
The urokinase plasminogen activator system as a target for prognostic studies in breast cancer
Upload
independentCategory
view
2download
0
2000;60:5839-5847. Cancer Res Linda S. Gutierrez, Alexis Schulman, Teresa Brito-Robinson, et al. Plasminogen Activator Inhibitor-1Urokinase-Type Plasminogen Activator or Its Inhibitor, Tumor Development Is Retarded in Mice Lacking the Gene for
Updated version
http://cancerres.aacrjournals.org/content/60/20/5839
Access the most recent version of this article at:
Cited Articles
http://cancerres.aacrjournals.org/content/60/20/5839.full.html#ref-list-1
This article cites by 43 articles, 16 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/60/20/5839.full.html#related-urls
This article has been cited by 37 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

[CANCER RESEARCH 60, 5839–5847, October 15, 2000]
Tumor Development Is Retarded in Mice Lacking the Gene for Urokinase-TypePlasminogen Activator or Its Inhibitor, Plasminogen Activator Inhibitor-1 1
Linda S. Gutierrez, Alexis Schulman, Teresa Brito-Robinson, Francisco Noria, Victoria A. Ploplis, andFrancis J. Castellino2
Walther Cancer Center, W. M. Keck Center for Transgene Research, and Department of Chemistry and Biochemistry, University of Notre Dame, Notre Dame, Indiana 46556
ABSTRACT
In vivo tumor progression in mice with targeted deficiencies in uroki-nase-type plasminogen activator(UPA2/2) and its inhibitor, plasminogenactivator inhibitor-1 (PAI-12/2), was studied using a fibrosarcoma tumormodel. Murine T241 fibrosarcoma cells were s.c. implanted into threegroups of mice with the following genotypes, wild-type (WT),UPA2/2, andPAI-12/2. A significantly diminished primary tumor growth in UPA2/2
and PAI-12/2 mice occurred, relative toWT mice. Tumors in UPA2/2 andPAI-12/2 mice displayed lower proliferative and higher apoptotic indicesand displayed a different neovascular morphology, as compared withWTmice. These results are consistent with the decreased growth rates of thistumor in these gene-deleted mice. Immunohistochemical analyses of thetumors revealed a decrease in vascularity and vascular endothelial growthfactor expression only in tumors inPAI-12/2 mice. Analyses of the relativeextents of corneal angiogenesis in these same animals, induced by basicfibroblast growth factor, corroborated the resistance ofPAI-12/2 mice toneovascularization. The results obtained suggest that the host fibrinolyticsystem plays an important role in tumor growth in this model. Alterationsin host expression of components of this system may alter tumor growthand dissemination by affecting the balance between tumor cell death andproliferation, as well as extracellular matrix changes needed for invasive-ness and angiogenesis.
INTRODUCTION
Activation of the fibrinolytic system results from the proteolyticconversion of plasma Pg3 to the serine protease, Pm. uPA, itself aserine protease, is one of the enzymes that catalyzes this process viacleavage of the Arg561-Val562peptide bond in Pg. PAI-1 is one naturalserpin-type inhibitor of this process that functions through its abilityto directly inhibit uPA (1). The localization of Pm activity at the siteof a developing thrombus, as well as the short-term protection of Pmand Pg activators from circulating inhibitors, are critical events thatregulate the rate of clot dissolution (2).
In addition to their roles in clot dissolution, components of thefibrinolytic system, including Pg, uPA, uPAR, PAI-1, and tPA havebeen shown to be important for other processes, such as ECM deg-radation, in some cases via activation of MMPs (3), wound healing(4), atherosclerosis (5), angiogenesis (6), and cancer (7). In this lattercase, observations that neoplastic cells express elevated levels of uPA,tPA, PAI-1, and uPAR suggest that these proteins may be involved in
tumor growth, metastasis, and angiogenesis (8–11). Angiogenesis-dependent tumor progression (12) has also been associated withinteraction of integrins with uPA and uPAR, thus linking cellularproteolysis and ECM interactions (13). Confirming the importance offibrinolytic components and the cancer phenotype, numerous clinicalstudies have demonstrated that high levels of uPA (14–17) and PAI-1(18, 19) are associated with a poorer prognostic outcome in certainhuman cancers, and recent data show a similar correlate with thereceptor for uPA, uPAR (20–22).
On a molecular level, uPA/uPAR complex formation on the surfaceof tumor cells can focus Pm-catalyzed proteolysis, which in turn canfacilitate tumor cell invasion and metastasis by either directly degrad-ing ECM proteins or indirectly performing this same function viaactivation of MMPs. Immunocytochemical studies of primary tumorand metastasis of Lewis lung carcinoma cells using uPA antibodieshave demonstrated intense immunoreactivity at the sites of invasivegrowth of the tumor and degradation of normal tissue (23).
Thus, although there appears to be a correlation between enhancedexpression of components of the fibrinolytic system and tumor growthand metastasis, the specific mechanisms that underlie the contribu-tions of this system to neoplasia are unclear. In an attempt to illumi-nate some of these issues, the current study used a fibrosarcoma modelin WT, UPA2/2, andPAI-12/2 C57BL/6J mice to directly assess therole of host expression of these proteins in tumorigenesis. The choiceof this tumor model was governed by its syngenicity with, and rapidgrowth characteristics in, the host C57BL/6J mice; its high degree ofvascularization; its expression of high levels of the angiogenic factor,VEGF; and its predictable route of metastasis. A summary of theresults of this investigation is presented herein.
MATERIALS AND METHODS
Mice. UPA2/2, PAI-12/2, andWTmice, 5–6 weeks of age, in a C57BL/6Jgenetic background (at least eight times back-crossed into this strain), wereused for this study. All animals were maintained in accordance with the NIHguide for the care and use of laboratory animals, and experimental protocolswere approved by the Institutional Animal Care and Use Committee.
Cell Culture. The C57BL/6J-derived murine fibrosarcoma tumor cell line,T241, was grown in McCoy’s modified withL-glutamine (RPMI 16290)containing essential vitamins, 10% fetal bovine serum (Life Technologies,Rockville, MD), and antibiotics. These cells were maintained in a humidifiedincubator at 37°C, 5% CO2.
Implantation of Tumor Cells in Mice. At confluency, adherent cells weredetached by incubation with 0.25% trypsin (Hyclone, Rockville, MD). Thecells were centrifuged and then resuspended in a sterile solution of PBS at afinal concentration of about 1.03 107 cells/ml. A 100-ml aliquot of resus-pended cells (about 1.03 106 cells) was injected s.c. between the shoulderblades;3 cm from the tail. Visible and small palpable tumors were measuredusing a digital caliper (MyCal; Thomas Scientific, Swedesboro, NJ). Tumorvolumes were calculated using the formula: (width2)(length)(0.52), as de-scribed (24).
RT-PCR. Total RNA was isolated from tumors implanted inWT, UPA2/2,andPAI-12/2 mice and from controlWTlivers, kidneys, intestines, spleen, andtestes. Additionally, total RNA was obtained from T241 fibrosarcoma cellsgrown in culture. The Ultraspec RNA isolation system (Biotech, Houston, TX)was used in each case for the first strand cDNA synthesis, which was carried
Received 5/1/00; accepted 8/29/00.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 This work was supported in part by NIH Grant HL13423 (to F. J. C.) and GrantHL63682 (to V. A. P.), a grant from the W. M. Keck Foundation, and the Kleiderer-Pezold family endowed professorship (to F. J. C.). Presented in part at the 41st AnnualMeeting of the American Society of Hematology, New Orleans, December 3–7, 1999.
2 To whom requests for reprints should be addressed, at University of Notre Dame, 229Nieuwland Science Hall, Notre Dame, IN 46556. Phone: (219) 631-6456; Fax: (219) 631-8017; E-mail: [email protected].
3 The abbreviations used are: Pg, plasminogen; Pm, plasmin; bFGF, basic fibroblastgrowth factor; BrdUrd, bromodeoxyuridine; DAB, 39,3-diaminobenzidine-HCl; ECM,extracellular matrix; MMP, matrix metalloprotein; PAI, plasminogen activator inhibitor;RT-PCR, reverse transcription-PCR; tPA, tissue plasminogen activator; TUNEL, terminaldUTP nick end labeling; MVC, microvessel count; uPA, urokinase plasminogen activator;uPAR, urokinase plasminogen activator receptor; VEGF, vascular endothelial growthfactor; vWF, von Willebrand factor; WT, wild type.
5839
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
out using the Ready-To-Go kit (Amersham Pharmacia Biotech, Piscataway,NJ). The entire volume of 33ml of cDNA was amplified by PCR using primersspecific for the gene of interest (Table 1). PCR conditions forPG,TPA,UPAR,andPAI-1 were: 94°C for 2 min (1 cycle) and 35 cycles at 94°C for 1 min,55°C for 1 min, 72°C for 2 min, and then a final incubation at 72°C for 8 min.PCR conditions for theUPA cDNA were similar, except the annealing tem-perature was 60°C.
