
Obesity-Associated Differentially Methylated Regions in Colon ...
Upload
independentCategory
view
0download
0
Int. J. Cancer: 58,650-657 (1994) Publication of the International Union Against Cancer Publication de I'Union Internationale Contre le Cancer 0 1994 Wiley-Liss, Inc.
TRANSCRIPTIONAL ACTIVATION OF THE UROKINASE RECEPTOR GENE IN INVASIVE COLON CANCER Heng WANG', John SKIBBER~, Jose JUAREZ' and Douglas BOYD1,3 Departments of 'Tumor Biology and 2Surgical Oncology, M.D. Anderson Cancer Center, Houston, TX 77030, USA.
The plasminogen activator urokinase promotes tumor inva- sion by converting plasminogen into plasmin, which degrades several extracellular matrix components. Urokinase can bind to a specific cell surface receptor, which leads to accelerated plasmin production. While there is good evidence indicating a role for this binding site in tumor invasion/metastasis, there is little information concerning the regulation of urokinase recep- tor expression in invasive cancer. To address this question a series of colon cancer cell lines, which demonstrate either a high or low ability to invade an extracellular matrix-coated porous filter, was characterized for receptor expression at the transcrip- tional and post-transcriptional levels. The invasive cell lines possessed 10-fold more receptors than their non-invasive coun- terparts as shown by cross-linking experiments and by Western blotting. Northern blotting indicated that this disparity in receptor number could be largely accounted for by a different amount of steady-state mRNA encoding the binding site. How- ever, neither gene amplification nor enhanced mRNA stability could account for the augmented receptor protein observed for the invasive colon cancer cell types. In contrast, nuclear run-on experiments with representative cell lines revealed that the 10-fold difference in receptor display between the invasive- competent and invasive-deficient cells could be largely ac- counted for by differences in transcription rates. Transcription of the U-PAR gene in the receptor-deficient GEO cells, but not in the receptor-rich RKO cells, could be augmented by protein kinase C stimulation. These findings provide a clear rationale for studies to determine if the urokinase receptor promoter in invasive colon cancer is activated in cis or in trans. o 1994 Wiley-Liss, Inc.
The ability of colon cancer to spread to distant sites, including the liver, is ultimately the cause of death for patients with this disease. Although the mechanism(s) responsible for tumor cell spread are extremely complex and not entirely understood at the present time, the degradation of the basement membrane and surrounding extracellular matrix represents the first step in this process (Tryggvason et al., 1987). This is accomplished by the action of one or more hydrolytic enzymes that cleave key constituents within these structures (Tryggvason et al., 1987).
The plasminogen activator urokinase (u-PA) promotes tu- mor invasion and metastasis (Ossowski and Reich, 1983) by converting the abundant zymogen, plasminogen, into plasmin, which degrades basement membrane laminin (Trygaason et al., 1987). Several studies have shown that u-PA derived from either the tumor cell or from the stromal compartment (Pyke et al., 1991) can bind to a cell surface receptor (u-PAR) specifi- cally and with high affinity (KD = 0.2-1.0 nM) (Nielsen et al., 1988). The receptor binds u-PA via the amino-terminal frag- ment (ATF) of the A chain of the PA (Appella et al., 1987), allowing for plasminogen activation by the catalytic site on the B chain. u-PA bound to its receptor does not internalize, thus concentrating the plasminogen activator at the cell surface. In fact, receptor-equipped cells have a distinct proteolytic advan- tage over their receptor-deficient counterparts since receptor- bound u-PA is a much more efficient converter of plasminogen than fluid-phase PA (Ellis et al., 1991).
There is a substantial body of evidence suggesting that receptor display is a critical determinant of tumor invasion and metastasis. Perhaps the most compelling evidence for this comes from a study by Kariko et al. (1993), who reported that the overexpression of an exogenous cDNA encoding the
binding site in a human osteosarcoma cell line led to a 4-fold increase in cell migration through an extracellular matrix- coated filter; this increase was paralleled by augmented lami- nin degradation. In a separate study, invasion of reconstituted basement membrane by the ovarian cancer cell line HOC-1 was inhibited by peptide fragments that corresponded to the receptor-binding fragment of u-PA (Kobayashi et al., 1993).
The receptor gene encodes a 1.4 kb mRNA (Roldan et al., 1990). The mature 313 amino acid urokinase receptor protein (45-55 kDa), which is heavily glycosylated, is comprised of 3 repeating domains (Nielsen et al., 1988; Behrendt et al., 1991). The amino terminal domain represents the ligand-binding portion of the receptor, while the carboy terminus covalently links the binding site to the cell membrane via a glycolipid anchor (Behrendt et al., 1991).
Studies of resected human tumors have demonstrated the presence of the receptor on the malignant cells. In this regard, Del Vecchio et al. (1993) showed by immunohistochemical staining that the binding site was localized to the invading tumor cells present in breast cancers. Similarly, Pyke et al. (1991) demonstrated the presence of u-PAR mRNA in invad- ing colon cancer cells but not in malignant cells distal to the leading edge of the tumor. Thus, it is becoming increasingly apparent that elevated urokinase receptor expression is a requirement for tumor invasion and, indeed, characterizes this phenotype both in vitro (Hollas et al., 1991) and in vivo (Del Vecchio et al., 1993; Pyke et al., 1991). However, the mecha- nism(s) that underscores this increased receptor synthesis in invasive cancer is poorly understood at the present time. This is an important issue to address from the perspective of developing novel anti-invasive/anti-metastatic agents that act by suppressing the production of the binding site. Conse- quently, we undertook a study to characterize, at the transcrip- tional and post-transcriptional levels, the expression of u-PAR in invasive-competent and invasive-deficient colon cancer cell lines previously shown by us to differ in receptor display by 10-fold (Schlechte et al., 1989; Hollas et al., 1991). We report, herein, that transcriptional activation of the u-PAR gene, rather than gene amplification or mRNA stability, is the primary mechanism for the elevated expression of the receptor in invasive colon cancer.
MATERIAL AND METHODS Cell culture
The colon cancer cell lines used in the proposed study were maintained in McCoy's medium 5A supplemented with 5% FCS. CBS, GEO and FET cells display a modest number (10,000/cell) of urokinase receptors (Schlechte et al., 1989) and are weakly invasive, with less than 1% of the cells
3To whom correspondence and reprint requests should be ad- dressed, at the Department of Tumor Biology, M.D. Anderson Cancer Center, 1515 Holcombe Blvd., Houston, TX 77030, USA. Fax: 713 794 0209
Dedicaiion: This communication is in remembrance of M. Gross, who dedicated his short life to scientific pursuit. May he rest in peace.
