
Examination of the Role of the Urokinase Receptor in Human Colon Cancer Mediated Laminin...
7
1989;49:6064-6069. Cancer Res Warren Schlechte, Genesio Murano and Douglas Boyd Colon Cancer Mediated Laminin Degradation Examination of the Role of the Urokinase Receptor in Human Updated version http://cancerres.aacrjournals.org/content/49/21/6064 Access the most recent version of this article at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications Research. on August 13, 2014. © 1989 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on August 13, 2014. © 1989 American Association for Cancer cancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Examination of the Role of the Urokinase Receptor in Human Colon Cancer Mediated Laminin...

1989;49:6064-6069. Cancer Res Warren Schlechte, Genesio Murano and Douglas Boyd Colon Cancer Mediated Laminin DegradationExamination of the Role of the Urokinase Receptor in Human
Updated version
http://cancerres.aacrjournals.org/content/49/21/6064
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on August 13, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on August 13, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

(CANCER RESEARCH 49, 6064-6069, November I, 1989]
Examination of the Role of the Urokinase Receptor in Human Colon CancerMediated Laminin Degradation1
Warren Schlechte, Genesio Murano2, and Douglas Boyd3
Department of Pharmacology, Bristol-Baylor Laboratory, Baylor College of Medicine, Houston, Texas 77030
ABSTRACT
The relevance of urokinase receptors to urokinase-mediated laminindegradation was investigated in cultured colon cancer. Six colon cancercell lines degraded laminin in a plasminogen-dependent manner. Theability of the individual cell lines to cleave the glycoprotein correlatedwell (/•= 0.9242) with the amount of urokinase recovered from the cell
surface by a mild acid treatment. A radioreceptor assay indicated thatcolon cancer cells most active in degrading the laminin, possessed thelargest number of urokinase receptors. Moreover, acid treatment whichdepletes the receptors of endogenous plasminogen activator augmentedthe specific binding of radioactive urokinase to the colon cancer cells by12-200%. A cell line (HCT 116) which displayed 1.1 x 10s receptors/
cell the majority of which were occupied with endogenous urokinase wasselected and the effects of a urokinase receptor antagonist on laminindegradation determined. The peptide antagonist reduced laminin turnoverby 60-80%. Morphological observations were consistent with thesefindings. Plasminogen-treated HCT 116 cells retracted from the culturedish and many cells were observed in the culture medium. This effectcould be largely reversed by simultaneous treatment with the peptideantagonist. A poor correlation was found between laminin degradationand soluble urokinase (r = 0.1074). These data strongly argue for a roleof the urokinase receptor in facilitating the action of the plasminogenactivator in colon cancer.
INTRODUCTION
The plasminogen activator, urokinase, is overexpressed inmany malignancies including those of the colon, prostate,breast, and lung (1, 2). The function of urokinase in tumorbiology is not entirely clear at the present time. However, it iswell documented (3, 4) that the production of the widely actingserine protease, plasmin, from the inert zymogen, plasminogenis mediated by urokinase. Plasmin has been reported to degradethe major basement membrane constituent, laminin (5, 6).Indeed, colonie tumors are characterized as having high levelsof urokinase and this is associated with reduced amounts oflaminin within a basement membrane structure (7-9). Depletion of laminin from a basement membrane could be one ofmany factors which allows for the expression of an invasivephenotype. In line with this, antiurokinase antibodies have beenreported to reduce tumor dissemination in several systems (10,11).
Recently, a binding site for urokinase has been identified onthe surface of a number of diverse tumor cells including thoseof the colon (12-14). It has been speculated that these receptorscould facilitate the actions of the plasminogen activator (13,15, 16). However, direct evidence in support of this premise isscant. Recently, in an elegant series of experiments, Ossowski(17) showed that tumor cells capable of binding urokinasedemonstrated a metastatic phenotype while those which se-
Received 3/21/89; revised 6/9/89; accepted 7/28/89.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by Grants CA45967 and CA34432.2 Present address: Center for Biologies Evaluation and Research. 8800 Rock-
ville Pike, Bethesda. MD 20892.3To whom requests for reprints should be addressed at Baylor College of
Medicine, Bristol-Baylor Laboratories, Texas Medical Center. Department ofPharmacology, Houston. T\ 77030.
creted urokinase in the absence of cell surface binding werenonmetastatic. Since the tumor cells were positive for urokinasereceptors, it was concluded that the interaction of urokinasewith its receptor was prerequisite for the actions of the plasminogen activator. The evidence, although highly suggestive ofsuch an interplay, cannot be regarded as irrefutable. In an earlierstudy, Quigley et al. (18) had demonstrated membrane boundplasminogen activator to be responsible for the morphologicaleffects of phorbol ester on transformed chick embryo fibro-blasts. Although the presence of urokinase receptors on thesecells was not examined, the membrane localization of theplasminogen activator is in line with such a possibility.
The present study was undertaken to determine the importance of the urokinase binding site for plasminogen-dependentlaminin degradation mediated by colon cancer in vitro. Theresults herein suggest an important role for the urokinasebinding site in promoting the actions of this plasminogenactivator in cultured colon cancer. In contrast, the contributionof soluble urokinase to laminin degradation appears to be onlyminor.
MATERIALS AND METHODS
Materials. High molecular weight (55,000) urokinase in the form of"Winkinase" was purified from a commercial preparation (14). A
peptide (UK 12-32) corresponding to amino acids 12-32 of high molecular weight urokinase was synthesized by Multi Peptide Systems, SanDiego, CA. ELISA4 antibodies were kindly provided by Drs. Collen
and DeClerk, Leuven, Belgium. lodogen was purchased from PierceChemicals, Rockford, IL. Collaborative Research (Bedford, MA) provided laminin. Plasminogen (<0.01 % plasmin present) was supplied byBoehringer Mannheim, Indianapolis, IN. Luteinizing hormone-releasing hormone was a generous gift from Dr. Hoosein (University ofTexas. Houston, TX).
