
The junctional epithelium originates from the odontogenic epithelium of an erupted tooth
Upload
independentCategory
view
0download
0
Graefe's Arch Clin Exp Ophthalmol (1992)230:468-475 Graefe's Archive for Clinical and Experimental
Ophthalmology © Springer-Verlag 1992
The ultrastrueture of transplanted rabbit retinal epithelium* Peter Gouras, Robert Lopez, Mariel Brittis, and Hild Kjeldbye
Edward S. Harkness Eye Institute, Columbia University, 630 W. 168 Street, New York, NY 10032, USA
Received May 17, 1991 / Accepted November 8, 1991
Abstract. Ret ina l ep i the l ia l cells f r om p i g m e n t e d rabb i t s have been cu l tu red a n d r ad io l abe l l ed du r ing d iv is ion wi th 3H- thymid ine a n d t r a n s p l a n t e d to the subre t ina l space o f a lb ino r abb i t s us ing a pa r s p l a n a t r ansv i t r ea l a p p r o a c h . The hos t r abb i t s were m a i n t a i n e d on cyclo- spor ine i m m u n o s u p p r e s s i o n af ter t r a n s p l a n t a t i o n sur- gery. The t r a n s p l a n t cells survive and m a i n t a i n a m o r - pho log i ca l l y n o r m a l a p p e a r a n c e for a t leas t 5 mon ths . The p i g m e n t e d epi the l ia l cells re ta in thei r p i g m e n t a t i o n and b e c o m e in t eg ra t ed wi th the hos t re t ina l ep i the l ium fo rming j u n c t i o n a l complexes wi th bo th the hos t and o the r t r a n s p l a n t cells. The t r a n s p l a n t cells con t ac t hos t p h o t o r e c e p t o r s wi th thei r ap ica l processes and p h a g o c y - tize ou te r segments . There a p p e a r s to be no s igni f icant cell d iv is ion a m o n g the t r a n s p l a n t cells.
Introduction
This p a p e r descr ibes the u l t r a s t r ue tu r a l a p p e a r a n c e o f t r i t i a ted t h y m i d i n e l abe led re t ina l ep i the l ia l cells t rans- p l a n t e d to the subre t ina l space o f r abb i t re t ina . The do- no r rabb i t s are p i g m e n t e d while the hos ts a re a lb inot ic , which a l lows c o n t i n u o u s e x a m i n a t i o n by o p h t h a l m o s - copy o f the t r a n s p l a n t site. P rev ious s tudies have shown tha t a lb ino r a b b i t re t ina l ep i the l ium can be t r a n s p l a n t e d to p i g m e n t e d hos ts and can survive and func t ion for several days [8]. The p resen t s tudy used cyc lospor ine i m m u n o s u p p r e s s i o n to faci l i ta te the longer t e rm surviva l and e x a m i n a t i o n o f such t r ansp lan t s . Some o f these re- sults have been pub l i shed in abs t r ac t f o rm [1] and in a s y m p o s i u m h o n o r i n g G u y Verriest [6]. Recent ly several o the r l abo ra to r i e s [2, 10] have also pub l i shed abs t rac t s showing s imi lar resul ts in r a b b i t r e t ina us ing o the r me th - ods , c a r b o n par t ic les [3] or a f luorescent s ta in [10] to ident i fy the t r a n s p l a n t e d cells.