Received: March 1,1994 and in revised form March 31, 1994.

UROKINASE RECEPTOR EXPRESSION IN COLON CANCER 65 1
penetrating an extracellular matrix-coated filter in a 72 hr period (Hollas et al., 1991). In contrast, HCT 116 and RKO cells, which are equipped with over 100,000 binding sites/cell (Schlechte et al., 1989), are highly invasive: 23 and 6% of the ce:lls invade this impermeable barrier, respectively (Hollas et al., 1991).
Quantitation of u-PAR in resected and in cultured colon cancer Resected tissue, verified pathologically as malignant or
normal, was washed thoroughly with ice-cold PBS and, after freezing with liquid nitrogen, pulverized and homogenized in ice-cold buffer containing 5 mM Tris-C1 (pH 7.5), 5 mM NaCI, 2.5 mM EGTA, 1 mM MgC12, 10 KIU/ml aprotinin and 50 yg/ml phenyl methane sulfonyl fluoride (PMSF). Tissue de- bris was removed at 5,OOOg and membranes recovered at 100,OOOg. The membranes were extracted in the above buffer containing 1% Triton-X 100 at 4°C for 30 min, after which the membranes were sedimented and the extract (supernatant) retained for analysis. Cultured cells were extracted with an identical buffer.
The tissue extracts were analyzed for u-PAR using a commercially available kit (IMUBINDe, American Diagnos- tics, Greenwich, CT).
Extracts from cultured colon cancer were subjected to immunoblotting. Equal amounts of protein were denatured at 100°C in SDS gel-loading buffer containing 62.5 mM Tris-C1 (pH 6.8), 2.0% SDS, 0.0025% bromophenol blue and 10% glycerol. The denatured material was electrophoresed in a 10.5% polyacrylamide gel and the resolved proteins trans- ferred to a nitrocellulose filter. The filter was blocked with PBS Containing 0.05% Tween 20 (PBS/Tween), and 3% BSA and incubated with 1 pg/ml of a polyclonal antibody (399-R, American Diagnostica) to the u-PA receptor. Subsequently, blots were incubated with 50,000 dpm/ml of radioactive Protein A, washed extensively with PBS/Tween and exposed to x-ray film.
Immunohistochemical staining of tumor sections for urokinase receptor
Tissue sections (4 pm) were, after blocking with normal goat serum, incubated with a monoclonal antibody (3936) to the u-PA receptor (50 pg/ml) for 1 hr at ambient temperature. Visualization of the immune complexes was accomplished using alkaline phosphatase conjugated to streptavadin and fast red chromogen as substrate. Sections were counterstained with hematoxylin.
Cross-linking of radioactive ATF to the cell surface of cultured colon cancer
The amino terminal fragment (ATF-amino acids 6-135 of u-PA) (Appella et al., 1987) prepared by autodigestion of two-chain u-PA and purified by reverse phase HPLC was kindly provided by Dr. J. Henkin (Abbott, Chicago, IL). Near confluent (95%) cells were acid pretreated and incubated with 2.0 nM of (ATF). In some instances, a 50-fold excess of unlabelled ATF was included in the incubation mixture. At the end of the incubation period, the cells were washed with PBS. The cross-linking agent DSS was added to achieve a final concentration of 1 mM and, after 15 min at ambient tempera- ture, the reaction was terminated with 10 mM ammonium acetate. The cells were extracted at 4°C for 1 hr with a solution containing 0.5% CHAPS, 0.1 M Tris (pH 8.1), 10 mM EDTA, 1 mM PMSF and 10 pg/ml of aprotinin. The extract was clarified by centrifugation, heated at 95°C in the presence of 2.3% SDS and 5% p-mercaptoethanol and equal amounts (10-50 pg) of protein electrophoresed in a 12.5% polyacryl- amide gel. The gel was subsequently dried and subjected to au tor adiogr aphy .
Northern blotting for u-PAR mRNA The level of steady-state u-PA receptor transcript expressed
by the colon cancer cells was determined by Northern analysis. Total cellular RNA was extracted from 90% confluent cultures using 5.0 M guanidinium isothiocyanate and purified on a cesium chloride cushion (5.7 M) by centrifugation at 150,OOOg for 20 hr. Purified RNA was electrophoresed in a 1.5% formaldehyde-agarose gel and transferred to Nytran-modified nylon by capillary action using 10 x SSC. The Northern blot was probed at 42°C with a random primed radiolabeled 0.65 kb cDNA specific for the u-PAR transcript and kindly provided by Dr. D. Cines, the University of Pennsylvania. The cDNA spans 650 nucleotides starting at the transcription initiation site and extending 3‘ (as delineated by an Apa I restriction site). The cDNA is in the pBC12B1 vector. Following hybrid- ization, stringency washes were undertaken at 65°C using 0.25 X SSC in the presence of 0.75% SDS. Loading efficiencies were checked by reprobing the blot with a radioactive cDNA that hybridizes with the glyceraldehyde phosphate dehydroge- nase (GAPDH) mRNA.
For mRNA stability studies, colon cancer cells were treated for the indicated times with 5 pg/ml of actinomycin D and extracted for RNA as described above. Southern blotting for the u-PAR gene
This was carried out essentially as described by this labora- tory (Hollas et al., 1991). Cell pellets were digested at 50°C overnight with 0.1 mg/ml proteinase K in a buffer containing 0.5% SDS, 25 mM EDTA, 10 mM NaCl and 10 mM Tris-C1 (pH 8.0). Genomic DNA was purified by multiple extractions with phenol/chloroform:isoamyl-alcohol(24: 1). Residual RNA was removed with 1 pg/ml DNAase-free RNAase (37”C, 1 hr) in the presence of 0.1% SDS. DNA (10 pg) was digested overnight at 37°C with 75 U of Nco I restriction endonuclease. The digested DNA was electrophoresed in a 1% agarose gel. The DNA in the gel was transferred to a nylon filter by capillary action after acid and base treatments. Probing of the blot for the u-PA receptor cDNA was accomplished using a radioactive cDNA as described in the section for Northern blotting. Nuclear run-on experiments
Nuclear run-on experiments were carried out as follows. Cells (lo8) were scraped into PBS and resuspended in a hypotonic buffer containing 10 mM Hepes, pH 7.9, 0.75 mM spermidine, 0.15 mM spermine, 0.1 mM EDTA, 0.1 mM EGTA, 1 mM D T I and 0.5% NP-40. The cells were lysed with the aid of a Dounce homogenizer and, following centrifuga- tion, the nuclear pellets resuspended in transcription buffer (150 mM KCI, 5 mM MgC12, 1 mM MnC12, 20 mM Hepes, pH 7.9, 10% glycerol and 5 mM DTT). Nuclei were incubated at 20°C for 30 min in the presence of 1 mM ATP, GTP, CTP and 200 pCi 32[P] dUTP. Nuclei were collected by centrifugation and treated twice with DNAase I and proteinase K. RNA was treated with phenol/chloroform and precipitated. Radioactive RNA was treated with DNAase I and proteinase K, again followed by phenol/chloroform. Newly transcribed RNA (6 x 107 cpm/reaction) was incubated at 42°C for 42 hr with nylon-immobilized cDNAs (5 kg/mm2) corresponding to u- PAR (identical sequence to that used for Northern blotting), a housekeeping mRNA (GAPDH) or the linearized plasmid. After hybridization, the filter was washed at 65°C with 0.1 X SSC in the presence of 1% SDS and exposed to x-ray film.