Histology and Immunohistochemistry. Mice were sacrificed 3 weeksafter tumor implantation. Tumors were excised and fixed overnight with 6%formalin in PBS. Mouse tissues were processed, embedded in paraffin, andsectioned. For the mouse cornea studies, whole eyes were bisected longitudi-nally posterior to the lens after a 10-min fixation in 10% neutral bufferedformalin. The frontal aspect was fixed for an additional 2 h and then routinelyprocessed and embedded in paraffin. The blocks were sectioned at 3mm forstaining with H&E.
For immunohistochemical analysis, 4-mm sections were placed on silane-coated slides, deparaffinized, and rehydrated. Following this, the slides wereblocked for endogenous peroxidase activity with Peroxoblock (Zymed, SanFrancisco, CA). Sections were then preincubated with 10% normal horseserum in PBS-5% BSA for 30 min to block nonspecific binding of theantibodies.
For vWF immunostaining, antihuman vWF antibody (DAKO EPOS,Carpinteria, CA), conjugated to horseradish peroxidase, was applied to theslides and allowed to incubate for 1 h. Detection of the antibody was accom-plished with the chromogen, 3-amino-9-ethylcarbazole (Vector, Burlingame,CA) or DAB (Immuno-Pure metal-enhanced DAB substrate kit; Pierce, Rock-ford, IL). For VEGF immunostaining, murine antihuman VEGF antibody(Oncogene, Research Products, Boston, MA) was used after pretreatment with10 mM Tris-HCl (pH 10). The slides were incubated with the antibodyovernight at room temperature in a humidified chamber. After washing, thesections were incubated for 30 min with a biotinylated horse antimouseantibody (Vector) and with the ABC Vector kit (Vector) for 30 min. Colordevelopment was achieved with 3-amino-9-ethylcarbazole kit (Vector).
For cell proliferation studies, BrdUrd (Amersham Pharmacia, Buckingham-shire, United Kingdom) injections were performed 1 h prior to sacrificing theanimal. BrdUrd immunohistochemistry was performed using a murine mono-clonal antibody (Dako). Before antibody application (1mg/ml), sections wereimmersed in a solution of 2N HCl at 37°C for 10 min, followed by trypsintreatment as above. Antibody was developed as described for anti-VEGF,except that DAB was used as the substrate.
TUNEL. The TUNEL assay was performed according to a publishedmethod (25). Tissue sections were incubated with 5mg/ml of proteinase K for15 min at room temperature to remove proteins. Sections were then coveredwith a buffer containing 30 mM Tris-HCl (pH 7.2), 140 mM sodium cacodylate,and 1 mM CoCl2. An aliquot of 0.2ml of terminal deoxynucleotidyl transferase(Boehringer Mannheim, Gaithersburg, MD) and 10mM biotinylated dUTP
(Boehringer Mannheim) were added to the sections. The slides were incubatedin a humidified chamber at 37°C for 60 min, washed with 50 mM Tris-HCl (pH7), and finally with PBS (0.06M sodium phosphate/0.15M NaCl, pH 7.3). Thesections were then incubated for 30 min with ABC system, and DAB was usedas the chromogen.
Analysis and Scoring. MVCs were determined by operational modifica-tions of a published method (26). MVCs were calculated by analyzing vWF-stained sections. Sections were scanned at low magnification (3100); threeareas per section were considered to have the highest microvessel densitieswithin the tumor, and adjacent peripheral tissues were defined as hot spots.Each hot spot was examined at high magnification (3400), and a computer-digitized image was taken and used for counting the vessels. The mean of theMVCs of three hot spots was designated as the total MVCs/hot spot. Twoinvestigators blinded to the genotype of the animal analyzed the same samples.Three microscopic fields/slide were counted.
Cell proliferation was determined by counting the number of BrdUrd-positive cells. The results were expressed as the percentage of the total numberof tumor cells within a field. The apoptotic index was obtained by dividing thenumber of apoptotic cells by the total number of cells counted within a fieldand then multiplying the results by 1000.
Electron Microscopy. Ultrastructural analyses were performed on tumorsfrom WT,as well asUPA2/2 andPAI-12/2 mice. The tumors were fixed withKarnovsky solution (27), rinsed twice with a solution of 0.1M sodium caco-dylate (pH 7.3), postfixed in 1% osmium tetroxide, dehydrated in a gradedseries of ethanol solutions, and then embedded in epoxy resins (Polysciences,Warrington, PA). Thick sections (0.5mm), stained with toluidine blue, wereused to choose the area of interest, after which corresponding ultrathin sections(90 nm) were cut and stained in 2% uranyl acetate and Reynold’s lead stain(28). Sections were viewed and photographed using a transmission electronmicroscope (Hitachi H600, Tokyo, Japan) at 75 KV accelerating voltage.
Corneal Micropocket Assay. Mice (4–12 weeks of age) of each genotypewere anesthetized with i.p injections of rodent cocktail containing 0.0135 mg
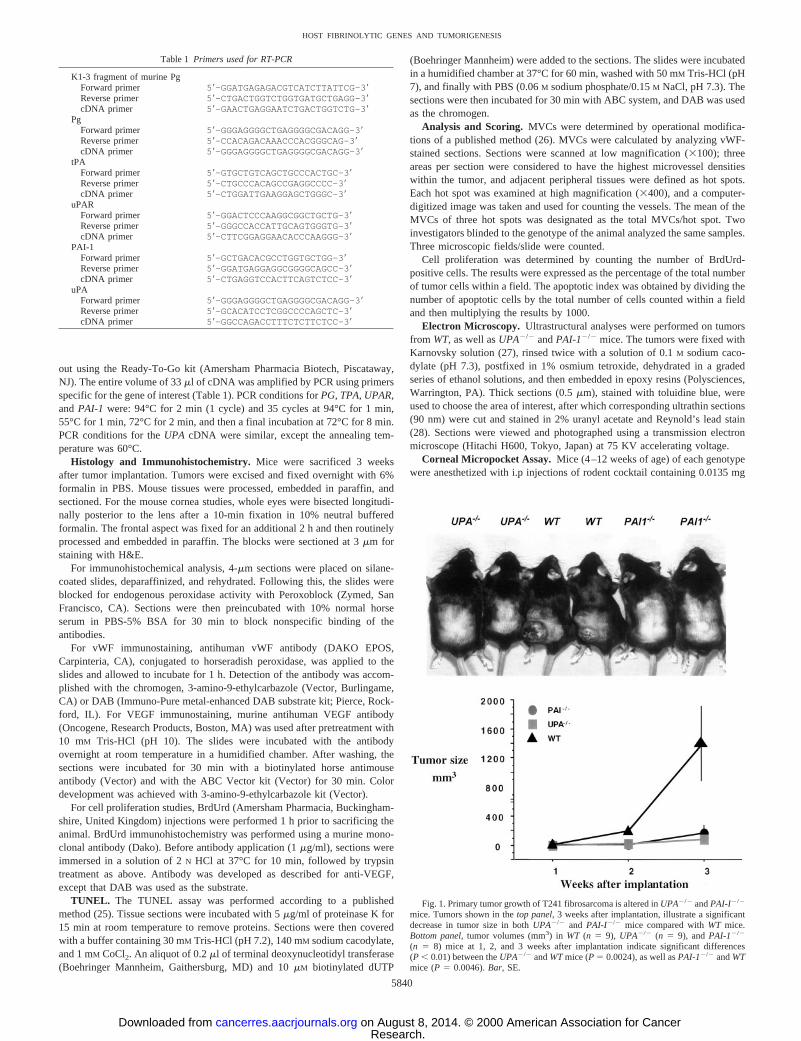
Fig. 1. Primary tumor growth of T241 fibrosarcoma is altered inUPA2/2 andPAI-I2/2
mice. Tumors shown in thetop panel,3 weeks after implantation, illustrate a significantdecrease in tumor size in bothUPA2/2 and PAI-I2/2 mice compared withWT mice.Bottom panel,tumor volumes (mm3) in WT (n 5 9), UPA2/2 (n 5 9), andPAI-12/2
(n 5 8) mice at 1, 2, and 3 weeks after implantation indicate significant differences(P , 0.01) between theUPA2/2 andWTmice (P5 0.0024), as well asPAI-12/2 andWTmice (P5 0.0046).Bar, SE.