Correspondence to: P. Gouras
* Supported by The G. Harold and Leila Y. Mathers Charitable Foundation, Research to Prevent Blindness, Inc., The Bedminster Fund, Inc., and The Alcon Research Institute
Materials and methods
Cultured retinal epithelium
The methodology for isolating and culturing retinal epithelial cells from pigmented rabbit eyes were similar to our methods for isolat- ing these cells from human eyes [4]. Rabbit eyes were kept at 4 ° C for about 18 h to facilitate the separation of the retina from the epithelial layer. After rinsing in sterile satine, the portion of the globe anterior to the ora serrata was dissected away and dis- carded. The vitreous and the neural retina were carefully removed, the latter cut at the optic nerve. The globe was placed on cotton in a sterile jar and the epithelium was rinsed 3-4 times with a balanced salt solution (BSS) without Mg ++ or Ca ++ and supple- mented with 0.1 mM EDTA. Then 2 ml of 0.25% trypsin dissolved in BSS was added and the eye cup incubated in a capped jar at 37°C for 0.1-1 h. The trypsin solution was gently aspirated and replaced with 1 ml of minimal essential medium (MEM) supple- mented with 20% fetal calf serum (FCS) and 5 g/1 glucose. The retinal epithelial cells were suspended in this solution by repeated pipetting. The cell solution was removed, the cell concentration determined with a hemocytometer and the cells plated at approxi- mately 150,000/cm z. The cells settle and attach to the surface of the culture plate; after 3 days the media was changed and unat- tached cells were washed away. Then the new medium was supple- mented with 0.2 gCi/ml 3H-thymidine. This medium was replaced every 2 days with new radioactive medium until the cells form a monolayer in about 2-3 weeks. Before transplantation surgery, these cells were dissociated by exposing them to a BSS solution without Mg ++ or Ca ++ containing 0.25% trypsin and i mM EDTA. The cells were taken up in a solution of MEM with 20% FCS to inactivate and wash out the trypsin. This solution was centrifuged at 800 RPM for 5 min; the supernatant was discarded and the pellet resuspended in MEM. About 5-10 gl containing approximately 105 cells was taken up into a micropipette for trans- plantation surgery (see below).
Surgery
Adult albino rabbits were anesthetized with an intravenous injec- tion of 20-30 mg/kg sodium pentobarbital with additional amounts given as needed. The pupil was dilated with 2% cyclopentolate and 2.5% neosynephrine and a 3 cc retrobulbar injection of 2% lidocaine was given to minimize eye movements. The eye was prop- tosed and held in position with a lid suture. A conjunctival flap was formed and a pars plana incision made with a stiletto blade.

469
Fig. 1. Photograph of the host albino rabbit fundus showing the site of the transplanted retinal pigmented epi- thelial cells that have spread in the subretinal space. The neural retina is reattached over the transplant site and there is not evidence of hemor- rhage or inflammation 2 weeks after transplantation. Two small satellite patches of transplants can also be seen below the main site
Fig. 2. Autoradiograph of pigmented retinN epithelial cell transplants in the subretinal space of the albino rabbit retina. The black grains pro- duced by 3H-thymidine are in the emulsion above the nuclei of three cells and demonstrate that the pig- mented cells are the transplanted cells
The dissociated epithelial cells were drawn into a glass micro-pi- pette filled with balanced salt solution (BSS). The pipette has an internal tip diameter of approximately 100 gm. The cell suspension was kept separate from the BSS by a small air bubble. An addition- al volume of 50 gl of clear BSS was drawn into the tip, also sepa- rated from the cell solution by an air bubble. The pipette was introduced through the pars plana incision and visualized in the vitreous cavity using a surgical microscope and a corneal contact lens. The pipette was guided to the neural retina several disc diame- ters from the optic nerve. A jet stream of clear BSS was ejected to produce a small retinal hole and bleb detachment. This often dislodged patches of retinal epithelial cells, a phenomenon more difficult to assess in the albinotic than in the pigmented retina [8]. With continued slow injection, the pigmented cell suspension was introduced into the subretinal space. Care was taken to keep the hole in the neural retina as small as possible and to avoid
seeding the vitreous with transplant cells as the pipette was with- drawn. The detachment volume was about 100-200 gl. The sclero- tomy site and conjunctivat flap were closed with 6-0 vicryl sutures.
Immunosuppression and tissue selection
In order to suppress any tissue rejection in these allogenic trans- plants, cyclosporine (15-25 mg/kg body wt, i.m./day) was admin- istered following surgery. These experiments involved 22 albino rabbits, but in only 14 was the transplantation surgery considered to be satisfactory enough to warrant detailed histological examina- tion. Factors that influenced this decision were either faulty injec- tion of the transplant cells, usually because a large fraction had entered the vitreous rather than the subretinal space or less fre- quently because of hemorrhage usually from the choroid and pro-

470
duced by excessive jet stream force. Earlier experiments on 97 pig- mented rabbits provided the background and experience that helped in evaluating what we considered to be a successful surgical procedure. Surgery was performed on one eye. Two animals were killed at 1 week; 4 at 2 weeks; 4 at 4 weeks and 1 at 2, 3, 4 and 5 months, respectively, after transplantation. All experiments adhered to the ARVO Resolution on the Use of Animals in Re- search. There was no evidence that this procedure caused discom- fort to the experimental animal or had a detrimental effect on vision in the operated eye.
