
What is the impact of granulocyte colony-stimulating factor (G ...
Upload
independentCategory
view
2download
0
1991 77: 841-848
PH Hart, GF Vitti, DR Burgess, GA Whitty, K Royston and JA Hamilton activitystimulating factor: increased urokinase-type plasminogen activator Activation of human monocytes by granulocyte-macrophage colony-
http://bloodjournal.hematologylibrary.org/content/77/4/841.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
For personal use only.on April 23, 2014. by guest bloodjournal.hematologylibrary.orgFrom For personal use only.on April 23, 2014. by guest bloodjournal.hematologylibrary.orgFrom

Activation of Human Monocytes by Granulocyte-Macrophage Colony-Stimulating Factor: Increased Urokinase-Type Plasminogen Activator Activity
By Prue H. Hart, Gerard F. Vitti, Diana R. Burgess, Genevieve A. Whitty, Keith Royston, and John A. Hamilton
Granulocyte-macrophage colony-stimulating factor (GM- CSF) raised the plasminogen activator (PA) activity of cul- tured human monocytes. This activity was characterized to be urokinase-PA (u-PA) by incubation with specific IgG and by sodium dodecyl sulfate-polyacrylamide gel electrophore- sis zymography. Increased U-PA activity reflected GM-CSF- induction of U-PA mRNA levels. The stimulatory properties of GM-CSF for monocyte PA activity differed from those of interleukin-4, which induced monocyte tissue-type PA (+PA) activity, and of interferon-y (IFN-y), which alone was not
RANULOCYTE-MACROPHAGE colony-stimulat- G ing factor (GM-CSF) was initially recognized as the glycoprotein that stimulated the proliferation and differen- tiation of GM precursor cells from bone marrow (BM).' However, additional biologic roles for this and other CSFs are becoming apparent and include the activation of ma- ture cells of the hematopoietic lineages."
For peripheral blood (PB) human monocytes, high af- finity receptors for GM-CSF have been well character- i~ed .~ , ' GM-CSF activates human monocytes in vitro by increased monocyte tumoricidal activity: HLA-DR expres- sion,' and killing of Leishmania donovani' and Trypanosoma cruzi.' We have previously found that GM-CSF alone does not stimulate interleukin-1 (IL-1) or tumor necrosis factor a (TNFa) activities, or prostaglandin E, (PGE,) levels of human monocytes," disagreeing with a study in which unfractionated mononuclear cells were used." However, our findings are consistent with those of others who reported GM-CSF-enhanced monocyte tumoricidal activ- ity only when a second stimulatory agent, viz endotoxin, was present." Large numbers of activated macrophages have been reported in the peritoneal and pleural cavities of GM-CSF transgenic mice and it has been suggested that products from these macrophages may be responsible for many of the biologic abnormalities observed in these ani mal^.'^"^
Plasminogen activators (PAS) are serine proteases that convert plasminogen to another serine protease, plasmin." There are two types of PA, urokinase-type PA (u-PA) and tissue-type PA (t-PA); the u-PNplasmin system is impli- cated in inflammation, tissue remodeling, and cell migra- tionI6 while the t-PNplasmin is considered to be responsi- ble for the removal of fibrin clots because the t-PA activity is potentiated by fibrin." WeLs and have found that unstimulated human monocytes produce u-PA; how- ever, its activity is not detected in culture supernatants without prior removal biochemically of a monocyte-derived u-PA inhibitor, presumably PA-inhibitor type 2 (PAI-2).2L322 The lymphokine, interferon-y (IFN-y), has been reported t o increase the PA activity of human monocytes cultured on a fibrin ~ubstrate.'~ We have reported that monocytes can also produce t-PA when stimulated with lipopolysaccharide (LPS)," with LPS and IFN--y together," or with another lymphokine, IL-4.24
Because PA expression by the macrophages of GM-CSF transgenic mice might explain much of their pathology, eg,
stimulatory but augmented lipopolysaccharide-induced t-PA activity. GM-CSF alone did not stimulate detectable mono- cyte t-PA activity but combined with IFN-7 to promote this activity. Plasmin formation arising from GM-CSF-induced u-PA in monocytes may contribute to the matrix turnover involved in, eg, cell migration and inflammation, and may explain some of the pathology seen in GM-CSF transgenic mice. o 1991 by The American Society of Hematology.
muscle wasting and peritoneal bleeding,".I4 and because we wanted to explore further the actions of GM-CSF on human monocytes, the PA activities of GM-CSF-treated human monocytes, with and without electrophoresis to remove u-PA inhibitors, were measured and characterized. We report here that for monocytes cultured on regular tissue culture surfaces, GM-CSF alone ( 2 10 U/mL; 2 10 pmol/L) can stimulate u-PA production, thus distinguishing this lymphokine from IFN-y" and IL-4F and distinguishing u-PA from the other monocyte mediators, viz IL-1, TNFa, and PGE,." However, incubation of monocytes with GM- CSF together with IFN-y resulted in the expression of t-PA activity. Unlike IFN-y" and IL-424 again, GM-CSF did not provide a strong, reproducible signal with LPS for en- hanced t-PA activity.
MATERIALS AND METHODS
Monocyte Isolation leukocyte-rich fractions collected after leuko-
phoresis of 500-mL donations of venous PB were obtained from the Melbourne Red Cross Blood Bank. Mononuclear cells were selected by centrifugation (17Og, 40 minutes) of the leukocyte fractions on pyrogen-tested Lymphoprep (Nycomed, Oslo, Nor- way). After multiple washings, the cells (3 x 108 to 8 x lo8) were suspended in Hanks Balanced Salt Solution (Commonwealth Serum Laboratories, Melbourne, Australia) containing 0.21% sodium citrate, 1 pg/mL polymyxin B sulfate (Sigma Chemical Co, St Louis, MO), and 70 pg/mL neomycin sulfate (Upjohn, Rydalm- ere, Australia) and separated by countercurrent centrifugal elutri- ation (Beckman J E d B elutriation system, Palo Alto, CA) with constant rotor speed (2,000 rpm) but increasing pump rates from 8 (loading flow rate) to 22 muminute. Monocytes were collected at
As
From the University of Melboume, Department of Medicine, Royal
Submitted November 20, 1989; accepted October 19, 1990. Supported by grants from the National Health and Medical Re-
search Council of Australia and the Arthritis Foundation of Australia. Address reprint requests to P.H. Hart, PhD, Clinical Microbiology
Unit, School of Medicine, Flinders University of South Australia, South Australia 5042, Australia.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section 1734 solely to indicate this fact.
Melboume Hospital, Parkville, Victoria, Australia.
