
Adrenaline induces chemoresistance in HT-29 colon adenocarcinoma cells
7
Adrenaline induces chemoresistance in HT-29 colon adenocarcinoma cells Herui Yao a, * , Zhaohui Duan b , Minghui Wang c , Awoniyi O. Awonuga d , Daniel Rappolee d,e , Yufen Xie d a Department of Oncology, The Second Affiliated Hospital of Zhongshan University, 107 West Yanjiang Road, Guangzhou, 510120, Guangdong, China b Laboratory of Clinical Immunology, The Second Affiliated Hospital of Zhongshan University, Guangzhou, Guangdong, China c Department of Surgery, The Second Affiliated Hospital of Zhongshan University, Guangzhou, Guangdong, China d Department of Obstetrics and Gynecology, Reproductive Sciences, Wayne State University School of Medicine, Detroit, MI 48201 e Karmanos Cancer Institute, Wayne State University School of Medicine, Detroit, MI 48201 Received 23 September 2008; received in revised form 2 December 2008; accepted 22 December 2008 Abstract Psychological distress and its ensuing chronic elevation of plasma catecholamines (adrenaline and noradrenaline) lead to poor response of tumors to chemotherapy, and constitute a poor prognostic factor for survival. Colorectal cancer patients suffer from various forms of psychological stress reflected in elevated plasma catecholamines, and their cancer cells express adrenergic receptors. Our objective was to investigate whether adrenergic activation contributes to the chemoresistance of colon cancers, and to explore the signal transduction pathway involved in the activation. The mRNA expression of the ABCB1 gene (previously MDR1) in human colon carcinoma HT-29 cell line was measured after treatment with an adrenergic receptor agonist (adrenaline) and various antagonists (propranolol, prazosin, and yohimbine). The function of P-glycoprotein, the protein product of the ABCB1 gene, was assessed by rhodamine 123 (Rh123)eretention assay, and chemo- sensitivity was determined by evaluating the cytotoxicity of 5-fluorouracil (5-FU) on the tumor cells. Increased ABCB1 mRNA expression and P-glycoprotein function levels in HT-29 cells by adrenaline was dose-dependent. This was accompanied by promotion of Rh123 efflux, and resis- tance to the growth-inhibiting effect of 5-FU in the tumor cells. The a 2 -adrenergic receptor antag- onist yohimbine completely abolished the induction of ABCB1 mRNA, the stimulatory effect of adrenaline on Rh123 efflux, and the growth-inhibiting effect of 5-FU. The a 1 -adrenergic receptor and b-adrenergic receptor antagonists did not inhibit the induction of ABCB1. The stimulating effects were coupled with extracellular receptor kinase 1/2 (Erk1/2) phosphorylation, but were not associated with protein kinase A activity. We conclude that adrenaline induces multidrug resis- tance in colon cancer cells by upregulating ABCB1 gene expression via a 2 -adrenergic receptors, and such effects were associated with the mitogen activated protein kinase (MAPK) pathway. Ó 2009 Elsevier Inc. All rights reserved. 1. Introduction Cancer patients undergoing chemotherapy experience associated heightened psychological distress, which in turn has a major effect on treatment efficacy [1e4]. The status of psychological distress with the ensuing chronic elevation of plasma catecholamines [5] predicts poor response of the tumors to chemotherapy, and may serve as an independent prognostic factor for survival [6]. Recent studies on a mouse model bearing MCF-7 breast cancers have shown that psychological stress induced by repetitive restraint of the animals results in chemoresistance of the tumors to chemo- therapeutic drugs via adrenergic upregulation of the Abcb1 gene (previously Mdr1) [7]. This was supported by studies on mice bearing 26-L5 colon carcinoma, Shionogi (SC115) mammary carcinoma, or TLX5 lymphoma that are exposed to rotational stress showing a decrease in the antitumor effects of chemotherapeutic drugs in terms of tumor burden, extent of metastasis, and survival time [8e11]. The P-glycoprotein encoded by ABCB1 is a plasma membrane glycoprotein that confers a multidrug-resistance (MDR) phenotype by virtue of its ability to exclude a wide range of chemotherapeutic drugs and other hydrophobic * Corresponding author. E-mail address: [email protected] (H. Yao). 0165-4608/09/$ e see front matter Ó 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.cancergencyto.2008.12.009 Cancer Genetics and Cytogenetics 190 (2009) 81e87
Transcript of Adrenaline induces chemoresistance in HT-29 colon adenocarcinoma cells

Cancer Genetics and Cytogenetics 190 (2009) 81e87
