
Aminobisphosphonates Cause Osteoblast Apoptosis and Inhibit Bone Nodule Formation In Vitro

Protoplasma (1986) 132:6%75 PROTOPLASMA �9 by Springer-Verlag 1986
Soybean Root Nodule Ultrastructure During Dark-Induced Stress and Recovery 1
HILLEL P. COHEN 2, GAUTAM SARATH 3, KIT LEE 3, and FRED W. WAGNER *
Department of Agricultural Biochemistry, East Campus, University of Nebraska-Lincoln
Received January 16, 1986 Accepted March 6, 1986
Summary
Root-nodules of soybean plants dark-stressed for 8 days and then allowed to recover for up to 17 days were examined by transmission electron microscopy. Control nodules possessed all the ultrastruc- rural features characteristic of infected and uninfected nodule cells. Minimal changes in the appearance of host cells and bacteroids occurred during the first four days of dark stress. After 8 days of dark stress, damage was observed in the cellular and organelle membranes; however, very few changes were observed in the bacteroids. Nodule structure continued to degrade during the first two days of recovery after which time nodules either recovered or completely degraded. In the former case, structural integrity returned to all nodule cells. In the ]latter case all structural integrity of the host cell disappeared; however, bacteroids appeared intact suggesting that they remained 'viable.
Keywords: Dark-stress; Recovery; Root-nodules; Soybean; Ultrastructure.
1. Introduction
Ultrastructurally, soybean nodule senescence begins at
numerous sites within the nodule (CHEN and THORNTON
1940) and the infected cells undergo several structural
changes. Membrane components , including the
plasma, organellar and peribacteroid membranes, de-
generate. The rough endoplasmic reticulum declines in
1 Published as Paper no. 7974, Journal Series, Nebraska Agriculture Research Division. 2 Present address: Hoffmann-La Roche Inc., Nutley, NJ 07110. 3 SchoolofBiologicalSciences, UniversityofNebraska-Lincoln, NE 68583. * Correspondence and Reprints: Department of Agricultural Bio- chemistry, University of Nebraska-Lincoln, Lincoln, NE 68583, U.S.A.
quant i ty and becomes fragmentary. Fewer r ibosomes
and Golgi bodies are present. Eventually, cellular
organelles degenerate and cannot be recognized ultra-
structurally (Tu 1975, KIJNE and PLANQUE 1979). Poly- 13-hydroxybutyrate granules disappear within bacteroids,
an event that correlates to a decline in levels o f poly-[3- hydroxybutyrate oxidoreductase in root nodule bacteroids
f rom naturally senescing plants (STRIPF and WERNER
1978). However, bacteroids do not appear to degenerate,
but remain constant in size and shape (Tu 1975, PAAU
and COWLES 1979) and probably remain viable.
Various types of stress induce premature roo t nodule
senescence. These include drought , heat, waterlogging, addit ion o f mineral nitrogen, and sudden reduct ion o f
photosyntha te supply by decapitat ion or by subjecting
the shoots to prolonged periods o f darkness (NEwCOMB 1981, VANCE et aL 1979). Earlier work f rom this labora-
tory (PFEIFFER e t a l . 1983) proposed that dark-stress induced premature senescence in soybean nodules was
characterized by biochemical changes which resembled those o f natural senescence. Thus it was o f interest to
determine whether the structural changes in these
nodules were similar to those reported to occur in
naturally senescing nodules.
2. Materials and Methods
2.1. Plant Material
Soybean seeds [Glycine max (L) Merr., cv. Woodworth] were inoculated with Rhizobium japonicum (strain 61A89) at the time of planting. Growth conditions were identical to those reported previ- ously (PFEIFFER et al. 1983) except that the light intensity averaged

70 H.P. COHEN etal.: Soybean Root Nodule Ultrastructure During Dark-Induced Stress and Recovery
450 gmol. m -2. s -1 (photosynthetically active radiation, 400- 700rim) Plants used for dark-induced senescence were in the R-I stage of reproductive growth (early flowering) (FEIqR etal. 1971). Plants were stressed by placing them in total darkness for eight days. After being dark stressed, plants were allowed to recover for up to 17 days on a 16-hour light, 8-hour dark regimen. Control plants were grown under a 16-hour light/8-hour dark regimen for 12 and 24 days after the experimental plants were placed in dark.
