
Endocytosis of a chimera between human pro-urokinase and the plant toxin saporin: an unusual...
10
Endocytosis of a chimera between human pro-urokinase and the plant toxin saporin: an unusual internalization mechanism RODOLFO IPPOLITI,* ,1 EUGENIO LENDARO,* PIER ALBERTO BENEDETTI, MARIA ROSARIA TORRISI, ‡ FRANCESCA BELLEUDI, ‡ DANIELA CARPANI, § MARCO RAFFAELLO SORIA, § AND MARIA SERENA FABBRINI § *Department of Biochemical Sciences ‘A. Rossi Fanelli’, University of Rome La Sapienza, Rome, Italy; ² C.N.R. Institute of Biophysics, Pisa, Italy; ‡ Department of Experimental Medicine and Pathology, University of Rome La Sapienza, Rome, Italy; and § DIBIT, Department of Biological and Technological Research-DIBIT, S. Raffaele Scientific Institute, Milano, Italy ABSTRACT A fluorescent derivative of a chimeric toxin between human pro-urokinase and the plant ribo- some-inactivating protein saporin (p-uPA-Sap TRITC ), has been prepared in order to study the endocytosis of this potentially antimetastatic conjugate in the murine model cell line LB6 clone19 (Cl19) trans- fected with the human urokinase receptor gene. The physiological internalization of urokinase-inhibitor complexes is triggered by the interaction of plasmin- ogen inhibitors (PAIs) with receptors belonging to the low density lipoprotein-related receptor protein (LRP) family, and involves a macro-quaternary struc- ture including uPAR, LRP, and PAIs. However, in contrast to this mechanism, we observed a two-step process: first, the urokinase receptor (uPAR) acts as the anchoring factor on the plasma membrane; sub- sequently, LRP acts as the endocytic trigger. Once the chimera is bound to the plasma membrane by interaction with uPAR, we suggest that a possible exchange may occur to transfer the toxin to LRP via the saporin moiety and begin the internalization. So an unusual endocytic process is described, where the toxin enters the cell via a receptor different from that used to bind the plasma membrane.—Ippoliti, R., Lendaro, E., Benedetti, P. A., Torrisi, M. R., Belleudi, F., Carpani, D., Soria, M. R., Fabbrini, M. S. Endocytosis of a chimera between human pro-urokinase and the plant toxin saporin: an un- usual internalization mechanism. FASEB J. 14, 1335–1344 (2000) Key Words: plasminogen activator z ribosome-inactivating proteins z receptors z ligand-passing z cancer Urokinase or urinary type plasminogen activator (uPA) is a serine protease that activates plasminogen to plasmin leading to cell matrix degradation. Sev- eral highly metastatic tumors overexpress the 55 kDa GPI-anchored uPA receptor (uPAR), such as breast cancers (1), melanomas (2), colon (3), and prostate carcinomas (4). uPAR is a receptor that preferen- tially binds species-specific Pro-uPA/uPA or its de- rived amino-terminal fragment (ATF) (5). However, ATF and the secreted precursor Pro-uPA (p-uPA) are not internalized, whereas uPAR-bound uPA internal- izes only if complexed to plasminogen activator inhibitors (PAIs). Endocytic receptors such as the a2-macroglobulin receptor or LDL-related receptor protein (LRP), the very low density lipoprotein re- ceptor, and the epithelial glycoprotein-330, are able to mediate the internalization of uPAR-bound uPA/ PAI-1 complexes (6, 7). Both uPAR and LRP are involved in the endocytosis (8); after ligand release, they recycle back to the plasma membrane (9). Saporin belongs to a class of monomeric plant seed ribosome-inactivating proteins (type I RIPs, 10) that catalyzes the in vitro depurination of a specific adenine residue in large ribosomal RNAs (11). Lack- ing a membrane binding subunit, such as in type II RIPs (i.e., ricin), they usually cannot enter cells by receptor-mediated endocytosis unless complexed to an appropriate carrier molecule; nevertheless, free saporin (and presumably other type I RIPs) can be internalized in some cells expressing LRP receptors (12), thereby acquiring cytotoxicity. A conjugate between active two-chain human uPA and native saporin was shown to internalize in the absence of plasminogen activator inhibitors (13) in cells ex- pressing both uPAR and LRP receptors. Moreover, preparation of an ATF-saporin recombinant chimera demonstrated that human ATF domain directs this chimera to human uPAR-bearing cells and that the toxin domain can mediate internalization (14). In fact, competition experiments showed that the sa- porin moiety is involved in the internalization of ATF-saporin through LRP receptors. Thus, neither 1 Correspondence: Department of Biochemical Sciences, University of Rome ‘La Sapienza’, P.le Aldo Moro 5, 00185 Rome, Italy. E-mail: [email protected] 1335 0892-6638/00/0014-1335/$02.25 © FASEB
-
Upload
universitalaquila -
Category
Documents
-
view
1 -
download
0
Transcript of Endocytosis of a chimera between human pro-urokinase and the plant toxin saporin: an unusual...

Endocytosis of a chimera between human pro-urokinaseand the plant toxin saporin: an unusualinternalization mechanism
RODOLFO IPPOLITI,*,1 EUGENIO LENDARO,* PIER ALBERTO BENEDETTI,†
MARIA ROSARIA TORRISI,‡ FRANCESCA BELLEUDI,‡ DANIELA CARPANI,§
MARCO RAFFAELLO SORIA,§ AND MARIA SERENA FABBRINI§
*Department of Biochemical Sciences ‘A. Rossi Fanelli’, University of Rome La Sapienza, Rome,Italy; †C.N.R. Institute of Biophysics, Pisa, Italy; ‡Department of Experimental Medicine andPathology, University of Rome La Sapienza, Rome, Italy; and §DIBIT, Department of Biological andTechnological Research-DIBIT, S. Raffaele Scientific Institute, Milano, Italy
ABSTRACT A fluorescent derivative of a chimerictoxin between human pro-urokinase and the plant ribo-some-inactivating protein saporin (p-uPA-SapTRITC),has been prepared in order to study the endocytosisof this potentially antimetastatic conjugate in themurine model cell line LB6 clone19 (Cl19) trans-fected with the human urokinase receptor gene. Thephysiological internalization of urokinase-inhibitorcomplexes is triggered by the interaction of plasmin-ogen inhibitors (PAIs) with receptors belonging tothe low density lipoprotein-related receptor protein(LRP) family, and involves a macro-quaternary struc-ture including uPAR, LRP, and PAIs. However, incontrast to this mechanism, we observed a two-stepprocess: first, the urokinase receptor (uPAR) acts asthe anchoring factor on the plasma membrane; sub-sequently, LRP acts as the endocytic trigger. Oncethe chimera is bound to the plasma membrane byinteraction with uPAR, we suggest that a possibleexchange may occur to transfer the toxin to LRP viathe saporin moiety and begin the internalization. Soan unusual endocytic process is described, where thetoxin enters the cell via a receptor different fromthat used to bind the plasma membrane.—Ippoliti,R., Lendaro, E., Benedetti, P. A., Torrisi, M. R.,Belleudi, F., Carpani, D., Soria, M. R., Fabbrini,M. S. Endocytosis of a chimera between humanpro-urokinase and the plant toxin saporin: an un-usual internalization mechanism. FASEB J. 14,1335–1344 (2000)
Key Words: plasminogen activator z ribosome-inactivatingproteins z receptors z ligand-passing z cancer
Urokinase or urinary type plasminogen activator(uPA) is a serine protease that activates plasminogento plasmin leading to cell matrix degradation. Sev-eral highly metastatic tumors overexpress the 55 kDaGPI-anchored uPA receptor (uPAR), such as breastcancers (1), melanomas (2), colon (3), and prostate
carcinomas (4). uPAR is a receptor that preferen-tially binds species-specific Pro-uPA/uPA or its de-rived amino-terminal fragment (ATF) (5). However,ATF and the secreted precursor Pro-uPA (p-uPA) arenot internalized, whereas uPAR-bound uPA internal-izes only if complexed to plasminogen activatorinhibitors (PAIs). Endocytic receptors such as thea2-macroglobulin receptor or LDL-related receptorprotein (LRP), the very low density lipoprotein re-ceptor, and the epithelial glycoprotein-330, are ableto mediate the internalization of uPAR-bound uPA/PAI-1 complexes (6, 7). Both uPAR and LRP areinvolved in the endocytosis (8); after ligand release,they recycle back to the plasma membrane (9).