Histology
The eyes were fixed in 3% glutaraldehyde in 0.05 M phosphate buffered saline at pH 7.2 for 24 h. The transplant site together with the surrounding retina was post-fixed in 1% osmic acid, dehy- drated with ethanol and embedded in epon. Sections were cut for autoradiography, dipped in Kodak NTB2 emulsion (Rochester, NY), and the coated slides were left in the dark at 4°C for 2 weeks. Selected blocks were trimmed and thin sections cut for ex- amination by electron microscopy.
Results
Figure 1 shows a pigmented patch of t ransplanted retinal epithelial cells in the fundus of an albino rabbit 2 weeks after surgery. Clusters of cells can be seen spread out into the subretinal space beneath of reattached neural retina. Typically, there was no evidence of hemorrhage or inf lammation in the retina or the ocular media so that the transplant site could be easily examined oph- thalmoscopically.
Figure 2 shows an autoradiogram of several trans- plant cells on Bruch's membrane in the host retina 2 weeks after surgery. The brownish pigmentat ion in these cells implies that they are the transplanted pigmented epithelial cells. The black grains in the emulsion over the nuclei of these cells confirm this conclusion.
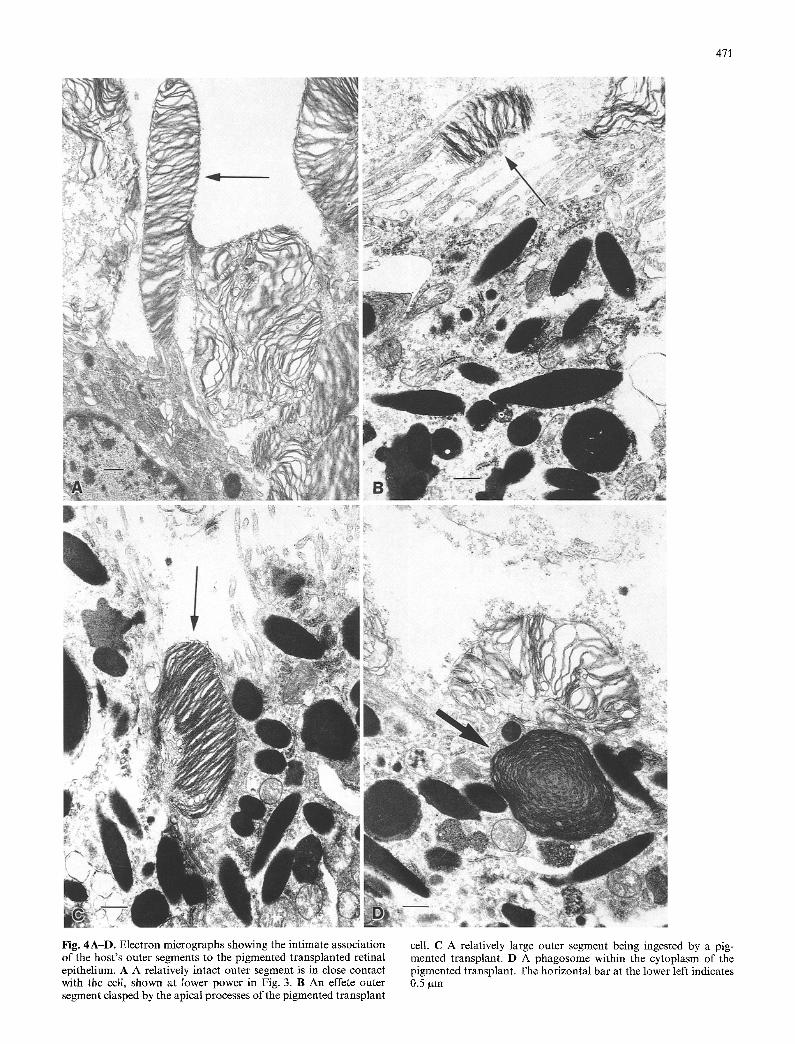
Figure 3 is an electron micrograph of the cell cluster labeled on the left side of Fig. 2. Serial sections through the labeled cells in the cluster are shown in Fig. 4 A - D . Close contact of a host outer segment with apical pro- cesses of the transplanted cell can be seen in Fig. 4A. The cell in contact with the outer segment (arrowed) is the same cell shown at lower magnification in the upper par t of Fig. 3 (labeled N). This cell has an abun- dance of melanin granules in its cytoplasm as well as a nucleus which is labelled by 3H-thymidine. The thin apical processes of the pigmented t ransplant cell in Fig. 4 B are enveloping an outer segment, and in Fig. 4 C a larger outer segment particle appears to be in the pro- cess of being internalized by the transplant cell. Figure 4D shows a phagosome within the cytoplasm of the pigmented transplant cell and another outer segment particle on the cell membrane. Since the transplant cell were in culture for 2-3 weeks before t ransplantat ion and therefore not in contact with outer segments for more than a month, the phagosome must be host outer-seg- ment material.