2.2. Selection of Root Nodules
Root nodules from three plants were harvested at each sampling date and placed on ice. Only nodules greater than 2 mm in diameter were harvested (PFEIFFER etal. 1983). Several nodules were selected at random at each sampling date and sliced directly into fixative for electron microscopy. Less than 30 minutes elapsed from the harvest of the first nodule to the completion of slicing and fixation.
2.3. Electron Microscopy
Nodules were processed for electron microscopy as described by BAIRD and WEBSTER (1982). Specimens were sectioned with a diamond knife on an LKB Ultratome III, stained for 30 minutes in 2% (w/v) aqueous uranyl acetate followed by 30 minutes in basic lead citrate (REYNOLDS 1963), and viewed in a Philips 201 electron microscope operating at 60 kV.
3. Results
3.1. Control Plants
Fresh sections of nodules from control plants were always red in color. Infected cells of control plants possessed dense cytoplasm containing little endo- plasmic reticulum, or Golgi bodies (Fig. 1). Numerous mitochondria occurred at the periphery of the infected cells. Plastids were commonly observed among the mitochondria. Plastids of infected cells rarely contained starch grains. Bacteroids were always enclosed within peribacteroid membranes and were round, oval, or rod shaped. Bacteroids contained a well defined nucleoid and some fibrillar material. Some dark granules, prob- ably polyphosphate deposits (WEe, NZR and MORSCHBI~ 1978), occurred near the center of the bacteroids. In addition, bacteroids contained electron lucent granules of poly-[3-hydroxybutyrate (Fig. 1). Uninfected cells had peroxisomes and a large central vacuole. Plastids generally possessed large starch grains (Fig. 1). The ultrastructural morphology of the control nodules in the 12 day light control and the 24 day light control was not appreciably different.
3.2. Dark Treatment
Nodules from plants kept in darkness for four days had few signs of ultrastructural degradation, and resembled nodules from control plants. After eight days of dark- stress, fresh sections of nodules were a green color.
Ultrastructurally, in the infected cells, some breakdown of peribacteroid membranes was observed. Lysis of the cytoplasm was evident by the appearance of extensive vacuolation of infected cells (Fig. 2). However, the bacteroids appeared to be intact, although they seemed to lack readily observable deposits of poly-13- hydroxybutyrate. Major changes were not evident in the uninfected cells except the cytoplasm was less dense
than that of control nodules (Fig. 2).
3.3. Nodule Recovery
Infected cells in nodules from plants placed in light for two days after dark stress were ultrastructurally similar to infected cells from nodules of eight-day dark-stressed plants. Occasional breaks were seen in the tonoplast of
the uninfected cells. Recovery of the structural integrity of infected cells was first detected after six days of normal photoperiod following the dark treatment. Peribacteroid mem-
branes completely surrounded almost all bacteroids. Bacteroids contained abundant fibrillar material as well as granules of poly-}-hydroxybutyrate (Fig. 3). After seventeen days of recovery, nodule ultrastructure was similar to that of control plants. All bacteroids were enclosed by peribacteroid membranes and showed abundant deposits ofpoly-[3-hydroxybutyrate. Numer- ous mitochondria occurred in the periphery of the infected cells. The uninfected cells contained well defined peroxisomes and plastids (Fig. 4).
3.4. Nodule Senescence
Some of the root nodules did not recover from dark stress. When sectioned, the interior of these nodules varied in color from light brown to dark green. After six days of a normal light regimen following stress, these nodules possessed small vesicles in the cytoplasm and in the nuclear matrix (Fig. 5). Bacteroid structure was
essentially similar to that of dark-stressed nodules (Fig. 2). With time, cellular disorganization became more pronounced in these nodules. There were breaks in virtually all peribacteroid membranes. The infected cells became flocculent and sparse. Lysis of the tono- plast of uninfected cells was apparent (Fig. 6). After seventeen days recovery, these nodules were soft and had a green color internally. While it was not possible to identify cellular organelles on the basis of their ultrastructural characteristics in either the in- fected or uninfected cells, bacteroids retained their structural integrity; however, it was difficult to distin- guish any deposits of poly-[3-hydroxybutyrate (Fig. 7).

