the parasitophorous vacuole mem- brane (Fig. 10). Mature merozoites contained a pellicle com- posed of an outer plasma membrane underlain by two closely apposed unit membranes, microtubules, a nucleus, a mito- chondrion with tubular cristae, a pair of electron-dense rhoptries and numerous micronemes. Merozoites also contained a few conspicuous electron-dense bodies that were approximately the same size as micronemes (Fig. 10).
Gametocytes. Gametocytes were bounded by a parasitopho- rous vacuole membrane and a pellicle composed of an outer unit membrane underlain by an irregularly spaced, discontin- uous pair of closely apposed unit membranes (Fig. 1 I). Using the morphological criteria of Aikawa et al. (2), we were able to identify both microgametocytes and macrogametocytes in thin- sectioned material. Microgametocytes contained a large, irreg- ularly shaped nucleus, osmiophilic bodies, and cytoplasm com- posed of fine granular material with scattered ribosomes (Fig. 1 1 ) . Mitochondria with tubular cristae, food vacuoles with pig- ment granules, and a cytostome were also observed. Macroga- metocytes were similar in structure, but contained larger num- bers of osmiophilic bodies and cytoplasmic ribosomes. Some macrogametocytes contained as many as three cytostomes in a single section.
Microgametocyte-infected erythrocytes occasionally con- tained unusual membrane-bounded bundles of microtubules from 200-600 nm in diameter (Fig. 11). These bundles were surrounded by the parasitophorous vacuole membrane, sug- gesting that they connected directly with the gametocyte cyto- plasm. Microtubules in the bundles appeared as pairs in cross- section and were arranged loosely in the 9+2 configuration of cilia and flagella (Fig. 11).
Host cell alterations. Electron-dense material (EDM) of Ai- kawa et al. (5) was commonly observed in the parasitophorous vacuole space and Maurer’s clefts of trophozoites and schizonts. The material was composed of a fine or coarsely granular matrix and present as a thin layer around trophozoites and early schiz- onts or in pocket-like indentations of the parasite cytoplasm (Fig. 12). Balls of amorphous EDM with a diameter of 40-50 nm were occasionally scattered in the granular matrix filling the parasitophorous vacuole space and associated Maurer’s clefts (Fig. 13). Scattered randomly throughout the cytoplasm of in- fected erythrocytes were similar balls of EDM surrounded by an electron-lucent halo. The balls of cytoplasmic EDM were often associated with the parasitophorous vacuole membrane next to the main body of trophozoites or along the Maurer’s clefts (Figs. 14-17). The electron-lucent halo appeared to de- velop as the material crossed the parasitophorous vacuole mem- brane, suggesting movement of EDM through this membrane into the erythrocyte cytoplasm.
Small balls of erythrocytic EDM were also found immediately under the erythrocyte plasma membrane and appeared to be associated with cup-like invaginations of the red cell membrane (Figs. 18-19). These invaginations appeared to fuse with the
t
thickened electron-dense membrane. Parasite nucleus contains a nucleolus (Nu) and it has shifted to the periphery of the parasite cytoplasm around the pocket of host cell cytoplasm. x 35,000. Inset: Higher magnification of the nuclear compaction domain. x 100,000.4. Oblique section through a pair of adjacent cytostomes (arrows). Cytostomes contain a pair of Concentric electron-dense rings. x 50,700. 5. Small cytostome with a developing food vacuole (arrow). x 78,000. 6 . Peripherally located food vacuoles in the trophozoite cytoplasm. Each vacuole (V) is bounded by a unit membrane and contains red cell cytoplasm and crystals of malarial pigment (P). Note pseudopodial extension of a Maurer’s cleft from the parasitophorous vacuole membrane (arrow), as well as several other clefts (Mc) in the host cell cytoplasm. x 32,800. 7. Mitochondrion (M) with tubular cristae and an electron-dense body (arrow) in the trophozoite cytoplasm. Note Maurer’s clefts (Mc) in the erythrocyte cytoplasm and a caveola in the host cell membrane (arrow). x 38,400.