Figure 5 A - D illustrates several examples of trans- planted retinal epithelial cells on or close to Bruch's membrane f rom 2 4 weeks after transplantation. There
Fig. 3. Electron mlcrograph obtained by serlal sectlomng ot the labeled cells shown on the left side of Fig. 2. The nucleus labelled (N) is the same nucleus labeled by 3H-thymidine at the upper aspect of this cluster of cells. The horizontal bar (lower left) indicates 1 gm
is close apposit ion of many host outer segments to the apical membranes of the transplanted cells. This apposi- tion is not as intimate or as organized as in the normal retina but par t o f this may be due to t rauma f rom the bleb detachment. In general most o f the transplant cells were in contact with outer segments. Sometimes clumps of t ransplant cells are formed in which the internalized ones are not in contact with outer segments; a tendency for such clumping can be seen in Fig. 5 A.
There are also close associations between the pig- mented transplant cells and the unpigmented host epi- thelium. Albinotic host epithelial cells can be seen mak- ing contact with transplant cells at the lower left o f Fig. 5A-D. In all cases a thin extension of the host epi- thelium is interposed between the transplants and Bruch's membrane. This was a very common occurrence. In fact, we were unable to find an example of a trans- plant cell making direct contact with Bruch's membrane. We believe that this is probably due to the difficulty in removing the host epithelium using a one-shot jet s tream pulse that produces a bleb detachment and dis- lodgement of the host epithelial layer simultaneously.
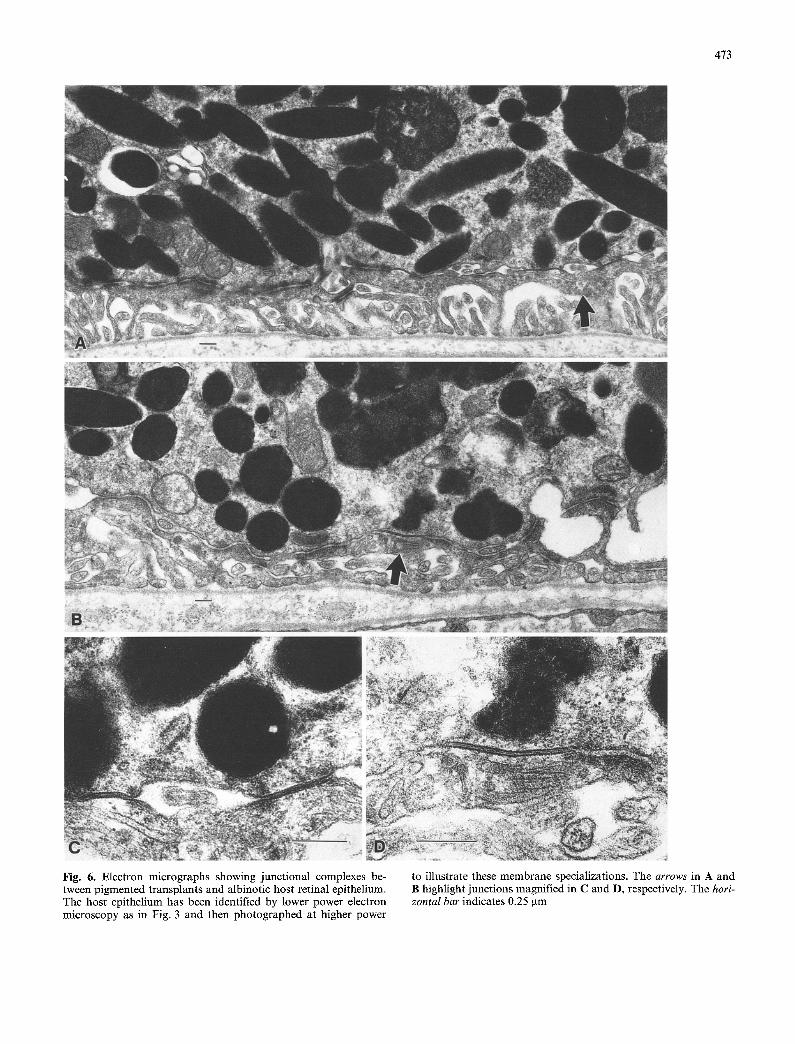
Higher power views of these processes interposed be- tween the transplant cells and Bruch's membrane are illustrated in Fig. 6A-D. These processes have been identified as belonging to the host epithelium by low-
471