Fig. i. Appearance of infected cells (IC) and uninfected cells (UC) at the start of the experimental protocol. Note the bacteroids (b) enclosed by intact peribacteroid membranes, and the absence of vacuoles in the infected cells, cw cell wall, g Golgi bodies, is intracellular space, m mitochondria~ pH poly-f3-hydroxybutyrate granules, PL plastid, Ps peroxisome, Vvacuole. x 10,000. Bar = 1.0 ~tm
Fig. 2. Section of a root nodule after 8 days of dark-stress, Note the extensive vacuolation due to lysis of the infected cell, and breaks in the peribacteroid membranes (solid arrows). Adjacent uninfected cells show less drastic effects. Note the sparser cytoplasm. Nu nucleus, other symbols as in Fig. 1. x 6,700. Bar = 1.0 gm
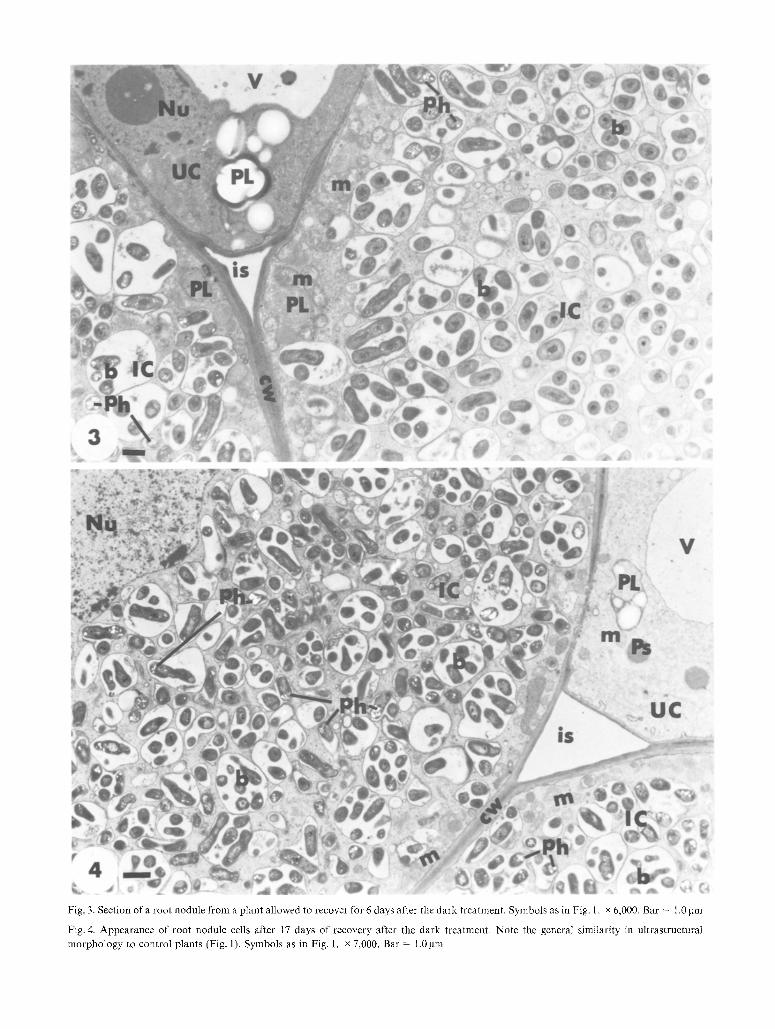
Fig. 3. Section of a root nodule from a plant allowed to recover for 6 days after the dark treatment. Symbols as in Fig. 1. x 6,000. Bar = !.0 gm
Fig.4. Appearance of root nodule cells after 17 days of recovery after the dark treatment. Note the general similarity in ultrastrnctural
morphology to control plants (Fig. 1). Symbols as in Fig. 1. x 7,000. Bar = 1.0 gm
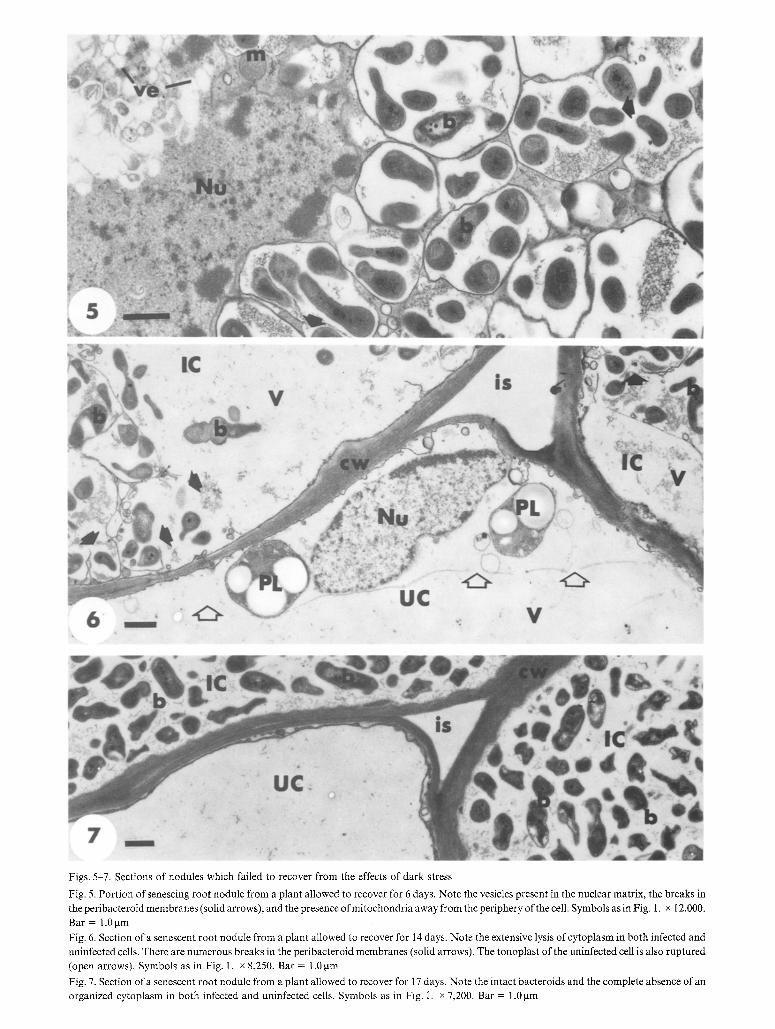
Figs. 5-7. Sections of nodules which failed to recover from the effects of dark stress
Fig. 5. Portion of senescing root nodule from a plant allowed to recover for 6 days. Note the vesicles present in the nuclear matrix, the breaks in the peribacteroid membranes (solid arrows), and the presence ofmitochondria away from the periphery of the cell. Symbols as in Fig. 1. x 12,000.