0 I991 by The American Society of Hematology. 0006-4971 I91 17704-001 2$3.00/0
84 1 Blood, Vol77, No 4 (February 15). 1991: pp 841-848
For personal use only.on April 23, 2014. by guest bloodjournal.hematologylibrary.orgFrom

842 HART ET AL
flow rates between 14.5 and 22 mL/minute; monocyte enrichment was confirmed by May-Griinwald-Giemsa staining of cytocentri- fuged smears and by nonspecific esterase staining.
Monocyte Culture
Cell fractions containing 2 90% monocytes were cultured as previously in a-Modified Eagle's Medium (a-MEM; Flow Laboratories, McLean, VA) supplemented with 20 mmol/L 3-(N-morpholino) propane sulfonic acid (Sigma), 13.3 mmol/L NaHCO,, 2 mmol/L glutamine, 5 x lo-' mol/L 2-mercaptoethanol, and 70 & n L neomycin sulfate (complete a-MEM), osmolarity 290 mmolkg. Cells (0.8 x lo6 to 1.0 x lo6 in 1 mL of medium supplemented with 1% fetal calf serum [FCS; Flow]) were cultured in 2-cm2 wells (Linbro 76-063-05) for up to 18 hours in 5% CO, at 37°C; triplicate cultures for each variable were examined. When indicated, the following reagents were added; recombinant human GM-CSF (rhuGM-CSF) from 1.0 to lo4 U/mL, rhu IFN-7 from 10 to 100 U/mL, and LPS from Escherichiu coli Olll:B4, purified by the Westphal method (Difco, Detroit, MI), at 100 ng/mL. To cultures to which LPS was not added, polymyxin B sulfate was added to a final concentration of 1 pg/mL. Cultures were termi- nated by removal of the supernatant, which was then centrifuged and stored at -20°C until assay. Cell monolayers were treated in one of two ways. For quantitation of the number of cells after 18 hours in culture, cell monolayers together with pelleted cells from the culture supernatants were lysed with Zaponin (Coulter Electron- ics, Bedford, UK) and nuclei measured in a Coulter Sampling Stand; for measurement of cell-associated PA, the same cells were washed twice with phosphate-buffered saline (PBS) before lysis with 0.5 mL 0.2% Triton X-100 (Ajax Chemicals, Sydney, Austra- lia) in Ca2+- and Mg2'-free PBS.
Maintenance of LPS-Free Conditions During monocyte isolation and culture, intense care was taken to
limit LPS c ~ n t a m i n a t i o n . ' ~ ~ ' ~ ~ ~ ~ All equipment was of a plastic disposable nature whenever possible. Glassware was soaked in 1% E-Toxiclean (Sigma) and, after multiple washings, heated to 240°C. All buffers and media were filtered through Zetapor membranes (AMF Cuno, Meriden, CT). LPS levels of less than 50 pg/mL in all reagents were confirmed in the Limulus lysate assay (Common- wealth Serum Laboratories) and by assay of induced procoagulant activity (ie, decreased plasma clotting time in the presence of 0.03 mol/L CaCI,) by mononuclear cells incubated with the reagents in question.lu
Assay of PA Activity By methods previously r e p ~ r t e d , ' ~ . ~ ~ 0.05 mL of culture superna-
tant or cell lysate were added to 0.28-cm2 wells, previously coated with I2'I-fibrin. Plasminogen (0.8 kg dissolved in 0.1 mL Tris-HC1, 0.1 mol/L, pH 8.1) was also added. The plasminogen was prepared from human plasma by Sepharose-lysine affinity chromatography.*' Monocyte-derived plasminogen-independent fibrinolytic activity was assayed in all experiments and was always 1 5 % of the plasminogen-dependent activity. Aliquots were withdrawn after 2 to 3 hours and soluble '"I-fibrin degradation products measured in a gamma counter; one or two further aliquots were taken at later times until significant levels of fibrinolysis were seen. The activities of both one- and two-chain forms of PA were measured because of low levels of contaminating plasmin in plasminogen preparations." PA activity was expressed according to the activity of u-PA standards (0.001 to 2.0 IU/mL; Leo Pharmaceutical Products, Ballerup, Denmark), dissolved in either culture medium or 0.2% Triton X-100 (in Ca*+- and Mg2'-free PBS). In the present experiments, allowances were not made for the lag period of
activation of single-chain to two-chain u-PA16 and activities have been expressed as International Units Equivalent (IUE). PA activities were also expressed according to the number of cells initially plated. As previously reported, in response to GM-CSF there was no significant change in the number of monocytes in any treatment group after 18 hours in culture.Iu
Immunologic Characterization of Monocyte PA Activity IgGs were isolated from rabbit antisera to human u-PA and to
human t-PA using Protein A Sepharose CL-4B (Pharmacia) as previously des~ribed'~.'~; the mouse myeloma IgG, HOPCY, was used as an irrelevant antibody.'* PA standards (0.1 IU/mL; u-PA from Leo Pharmaceutical Products, t-PA as a culture supernatant from the MM138 melanoma cell line26) and pooled supernatants from duplicate 18-hour monocyte cultures were incubated with the IgGs (1 pg/mL final concentration) for 1 hour at 37°C before assay of residual PA activity by the techniques outlined above. PA standards from the National Institute for Biological Standards and Control, London, UK, were used to confirm the definition of PA units.
Characterization of PA by Sodium Dodecyl Sulfate-Polyacylamide Gel Electrophoresis (SDS-PAGE) Zymography
SDS-PAGE was performed essentially as p~b l i shed . '~~~ ' The resolving gel (10%) contained casein (2 mg/mL, Sigma) and human plasminogen (6 kg/mL) and was pre-electrophoresed at 15 mA for 1 hour at room temperature. After loading onto a 4% stacking gel, the samples (0.02 mL), equilibrated in 0.0625 mol/L Tris-HC1, pH 6.8,1.25% SDS, 10% glycerol, were electrophoresed at 12 mA for 2 hours using the buffer system of Laemmli.'x After electrophoresis, the gels were washed for 1 hour in 2.5% Triton X-100, then rinsed and incubated in 0.1 mol/L Tris, pH 8.0, for 18 hours at 37°C. Gels were stained by immersion in 0.25% Coomassie Blue R-250 in 50% methanol/ 7% acetic acid for 60 minutes, then destained for 2 hours in 30% methanol/ 10% acetic acid. Clear bands of lysis appeared where PA had converted plasminogen to plasmin, which then degraded the casein. The hands provided a semi-quantitative measure of PA levels; this was particularly pertinent for superna- tant u-PA, whose activity upon conversion to the two-chain form in the presence of low levels of plasmin" was inhibited in nonelectro- phoresed samples by PAI-2." Low molecular weight markers (Pharmacia) were used to identify the PA standards (0.1 IU/mL); the main lytic bands for u-PA and for t-PA were detected at 52 Kd and 70 Kd, respectively. As published previously,'* immunoprecipi- tation experiments using the specific IgGs described above also confirmed the identity of the standard PA bands.