Table 1 Primers used for RT-PCR
K1-3 fragment of murine PgForward primer 59-GGATGAGAGACGTCATCTTATTCG-39Reverse primer 59-CTGACTGGTCTGGTGATGCTGAGG-39cDNA primer 59-GAACTGAGGAATCTGACTGGTCTG-39
PgForward primer 59-GGGAGGGGCTGAGGGGCGACAGG-39Reverse primer 59-CCACAGACAAACCCACGGGCAG-39cDNA primer 59-GGGAGGGGCTGAGGGGCGACAGG-39
tPAForward primer 59-GTGCTGTCAGCTGCCCACTGC-39Reverse primer 59-CTGCCCACAGCCGAGGCCCC-39cDNA primer 59-CTGGATTGAAGGAGCTGGGC-39
uPARForward primer 59-GGACTCCCAAGGCGGCTGCTG-39Reverse primer 59-GGGCCACCATTGCAGTGGGTG-39cDNA primer 59-CTTCGGAGGAACACCCAAGGG-39
PAI-1Forward primer 59-GCTGACACGCCTGGTGCTGG-39Reverse primer 59-GGATGAGGAGGCGGGGCAGCC-39cDNA primer 59-CTGAGGTCCACTTCAGTCTCC-39
uPAForward primer 59-GGGAGGGGCTGAGGGGCGACAGG-39Reverse primer 59-GCACATCCTCGGCCCCAGCTC-39cDNA primer 59-GGCCAGACCTTTCTCTTCTCC-39
5840
HOST FIBRINOLYTIC GENES AND TUMORIGENESIS
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
of xylazine, 0.0675 mg of ketamine, and 0.00225 mg of ACE prozamine/gweight of animal. A corneal pocket was made in one eye by a transverseincision, centrally, halfway through the cornea. The pocket was;1 mm fromthe limbic vessel toward the corneal center (29). Hydron-coated pellets ofsucrose aluminum sulfate containing 100 ng of bFGF were placed in thecorneal pocket. On days 3 and 7, bFGF-induced corneal angiogenesis wasevaluated using a slit-lamp ophthamology scope (Nikon) and photographedwith a digital camera (Olympus DP-10). Through the use of a 180° reticule inthe slit-lamp, two measurements were made: maximal vessel length (mm)between the limbic vessel and the bFGF pellet and the circumference ofneovascularization in clock hours (15° of arc5 1 clock hour; Ref. 29). Theneovascular area (mm2) was calculated as described (29, 30).
Statistical Analysis. Statistical analyses of the tumor growth measure-ments and immunohistochemistry scores were performed with nonparametricANOVA and Fisher’s test. The calculations were performed using the Stat-View system for Macintosh (Abacus Concepts, Berkeley, CA).P , 0.05 wasconsidered significant. Where appropriate, values were expressed as themean6 SE.
RESULTS
T241 Fibrosarcoma Tumor Cells Express Fibrinolytic Proteinsin Vitro. This investigation was focused on the role of uPA and PAI-1in tumor growth and dissemination by use ofUPA- and PAI-1-deficient mice in which the murine fibrosarcoma tumor cell line,T241, had been implanted. This is a rapidly growing hypervascularmetastatic tumor that offers advantages in studying tumor angiogen-esis. Analyses of the expression of these components of the fibrino-lytic system were performed by RT-PCR on cultured cells (notshown). mRNA for uPA, PAI-1, u-PAR, and tPA was observed in thiscell line. However, Pg was detected only in the liver control and notin the fibrosarcoma cell line.
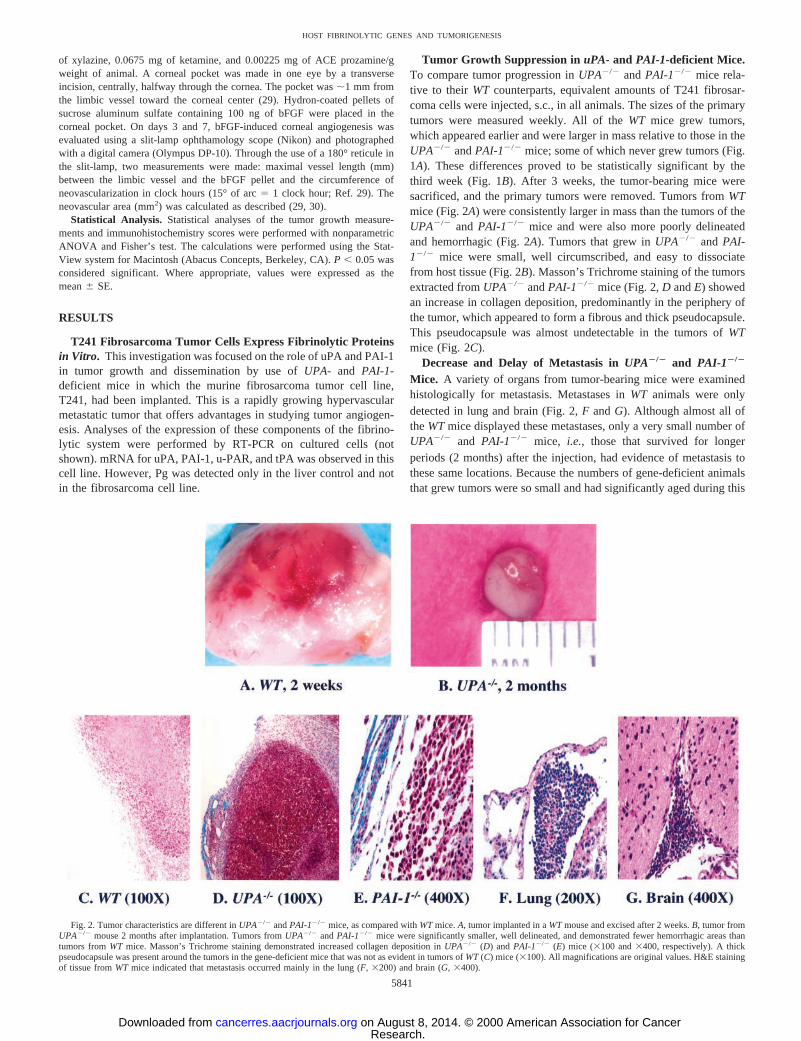
Tumor Growth Suppression in uPA- and PAI-1-deficient Mice.To compare tumor progression inUPA2/2 andPAI-12/2 mice rela-tive to their WT counterparts, equivalent amounts of T241 fibrosar-coma cells were injected, s.c., in all animals. The sizes of the primarytumors were measured weekly. All of theWT mice grew tumors,which appeared earlier and were larger in mass relative to those in theUPA2/2 andPAI-12/2 mice; some of which never grew tumors (Fig.1A). These differences proved to be statistically significant by thethird week (Fig. 1B). After 3 weeks, the tumor-bearing mice weresacrificed, and the primary tumors were removed. Tumors fromWTmice (Fig. 2A) were consistently larger in mass than the tumors of theUPA2/2 and PAI-12/2 mice and were also more poorly delineatedand hemorrhagic (Fig. 2A). Tumors that grew inUPA2/2 and PAI-12/2 mice were small, well circumscribed, and easy to dissociatefrom host tissue (Fig. 2B). Masson’s Trichrome staining of the tumorsextracted fromUPA2/2 andPAI-12/2 mice (Fig. 2,D andE) showedan increase in collagen deposition, predominantly in the periphery ofthe tumor, which appeared to form a fibrous and thick pseudocapsule.This pseudocapsule was almost undetectable in the tumors ofWTmice (Fig. 2C).
Decrease and Delay of Metastasis inUPA2/2 and PAI-12/2
Mice. A variety of organs from tumor-bearing mice were examinedhistologically for metastasis. Metastases inWT animals were onlydetected in lung and brain (Fig. 2,F andG). Although almost all oftheWTmice displayed these metastases, only a very small number ofUPA2/2 and PAI-12/2 mice, i.e., those that survived for longerperiods (2 months) after the injection, had evidence of metastasis tothese same locations. Because the numbers of gene-deficient animalsthat grew tumors were so small and had significantly aged during this
Fig. 2. Tumor characteristics are different inUPA2/2 andPAI-12/2 mice, as compared withWTmice.A, tumor implanted in aWTmouse and excised after 2 weeks.B, tumor fromUPA2/2 mouse 2 months after implantation. Tumors fromUPA2/2 andPAI-12/2 mice were significantly smaller, well delineated, and demonstrated fewer hemorrhagic areas thantumors fromWT mice. Masson’s Trichrome staining demonstrated increased collagen deposition inUPA2/2 (D) and PAI-12/2 (E) mice (3100 and3400, respectively). A thickpseudocapsule was present around the tumors in the gene-deficient mice that was not as evident in tumors ofWT(C) mice (3100). All magnifications are original values. H&E stainingof tissue fromWT mice indicated that metastasis occurred mainly in the lung (F,3200) and brain (G,3400).
5841
HOST FIBRINOLYTIC GENES AND TUMORIGENESIS
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
time, more meaningful commentary on this aspect of the study is notpossible.