RESULTS Elevated production of u-PAR by invasive colonic cancer
We quantitated u-PAR in resected tumor tissue and matched adjacent “normal” tissue by an ELISA. In 8 of the 10 colonic tumors (Table I) examined, the malignant tissue had 30400% more u-PAR than the non-malignant adjacent tissue, as determined by the ELISA (Fig. 1). Interestingly, all of these

652 WANG ETAL.
TABLE I - DESCRIPTION OF RESECTED COLORECTAL TUMORS
Patient Tumor histology Tumor staging number site
278622 Colon
268867
282832 Colonic
282015 Rectal
288131 Colon
260724 Colon
287818 Rectum
284505 Colon
281981 Colon
292130 Colon
Moderately differ- entiated
Moderately differ- entiated
Moderately to poorly differenti- ated
Arising from tubu-
Moderately differ-
Moderately differ-
Moderately differ-
Tubular and tubu-
Moderately differ-
Poorly differenti-
lovillous ad e n o m a
entiated
entiated
entiated
lovillous adenoma
entiated
ated
2 cm depth, inva- sion through muscularis pro- pria
Extends through muscle wall, perineural inva- sion
Metastatic carci- noma with colon primary in liver parenchyma, car- cinoma in peri- colonic adipose tissue
Extension through muscularis pro- pria
Transmurally inva- sive w i metastasis Infiltrating through muscularis propria
Invasive with metastasis in 1 of 3 lymph nodes
Lymphatic, vas- cular invasion, extension to renal causule
::1 20 UROKINASE RECEPTOR Inaimgl TUMOR
UROKINASE RECEPTOR Ingimgl NORMAL
PATIENT NUMBER
FIGURE 1 - Determination of u-PA receptor levels in extracts of resected colon cancer by an ELISA. Membranes from tumor and paired normal adjacent mucosa were extracted into a buffer containing 1.0% Triton X-100 detergent and centrifuged at 100,000 x gav. The extracted material was analyzed for receptor levels by an ELISA as described by the manufacturer.
tumors were determined to be highly invasive, especially that of 292130, which had pronounced lymphatic and vascular invasion as well as extension to the renal capsule. In contrast, specimens 282015 and 284505, which were tubulovillous adeno- mas, had low levels of the binding site in both the tumor and the “normal” adjacent tissue. To determine the localization of u-PAR in the resected colonic specimens, sections of resected tissue were stained immunohistochemically using a mono- clonal antibody that recognizes an epitope at, or near to, the ligand-binding domain of the binding site as evidenced by its ability to inhibit the binding of radioactive sc-uPA to a colon
cancer cell line (Fig. 2a). Examination of sections of resected invasive tumor indicated that the u-PAR protein was exclu- sively localized to the malignant cells of the invasive tumor (Fig. 2b, c). In contrast, there was little evidence of staining of non-involved mucosa or of fibroblasts (Fig. 2b, c). The majority of u-PAR staining was localized to the basal aspect of the malignant cells (see arrows in Fig. 2d), as would be expected for a molecule involved in cell detachment. These results extend the finding of Pyke et al. (1991), who demonstrated the presence of u-PAR mRNA in invasive colonic cancer. Put together, these findings indicate that an elevated level of u-PAR is an accompaniment of the invasive phenotype of colon cancer.
Differential display of u-PAR by highly and by poorly invasive colon cancer
While there is compelling evidence implicating the u-PA receptor in the invasive phenotype demonstrated by a number of malignancies, including colon cancer, little however is known concerning the molecular mechanism(s) responsible for the high levels of urokinase receptor observed in infiltrating cancers. Such information could, ultimately, lead to the design of novel anti-invasive agents that act by repressing the synthe- sis of the binding site.
To determine the mechanism(s) by which u-PAR produc- tion is augmented in invasive colon cancer, we made use of a bank of cultured colon cancer cell lines comprised of cell types that demonstrate either a high or a low ability to invade an extracellular matrix-coated filter (Hollas et al., 1991). GEO, FET and CBS, which are poorly invasive, display 110,000 receptors/cell, while the invasive HCT 116 and the RKO cell lines are equipped with > 100,000 binding sites/cell as deter- mined by radioligand binding assay (Schlechte et al., 1989).