Detection of mRhLA in Human Monocytes As described,18 total RNA was prepared using the method of
Chirgwin et al.2R RNA (5 pgllane) was fractionated on a formalde- hyde-containing 1% agarose gel before transfer to Genescreen Plus nylon membrane (DuPont, Boston, MA). The filter was hybridized overnight at 60°C in a standard hybridization buffer containing greater than 2 x lo6 cpm/mL of "P-labeled u-PA cDNA, or t-PA cRNA. For u-PA cDNA, hybridization was essentially as previously described? except that it was performed at 45°C. The t-PA cRNA was prepared as before," and after hybridization, the filter was washed and treated with 1 pg/mL RNaseA.'* Loading of mRNA for all lanes was generally shown to be approximately equal by probing with a "P-labeled cDNA for glyceraldehyde-3-phosphate dehydrogenase.
For personal use only.on April 23, 2014. by guest bloodjournal.hematologylibrary.orgFrom

GM-CSF INCREASES MONOCYTE U-PA ACTIVITY 843
Reagents
Bacterially-synthesized huGM-CSF was a kind gift of Dr J.F. DeLamarter, Biogen SA, Geneva, Switzerland; it had been pre- pared and purified as described by Burgess et al.” Batches CAE 392-32 and AS 5/5 were used; the stock solutions of 100 pg/mL and 20 pg/mL had activities of 8 x lo6 U/mL and lo6 U/mL, respec- tively, as judged by day 14 colony growth of normal human BM samples.”’ With a molecular weight of 14.5 Kd, 1,000 U GM- CSF/mL approximated 1.1 nmol/L or 16 ng/mL. The following reagents were kindly supplied by those listed: rhu IFN-.I (1.4 X lo7 U/mg protein), Dr E. Hochuli, Hofkann-La Roche, Basel, Switzer- land; rabbit antisera to human PAS, Dr W-D. Schleuning, Schering AG, Berlin, Germany; MM138 melanoma cell line, Dr R. White- head, Ludwig Institute for Cancer Research, Melbourne, Austra- lia; mouse myeloma IgG, Dr A. Burgess, Ludwig Institute for Cancer Research, Melbourne, Australia; cDNA probe for u-PA and for t-PA, Dr F. Blasi, University of Copenhagen, Denmark, and Dr W-D. Schleuning, respectively.
Expression of Results Unless otherwise indicated, mean values f SEM are shown. The
significance of differences was assessed using a two-tailed Student’s t-test; results were considered significantly different ifP < .05.
RESULTS
PA Activity of GM-CSF-Treated Human Monocytes
To measure (this section) and to characterize (see below) cell-associated PA activity, either a net intracellular activity that allows for inactivation by PA inhibitors, or one associ- ated with specific cell membrane receptor^,'^." lysates of cells treated for 18 hours with GM-CSF were prepared using 0.2% Triton X-100 as the lysing agent. Table 1 shows the mean cell-associated PA activities for triplicate cultures from each of 15 monocyte donors. Although there was variability between donors for the low levels of detectable cell-associated PA activity in both control and stimulated
Table 1. PA Activities of GM-CSF-Treated Monocytes From Several Donors
Addition to Monocyte Cultures.
IFN-y GM-CSF GM-CSF + IFN-y LPS LPS + GM-CSF LPS + IFN-y
PA Activity (IUE/106 cells)t Mean ? SEM
Cell Lysates (n = 15)
0.004 f 0.001 0.005 2 0.002 (NS) 0.036 f 0.017 (P < .01) 0.021 * 0.010 (P < .01) 0.004 f 0.002 (NS) 0.005 t 0.002 (NS) 0.004 -c 0.002 (NS)
Culture Supernatants (n = 19)
ND ND ND
0.14 f 0.04 0.16 f 0.04 0.20 t 0.05 (P* < .05) 0.46 -c 0.07 (P* < .01)
Abbreviations: ND, not detected; NS, not significant (P < .05). *Monocytes were cultured for 18 hours with no added stimulus, or
with the following alone or in combination: IFNy, 100 U/mL; GM-CSF. 1,000 U/mL; LPS, 100 ng/mL.
tPA activities were assayed as described in Materials and Methods and expressed according to the activity of U-PA standards. P values were determined by comparison with control cultures. PS values were determined by comparison with LPS-treated cultures. For the cell lysates, the differences between the mean PA activities forthe GM-CSF- treated and the GM-CSF + IFN-y-treated groups were not statistically significant.
cells, GM-CSF (1,000 U/mL; - 1 nmol/L) consistently enhanced such PA activity. The effects of other monocyte- stimulating agents were also examined; IFN-y (100 U/mL) was inactive by itself and did not alter the stimulatory effect of GM-CSF (Table 1). As we have reported previously,” LPS (100 ng/mL) did not increase the net PA activity. There was no significant difference between LPS-treated cultures and those incubated with LPS and GM-CSF, ie, LPS completely inhibited the stimulatory effect of GM-CSF for cell-associated PA activity. The combination of LPS + IFN-y, which is potent at increasing the synthesis of other monocyte products,’o,L8 was not stimulatory for cell- associated PA activity (Table 1).
We have previously shown that there is no detectable PA activity in the supernatants of unstimulated or IFN-y- treated monocytes cultured for 18 hours on plastic tissue culture dishes.” We now report that PA activity was also not detected in the culture medium of monocytes from 19 donors treated with 1,000 U GM-CSF/mL (Table 1). Even though IFN-y (100 U/mL) was also not stimulatory, simulta- neous incubation of monocytes with GM-CSF and IFN-y gave detectable PA activity in the supernatants of cells from 16 of the 19 donors investigated with a mean activity (+SEM) of 0.14 (k0.04) IUE/106 cells. LPS alone was active for all donors. GM-CSF augmented, although weakly, LPS-induced activities (P < .05, Table 1); however, this increase was significant for only 7 of 19 donors examined. In contrast, IFN-y (100 U/mL) consistently interacted strongly with LPS for enhanced PA activity (Table 1).
Thus, the detectable responses to GM-CSF varied de- pending whether cell-associated or supernatant PA activity was monitored.