270 J . PROTOZOOL., VOL. 34, NO. 3, AUGUST 1987
Figs. 8-1 I . Electron micrographs of schizonts and a microgametocyte of P. rnalariue. 8. Striated, highly organized merozoite surface coat precursors (S) in the cytoplasm of an early schizont. The parasitophorous vacuole space (Pv) contains the “zipper-like” merozoite surface coat on the surface of the schizont (arrows) and a coarse granular matrix. x 74,000. 9. “Zipper-like” material (arrows) fills the parasitophorous vacuole space (Pv) between a budding merozoite (M) and the parasite cytoplasm (P). Note coarse granular material in the parasitophorous vacuole space. x 74,000.10. Mature segmenter. Merozoites (M) are embedded in an electron-dense matrix that fills the parasitophorous vacuole space. Merozoites are bounded by a three-layered pellicle (arrows) and contain rhoptries (R), micronemes (Mi), electron-dense bodies (Db), and a single nucleus (N). x 31,200. 11. Microgametocyte bounded by a pellicle consisting of an outer plasma membrane and an interrupted pair of closely apposed unit membranes (arrows). Osmiophilic bodies (0s) are scattered in the cytoplasm. Two membrane-bounded bundles of microtubules (Mt) are surrounded by a parasitophorous vacuole membrane in the host cell cytoplasm. The microtubules appear as doublets in cross section (arrowheads). ~45,500.
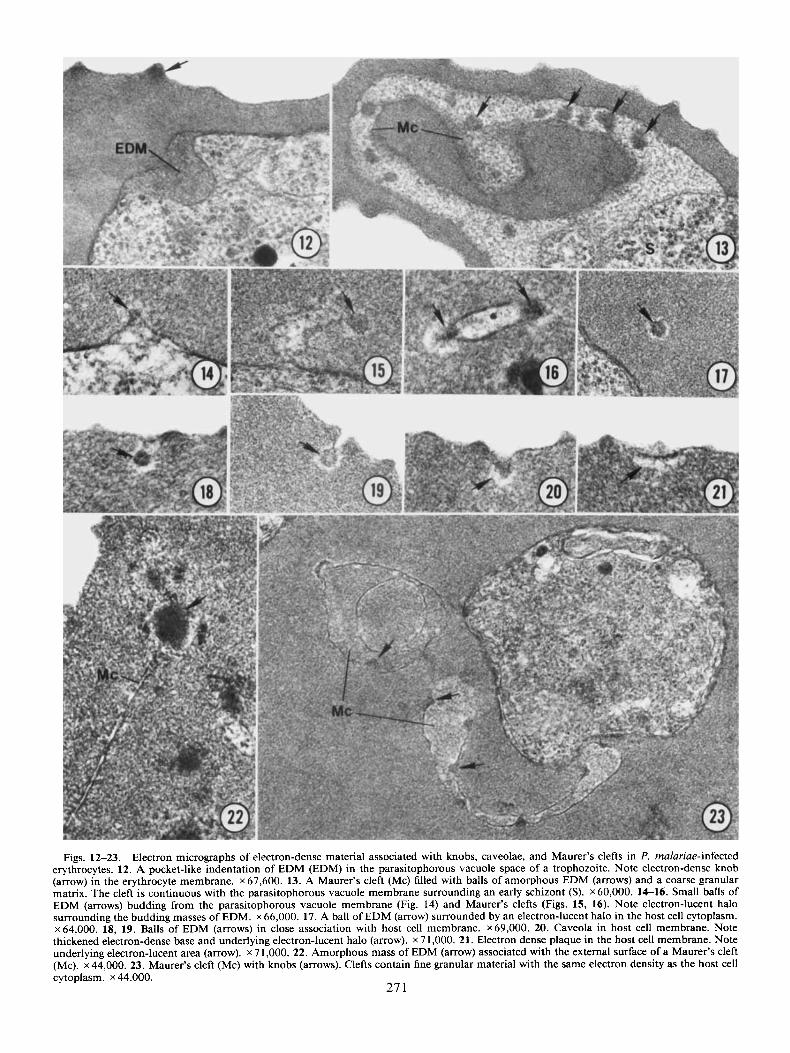
Figs. 12-23. Electron micrographs of electron-dense material associated with knobs, caveolae, and Maurer’s clefts in P. rnalariae-infected erythrocytes. 12. A pocket-like indentation of EDM (EDM) in the parasitophorous vacuole space of a trophozoite. Note electron-dense knob (arrow) in the erythrocyte membrane. x 67,600. 13. A Maurer’s cleft (Mc) filled with balls of amorphous EDM (arrows) and a coarse granular matrix. The cleft is continuous with the parasitophorous vacuole membrane surrounding an early schizont (S). x 60,000. 14-16. Small balls of EDM (arrows) budding from the parasitophorous vacuole membrane (Fig. 14) and Maurer’s clefts (Figs. 15, 16). Note electron-lucent halo surrounding the budding masses of EDM. x 66,000. 17. A ball of EDM (arrow) surrounded by an electron-lucent halo in the host cell cytoplasm. x 64,000. 18, 19. Balls of EDM (arrows) in close association with host cell membrane. x 69,000. 20. Caveola in host cell membrane. Note thickened electron-dense base and underlying electron-lucent halo (arrow). x 7 1,000. 21. Electron dense plaque in the host cell membrane. Note underlying electron-lucent area (arrow). x 7 1,000. 22. Amorphous mass of EDM (arrow) associated with the external surface of a Maurer’s cleft (Mc). ~44,000. 23. Maurer’s cleft (Mc) with knobs (arrows). Clefts contain fine granular material with the same electron density as the host cell cytoplasm. x 44,000.