Saporin belongs to a class of monomeric plantseed ribosome-inactivating proteins (type I RIPs, 10)that catalyzes the in vitro depurination of a specificadenine residue in large ribosomal RNAs (11). Lack-ing a membrane binding subunit, such as in type IIRIPs (i.e., ricin), they usually cannot enter cells byreceptor-mediated endocytosis unless complexed toan appropriate carrier molecule; nevertheless, freesaporin (and presumably other type I RIPs) can beinternalized in some cells expressing LRP receptors(12), thereby acquiring cytotoxicity. A conjugatebetween active two-chain human uPA and nativesaporin was shown to internalize in the absence ofplasminogen activator inhibitors (13) in cells ex-pressing both uPAR and LRP receptors. Moreover,preparation of an ATF-saporin recombinant chimerademonstrated that human ATF domain directs thischimera to human uPAR-bearing cells and that thetoxin domain can mediate internalization (14). Infact, competition experiments showed that the sa-porin moiety is involved in the internalization ofATF-saporin through LRP receptors. Thus, neither
1 Correspondence: Department of Biochemical Sciences,University of Rome ‘La Sapienza’, P.le Aldo Moro 5, 00185Rome, Italy. E-mail: [email protected]
13350892-6638/00/0014-1335/$02.25 © FASEB

catalytically active urokinase nor PAIs are needed toinitiate these internalization pathways.
In this paper we focus on the intracellular pathwayof a related chimeric toxin (p-uPA-SapTRITC), be-tween fluorescently labeled saporin and human pro-urokinase in murine cells transfected (15) with thehuman uPAR gene (Cl19). Again, this conjugate wasfound to be targeted to receptors (uPAR and LRP)that do not colocalize in control cells. uPAR is in fact,as previously shown, mostly confined to the leadingedge of migrating cells (16, 17), whereas LRP isdistributed among the whole plasma membrane (18)and inside clathrin-coated invaginations. Fluores-cence microscopy experiments show that bindingto the cell membrane is indeed mediated by uPAR,but on triggering of the endocytic process, p-uPA-SapTRITC seems to follow the fate of internalizedLRP.
In contrast to what is proposed for the endocytosisof uPA-PAI complexes, where uPAR and LRP recep-tors remain part of the same quaternary complex(presumably through the presence of PAI), we pro-pose that endocytosis of p-uPA-SapTRITC follows atwo-step process. Initially the p-uPA domain of thetoxin binds to uPAR, presumably triggering a signalthat induces migration of LRP toward uPAR; byvirtue of some conformational change, this is subse-quently coupled to an exchange of the chimerictoxin (mediated by the saporin domain) between thetwo receptors such as p-uPA-SapTRITC follows LRP inthe endocytic process.
MATERIALS AND METHODS
Standard recombinant human pro-uPA was courtesy of Dr. S.Toma, Pharmacia and Upjohn, Italy. Monoclonal R2 antibod-ies to human uPAR were a generous gift of Dr. K. Dano,Rigshospitalet, Denmark. Rabbit polyclonal serum againsthuman LRP was a generous gift from Dr. A. Nykjaer (Univer-sity of Aarhus, Denmark). Rhodamine isothiocyanate (mixedisomers), 2-iminothiolane, SPDP, chlorpromazine, and filipinIII were from Sigma/Aldrich. Fluorescein or rhodamine-labeled secondary antibodies were from Dako or Sigma/Aldrich.
Synthesis of the conjugate
Saporin was purified and labeled with rhodamine as de-scribed (19). The rhodaminated toxin was conjugated withp-uPA following the procedure of Cavallaro et al. (13) withsome modifications. SapTRITC was modified with 2.5-foldmolar excess of 2-iminothiolane (instead of SPDP) to intro-duce a free sulfydril group and allow the following reactionwith SPDP-modified p-uPA.
The p-uPA-SapTRITC conjugate was purified by HPLC on aion exchange column (Vydac VHP575) as described (13).
In vitro ribosome-inhibiting activity
The RIP activities of rhodaminated saporin and p-uPA-SapTRITC were measured in nuclease-treated rabbit reticulo-
cyte lysates (Promega, Madison, Wis.) measuring BMV mRNAtranslation as described (20). Values were expressed as per-cent of control 3H-leucine incorporation by the untreatedlysates. Activity was calculated by measuring the concentra-tion of toxin inhibiting mRNA translation by 50% (IC50).
Cell-killing experiments
At least two independent experiments were performed, usingmurine fibroblast LB6 and Cl19 expressing human uPARtreated as described (13). Cytotoxicity was calculated bymeasuring the dose of toxin inhibiting by 50% the incorpo-ration of the untreated cells (ID50). For the competitionexperiments, Cl19 cells were plated at 104 cells in 80 ml andexposed to 5 3 1029 M p-uPA-SapTRITC conjugate in theabsence or in the presence of increasing concentrations ofhuman recombinant pro-uPA (from 2531029 M to 25031029
M), for a total exposure of 2 h. Cells were acid-washed,incubated 16 h at 37°C, then pulse-labeled with 3H-leucinefor 4 h. Protein synthesis was not affected by pro-uPA expo-sure. Each experiment was performed in four replicates.