Kinetics and Dose Response to GM-CSF
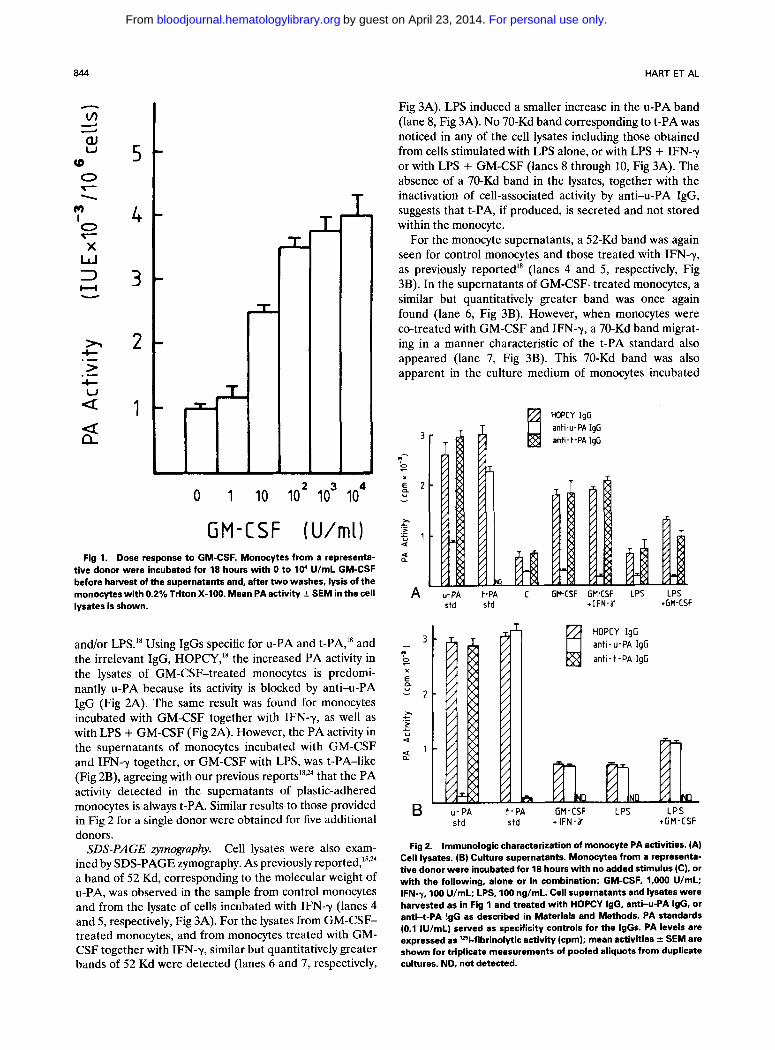
A significant enhancement by GM-CSF of cell-associated PA activity was found in response to 10 U GM-CSFlmL (10 pmol/L) (P < .02), with maximal stimulation detected for this, and other donors, in response to 100 U GM-CSF/mL (Fig 1). For the two donors investigated, increases in cell-associated PA activity were first detected after expo- sure to GM-CSF (1,000 U/mL) for 4 and 6 hours, respec- tively.
PA activity was not detected in the culture medium over a concentration range of 1 to lo4 U GM-CSF/mL (data not shown). When the effects of varying both GM-CSF and IFN-y concentrations on the supernatant PA activity were monitored for two donors, IFN-y at 10 U/mL was less potent than at 100 U/mL for combining with GM-CSF (100 and 1,000 U/mL) (data not shown).
Characterization of the PA Activities of GM-CSF-Treated Human Monocytes
We next determined the type(s) of PA activity that were being measured in the cell lysates and supernatants of monocytes subjected to the various treatment regimes.
We have previously reported that cell- associated PA activities are predominantly u-PA for un- treated monocytes and for those incubated with IFN-y
Antigenicity.
For personal use only.on April 23, 2014. by guest bloodjournal.hematologylibrary.orgFrom
844
h
m aJ d
U
0 0 - \
m I 0
X W 3
7
H Y
)r + > + U
.-
.-
Q
2
Fig 1.
5
4
3
2
1 T n
0 1 10
I.
-
1
- i o 2 lo3 IO*
GM-CSF W m l ) Dose resDonse to GM-CSF. Monocytes from a representa-
tive donor were incubated for 18 hours with.0 to 10' U/mL GM-CSF before harvest of the supernatants and, after two washes, lysis of the monocytes with 0.2% Triton X-100. Mean PA activity t SEM in the cell lysates is shown.
and/or LPS.'* Using IgGs specific for u-PA and t-PA,'* and the irrelevant IgG, HOPCY," the increased PA activity in the lysates of GM-CSF-treated monocytes is predomi- nantly u-PA because its activity is blocked by anti-u-PA IgG (Fig 2A). The same result was found for monocytes incubated with GM-CSF together with IFN-y, as well as with LPS + GM-CSF (Fig 2A). However, the PA activity in the supernatants of monocytes incubated with GM-CSF and IFN-y together, or GM-CSF with LPS, was t-PA-like (Fig 2B), agreeing with our previous that the PA activity detected in the supernatants of plastic-adhered monocytes is always t-PA. Similar results to those provided in Fig 2 for a single donor were obtained for five additional donors.
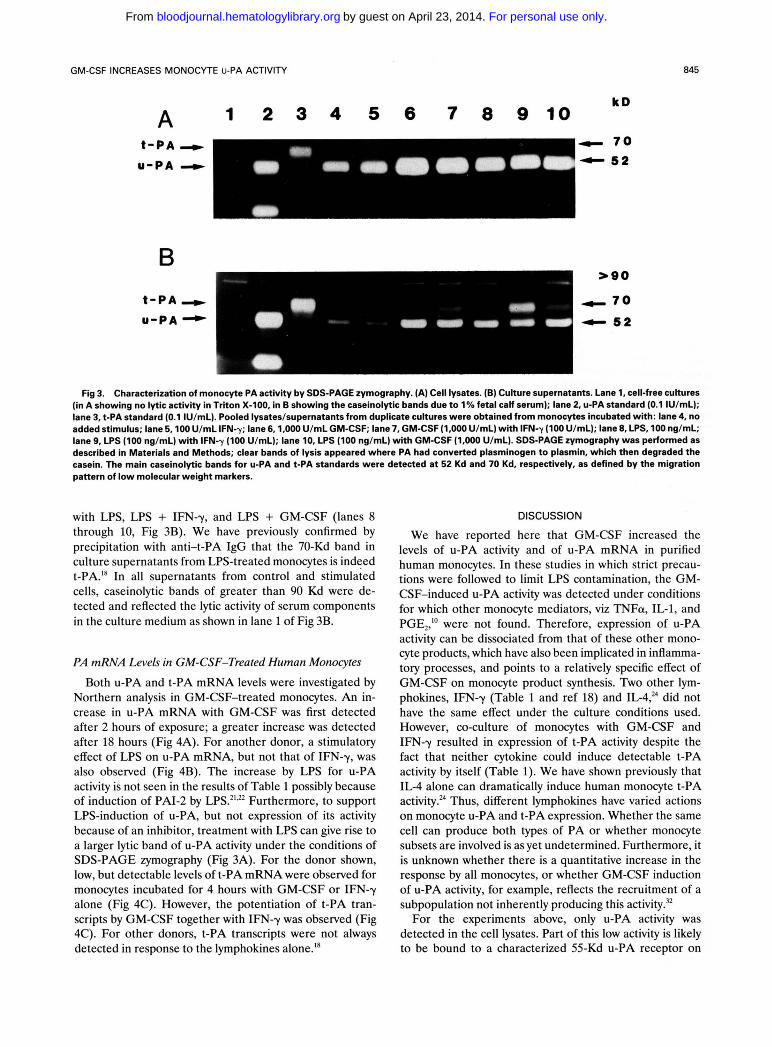
Cell lysates were also exam- ined by SDS-PAGE zymography. As previously rep~rted, '~ ' '~ a band of 52 Kd, corresponding to the molecular weight of u-PA, was observed in the sample from control monocytes and from the lysate of cells incubated with IFN-y (lanes 4 and 5, respectively, Fig 3A). For the lysates from GM-CSF- treated monocytes, and from monocytes treated with GM- CSF together with IFN-y, similar but quantitatively greater bands of 52 Kd were detected (lanes 6 and 7, respectively,
SDS-PAGE zymogruphy.