Fig. 4A-D. Electron micrographs showing the intimate association of the host's outer segments to the pigmented transplanted retinal epithelium. A A relatively intact outer segment is in close contact with the cell, shown at lower power in Fig. 3. B An effete outer segment clasped by the apical processes of the pigmented transplant
cell. C A relatively large outer segment being ingested by a pig- mented transplant. D A phagosome within the cytoplasm of the pigmented transplant. The horizontal bar at the lower left indicates 0.5 gm

472
Fig. 5A-D. Electron micrographs of transplanted pigmented retinal epithelium at 2 (A, B) and 4 (C, D) weeks after transplantation. The transplants are in close apposition to both the host's outer segments and Bruch's membrane. The albinotic retinal epithelium of the host can be distinguished from the pigmented transplants by the absence of melanin granules in their cytoplasm. There are
some small pigmented lysosomal granules present in the albinotic cells but the dense, often elliptical melanin granules are absent, (see D in particular). In all cases the transplant cells are not making direct contact with Bruch's membrane but are separated from it by relatively thin basal processes of the host retinal epithelium. The horizontal bar indicates 1 Ixm
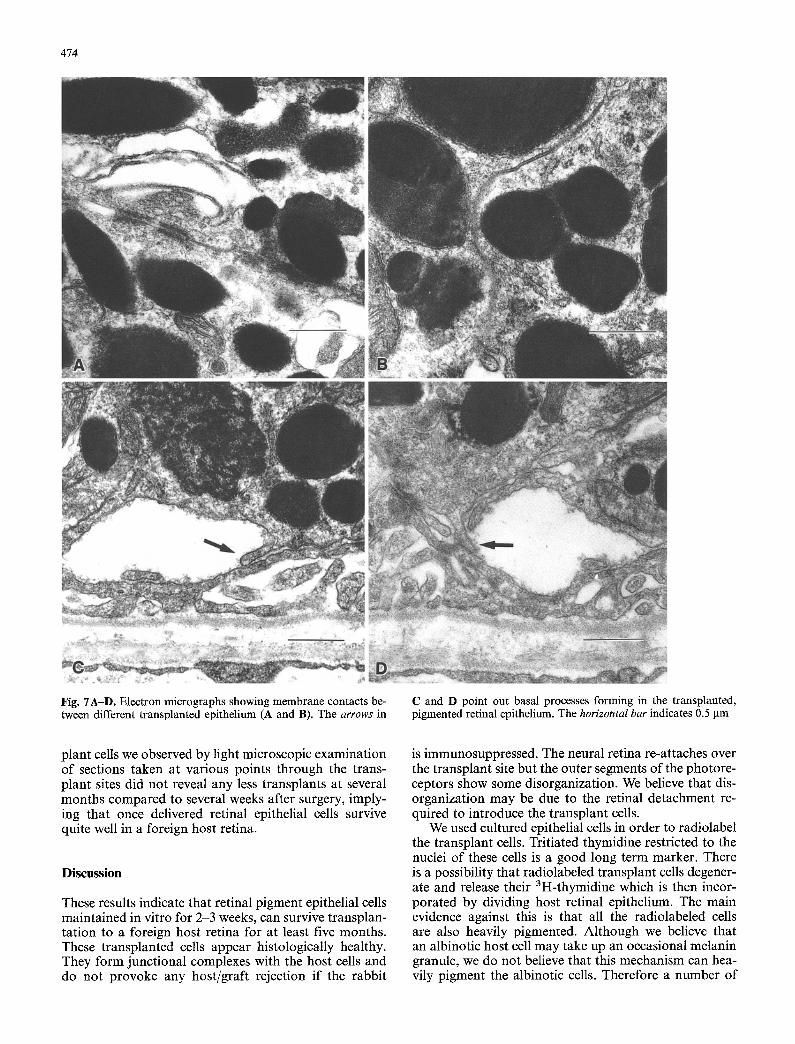
power electron microscopy as in Fig. 5 A - D and then photographed at higher power to show the desmosomal and gap junctions (arrowed) that form between the host and t ransplant epithelial cells. Magnified examples of several o f these contacts are shown in Fig. 6 C and D. It was more difficult to find junctional complexes be- tween t ransplant cells, probably because there is a greater chance for a t ransplant cell to come in contact with host than with fellow transplant cells. Two exam- ples of junctional complexes occurring between trans- plant cells are illustrated in Fig. 7A and B.
The lack of contacts between any of the transplant cells and Bruch's membrane may be responsible for the