FACS scan analysis
Fluorescence-activated cell sorting was performed essentiallyas described (8) using aliquots of 106 U937 cells exposed 90min at 4°C either to 10 nM p-uPA alone and a preformeduPA-PAI1 complex as control or to 10 nM p-uPA-Sap TRITC.Briefly, after exposure to the ligands, the U937 cells werewashed extensively and duplicate samples warmed up to 37°Cfor 0, 5, 10, 15, 30, 45, 60, and 90 min. After transfer to an icebath to stop the internalization process, the samples werewashed and further incubated at 4°C with R2 MoAb (30mg/ml) or anti-LRP polyclonal antibodies, followed by incu-bation with FITC anti-mouse or FITC anti-rabbit secondaryantibodies, respectively. Background fluorescence was esti-mated with samples of cells exposed to the ligand (s) andincubated only with the secondary antibodies. Fluorescenceof 0.5–1 3 104 cells per sample was usually read in duplicatesamples.
Fluorescence microscopy
Topology of uPAR and LRP in control and p-uPA-SapTRITC-treated cells
Cl19 cells grown on coverslips were incubated with nothing orp-uPA-SapTRITC at a 300 nM concentration in D-MEM (LifeTechnologies, Inc.) containing geneticin sulfate G418 for 60min at 4°C. The cells were then washed with cold mediumand further incubated at 37°C in the humidified chamber (inthe presence of 5% CO2) for different times. The cells werethen fixed with 4% p-formaldehyde in phosphate-bufferedsaline (PBS) and permeabilized with 0.1% Triton X-100.Immunochemical staining of the two receptors was done byincubation of the cells with the monoclonal anti-uPAR (R2,0.125 mg/ml) or the rabbit polyclonal anti-LRP (0.05 mg/ml), followed by secondary antibodies conjugated with fluo-rescein or rhodamine.
Coverslips were then mounted on slides with Aqua poly-mount (Polyscience Inc., Warrington, Pa.) and observed witheither a Zeiss-Axiophot microscope connected to aHamamatsu Argus 20 video recording system or a confocalmicroscope (custom assembled by P. A. Benedetti).
1336 Vol. 14 July 2000 IPPOLITI ET AL.The FASEB Journal

Endocytosis of p-uPA-SapTRITC in the presence of R2 MoAbanti-uPAR
An experiment in the presence of the R2 MoAb anti-uPAR wascarried out incubating cells grown on coverslips with p-uPA-SapTRITC in the presence of the R2 MoAb anti-uPAR (125mg/ml) at 4°C for 60 min. After washing, cells were immedi-ately fixed or transferred to 37°C for further incubation andthen fixed. uPAR was visualized by addition of a secondaryanti-mouse antibody FITC-labeled.
Endocytosis of p-uPA-SapTRITC in the presence of filipin orchlorpromazine
Cells grown on coverslips were incubated when indicated with1 mg/ml Filipin III (Sigma) or with 10 mg/ml chlorpromazine(Sigma) in the culture medium for 30 min at 37°C. The cellswere incubated with 300 nM p-uPA-SapTRITC for another 30min, washed, fixed, and observed under the microscope.Where indicated, a fluorescent derivative of the toxin lectinRicin (RCAFITC, 300 nM) was used for the second incubation.
Endocytosis of p-uPA-SapTRITC followed in Cl19 living cells
In some experiments cells were incubated with p-uPA-SapTRITC as described above and further incubated withNBD-ceramide (21) for visualization of the Golgi apparatus inliving cells.
Immunoelectron microscopy
Cl19 cells were treated with p-uPA-SapTRITC 300 nM for 1 h at4°C and immediately fixed or warmed to 37°C for 5 min toallow internalization. Cl19 untreated cells were used as acontrol. All cells were washed twice with PBS and fixed in 4%paraformaldehyde for 30 min at 25°C. For single labelingexperiments, cells were scraped and incubated with anti-uPAR monoclonal antibody (0.2 mg/ml in PBS 1% bovineserum albumin (BSA) for 1 h at 4°C) or with anti-LRPpolyclonal antibody (0.5 mg/ml in PBS 1% BSA for 1 h at4°C). Cells were then washed extensively and labeled for 3 hat 4°C with colloidal gold (18 nm, prepared by citratemethod) conjugated with protein A (Pharmacia/Upjohn).For double labeling experiments, cells were scraped andincubated with anti-uPAR monoclonal antibody (0.2 mg/mlin PBS 1% BSA for 1 h at 4°C), followed by 10 nm goatanti-mouse immunoglobulin G gold conjugates and then withanti-LRP polyclonal antibody (0.5 mg/ml in PBS 1% BSA for1 h at 4°C), followed by 18 nm protein A gold particles.Samples were postfixed in 1% osmium tetroxide in Veronalacetate buffer, pH 7.4, for 2 h at 4°C, stained with uranylacetate (5 mg/ml), dehydrated in acetone, and embedded inEpon 812.
RESULTS
Cytotoxicity of p-uPA-SapTRITC
Both free SapTRITC and p-uPA-SapTRITC were testedfor their ability to inhibit protein synthesis in acell-free inhibition assay. They were found to havethe same activity (IC505 40 pM), demonstrating thatthe chemical modifications introduced in both sa-porin and pro-urokinase scarcely affected their bio-logical activity (see Table 1).
In cell killing experiments, the conjugate wasextremely cytotoxic for murine-transfected Cl19 cellsexpressing uPAR, with an ID50 value of 0.015 nM;free saporin showed an ID50 of 100 nM on the samecells, as previously shown (20).
When tested on the untransfected wild-type cells(LB6), p-uPA-SapTRITC had an ID50 of 2 nM (133-fold higher than in Cl19 cells) while Sap toxicity, asexpected, remained unchanged (ID505100 nM, Ta-ble 1). Furthermore, p-uPA-SapTRITC toxicity (overthree orders of magnitude higher in potency thanfree saporin) may be competed by addition of anexcess of free human p-uPA to Cl19 cells (data notshown).
The topology of uPAR and LRP in LB6 and Cl19cells
Immunocytochemical staining on wild-type LB6showed that nontransfected LB6 cells do not expresshuman uPAR, and indeed no staining was observedeither on the cell surface (not permeabilized cells)or intracellularly (with Triton X-100 permeabilizedcells). Instead, LRP was found throughout the mem-brane of both LB6 and Cl19 cells and heavily stainedthe perinuclear (ER) region in permeabilized cells(data not shown).
In Cl19 cells, human uPAR is overexpressed andappears to be localized in permeabilized cells in theperinuclear region, where it is colocalized with LRP(presumably as newly synthesized polypeptides). Onthe plasma membrane of nonpermeabilized cells,however, the two receptors are not colocalized,uPAR being mostly confined to the focal adhesion
TABLE 1. Comparison between the RIP activities and cytotoxicities of p-uPA-SapTRITC and SapTRITCa
Toxin p-uPA-SapTRITC SapTRITC
Cell free inhibition onrabbit reticulocytelysate
IC50 (nM) Means 6 sem 0,040 6 0,004 0,040 6 0,010
Cytotoxicity to C119 cells ID50 (nM) 6 sdb 0,015 6 0,005 100 6 10Cytotoxicity to w.t. LB6
cellsID50 (nM) 6 sdb 2 6 1 100 6 15
a Values are the means of two experiments each performed in duplicate. b Means of one representativeexperiment performed in quadruplicate.
