
Plasminogen activators in human corneal fibroblasts: secretion, cellular localization, and...
8
Fibdnolysis (1996) 10(4), 255-262 © PearsonProfessionalLtd 1996 Plasminogen activators in human corneal fibroblasts: secretion, cellular localization, and regulation S. MirshahP '2, J. Soria 2, L. Nelles s, C. Soria 4, J. P. Faure ~, Y, Pouliquen ~, M. MirshahP 1Unit4 de Recherche d'Ophtalmologie, INSERM U 86, H6teI-Dieu, 1 Place du Parvis Notre-Dame, 75181 Paris 04, France 2Laboratoire Sainte-Marie, H6teI-Dieu, Paris, France 3Laboratory of Molecular Biology, University of Leuven, Belgium 4Dif4ma, Universite de Rouen, France Summary Plasminogen activators (PA) play an important role not only in fibrinolysis but also in a variety of processes including tissue remodelling. The stroma of the cornea is a dense connective tissue characterized by its transparency. Healing, degenerative, and inflammatory processes lead to corneal opacification. In an attempt to determine the role of PA in corneal physiology, we have analysed the secretion of PA and plasminogen activator inhibitor (PAl-l) by corneal fibroblasts in vitro. We show that in contrast to other adult fibroblasts, corneal fibroblasts secrete both tissue-type plas- minogen activator (t-PA) and urokinase-plasminogen activator (u-PA). Epithelial growth factor and basic fibroblast growth factor stimulated t-PA secretion whereas transforming growth factor-13 decreased t-PA and increased PAl-1 secretion, t-PA was secreted in the surrounding medium while u-PA remained mostly associated to the cell surface. The production and secretion of t-PA are characteristic of corneal fibroblasts and could be implicated in matrix remodelling and the maintenance of corneal transparency. INTRODUCTION The cornea is composed of three layers: the external layer (epithelium) is in contact with the lacrimal film, the medium layer (stroma) is composed of a matrix syn- thesized by corneal fibroblasts (keratocytes), and the inner layer (endothelium) is in contact with aqueous humor in the anterior chamber of the eye. Cells from both corneal epithelium and endothelium contain plas- minogen activators) These cells produce both tissue- type plasminogen activator (t-PA) and urokinase-plas- minogen activator (u-PA). 2-4 It was suggested that the major role of t-PA was to degrade fibrin in the tear film or in aqueous humor in order to assure their trans- parency and that the major role of u-PA was to help re- epithelialization after a superficial injuryfi -7 In this study, we examined the secretion of PA by the fibroblasts of corneal stroma. It is well established that Received21 January 1996 Accepted after revision 8July 1996 Correspondence to: Dr M. Mirshahi, INSERM U 86, Institut des Cordeliers, 15 rue de I'Ecole de M4decine, 75270 Paris 06, France. Fax. (33)1 4407 1052. fibroblasts secrete PA of u-PA type which is involved in tissue remodelling by two mechanisms. (1) Activation of plasminogen to plasmin at sites of cell-bound u-PA leads to focal degradation of some components of the extracellular matrix (ECM). Moreover, plasmin acti- vates some metalloproteinases, allowing the degrada- tion of other components of the ECM. Plasmin, there- fore, is involved in ceil migrationfi -~1 (2) Plasmin activates growth factors from the ECM: basic fibroblast growth factor (bFGF) is released from its compIex with heparan sulfate proteoglycans, allowing bFGF to dif- fuse in the tissues and to interact with its membrane receptors, j2-~4 Latent transforming growth factor-~ (TGF-~) is activated by plasmin 15 and hepatocyte growth factor (HGF or scatter factor) is directly acti- vated by u-PA. 16 In addition to maintaining homeostasis in cornea, corneal fibroblasts must respond to trauma or other dam- aging stimuli to initiate or participate in repair processes. Corneal fibroblast proliferation, matrix synthesis, and secretion, as well as expression of proteinases capable of tissue remodelling, must be tightly regulated in order to save transparency. 255
Transcript of Plasminogen activators in human corneal fibroblasts: secretion, cellular localization, and...

Fibdnolysis (1996) 10(4), 255-262 © Pearson Professional Ltd 1996
Plasminogen activators in human corneal fibroblasts: secretion, cellular localization, and regulation
S. MirshahP '2, J. Soria 2, L. Nelles s, C. Soria 4, J. P. Faure ~, Y, Pouliquen ~, M. MirshahP 1Unit4 de Recherche d'Ophtalmologie, I NSERM U 86, H6teI-Dieu, 1 Place du Parvis Notre-Dame, 75181 Paris 04, France 2Laboratoire Sainte-Marie, H6teI-Dieu, Paris, France 3Laboratory of Molecular Biology, University of Leuven, Belgium 4Dif4ma, Universite de Rouen, France
Summary Plasminogen activators (PA) play an important role not only in fibrinolysis but also in a variety of processes including tissue remodelling. The stroma of the cornea is a dense connective tissue characterized by its transparency. Healing, degenerative, and inflammatory processes lead to corneal opacification. In an attempt to determine the role of PA in corneal physiology, we have analysed the secretion of PA and plasminogen activator inhibitor (PAl-l) by corneal fibroblasts in vitro. We show that in contrast to other adult fibroblasts, corneal fibroblasts secrete both tissue-type plas- minogen activator (t-PA) and urokinase-plasminogen activator (u-PA). Epithelial growth factor and basic fibroblast growth factor stimulated t-PA secretion whereas transforming growth factor-13 decreased t-PA and increased PAl-1 secretion, t-PA was secreted in the surrounding medium while u-PA remained mostly associated to the cell surface. The production and secretion of t-PA are characteristic of corneal fibroblasts and could be implicated in matrix remodelling and the maintenance of corneal transparency.