HART ET AL
Fig 3A). LPS induced a smaller increase in the u-PA band (lane 8, Fig 3A). No 70-Kd band corresponding to t-PA was noticed in any of the cell lysates including those obtained from cells stimulated with LPS alone, or with LPS + IFN-y or with LPS + GM-CSF (lanes 8 through 10, Fig 3A). The absence of a 70-Kd band in the lysates, together with the inactivation of cell-associated activity by anti-u-PA IgG, suggests that t-PA, if produced, is secreted and not stored within the monocyte.
For the monocyte supernatants, a 52-Kd band was again seen for control monocytes and those treated with IFN-y, as previously reported18 (lanes 4 and 5, respectively, Fig 3B). In the supernatants of GM-CSF-treated monocytes, a similar but quantitatively greater band was once again found (lane 6, Fig 3B). However, when monocytes were co-treated with GM-CSF and IFN-y, a 70-Kd band migrat- ing in a manner characteristic of the t-PA standard also appeared (lane 7, Fig 3B). This 70-Kd band was also apparent in the culture medium of monocytes incubated
A U-PA t-PA C GMCSF GMCSF LPS LPS std std + I F N - X +GM-CSF
3 I
? 0,
e - 2
x E
x c .- c Y
a 1 B
B
ant i - t -PA I&
m
u- PA t - P A GM-CSF LPS L P S std std + IFN-X +GM-CSF
Fig 2. Immunologic characterization of monocyte PA activities. (A) Cell lysates. (B) Culture supernatants. Monocytes from a representa- tive donor were incubated for 18 hours with no added stimulus (C), or with the following, alone or in combination: GM-CSF, 1,000 U/mL; IFN-y.100 U/mL; LPS, 100 ng/mL. Cell supernatants and lysates were harvested as in Fig 1 and treated with HOPCY IgG, anti-u-PA IgG, or anti-t-PA !.go as described in Materials and Methods. PA standards (0.1 IU/mL) sewed as specificity controls for the IgGs. PA levels are expressed as '%fibrinolytic activity (cpm); mean activities t SEM are shown for triplicate measurements of pooled aliquots from duplicate cultures. ND, not detected.
For personal use only.on April 23, 2014. by guest bloodjournal.hematologylibrary.orgFrom
GM-CSF INCREASES MONOCYTE U-PA ACTIVITY 845
A kD
1 2 3 4 5 6 7 8 9 1 0
B
Fig 3. Characterization of monocyte PA activity by SDS-PAGE rymography. (A) Cell lysates. (B) Culture supernatants. Lane 1, cell-free cultures (in A showing no lytic activity in Triton X-100. in B showing the caseinolytic bands due to 1 % fetal calf serum); lane 2, U-PA standard (0.1 IU/mL); lane 3, t-PA standard (0.1 IU/mL). Pooled lysates/supernatants from duplicate cultures were obtained from monocytes incubated with: lane 4, no added stimulus; lane 5,100 U/mL IFN-y; lane 6,1,000 U/mL GM-CSF; lane 7, GM-CSF (1,000 U/mL) with IFN-y (100 U/mL); lane 8, LPS, 100 ng/mL; lane 9, LPS (100 ng/mL) with IFN-y (100 U/mL); lane 10, LPS (100 ng/mL) with GM-CSF (1.000 U/mL). SDS-PAGE zymography was performed as described in Materials and Methods; clear bands of lysis appeared where PA had converted plasminogen to plasmin, which then degraded the casein. The main caseinolytic bands for U-PA and t-PA standards were detected at 52 Kd and 70 Kd, respectively, as defined by the migration pattern of low molecular weight markers.
with LPS, LPS + IFN-y, and LPS + GM-CSF (lanes 8 through 10, Fig 3B). We have previously confirmed by precipitation with anti-t-PA IgG that the 70-Kd band in culture supernatants from LPS-treated monocytes is indeed t-PA.I8 In all supernatants from control and stimulated cells, caseinolytic bands of greater than 90 Kd were de- tected and reflected the lytic activity of serum components in the culture medium as shown in lane 1 of Fig 3B.