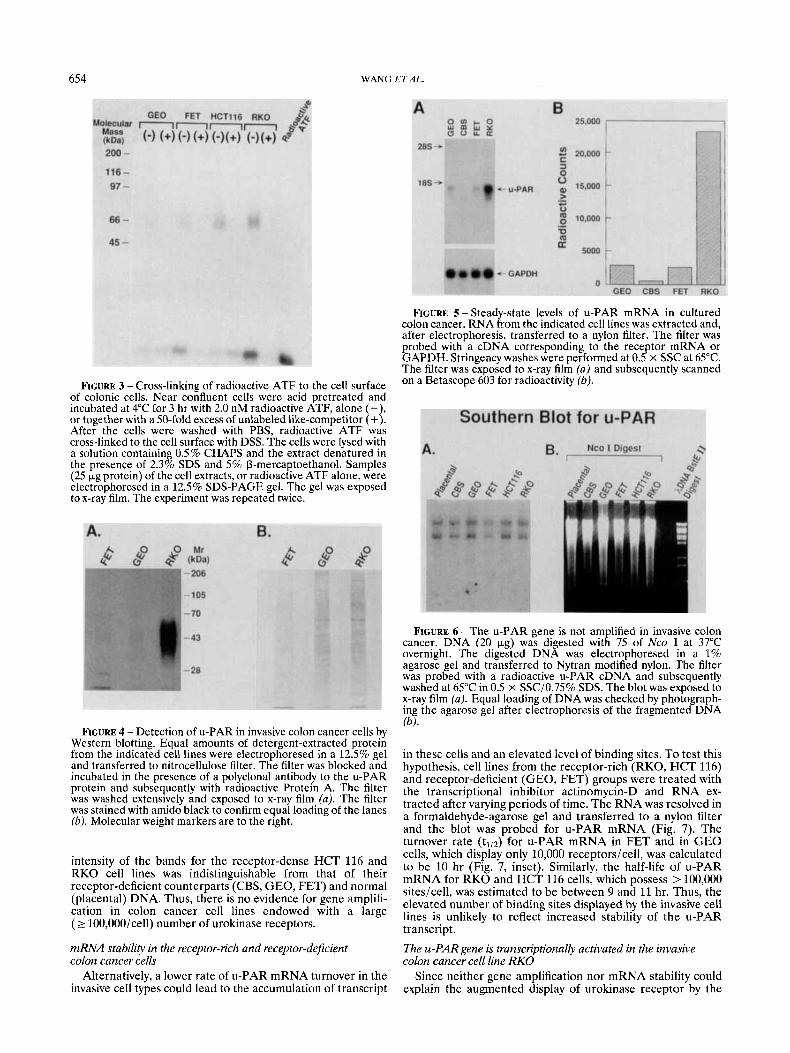
To confirm the identity of the receptor, we performed cross-linking and Western blotting experiments. The ATF of u-PA, which represents the receptor-binding portion of the plasminogen activator (Appella et al., 1987), was cross-linked to acid-pretreated cultured colon cancer cells and the cells subsequently lysed with detergent. Equal amounts of protein were subjected to SDS-PAGE and the gel autoradiographed (Fig. 3). A band of 60-65 kDa, which could be abolished with an excess of unlabeled ATF, was detected in the extracts of the invasive RKO and HCT 116 cell lines. The 45-50 kDa molecular size of the u-PAR (after subtraction of the 15 kDa of the ATF) is indistinguishable from that published for u-PAR displayed on the surface of U937 monocytes (Nielsen et al., 1988). In contrast, this band was either absent or barely visible in extracts of the poorly invasive GEO and FET cell lines. These results agree well with the Western blotting data, which showed that the invasive RKO cell line had a substan- tially higher amount of u-PAR than its poorly invasive GEO and FET counterparts (Fig. 4). These data suggest that the highly and the poorly invasive colon cancer cell lines can, indeed, be distinguished on the basis of their different number of urokinase receptors.
u-PAR display correlates with mRNA levels To determine if the differential u-PAR display by the
invasive-competent and invasive-deficient colon cancer cells was a consequence of a different amount of receptor tran- script, RNA was extracted from the cells and blotted for the u-PAR mRNA (Fig. 5). A 1.4 kb band, identical in size to that reported for u-PAR mRNA from U937 cells (Roldan et al., 1990), was detected in all of the cell lines examined. However, the RKO cell line, which displays > 100,000 binding sitesicell, had > 10 times as much u-PAR mRNA than GEO, CBS and FET cells, which are equipped with only 10,000 receptorsicell. Thus, u-PAR receptor display appears to be a function of the amount of steady-state mRNA encoding the binding site.
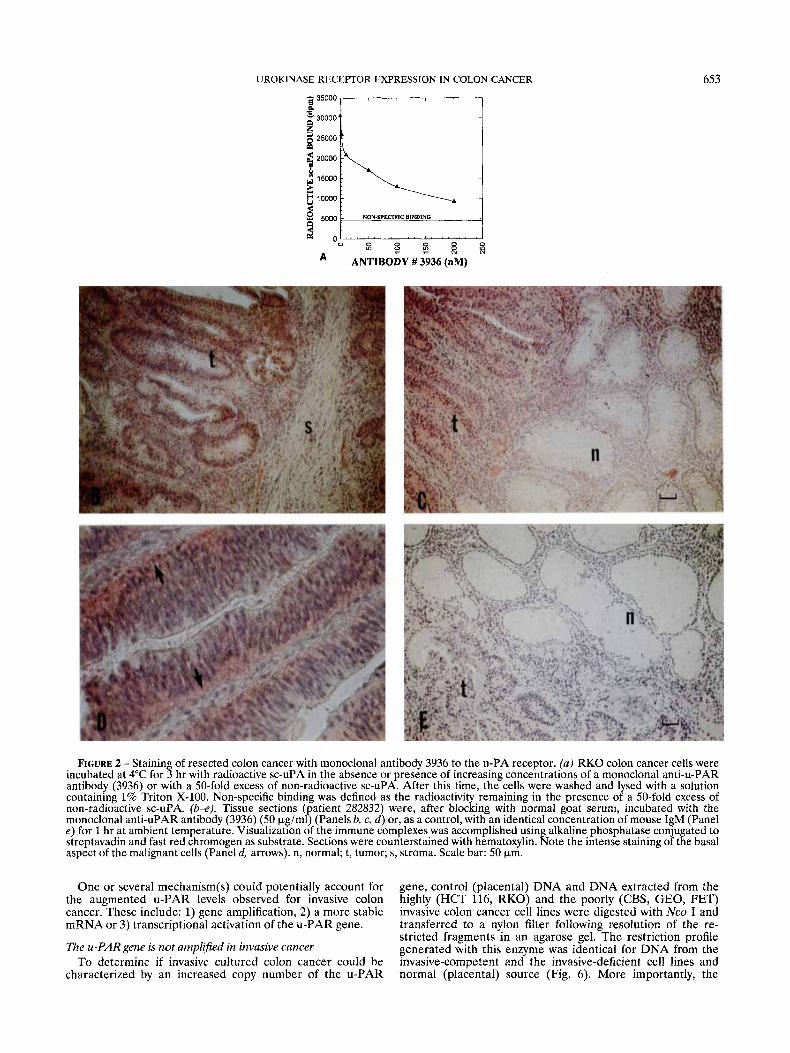
UROKINASE RECEPTOR EXPRESSION IN COLON CANCER 653 p 35000
30000
25000 I
5
A 4
L NON-SPECIFIC BINDING
" Z W W 00' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' '
A ANTIBODY # 3936 (nM)
FIGURE 2 - Staining of resected colon cancer with monoclonal antibody 3936 to the u-PA receptor. (a) RKO colon cancer cells were incubated at 4°C for 3 hr with radioactive sc-uPA in the absence or presence of increasing concentrations of a monoclonal anti-u-PAR antibody (3936) or with a 50-fold excess of non-radioactive sc-uPA. After this time, the cells were washed and lysed with a solution containing 1% Triton X-100. Non-specific binding was defined as the radioactivi remaining in the presence of a 50-fold excess of
monoclonal anti-uPAR antibody (3936) (50 p @ ~ l ~ (Panels b, c, d ) or, as a control, with an identical concentration of mouse IgM (Panel e ) for 1 hr at ambient temperature. Visualization of the immune complexes was accomplished using alkaline phosphatase conjugated to streptavadin and fast red chromogen as substrate. Sections were counterstained with hematoxylin. Note the intense staining of the basal aspect of the malignant cells (Panel d, arrows). n, normal; t, tumor; s, stroma. Scale bar: 50 pm.