Bar = 1.0 gm
Fig. 6. Section of a senescent root nodule from a plant allowed to recover for 14 days. Note the extensive lysis of cytoplasm in both infected and
uninfected cells. There are numerous breaks in the peribacteroid membranes (solid arrows). The tonoplast of the uninfected cell is also ruptured
(open arrows). Symbols as in Fig. 1. x 8,250. Bar = 1.0 gm
Fig. 7. Section of a senescent root nodule from a plant allowed to recover for 17 days. Note the intact bacteroids and the complete absence of an organized cytoplasm in both infected and uninfected cells. Symbols as in Fig. i. x 7,200. Bar = 1.0 gm

74 H.P. COHEN et at.: Soybean Root Nodule Ultrastructure During Dark-Induced Stress and Recovery
4. Discussion
The ultrastructure of dark-stressed nodules is similar to the early stages of natural nodule senescence of soybean (Tu 1975, KIJNE and PLANQUE 1979), and Phaseolus
vulgaris, another species with determinate nodules (BAIRD and WEBSTER 1982). The earliest ultrastructural changes observed in dark stressed root nodules are breaks in peribacteroid membranes, the formation of numerous small vesicles, and the development of irregularities in bacteroid shape (Figs. 5-6 A). WERNER etal. (1984) have shown that the stability of the peribacteroid membrane is due to the interaction of the host cell and the bacteroid(s). In ineffective soybean symbioses, breakdown of the peribacteroid membrane leads to premature senescence (BASSETT etal. 1977, WEANER el al. 1984, MELLOR et aI. 1985, WEANER et al.