Cell Culture. The well (CBS, GEO, FET) and the poorly differentiated (HCT 116, RKO, MOSER) colon cancer cell lines have beenextensively characterized (19, 20). In monolayer, the well-differentiatedcell lines are polarized and form numerous tight junctions. A numberof these cell lines form secretory domes at confluency. In vivo, theyform glandular type tumors at low rates of incidence. In contrast, thepoorly differentiated colon cancer cell lines lack polarity in vitro, anddo not manifest tight junctions nor secretory domes. In vivo, they formanaplastic tumors at a high rate of incidence.
Both well and poorly differentiated colon cancer cell lines have beenadapted to grow continuously in McCoy's medium 5A supplemented
with 4 Mg/ml transferrin, 5 Mg/ml insulin, and 10 ng/ml EGF (serumfree medium) (21, 22). At approximately 80% confluency, the cellswere subcultured using 0.125% and 0.025% trypsin for the well andpoorly differentiated cell types, respectively.
Collection of conditioned medium was as follows. Cells were subcultured in 35-mm dishes (Falcon) and at 90% confluency, replenishedwith fresh serum free medium. The cultures were maintained for anadditional 72 h, the spent medium harvested and cells enumerated.
Colon cancer cell surface extracts were obtained by treating confluentcultures with an acid buffer (50 mm glycine, 0.1 M NaCl, pH 3.0) for 3min at room temperature (13, 14). The extract was neutralized with asolution containing 0.5 M HEPES, 0.1 M NaCl (pH 7.4) and dialyzed
4The abbreviations used are: ELISA, enzyme-linked immunosorbent assay:EGF. epidermal growth factor; HEPES. 4-(2-hydroxyethyl)-l-piperazineethane-sulfonic acid; HMW, high molecular weight; LH-RH. luteinizing hormone-releasing hormone; PBS, phosphate-buffered saline: BSA. bovine serum albumin.
6064
Research. on August 13, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

UROKINASE RECEPTOR FUNCTION IN COLON CANCER
against PBS. The dialyzed material was concentrated 10-fold by freeze
drying.Laminili Degradation Studies. Laminin (100 Mg)was iodinated by a
chloramine T method as described previously (23). Radioactive lamininwas separated from unincorporated I25I by extensive dialysis againstPBS. I25l-labeled laminin was filter sterilized and air dried on Falcon35-mm dishes. Tissue culture dishes were washed 3x with serum freemedium and stored at 4°Cuntil required. Routinely, each dish contained = 107 dpm of radioactive laminin (=2 ^g). Colon cancer cells
were trypsinized, washed 3x with serum free medium and dispensedinto radioactive laminin coated dishes. At 100% confluency (=30 hafter subculture), the cells were supplemented with 10 pg/m\ plasmi nogen (final concentration). Subsequently, aliquots of the culture medium were withdrawn every 3 h and radioactivity determined. In somecases, aliquots of the supernatant were denatured and analyzed on a7.5% sodium dodecyl sulfate-polyacrylamide gel electrophoresis gelunder reducing conditions (24). In experiments in which the effect ofUK12-32 was under study, the peptide was added both 12 h prior to,and at the time of plasminogen addition. Experiments were terminatedwhen 20% of the radioactive substrate had been exhausted.
Determination of Urokinase Levels in Cell Surface Acid Extracts andin the Conditioned Medium. An ELISA was used to assess the amountof this plasminogen activator in the spent medium or in the cell surfaceextracts. The method has been reported elsewhere (14, 25). The assaydetects both the single and the two chain forms of high molecularweight urokinase but not the M, 33,000 fragment. The detection limitis arbitrarily set at 20 pg and the assay is linear up to 600 pg of antigen.
Urokinase Receptor Assays and Competition Studies. Affinity purifiedHMW-urokinase (14) was iodinated, as described previously (14) using"iodogen" as the oxidant. The radioactive protein was desalted on a
Pharmacia PD10 column equilibrated with 0.1% BSA in 30 mM phosphate buffer (pH 7.4).
The receptor assay has been detailed elsewhere (14). Briefly, confluent cultures were acid pretreated (glycine 50 mM, 0.1 M NaCl, pH3.0) for 3 min (unless stated otherwise) and the pH restored with asolution containing 0.5 M HEPES, 0. l M NaCl (pH 7.4). The cells wereincubated at 37°Cfor 30 min in binding buffer (20 mM HEPES, 0.1%BSA in McCoy's medium 5A, pH 7.4) in the presence of a single (2.0nM) or a range (0.25-12.5 nM) of '"I-labeled urokinase concentrations.Parallel cultures contained unlabeled HMW-urokinase in a 20-foldexcess to determine nonspecific ligand interactions. For competitionstudies, the cultures were incubated with 2.0 nM radioligand in theabsence or presence of increasing concentrations of the UK 12-32 pep-tide. After incubation at 37°C,cultures were washed 2x with PBS
containing 0.1% BSA and lysed with a 1% Triton XI00 solution.To obtain an estimate of the number of occupied receptors, cultures
were either acid pretreated (to remove endogenous urokinase) or washedwith PBS and analyzed for the specific binding of radioactive urokinase.The amount of radioligand bound, in each case, is a crude measure ofthe total number and the number of the vacant binding sites, respectively. The number of cryptic receptors was estimated by subtraction.
RESULTS
Neither the well nor the poorly differentiated colon cancercells solubilized radioactive laminin in the absence of plasminogen (Fig. 1). However, supplementation of the cultured cellswith the zymogen caused a marked solubilization of the radioactive basement membrane constituent (Fig. 1). In this respect,the poorly differentiated colon cancer cells were 2-32x morepotent than the well-differentiated cells. Analysis of the solubilized products in the culture medium (Fig. 2) revealed theabsence of the Mr 2 x IO5and the Mr 4 x IO5laminin subunits.