non-radioactive sc-uPA. (k), Tissue sections ( atient 282832) were, after bloc 'i; ing with normal goat serum, incubated with the
One or several mechanism(s) could potentially account for the augmented u-PAR levels observed for invasive colon cancer. These include: 1) gene amplification, 2) a more stable mRNA or 3) transcriptional activation of the u-PAR gene.
The u-PAR gene is not amplified in invasive cancer To determine if invasive cultured colon cancer could be
characterized by an increased copy number of the u-PAR
gene, control (placental) DNA and DNA extracted from the highly (HCT 116, RKO) and the poorly (CBS, GEO, FET) invasive colon cancer cell lines were digested with Nco I and transferred to a nylon filter following resolution of the re- stricted fragments in an agarose gel. The restriction profile generated with this enzyme was identical for D N A from the invasive-competent and the invasive-deficient cell lines and normal (placental) source (Fig. 6). More importantly, the
6.54 WANG ETAL.
FIGURE 5 - Steady-state levels of u-PAR mRNA in cultured colon cancer. RNA from the indicated cell lines was extracted and, after electrophoresis, transferred to a nylon filter. The filter was probed with a cDNA corresponding to the receptor mRNA or GAPDH. Stringency washes were performed at 0.5 x SSC at 65°C. The filter was exposed to x-ray film (a) and subsequently scanned On a Betascope ‘03 for radioactivity (b)’ FIGURE 3 - Cross-linking of radioactive ATF to the cell surface
of colonic cells. Near confluent cells were acid pretreated and incubated at 4°C for 3 hr with 2.0 nM radioactive ATF, alone (- , or together with a 50-fold excess of unlabeled like-competitor (+{ After the cells were washed with PBS, radioactive ATF was cross-linked to the cell surface with DSS. The cells were lysed with a solution containing 0.5% CHAPS and the extract denatured in the presence of 2.3% SDS and 5% P-mercaptoethanol. Samples (25 bg protein) of the cell extracts, or radioactive ATF alone, were electrophoresed in a 12.5% SDS-PAGE gel. The gel was exposed to x-ray film. The experiment was repeated twice.
FIGURE 4 - Detection of u-PAR in invasive colon cancer cells by Western blotting. Equal amounts of detergent-extracted protein from the indicated cell lines were electrophoresed in a 12.5% gel and transferred to nitrocellulose filter. The filter was blocked and incubated in the presence of a polyclonal antibody to the u-PAR protein and subsequently with radioactive Protein A. The filter was washed extensively and exposed to x-ray film (a). The filter was stained with amido black to confirm equal loading of the lanes (b). Molecular weight markers are to the right.
intensity of the bands for the receptor-dense HCT 116 and RKO cell lines was indistinguishable from that of their receptor-deficient counterparts (CBS, GEO, FET) and normal (placental) DNA. Thus, there is no evidence for gene amplifi- cation in colon cancer cell lines endowed with a large ( 2 100,000/cell) number of urokinase receptors.
mRNA stability in the receptor-rich and receptor-deficient colon cancer cells
Alternatively, a lower rate of u-PAR mRNA turnover in the invasive cell types could lead to the accumulation of transcript
FIGURE 6 -The u-PAR gene is not amplified in invasive colon cancer. DNA (20 pg) was digested with 7.5 of Nco 1 at 37°C overnight. The digested DNA was electrophoresed in a 1% agarose gel and transferred to Nytran modified nylon. The filter was probed with a radioactive u-PAR cDNA and subsequently washed at 65°C in 0.5 x SSC/O.7.5% SDS. The blot was exposed to x-ray film (a). Equal loading of DNA was checked by photograph- ing the agarose gel after electrophoresis of the fragmented DNA (bl .
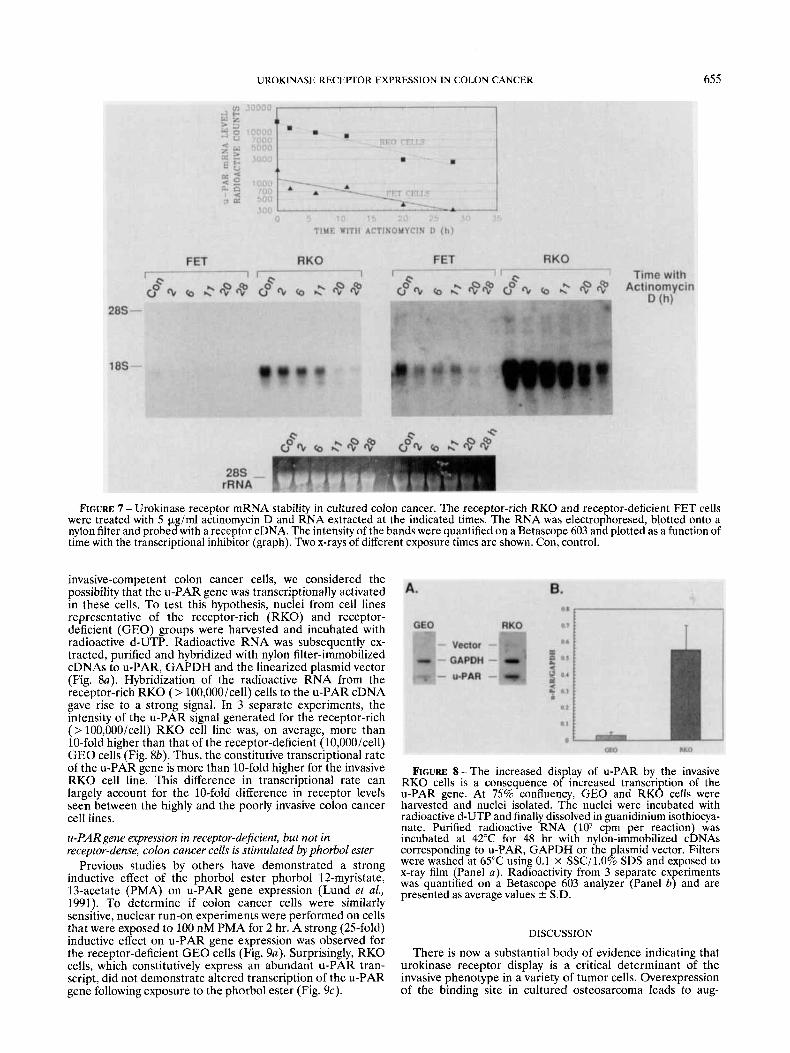
in these cells and an elevated level of binding sites. To test this hypothesis, cell lines from the receptor-rich (RKO, HCT 116) and receptor-deficient (GEO, FET) groups were treated with the transcriptional inhibitor actinomycin-D and R N A ex- tracted after varying periods of time. The R N A was resolved in a formaldehyde-agarose gel and transferred to a nylon filter and the blot was probed for u-PAR mRNA (Fig. 7). The turnover rate ( t lp) for u-PAR mRNA in FET and in GEO cells, which display only 10,000 receptors/cell, was calculated to be 10 hr (Fig. 7, inset). Similarly, the half-life of u-PAR mRNA for RKO and HCT 116 cells, which possess > 100,000 sites/cell, was estimated to be between 9 and 11 hr. Thus, the elevated number of binding sites displayed by the invasive cell lines is unlikely to reflect increased stability of the u-PAR transcript.