INTRODUCTION
The cornea is composed of three layers: the external layer (epithelium) is in contact with the lacrimal film, the medium layer (stroma) is composed of a matrix syn- thesized by corneal fibroblasts (keratocytes), and the inner layer (endothelium) is in contact with aqueous humor in the anterior chamber of the eye. Cells from both corneal epithelium and endothel ium contain plas- minogen activators) These cells produce both tissue- type plasminogen activator (t-PA) and urokinase-plas- minogen a c t i v a t o r (u-PA). 2-4 It was suggested that the major role of t-PA was to degrade fibrin in the tear film or in aqueous humor in order to assure their trans- parency and that the major role of u-PA was to help re- epithelialization after a superficial injuryfi -7
In this study, we examined the secretion of PA by the fibroblasts of corneal stroma. It is well established that
Received21 January 1996 Accepted after revision 8July 1996
Correspondence to: Dr M. Mirshahi, INSERM U 86, Institut des Cordeliers, 15 rue de I'Ecole de M4decine, 75270 Paris 06, France. Fax. (33)1 4407 1052.
fibroblasts secrete PA of u-PA type which is involved in tissue remodelling by two mechanisms. (1) Activation of plasminogen to plasmin at sites of cell-bound u-PA leads to focal degradation of some components of the extracellular matrix (ECM). Moreover, plasmin acti- vates some metalloproteinases, allowing the degrada- tion of other components of the ECM. Plasmin, there- fore, is involved in ceil migrationfi -~1 (2) Plasmin activates growth factors from the ECM: basic fibroblast growth factor (bFGF) is released from its compIex with heparan sulfate proteoglycans, allowing bFGF to dif- fuse in the tissues and to interact with its membrane receptors, j2-~4 Latent transforming growth factor-~ (TGF-~) is activated by plasmin 15 and hepatocyte growth factor (HGF or scatter factor) is directly acti- vated by u - P A . 16
In addition to maintaining homeostasis in cornea, corneal fibroblasts must respond to trauma or other dam- aging stimuli to initiate or participate in repair processes. Corneal fibroblast proliferation, matrix synthesis, and secretion, as well as expression of proteinases capable of tissue remodelling, must be tightly regulated in order to save transparency.
255
256 Mirshahi et al
Recently, we demonstrated the binding of plasminogen to corneal fibroblasts and their extracellular matrix. ~ 7 In this work, we show that normal human corneal fibro- blasts synthesize both t-PA and u-PA in vitro, t-PA is released in the matrix while u-PA remains associated to the cell surface.
MATERIALS AND METHODS
Reagents
RPMI 1640, fetal calf serum, penicillin, and streptomycin were purchased from Gibco-BRL (Paisley, Scotland). Tween-80, diaminobenzine, and bovine serum albumin (BSA) were from Sigma (St Louis, MO, USA). The plasmin- sensitive substrate $2251 (Val-Leu-Lys-paranitroanilin) was from Kabi Vitrum (Stockholm, Sweden). Lys-plas- minogen (Substrene) was from Choay (Paris, France). Before use it was depleted from plasmin contamination by adsorption through aprotinin-agarose. Growth fac- tors: epidermal growth factor (EGF), TGF-~, TGF-[3, and bFGF were from Genzyme (Cambridge, MA, USA). Rabbit anti-u-PA serum and recombinant two-chain-urokinase of high molecular weight were gifts from Dr R. Lijnen (University of Leuven, Belgium). Mouse monoclonal anti- u-PA, anti-t-PA, and anti-PAI-1 antibodies were from American Diagnostics (Greenwich, CT, USA).
Tissue culture
Fibroblasts were isolated from the peripheral ring of human donor corneas that remains after excision of a corneal transplant, and cultured as previously de- scribed. 17 The tissue was divided in small fragments with scissors and the fragments were seeded into uncoated 75 cm 2 culture dishes (Nunc, Naperville, IL, USA) in RPMI 1640 medium supplemented with glutamine (2 x 10 -3 M), fetal calf serum (10%), penicillin (100 U/ml), and strepto- mycin (100U/ml), and incubated at 37°C in a 5% CO2 humidified atmosphere. After a few days, fibroblasts migrate from the explants whereas other cell types, i.e. epithelial and endothelial cells, do not proliferate in this medium. The cultures were expanded by trypsination and fibroblast monolayers were used after two passages. The ceils were grown in either 75 cm 2 culture flasks in order to obtain conditioned supernatant medium or in 96-well tissue culture plate (3 x 104 cells/well) for cell sur- face analysis. The conditioned media, with or without stimulation by growth factors, were prepared in 10 cm 2 culture dishes. The cell monolayers at approximately 80% confluence were rinsed twice with fresh serum-free medium and then incubated with serum-free medium for 18h. The conditioned media were collected and cen- trifuged, and the supernatants were frozen at -20°C.