Adrenaline induces chemoresistance in HT-29colon adenocarcinoma cells
Herui Yaoa,*, Zhaohui Duanb, Minghui Wangc, Awoniyi O. Awonugad,Daniel Rappoleed,e, Yufen Xied
aDepartment of Oncology, The Second Affiliated Hospital of Zhongshan University, 107 West Yanjiang Road, Guangzhou, 510120, Guangdong, ChinabLaboratory of Clinical Immunology, The Second Affiliated Hospital of Zhongshan University, Guangzhou, Guangdong, China
cDepartment of Surgery, The Second Affiliated Hospital of Zhongshan University, Guangzhou, Guangdong, ChinadDepartment of Obstetrics and Gynecology, Reproductive Sciences, Wayne State University School of Medicine, Detroit, MI 48201
eKarmanos Cancer Institute, Wayne State University School of Medicine, Detroit, MI 48201
Received 23 September 2008; received in revised form 2 December 2008; accepted 22 December 2008
Abstract Psychological distress and its ensuing chronic e
* Corresponding a
E-mail address: y
0165-4608/09/$ e see
doi:10.1016/j.cancerg
levation of plasma catecholamines (adrenaline andnoradrenaline) lead to poor response of tumors to chemotherapy, and constitute a poor prognosticfactor for survival. Colorectal cancer patients suffer from various forms of psychological stressreflected in elevated plasma catecholamines, and their cancer cells express adrenergic receptors.Our objective was to investigate whether adrenergic activation contributes to the chemoresistanceof colon cancers, and to explore the signal transduction pathway involved in the activation. ThemRNA expression of the ABCB1 gene (previously MDR1) in human colon carcinoma HT-29 cellline was measured after treatment with an adrenergic receptor agonist (adrenaline) and variousantagonists (propranolol, prazosin, and yohimbine). The function of P-glycoprotein, the proteinproduct of the ABCB1 gene, was assessed by rhodamine 123 (Rh123)eretention assay, and chemo-sensitivity was determined by evaluating the cytotoxicity of 5-fluorouracil (5-FU) on the tumorcells. Increased ABCB1 mRNA expression and P-glycoprotein function levels in HT-29 cells byadrenaline was dose-dependent. This was accompanied by promotion of Rh123 efflux, and resis-tance to the growth-inhibiting effect of 5-FU in the tumor cells. The a2-adrenergic receptor antag-onist yohimbine completely abolished the induction of ABCB1 mRNA, the stimulatory effect ofadrenaline on Rh123 efflux, and the growth-inhibiting effect of 5-FU. The a1-adrenergic receptorand b-adrenergic receptor antagonists did not inhibit the induction of ABCB1. The stimulatingeffects were coupled with extracellular receptor kinase 1/2 (Erk1/2) phosphorylation, but werenot associated with protein kinase A activity. We conclude that adrenaline induces multidrug resis-tance in colon cancer cells by upregulating ABCB1 gene expression via a2-adrenergic receptors, andsuch effects were associated with the mitogen activated protein kinase (MAPK) pathway. � 2009Elsevier Inc. All rights reserved.
1. Introduction
Cancer patients undergoing chemotherapy experienceassociated heightened psychological distress, which in turnhas a major effect on treatment efficacy [1e4]. The statusof psychological distress with the ensuing chronic elevationof plasma catecholamines [5] predicts poor response of thetumors to chemotherapy, and may serve as an independentprognostic factor for survival [6]. Recent studies on a mousemodel bearing MCF-7 breast cancers have shown that
uthor.
[email protected] (H. Yao).
front matter � 2009 Elsevier Inc. All rights reserved.
encyto.2008.12.009
psychological stress induced by repetitive restraint of theanimals results in chemoresistance of the tumors to chemo-therapeutic drugs via adrenergic upregulation of the Abcb1gene (previously Mdr1) [7]. This was supported by studieson mice bearing 26-L5 colon carcinoma, Shionogi (SC115)mammary carcinoma, or TLX5 lymphoma that are exposedto rotational stress showing a decrease in the antitumoreffects of chemotherapeutic drugs in terms of tumorburden, extent of metastasis, and survival time [8e11].
The P-glycoprotein encoded by ABCB1 is a plasmamembrane glycoprotein that confers a multidrug-resistance(MDR) phenotype by virtue of its ability to exclude a widerange of chemotherapeutic drugs and other hydrophobic

82 H. Yao et al. / Cancer Genetics and Cytogenetics 190 (2009) 81e87
compounds such as the fluorescent dye rhodamine 123(Rh123) from tumors cell in an ATP-dependent manner.P-glycoprotein function in tumor cells can be assessed withflow-cytometric analysis using Rh123-retention assay.