The u-PAR gene is transcriptionally activated in the invasive colon cancer cell line RKO
Since neither gene amplification nor mRNA stability could explain the augmented display of urokinase receptor by the
UROKINASE RECEPTOR EXPRESSION IN COLON CANCER 655
FIGURE 7 - Urokinase receptor mRNA stability in cultured colon cancer. The receptor-rich RKO and receptor-deficient FET cells were treated with 5 pg/ml actinomycin D and RNA extracted at the indicated times. The RNA was electrophoresed, blotted onto a nylon filter and probed with a receptor cDNA. The intensity of the bands were quantified on a Betascope 603 and plotted as a function of time with the transcriptional inhibitor (graph). Two x-rays of different exposure times are shown. Con, control.
invasive-competent colon cancer cells, we considered the possibility that the u-PAR gene was transcriptionally activated in these cells. To test this hypothesis, nuclei from cell lines representative of the receptor-rich (RKO) and receptor- deficient (GEO) groups were harvested and incubated with radioactive d-UTP. Radioactive RNA was subsequently ex- tracted, purified and hybridized with nylon filter-immobilized cDNAs to u-PAR, GAPDH and the linearized plasmid vector (Fig. 8a). Hybridization of the radioactive RNA from the receptor-rich RKO (> 100,00O/cell) cells to the u-PAR cDNA gave rise to a strong signal. In 3 separate experiments, the intensity of the u-PAR signal generated for the receptor-rich (> 100,00O/cell) RKO cell line was, on average, more than 10-fold higher than that of the receptor-deficient (10,00O/cell) GEO cells (Fig. 8b). Thus, the constitutive transcriptional rate of the u-PAR gene is more than 10-fold higher for the invasive RKO cell line. This difference in transcriptional rate can largely account for the 10-fold difference in receptor levels seen between the highly and the poorly invasive colon cancer cell lines. u-PAR gene expression in receptor-deficient, but not in receptor-dense, colon cancer cells is stimulated by phorbol ester
Previous studies by others have demonstrated a strong inductive effect of the phorbol ester phorbol 12-myristate, 13-acetate (PMA) on u-PAR gene expression (Lund et al., 1991). To determine if colon cancer cells were similarly sensitive, nuclear run-on experiments were performed on cells that were exposed to 100 nM PMA for 2 hr. A strong (25-fold) inductive effect on u-PAR gene expression was observed for the receptor-deficient GEO cells (Fig. 9a). Surprisingly, RKO cells, which constitutively express an abundant u-PAR tran- script, did not demonstrate altered transcription of the u-PAR gene following exposure to the phorbol ester (Fig. 9c).
FIGURE 8-The increased display of u-PAR by the invasive RKO cells is a consequence of increased transcri tion of the u-PAR gene. At 75% confluency, GEO and R K 8 cells were harvested and nuclei isolated. The nuclei were incubated with radioactive d-UTP and finally dissolved in guanidinium isothiocya- nate. Purified radioactive RNA (lo7 cpm per reaction) was incubated at 42°C for 48 hr with nylon-immobilized cDNAs corresponding to u-PAR, GAPDH or the plasmid vector. Filters were washed at 65°C using 0.1 X SSC/l.O% SDS and exposed to x-ray film (Panel a). Radioactivity from 3 separate ex eriments was quantified on a Betascope 603 analyzer (Panel by and are presented as average values k S.D.
DISCUSSION
There is now a substantial body of evidence indicating that urokinase receptor display is a critical determinant of the invasive phenotype in a variety of tumor cells. Overexpression of the binding site in cultured osteosarcoma leads to aug-

WANG ETAL. 656
FIGURE 9 - Treatment of GEO cells, but not RKO cells, with PMA stimulates transcription of the u-PAR gene. GEO (Panels a and b ) and RKO (Panel c) cells were treated, at 75% confluency, with 100 nM PMA for 2 hr. The cells were harvested and nuclei were collected and run on experiments conducted as described in the legend to Figure 8. Radioactivity for the GEO blot was quantified with a Betascope 603 analyzer and the data represented as a bar graph (Panel b).
mented laminin degradation as well as an increased penetra- tion of an extracellular matrix barrier (Kariko et al., 1993). Conversely, interfering with the binding of u-PA with its receptor has been shown to reduce extracellular matrix turn- over and in vitro invasion of cultured colon cancer and ovarian cancer (Schlechte et al., 1989; Hollas et al., 1991; Kobayashi et al., 1993). The contribution of the urokinase receptor to the invasive phenotype probably reflects the accelerated produc- tion of plasmin by receptor-bound u-PA, as reported by Ellis et al. (1991).
Although both invasive colon and breast cancers have been characterized by their increased u-PAR display, little is cur- rently known regarding the molecular mechanism(s) respon- sible for the elevated levels of u-PAR seen in these infiltrative cancers. This is a crucial issue to address from the perspective of developing novel anti-invasive and anti-metastatic strategies that exert their therapeutic effect via the reduced production of the binding site. To determine the cause of the elevated level of u-PAR in invasive colon cancer in vivo, 5 colon cancer cell lines, which exhibit either a high or low capacity to invade an extracellular matrix-coated porous filter and which can be readily distinguished by their different receptor number, were characterized for u-PAR receptor expression at the transcrip- tional and post-transcriptional level.