Amidolytic activity of PA in supernatants and on cell surfaces
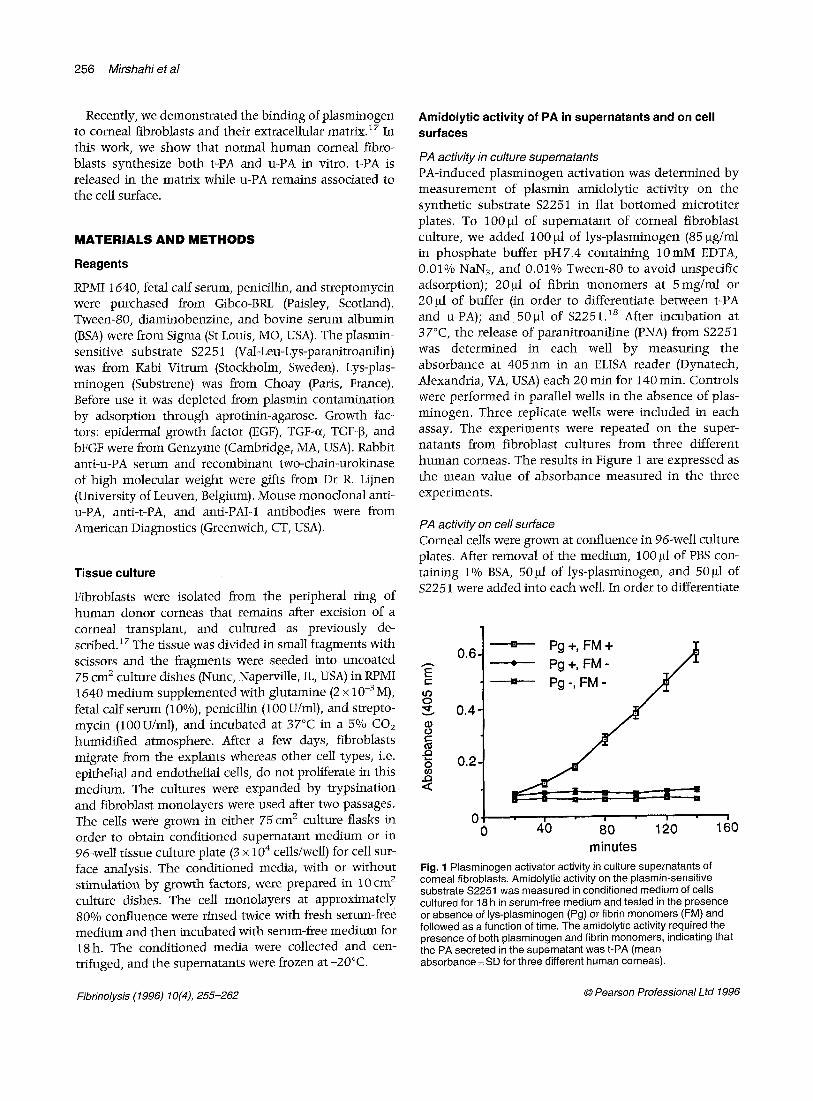
PA activity in culture supematants PA-induced plasminogen activation was determined by measurement of plasmin amidolytic activity on the synthetic substrate $2251 in flat bot tomed microtiter plates. To 100 gl of supernatant of corneal fibroblast culture, we added 100 ~1 of lys-plasminogen (85 gg/ml in phosphate buffer pH7.4 containing 10mM EDTA, 0.01% NaN3, and 0.01% Tween-80 to avoid unspecific adsorption); 20gl of fibrin monomers at 5mg/ml or 20 gl of buffer (in order to differentiate between t-PA and u-PA); and 50gl of $2251. is After incubation at 37°C, the release of paranitroaniline (PNA) from $2251 was determined in each well by measuring the absorbance at 405 nm in an ELISA reader (Dynatech, Alexandria, VA, USA) each 20 min for 140 min. Controls were performed in parallel wells in the absence of plas- minogen. Three replicate wells were included in each assay. The experiments were repeated on the super- natants from fibroblast cultures from three different human corneas. The results in Figure 1 are expressed as the mean value of absorbance measured in the three experiments.
PA activity on cell surface
Corneal cells were grown at confluence in 96-well culture plates. After removal of the medium, 100 gl of PBS con- taining 1% BSA, 50gl of lys-plasminogen, and 50gl of $2251 were added into each well. In order to differentiate
0.6- [] Pg +' FM + 2]~ --- • Pg +, FM - . , J "
o 0.4
o ¢:
"~ 0 . 2
0 o 4'o ;o 14o minutes
Fig. 1 Plasminogen activator activity in culture supernatants of corneal fibroblasts. Amidolytic activity on the plasmin-sensitive substrate $2251 was measured in conditioned medium of cells cultured for 18 h in serum-free medium and tested in the presence or absence of lys-plasminogen (Pg) or fibrin monomers (FM) and followed as a function of time. The amidolytic activity required the presence of both plasminogen and fibrin monomers, indicating that the PA secreted in the supernatant was t-PA (mean absorbance _+ SD for three different human corneas).
Fibrinolysis (1996) 10(4), 255-262 © Pearson Professional Ltd 1996

Plasminogen activators in human corneal fibroblasts 257
between u-PA and t-PA associated to the cells (on cells, fibrin monomers are not needed for p lasminogen acti- vation by t-PA) the incubation was performed in the presence or in the absence of an u-PA inhibitor, amiloride (1 mM, final concentration). 19 The release of PNA was tested as described for the supernatants. Controls were performed in the absence of plasmino- gen. This assay was performed on fibroblasts from five different corneas. The data presented in Figure 2 are the mean values of absorbance measured for these five specimens.
Fibrin zymography
Fibroblast culture supernatants were electrophoresed in SDS-10% polyacrylamide gels according to Laemmli. 2° After washing in 2.5% Triton X-100 to eliminate SDS, the gels were transferred onto precast plasminogen-rich fib- rin plates and incubated in a moist a tmosphere at 37°C for 24h, according to Granelli-Piperno and Reich. 21 A negative control assay was also performed using plas- minogen depleted fibrin plates. Plasminogen-free fibrino- gen used was from Diagnostica-Stago (Asni6res, France). Assays were performed in culture medium previously concentrated five times by freeze drying. Photography of the plates was performed with indirect illumination on a black background.
Table 1 Amounts of t-PA and PAl-1 (ng/105 cells -+ SD, N= 5) released in the supernatants of corneal fibroblasts cultured with or without growth factors
Growth Concentration in factors medium (ng/ml) t-PA PAl-1
none - 7.3 + 0.4 0.4 _+ 0,02 EGF 4 12.0_+1.2(*) 0.6_+ 0.09 (NS) b-FGF 6 13.2 + 2.5 (*) 0.4 __ 0.04 (NS) TGF-J3 2 1.8 _+ 0.3 (**) 1.6 _+ 0.08 (*) TGF-c~ 4 8.1 +_ 1 (NS) 1.0 -4- 0.02 (*)
(*): Significantly higher than control (P< 0.001 ), (**): Significantly lower than control (P< 0.001 ), NS: Not significantly different from control (Mann-Whitney U test).