fact that the basal surface of all the transplant cells we examined were not convoluted. Extensive convolutions of the basal surface is a striking and general characteris- tic of retinal epithelium. Several suggestions of abortive basal infoldings were observed and these are illustrated in Fig. 7 C and D.
We have not observed any significant difference in the morphological appearance of the transplant cells with time up to 5 months after t ransplantat ion or any evidence of cell division. The transplanted epithelium retains a considerable amount of melanin and does not show the pleomorphic appearance associated with rela- tively rapid cell division. The relative number of trans-
473
Fig. 6. Electron micrographs showing junctional complexes be- tween pigmented transplants and albinotic host retinal epithelium. The host epithelium has been identified by lower power electron microscopy as in Fig. 3 and then photographed at higher power
to illustrate these membrane specializations. The arrows in A and B highlight junctions magnified in C and D, respectively. The hori- zontal bar indicates 0.25 gm
474
Fig. 7A-D. Electron micrographs showing membrane contacts be- tween different transplanted epithelium (A and B). The arrows in
C and D point out basal processes forming in the transplanted, pigmented retinal epithelium. The horizontal bar indicates 0.5 gm
plant cells we observed by light microscopic examination of sections taken at various points through the trans- plant sites did not reveal any less transplants at several months compared to several weeks after surgery, imply- ing that once delivered retinal epithelial cells survive quite well in a foreign host retina.
Discussion
These results indicate that retinal pigment epithelial cells maintained in vitro for 2-3 weeks, can survive transplan- tation to a foreign host retina for at least five months. These transplanted cells appear histologically healthy. They form junctional complexes with the host cells and do not provoke any host/graft rejection if the rabbit
is immunosuppressed. The neural retina re-attaches over the transplant site but the outer segments of the photore- ceptors show some disorganization. We believe that dis- organization may be due to the retinal detachment re- quired to introduce the transplant cells.