A good correlation existed between the amount of steady- state mRNA levels encoding the receptor and the number of surface binding sites. These data suggest that the disparity in receptor number observed between the 2 groups of colon cancer cell lines is a consequence of different amounts of u-PAR mRNA. Conversely, the data argue against the possibil- ity that the receptor disparity seen between the highly and the
poorly invasive groups is attributed to post-translational events, as has been reported for u-PAR in ovarian cancer (Pedersen et al., 1993) and in paroxysmal nocturnal hemoglobinuria (Ploug et al., 1992). In both instances, the receptor is secreted by virtue of its inability to anchor to the cell surface, possibly as a consequence of an absent glycolipid chain required for mem- brane anchoring. In the case of paroxysmal nocturnal hemoglo- binuria, leukocytes are positive for u-PAR mRNA while lacking cell surface receptors. This situation was not evident with the cultured colon cancer cell lines examined insofar as the receptor-deficient colon cancer cell lines, including FET, GEO and CBS, had little mRNA encoding the binding site. Thus, it is unlikely that u-PAR secretion is the underlying cause of receptor insufficiency in the poorly invasive cell lines. We also considered the possibility that the decrease in u-PAR display observed for GEO and CBS cells might be a conse- quence of an increased internalization of a ternary complex of u-PA-plasminogen activator inhibitor (PA1)-1 and the binding site. However, this possibility appears remote, since we have previously shown by reverse zymography and by Western blotting that GEO and CBS cells secrete substantially lower levels of PAI-1 than RKO and HCT 116 cells (Schlechte and Boyd, 1990).
The possibility that multiple copies of the u-PAR gene were contributing to an increased pool of receptor mRNA/protein, as has been reported for the HER2/neu receptor oncoprotein in breast cancer (Slamon et al., 1987), was considered. How- ever, Southern blotting of DNA derived from the invasive- competent and invasive-deficient cell lines revealed no evi- dence of gene amplification in any of the cell lines examined when compared with DNA from a normal (placental) source. Although, to our knowledge, there have been no published studies on u-PAR gene amplification in any cancer, a previous study has shown that the gene for the protease (u-PA) itself may be a target for DNA amplification. Thus, the PC-3 prostate cancer cell line, which secretes approximately 1 pg/ml of the plasminogen activator, was found to have 3 copies of the u-PA gene (Hollas et al., 1992).
Alternatively, we hypothesized that a more stable transcript could lead to the accumulation of mRNA/protein in invasive colon cancer. Indeed, both the constitutive and the inducible expression of several proteases and their inhibitors have been ascribed to message stabilization. For example, the increased level of u-PA observed in metastatic mammary adenocarci- noma was found to be a consequence of a more stable mRNA encoding the plasminogen activator (Henderson et al., 1992). Likewise, the induction of tissue inhibitor of metalloprotein- ases (TIMP) by phorbol ester could be partly attributed to a more stable transcript (Shapiro et al., 1993). In contrast, our mRNA stability studies revealed no difference in the half-life of the u-PAR message between the receptor-rich (RKO, HCT 116) and the receptor-deficient (GEO, CBS, FET) colon cancer cells. Thus, it is unlikely that the large number of u-PAR receptors, which are characteristic of the invasive colon cancer cells, reflects a decreased turnover of the mRNA.
Since neither gene amplification nor mRNA stability could explain the 10-fold disparity in receptor number seen between the invasive-competent and invasive-deficient cell types, we hypothesized that transcriptional rates were elevated in the former cells. Indeed, nuclear run-on experiments with repre- sentative cell lines indicated that the difference in the amount of steady-state u-PAR mRNA and protein could be largely explained by the different transcription rates of the u-PAR gene. While these studies suggest that transcriptional mecha- nisms are the main cause of increased u-PAR expression in invasive colon cancer, they provide little insight into the mechanisms by which u-PAR expression is regulated in infiltra- tive cancers. Three possibilities exist. First, transcription factors selectively produced by the invasive cells may activate the u-PAR promoter in trans. Second, the promoter may be activated as a consequence of a mutation(s) in the 5’ flanking

UROKINASE RECEPTOR EXPRESSION IN COLON CANCER 657
sequence. A third possibility, which has been documented for the human epidermal growth factor (EGF) receptor, is that a pause in transcription may occur leading to premature termina- tion of the mRNA in the receptor-deficient colon cancer cells. In the case of the EGF receptor, this transcriptional block is relieved by phorbol ester, culminating in a 5-fold induction in EGF receptor transcription.
Clearly, discrimination between the first 2 possibilities will have to await the cloning and sequencing of the regulatory sequences of the u-PAR gene. Several agents of diverse structure including TGF-p, the phosphatase inhibitor okadaic acid and protein kinase C activators have been reported to be strong inducers of u-PAR gene transcription (Lund et al., 1991, 1992). Perhaps the best documented, to date, is the inductive effect of PMA on u-PAR gene transcription. A SO-fold increase in the steady-state amount of u-PAR mRNA, which was paralleled by an increase in u-PAR gene transcription as determined by nuclear run-on experiments, was observed for PMA-stimulated U937 cells (Lund et al., 1991). This induction of u-PAR by PMA might, in fact, suggest that the expression of the u-PAR gene is regulated by protein kinase C pathways. Interestingly, while we were able to stimulate u-PAR gene transcription in the receptor-deficient GEO cells, we were unable to reproduce this finding for the invasive RKO cell line, which already produces a large number of u-PA binding sites. It i s tempting to speculate that the insensitivity of u-PAR gene
transcription to PMA in R K O cells reflects the maximal activation of protein kinase C pathways in these receptor-rich invasive cell lines. However, an alternative possibility is that the expression of the u-PAR gene in RKO cells is already maximally activated and thus incapable of being further augmented by phorbol ester.
In conclusion, we have shown that the elevated number of urokinase receptors displayed by invasive colon cancer is a consequence, at least in part, of transcriptional activation of the gene. In contrast, we found no evidence that gene amplification or increased mRNA stability were contributing factors to the high level of u-PAR mRNA/protein observed for invasive colon cancer cell lines. Our findings provide a clear rationale for further studies to determine the role of cis- and trans-activating mechanisms in u-PAR gene activation in infiltrating colon cancer.
ACKNOWLEDGEMENTS
We express our gratitude t o Dr. R. Hart at American Diagnostica for the generous support of this project. We also thank Dr. G. Nicolson for providing assistance with the immunohistochemical staining for u-PAR. This work was supported by grants to D.B. from the NIH (R29CA51539 and ROlDE1084.5) and from the Council for Tobacco Research (3438).