Enzyme immunoassays
t-PA and PAI-1 were determined by ELISA, using Asserachrom kits from Diagnostica-Stago (Asni6res, France), in the conditioned medium from cells cultured for 18h without fetal calf serum, t-PA and PAI-1 were determined without stimulation and after stimulation of the cells by growth factors EGF, bFGF, TGF-[~, and TGF-~z at the concentrations indicated in Table 1. The test was performed on the fibroblasts from five different human corneas and the results expressed as the mean value of t- PA or PAI-1 concentration (ng/105 cells+SD). Statistical significance of the results on growth factor stimulation or inhibition was calculated using the Mann-Whitney U test.
~= 0.9
8 • pg-
t ~ I P g + o e 0.6 A~ m+ o
~ 0.3
0 Fig. 2 Plasminogen activator activity associated with corneal fibroblasts. Amidolytic activity on $2251 was measured on the cultured fibroblasts after incubation in the presence or absence of plasminogen and amiloride. The PA activity required the addition of plasminogen. It was related to u-PA because it was totally inhibited by the addition of amileride (mean absorbance _+ SD for five different human corneas). The differences in amidolytic activity of the cel ls with or without Pg or with or without amileride are highly significant (P< 0.001).
t-PA, u-PA, and PAl mRNAs
RNA from corneal cells was extracted using the guanidine thiocyanate/acid phenol extraction procedure. 22 PolyA + RNA was purified from cell lysates using the Quick-Prep mRNA purification kit (Pharmacia/LKB, Uppsala, Sweden) according to the manufacturer 's instructions. 2 gg of gly- oxal denatured total RNA were fractionated on a 1% agarose gel and blotted onto a nylon membrane. Parallel gels were probed either for t-PA, u-PA, or PAI-1 using spe- cific riboprobes. Antisense riboprobes were prepared using SP6 or T7 polymerase on linearized plasmids con- taining the appropriate promotor in front of a restriction fragment of the corresponding cDNAs. These fragments were nucleotides 1045 to 1481 of the human PAI-1 eDNA, 23 nucleotides 278 to 885 of the human t-PA eDNA, 24 and nucleotides 1089 to 1448 of the human u- PA eDNA. 25 Hybridization proceeded at 65°C for 18 h in 0.2% BSA, 2°/0 polyanyl, 2°/o 3-pyrrolidine 0.1 mM phos- phate, pH 7.6, 5% dextran sulfate, 1% SDS, 1 mM EDTA, 50% formamide, 200 gg/ml yeast t-RNA, and 200 btg heat- denatLtred herring sperm DNA. Final washing conditions for the membrane were 70°C in 0.015M NaC1, 0.0015M Naa-citrate, pH7.0. Membranes were then exposed for 3 days at -80°C using intensifying screens.
© Pearson Professional Ltd 1996 Fibrinolysis (1996) 10(4), 255-262
258 Mirshahi et al
Immunocytochemistry
Corneal fibroblasts (3 × 104 cells) were cultured in a chamber slide (Lab Tek, Nunc, Naperville, IL, USA) in RPMI 1640 medium with fetal calf serum, glutamine, and antiobiotics. After 2 days the cells were carefully washed three times with PBS containing 1% BSA.
For intracellular detection of u-PA, t-PA or PAI-1, the cells were fixed and permeabifized on the slide by a mix- ture of methanol (95°/o) and acetic acid (5%) at -20°C for 1 min, followed by three washes with PBS. The slides were incubated for 45 min at room temperature with monoclonal antibodies (5 ~tg/mlIg) specific for u-PA, t- PA, or PAI-1, or with a control monoclonal antibody ($9E2, an antibody against bovine arrestin26), then washed three times with PBS. The slides were then incu- bated successively with biotinylated anti-mouse Ig anti- body (Amersham, Amersham, Bucks., UK) diluted 1/50 and streptavidin-biotinylated horseradish peroxidase complex (Amersham) diluted 1/50. Bound peroxidase was detected by microscopic observation after 5 min col- oration with diaminobenzidine (0.4 g/1 distilled water) in the presence of perhydrol.
For detection of u-PA on external cell membrane, the living cells were incubated for 1 h at 4°C with rabbit anti- u-PA antiserum diluted 1/100. After three washes, the anti-u-PA antibodies were detected using two successive incubations for 30 min at 4°C: the first with biotinylated anti-rabbit Ig diluted 1/50, and the second with FITC- conjugated streptavidin diluted 1/50 according to Amersham procedure. Control was performed using nor- real rabbit serum instead of anti-u-PA serum.
RESULTS
Amidolytic activity on S2251
Detection of amidolytic activity in the supematant of corneal fibroblasts needed the presence of both plas- minogen and fibrin monomers indicating that the PA secreted was t-PA (Fig. 1). On the contrary, PA activity associated to cell membrane was u-PA because it needed the presence of plasminogen but was totally inhibited by 1 mM amiloride (Fig. 2). The results were highly repro- ducible for the culture supernatants from three different corneas and the ceils from five different corneas.