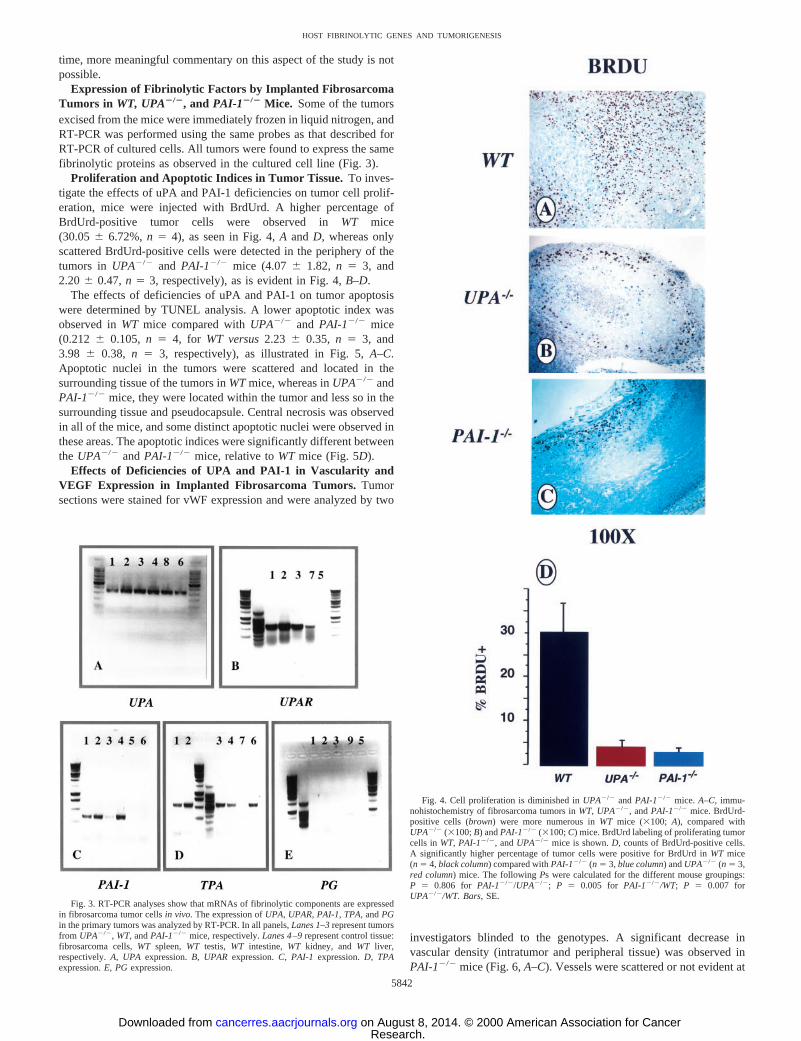
Expression of Fibrinolytic Factors by Implanted FibrosarcomaTumors in WT, UPA2/2, and PAI-12/2 Mice. Some of the tumorsexcised from the mice were immediately frozen in liquid nitrogen, andRT-PCR was performed using the same probes as that described forRT-PCR of cultured cells. All tumors were found to express the samefibrinolytic proteins as observed in the cultured cell line (Fig. 3).
Proliferation and Apoptotic Indices in Tumor Tissue. To inves-tigate the effects of uPA and PAI-1 deficiencies on tumor cell prolif-eration, mice were injected with BrdUrd. A higher percentage ofBrdUrd-positive tumor cells were observed inWT mice(30.056 6.72%,n 5 4), as seen in Fig. 4,A and D, whereas onlyscattered BrdUrd-positive cells were detected in the periphery of thetumors in UPA2/2 and PAI-12/2 mice (4.076 1.82, n 5 3, and2.206 0.47,n 5 3, respectively), as is evident in Fig. 4,B–D.
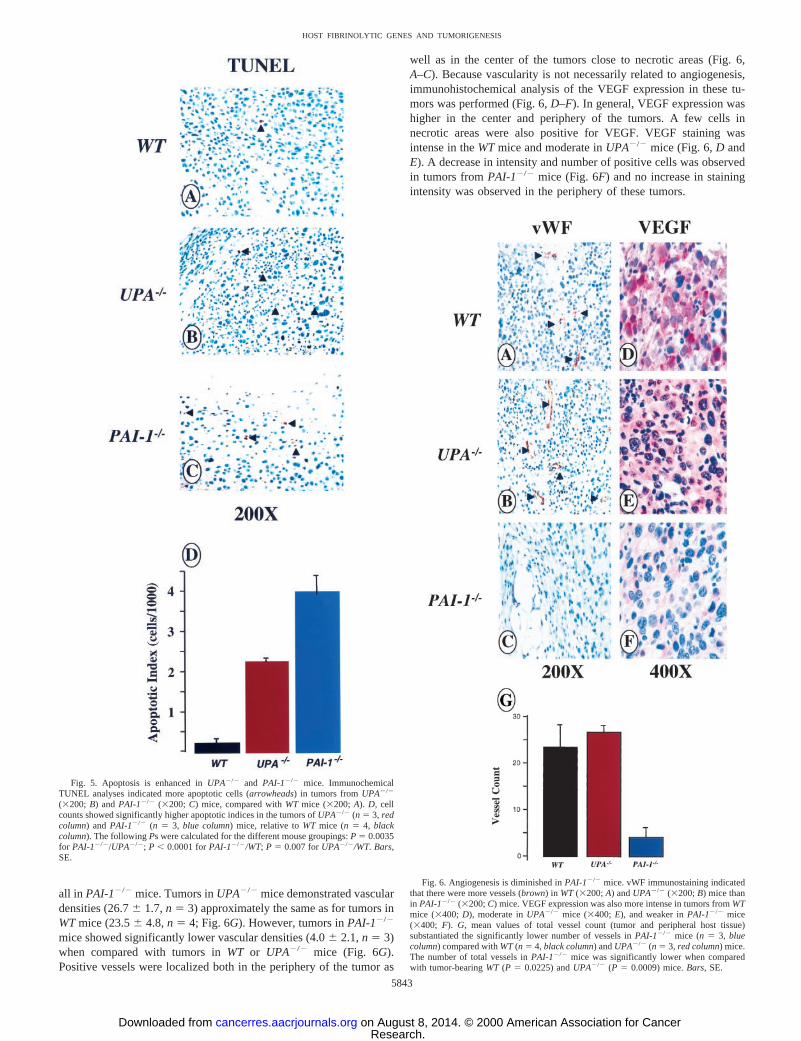
The effects of deficiencies of uPA and PAI-1 on tumor apoptosiswere determined by TUNEL analysis. A lower apoptotic index wasobserved inWT mice compared withUPA2/2 and PAI-12/2 mice(0.212 6 0.105, n 5 4, for WT versus2.23 6 0.35, n 5 3, and3.98 6 0.38, n 5 3, respectively), as illustrated in Fig. 5,A–C.Apoptotic nuclei in the tumors were scattered and located in thesurrounding tissue of the tumors inWTmice, whereas inUPA2/2 andPAI-12/2 mice, they were located within the tumor and less so in thesurrounding tissue and pseudocapsule. Central necrosis was observedin all of the mice, and some distinct apoptotic nuclei were observed inthese areas. The apoptotic indices were significantly different betweenthe UPA2/2 andPAI-12/2 mice, relative toWT mice (Fig. 5D).
Effects of Deficiencies of UPA and PAI-1 in Vascularity andVEGF Expression in Implanted Fibrosarcoma Tumors. Tumorsections were stained for vWF expression and were analyzed by two
investigators blinded to the genotypes. A significant decrease invascular density (intratumor and peripheral tissue) was observed inPAI-12/2 mice (Fig. 6,A–C). Vessels were scattered or not evident at
Fig. 3. RT-PCR analyses show that mRNAs of fibrinolytic components are expressedin fibrosarcoma tumor cellsin vivo. The expression ofUPA, UPAR,PAI-1, TPA, andPGin the primary tumors was analyzed by RT-PCR. In all panels,Lanes 1–3represent tumorsfrom UPA2/2, WT,andPAI-12/2 mice,respectively.Lanes 4–9represent control tissue:fibrosarcoma cells,WT spleen,WT testis, WT intestine, WT kidney, andWT liver,respectively.A, UPA expression.B, UPAR expression.C, PAI-1 expression.D, TPAexpression.E, PGexpression.
Fig. 4. Cell proliferation is diminished inUPA2/2 andPAI-12/2 mice. A–C, immu-nohistochemistry of fibrosarcoma tumors inWT,UPA2/2, andPAI-12/2 mice. BrdUrd-positive cells (brown) were more numerous inWT mice (3100; A), compared withUPA2/2 (3100;B) andPAI-12/2 (3100;C) mice. BrdUrd labeling of proliferating tumorcells in WT,PAI-12/2, andUPA2/2 mice is shown.D, counts of BrdUrd-positive cells.A significantly higher percentage of tumor cells were positive for BrdUrd inWT mice(n 5 4,black column) compared withPAI-12/2 (n 5 3,blue column) andUPA2/2 (n 5 3,red column) mice. The followingPs were calculated for the different mouse groupings:P 5 0.806 for PAI-12/2/UPA2/2; P 5 0.005 for PAI-12/2/WT; P 5 0.007 forUPA2/2/WT. Bars,SE.
5842
HOST FIBRINOLYTIC GENES AND TUMORIGENESIS
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
all in PAI-12/2 mice. Tumors inUPA2/2 mice demonstrated vasculardensities (26.76 1.7,n 5 3) approximately the same as for tumors inWTmice (23.56 4.8,n 5 4; Fig. 6G). However, tumors inPAI-12/2
mice showed significantly lower vascular densities (4.06 2.1,n 5 3)when compared with tumors inWT or UPA2/2 mice (Fig. 6G).Positive vessels were localized both in the periphery of the tumor as
well as in the center of the tumors close to necrotic areas (Fig. 6,A–C). Because vascularity is not necessarily related to angiogenesis,immunohistochemical analysis of the VEGF expression in these tu-mors was performed (Fig. 6,D–F). In general, VEGF expression washigher in the center and periphery of the tumors. A few cells innecrotic areas were also positive for VEGF. VEGF staining wasintense in theWTmice and moderate inUPA2/2 mice (Fig. 6,D andE). A decrease in intensity and number of positive cells was observedin tumors fromPAI-12/2 mice (Fig. 6F) and no increase in stainingintensity was observed in the periphery of these tumors.