We used cultured epithelial cells in order to radiolabel the transplant cells. Tritiated thymidine restricted to the nuclei of these cells is a good long term marker. There is a possibility that radiolabeled transplant cells degener- ate and release their 3H-thymidine which is then incor- porated by dividing host retinal epithelium. The main evidence against this is that all the radiolabeled cells are also heavily pigmented. Although we believe that an albinotic host cell may take up an occasional melanin granule, we do not believe that this mechanism can hea- vily pigment the albinotic cells. Therefore a number of

475
unique events must occur. The host epithelial cells must not only incorporate 3H-thymidine and enter into cell division but also completely fill its cytoplasm with me- lanin while all its neighbors remain albinotic. This seems highly unlikely. There is also no evidence for any signifi- cant cell division among the host retinal epithelial cells. In addition the concentrations of 3H-thymidine resulting f rom the breakdown of t ransplant cells would be ex- pected to be very low and variable leading to weak and variable labeling of the labeled cells, which is not the case. The labeling is relatively intense and similar in the labeled cells. Recently several other laboratories have reported other labels such as carbon particles [2] or the fluorescent marker , 5 bromodeoxyuridine [10]. The lat- ter has the advantage of being a nuclear label which is less prone to ambiguity by host cell phagocytosis and in this sense similar to 3H-thymidine. It is, however, less stringent than tritiated thymidine which requires in addition, mitosis, for its incorporat ion into the nucleus.
Retinal cell t ransplantat ion has been shown to slow the progress of the retinal degeneration in the Royal College of Surgeon (RCS) strain of rats [5, 7, 9], where there is a known genetic defect in the retinal epithelium. It is conceivable that retinal epithelial cell transplanta- tion could also influence some hereditary retinal dystro- phies in man in which retinal epithelium may be defec- tive such as age related macular degeneration or Star- gardt 's disease. The results of the present study indicate that such transplants can survive and function for rela- tively long periods of time in mammal ian retina without leading to inf lammation or tumorogenesis.
References
1. Brittis M, Gouras P, Lopez R, Sullivan B, Kjeldbye H (1988) Retinal epithelial allografts facilitated by cyclosporine. Invest Ophthalmol Vis Sci 29 [Suppl] :241
2. E1 Dirini AA, Wang H, Ogden TE, Ryan SJ (1992) Retinal pigment epithelium implantation in the rabbit: technique and morphology. Invest Ophthalmol Vis Sci 32:668
3. Flood MT, Gouras P, Kjeldbye H (1980) Growth characteris- tics and ultrastructure of human retinal pigment epithelium in vitro. Invest Ophthalmol Vis Sci 19:1309
4. Gouras P, Flood MT, Kjeldbye H, Bilek MK, Eggers H (1985) Transplantation of cultured human retinal epithelium to Bruch's membrane of the owl monkey's eye. Curr Eye Res 4: 253-265
5. Gouras P, Lopez R, Brittis M, Kjeldbye H, Sullivan B (1988) The experimental route to curing a rat retinal dystrophy by transplantation. 5th International Retinitis Pigmentosa Con- gress Proceedings, pp 267-276
6. Gouras P, Lopez R, Du J, Gelanze M, Kwun R, Brittis M, Kjeldbye H (1990) Transplantation of retinal cells. Neurooph- thalmol 10:165-176
7. Li L, Turner JE (1988) Inherited retinal dystrophy in the RCS rat: photoreceptor cell rescue by RPE cell transplantation. Exp Eye Res 47:911
8. Lopez R, Gouras P, Brittis M, Kjeldbye H (1987) Transplanta- tion of cultured rabbit retinal epithelium to rabbit retina using a closed eye method. Invest Ophthalmol Vis Sci 28:1131
9. Lopez R, Gouras P, Kjeldbye H, Sullivan B, Reppucci V, Brittis M, Wapner F, Goluboff E (1989) Transplanted retinal pigment epithelium modifies the retinal degeneration in the RCS rat. Invest Ophthalmol Vis Sci 30 : 586-588
10. Young RW, Gaur VP, Li L, Turner JE (1991) Transplantation of 5-bromodeoxyuridine labeled retinal pigmented epithelial cells on Bruch's membrane in the rabbit. Invest Ophthalmol Vis Sci 32: 982
Copyright © 2022 FDOKUMEN




