I 2 3 4
PA-PAI
t-PA
u-PA
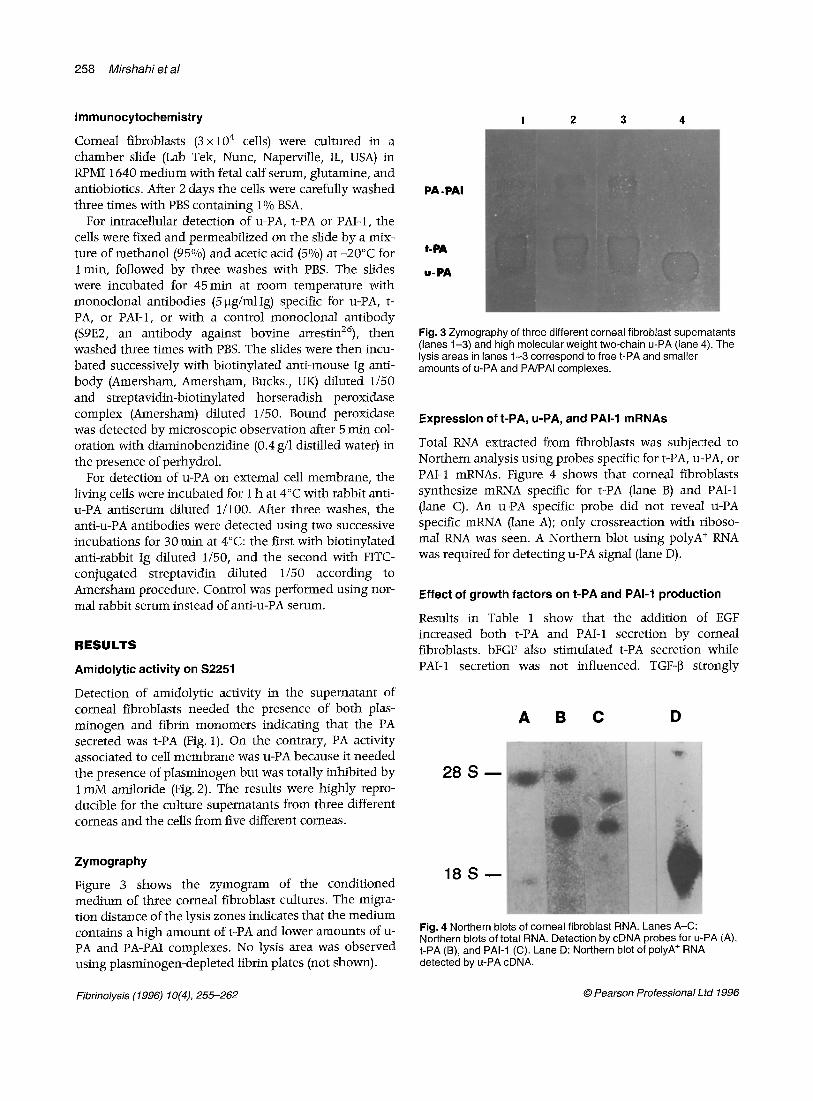
Fig. 3 Zymography of three different corneal fibroblast supernatants (lanes 1-3) and high molecular weight two-chain u-PA (lane 4). The lysis areas in lanes 1-3 correspond to free t-PA and smaller amounts of u-PA and PA/PAI complexes.
Expression of t-PA, u-PA, and PAl-1 mRNAs
Total RNA extracted from fibroblasts was subjected to Northern analysis using probes specific for t-PA, u-PA, or PAI-1 mRNAs. Figure 4 shows that corneal fibroblasts synthesize mRNA specific for t-PA (lane B) and PAI-1 (lane C). An u-PA specific probe did not reveal u-PA specific mRNA (lane .4); only crossreaction with riboso- mal RNA was seen. A Northern blot using polyA + RNA was required for detecting u-PA signal (lane D).
Effect of growth factors on t-PA and PAl-1 production
Results in Table 1 show that the addition of EGF increased both t-PA and PAI-1 secretion by corneal fibroblasts, bFGF also stimulated t-PA secretion while PAI-1 secretion was not influenced. TGF-[3 strongly
A B C D
28 S m
Zymography
Figure 3 shows the zymogram of the conditioned medium of three corneal fibroblast cultures. The migra- tion distance of the lysis zones indicates that the medium contains a high amount of t-PA and lower amounts of u- PA and PA-PAI complexes. No lysis area was observed using plasminogen-depleted fibrin plates (not shown).
1 8 S - -
Fig. 4 Northern blots of corneal fibroblast RNA. Lanes A-C: Northern blots of total RNA. Detection by cDNA probes for u-PA (A), t-PA (B), and PAl-1 (C). Lane D: Northern blot of polyA ÷ RNA detected by u-PA cDNA.
Fibrinolysis (1996) 10(4), 255-262 © Pearson Professional Ltd 1996

Plasminogen activators in human cornea/fibroblasts 259
increased PAI-1 secretion (four times the basal value) and decreased t-PA secretion (a quarter of the basal value). TGF-cz increased PAI-1 (more than twofold the control value), but did not change t-PA level.