Fig. 5. Apoptosis is enhanced inUPA2/2 and PAI-12/2 mice. ImmunochemicalTUNEL analyses indicated more apoptotic cells (arrowheads) in tumors fromUPA2/2
(3200; B) andPAI-12/2 (3200; C) mice, compared withWT mice (3200;A). D, cellcounts showed significantly higher apoptotic indices in the tumors ofUPA2/2 (n 5 3, redcolumn) andPAI-12/2 (n 5 3, blue column) mice, relative toWT mice (n 5 4, blackcolumn). The followingPs were calculated for the different mouse groupings:P 5 0.0035for PAI-12/2/UPA2/2; P , 0.0001 forPAI-12/2/WT;P 5 0.007 forUPA2/2/WT. Bars,SE.
Fig. 6. Angiogenesis is diminished inPAI-12/2 mice. vWF immunostaining indicatedthat there were more vessels (brown) in WT(3200;A) andUPA2/2 (3200;B) mice thanin PAI-12/2 (3200;C) mice. VEGF expression was also more intense in tumors fromWTmice (3400;D), moderate inUPA2/2 mice (3400;E), and weaker inPAI-12/2 mice(3400; F). G, mean values of total vessel count (tumor and peripheral host tissue)substantiated the significantly lower number of vessels inPAI-12/2 mice (n 5 3, bluecolumn) compared withWT(n 5 4, black column) andUPA2/2 (n 5 3, red column) mice.The number of total vessels inPAI-12/2 mice was significantly lower when comparedwith tumor-bearingWT (P 5 0.0225) andUPA2/2 (P 5 0.0009) mice.Bars,SE.
5843
HOST FIBRINOLYTIC GENES AND TUMORIGENESIS
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
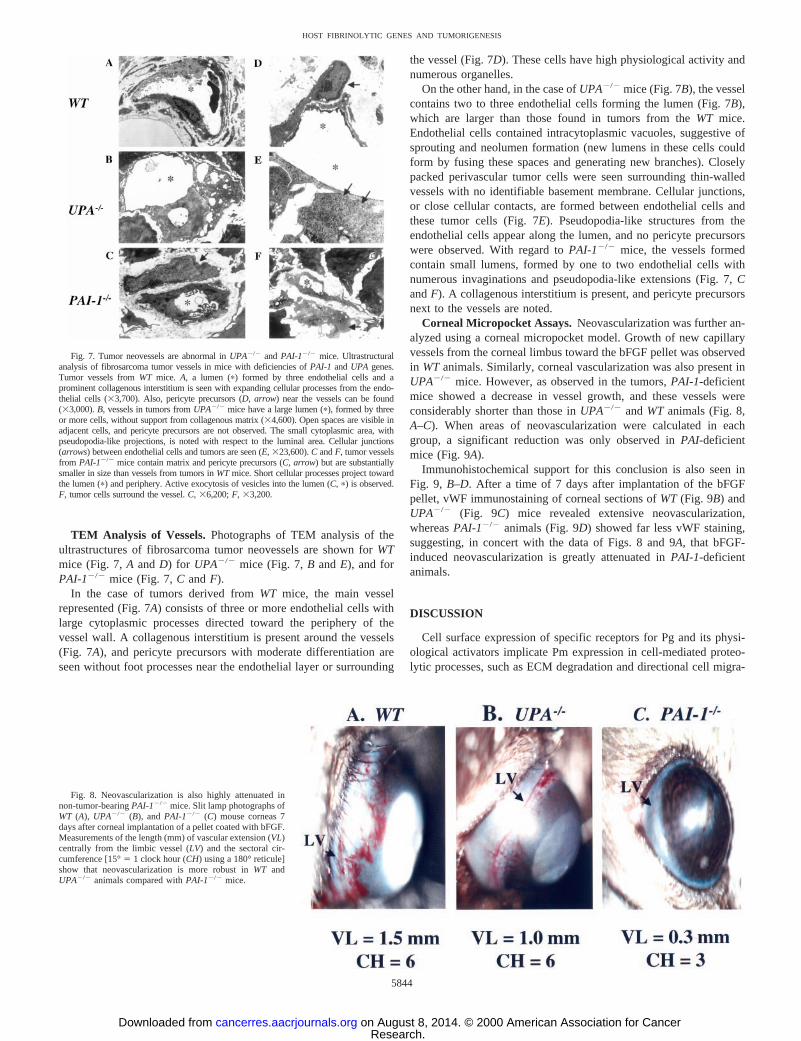
TEM Analysis of Vessels.Photographs of TEM analysis of theultrastructures of fibrosarcoma tumor neovessels are shown forWTmice (Fig. 7,A andD) for UPA2/2 mice (Fig. 7,B andE), and forPAI-12/2 mice (Fig. 7,C andF).
In the case of tumors derived fromWT mice, the main vesselrepresented (Fig. 7A) consists of three or more endothelial cells withlarge cytoplasmic processes directed toward the periphery of thevessel wall. A collagenous interstitium is present around the vessels(Fig. 7A), and pericyte precursors with moderate differentiation areseen without foot processes near the endothelial layer or surrounding
the vessel (Fig. 7D). These cells have high physiological activity andnumerous organelles.
On the other hand, in the case ofUPA2/2 mice (Fig. 7B), the vesselcontains two to three endothelial cells forming the lumen (Fig. 7B),which are larger than those found in tumors from theWT mice.Endothelial cells contained intracytoplasmic vacuoles, suggestive ofsprouting and neolumen formation (new lumens in these cells couldform by fusing these spaces and generating new branches). Closelypacked perivascular tumor cells were seen surrounding thin-walledvessels with no identifiable basement membrane. Cellular junctions,or close cellular contacts, are formed between endothelial cells andthese tumor cells (Fig. 7E). Pseudopodia-like structures from theendothelial cells appear along the lumen, and no pericyte precursorswere observed. With regard toPAI-12/2 mice, the vessels formedcontain small lumens, formed by one to two endothelial cells withnumerous invaginations and pseudopodia-like extensions (Fig. 7,CandF). A collagenous interstitium is present, and pericyte precursorsnext to the vessels are noted.
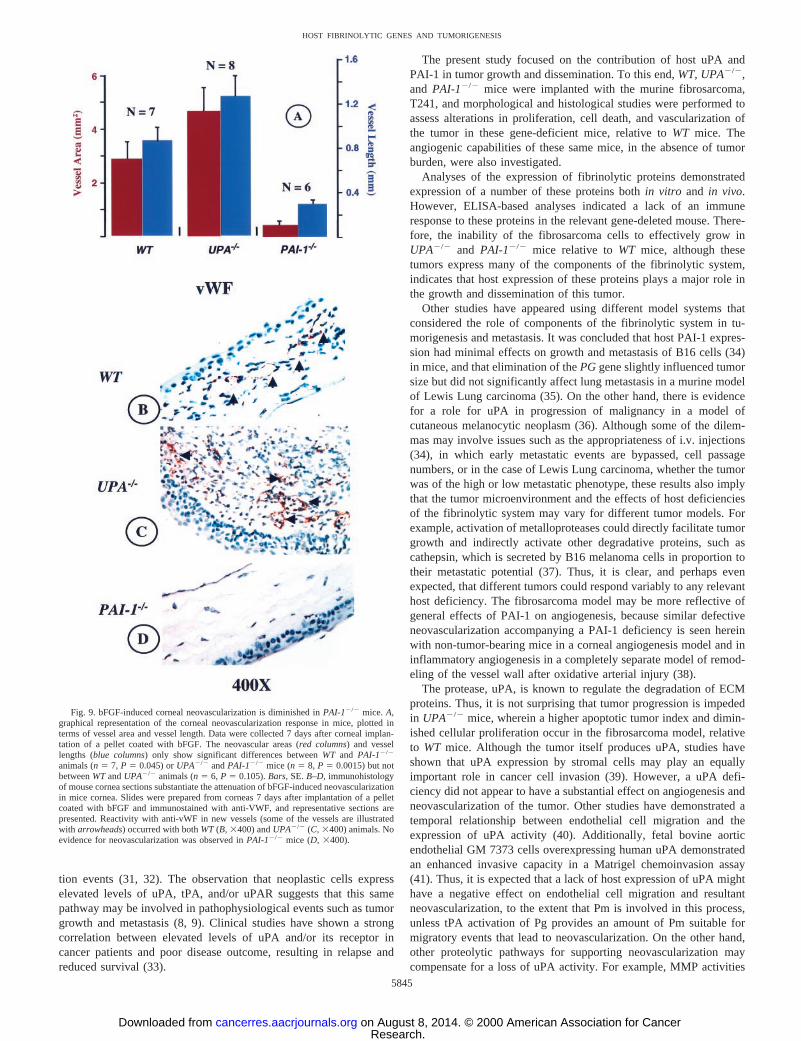
Corneal Micropocket Assays.Neovascularization was further an-alyzed using a corneal micropocket model. Growth of new capillaryvessels from the corneal limbus toward the bFGF pellet was observedin WTanimals. Similarly, corneal vascularization was also present inUPA2/2 mice. However, as observed in the tumors,PAI-1-deficientmice showed a decrease in vessel growth, and these vessels wereconsiderably shorter than those inUPA2/2 andWT animals (Fig. 8,A–C). When areas of neovascularization were calculated in eachgroup, a significant reduction was only observed inPAI-deficientmice (Fig. 9A).