PA mRNA Levels in GM-CSF-Treated Human Monocytes Both u-PA and t-PA mRNA levels were investigated by
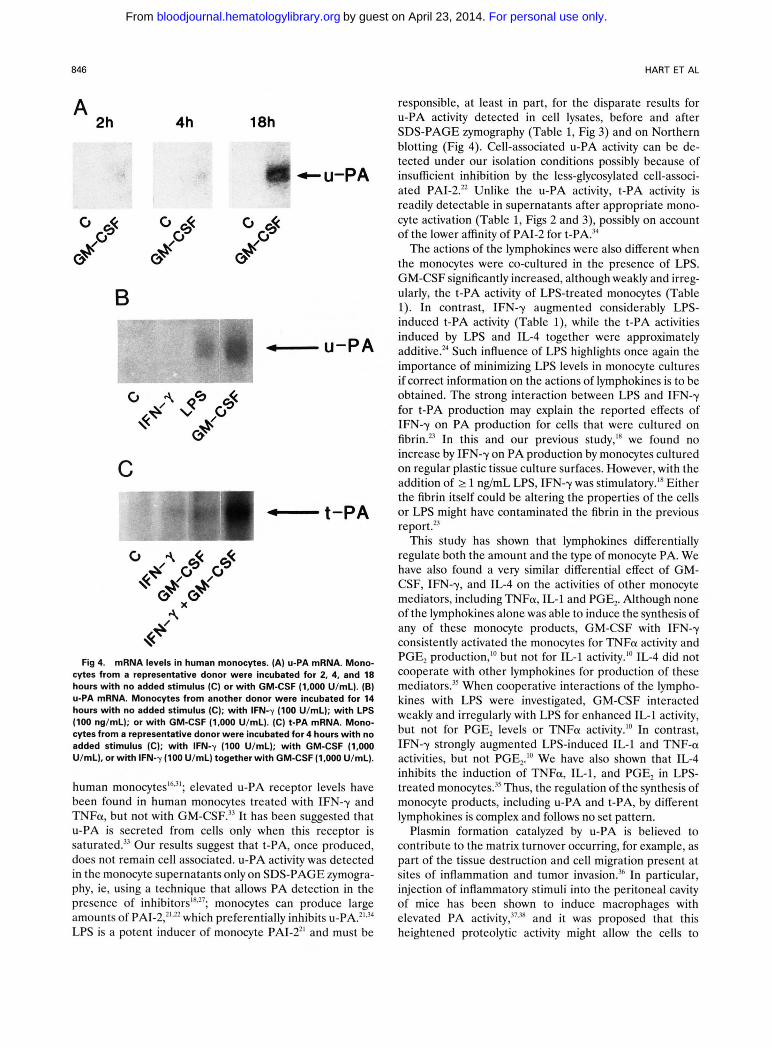
Northern analysis in GM-CSF-treated monocytes. An in- crease in u-PA mRNA with GM-CSF was first detected after 2 hours of exposure; a greater increase was detected after 18 hours (Fig 4A). For another donor, a stimulatory effect of LPS on u-PA mRNA, but not that of IFN-y, was also observed (Fig 4B). The increase by LPS for u-PA activity is not seen in the results of Table 1 possibly because of induction of PAI-2 by LPS?1.22 Furthermore, to support LPS-induction of u-PA, but not expression of its activity because of an inhibitor, treatment with LPS can give rise to a larger lytic band of u-PA activity under the conditions of SDS-PAGE zymography (Fig 3A). For the donor shown, low, but detectable levels of t-PA mRNA were observed for monocytes incubated for 4 hours with GM-CSF or IFN-y alone (Fig 4C). However, the potentiation of t-PA tran- scripts by GM-CSF together with IFN-y was observed (Fig 4C). For other donors, t-PA transcripts were not always detected in response to the lymphokines alone."
DISCUSSION
We have reported here that GM-CSF increased the levels of u-PA activity and of u-PA mRNA in purified human monocytes. In these studies in which strict precau- tions were followed to limit LPS contamination, the GM- CSF-induced u-PA activity was detected under conditions for which other monocyte mediators, viz TNFa, IL-1, and PGE2," were not found. Therefore, expression of u-PA activity can be dissociated from that of these other mono- cyte products, which have also been implicated in inflamma- tory processes, and points to a relatively specific effect of GM-CSF on monocyte product synthesis. Two other lym- phokines, I F N y (Table 1 and ref 18) and IL-4Y did not have the same effect under the culture conditions used. However, co-culture of monocytes with GM-CSF and IFN-y resulted in expression of t-PA activity despite the fact that neither cytokine could induce detectable t-PA activity by itself (Table 1). We have shown previously that IL-4 alone can dramatically induce human monocyte t-PA a~tivity.2~ Thus, different lymphokines have varied actions on monocyte u-PA and t-PA expression. Whether the same cell can produce both types of PA or whether monocyte subsets are involved is as yet undetermined. Furthermore, it is unknown whether there is a quantitative increase in the response by all monocytes, or whether GM-CSF induction of u-PA activity, for example, reflects the recruitment of a subpopulation not inherently producing this activity."
For the experiments above, only u-PA activity was detected in the cell lysates. Part of this low activity is likely to be bound to a characterized 55-Kd u-PA receptor on
For personal use only.on April 23, 2014. by guest bloodjournal.hematologylibrary.orgFrom
HART ET AL 846
2h A
B
4h responsible, at least in part, for the disparate results for U-PA activity detected in cell lysates, before and after SDS-PAGE zymography (Table 1, Fig 3) and on Northern blotting (Fig 4). Cell-associated u-PA activity can be de- tected under our isolation conditions possibly because of insufficient inhibition by the less-glycosylated cell-associ- ated PAI-2." Unlike the u-PA activity, t-PA activity is readily detectable in supernatants after appropriate mono- cyte activation (Table 1, Figs 2 and 3), possibly on account of the lower affinity of PAI-2 for t-PA.34
18h
U-PA
CJ& 6 &
CJ
C
The actions of the lymphokines were also different when the monocytes were co-cultured in the presence of LPS. GM-CSF significantly increased, although weakly and irreg- ularly, the t-PA activity of LPS-treated monocytes (Table 1). In contrast, IFN-y augmented considerably LPS- induced t-PA activity (Table l) , while the t-PA activities induced by LPS and IL-4 together were approximately additi~e.2~ Such influence of LPS highlights once again the U-PA importance of minimizing LPS levels in monocyte cultures if correct information on the actions of lymphokines is to be obtained. The strong interaction between LPS and IFN-y for t-PA production may explain the reported effects of IFN-y on PA production for cells that were cultured on fibrin." In this and our previous study,'" we found no increase by IFN-y on PA production by monocytes cultured on regular plastic tissue culture surfaces. However, with the addition of 2 1 ng/mL LPS, IFN-y was stimulatory." Either the fibrin itself could be altering the properties of the cells or LPS might have contaminated the fibrin in the previous report?
This study has shown that lymphokines differentially regulate both the amount and the type of monocyte PA. We have also found a very similar differential effect of GM- CSF, IFN-y, and IL-4 on the activities of other monocyte mediators, including TNFa, IL-1 and PGE,. Although none of the lymphokines alone was able to induce the synthesis of any of these monocyte products, GM-CSF with IFN-y consistently activated the monocytes for TNFa activity and
t-PA
Fig 4. mRNA levels in human monocytes. (A) U-PA mRNA. Mono- cytes from a representative donor were incubated for 2, 4, and 18 hours with no added stimulus (C) or with GM-CSF (1.000 U/mL). (E) U-PA mRNA. Monocytes from another donor were incubated for 14 hours with no added stimulus (C); with IFN-y (100 U/mL); with LPS (100 ng/mL); or with GM-CSF (1.000 U/mL). (C) t-PA mRNA. Mono- cytes from a representative donor were incubated for 4 hours with no added stimulus (C); with IFN-y (100 U/mL); with GM-CSF (1.000 U/mL), or with IFN-y (100 U/mL) together with GM-CSF (1.000 U/mL).
human monocytes""'; elevated u-PA receptor levels have been found in human monocytes treated with IFN-y and TNFa, but not with GM-CSF..'j It has been suggested that u-PA is secreted from cells only when this receptor is saturated." Our results suggest that t-PA, once produced, does not remain cell associated. u-PA activity was detected in the monocyte supernatants only on SDS-PAGE zymogra- phy, ie, using a technique that allows PA detection in the presence of inhibitor^"^^'; monocytes can produce large amounts of PAI-2,2',22 which preferentially inhibits u-PA? ' .~ LPS is a potent inducer of monocyte PAI-2*' and must be
PGE, production," but not for IL-1 activity.'" IL-4 did not cooperate with other lymphokines for production of these mediators." When cooperative interactions of the lympho- kines with LPS were investigated, GM-CSF interacted weakly and irregularly with LPS for enhanced IL-1 activity, but not for PGE, levels or TNFa activity." In contrast, IFN-y strongly augmented LPS-induced IL-1 and TNF-a activities, but not PGE,." We have also shown that IL-4 inhibits the induction of TNFa, IL-1, and PGE, in LPS- treated monocytes.'s Thus, the regulation of the synthesis of monocyte products, including U-PA and t-PA, by different lymphokines is complex and follows no set pattern.