The human colon is particularly sensitive to stressbecause of the close distribution of noradrenergic fibersto its basement lamina [12], and both human colonicepithelial cells and adenocarcinoma cells express adren-ergic receptors [13], notably including a2-adrenergic recep-tors on the cell surface [14,15]. Furthermore, the number ofa2-adrenergic receptors in immature colonic epithelial orcarcinoma cells is severalfold higher than in mature cells[16]. In this study, we investigated whether a human colonadenocarcinoma cell line, HT-29, which is abundant in a2-adrenergic receptors, responds to adrenaline treatment anddevelops chemoresistance. The objectives were to deter-mine whether adrenergic activation contributes to the che-moresistance of colon cancers and to explore the signaltransduction pathway underlying the response.
2. Materials and methods
2.1. Cell culture and stimulation
Human colon adenocarcinoma HT-29 cells (AmericanType Culture Collection [ATCC], Manassas, VA) werecultured in Dulbecco’s modified Eagle’s medium (DMEM;Gibco, Grand Island, NY) supplemented with 10% heat-inactivated fetal calf serum (FCS; BoehringereMannheim,Mannheim, Germany), 25 mmol/L glucose, 0.01 mg/mLgentamycin, and 0.1 mmol/L nonessential amino acid(Gibco) at 37�C and 5% CO2. For catecholamine stimula-tion, adrenaline (adrenaline bitartrate; SigmaeAldrich,Munich, Germany), a pan-adrenergic receptor agonist,was added to the medium at various concentrations for indi-cated times. To block the catecholamine effect, the b-adren-ergic receptor antagonist propranolol (10 mmol/L) andeither the a1-adrenergic receptor antagonist prazosin (10mmol/L) or the selective a2-adrenergic receptor blockeryohimbine (1 mmol/L) (yohimbine HCl, SigmaeAldrich)were added to the medium at the peak concentration ofadrenaline for 6 hours prior to adrenaline stimulation.
2.2. Real time reverse transcriptaseepolymerase chainreaction for ABCB1 mRNA expression
The mRNA expression of the multiple drug resistance geneABCB1 in HT-29 cells was quantified 24 hours after adrenalinestimulation using a TaqMan quantitative real-time reversetranscriptaseepolymerase chain reaction (real time RT-PCR)assay. Briefly, total RNAwas isolated from the cells using Tri-zol reagent (Life Technologies, Gaithersburg, MD). We de-signed primers for ABCB1 and GAPDH according topublished sequences [7], and performed a one-step real-timeRT-PCR using SYBR green reagent (Applied Biosystems,Foster City, CA). All reactions were done in triplicate in
a 25-mL reaction volume according to the manufacturer’sinstructions. After reverse transcription for 30 minutes at48�C and Taq activation for 10 minutes at 95�C, we performed40 cycles of PCR at 95�C for 20 seconds (melt), 55�C for 30seconds (anneal), and 72�C for 30 seconds (extension).Real-time expression of ABCB1 and GAPDH mRNA wasmeasured on an ABI Prism 5700 sequence detection system(Applied Biosystems) by calculating the threshold cycles.Relative amount of ABCB1 mRNA was standardized to theexpression of GAPDH mRNA. A positive control with cDNAfrom MCF-7/Adr cells and negative controls with no templateand genomic DNA as templates were added in every experi-ment. To block the adrenaline effect, the cells were pretreatedwith yohimbine for 6 hours prior to adrenaline stimulation inanother group. GAPDH and ABCB1 mRNA was detected atthe same linear part of the PCR cycle.
2.3. Rh123-retention assay for P-glycoprotein function
The fluorescent compound Rh123 is a known substrate forP-glycoprotein, and efflux of intracellular Rh123 out ofliving cells relies on normal function of the protein. Hence,we evaluated P-glycoprotein function of the tumor cells byanalyzing Rh123 retention in the cells. After 24 hours ofadrenergic stimulation (to block the adrenaline effect,pretreatment with yohimbine was undertaken for 6 hoursprior to adrenaline in another group), cells were seeded insix-well culture plates at a concentration of 4 � 105 cells/mL. The Rh123 (SigmaeAldrich) was added to yield a finalconcentration of 200 ng/mL. After incubation for 45 minutes,cells were washed and fed with Rh123-free culture mediumat 37�C. Cellular efflux of Rh123 was measured over 60minutes at 10-minute time intervals by monitoring its fluores-cence decrease at an emission wavelength of 525 nm on a flowcytometer (BD Biosciences, San Jose, CA). The results wereexpressed as the mean of fluorescence intensity.