27 1

272 J. PROTOZOOL., VOL. 34, NO. 3, AUGUST 1987
EDM to form small, membrane-bounded caveolae that were continuous with the erythrocyte plasma membrane (Fig. 20). The membrane forming the base of the cup-like caveolae was thickened and electron dense and underlain by an electron- lucent area that corresponded to the halo surrounding the cy- toplasmic balls of EDM (Fig. 20). Some caveolae appeared to have opened outward onto the surface of the infected erythro- cyte, creating electron-dense plaques in the erythrocyte mem- brane (Fig. 21).
A second type of EDM was observed more rarely in the cy- toplasm of infected erythrocytes. This EDM formed amorphous masses, without an associated halo, that were associated with the external surface ofMaurer’s clefts (Fig. 22). In some sections, this material formed electron-dense thickenings associated with the membranes of Maurer’s clefts, morphologically identical to knobs found on the erythrocyte surface (Fig. 23). Maurer’s clefts containing these internal knobs contained fine granules with an electron density similar to that of the host cell cytoplasm (Fig. 23) indicating that they were not pocket-like identations of the erythrocyte membrane.
The surface of trophozoite-, schizont- and gametocyte-in- fected erythrocytes was modified into numerous, regularly spaced, electron-dense knobs. Oblique sections demonstrated that the knobs were spaced approximately 120 nm apart on the eryth- rocyte surface. They measured 46 (34-57) nm in height and 80 (65-98) nm in diameter at their base (n = 25). Knobs were composed of electron-dense material that fused gradually into the host cell cytoplasm (Fig. 12).
DISCUSSION Ultrastructural observations of P. malariae were similar to
those of Smith & Theakston (20). These authors briefly de- scribed trophozoites and schizonts and reported a nucleus with a nucleolus, mitochondria with tubular cristae, and pigment granules in trophozoites from human infections that were ac- quired naturally in The Gambia and The Cameroons in Western Africa. Merozoites in mature segmenters contained organelles characteristic of this invasive stage, including a pair of rhoptries and numerous micronemes (20). We observed similar organelles in this study and also identified the presence of a small cyto- stome, membrane-bounded peripheral food vacuoles, and nu- clear compaction domains. Wiinderlich et al. (23) described similar nuclear compaction domains in early ring stages and trophozoites of P. knowlesi and P. chabaudi and was able to resolve a heptalaminate structure in the closely apposed nuclear and plasma membranes. They noted that this structure disap- peared as parasites matured to form schizonts and speculated that it may play an undetermined role in facilitating the ex- change of substances directly between the parasite nucleus and parasitophorous vacuole space. We observed these structures primarily in older trophozoites during margination of the par- asite nucleus with the plasma membrane and development of electron-dense plaques in the nuclear envelope, suggesting that it may play some as yet undetermined role in the early events of nuclear division.
The masses of rhomboidal or cuboidal striated material we observed in early schizonts of P. malariae and the zipper-like material on the surface of budding merozoites are identical to the highly structured merozoite surface coat described by Ban- nister et al. (8) in erythrocytic schizonts of P. knowlesi. These workers first detected coat formation as patches of regular, 20- nm high denticulations on the surface of early binucleated schiz- onts, but did not describe early precursors of this material in the schizont cytoplasm. The morphological similarities between
the striated material we observed in the schizont cytoplasm and the zipper-like material on the surface of budding merozoites of P. ma(ariae suggests that the merozoite surface coat is syn- thesized in large amounts in the schizont cytoplasm before trans- port to the surface of the parasite. Precisely how this material reaches the surface of the developing schizont remains to be determined.