Plasmin formation catalyzed by u-PA is believed to contribute to the matrix turnover occurring, for example, as part of the tissue destruction and cell migration present at sites of inflammation and tumor invasion.j6 In particular, injection of inflammatory stimuli into the peritoneal cavity of mice has been shown to induce macrophages with elevated PA acti~ity,~'.'' and it was proposed that this heightened proteolytic activity might allow the cells to
For personal use only.on April 23, 2014. by guest bloodjournal.hematologylibrary.orgFrom

GM-CSF INCREASES MONOCYTE U-PA ACTIVITY 847
transgress from the bloodstream into neighboring tissue by degrading interstitial matrix. It has also been shown that GM-CSF activates murine peritoneal macrophages in vitro to produce more PA activity." Elevated levels of GM-CSF have been found in the synovial fluids of patients with rheumatoid arthritis7 Because raised u-PA levels have also been detected in the synovial fluids of such patients:' GM-CSF may contribute to the pathogenesis of this disease by activating monocyte u-PA production, leading to en- hanced monocyte/macrophage migration, cartilage destruc- tion, and inflammation. The same possibility would exist at other inflammatory sites.
It has been suggested that the various complications present in GM-CSF transgenic mice were induced by GM-CSF-stimulated products of macrophages, such as TNFa, IL-1, or PA.'3,14 Our results above showing that GM-CSF by itself stimulates u-PA production, but not that of TNFa or IL-1," indicates that u-PA activity might be of more significance in this regard. GM-CSF alone also does not stimulate IL-1 or TNFa activity or mRNA levels in cultured murine macrophage^.^' PA production by macroph- ages in the GM-CSF transgenic mice may contribute to the necrotic lesions of the eye and striated muscle; a role for PNplasmin in the degradation of the adhesive basement membrane macromolecules of muscle has been ~uggested.~' In certain GM-CSF transgenic mice, there was the develop-
ment of ascites with evidence of chronic bleeding associated with the presence of unusually large numbers of peritoneal macrophage^.'^ Cells produce u-PA, in fact, as a latent or single-chain form (scu-PA) that is activated by trace amounts of plasmin to the two-chain form.43 Because scu-PA re- quires fibrin for its activity, it is being used clinically, in addition to t-PA, as a thrombolytic agent to lyse clots formed, for example, during myocardial in far~ t ion .~~ The bleeding noted in the GM-CSF transgenic miceI4 could be because of plasmin arising from the scu-PA, and therefore u-PA, produced by the activated macrophages. This obser- vation with the transgenic mice suggests that controlled short-term in vivo infusion of GM-CSF might lead to elevated scu-PA levels around monocyte/macrophages which, because of its fibrin-dependent a~tivity:~ could lead to lysis of local fibrin clots. In this connection, we have proposed before that IL-4, on account of its ability to stimulate monocyte production of the thrombolytic t-PA? might have a role in thrombolysis; GM-CSF may function likewise.
This study highlights the complexities of the activation processes in monocytes; further experimentation into the control of expression of PA in monocytes may contribute to a better understanding of GM-CSF-mediated control of inflammatory mediator production by monocytes.
REFERENCES
1. Medcalf D: The molecular biology and functions of the granulocyte macrophage colony-stimulating factors. Blood 67:257, 1986
2. Weisbart RH, Golde DW, Clark SC, Wong GG, Gasson J C Human granulocyte macrophage colony-stimulating factor is a neutrophil activator. Nature 314361,1985
3. Lopez AF, Williamson DJ, Gamble JR, Begley CG, Harlan JH, Klebanoff SJ, Waltersdorf A, Wong G, Clark SC, Vadas M A Recombinant human granulocyte-macrophage colony-stimulating factor stimulates in vitro mature human neutrophil and eosinophil function, surface receptor expression, and survival. J Clin Invest 78:1220,1986
4. Park LS, Friend D, Gillis S, Urdal D L Characterization of the cell surface receptor for human granulocyte/macrophage colony- stimulating factor. J Exp Med 164:251,1986
5. DiPersio J, Billing P, Kaufman S, Eghtesady P, Williams RE, Gasson JC: Characterization of the human granulocyte-macro- phage colony-stimulating factor receptor. J Biol Chem 263:1834, 1988
6. Grabstein KH, Urdal DL, Tushinski RJ, Mochizuki DY, Price VL, Cantrell MA, Gillis S, Conlon PJ: Induction of macrophage tumoricidal activity by granulocyte-macrophage colony-stimulating factor. Science 232506, 1986
7. Alvaro-Gracia JM, Firestein GS, Xu WD, Taetle R, Zvaifler NJ: GM-CSF is a major macrophage activating factor (MAF) in rheumatoid synovitis. Arthritis Rheum 31:S27, 1988 (abstr)
8. Weiser WY, Van Niel A, Clark SC, David JR, Remold HG: Recombinant human granulocyte/macrophage colony-stimulating factor activates intracellular killing of Leishmania donovani by human monocyte-derived macrophages. J Exp Med 166: 1436,1987
9. Reed SG, Nathan CF, Pihl DL, Rodricks P, Shanebeck K, Conlon PJ, Grabstein KH: Recombinant granulocyte/macrophage colony-stimulating factor activates macrophages to inhibit Trypano-
soma cnui and release hydrogen peroxide. Comparison with interferon y. J Exp Med 166:1734,1987
10. Hart PH, Whitty GA, Piccoli DS, Hamilton J A Synergistic activation of human monocytes by granulocyte-macrophage colony- stimulating factor and IFN-y. Increased TNF-a but not IL-1 activity. J Immunol 141:1516,1988
11. Sisson SD, Dinarello C A Production of interleukin-la, interleukin-lp and tumor necrosis factor by human mononuclear cells stimulated with granulocyte-macrophage colony-stimulating factor. Blood 72:1368, 1988
12. Cannistra SA, Vellenga E, Groshek P, Rambaldi A, Griffin JD: Human granulocyte-macrophage colony-stimulating factor and interleukin 3 stimulate monocyte cytotoxicity through a tumor necrosis factor-dependent mechanism. Blood 71:672,1988