2.4. MTT assay
The influence of adrenaline stimulation on chemosensitiv-ity of HT-29 cells was investigated by first treating stimulatedtumor cells with 5-fluorouracil (5-FU) and then evaluatingthe viability of the tumor cells with a 3-(4,5-dimethyl-2-thia-zole)-2,5-diphenyl-[2H]-tetrazolium bromide (MTT) assay.For treatment with 5-FU, the HT-29 cells stimulated byadrenaline with or without yohimbine and the nonstimulatedcontrols were cultured in fresh medium, with 5-FU(SigmaeAldrich) at various concentrations, and then incu-bated for 72 hours at 37�C. After drug treatment, cells wereharvested and used for MTT assay.
The inhibitory effect of chemotherapy on tumor cellviability was investigated using MTT assay as previouslydescribed. Briefly, after treatment, 1 � 103 cells were platedinto each well of 96-well microtiter plates. Medium wasremoved, and the tumor cells were washed once with phos-phate-buffered saline; 100 mL 0.4% MTT (SigmaeAldrich)

83H. Yao et al. / Cancer Genetics and Cytogenetics 190 (2009) 81e87
in phosphate-buffered saline was added, and the cells werethen incubated for 3 hours at 37�C. The formazan dye wasdissolved in 150 mL dimethylsulfoxide (DMSO; SigmaeAldrich), and the absorbance was measured at 540 nm ina microplate reader, with the absorbance of untreatedcontrols taken as 100% survival.
2.5. Mitogen activated protein kinase activation
Activation of extracellular receptor kinase (Erk) formsof mitogen activated protein kinase (MAPK) was evaluatedby immunoblot analysis of Erk1/2 phosphorylation. Briefly,after stimulation for 15 minutes with adrenaline with orwithout yohimbine, these cells and those of nonstimulatedcontrols, were harvested, and proteins were separated byelectrophoresis on 12% polyacrylamide gel. The gel wasthen electrotransferred overnight onto Hybond-C Extranitrocellulose membranes (pore size 0.45 mm; AmershamPharmacia Biotech, Little Chalfont, UK) in transfer buffer(20% methanol; 0.19 mol/L glycine; 25 mmol/L Tris base).To assess activated MAPK content, the nitrocellulose blotwas incubated in blocking buffer (5% nonfat milk, 0.1%Tween 20 in Tris-buffered saline) for 1 hour at roomtemperature to quench nonspecific binding, and then probedovernight at 4�C with a rabbit polyclonal primary antibodythat recognizes dually phosphorylated (Thr/Tyr) Erk formsof MAP kinase (1:500 dilution in blocking buffer; NewEngland Biolabs, Beverly, MA). After washing in TBST(2.42 g/L Tris base; 8.0 g/L NaCl; 0.1% Tween 20; pH7.6), the primary antibody was detected with a goat anti-rabbit horseradish peroxidase-linked secondary antibody(1:2000 dilution in blocking buffer; New England Biolabs)and enhanced chemiluminescence (ECL; Amersham Phar-macia Biotech). The blots were then stripped with strippingbuffer (62.5 mmol/L Tris-HCl, pH 6.8; 2% SDS; 100 mmol/L2-mercaptoethanol) for 30 minutes at 65�C and reprobedovernight at 4�C with an anti-rabbit polyclonal primaryantibody to total MAPK (1:500 dilution in blocking buffer;New England Biolabs), and then by goat anti-rabbitsecondary antibody as described above. The blots wereexposed to Kodak X-Omat RP films (Rochester, NY).
2.6. Protein kinase A assays
To measure the PKA activity of adrenaline-treated HT-29cells, an enzyme-linked immunosorbent assay (ELISA)ebased kit for PKA activity (Calbiochem-Novabiochem, SanDiego, CA), employing a synthetic PKA pseudosubstrateand a monoclonal antibody, were used according to the instruc-tion manual. Briefly, cell extracts were obtained by lysing thecells in 500 mL lysis buffer: 20 mmol/L tris-HCl, pH 7.5, 5mmol/L ethylene-diamine-tetraacetic acid (EDTA), 10mmol/L ethylene-glycol-bis-tetraacetic acid (EGTA), 0.3%b-mercaptoethanol, 1 mmol/L phenylmethylsulphonyl fluo-ride, and 10 mmol/L benzamidine. The protein was extractedusing sonication for 30 seconds, followed by centrifugation at
16,000 g for 45 minutes at 4�C, recovering the supernatant. Tomeasure PKA activity, protein was incubated with componentmixture contained in the kit in the pseudosubstrate-coatedwells at room temperature for 20 minutes. A biotinylated anti-body for the phosphorylated substrate was added to the wellsafter stringent washing, and incubated at room temperaturefor 60 minutes. Subsequently, a peroxidase-conjugated strep-tavidin was added and incubated for 60 minutes at roomtemperature. For color development, a phenylenediaminesubstrate solution was added, and the optical density valueof each well was measured at 492 nm with a microplatereader. Values were expressed as percentage increase overresting activity of the untreated controls.