Large bundles of membrane-bounded microtubules similar to those we occasionally observed in microgametocytes of P. malariae have not been described in other ultrastructural studies of malarial parasites. These bundles were similar enough in structure to the differentiating flagella of microgametes (4), how- ever, to suggest that they represent some form of aberrant ex- flagellation, perhaps induced by temperature shock when the blood sample was fixed.
Other species of primate Plasmodium that possess a promi- nent nucleolus include P. ovule of humans and P. brasilianum of New World monkeys (1 6, 2 1). Of these two species, tropho- zoites of P. brasilianum also possess mitochondria with tubular cristae (21). Several authors have suggested that infections of P. brasilianum may be anthroponoses of P. malariae that have become established in New World monkeys (1 0). Evidence has included similarities in quartan periodicity, morphology of the erythrocytic and exoerythrocytic stages, and antigenic similar- ities between both species. We noted a striking similarity be- tween the erythrocytic stages of P. malariae and P. brasilianum. Important differences were evident, however. The inside di- ameter of cytostomes of P. malariae was considerably smaller than those of P. brasilianum (60 nm vs. 170 nm). In addition, mitochondria of P. malariae lacked an associated spherical body as described by Sterling et al. (21) in trophozoites of P. brasi- lianum. While the size and density of knobs on P. brasilianum- infected erythrocytes were comparable to those in our study, Sterling et al. (21) did not describe EDM in their investigation although their micrographs (Figs. 11, 22 in ref. 21) illustrate both large and small masses ofEDM. Significantly, these authors did not detect the presence of caveolae. It is uncertain whether these differences represent host-induced morphological varia- tions or strain or species differences between the two parasites.
During their growth and development, malaria parasites in- duce several alterations in host erythrocyte morphology includ- ing cytoplasmic clefts, nodules, knobs, and caveola-vesicle com- plexes in the erythrocyte plasma membrane (1, 3, 17). Smith & Theakston (20) observed numerous knobs and occasional cy- toplasmic clefts in P. malariae-infected erythrocytes. Their lim- ited study failed to detect caveolae in the membrane of host erythrocytes. The caveolae that we observed lacked the small alveolar-like vesicles of the caveolae-vesicle complex described in vivax- and ovule-type primate malarias (1,3). They were more similar to the caveolae described in the primate parasites, P. coatneyi and P. knowlesi (19, 24). Aikawa et al. (3) suggested that the caveolae-vesicle complexes of vivax- and ovule-type malarias correspond to the Schiiffner’s dots visible in Giemsa- stained blood smears. James (1 4) described a similar stippling, termed Ziemann’s stippling, in P. malariae-infected erythro- cytes when blood smears were stained for a long time by Ro- manowsky stains. The stippling consisted of dots and points that were smaller and less distinct than Schiiffner’s dots. The caveolae we observed may correspond to Ziemann’s stippling and result from the deposition of stain in the flask-shaped cav- eolae of the erythrocyte membrane.
The large masses of EDM we observed in association with Maurer’s clefts in this study appear to correspond to the EDM described by Aikawa et al. (5) in the parasitophorous vacuole space and adjacent to Maurer’s clefts in the erythrocytic stages

ATKINSON ET AL.-FINE STRUCTURE OF PLASMODIUM MALARIAE 273
of P. falciparum. These authors suggested that movement of EDM from the parasitophorous vacuole to the erythrocyte membrane via the Maurer’s clefts may provide a system of intracellular transport for the movement of histidine-rich knob material and other knob-associated proteins to the host cell membrane. Our observations of unusual internal knobs in the Maurer’s clefts of P. malariae-infected erythrocytes and the presence of similar EDM in the parasitophorous vacuole space and erythrocyte cytoplasm suggest that a similar mechanism may be important in the formation of knobs in this species of Plasmodium.
Small vesicular structures, corresponding in size and electron density to the small balls of EDM in the cytoplasm of P. mala- riae-infected erythrocytes, have been observed in studies of P. knowlesi, P. simium, P.fieldi, P. vivax, P. ovale, P. cynomolgi, and P. simiovale (1, 3 , 16, 24). Each of these species produce flask-shaped caveolae or caveolae-vesicle complexes with thick- ened electron-dense bases that are morphologically similar to those found in P. malariae-infected erythrocytes. Several au- thors have suggested that the small vesicles may fuse with the host cell membrane to form caveolae or develop as micropi- nocytotic vesicles from caveolae as they complete their invag- ination from the host cell membrane (22,24). Either route would provide a mechanism for the import or export of host- or par- asite-derived polypeptides, independent of Maurer’s clefts, and would provide a mechanism for the insertion of plasmodia1 antigens into the erythrocyte membrane.