1337TWO-STEP ENDOCYTOSIS OF A TOXIC CHIMERA

regions of the cell whereas LRP stained uniformlythe membrane and scarcely overlapped with uPAR(see Fig. 1).
p-uPA-SapTRITC binding and internalization in Cl19 cells
p-uPA-SapTRITC binds to Cl19 cells and israpidly internalized at 37°C (Fig. 2a). AlthoughsaporinTRITC is toxic to both LB6 and Cl19 cells (seeTable 1), we could not reveal its presence in fluores-cent spots as with p-uPA-SapTRITC-treated cells (seeFig. 2b). This difference can be attributed to thehigher efficiency of the binding and internalizationmediated by the presence of the pro-urokinase do-main conjugated to saporin. The fluorescence signaldue to the saporin molecule is quite clear (Fig. 2a)and cannot be confused with that due to autofluo-rescence (Fig. 2c).
As shown by confocal microscopy, after binding at4°C (see Fig. 3) the chimeric toxin (red fluores-cence) was mostly colocalized (Fig. 3a) with uPAR(green fluorescence) but scarcely (if at all; Fig. 3b)with LRP (green fluorescence).
When the temperature was raised to 37°C thechimera was internalized, and after 30 min (Fig. 3d)appeared in clearly visible spots (presumably endo-somes) that largely overlapped with the fluorescencesignal due to LRP localization. On the contrary,uPAR still seemed mostly confined to the edges ofthe membrane and did not appear to enter the cellafter endocytosis (Fig. 3c). Even when shorter timesof incubation at 37°C were used we could notobserve colocalization between uPAR and p-uPA-SapTRITC. The conclusion of these observations leadsus to hypothesize a different possible role of the tworeceptors, with uPAR being essential for binding tothe cell membrane and LRP for internalization.
To test this hypothesis, we followed binding andinternalization of p-uPA-SapTRITC in the presence ofa MoAb anti-uPAR (R2). This antibody does notcompete for the ligand binding site of p-uPA and hasno effect on the endocytic process as it has beenwidely used to follow the down-regulation of uPAR(8, 9). As shown in Fig. 4, the R2Ab signal (green)overlaps that of the toxin (red) at 4°C (Fig. 4a), butwhen p-uPA-SapTRITC is internalized as a function of
Figure 2. Internalization of p-uPA-SapTRITC conjugate fol-lowed in living Cl19 cells by fluorescence microscopy ascompared to free saporin. Cl19 cells were incubated with 300nM p-uPA-SapTRITC (A), 1 mM SapTRITC (B), or none (C) for30 min at 37°C in D-MEM containing G418. Fluorescencephotomicrographs (12003 magnification) were obtainedfor rhodamine-labeled saporin in (A, B) and autofluores-cence (C).
Figure 1. Immunolocalization of LRP and uPAR receptors onthe surface of Cl19 cells. The two receptors involved in theinternalization process of p-uPA-SapTRITC have been immu-nolocalized in nonpermeabilized control Cl19 cells and visu-alized by confocal fluorescence microscopy (20003 magnifi-cation); the color-merged image represents uPAR (greenfluorescence) and LRP (red fluorescence).
1338 Vol. 14 July 2000 IPPOLITI ET AL.The FASEB Journal

time at 37°C (Fig. 4b–d), the green signal remainsmainly confined to the edges of the membrane. Thepresence of the antibody does not induce anychange on the distribution nor does it trigger endo-cytosis of the receptor (data not shown).
We followed as control the cellular localization ofuPAR and LRP on binding and internalization of thecomplex uPA-PAI1, known conversely to induceuPAR endocytosis and recycling (8, 9). Indeed, thetwo receptors (red uPAR, green LRP) still appearedsegregated into different regions of the cell mem-brane after binding of uPA-PAI1 complex at 4°C(Fig. 4e), but showed a distinct colocalization after 30min at 37°C (Fig. 4f ).
Moreover, further evidences were obtained bycytofluorometric analysis of U937 human monocytesexposed to p-uPA-SapTRITC or the uPA-PAI1 com-plex, as a control. Down-regulation of uPAR afterbinding and internalization of the uPA-PAI1 com-plex was indeed observed as previously reported (9),whereas neither p-uPA alone nor p-uPA-SapTRITC
could induce any change in the levels of uPAR on
the cell membrane even at the long incubation times(data not shown).
Fluorescence microscopy in the presence of filipinand chlorpromazine
To further demonstrate that the internalization ofp-uPA-SapTRITC is mediated by endocytosis throughcoated pits and not by caveolae, the following exper-iments have been carried out in the presence offilipin and chlorpromazine. Filipin is a drug thatinterferes with the normal distribution of cholesterolby its sequestration. This process induces a substan-tial loss of ‘caveolae-like’ structures blocking trans-cellular transport in endothelial cells (22) andblocks the intracellular absorption of cholera toxin(23), a protein known to interact with gangliosideGM1 in the membrane and to follow the ‘clathrin-independent’ endocytosis pathway. Since uPAR is aGPI-anchored receptor localized in caveolae in somecells (24), we tested whether the presence of filipincould induce changes in the normal distribution of
Figure 4. Endocytosis of p-uPA-SapTRITC in Cl19 cells in thepresence of the MoAb anti-uPAR. a) Distribution after incu-bation at 4°C of both the p-uPA-SapTRITC conjugate (red) andthe R2 antibody (green); the same molecules are then shownafter incubation at 37°C as a function of time (b515, c530,d560) in minutes. In panels e (4°C) and f (309 at 37°C) arereported control experiments carried out by incubation ofthe cells with a preformed uPA-PAI1 complex, which is knownto induce uPAR internalization together with LRP. Thetwo receptors have been immunolocalized (green5LRP,red5uPAR). All images have been obtained at 12003 magni-fication.
Figure 3. Colocalization of p-uPA-SapTRITC with uPAR or LRPafter binding to the plasma membrane of Cl19 cells at 4°C,followed or not by internalization at 37°C. The confocalmicroscopic image (12003 magnification) shows the bindingof p-uPA-SapTRITC in Cl19 cells and its colocalization withLRP and uPAR receptors; the red signal refers to p-uPA-SapTRITC in all panels and the green signal refers to uPAR inpanels a, c and to LRP in panels b, d. As clearly visible, at 4°Cmost of the conjugate is overlapping (as evidenced by theyellow fluorescence) uPAR receptor (a) whereas it scarcelycolocalizes with LRP receptor (b), being the toxin decoratingthe edges of the cell membrane. On internalization at 37°C,the situation is reverted and p-uPA-SapTRITC fluorescenceoverlaps that of LRP (d) but scarcely that of uPAR (c).