Immunocytochemical study
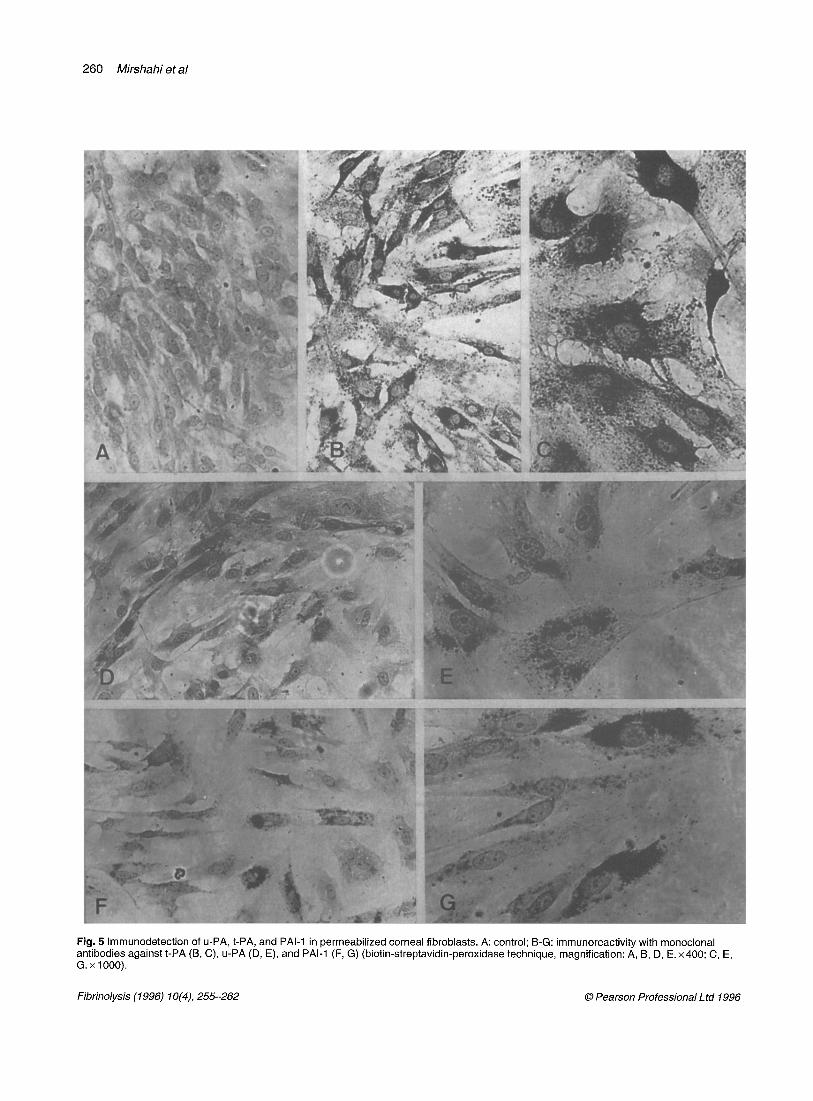
On permeabilized cells, a specific positive reaction in perinuclear cytoplasm was obtained with anti-t-PA (Fig. 5B, C), anti-u-PA (Fig. 5D, E), and anti-PAI-1 (Fig. 5F, G). On living cells, u-PA immunoreactivity was localized on the external cell surface (Fig. 6). On isolated cells, u-PA was set out mainly on the periphery, while on confluent cells u-PA was distributed on the total surface but inho- mogeneously, suggesting that it was located at sites of contact of the cell surface with the substratum, t-PA and u-PA immunoreactivities were very strong in all the cells, but, for PAId, only some of the cells displayed an intense labelling.
DISCUSSION
In this study, we demonstrated that human corneal fibroblasts synthesize both PA (t-PA and u-PA) and their inhibitor PAI-1, as shown by their fibrinolytic and ami- dolytic activities, their immunodetection in cells, and the expression of their mRNA. t-PA was secreted in the super- natant and most of it was in the active free form, as shown by both fibrin zymography (low amount of PA- PAI complex) and amidolytic activity on the plasmin-sen- sitive substrate $2251. u-PA, on the contrary, was expressed on the cell surface, as demonstrated by immunodetection on cultured cells, and by the ami- dolytic activity of living cells, activity which disappears by adding the u-PA inhibitor, amiloride, u-PA associated to cell membrane is probably bound to the membrane via a specific receptor anchored to the cell membrane by a phosphatidyl inositide. 2z On confluent cells, u-PA was concentrated at multiple sites on all the surface of the cell, whereas, when the cells were not confluent, u-PA was located on the edge of cells. This focal distribution probably corresponds to the focal cell-substratum or cell- cell contacts where u-PA and its receptor are concen- trated. 2s'29 Our findings indicate that fibroblasts from cornea look like both embryonic fibroblasts which secrete t-PA, and mature fibroblasts which secrete u-PA remaining associated to cell membrane through specific u-PA receptor. The PA secreted by corneal fibroblasts could induce the degradation of ECM, and therefore the remodelling of the matrix, since plasmin generated by PA-induced plasminogen activation degrades fibronec- tin 3° and a 66kDa ECM-dependent protein. 31 Further- more, plasmin also activates a procollagenase which is secreted in large amounts by corneal fibroblasts. 32
Therefore, the secretion of both collagenase and PA con- tributes to ECM degradation and cell migration. PAI-1 was also secreted by these corneal cells. It is common that PA-producing cells also express their inhibitor PAI-1. Since PAI-1 binds to vitronectin, a component of the E C M 33'34 that we have immunodetected in the corneal stroma (unpublished observation), PAId seems impli- cated in the adhesion of cells to ECM.
As already described for the secretion of PA by vascu- lar endothelial cells, 1°'23 t-PA and PAI-1 secretion by corneal fibroblasts was also regulated by growth factors. EGF and bFGF were inducers of t-PA secretion while TGF-~ inhibited this secretion and increased the secre- tion of the inhibitor PAI-1. One can suggest that endogenous growth factors could modulate PA and PAI activity in vivo and thus contribute to corneal connec- tive tissue remodelling and corneal wound healing. Though the presence of the factors in the corneal stroma in situ was not ascertained, the expression of mRNAs for EGF, EGF receptor, bFGF, and TGF-[~I has been evidenced in both corneal epithelial cells and stro- mal fibroblasts in primary culture. 3s Healing of corneal wounds is a complex biological process involving migration, mitosis and differentiation of epithelial cells and stromal fibroblasts, and synthesis of collagen and other connective tissue components. Several experi- ments (reviewed in references 35 and 36) have shown that exogenous EGF, FGF, TGF-a, and TGF-[~ are potent modulators of corneal wound healing. Our observations suggest that the activity of these factors in the healing process can be mediated by PA or PAI activation. Furthermore, the secretion of t-PA by corneal fibrob- lasts, by activating plasminogen associated to ECM into active plasmin, can be responsible for the release of growth factors from their complexes with glycosamino- glycans of the ECM. Following corneal injury, one can speculate a bimodal regulation in wound healing. Stromal fibroblasts migrate to the wound. The migra- tion is facilitated by an increased expression of u-PA on the cell surface, u-PA can activate the scatter factor that is secreted by corneal fibroblasts and probably con- tributes to the epithelial wound healing process. 37 u-PA also induces a local generation of plasmin which degrades some components of the ECM. However, in turn, u-PA could activate growth factors such as TGFq3 which, on the contrary, blocks the destruction of newly synthesized ECM components by up-regulating the syn- thesis of protease inhibitors and down-regulating the synthesis of proteases involved in the degradation of the matrix. However, the secretion of u-PA is up-regu- lated by a positive feedback: u-PA also mediates the release of active form of bFGF from its complex with glycosaminoglycans of the E C M . 13 Active bFGF then binds to its cellular receptor, stimulating u-PA secretion.