Observation of the balls of EDM in close association with the parasitophorous vacuole membrane and Maurer’s clefts in this study suggests that material may be originating from or is being added to the parasitophorous vacuole space. The electron-lucent halo around the balls of EDM and at the base of caveolae cor- responds in location to the clathrin coating of coated pits and vesicles that facilitates the intracellular transport of material in eukaryotic cells, but lacks the fibrous, electron-dense structure of this protein ( 1 2). The nature of the electron-lucent halo is currently unknown, but it may facilitate the movement of hy- drophilic parasite- or host-derived substances through the hy- drophobic host cell and parasitophorous vacuole ‘membranes. Thus, Maurer’s clefts and caveolae may provide different mech- anisms for the intracellular transport of host- and parasite-de- rived proteins in P. malariae-infected erythrocytes-one asso- ciated with the Maurer’s clefts for movement of membrane associated knob materials and a second associated with caveolae for transport of water-soluble substances.
Howard et al. ( 1 3) , recently demonstrated the export of a water soluble histidine-rich protein (Pf HRP 2) from P. falciparum- infected erythrocytes. These workers observed electron-dense “packets” of Pf HRP 2 in the host cytoplasm and under the plasma membrane of infected erythrocytes in cryo-sectioned material labeled with monoclonal antibody specific for Pf HRP 2. Although there are morphological similarities in the appear- ance of the electron-dense packets which export Pf HRP 2 in P. falciparum-infected cells (1 3 ) and the balls of EDM described here for P. malariae, preliminary experiments using Western- blotted, SDS-polyacrylamide gel-separated polypeptides of P. malariae have shown no reaction with specific antibodies to Pf HRP 2 (Rock & Howard, unpubl.).
This study highlights the diversity of morphological structures and alterations that are associated with the host cell cytoplasm and membrane in P. malariae-infected erythrocytes. The Mau- ref’s clefts and associated EDM, the balls of EDM and their surrounding electron-lucent halo, caveolae, and knobs all reflect parasite-induced changes in structure that have functional sig- nificance for this malarial parasite. Comparative studies be-
tween P. malariae and other knob- and caveolae-producing par- asites may help to reveal some of these structure-function relationships.
LITERATURE CITED 1. Aikawa, M., Hsieh, C.-L. & Miller, L. H. 1977. Ultrastructural
changes of the erythrocyte membrane in ovale-type malarial parasites. J. Parasitol., 63: 152-154.
2. Aikawa, M., Huff, C. G. & Sprinz, H. 1969. Comparative fine structure study of the gametocytes of avian, reptilian and mammalian malarial parasites. J. Ultrastruct. Res., 26: 316-331.
3. Aikawa, M., Miller, L. H. & Rabbege, J. 1975. Caveola-vesicle complexes in the plasmalemma of erythrocytes infected by Plasmodium vivax and P. cynomolgi. Am. J. Path., 79: 285-300.
4. Aikawa, M. & Sterling, C . R. 1974. Intracellular Parasitic Pro- tozoa. Academic Press, Inc., New York.
5. Aikawa, M., Uni, Y., Andrutis, A. T. & Howard, R. J. 1986. Membrane-associated electron-dense material of the asexual stages of Plasmodium falciparum: evidence for movement from the intracellular parasite to the erythrocyte membrane. Am. J. Trop. Med. Hyg., 35: 30- 36.
6. Aley, S. B., Sherwood, J. A,, Marsh, R., Eidelman, 0. & Howard, R. J. 1986. Identification of isolate-specific proteins on sorbitol-en- riched Plasmodium faleiparum-infected erythrocytes from Gambian pa- tients. Parasitology, 92: 51 1-525.
7. Andrysiak, P. M., Collins, W. E. & Campbell, G. H. 1986. Con- centration of Plasmodium ovale- and Plasmodium viva-infected eryth- rocytes from non-human primate blood using Percoll gradients. Am. J. Trop. Med. Hyg., 35: 25 1-254.