1339TWO-STEP ENDOCYTOSIS OF A TOXIC CHIMERA
the receptor inside the membrane and whether theendocytic process of p-uPA-SapTRITC conjugate inCL19 cells could be specifically inhibited.
Filipin did not induce any significant redistribu-tion of either uPAR or LRP after 309 incubation at37°C (data not shown); furthermore, the drug didnot inhibit the binding and internalization of p-uPA-SapTRITC conjugate, as shown in Fig. 5c (comparewith Fig. 5a, as a control).
Chlorpromazine is a known inhibitor of clathrincoat assembly and completely blocks receptor-medi-ated endocytosis of many molecules (23, 25). In ourexperimental conditions, this drug could block in-ternalization (but not binding) of p-uPA-SapTRITC inCL19 cells (see Fig. 5e) as visualized by the appear-ance of patches on the cell membrane that con-tained the conjugate still bound to uPAR; indeed,uPAR distribution also changed after exposure tochlorpromazine at 37°C, with the appearance ofpatches (not shown). Surprisingly, LRP was notaffected by the presence of chlorpromazine in theabsence of ligand.
As a control we used fluorescently labeled ricin to
verify the activity of chlorpromazine and filipin. Asexpected, most of the toxin remains on the plasmamembrane in the presence of chlorpromazine (seeFig. 5f), whereas filipin seemed to have scarce effect(if any) on the endocytosis of the toxin (see Fig. 5d)that accumulated in the Golgi apparatus as in thecontrol cells (Fig. 5b).
Immunoelectron microscopy
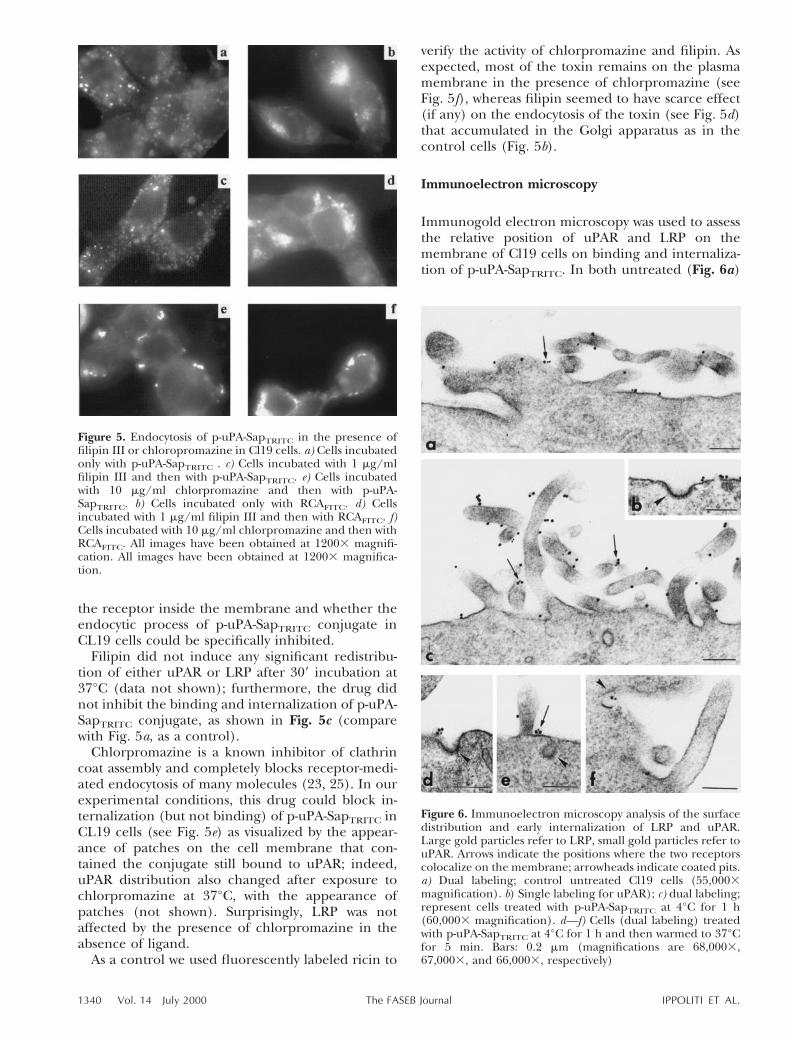
Immunogold electron microscopy was used to assessthe relative position of uPAR and LRP on themembrane of Cl19 cells on binding and internaliza-tion of p-uPA-SapTRITC. In both untreated (Fig. 6a)
Figure 6. Immunoelectron microscopy analysis of the surfacedistribution and early internalization of LRP and uPAR.Large gold particles refer to LRP, small gold particles refer touPAR. Arrows indicate the positions where the two receptorscolocalize on the membrane; arrowheads indicate coated pits.a) Dual labeling; control untreated Cl19 cells (55,0003magnification). b) Single labeling for uPAR); c) dual labeling;represent cells treated with p-uPA-SapTRITC at 4°C for 1 h(60,0003 magnification). d—f) Cells (dual labeling) treatedwith p-uPA-SapTRITC at 4°C for 1 h and then warmed to 37°Cfor 5 min. Bars: 0.2 mm (magnifications are 68,0003,67,0003, and 66,0003, respectively)
Figure 5. Endocytosis of p-uPA-SapTRITC in the presence offilipin III or chloropromazine in Cl19 cells. a) Cells incubatedonly with p-uPA-SapTRITC . c) Cells incubated with 1 mg/mlfilipin III and then with p-uPA-SapTRITC. e) Cells incubatedwith 10 mg/ml chlorpromazine and then with p-uPA-SapTRITC. b) Cells incubated only with RCAFITC. d) Cellsincubated with 1 mg/ml filipin III and then with RCAFITC. f)Cells incubated with 10 mg/ml chlorpromazine and then withRCAFITC. All images have been obtained at 12003 magnifi-cation. All images have been obtained at 12003 magnifica-tion.
1340 Vol. 14 July 2000 IPPOLITI ET AL.The FASEB Journal

or treated with p-uPA-SapTRITC at 4°C (Fig. 6b, c)Cl19 cells, the two immunogold-labeled receptorsappeared to be localized on the surface microvilli(Fig. 6, arrows) and they partly colocalized (largegolds refer to LRP, small golds to uPAR); in addition,the receptors appeared to be excluded from clath-rin-coated pits as shown in single labeling for uPAR(Fig. 6b, arrowhead).
During 59 at 37°C internalization after the bindingof the conjugate, we observed a colocalization of thetwo receptors in close proximity to clathrin-coatedpits (Fig. 6d, e, arrowheads), at the base of microvilli(Fig. 6e, arrows) and inside endocytic invaginations(Fig. 6f, arrowhead). Thus, at least at the verybeginning of the endocytic process, uPAR and LRPare moving together toward that region of the mem-brane where the endocytic pits are forming.