REFERENCES
APPELLA, E., ROBINSON, E., ULLRICH, S., STOPPELLI, M., CORTI, A., CASSANI, G. and BLASI, E., The receptor-binding sequence of uroki- nase. J. biol. Chem., 262,4437-4440 (1987). BEHRENDT, N., PLOUG, M., PATTHY, L., HOUEN, G., BLASI, F. and DAN0, K., The ligand-binding domain of the cell surface receptor for urokinase-type plasminogen activator. J. biol. Chem., 266, 7842-7847 (1991). DEL VECCHIO, S., STOPPELLI, M.P., CARRIERO, M.V., FONTI, R., MASSA, O., LI, P., BOTTI, G., CERRA, M., D’AIUTO, G., ESPOSITO, G. and SALVATORE, M., Human urokinase receptor concentration in malignant and benign breast tumors by in vitro quantitative autoradiog- raphy: Comparison with urokinase levels. Cancer Res., 53,3198-3206 (1993). ELLIS, V., BEHRENDT, N. and DAN@, K., Plasminogen activation by receptor-bound urokinase. J. biol. Chem., 266,12,752-12,758 (1991). HENDERSON, B.R., TANSEY, W.P., PHILLIPS, S.M., RAMSHAW, I.A. and KEFFORD, R.E., Transcriptional and posttranscriptional activation of urokinase plasminogen activator gene expression in metastatic tumor cells. CancerRes., 52,2489-2496 (1992). HOLLAS, W., BLASI, F. and BOYD, D., Role of the urokinase receptor in facilitating extracellular matrix invasion by cultured colon cancer. Cancer Res., 51,3690-3695 (1991). HOLLAS, W., HOOSEIN, N., CHUNG, L.W.K., MAZAR, A., HENKIN, J., KARIKO, K., BARNATHAN, E. and BOYD, D., Expression of urokinase and its receptor in invasive and non-invasive prostate cancer cell lines. Thrombosis Haemostasis, 68,662-666 (1992). KAKIKO, K., Kuo, A., BOYD, D., OKADA, S., CINES, D. and BAR- NATHAN, E., Overexpression of urokinase receptor increases matrix invasion without altering cell migration in a human osteosarcoma cell line. Cancer Res., 53,3109-3117 (1993). KORAYASHI, H., OHI, H., SHINOHARA, H., FUJII, T., SCHMITT, M., GOIIETZKI, L., CHUCHOLOWSKI, N., JANICKE, F. and GRAEFF, H., Saturation of tumour cell surface receptors for urokinase-type plas- minogen activator by amino-terminal fragment and subsequent effect of on reconstituted basement membrane invasion. Brif. J. Cancer, 67,
LUND, L., RONNE, E., ROLDAN, A., BEHRENDT, N., ROMER, J., BLASI, F. and DAN@, K., Urokinase receptor mRNA level and gene transcrip- tion are strongly and rapidly increased by phorbol myristate acetate in human monocyte-like U937 cells. J. biol. Chem., 266, 5177-5181 (1991). LUND, L.R., ROMER, J., ELLIS, V., BLASI, F. and DAN^,, K., Urokinase- receptor biosynthesis, mRNA levels and gene transcription are in- creased by transforming growth factor-pl in human A549 lung carcinoma cells. EMBO J., 10,3399-3407 (1992).
537--544 (1993).
NIELSEN, L., KELLERMAN, G.M., BEHRENDT, N., PICONE, R., DAN^, K. and BLASI, F., A 55,000-60,000 Mr receptor protein for urokinase-type plasminogen activator. J. biol. Chem., 263,2358-2363 (1988). OSSOWSKI, L. and REICH, E., Antibodies to plasminogen activator inhibit human tumor metastasis. Cell, 35,611-619 (1983). PEDERSEN, N., SCHMIIT, M., RONNE, E., INES NICOLETTI, M., HOYER- HANSEN, G., CONESE, M., GIAVAZZI, R., DAN^, K., KUHN, W., JANICKE, F. and BLASI, F., A ligand-free, soluble urokinase receptor is present in the ascitic fluid from patients with ovarian cancer. J. clin. Invest., 92,2160-2167 (1993). PLOUG, M., ERIKSEN, J., PLESNER, T., HANSEN, N.E. and DAN@, K., A soluble form of the glycolipid-anchored receptor for urokinase-type plasminogen activator is secreted from peripheral blood leukocytes from patients with paroxysmal nocturnal hemoglobinuria. Europ. J. Biochem., 208,397-404 (1992). PYKE, C., KRISTENSEN, P., RALFKIAER, E., GRONDAHL-HANSEN, J., EIRKSEN, J., BLASI, F. and DAN0, K., Urokinase-type plasminogen activator is expressed in stromal cells and its receptor in cancer cells at invasive foci in human colon adenocarcinomas. Amer. J. Pathol., 138, 1059-1067 (1991). ROLDAN, A,, CUBELLIS, M., MASUCCI, M., BEHRENDT, N., LUND, L., DAN0, K., APPELLA, E. and BLASI, F., Cloning and expression of the receptor for urokinase plasminogen activator, a central molecule in cell surface, plasmin dependent proteolysis. EMBO J., 9, 467-474 (1990). SCHLECHTE, W. and BOYD, D., Insensitivity of laminin degradation by receptor-bound urokinase to PAL1 in cultured colon cancer. Cancer Comm., 2,261-270 (1990). SCHLECHTE, W., MURANO, G. and BOYD, D.D., Examination of the role of the urokinase receptor in human colon cancer mediated laminin degradation. Cancer Res., 49,6064-6069 (1989). SHAPIRO, S.D., DOYLE, G.A., LEY, T.J., PARKS, W.C. and WELGUS, H.G., Molecular mechanisms regulating the production of collagenase and TIMP in U937 cells: Evidence for involvement of delayed transcriptional activation and enhanced mRNA stability. Biochemisty, 32,42864292 (1993). SLAMON, D.J., CLARK, G.M., WONG, S.G., LEVIN, W.J., ULLRICH, A. and MCGUIRE, W.L., Human breast cancer: Correlation of relapse and survival with amplification of the HER2/neu oncogene. Science, 235,
TRYGGVASON, K., HOYHTYA, M. and SALO, T., Proteolytic degradation of extracellular matrix in tumor invasion. Biochim. biophys. Acta, 907,
177-182 (1987).
191-217 (1987).
Copyright © 2022 FDOKUMEN




