© Pearson Professional Ltd 1996 Fibrinolysis (1996) 10(4), 255-262
260 Mirshahi et al
Fig. 5 Immunodetection of u-PA, t-PA, and PAl-1 in permeabilized corneal fibroblasts. A: control; B-G: immunoreactivity with monoclonal antibodies against t-PA (B, C), u-PA (D, E), and PAl-1 (F, G) (biotin-streptavidin-peroxidase technique, magnification: A, B, D, E. x400; C, E, G. x 1000).
Fibrinolysis (1996) 10(4), 255-262 © Pearson Professional Ltd 1996

Plasminogen activators in human corneal fibroblasts 261
Fig. 6 Immunodetection of u-PA on the surface of living corneal fibroblasts, u-PA immunoreactivity was maximal on the edge of isolated cells (A) and distributed on all the cell surface of confluent cells (B: two cells, C: cluster of cells) (magnification: x 1000).
We therefore argue tha t the h igh synthes is and secre- t ion of PA by the corneal f ibroblasts shou ld be impor- t an t for connec t ive t issue r emode l l ing by ac t iva t ion or release of several g rowth factors secreted locally b y the corneal cells.
We sugges t tha t the role of u-PA is l imi ted to the close v ic in i ty of the cells, s ince u-PA rema ins associ- a ted to the cell m e m b r a n e , and tha t it shou ld p lay a major role in cell m ig ra t i on especial ly d u r i n g the heal- ing process, t-PA b e i n g secreted in the m e d i u m could be impl ica ted in mat r ix remode l l ing , an i m p o r t a n t phe- n o m e n o n impl ica ted in the m a i n t e n a n c e of corneal t r ansparency .
REFERENCES
1. Pandolfi M, Astedt B, Dyster-Aas K. Release of fibfinolytic activators from human cornea. Acta Ophthalmol 1972; 50: 199-205.
2. Fehrenbacher L, Gospodarowicz D, Shuman M A. Synthesis of plasminogen activator in bovine corneal endothelial cells. Exp Eye Res 1979; 29: 216-228.
3. Mirshahi M, Mirshahi S, Sofia C, Soria J, ThomaJdis A, Pouliquen Y, Fanre J P. Production of proteinase type plasminogen activators and their inhibitor in cornea. Biochem Biophys Res Commun 1989; 160: 1021-1025.
4. Ramby M L, Kreutzer D L. Fibrin induction of tissue plasminogen activator expression in corneal endothelial cells in vitro. Invest Ophthalmol Vis Sci 1993; 34: 3207-3219.
5. Berman M, Kenyon K, Hayashi K, L'Hernault N. The pathogenesis of epithelial defects and stromal ulceration. In: Cavanagh H D, ed. The Cornea. Transactions of the World Congress on the Cornea Ill. New York: Raven Press, 1988; 35-43.
6. Stevens J D, Marshall J M, Benjamin L, Cederholm-Williams S A, Bron J B. Plasminogen activator in human tears. Eye 1992; 6: 653-658.
7. Morimoto K, Mishima H, Nishida T, Otori T. Role of urokinase type plasminogen activator (u-PA) in corneal epithelial migration. Thromb Haemost 1993; 69: 387-391.
8. Dano K, Andreasen P A, Grondahl-Hasen J, Ktistensen P, Nielsen L S. Plasminogen activators, tissue degradation and cancer. Adv Cancer Res 1985; 44: 139-266.
9. Hart D A, Rehemutulla A. Plasminogen activators and their inhibitors: Regulators of extracellular proteolysis and cell function. Comp Biochem Physiol 1988; 90B: 691-708.
10. Saksela O, Rifkin D B. Cell-associated plasrninogen activator: regulation and physiological function. Annu Rev Cell Biol 1988; 4: 93-125.
11. Vassalli J D, Sappino A P, Belin D. The plasminogen activator/plasmin system. J Clin Invest 1991; 88: 1067-1072.
12. Flaumenhaft R, Moscatelli D, Saksela O, Rifkin D B. The role of extracellular matrix in the action of basic fibroblast growth factor: matrix as a source of growth factor for long-term stimulation of plasminogen activator production and DNA synthesis. J Cell Physiol 1989; 140: 75-81.
13. Saksela O, Rifldn D B. Release of basic fibroblast growth factor- heparan sulfate complexes from endothelial cells by plasminogen activator-mediated proteolytic activity. J Cell Biol 1990; 110: 767-775.
14. Vlodavsk I, Fuks Z, Michaeli R I, Bashkin P, Levi E, Korner G, Bar-Shavit R, Klagsbrun M. Extracellular matrix-resident basic fibroblast growth factor: implication for the control of angiogenesis. J Cell Biochem 1991; 45:167-176.