8. Bannister, L. H., Mitchell, G. H., Butcher, G. A,, Dennis, E. D. & Cohen, S. 1986. Structure and development of the surface coat of erythrocytic merozoites of Plasmodium knowlesi. Cell Tissue Rex, 245:
9. Beutler, E., West, C. & Blume, K. G. 1976. Removal ofleucocytes and platelets from whole blood. J. Lab. Clin. Med., 88: 328-333.
10. Coatney, G. R., Collins, W. E., Warren, M. & Contacos, P. G. 197 1. The Primate Malarias. U.S. Gov. Print. Off., Washington, D.C.,
1 1. Collins, W. E., Schwartz, I. K., Skinner, J. C. & Broderson, J. R. 1984. Studies on the Uganda IKDC strain of Plasmodium malariae in Bolivian Aotus monkeys and various anophelines. J. Parasitol., 70:
12. Goldstein, J. L., Anderson, R. G. W. & Brown, M. S. 1979. Coated pits, coated vesicles and receptor-mediated endocytosis. Nature,
13. Howard, R. J., Uni, S., Aikawa, M., Aley, S. B., Leech, J. H., Lew, A. M., Wellems, T. E., Rener, J. & Taylor, D. 1986. Secretion of a malarial histidine-rich protein (Pf HRP 11) from Plasmodium fal- ciparum-infected erythrocytes. J. Cell Biol., 103: 1269-1 277.
14. James, S. P. 1929. A note on the Shute technique for staining malarial parasites with Leishman’s stain and on the stippling in infected red blood corpuscles which it reveals. Trans. Roy. SOC. Trop. Med. Hyg.,
15. Kibukamasoke, J. W. 1984. Renal disease in the tropics, in Woodruff, A. W. &Wright, S. G., eds., Medicinein the Tropics. Churchill Livingston, New York, pp. 630-636.
16. Matsumoto, Y., Matsuda, S. & Yoshida, Y. 1986. Ultrastruc- ture of erythrocytic stages of Plasmodium ovale in humans. Am. J. Trop. Med. Hyg., 35: 689-696.
17. - 1986. Ultrastructure of human erythrocytes infected with Plasmodium ovule. Am. J. Trop. Med. Hyg., 35: 697-703.
18. Pasvol, G. 1978. Separation of viable schizont-infected red cells of Plasmodium falciparum from human blood. Ann. Trop. Med. Par- asitol., 72: 87-88.
19. Rudzinska, M. A. & Trager, W. 1968. The fine structure of trophozoites and gametocytes in Plasmodium coatneyi. J. Protozool.,
20. Smith, D. H. & Theakston, R. D. G. 1970. Comments on the ultrastructure of human erythrocytes infected with Plasmodium ma- lariae. Ann. Trop. Med. Parasitol., 64: 329-330.
21. Sterling, C. R., Aikawa, M. & Nussenzweig, R. S. 1972. Mor-
28 1-290.
pp. 209-229.
677-68 1.
279: 679-685.
23: 269-278.
15: 73-88.

274 J . PROTOZOOL., VOL. 34, NO. 3, AUGUST 1987
phological divergence in a mammalian malarial parasite: the fine struc- ture ofPlasmodium brasilianum. Proc. Helm. SOC. Wash., 39: 109-1 29.
Erythrocyte membrane alterations induced by Plasmodium simium in- fection in Saimiri sciureus: relation to Schiiffner’s dots. J. Parasitol.,
23. Wiinderlich, F., Stubig, H. & Konigk, E. 1980. Nuclear enve-
lope-plasma membrane-compaction domain in malaria parasites. Cell Biol. Int. Rep., 4: 51 9-524.
22. Sterling, C. R., Seed, T. M., Aikawa, M. & Rabbege, J. 1975. 24. - 1982. Intraerythrocytic development of Plasmodium knowlesi: structure, temperature and Ca2+-response of the host and par- asite membranes. J. Protozool., 29: 49-59.