Smaller fragments, presumably reflecting proteolytic end products, were observed in the conditioned medium derived fromHCT 116 cells supplemented with plasminogen. Urokinase isthe predominant plasminogen activator elaborated by culturedcolon cancer cells since over 80% of plasminogen activationcan be blocked by antiurokinase antibody (23). While we cannotexclude the possibility that trace trypsin retained by the cells
a 1.6
1 I = 1-4
fi » 1J
11? 1.0•¿�DO»S « ° 0.8
HE0'6
f «f 0.4
ü 0.2
*/* ffi
WellDifferentiated
PoorlyDifferentiated
Fig. 1. Radioactive laminin solubilization by colon cancer cell lines. Cells wereplated onto dishes coated with =107 dpm radioactive laminin. At approximately
100% confluency, the experiment was initiated by the addition of 10 Mg/mlplasminogen. After 3 h of incubation, aliquots were withdrawn and counted forradioactivity. Spontaneous release of intact laminin did not exceed 2% of thetotal radioactivity. Control experiments indicated that plasminogen alone wasinsufficient to solubilize the radioactive substrate. The experiment was terminatedwhen 20% of the substrate had been exhausted. Data are presented as mean ±range values of two separate experiments. @, plasminogen supplemented cultures;D, plasminogen absent.
A ÃŽ*-*T
Z//Molecular
WeightStandards .
(DaltonsXKT)
200-
116-
97-
66-
42-
Fig. 2. Degradation of '"I-labeled laminin by HCT 116 cells—analysis bysodium dodecyl sulfate-polyacrylamide gel electrophoresis. HCT 116 cells weresubcultured on radioactive laminin-coated dishes and at =100% confluency,plasminogen added. Parallel dishes contained fresh serum free medium supplemented with plasminogen and 5 ng/ml authentic urokinase. After 3 h of incubation at 37'C, aliquots of the supernates were denatured in the presence of ß-mercaptoethanol. The samples (2-4 x IO4dpm) were electrophoresed on a 7.5%sodium-dodecyl sulfate-polyacrylamide electrophoresis gel. The gel was autora-diographed. Molecular weight markers are indicated.
after extensive washing contributes to laminin solubilization,these effects are probably only minor. In the absence of plasminogen (Fig. 1) laminin degradation was negligible. Further,a nonspecific interaction of trypsin with these cells is unlikelyto account for the differential abilities of the well and poorlydifferentiated cells to cleave the radioactive substrate in thepresence of plasminogen (Fig. 1).
Laminin degradation might reflect cell surface bound urokinase. To examine this possibility, the colon cancer cells were
6065
Research. on August 13, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
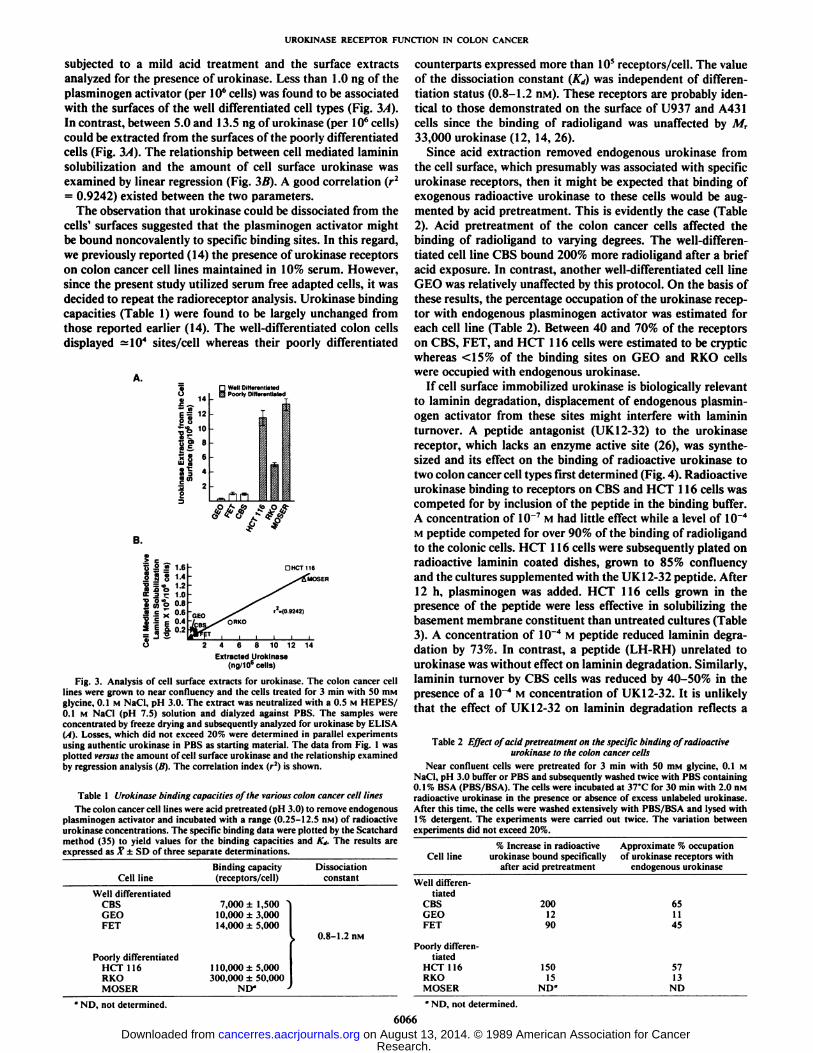
UROKINASE RECEPTOR FUNCTION IN COLON CANCER
subjected to a mild acid treatment and the surface extractsanalyzed for the presence of urokinase. Less than 1.0 ng of theplasminogen activator (per IO6cells) was found to be associated
with the surfaces of the well differentiated cell types (Fig. 3A).In contrast, between 5.0 and 13.5 ng of urokinase (per IO6cells)
could be extracted from the surfaces of the poorly differentiatedcells (Fig. 3/4). The relationship between cell mediated lamininsolubilization and the amount of cell surface urokinase wasexamined by linear regression (Fig. 3B). A good correlation (r2= 0.9242) existed between the two parameters.
The observation that urokinase could be dissociated from thecells' surfaces suggested that the plasminogen activator might
be bound noncovalently to specific binding sites. In this regard,we previously reported (14) the presence of urokinase receptorson colon cancer cell lines maintained in 10% serum. However,since the present study utilized serum free adapted cells, it wasdecided to repeat the radioreceptor analysis. Urokinase bindingcapacities (Table 1) were found to be largely unchanged fromthose reported earlier (14). The well-differentiated colon cellsdisplayed = 104 sites/cell whereas their poorly differentiated
well Differentiated
a 14
•¿�I «*•£10
118.158 6
:¿ii
[3 PoorlyDitf_ñnr«iiiated
B.