Immunohistochemical support for this conclusion is also seen inFig. 9, B–D. After a time of 7 days after implantation of the bFGFpellet, vWF immunostaining of corneal sections ofWT (Fig. 9B) andUPA2/2 (Fig. 9C) mice revealed extensive neovascularization,whereasPAI-12/2 animals (Fig. 9D) showed far less vWF staining,suggesting, in concert with the data of Figs. 8 and 9A, that bFGF-induced neovascularization is greatly attenuated inPAI-1-deficientanimals.
DISCUSSION
Cell surface expression of specific receptors for Pg and its physi-ological activators implicate Pm expression in cell-mediated proteo-lytic processes, such as ECM degradation and directional cell migra-
Fig. 7. Tumor neovessels are abnormal inUPA2/2 andPAI-12/2 mice. Ultrastructuralanalysis of fibrosarcoma tumor vessels in mice with deficiencies ofPAI-1 andUPA genes.Tumor vessels fromWT mice. A, a lumen (p) formed by three endothelial cells and aprominent collagenous interstitium is seen with expanding cellular processes from the endo-thelial cells (33,700). Also, pericyte precursors (D, arrow) near the vessels can be found(33,000).B, vessels in tumors fromUPA2/2 mice have a large lumen (p), formed by threeor more cells, without support from collagenous matrix (34,600). Open spaces are visible inadjacent cells, and pericyte precursors are not observed. The small cytoplasmic area, withpseudopodia-like projections, is noted with respect to the luminal area. Cellular junctions(arrows) between endothelial cells and tumors are seen (E, 323,600).C andF, tumor vesselsfrom PAI-12/2 mice contain matrix and pericyte precursors (C, arrow) but are substantiallysmaller in size than vessels from tumors inWTmice. Short cellular processes project towardthe lumen (p) and periphery. Active exocytosis of vesicles into the lumen (C, p) is observed.F, tumor cells surround the vessel.C, 36,200;F, 33,200.
Fig. 8. Neovascularization is also highly attenuated innon-tumor-bearingPAI-12/2 mice. Slit lamp photographs ofWT (A), UPA2/2 (B), andPAI-12/2 (C) mouse corneas 7days after corneal implantation of a pellet coated with bFGF.Measurements of the length (mm) of vascular extension (VL)centrally from the limbic vessel (LV) and the sectoral cir-cumference [15°5 1 clock hour (CH) using a 180° reticule]show that neovascularization is more robust inWT andUPA2/2 animals compared withPAI-12/2 mice.
5844
HOST FIBRINOLYTIC GENES AND TUMORIGENESIS
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
tion events (31, 32). The observation that neoplastic cells expresselevated levels of uPA, tPA, and/or uPAR suggests that this samepathway may be involved in pathophysiological events such as tumorgrowth and metastasis (8, 9). Clinical studies have shown a strongcorrelation between elevated levels of uPA and/or its receptor incancer patients and poor disease outcome, resulting in relapse andreduced survival (33).
The present study focused on the contribution of host uPA andPAI-1 in tumor growth and dissemination. To this end,WT,UPA2/2,and PAI-12/2 mice were implanted with the murine fibrosarcoma,T241, and morphological and histological studies were performed toassess alterations in proliferation, cell death, and vascularization ofthe tumor in these gene-deficient mice, relative toWT mice. Theangiogenic capabilities of these same mice, in the absence of tumorburden, were also investigated.
Analyses of the expression of fibrinolytic proteins demonstratedexpression of a number of these proteins bothin vitro and in vivo.However, ELISA-based analyses indicated a lack of an immuneresponse to these proteins in the relevant gene-deleted mouse. There-fore, the inability of the fibrosarcoma cells to effectively grow inUPA2/2 and PAI-12/2 mice relative toWT mice, although thesetumors express many of the components of the fibrinolytic system,indicates that host expression of these proteins plays a major role inthe growth and dissemination of this tumor.
Other studies have appeared using different model systems thatconsidered the role of components of the fibrinolytic system in tu-morigenesis and metastasis. It was concluded that host PAI-1 expres-sion had minimal effects on growth and metastasis of B16 cells (34)in mice, and that elimination of thePGgene slightly influenced tumorsize but did not significantly affect lung metastasis in a murine modelof Lewis Lung carcinoma (35). On the other hand, there is evidencefor a role for uPA in progression of malignancy in a model ofcutaneous melanocytic neoplasm (36). Although some of the dilem-mas may involve issues such as the appropriateness of i.v. injections(34), in which early metastatic events are bypassed, cell passagenumbers, or in the case of Lewis Lung carcinoma, whether the tumorwas of the high or low metastatic phenotype, these results also implythat the tumor microenvironment and the effects of host deficienciesof the fibrinolytic system may vary for different tumor models. Forexample, activation of metalloproteases could directly facilitate tumorgrowth and indirectly activate other degradative proteins, such ascathepsin, which is secreted by B16 melanoma cells in proportion totheir metastatic potential (37). Thus, it is clear, and perhaps evenexpected, that different tumors could respond variably to any relevanthost deficiency. The fibrosarcoma model may be more reflective ofgeneral effects of PAI-1 on angiogenesis, because similar defectiveneovascularization accompanying a PAI-1 deficiency is seen hereinwith non-tumor-bearing mice in a corneal angiogenesis model and ininflammatory angiogenesis in a completely separate model of remod-eling of the vessel wall after oxidative arterial injury (38).
The protease, uPA, is known to regulate the degradation of ECMproteins. Thus, it is not surprising that tumor progression is impededin UPA2/2 mice, wherein a higher apoptotic tumor index and dimin-ished cellular proliferation occur in the fibrosarcoma model, relativeto WT mice. Although the tumor itself produces uPA, studies haveshown that uPA expression by stromal cells may play an equallyimportant role in cancer cell invasion (39). However, a uPA defi-ciency did not appear to have a substantial effect on angiogenesis andneovascularization of the tumor. Other studies have demonstrated atemporal relationship between endothelial cell migration and theexpression of uPA activity (40). Additionally, fetal bovine aorticendothelial GM 7373 cells overexpressing human uPA demonstratedan enhanced invasive capacity in a Matrigel chemoinvasion assay(41). Thus, it is expected that a lack of host expression of uPA mighthave a negative effect on endothelial cell migration and resultantneovascularization, to the extent that Pm is involved in this process,unless tPA activation of Pg provides an amount of Pm suitable formigratory events that lead to neovascularization. On the other hand,other proteolytic pathways for supporting neovascularization maycompensate for a loss of uPA activity. For example, MMP activities
Fig. 9. bFGF-induced corneal neovascularization is diminished inPAI-12/2 mice.A,graphical representation of the corneal neovascularization response in mice, plotted interms of vessel area and vessel length. Data were collected 7 days after corneal implan-tation of a pellet coated with bFGF. The neovascular areas (red columns) and vessellengths (blue columns) only show significant differences betweenWT and PAI-12/2
animals (n5 7, P 5 0.045) orUPA2/2 andPAI-12/2 mice (n5 8, P 5 0.0015) but notbetweenWTandUPA2/2 animals (n5 6, P 5 0.105).Bars,SE.B–D, immunohistologyof mouse cornea sections substantiate the attenuation of bFGF-induced neovascularizationin mice cornea. Slides were prepared from corneas 7 days after implantation of a pelletcoated with bFGF and immunostained with anti-VWF, and representative sections arepresented. Reactivity with anti-vWF in new vessels (some of the vessels are illustratedwith arrowheads) occurred with bothWT(B, 3400) andUPA2/2 (C, 3400) animals. Noevidence for neovascularization was observed inPAI-12/2 mice (D,3400).
5845
HOST FIBRINOLYTIC GENES AND TUMORIGENESIS
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

are tightly regulated during angiogenesis, and their activation, andresulting function in this regard, may not necessarily be dependent onPm activity (42). Clinically, it has been shown in patients withcolorectal adenocarcinoma that MMP-2 and MMP-9 expression pos-itively correlated with angiogenesis (43). Additionally, the transcrip-tion factor, ETS-1, has been implicated in regulation of endothelialcell conversion to an angiogenic phenotype. Cells that overexpressETS-1 are highly invasive and demonstrate increased expression ofMMP-1, MMP-3, and MMP-9 (44).