61: 177-188. Received 19 I87; accepted 10 III 87
J . Protozool, 34(3). 1987, pp. 274-277 (I\ 1987 by the Society o f Protozoologists
An Intranuclear Microsporidium Associated with Acute Anemia in the Chinook Salmon, Oncorhynchus tsha wytscha
R. A. ELSTON,* M. L. KENT,* and L. H. HARRELL** *Center for Marine Disease Control, Battelle/Marine Research Laboratory,
439 West Sequim Bay Road, Sequim, Washington 98382 and **Coastal Zone and Estuarine Studies Division, Northwest and Alaska Fisheries Center, National Marine Fisheries Service,
National Oceanic and Atmospheric Administration, 2725 Montlake Boulevard East, Seattle, Washington 981 12
ABSTRACT. An intranuclear microsporidium is described from hemoblastic cells of the chinook salmon, Oncorhynchus tshawytscha. The infection is associated with an acute anemia in the fish. Up to 47% of the hemoblast nuclei were infected in anemic fish. The organisms, found only in spleen and kidney tissues, were 1-2 fim in diameter and consisted of vegetative and early sporulation forms. This microsporidium differs from known species which parasitize fish in its tissue location; however, the absence of mature spores and other life cycle stages precludes determination of its precise taxonomic identity.
HE chinook salmon, Oncorhynchus tshawytscha, is one of T several species of Pacific salmon that can be reared throughout its entire life cycle in captivity. In commercial or research husbandry of the chinook salmon, this includes three to four years ofresidence in saltwater net pens or cages. Although research on salmonid diseases has emphasized diseases occur- ring in the fish in their freshwater phase of development, infec- tious diseases of the fish during the saltwater phase can have a substantial impact on the health, survival, and growth of the fish (3,4). Of particular concern is an anemia of unknown cause which has been associated with mortality in chinook salmon broodstocks at the National Marine Fisheries Service’s Man- Chester Marine Experimental Station on central Puget Sound, Washington.
Over 70 species of microsporidia have been reported from fishes (1, 8). some of which are significant pathogens of wild or cultured species (1). Most microsporidia of fishes form large, often macroscopically visible, xenomas or cysts originating from intracytoplasmic reproduction, and the most significant patho- logical changes that they induce are usually associated with pres- sure atrophy or tissue replacement (2).
Microsporidia are intracellular parasites and most occur only in the host cell cytoplasm (1 0, 1 1). However, at least one species, Microsporidium rhabdophilia. has been described that develops exclusively in the nuclei of rodlet cells in the chinook salmon as well as in other salmonid species (5). We report here on the occurrence of another intranuclear microsporidium of chinook salmon. The parasite was observed in hemoblastic cells in the hematopoietic tissues of anemic salmon, namely the spleen and kidney. Thus, this preliminary report notes the occurrence of
I The Marine Research Laboratory is part of the Pacific Northwest Laboratory, which is operated for the Department of Energy by Battelle Memorial Institute under Contract DE-AC06-76RLO 1830. We thank Dr. E. Canning for suggestions on the electron micrographs. Ms. Marilyn Wilkinson assisted in the preparation of specimens for histology and electron microscopy, and Ms. Ann Trelstad assisted in the preparation of the manuscript.
the microorganism in association with significant anemic dis- ease of the fish and is the second report of an intranuclear microsporidium of vertebrates.
MATERIALS AND METHODS Six 3-year old salmon, four of which were clinically anemic
as determined by pallor of the gills, were collected in September 1983 from the National Marine Fisheries Service’s Manchester Marine Experimental Station. Hematocrit values were deter- mined by centrifuging blood samples in capillary hematocrit tubes. Spleen, kidney, liver, and gill tissues were excised and processed for histological and electron microscopic analysis us- ing previously reported methods (4). Histological sections were stained with hematoxylin and eosin. The proportion of hemo- blast cells in the spleen containing the intranuclear inclusions was enumerated by examining 100 randomly selected cells from the spleen of each fish using a 40 x objective. Thin sections for electron microscopy were examined on a Philips EM 300 using an accelerating voltage of 80 kV.
RESULTS Intranuclear inclusions, later shown to be microsporidian stages
by electron microscopy, were detected in hemoblast cells of the spleen and kidney (i.e. cells which did not comprise the fibrous stroma, endothelium, excretory tubules, or mature erythro- cytes). They were not observed in other cell types, including rodlet cells, or in other organs or in the spleen or kidney ofwild- caught clinically healthy fish (Fig. 1). The inclusions were rough- ly spherical, about 1.5 pm in diameter, and eosinophilic (Fig.
Table I shows the hematocrit values of the fish from the experimental net pens and the corresponding proportion of spleen cells with the intranuclear inclusions. Two of the fish with the lowest numbers of inclusions (8 and 13% of the cells) had he- matocrit values in the normal range. The other fish with sub- normal hematocrit values had up to 47% of the hemoblast cells infected with microsporidia.
Examination of the spleen and kidney interstitium by electron
2).




