15. Lyons R M, Keski-Oja J, Moses H L. Proteolytic activation of latent transforming growth factor-beta from fibroblast conditioned medium. J Cell Biol 1988; 106: 1659-1665.
16. Naldini L, Tamagonen L, Vigna E, Sachs M, Hartmann G, Birchmeier W, Daikuhara Y, Tsubouchi H, BIasi F, Comoglio P M. Extracellular proteolytic cleavage by urokinase is required for activation of hepatocyte growth factor/scatter factor. ]EMBO J 1992; 11 : 4825-4833.
17. Lopez-Alemany R, Mirshahi S, Faure J P, Pouliquen Y, Burtin P, Mirshahi M. Binding of plasminogen to corneal, fibroblasts and their extracellular matrix, evidence for a receptor in cell membranes. Fibrinolysis 1995; 9: 223-229.
18. Friberger P, Knose M. Plasminogen determination in human plasma chromogenic peptide substrates. In: ScuUy M F, Kakar V V, eds. Chemistry and Clinical Usage. London: Churchill Livingstone, 1979: 128-140.
19. Montesano R, Pepper M S, Mohle-Stenlein U, Risau W, Wagner E F, Orci L. Increased proteolytic activity is responsible for the aberrant morphogenetic behavior of endothelial cells expressing the middle T oncogene. Cell 1990; 62: 435-445.
20. Laemmli U K. Cleavage of structural proteins during the assembly of the head of bactetiophage T4. Nature 1970; 227: 680-685.
© Pearson Professional Lid 1996 Fibrinolysis (1996) 10(4), 255-262

262 Mirshahi et al
21. Granelli-Pipemo A, Reich E A. Study ofprotease and protease inhibitor complexes in biological fluids. J Exp Med 1978; 148: 223-234.
22. Chomczynski P, Sacchi N. Single step method of RNA isolation by acid guanidinium. Anal Biochem 1987; 162: 156-159.
23. Pannekoek H, Veerman H, Lambers H, Diergaarde P, Verweij C L, Van Zonneveld A J, Van Mourik J A. Endothelial plasminogen activator inhibitor (PAl): a new member of the serpin gene family. EMBO J 1986; 5: 2539-2544.
24. Pennica D, Holmes W E, Kohr W J, Harkins R N, Vehar G A, Ward C A, Bennett W F, Yelverton E, Seeburg P H, Heyneker H L, Goddel D, Collen D. Cloning and expression of human tissue-type plasminogen activator cDNA in E. coli. Nature 1993; 301: 214-221.
25. Holmes W E, Pennica D, Blaber M, Rey M W, Guenzler W A, Steffens J G, Heyneker H L. Cloning and expression of the gene for pro-urokinase in Escherichia coll. Biotechnology 1985; 3: 923-929.
26. Faure J P, Mirshahi M, Dorey C, Thillaye B, de Kozak Y, Boucheix C. Production and specificity of monoclonal antibodies to retinal S-antigen. Curr Eye Res 1984; 3: 867-872.
27. Plough M, R6nne E, Behrendt N, Jensen A L, Blasi F, Dano K. Cellular receptor for urokinase-plasminogen activator. Carboxy terminal processing and membrane anchoring by glycosyl- phosphatidyl inositol. J Biol Chem 1991; 266: 1926-1933.
28. Pollanen J, Hedman K, Nielsen L S, Dano K, Vaheri A. Uhrastructural localization of plasma membrane-associated urokinase-type plasminogen activator at focal contact. J Cell Bio11988; 106: 87-95.
29. Myohanen H T, Stephens R W, Hedman K, Tapiovaara H, Ronne E, Hoyer-Hansen G, Dano K, Vaheri A. Distribution and
lateral mobility of the urokinase-receptor complex at the cell surface. J Histochem Cytochem 1993; 41 : 1291-1301.
30. Quigley J P, Gold L I, Schwimmer R, Sullivan L M. Limited cleavage of cellular fibronectin by plasminogen activator purified from transformed cells. Proc Natl Acad Sci USA, 1987; 84: 2776-2780.
31. Keski-Oja J, Vaheri A. The cellular target for the plasminogen activator, urokinase, in human fibroblasts-66000 dalton protein. Biochim Biophys Acta 1982; 720: 141-146.
32. Matsubara M, Girard M T, Kublin C L, Cintron C, Fini M E. Differential roles for two gelatinolytic enzymes of the matrix metalloproteinase family in the remodelling of cornea. Dev Biol 1991; 147: 425-439.
33. Knudsen B S, Harpel P C, Nachman R L. Plasminogen activator inhibitor is associated with the extracellular matrix of cultured bovine smooth muscle cells. J Clin Invest 1987; 80: 1082-1089.
34. Knudsen B S, Nachman R L. Matrix plasminogen activator inhibitor modulation of the extracellular proteolytic environment. J Biol Chem 1988; 263: 9476-9481.
35. Wilson S E, He Y G, Lloyd S A. EGF, EGF receptor, basic FGF, TGF beta-l, and II-1 alpha mRNA in human corneal epithelial cells and stromal fibroblasts. Invest Ophthalmol Vis Sci 1992; 33: 1756-1765.
36. Schultz G, Chegini N, Grant M, Khaw P, MacKay S. Effects of growth factors on corneal wound healing. Acta Ophthalmol 1992; 70, Suppl. 202: 60-66.
37. Li O~ Weng J, Mohan R R, Bennett G L, Schwall R, Wang Z F, Tabor K, Kim J, Hargrave S, Cuevas K H, Wilson S E. Hepatocyte growth factor and hepatocyte growth factor receptor in the lacrimal gland, tear and cornea. Invest Ophthalmol Vis Sci 1996; 37: 727-739.
Fibrinolysis (1996) 10(4), 255-262 © Pearson Professional Ltd 1996




