VEGF is a potent neovascularization agent in some cells (45–48),and its expression is up-regulated by bFGF and other growth factors(49, 50). VEGF, in turn, also up-regulates the expression of Pgactivators and PAI-1 (51), thus providing a possible link between anarray of growth factors, angiogenesis, and extracellular protease ac-tivity. Furthermore, expression of VEGF, itself, is up-regulated bybFGF. Importantly,PAI-12/2 mice displayed greatly reduced expres-sion of VEGF and, probably linked to this effect, diminished angio-genesis and neovascularization of the tumor. As a result, the apoptoticindex was higher in the tumors of these mice than inWTanimals, andcellular proliferation was diminished, thus offering an explanation asto the lack of growth of tumors cells in these gene-deficient animals.This is coupled with ultrastructural analysis, suggesting that neovesselformations inUPA2/2 andPAI2/2 mice have abnormal features, suchas larger and less mature vessels in theUPA2/2 mice and small shortneovessels inPAI-12/2 mice. Without appropriate angiogenesis in thetumor cells, tumors cannot grow and spread effectively.
Previous studies have also shown that PAI-1 blocks cellular bindingof uPAR to vitronectin and thus promotes detachment of uPAR-bearing cells from vitronectin, with a resultant increase in cell motility(52). A loss of host PAI-1 expression could result in enhancedadhesion of uPAR-bearing endothelial cells to the matrix protein,vitronectin, and thus adversely affect cell motility and resultant neo-vascularization. Additionally, balanced cell adhesion and proteolytic-mediated detachment through regulation of uPA activity and inhibi-tion could be necessary for protecting the neovascularizing tissue (53).All of these mechanisms may be functioning to enhance angiogenesisin the presence of host PAI-1, and the absence of this protein wouldthen attenuate neovascularization in tumors and other types of cells.
In summary, the studies presented in this report underscore theimportance of host expression of components of the fibrinolyticsystem in supporting growth and dissemination of the murine fibro-sarcoma tumor, T241. Lack of host expression of either uPA or itsphysiological inhibitor, PAI-1, attenuates growth and survival of thistumor. uPA can provide the necessary targeted proteolysis, amplifiedthrough Pg activation, to allow the cellular migration and proliferationneeded for angiogenesis to occur. However, PAI-1 can provide acontrol to allow this process to occur in a highly regulated manner. Inthe absence of PAI-1, unregulated proteolysis may be detrimental toangiogenesis. Furthermore, the absence of PAI-1 may inhibit endo-thelial cell motility via effects on the uPAR-vitronectin interaction.Additionally, we show that the PAI-1-deficient state may also inhibitneovascularization by affecting VEGF levels, which are needed forthis process to occur. Thus, PAI-1 is a complex regulator of neovas-cularization, and the influence of these different mechanisms likelydepends on the nature of the particular angiogenic process that isoccurring in different cell types.
ACKNOWLEDGMENTS
These studies were performed in collaboration with the Walther CancerFoundation, Indianapolis, IN. We thank Mayra J. Sandoval-Cooper and SunLonghua for assistance with the histology and immunohistochemistry and
Stacey Raje and Melanie E. DeFord for management of veterinary care andbreeding. The input into this research of Drs. Morris Pollard and RudolphNavari is gratefully acknowledged.
REFERENCES
1. Castellino, F. J., Urano, T., deSerrano, V., Morris, J. P., and Chibber, B. A. K. Controlof human plasminogen activation. Haemostasis,18: 15–23, 1988.
2. Collen, D. On the regulation and control of fibrinolysis. Thromb. Haemost.,43:77–89, 1980.
3. Carmeliet, P., Moons, L., Lijnen, R., Baes, M., Lemaitre, V., Tipping, P., Drew, A.,Eeckhout, Y., Shapiro, S., Lupu, F., and Collen, D. Urokinase-generated plasminactivates matrix metalloproteinases during aneurysm formation. Nat. Genet.,17:439–444, 1997.
4. Carmeliet, P., and Collen, D. Evaluation of the plasminogen/plasmin system intransgenic mice. Fibrinolysis,8: 269–276, 1994.
5. Moons, L., Shi, C., Ploplis, V., Plow, E., Haber, E., Collen, D., and Carmeliet, P.Reduced transplant arteriosclerosis in plasminogen-deficient mice. J. Clin. Investig.,102: 1788–1797, 1998.
6. Mignatti, P. The fibrinolytic system and angiogenesis. Fibrinolysis,7: 20–21, 1993.7. DeVries, T. J., Vanmuijen, G. N. P., and Ruiter, D. J. The plasminogen activation
system in tumour invasion and metastasis. Pathol. Res. Pract.,192: 718–733, 1996.8. Ossowski, L., and Reich, E. Antibodies to plasminogen activator inhibit human tumor
metastasis. Cell,35: 611–619, 1983.9. Dano, K., Andraesen, P. A., Grondahl-Hansen, J., Kristensen, P., Nielsen, L. S., and
Skriver, L. Plasminogen activators, tissue degradation, and cancer. Adv. Cancer Res.,44: 139–266, 1985.
10. Bajou, K., Noel, A., Gerard, R. D., Masson, V., Brunner, N., Holst-Hansen, C.,Skobe, M., Fusenig, N. E., Carmeliet, P., Collen, D., and Foidart, J. M. Absence ofhost plasminogen activator inhibitor 1 prevents cancer invasion and vascularization.Nature (Lond.), Med.,4: 923–928, 1998.
11. Blasi, F. Proteolysis, cell adhesion, chemotaxis, and invasiveness are regulated by theu-PA-u-PAR-PAI-1 system. Thromb. Haemost.,82: 298–304, 1999.
12. Folkman, J. What is the evidence that tumors are angiogenesis dependent? J. Natl.Cancer Inst.,82: 4–6, 1989.
13. May, A. E., Kanse, S. M., Chavakis, T., and Preissner, K. T. Molecular interactionsbetween the urokinase receptor and integrins in the vasculature. Fibrinolysis Prote-olysis,12: 205–210, 1998.
14. Duffy, M. J., Duggan, C., Maguire, T., Mulcahy, K., Elvin, P., McDermott, E.,Fennelly, J. J., and O’Higgins, N. Urokinase plasminogen activator as a predictor ofaggressive disease in breast cancer. Enzyme Protein,49: 85–93, 1996.
15. Nekarda, H., Schmitt, M., Ulm, K., Wenninger, A., Vogelsang, H., Becker, K., Roder,J. D., Fink, U., and Siewert, J. R. Prognostic impact of urokinase-type plasminogenactivator and its inhibitor PAI-1 in completely resected gastric cancer. Cancer Res.,54: 2900–2907, 1994.
16. Miyake, H., Hara, I., Yamanaka, K., Gohji, K., Arakawa, S., and Kamidono, S.Elevation of serum levels of urokinase-type plasminogen activator and its receptor isassociated with disease progression and prognosis in patients with prostate cancer.Prostate,39: 123–129, 1999.
17. Foekens, J. A., Peters, H. A., Look, M. P., Portengen, H., Schmitt, M., Kramer, M. D.,Brunner, N., Janicke, F., Meijervan Gelder, M. E., Henzen-Logmans, S. C., vanPutten, W. L. J., and Klijn, J. G. M. The urokinase system of plasminogen activationand prognosis in 2780 breast cancer patients. Cancer Res.,60: 636–643, 2000.
18. Pedersen, H., Brunner, N., Francis, D., Osterlind, K., Ronne, E., Hansen, H. H., Dano,K., and Grondahl-Hansen, J. Prognostic impact of urokinase, urokinase receptor, andtype 1 plasminogen activator inhibitor in squamous and large cell lung cancer tissue.Cancer Res.,54: 4671–4675, 1994.
19. Pedersen, H., Grondahl-Hansen, J., Francis, D., Osterlind, K., Hansen, H. H., Dano,K., and Brunner, N. Urokinase and plasminogen activator inhibitor type-1 in pulmo-nary adenocarcinoma. Cancer Res.,54: 120–123, 1994.
20. Duggan, C., Maguire, T., McDermott, E., O’Higgins, N., Fennelly, J. J., and Duffy,M. J. Urokinase plasminogen activator and urokinase plasminogen activator receptorin breast cancer. Int. J. Cancer,61: 597–600, 1995.
21. Stephens, R. W., Pedersen, A. N., Nielsen, H. J., Hamers, M. J. A. G., Hoyer-Hansen,G., Ronne, E., Dybkjaer, E., Dano, K., and Brunner, N. ELISA determination ofsoluble urokinase receptor in blood from healthy donors and cancer patients. Clin.Chem.,43: 1868–1876, 1997.
22. Brunner, N., Nielsen, H. J., Hamers, M., Christensen, I. J., Thorlacius-Ussing, O., andStephens, R. W. The urokinase plasminogen activator receptor in blood from healthyindividuals and patients with cancer. Acta Pathol. Microbiol. Immunol. Scand.,107:160–167, 1999.