C .2 J5 1.6.i S g 1.4•¿�O= w 1 ?B J3 O ' ~
Å’ 2 !T 1.0
S 5 o 0.8« <= 3 0.6Sip 0.45 I I 0.2
u 2 4 6 8 10 12 14
Extracted Urokinase(ng;i06cells)
Fig. 3. Analysis of cell surface extracts for urokinase. The colon cancer celllines were grown to near confluency and the cells treated for 3 min with 50 HIMglycine. 0.1 M NaCI, pH 3.0. The extract was neutralized with a 0.5 M HEPES/0.1 M NaCI (pH 7.5) solution and dialyzed against PBS. The samples wereconcentrated by freeze drying and subsequently analyzed for urokinase by ELISA(A). Losses, which did not exceed 20% were determined in parallel experimentsusing authentic urokinase in PBS as starting material. The data from Fig. 1 wasplotted versus the amount of cell surface urokinase and the relationship examinedby regression analysis (B). The correlation index (r2) is shown.
Table 1 Urokinase binding capacities of the various colon cancer cell linesThe colon cancer cell lines were acid pretreated (pH 3.0) to remove endogenous
plasminogen activator and incubated with a range (0.25-12.5 nM) of radioactiveurokinase concentrations. The specific binding data were plotted by the Scatchardmethod (35) to yield values for the binding capacities and K* The results areexpressed as X ±SD of three separate determinations.
counterparts expressed more than IO5receptors/cell. The value
of the dissociation constant (Kd) was independent of differentiation status (0.8-1.2 n\i). These receptors are probably identical to those demonstrated on the surface of U937 and A431cells since the binding of radioligand was unaffected by M,33,000 urokinase (12, 14, 26).
Since acid extraction removed endogenous urokinase fromthe cell surface, which presumably was associated with specificurokinase receptors, then it might be expected that binding ofexogenous radioactive urokinase to these cells would be augmented by acid pretreatment. This is evidently the case (Table2). Acid pretreatment of the colon cancer cells affected thebinding of radioligand to varying degrees. The well-differentiated cell line CBS bound 200% more radioligand after a briefacid exposure. In contrast, another well-differentiated cell lineGEO was relatively unaffected by this protocol. On the basis ofthese results, the percentage occupation of the urokinase receptor with endogenous plasminogen activator was estimated foreach cell line (Table 2). Between 40 and 70% of the receptorson CBS, FET, and HCT 116 cells were estimated to be crypticwhereas <15% of the binding sites on GEO and RKO cellswere occupied with endogenous urokinase.
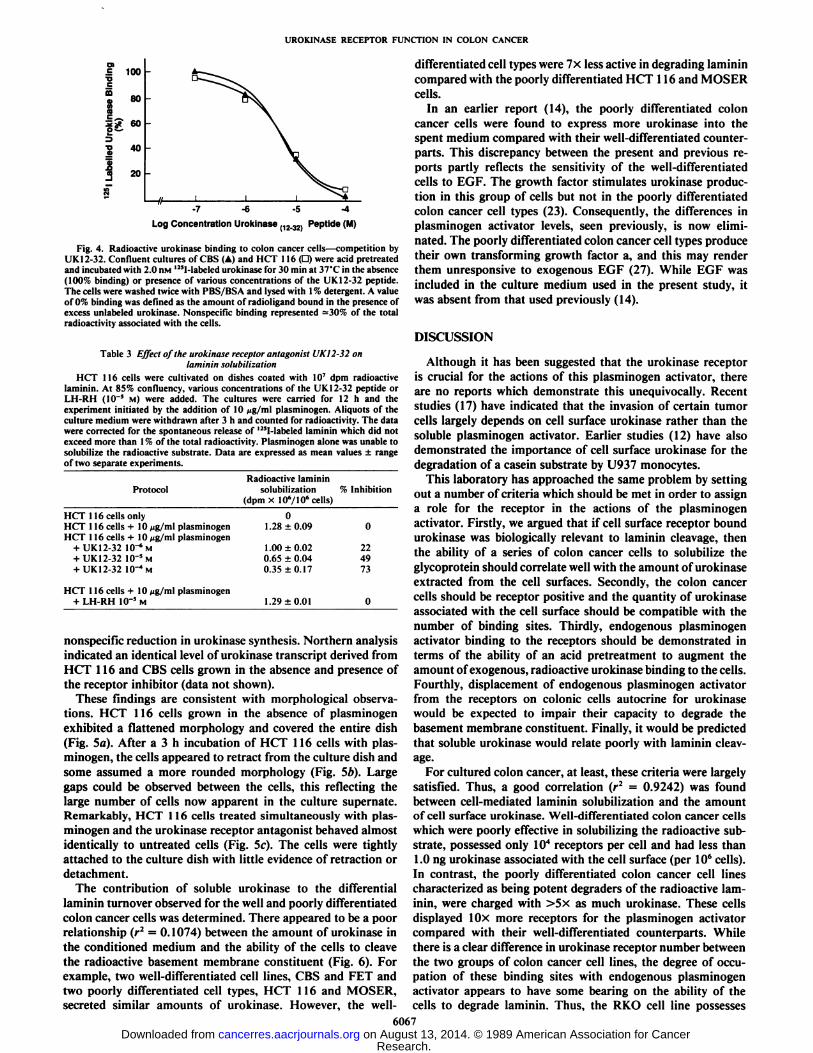
If cell surface immobilized urokinase is biologically relevantto laminin degradation, displacement of endogenous plasminogen activator from these sites might interfere with lamininturnover. A peptide antagonist (UK 12-32) to the urokinasereceptor, which lacks an enzyme active site (26), was synthesized and its effect on the binding of radioactive urokinase totwo colon cancer cell types first determined (Fig. 4). Radioactiveurokinase binding to receptors on CBS and HCT 116 cells wascompeted for by inclusion of the peptide in the binding buffer.A concentration of 10~7M had little effect while a level of 10~4
M peptide competed for over 90% of the binding of radioligandto the colonie cells. HCT 116 cells were subsequently plated onradioactive laminin coated dishes, grown to 85% confluencyand the cultures supplemented with the UK12-32 peptide. After12 h, plasminogen was added. HCT 116 cells grown in thepresence of the peptide were less effective in solubili/iny thebasement membrane constituent than untreated cultures (Table3). A concentration of 10~4 M peptide reduced laminin degra
dation by 73%. In contrast, a peptide (LH-RH) unrelated tourokinase was without effect on laminin degradation. Similarly,laminin turnover by CBS cells was reduced by 40-50% in thepresence of a 10~4 M concentration of UK 12-32. It is unlikely
that the effect of UK 12-32 on laminin degradation reflects a
Table 2 Effect of acid pretreatment on the specific binding of radioactiveurokinase to the colon cancer cells
Near confluent cells were pretreated for 3 min with 50 niM glycine. 0.1 MNaCI, pH 3.0 buffer or PBS and subsequently washed twice with PBS containing0.1% BSA (PBS/BSA). The cells were incubated at 37'C for 30 min with 2.0 nM
radioactive urokinase in the presence or absence of excess unlabeled urokinase.After this time, the cells were washed extensively with PBS/BSA and lysed with1% detergent. The experiments were carried out twice. The variation betweenexperiments did not exceed 20%.