In vivo localization of p-uPA-SapTRITC
As shown in Fig. 7, in living (not fixed) cells the toxicp-uPA-SapTRITC was accumulated in vesicular struc-tures that did not overlap the fluorescence of theGolgi marker NBD-ceramide (21); the fluorescentspots varied in dimension and positions, startingfrom the plasma membrane down to the TGN,allowing one to hypothesize a bulk transport alongthe endosomal system to a possible final degradationin the lysosomes. No direct evidences were availableat this resolution as to a possible escape of freesaporin or the intact conjugate from the endocyticcompartment(s) to the cell cytoplasm.
DISCUSSION
UPAR-targeted chimeric toxins may have anti-cancerpotential (26) and recently a strategy has been
devised allowing to express a secretory version of arecombinant chimera between the amino-terminalfragment of human urokinase and saporin (ATF-Sap) in high amounts in eukaryotic cells (27). Thepossibility of investigating the intracellular pathways,followed by these chimeras represents a prerequisitefor optimizing and modulating their therapeutic use.To dissect and understand the sequence of stepsinvolved in binding and internalization and to studythe transport along the endomembrane system, weused the model cell line of murine cells Cl19 trans-fected with human uPAR, for which data are avail-able (3, 15) on uPAr and LRP levels (1.2310 5
and 1–5310 4 per cell respectively) and followedcell-surface binding and internalization of p-uPA-SapTRITC conjugate. This chimera represents aunique tool to study the interactions between uPARand LRP receptors and to obtain insights on intra-cellular routing of single chain RIPs such as saporin.The presence of uPAR confers to p-uPA-SapTRITC anextreme toxicity toward cells overexpressing thisreceptor, thus making the system very sensitive.
The receptor of urokinase-type plasminogen acti-vator is an adhesion/migration receptor that focusesthe urokinase activity at the leading edge of activelymigrating cell (28); it behaves as an activable cell-surface chemokine capable of triggering signaling ina variety of cell types (29–31); moreover, it hasrecently been demonstrated that cooperation be-tween uPA/uPAR and metalloproteinase MMP-9 isrequired for metastasis spread (32). uPAR and theendocytic receptor LRP normally do not colocalize,and indeed some signaling or ‘cross-talk’ betweenthese two receptors might exist: ATF has been shownto induce Cl19 cell migration, and these phenomenaare mediated by a signal transduction cascade (33,34). Moreover, ATF-Sap is efficiently endocytosed(14), and down-regulation of LRP by FACS-scananalysis after binding and internalization of therecombinant chimera to U-937 cells could be ob-served (M. S. Fabbrini, unpublished results), indicat-ing that LRP is actively involved in the endocytosis ofATF-Sap (and presumably of p-uPA-SapTRITC).
Physiological regulation of uPA activity on the cellmembrane is mediated by internalization of inhibi-tor complexes, such as uPA-PAI1, with a mechanisminvolving cointernalization of both uPAR and LRPreceptors (9); the saporin domain X-linked to p-uPAis expected to mimic the PAI1 function in anchoringthe complex to LRP, and thus one would expect thatp-uPA-SapTRITC followed an endocytic pathway simi-lar to uPA-PAI1. However, both fluorescence micros-copy on Cl19 and experiments by FACS scan analysisof U937 cells after exposure to p-uPA-SapTRITC indi-cated no down-regulation of uPAR. Altogether, ourresults seem to indicate that only LRP is responsiblefor the internalization of p-uPA-SapTRITC.
Figure 7. in vivo localization of p-uPA-SapTRITC in Cl19 cells.The red fluorescence refers to p-uPA-SapTRITC, and the greenone is due to NBD fluorescence staining the Golgi apparatusof living cells. (12003 magnification).
1341TWO-STEP ENDOCYTOSIS OF A TOXIC CHIMERA

Whereas the intracellular trafficking of chimerictoxins is generally dictated by the ligand moieties(19), in this case internalization seems to take placejust because of the presence of the toxin moiety,which binds to the endocytic receptor LRP. uPARbinding to the uPA moiety allows p-uPA-SapTRITC tofix on the plasma membrane, and the subsequenttransfer of the complex to LRP may require apossible cross-signaling between the two receptors.These results are indeed supported by data previ-ously available (35) on the relative affinities of p-uPAfor uPAr and LRP (5310211 and 1310 28 M, respec-tively) and by the observation that the presence ofuPAr completely abolishes the interaction of p-uPA-PAI1 complex with LRP.
Direct observation of p-uPA-SapTRITC in singlecells allowed us to follow all the phases involved inthe endocytic process, starting from the binding tothe plasma membrane. As clearly shown by confocalmicroscopy, the toxic conjugate binds only to uPARat 4°C, since binding does not overlap with LRPdistribution. By increasing the temperature to 37°C,endocytosis starts and the conjugate is internalized,but inside the cell it is mostly observed to coincidewith LRP, whereas uPAR is still confined at theleading edges of the cell membrane. The endocytosisof p-uPA-SapTRITC conjugate presumably proceedsfollowing the classical pathway mediated by theclathrin coat formation, as demonstrated by theinhibition induced by chlorpromazine. Our datamight indicate an interaction of uPAR with clathrin
components prior to ligand binding; on the con-trary, filipin alters the structure of the caveolae butdoes not inhibit p-uPA-SapTRITC internalization. Al-together, these results favor LRP as the main recep-tor involved in the endocytic process.
More evidence that uPAR may not be involvedin the intracellular pathway, followed by p-uPA-SapTRITC, is provided by the experiment in whichliving Cl19 cells were incubated with both the con-jugate and the monoclonal anti-uPAR at 4°C. In thiscondition the conjugate was internalized after in-creasing the temperature to 37°C, but the antibody(and hence the receptor) remained on the cellsurface.
Immunoelectron microscopy revealed that afterexposure at 4°C to p-uPA-SapTRITC, the two receptorsappear to be excluded from endocytic invaginations.Once the cells exposed to p-uPA-SapTRITC have beenwarmed to 37°C for 5 min, both LRP and uPAR movetoward the endocytic-forming pits and are present inunclosed invaginations, which may be defined ascoated pits.
We have never observed by fluorescence micros-copy any clear colocalization of uPAR and p-uPA-SapTRITC during internalization; however, the higherresolution of electron microscopy allows us to betterdiscriminate the earliest phases of this process. Apossible recycling of uPAR just below the plasmamembrane on triggering of endocytosis might besuggested, being the p-uPA-SapTRITC molecule pre-sumably transferred to LRP inside the forming en-
Figure 8. Schematic representation of the endocytosis of p-uPA-SapTRITC. a) The conjugate binds to uPAR via p-uPA domain;b) after binding, LRP could form a bridge with uPAR via the saporin moiety; c) endocytosis is triggered and coated vesicles areforming; d) uPAR may be rapidly released from the complex before (or immediately after) internalization and follow anindependent and rapid recycling just below the plasma membrane; e) the p-uPA-SapTRITC–LRP complex is accumulated inspotted structures (presumably endosomes) and larger vesicles (presumably lysosomes); f) most of the conjugate is presumablydegraded and LRP is recycled back to the plasma membrane. Somewhere along this pathway, part of the conjugated/freesaporin escapes from the vesicles, reaches the cytoplasm, and intoxicates the cell.