13. Lang RA, Metcalf D, Cuthbertson RA, Lyons I, Stanley E, Kelso A, Kannourakis G, Williamson DJ, Klintworth GK, Gonda TJ, Dunn AR: Transgenic mice expressing a hemopoietic growth factor gene (GM-CSF) develop accumulations of macrophages, blindess, and a fatal syndrome of tissue damage. Cell 51:675, 1987
14. Metcalf D, Moore JG: Divergent disease patterns in granulo- cyte-macrophage colony-stimulating factor transgenic mice associ- ated with different transgene insertion sites. Proc Natl Acad Sci USA 85:7767,1988
15. Saksela 0: Plasminogen activation and regulation of pericel- lular proteolysis. Biochim Biophys Acta 823:35, 1985
16. Blasi F, Vassalli J-D, Dan0 K Urokinase-type plasminogen activator: Proenzyme, receptor, and inhibitors. J Cell Biol 104:801, 1987
17. Loscalzo J, Braunwald E: Tissue plasminogen activator. N Engl J Med 319:925,1988
18. Hart PH, Vitti GV, Burgess DR, Singleton DK, Hamilton J A Human monocytes can produce tissue-type plasminogen activa- tor. J Exp Med 169:1509,1989
For personal use only.on April 23, 2014. by guest bloodjournal.hematologylibrary.orgFrom

HART ET AL
19. Vassalli J-D, Dayer J-M, Wohlwend A, Belin D: Concomi- tant secretion of prourokinase and of a plasminogen activator- specific inhibitor by cultured human monocytes-macrophages. J Exp Med 159:1653,1984
20. Saksela 0, Hovi T, Vaheri A: Urokinase-type plasminogen activator and its inhibitor secreted by cultured human monocyte- macrophages. J Cell Physiol122:125, 1985
21. Schwartz BS, Monroe MC, Levin EG: Increased release of plasminogen activator inhibitor type 2 accompanies the human mononuclear cell tissue factor response to lipopolysaccharide. Blood 71:734,1988
22. Wohlwend A, Belin D, Vassalli J-D: Plasminogen activator- specific inhibitors produced by human monocytesimacrophages. J Exp Med 165:320,1987
23. Mokoena T, Gordon S: Human macrophage activation. Modulation of mannosyl, fucosyl receptor activity in vitro by lymphokines, gamma and alpha interferons, and dexamethasone. J Clin Invest 75:624,1985
24. Hart PH, Burgess DR, Vitti GV, Hamilton J A Interleukin-4 stimulates human monocytes to produce tissue-type plasminogen activator. Blood 74:551,1989
25. Hamilton JA, Slywka J: Stimulation of human synovial fibroblast plasminogen activator production by mononuclear cell supernatants. J Immunol126:851,1981
26. Rijken DC, Collen D: Purification and characterization of the plasminogen activator secreted by human melanoma cells in culture. J Biol Chem 256:7035,1981
27. Roche PC, Campeau JD, Shaw ST: Comparative electro- phoretic analysis of human and porcine plasminogen activators in SDS-polyacrylamide gels containing plasminogen and casein. Bio- chim Biophys Acta 745:82,1983
28. Chirgwin JM, Przybyla AFi, MacDonald RJ, Rutter WJ: Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18:5292,1979
29. Vitti GF, Hamilton JA: Modulation of urokinase-type plas- minogen activator messenger RNA levels in human synovial fibroblasts by interleukin-1, retinoic acid, and a glucocorticoid. Arthritis Rheum 31:1046,1988
30. Burgess AW, Begley CG, Johnson GR, Lopez AF, William- son DJ, Mermod JJ, Simpson RJ, Schmitz A, Delamarter JF: Purification and properties of bacterially synthesized human gran- ulocyte-macrophage colony-stimulating factor. Blood 69:43,1987
31. Vassalli J-D, Baccino D, Belin D: A cellular binding site for the Mr 55,000 form of the human plasminogen activator, uroki- nase. J Cell Biol 100:86.1985
32. Fletcher MP, Gasson JC: Enhancement of neutrophil func- tion by granulocyte-macrophage colony-stimulating factor involves recruitment of a less responsive subpopulation. Blood 71:652,1988
33. Kirchheimer JM, Nong Y-H, Remold HG: IFN-7, tumor necrosis factor-a, and urokinase regulate the expression of uroki- nase receptors on human monocytes. J Immunol141:4229,1988
34. Thorsen S, Philips M, Selmer J, Lecander I, Astedt B: Kinetics of inhibition of tissue-type and urokinase-type plasmino- gen activator by plasminogen-activator inhibitor type 1 and type 2. Eur J Biochem 175:33,1988
35. Hart PH, Vitti GV, Burgess DR, Whitty GA, Piccoli DS, Hamilton J A Potential antiinflammatory effects of interleukin 4: Suppression of human monocyte tumor necrosis factor a, interleu- kin 1, and prostaglandin E,. Proc Natl Acad Sci USA 86:3803, 1989
36. Reich E: Activation of plasminogen: A general mechanism for producing localized extracellular proteolysis, in Berlin RD, Herrmann H, Lepow IH, Tanzer JM (eds): Molecular Basis of Biological Degradative Processes. San Diego, CA, Academic, 1978, p 155
37. Vassalli J-D, Hamilton JA, Reich E: Macrophage plasmino- gen activator: Modulation of enzyme production by anti-inflamma- tory steroids, mitotic inhibitors, and cyclic nucleotides. Cell 8:271, 1976
38. Hamilton JA, Vassalli JD, Reich E: Asbestos-induced secre- tion of plasminogen activator from mouse macrophages. J Exp Med 144:1689,1976
39. Hamilton JA, Stanley ER, Burgess AW, Shadduck R K Stimulation of macrophage plasminogen activator activity by colony- stimulating factors. J Cell Physiol 103:435,1980
40. Mochan E, Uhl J: Elevation in synovial fluid plasminogen activator in patients with rheumatoid arthritis. J Rheumatol 11:123,1984
41. Dunn AR, Lang RA, Cuthbertson RA, Metcalf D: Aberrant expression of a murine hemopoietic growth factor (granulocyte macrophage colony stimulating factor) in transgenic mice, in Ross R, Burgess AW, Hunter T (eds): UCLA Symposia on Molecular and Cellular Biology New Series, vol102. Growth Factors and their Receptors: Genetic Control and Rational Application. New York, NY, Lis, 1989, p 219
42. Hantai D, Festoff BW: Degradation of muscle basement membrane zone by locally generated plasmin. Exp Neurol 95:44, 1981
43. Collen D: Molecular mechanisms of fibrinolysis and their application to fibrin-specific thrombolytic therapy. J Cell Biochem 33:11,1987
For personal use only.on April 23, 2014. by guest bloodjournal.hematologylibrary.orgFrom
Copyright © 2022 FDOKUMEN




