Cell line
CelllineWell
differentiatedCBSGEOFETPoorly
differentiatedHCT116RKOMOSER"
ND, not determined.Binding
capacity Dissociation after acidpretreatment(receptors/cell)constant Wp||Him.rpn.tiated7.000
±1,500'1
0,000 ±3,00014.000 ±5.0001
10.000 ±5,000300.000±50.000CBSGEOFET0.8-1.
2 nMPoorlydifferen
tiatedHCT116RKONLV
J MOSER•
ND. notdetermined.60662001290ISO15ND"endogenous
urokinase65II455713ND
Research. on August 13, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
UROKINASE RECEPTOR FUNCTION IN COLON CANCER
O>.= 100TJ
80
40
20
-7 -6 -5 -4
Log Concentration Urokinase (12.32. Peptide (M)
Fig. 4. Radioactive urokinase binding to colon cancer cells—competition byUK 12-32. Confluent cultures of CBS (A) and HCT 116 (D) were acid pretreatedand incubated with 2.0 nw 12!I-labeled urokinase for 30 min at 37°Cin the absence(100% binding) or presence of various concentrations of the UK12-32 peptide.The cells were washed twice with PBS/BSA and lysed with 1% detergent. A valueof 0% binding was defined as the amount of radioligand bound in the presence ofexcess unlabeled urokinase. Nonspecific binding represented =30% of the totalradioactivity associated with the cells.
Table 3 Effect of the urokinase receptor antagonist UKI2-32 onlaminiti solubilization
HCT 116 cells were cultivated on dishes coated with IO7 dpm radioactivelaminin. At 85% confluency, various concentrations of the UK 12-32 peptide orLH-RH (10~5 M) were added. The cultures were carried for 12 h and the
experiment initiated by the addition of 10 Mg/ml plasminogen. Aliquots of theculture medium were withdrawn after 3 h and counted for radioactivity. The datawere corrected for the spontaneous release of 125l-labeled laminin which did not
exceed more than \% of the total radioactivity. Plasminogen alone was unable tosolubilize the radioactive substrate. Data are expressed as mean values ±rangeof two separate experiments.
Radioactive lamininProtocol solubilization % Inhibition
(dpm x 10'/106 cells)
HCT 116 cells only 0HCT 116 cells + 10 Mg/ml plasminogen 1.28 ±0.09 0HCT 116 cells -I-10 /jg/ml plasminogen
+ UK 12-32 10"6 M 1.00 ±0.02 22+ UK 12-32 10~5M 0.65 ±0.04 49+ UK12-32 lO-'M 0.35 ±0.17 73
HCT 116 cells + 10 /-u. ml plasminogen+ LH-RH IO"5 M 1.29 ±0.01 0
nonspecific reduction in urokinase synthesis. Northern analysisindicated an identical level of urokinase transcript derived fromHCT 116 and CBS cells grown in the absence and presence ofthe receptor inhibitor (data not shown).