1985). In Phaseolus vulgaris, degeneration of the peri- bacteroid membrane is also an early event associated with nodule senescence (BAIRD and WEBSTER 1982). As nodule degradation continued, numerous vesicles were detected in the cytosol and within cellular organelles of infected cells. Similar vesicle formation was detected in senescing infected cells of pea and alfalfa nodules (TRUCHET and COULOMB 1973, VANCE etal. 1980). TRUCHET and COULOMn (1973) have termed these vesicles "phytolysosomes", and suggest that they play a lyric role in nodule senescence. The fact that bacteroids of soybean nodules did not degrade during dark-induced senescence, coupled with the evidence that Rhizobium bacteroids of all nodule ages retain their ability to form colonies in culture, (GRESSHOFF and ROLFE 1978, TSIEN et al. 1977), and the presence of structurally intact bacteroids in senescent nodules (Figs. 5-7) support the claim that the Rhizobium bacteroids can be released from the decaying nodule into the soil (REYES and SCHMIDT 1979). Imposition of dark stress on healthy nodulated soybean plants causes root nodules to undergo premature senescence (PFEIFFER et al. 1983). Dark treatment also causes a decline in nodule levels of poly-13- hydroxybutyrate (WONG and EVANS 1971). These physiological changes were detected after four days of dark stress, before any morphological changes were observed. When ultrastructural changes could be de- tected after eight days of dark stress (Fig. 2), soybean plants were no longer capable of reducing acetylene and showed extensive loss of plant cytosolic proteins (PFEIFFER etal. 1983). Similar physiological changes have been detected in naturally and stress-induced senescing root nodules of many plants (SUTTON 1983). In their work, PEEIFFER et al. (1983) demonstrated that
recovery of acetylene-reducing capacity of dark- stressed plants resulted from the recovery of existing nodules rather than from renodulation. Thus, recovery of physiological and biochemical function after dark- stress (PFEIFFER et aI. 1983), correlates temporally to the recovery of infected cell morphology shown in this report. Recovery from stress-induced senescence has also been reported for root nodules of alfalfa, birdsfoot trefoil, and clover (MouSTAFA etal. 1969, VANCE etal. 1979, VANCE etal. 1980, CRALLE and HEICHEL 198t). However, root nodules of these three species have persistent meristems (DART 1977), and recovery is due to the formation of new infected cells by the continued growth of the meristematic region (VANCE et al. 1980). In contrast, mature soybean root nodules lack meris- tematic cells (DART 1977). Furthermore, we did not observed any root nodule cell during recovery that had meristematic characteristics. Our data also indicate that a small subset of root- nodules is unable to recover from the effects of dark- stress, in accord with earlier work on this system (PFEIFFER etal. 1983). Results of this ultrastructural study provide evidence that dark-stress induces a set of conditions in soybean root nodules that mimics natural senescence. Further, our data also show that the initial morphological changes induced by dark-stress can be reversed within individual cells of soybean root nodules and that bacteroids remain stable during stress and subsequent recovery. Our results suggest that the regulation of soybean root nodule recovery from stress, and the regulation of premature nodule senescence induced by stress, is coordinated at the whole nodule level.
Acknowledgements
We thank Dr. L. M. BAIRD for many useful suggestions and criticisms during the course of this work. We also thank Dr. W. LANGENBERG for the use of dark-room facilities. This work was supported in part by the U.S. Department of AgricuIture under Grant no. 83-CRCR-1- 1293 and a grant from the Nebraska Soybean Development, Utiliza- tion and Marketing Board.