1342 Vol. 14 July 2000 IPPOLITI ET AL.The FASEB Journal

docytic vesicle. LRP might then recycle to the plasmamembrane whereas p-uPA-SapTRITC is presumablydelivered via the endosomal system to lysosomes,where degradation might take place. Indeed, bafilo-mycin A and chloroquine treatment of intoxicatedcells potentiated ATF-saporin cytotoxicity (14), sug-gesting a possible involvement of acidic compart-ments in the routing of the toxin. Part (not detect-able) of the conjugated or free saporin must escapethe vesicles and reach the ribosomes in a pathwaynot yet clarified. It must be stressed that the bulk ofp-uPA-SapTRITC does not to seem to reach the Golgicomplex. Also, recombinant ATF-Sap to which aKDEL sequence has been added was no more cyto-toxic than the parental ATF-Sap (M. S. Fabbrini etal., unpublished results) in U937 cells, suggestingthat the Golgi complex indeed might not be involvedin toxin routing, in contrast to what is known in thecase of ricin (36).
Based on the summary reported above, our hy-pothesis about the endocytic process of the toxicp-uPA-SapTRITC conjugate may be depicted as inFig. 8.
This mechanism of binding to the plasma mem-brane might resemble that described for tumornecrosis factor (TNF), where a dual step processinvolving two receptors for the TNF has been iden-tified and called ‘ligand passing’ (37): TNF binds onthe plasma membrane through the first receptor(P55); once transfer has occurred, the second recep-tor (P75) triggers the intracellular second messengercascade.
In this work we describe a somewhat similar mech-anism, although the ‘ligand passing’ event involvestwo different receptors: uPAR, which recognizes thepro-urokinase moiety warranting a correct anchor-age and orientation of p-uPA-SapTRITC on the plasmamembrane; and LRP, which binds saporin and ac-tively transports the conjugate inside the cell.
In conclusion, we have described a peculiar andnovel mechanism of endocytosis of a toxic conjugatediffering from that of the ligand used for cellulartargeting. Since many of the conjugated (immuno-or hormono-) toxins used in cancer therapy ap-proaches contain saporin (or similar type I RIPs; ref38), it is important to assess whether their targetingand toxicity may be influenced by the presence ofcooperative effects between receptors. In fact, al-though most research in this field has now con-verged to a careful molecular design to obtain re-combinant toxins (39) with a higher degree ofspecificity and toxicity, scarce attention has beendevoted to the study of molecular interactions at thelevel of the plasma membrane.
This work was partially supported by grants from MURST,C.N.R. (ctb. 97.04161.04, and Target Project on Biotechnol-ogy), and Associazione Italiana per la Ricerca sul Cancro,
Italy. We thank Prof. Maurizio Brunori for critical reading ofthe manuscript and Dr. Massimo Conese for useful discus-sions. We also thank Alessandra Masci for help in someexperiments, Sier Kees for recombinant PAI1, and SalvatoreToma for human recombinant p-uPA.
REFERENCES
1. Gronolahl-Hansen, J., Peters, H. A., van Putten, W. L. J., Look,M. P., Pappot, H., Ronne, E., Dano, K., Klijn, J. M. G., Brunner,N., and Foekens, J. A. (1995) Prognostic significance of thereceptor for urokinase plasminogen activator in breast cancer.Clin. Cancer Res. 1, 1079–1087
2. de Vries, T., Mooy, C., van Bakken, M., Luyten, G., Quax, P.,Verspaget, H., Weidle, U., Ruiter, D. J., and van Muijen, G.(1995) Components of the plasminogen activation system inuveal melanoma: a clinico-pathological study. J. Pathol. 175,59–67
3. Verspaget, H. W., Sier, C. F., Gonesh, S., Griffionen, G., andLamers, C. B. (1995) Prognostic value of plasminogen activatorsand their inhibitors in colorectal cancer. Eur. J. Cancer 31A,1105–1109
4. Crowley, C. W., Cohen, R. L., Lucas, B. K., Liu, G., Shuman,M. A., and Levinson, A. D. (1993) Prevention of metastasis byinhibition of the urokinase receptor. Proc. Natl. Acad. Sci. USA,90, 5021–5025
5. Blasi, F., Vassalli, J. D., and Dano, K. (1987) Urokinase-typeplasminogen activator: pro-enzyme, receptor and inhibitors.J. Cell Biol. 104, 801–804
6. Andreasen, P. A., Sottrup-Jensen, L., Kjoner, L., Nykjaer, A.,Moestrup, S. K., Petersen, C. M., and Gliemann, J. (1994)Receptor mediated endocytosis of plasminogen activators andactivator/inhibitor complexes. FEBS Lett. 338, 239–245
7. Heegaard, C. W., Simonsen, A. C., Oka, K., Kjoller, L., Chris-tensen, A., Madsen, B., Ellgard, L., Chan, L., and Andreasen,P. A. (1995) Very low density lipoprotein receptor binds andmediates endocytosis of urokinase-type plasminogen activator-type I plasminogen activator inhibitor complex. J. Biol. Chem.270, 20855–20861
8. Conese, M., Nykjaer, A., Petersen, C. M., Cremona, O., Pardi, R.,Andreasen, P.A., Gliemann, J., Christensen, E. I., and Blasi, F.(1995) a-2 macroglobulin receptor/LDL receptor-related pro-tein (Lrp)-dependent internalization of the urokinase receptor.J. Cell Biol. 131, 1609–1622
9. Nykjaer, A., Conese, M., Christensen, E. I., Olson, D., Cremona,O., Glieman, J., and Blasi, F. (1997) Recycling of the urokinasereceptor upon internalization of the uPA:serpin complexes.EMBO J. 16, 2610–2620
10. Barbieri, L., Battelli, M. G., and Stirpe, F. (1993) Ribosome-inactivating proteins from plants. Biochim. Biophys. Acta 1154,237–282
11. Endo, Y., Mitsui, K., Motizuki, M., and Tsurugi, K. (1987)Mechanism of action of the toxin lectin ricin on eukaryotic cells.The site and characteristics of the modification in 28S RNAcaused by the toxin. J. Biol. Chem. 262, 5908–5912
12. Cavallaro, U., Nykjaer, A:, Nielsen, M., and Soria, M. R. (1995)a-2 macroglobulin receptor mediates binding and cytotoxicityof plant ribosome-inactivating proteins. J. Biochem. 232, 165–171