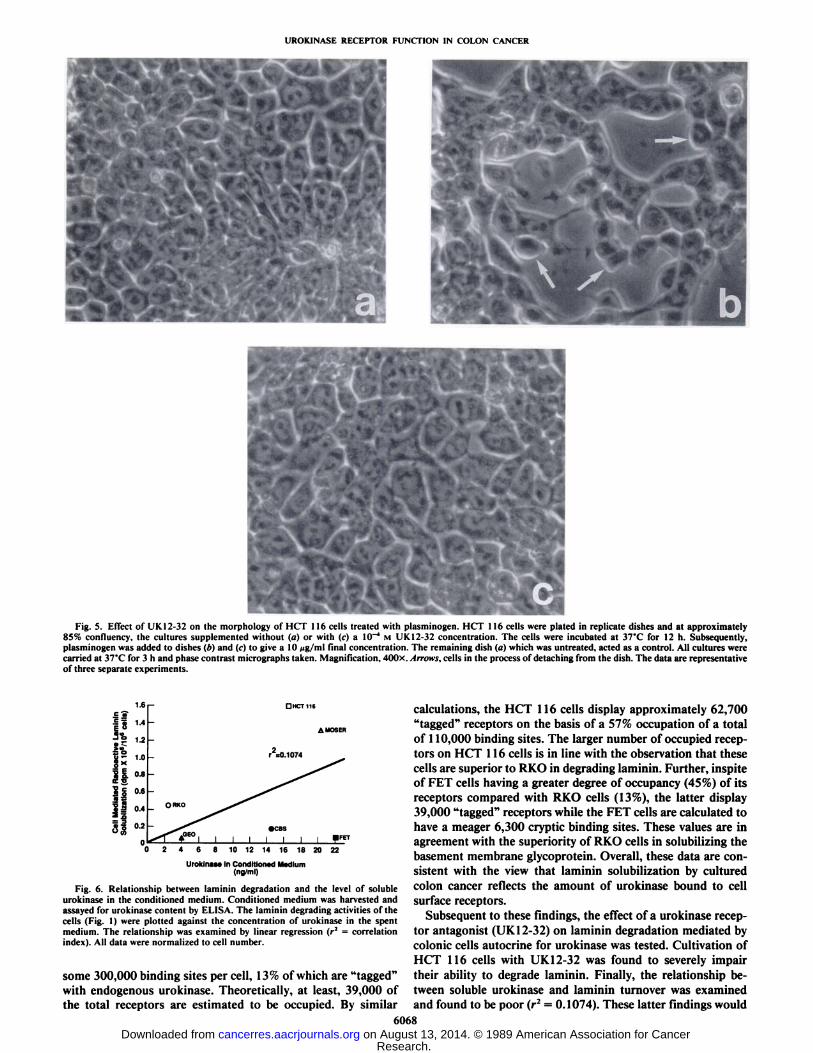
These findings are consistent with morphological observations. HCT 116 cells grown in the absence of plasminogenexhibited a flattened morphology and covered the entire dish(Fig. 5a). After a 3 h incubation of HCT 116 cells with plasminogen, the cells appeared to retract from the culture dish andsome assumed a more rounded morphology (Fig. 56). Largegaps could be observed between the cells, this reflecting thelarge number of cells now apparent in the culture supernate.Remarkably, HCT 116 cells treated simultaneously with plasminogen and the urokinase receptor antagonist behaved almostidentically to untreated cells (Fig. 5c). The cells were tightlyattached to the culture dish with little evidence of retraction ordetachment.
The contribution of soluble urokinase to the differentiallaminin turnover observed for the well and poorly differentiatedcolon cancer cells was determined. There appeared to be a poorrelationship (r2 = 0.1074) between the amount of urokinase in
the conditioned medium and the ability of the cells to cleavethe radioactive basement membrane constituent (Fig. 6). Forexample, two well-differentiated cell lines, CBS and FET andtwo poorly differentiated cell types, HCT 116 and MOSER,secreted similar amounts of urokinase. However, the well-
differentiated cell types were 7x less active in degrading laminincompared with the poorly differentiated HCT 116 and MOSERcells.
In an earlier report (14), the poorly differentiated coloncancer cells were found to express more urokinase into thespent medium compared with their well-differentiated counterparts. This discrepancy between the present and previous reports partly reflects the sensitivity of the well-differentiatedcells to EGF. The growth factor stimulates urokinase production in this group of cells but not in the poorly differentiatedcolon cancer cell types (23). Consequently, the differences inplasminogen activator levels, seen previously, is now eliminated. The poorly differentiated colon cancer cell types producetheir own transforming growth factor a, and this may renderthem unresponsive to exogenous EGF (27). While EGF wasincluded in the culture medium used in the present study, itwas absent from that used previously (14).
DISCUSSION
Although it has been suggested that the urokinase receptoris crucial for the actions of this plasminogen activator, thereare no reports which demonstrate this unequivocally. Recentstudies (17) have indicated that the invasion of certain tumorcells largely depends on cell surface urokinase rather than thesoluble plasminogen activator. Earlier studies (12) have alsodemonstrated the importance of cell surface urokinase for thedegradation of a casein substrate by U937 monocytes.
This laboratory has approached the same problem by settingout a number of criteria which should be met in order to assigna role for the receptor in the actions of the plasminogenactivator. Firstly, we argued that if cell surface receptor boundurokinase was biologically relevant to laminin cleavage, thenthe ability of a series of colon cancer cells to solubilize theglycoprotein should correlate well with the amount of urokinaseextracted from the cell surfaces. Secondly, the colon cancercells should be receptor positive and the quantity of urokinaseassociated with the cell surface should be compatible with thenumber of binding sites. Thirdly, endogenous plasminogenactivator binding to the receptors should be demonstrated interms of the ability of an acid pretreatment to augment theamount of exogenous, radioactive urokinase binding to the cells.Fourthly, displacement of endogenous plasminogen activatorfrom the receptors on colonie cells autocrine for urokinasewould be expected to impair their capacity to degrade thebasement membrane constituent. Finally, it would be predictedthat soluble urokinase would relate poorly with laminin cleavage.
For cultured colon cancer, at least, these criteria were largelysatisfied. Thus, a good correlation (r2 = 0.9242) was foundbetween cell-mediated laminin solubilization and the amountof cell surface urokinase. Well-differentiated colon cancer cellswhich were poorly effective in solubilizing the radioactive substrate, possessed only 104 receptors per cell and had less than1.0 ng urokinase associated with the cell surface (per IO6cells).
In contrast, the poorly differentiated colon cancer cell linescharacterized as being potent degraders of the radioactive laminin, were charged with >5x as much urokinase. These cellsdisplayed 10x more receptors for the plasminogen activatorcompared with their well-differentiated counterparts. Whilethere is a clear difference in urokinase receptor number betweenthe two groups of colon cancer cell lines, the degree of occupation of these binding sites with endogenous plasminogenactivator appears to have some bearing on the ability of thecells to degrade laminin. Thus, the RKO cell line possesses
6067
Research. on August 13, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
UROKINASE RECEPTOR FUNCTION IN COLON CANCER
Fig. 5. Effect of UK12-32 on the morphology of HCT 116 cells treated with plasminogen. HCT 116 cells were plated in replicate dishes and at approximately85% confluency, the cultures supplemented without (fl) or with (c) a 10~4 M UK 12-32 concentration. The cells were incubated at 37°Cfor 12 h. Subsequently,
plasminogen was added to dishes (A)and (c) to give a 10 Mg/ml limi concentration. The remaining dish (a) which was untreated, acted as a control. All cultures werecarried at 37°Cfor 3 h and phase contrast micrographs taken. Magnification, 400x. Arrows, cells in the process of detaching from the dish. The data are representative
of three separate experiments.