23. Skriver, L., Larsson, L. I., Kielberg, V., Nielsen, L. S., Andresen, P. B., Kristensen,P., and Dano, K. Immunocytochemical localization of urokinase-type plasminogenactivator in Lewis lung carcinoma. J. Cell Biol.,99: 752–757, 1984.
24. Cao, R. H., Wu, H. L., Veitonmaki, N., Linden, P., Farnebo, J., Shi, G. Y., and Cao,Y. H. Suppression of angiogenesis and tumor growth by the inhibitor K1–5 generatedby plasmin-mediated proteolysis. Proc. Natl. Acad. Sci. USA,96: 5728–5733, 1999.
25. Gavrieli, Y., Sherman, Y., and Ben-Sasson, S. Identification of programmed celldeath in situ specific labeling of nuclear DNA fragmentation. J. Cell Biol.,119:493–501, 1992.
26. Zanetta, L., Marcus, S. G., Vasile, J., Dobryansky, M., Cohen, H., Eng, K.,Shamamian, P., and Mignatti, P. Expression of Von Willebrand factor, an endothelialcell marker, is up-regulated by angiogenesis factors: a potential method for objectiveassessment of tumor angiogenesis. Int. J. Cancer,85: 281–288, 2000.
5846
HOST FIBRINOLYTIC GENES AND TUMORIGENESIS
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

27. Hopwood, D. Fixative.In: J. R. Harris (ed.), Electron Microscopy in Biology. APractical Approach, p. 7. Oxford, United Kingdom: Oxford University Press, 1991.
28. Hincherick, F. R. Transmission electron microscopy.In: E. B. Prophet, B. Mills, J. B.Arrington, and L. H. Sobin (eds.), Laboratory Methods in Histotechnology, p. 262.Washington, DC: American Registry of Pathology, 1994.
29. Kenyon, B. M., Voest, E. E., Chen, C. C., Flynn, E., Folkman, J., and D’Amato, R. J.A model of angiogenesis in the mouse cornea. Investig. Ophthalmol. Vis. Sci.,37:1625–1632, 1996.
30. O’Leary, J. J., Shapiro, R. L., Ren, C. J., Chuang, N., Cohen, H. W., and Potmesil,M. Antiangiogenic effects of camptothecin analogues 9-amino-20(S)-camptothecin,topotecan, and CPT-11 studied in the mouse cornea model. Clin. Cancer Res.,5:181–187, 1999.
31. Valinsky, J. E., Reich, E., and Le Douarin, N. M. Plasminogen activator in the bursaof Fabricius: correlations with morphogenetic remodeling and cell migrations. Cell,25: 471–476, 1981.
32. Vassalli, J-D., Baccino, D., and Belin, D. A cellular binding site for theMr 55,000form of the human plasminogen activator, urokinase. J. Cell Biol.,100: 86–92, 1985.
33. Kwaan, H. C. Plasminogen-plasmin system in tumor invasion. Jpn. J. Thromb.Haemost.,5: 79–97, 1994.
34. Eitzman, D. T., Krauss, J. C., Shen, T., Cui, J., and Ginsburg, D. Lack of plasminogenactivator inhibitor-1 effect in a transgenic mouse model of metastatic melanoma.Blood, 87: 18–22, 1996.
35. Bugge, T. H., Kombrinck, K. W., Xiao, Q., Holmback, K., Daugherty, C. C., Witte,D. P., and Degen, J. L. Growth and dissemination of Lewis lung carcinoma inplasminogen-deficient mice. Blood,90: 4522–4531, 1997.
36. Shapiro, R. L., Duquette, J. G., Roses, D. F., Nunes, I., Harris, M. N., Kamino, H.,Wilson, E. L., and Rifkin, D. B. Induction of primary cutaneous melanocytic neo-plasms in urokinase-type plasminogen activator (uPA)-deficient and wild-type mice:cellular blue nevi invade but do not progress to malignant melanoma in uPA-deficientanimals. Cancer Res.,56: 3597–3604, 1994.
37. Qian, F., Chan, S-J., Achkar, C., Steiner, D. F., and Frakfater, A. Transcriptionalregulation of cathepsin B expression in B16 melanomas of varying metastatic poten-tial. Biochem. Biophys. Res. Commun.,202: 429–436, 1994.
38. Ploplis, V. A., Cornelissen, I., Sandoval-Cooper, M., Weeks, L., Noria, F., andCastellino, F. J. Remodeling of the vessel wall after copper-induced injury is highlyattenuated in mice with a total deficiency of plasminogen activator inhibitor-1. Am. J.Pathol., in press, 2000.
39. Okusa, Y., Ichikura, T., and Mochizuki, H. Prognostic impact of stromal cell-derivedurokinase-type plasminogen activator in gastric carcinoma. Cancer (Phila.),85:1033–1038, 1999.
40. Pepper, M. S., Vassalli, J. D., Montesano, R., and Orci, L. Urokinase-type plasmin-ogen activator is induced in migrating capillary endothelial cells. J. Cell Biol.,105:2535–2541, 1987.
41. Gualandris, A., Conejo, T. L., Giunciuglio, D., Albini, A., Sabini, E., Rusnati, M.,Dell Era, P., and Presta, M. Urokinase-type plasminogen activator overexpression
enhances the invasive capacity of endothelial cells. Microvasc. Res.,53: 254–260,1997.
42. Lijnen, H. R., Silence, J., Lemmens, G., Frederix, L., and Collen, D. Regulation ofgelatinase activity in mice with targeted inactivation of components of the plasmin-ogen/plasmin system. Thromb. Haemost.,79: 1171–1176, 1998.
43. Kim, T. S., and Kim, Y. B. Correlation between expression of matrix metallopro-teinase-2 (MMP-2), and matrix metalloproteinase-9 (MMP-9) and angiogenesis incolorectal adenocarcinoma. J. Korean Med. Sci.,14: 263–270, 1999.
44. Oda, N., Abe, M., and Sato, Y. ETS-1 converts endothelial cells to the angiogenicphenotype by inducing the expression of matrix metalloproteinases and integrinb(3).J. Cell. Physiol.,178: 121–132, 1999.
45. Bikfalvi, A., Sauzeau, C., Moukadiri, H., Maclouf, J., Busso, N., Bryckaert, M.,Plouet, J., and Tobelem, G. Interaction of vasculotropin/vascular endothelial cellgrowth factor with human umbilical vein endothelial cells: binding, internalization,degradation, and biological effects. J. Cell. Physiol.,149: 50–59, 1991.
46. Pepper, M. S., Wasi, S., Ferrara, N., Orci, L., and Montesano, R.In vitro angiogenicand proteolytic properties of bovine lymphatic endothelial cells. Exp. Cell Res.,210:298–305, 1994.
47. Nakata, S., Ito, K., Fujimori, M., Shingu, K., Kajikawa, S., Adachi, W., Matsuyama,I., Tsuchiya, S., Kuwano, M., and Amano, J. Involvement of vascular endothelialgrowth factor and urokinase-type plasminogen activator receptor in microvesselinvasion in human colorectal cancers. Int. J. Cancer,79: 179–186, 1998.
48. Sandstrom, M., Johansson, M., Sandstrom, J., Bergenheim, A. T., and Henriksson, R.Expression of the proteolytic factors, tPA and uPA, PAI-1 and VEGF during malig-nant glioma progression. Int. J. Dev. Neurosci.,17: 473–481, 1999.
49. Mandriota, S. J., and Pepper, M. S. Vascular endothelial growth factor-inducedinvitro angiogenesis and plasminogen activator expression are dependent on endoge-nous basic fibroblast growth factor. J. Cell Sci.,110: 2293–2302, 1997.
50. Wojta, J., Kaun, C., Breuss, J. M., Koshelnick, Y., Beckmann, R., Hattey, E., Mildner,M., Weninger, W., Nakamura, T., Tschachler, E., and Binder, B. R. Hepatocytegrowth factor increases expression of vascular endothelial growth factor and plas-minogen activator inhibitor-1 in human keratinocytes and the vascular endothelialgrowth factor receptor flk-1 in human endothelial cells. Lab. Investig.,79: 427–438,1999.
51. Pepper, M. S., Ferrara, N., Orci, L., and Montesano, R. Vascular endothelial growthfactor (VEGF) induces plasminogen activators and plasminogen activator inhibitor-1in microvascular endothelial cells. Biochem. Biophys. Res. Commun.,181: 902–906,1991.
52. Waltz, D. A., Natkin, L. R., Fujita, R. M., Wei, Y., and Chapman, H. A. Plasmin andplasminogen activator inhibitor type 1 promote cellular motility by regulating the inter-action between urokinase receptor and vitronectin. J. Clin. Investig.,100: 58–67, 1997.
53. Bacharach, E., Itin, A., and Keshet, E.In vivo patterns of expression of urokinase andits inhibitor PAI-1 suggest a concerted role in regulating physiological angiogenesis.Proc. Natl. Acad. Sci. USA,89: 10686–10690, 1992.
5847
HOST FIBRINOLYTIC GENES AND TUMORIGENESIS
Research. on August 8, 2014. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
Copyright © 2022 FDOKUMEN




