References
BAIRD LM, WEBSTER BD (1982) Morphogenesis of effective and ineffective root nodules in Phaseolus vulgaris L. Bot Gaz 143: 41- 51
BASSETT B, GOODMAN RN, NOVACKY A (1977) Ultrastructure of soybean nodules. II. Deterioration of the symbiosis in ineffective nodules. Can J Microbiol 23:873-883

H. P. COHEN et al.: Soybean Root Nodule Ultrastructure During Dark-Induced Stress and Recovery "75
CHEN HK, THORNTON HG (1940) The structure of "ineffective nodules" and its influence on nitrogen fixation. Proc Roy Soc London B 129:208-229
CRALLE HT, HEICHEL GH (1981) Nitrogen fixation and vegetative regrowth of alfalfa and birdsfoot trefoil after successive harvests or floral debudding. Plant Physiol 67:898-905
DART PJ (1977) Infection and development in leguminous nodules. In: HARDY RWF (ed) A treatise on dinitrogen fixation. Wiley Interscience, New York, pp 367-472
FEHR WR, CAVINESS CE, BURMOOD DT, PENNINGTON JS (1971) Stage of development descripitions for soybeans, Glycine max (L.) Merrill Crop Sci 11:929-931
GRESSHOFF PM, ROLFE BG (I978) Viability ofRhizobium bacteroids isolated from soybean nodule protoplasts. Planta 142:329-333
K_IJNE JW, PLANQUE K (1979) Ultrastructural study of the endomem- brahe system in infected cells of pea and soybean root nodules. Physiol Plant Path 14:339-345
MELLOR RB, CHRISTENSEN TMEE, BASSARAB S, WERNER D (1985) Phospholipid transfer from ER to the peribacteroid membrane in soybean nodules. Z Naturforsch 40 C: 73-79
MOUSTAFA E, BALL R, F~ELD RRO (1969) The use of acetylene reduction to study the effect of nitrogen fertilizer and defoliation on nitrogen fixation by field-grown white clover. N Z J Agric Res 12:691-696
NEWCOMB W (1981) Nodule morphogenesis and differentiation. In: GILES KL, ATHERLY AG (eds) Int Rev Cytol Supp 1: Biology of the Rhizobacea. Academic Press, New York, pp 247-297
PAAU AS, COWLES JR (1979) Effect of induced nodule senescence on parameters related to dinitrogen fixation, bacteroid size, and nucleic acid content. J Gen Microbiol 111:101-107
PEIFFER NE, MALIK NSA, WAGNER FW (1983) Reversible dark- induced senescence of soybean root nodules. Plant Physiol 71: 393-399
REYES VG, SCHMIDT EL (1979) Population densities of Rhizobium japonicum strain 123 estimated directly in soil and rhizospheres, Appl Env Microbiol 37:854-858
REYNOLDS ES (1963) The use of lead citrate at high pH as an electron- opaque stain in electron microscopy. J Cell Biol 17:108
STR1PF R, WERNER D (1978) Differentiation of Rhizobiumjaponicum. II. Enzymatic activities in bacteroids and plant cytoplasm during development of nodules of Glyeine max. Z Naturforsch 33 C: 373-381
SUTTON WD (1983) Nodule development and senescence. In: BROUGHTON WJ (ed) Nitrogen Fixation, Vol 3, Legumes. Cla- rendon Press, Oxford, pp 144-212
TRUCHET G, COULOMB P (1973) Mise en 6vidence et evolution du syst6me phytolysosomal dans les cellules des diff6rentes zones de nodules radiculaires de pois (Pisum sativum L.). Notion d'h&6rophagie. J Ultrastruct Res 43:36-57
TSIEN HC, CAIN PS, SCHMIDT EL (1977) Viability of Rhizobium bacteroids. App Env Microbiol 34:854-856
Tu JC (1975) Rhizobial root nodules of soybean as revealed by scanning an transmission electron microscopy. Phytopath 65: 447-454
VANCE CP, HEICHEL GH, BARNES DK, BRYAN JW, JOHNSON LE (1979) Nitrogen fixation, nodule development, and vegetative regrowth of alfalfa (Medicago sativa L.) following harvest. Plant Physiol 64:1-8
- - JOHNSON LEB, HALVERSEN AM, HEICHEL GH, BARNES I)K (1980) Histological and ultrastructural observations of Medic~go sativa root nodule senescence after foliage removal. Can J Bot 58: 295-309
WERNER D, MELLOR RB, HAHN MG, GRISEBACH H (1985) Soybean root response to symbiotic infection. Glyceollin accumulation in an ineffective type of soybean nodules with an early loss of the peribacteroid membrane. Z Naturforsch 40 C: 179-181
- - MORSCHEL E (1978) Differentiation of nodules of Glycine max: Ultrastructural studies of plant cells and bacteroids. Planta i[41: 169-177
- - - - KORT R, MELLOR RB, BASSARAB S (1984) Lysis ofbacteroids in the vicinity of the host cell nucleus in an ineffective (fix--) root nodule of soybean (Glycine max). Planta 162:8-16
WONG PP, EVANS HJ (1971) Poly-l~-hydroxybutyrate utilization by soybean (Glycine max Merr) Nodule and assessment of its role in maintance of nitrogenase activity. Plant Physiol 47:750-755
Copyright © 2022 FDOKUMEN




