|i 0.4
0 2 4 6 8 10 12 14 16 18 20 22
Urokinase In Conditioned Medium(ng/ml)
Fig. 6. Relationship between laminin degradation and the level of solubleurokinase in the conditioned medium. Conditioned medium was harvested andassayed for urokinase content by ELISA. The laminin degrading activities of thecells (Fig. I ) were plotted against the concentration of urokinase in the spentmedium. The relationship was examined by linear regression (r2 = correlation
index). All data were normalized to cell number.
some 300,000 binding sites per cell, 13% of which are "tagged"
with endogenous urokinase. Theoretically, at least, 39,000 ofthe total receptors are estimated to be occupied. By similar
calculations, the HCT 116 cells display approximately 62,700"tagged" receptors on the basis of a 57% occupation of a total
of 110,000 binding sites. The larger number of occupied receptors on HCT 116 cells is in line with the observation that thesecells are superior to RKO in degrading laminin. Further, inspiteof FET cells having a greater degree of occupancy (45%) of itsreceptors compared with RKO cells (13%), the latter display39,000 "tagged" receptors while the FET cells are calculated to
have a meager 6,300 cryptic binding sites. These values are inagreement with the superiority of RKO cells in solubilizing thebasement membrane glycoprotein. Overall, these data are consistent with the view that laminin solubilization by culturedcolon cancer reflects the amount of urokinase bound to cellsurface receptors.
Subsequent to these findings, the effect of a urokinase receptor antagonist (UK 12-32) on laminin degradation mediated bycolonie cells autocrine for urokinase was tested. Cultivation ofHCT 116 cells with UK 12-32 was found to severely impair
their ability to degrade laminin. Finally, the relationship between soluble urokinase and laminin turnover was examinedand found to be poor (r2 = 0.1074). These latter findings would
6068
Research. on August 13, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

UROKINASE RECEPTOR FUNCTION IN COLON CANCER
suggest the relative unimportance of "free" urokinase in laminin
cleavage.Receptor bound urokinase may be superior to the soluble
form for one or more reasons. Immobilization of urokinase onthe cell surface could represent a concentrating mechanism,bringing it in close proximity with laminin located at thebasolateral surface. A variation of this proposal is that bothurokinase and plasminogen are juxtaposed on the tumor cellsurface, allowing for the generation of high levels of plasmin inthe immediate vicinity of the laminin. In line with this, receptorsfor plasminogen and urokinase have been identified on thesame cell types (28). An alternative hypothesis put forward byChapman et al. (29) is that soluble urokinase but not cell surfacebound plasminogen activator is vulnerable to serum or cell-derived inhibitors. Urokinase present in conditioned mediummay represent plasminogen activator which has escaped entrapment by surface receptors.
Laminin is a major constituent of basement membranes invivo (30), and its removal may be one of a series of eventsnecessary for the expression of an invasive phenotype. However,it has been well documented (31, 32) that other key extracellularmatrix constituents must be cleaved to permit tumor cell traverse of the basement membrane. To achieve this end, cancercells produce, in a coordinate fashion, an array of hydrolyticenzymes including collagenases and heparanases (32-34). Ourstudies have not addressed the expression of these enzymes incultured colon cancer. For this reason, caution should be exercised in drawing conclusions as to the in vivo behavior of coloniecancer cells based on the amount of urokinase immobilized tocell surface receptors. Notwithstanding these limitations, thedata herein provide strong support for the contention that theurokinase receptor facilitates the laminin degrading actions ofthis plasminogen activator in cultured colon cancer.
ACKNOWLEDGMENTS
We are indebted to Dr. D. Collen and Dr. P. DeClerk for the generousgifts of the ELISA antibodies. The authors also wish to thank Drs.Brattain and Yeoman for critical appraisal of the manuscript.
REFERENCES
1. Markus, G., Camiolo, S., Kohga. S., Madeja, J., and Mittleman, A. Plasmin-ogen activator secretion of human tumors in short-term organ culture including a comparison of primary and metastatic tumors. Cancer Res., 43: 5517-5525, 1983.
2. Sappine, A., Busso. N., Belin. D., and Vassalli, J. Increase of urokinase-typeplasminogen activator gene expression in human lung and breast carcinomas.Cancer Res., 47:4043-4046, 1987.
3. Nelson, L., Hansen, J., Skriver, L., Wilson, E., Kaltoft, K., Zenthen, J., andDaño.K. Purification of zymogen to plasminogen activator from humanglioblastoma cells by affinity chromatography with monoclonal antibody.Biochemistry, 21:6410-6415. 1982.
4. Stump. D.. Thienpont. M.. and Collen. D. Urokinase-related proteins inhuman urine. J. Biol. Chem., 261: 1267-1273, 1986.
5. Liotta, L., Goldfarb, R., Brundage, R.. Siegal, G., Terranova. V., and Garbisa,S. Effect of plasminogen activator (urokinase). plasmin, and thrombin onglycoprotein and collagenous components of basement membrane. CancerRes., 41: 4629-4636, 1981.
6. Goldfarb, R., Murano. G., Brundage. R.. Siegal. G.. Terranova, V., Garbisa,S., and Liotta, L. Degradation of glycoprotein and collagenous componentsof the basement membrane: studies with urokinase-type plasminogen activator, thrombin and plasmin. Sem. Thrombosis Hemostasis, 12: 335-336,1986.
7. de Bruin. P.. Griffioen, G., Verspaget, H., Verheijen, I., and Lamers, C.Plasminogen activators and tumor development in the human colon: activity
levels in normal mucosa, adenomatous polyps and adenocarcinomas. CancerRes., 47: 4654-4657, 1987.
8. Barsky. S., Siegal, G.. Jannotta, F., and Liotta. L. Loss of basement membrane components by invasive tumors but not by their benign counterparts.Lab. Invest., 49: 140-147. 1983.
9. Burtin, P., Chavanel, G.. Foidart, J.. and Martin, E. Antigens of the basementmembrane and the peritumoral stroma in human colonie adenocarcinomas:an immunofluorescencc study. Int. J. Cancer, 30: 13-20. 1982.