13. Cavallaro, U., del Vecchio, A., Lappi, D. A., and Soria, M. R.(1993) A conjugate between human urokinase and a type Iribosome inactivating protein, is selectively cytotoxic in uroki-nase expressing cells. J. Biol. Chem. 268, 23186–23190
14. Fabbrini, M. S., Carpani, D., Bello-Riveiro, I., and Soria, M. R.(1997) The amino-terminal fragment of human urokinase di-rects a recombinant chimeric toxin to target cells: internaliza-tion is toxin mediated. FASEB J. 11, 1169–1176
15. Roldan, A. L., Cubellis, M. V., Masucci, M. T., Behrendt, N.,Lund, L. R., Dano, K., Appella, E., and Blasi, F. (1990) Cloningand expression of the receptor for human urokinase plasmino-gen activator, a central molecule in cell surface, plasmin depen-dent proteolysis. EMBO J. 9, 467–474
16. Myohanen, H. T., Stepheus, R. W., Hedman, K., Tapiovaara, H.,Ronne, E., HoyerHansen, G., Dano, K., and Vaheri, A. (1993)
1343TWO-STEP ENDOCYTOSIS OF A TOXIC CHIMERA

Distribution and lateral mobility of the urokinase-receptorcomplex at the cell surface. J. Histochem. Cytochem. 41, 1291–1301
17. Okada, S. S., Tomaszewski, J. E., and Barnathan, E. S. (1995)Migrating vascular smooth muscle cells polarize cell surfaceurokinase receptors after injury in vitro. Exp. Cell Res. 217,180–187
18. Moestrup, S. K. (1994) The a-2 macroglobulin receptor andepithelial glycoprotein-330: two giant receptors mediating en-docytosis of multiple ligands. Biochim. Biophys. Acta 1197, 197–213
19. Ippoliti, R., Lendaro, E., D’Agostino, I., Fiani, M. L., Guidarini,D., Vestri, S., Benedetti, P. A., and Brunori, M. (1995) Achimeric saporin-transferrin conjugate compared to ricin toxin:role of the carrier in intracellular transport and toxicity. FASEBJ. 9, 1220–1225
20. Fabbrini, M. S., Rappocciolo, E., Carpani, D., Solinas, M.,Valsasina, B., Breme, U., Nykjaer, A., Rovida, E., Legname, G.,and Soria, M. R. (1997b) Characterization of a saporin isoformwith lower ribosome-inactivating activity. Biochem J. 322, 719–727
21. Sleight, R. G., and Pagano, R. E. (1984) Transport of a fluores-cent phosphatidylcholine analog from the plasma membrane tothe Golgi apparatus. J. Cell Biol. 99, 742–751
22. Schnitzer, J. E., Oh, P., Pinney, E., and Allard, J. (1994)Filipin-sensitive caveolae-mediated transport in endothelium:reduced transcytosis, scavenger endocytosis, and capillarypermeability of select macromolecules. J. Cell Biol. 127,1217–1232
23. Orlandi, P. A., and Fishman, P. H. (1998) Filipin-dependentinhibition of cholera toxin: evidence for toxin internalizationand activation through caveolae-like domains. J. Cell Biol. 141,905–915
24. Stahl, A., and Mueller, B. M. (1995) The urokinase-type plas-minogen activator receptor, a GPI-linked protein, is localized incaveolae. J. Cell Biol. 129, 335–344
25. Wang, L. H., Rothberg, K. G., and Anderson, R. G. W. (1993)Mis-assembly of clathrin lattices on endosomes reveals aregulatory switch for coated pit formation. J. Cell Biol. 123,1107–1117
26. Andreasen, P. A., Kjoller, L., Christensen, L., and Duffy, M. J.(1997) The urokinase-type plasminogen activator system incancer metastasis: a review. Int. J. Cancer 72, 1–22
27. Fabbrini, M. S., Carpani, D., Soria, M. R., and Ceriotti, A. (2000)Cytosolic immunization allows expression at pre-ATF-Saporinchimeric toxin in eukaryotic cells. FASEB J. 14, 391–398
28. Conese, M., and Blasi, F. (1995) Urokinase/urokinase receptorsystem: internalization/degradation of urokinase-serpin com-plexes: mechanism and regulation. Biol. Chem. Hoppe-Seyler 376,143–155
29. Blasi, F., and Stoppelli, M. P. (1998) Proteases and cancerinvasion: from belief to certainty. Biochim. Biophys. Acta 1423,R35–R44
30. Nguyen, D. H., Hussaini, I. M., and Gonias, S. L. (1998) Bindingof urokinase-type plasminogen activator to its receptor in MCF7cells activates extracellular signal-regulated kinase 1 and 2 whichis required for increased cellular motility. J. Biol. Chem. 273,8502–8507
31. Konakova, M., Hucho, F., and Schleuning, W. D. (1998) Down-stream targets of urokinase-type plasminogen-activator-medi-ated signal transduction. Eur. J. Biochem. 253, 421–429
32. Kim, J., Yu, K., and Ossowski, L. (1998) Requirement for specificproteases in cancer cell intravasation as revealed by a novelsemiquantitative PCR-based assay. Cell 94, 353–362
33. Besser, D., Verde, P., Nagamine, Y., and Blasi, F. (1996) Signaltransduction and the uPA/uPAR system. Fibrinolysis 10, 215–237
34. Resnati, M., Guttinger, M., Valcamonica, S., Sidenius, N., Blasi,F., and Fazioli F. (1996) Proteolytic cleavage of the urokinasereceptor substitutes for the agonist-induced chemotactic effect.EMBO J. 15, 1572–1580
35. Nykjaer, A., Kjoller, L., Cohen, R. L., Lawrence, D. A., Garni-Wagner, B. A., Todd, R. F., van Zonneveld, A.-J., Gliemann, J.and Andreasen, P. A. (1994) Regions involved in binding ofurokinase-type I inhibitor complex and pro-urokinase to theendocytic a2-macroglobulin receptor/low density lipoproteinreceptor-related protein. J. Biol. Chem. 269, 25668–25676
36. Lord, M. J., and Roberts, L. (1998) Toxin entry—retrogradetransport through the secretory pathway. J. Cell Biol. 140,733–736
37. Pinckard, J. K., Sheehan, K. C. F., and Schneiber, R. D. (1997)Ligand-induced formation of P55 and P75 tumor necrosis factorreceptor heterocomplexes on intact cells. J. Biol. Chem. 272,10784–10789
38. Flavell, D. J. (1998) Saporin immunotoxins. Curr. Top. Microbiol.Immunol. 234, 57–61
39. Reiter, Y., and Pastan, I. (1998) Recombinant FV immunotoxinsand FV fragments are novel agents for cancer therapy anddiagnosis. Trends Biotech. 16, 513–520
Received for publication August 25, 1999.Revised for publication December 16, 1999.
1344 Vol. 14 July 2000 IPPOLITI ET AL.The FASEB Journal




