10. Ossowski, L., and Reich, E. Antibodies to plasminogen activator inhibithuman tumor métastases.Cell. 35: 611-619. 1983.
11. Hearing, V., Law, L., Corti, A., Appella, E., and Blasi. F. Modulation ofmetastatic potential by cell surface urokinase of murine melanoma cells.Cancer Res., 48: 1270-1278. 1988.
12. Vassalli. J.. Baccino, D.. and Belin, D. A cellular binding site for the M,55,000 form of the human plasminogen activator, urokinase. J. Cell. Biol.,700:86-92, 1985.
13. Stoppelli. M.. Tacchetti, C., Cubellis, M., Corti. A., Hearing. V.. Cassani.G., Apella, E., and Blasi. F. Autocrine saturation of pro-urokinase receptorson human A431 cells. Cell. 45: 675-684. 1986.
14. Boyd, D., Florent. G., Kim. P.. and Brattain. M. Determination of the levelsof urokinase and its receptor in human colon carcinoma cell lines. CancerRes.. 48: 3112-3116. 1988.
15. Hébert.C., and Baker, J. Linkage of extracellular plasminogen activator tothe fibroblast cytoskeleton: colocalization of cell surface urokinase withvinculin. J. Cell. Biol.. 106: 1241-1247. 1988.
16. Blasi, F., Vassalli, J., and Dano, K. Urokinase-type plasminogen activator:proenzyme. receptor and inhibitors. J. Cell. Biol.. 104: 801-804. 1987.
17. Ossowski, L. In vivo invasion of modified chorioallantoic membrane by tumorcells: the role of cell surface-bound urokinase. J. Cell. Biol., 107: 2437-2445.1988.
18. Quigley. J.. Goldfarb, R., Scheiner, C., O'Donnell-Tormey. J., and Yeo, T.
Plasminogen activator and the membrane of transformed cells. In: R. Hynesand C. Fox (eds.). Tumor Cell Surfaces and Malignancy. Vol. 41. pp. 773-796. New York: Alan R. Liss. Inc., 1980.
19. Brattain. M.. Levine. A.. Chakrabarty, S., Yeoman, L., Willson, J., and Long,B. Heterogeneity of human colon carcinoma. Cancer MétastasesRev.. 3:177-191. 1984.
20. Chantret, L, Barbat, A., Dussaulx. E., Brattain, M.. and Zweibaum, A.Epithelial polarity, villin expression and enterocytic differentiation of cultured colon carcinoma cells: A survey of twenty cell lines. Cancer Res.. 48:1936-1942, 1988.
21. Brattain, D.. Matthews. K., and Brattain. M. Growth factor requirements ofhuman colon carcinoma cells. Proc. Am. Assoc. Cancer Res., 27:42, 1986.
22. Boyd. D., Levine, A., Brattain. D., McKnight. M., and Brattain, M. Comparison of growth requirements of two human intratumoral colon carcinomacell lines in monolayer and soft agarose. Cancer Res., 48: 2469-2474. 1988.
23. Boyd, D.. Ziober. B.. Chakrabarty. S.. and Brattain. M. Examination ofurokinase protein/transcript levels and their relationship with laminin degradation in cultured colon carcinoma. Cancer Res., 49: 816-820, 1989.
24. Laemmli, U. Cleavage of structural proteins during the assembly of the headof bacteriophage T4. Nature (Lond.), 227: 680-685. 1970.
25. Darras, V., Thienpont, M., Stump, D., and Collen, D. Measurement ofurokinase-type plasminogen activator (U-PA) with an enzyme linked immu-nosorbent assay (ELISA) based on three murine monoclonal antibodies.Thrombosis Haemostasis, 56:411-414. 1986.
26. Appella. E.. Robinson, E., Ullrich. S.. Stoppelli. M., Corti. A., Cassani, G..and Blasi, F. The receptor-binding sequence of urokinase. J. Biol. Chem.,262:4437-4440,1987.
27. Watkins, L., Brattain, M., and Levine, A. Modulation of a high molecularweight form of transforming growth factor-a in human colon carcinoma celllines. Cancer Lett., 40: 59-70, 1988.
28. Plow. E.. Freaney, D., Plescia, J., and Miles, L. The plasminogen systemand cell surfaces: evidence for plasminogen and urokinase receptors on thesame cell type. J. Cell. Biol.. 103: 2411-2420, 1986.
29. Chapman, H., Vavrin, Z.. and Hibbs. J. Macrophage fibrinolytic activity:identification of two pathways of plasmin formation by intact cells and of aplasminogen activator inhibitor. Cell. 28: 653-662, 1982.
30. Kleinman, H., Cannon, F., Laurie, G., Hassell, J., Aumailley, M., Terranova,V., Martin, G., and DuBois-Dalcq, M. Biological activities of laminin. J.Cell Biochem., 27: 317-325, 1985.
31. Tryggvason, K., Hoyhtya, M.. and Salo, T. Proteolytic degradation of extracellular matrix in tumor invasion. Biochim. Biophys. Acta. 907: 191-217,1987.
32. Jones, P., and DeClerck. Y. Extracellular matrix destruction by invasivetumor cells. Cancer MétastasesRev., 3: 177-191, 1984.
33. Niedbala, M., Madiyalakan, R., Matta, K.. Crickard, K., Sharma, M., andBernacki, R. Role of glycosidases in human ovarian carcinoma cell mediateddegradation of subendothelial extracellular matrix. Cancer Res., 47: 4634-4641. 1987.
34. Rozhin. J.. Robinson, D., Stevens, M., Lah, T., Honn, K., Ryan. R., andSloane. B. Properties of a plasma membrane-associated cathepsin B-likecysteine proteinase in metastatic B16 melanoma variants. Cancer Res., 47:6620-6628. 1987.
35. Scatchard, G. The attractions of proteins for small molecules and ions. Ann.NY Acad. Sci.. 51: 660-672. 1949.
6069
Research. on August 13, 2014. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from




