2.7. Statistical analysis
Data were expressed as mean 6 standard error of themean. Statistical analysis was performed by one-way anal-ysis of variance (ANOVA) and comparisons among groupswere performed by Bonferroni’s multiple-comparisont-test. The significance level was set at P ! 0.05.
3. Results
3.1. Adrenergic modulation of ABCB1 expressionin HT-29 cells
To evaluate the effect of adrenaline on ABCB1 expressionin HT-29 cells, we treated the cultures with adrenaline for 24hours and measured the expression of ABCB1 by means ofquantitative real time RT-PCR. Adrenaline induced ABCB1mRNA expression in a dose-dependent manner; ABCB1mRNA levels increased with increasing concentrations ofadrenaline (Fig. 1), reaching a maximal level of O60-foldrelative to the control (2.47 6 0.46 vs. 0.041 6 0.024 timesGAPDH; P ! 0.001) at 1.25 mmol/L adrenaline. Furtherincreasing the concentration of adrenaline up to 12.5 mmol/L did not enhance ABCB1 expression, indicating that theadrenergic effect reaches a plateau at 1.25 mmol/L.
To test which isoform of adrenergic receptor mediated theadrenaline-induced upregulation of ABCB1, different adren-ergic receptor blockers were used (Fig. 2). The b-adrenergicreceptor antagonist (propranolol; 10 mmol/L) and thea1-adrenergic receptor antagonist (prazosin; 10 mmol/L)did not significantly influence adrenaline-induced ABCB1expression (P O 0.05), but the a2-adrenergic receptor antag-onist (yohimbine; 1 mmol/L) completely abolished the stim-ulatory effect of adrenaline on ABCB1 expression (P !0.01). Thus, a2-adrenergic receptors alone mediated theadrenaline-induced ABCB1 expression in HT-29 cells.
3.2. Effect of adrenaline on Rh123 retentionin HT-29 cells
To examine the functional importance of the adrenalineeffects on ABCB1 expression, retention of Rh123 was

0
0.5
1
1.5
2
2.5
3
3.5
0 0.01 0.05 0.25 1.25 6.25 12.5Adrenaline concentrations (uM)
AB
CB
1 m
RN
A /G
AP
DH
m
RN
A
*
*
*
*
Fig. 1. Adrenaline induces expression of ABCB1 (previously MDR1). Error
bars represent the standard deviation of the mean. All experiments were per-
formed in triplicate. HT-29 colon adenocarcinoma cells treated with indi-
cated concentrations of adrenaline for 24 hours. Total RNA was extracted
from the cells, and real-time RT-PCR was performed for ABCB1 mRNA.
Treatment with adrenaline increases the expression of the ABCB1 gene at
mRNA level in a dose-dependent manner. * Significant at P ! 0.001
compared with cells treated with drugs at a lower concentration level.
0
20
40
60
80
100
0 10 20 30 40 50 60Time(min)
Rh
123 reten
tio
n(%
)
ControlAdrAdr+Yh
Fig. 3. Adrenaline enhances ABCB1 function via a2-adrenergic receptors
by retaining a fluorescent compound, rhodamine 123 (Rh123) in the HT-29
colon adenocarcinoma cells. Error bars represent the standard deviation of
the mean. All experiments were performed in triplicate. The Rh123 reten-
tion in HT-29 cells was determined by FACScan analysis after treatment
with media alone as control, adrenaline (1.25 mmol/L), adrenaline (1.25
mmol/L) þ yohimbine (1 mmol/L) for different periods of time. Abbrevia-tions: Adr, adrenaline; Yh, yohimbine.
84 H. Yao et al. / Cancer Genetics and Cytogenetics 190 (2009) 81e87
determined by flow cytometric analysis, exploiting the fluo-rescent properties of this compound to show the function ofP-glycoprotein that is encoded by ABCB1. Rh123 retention,expressed as mean fluorescence intensity of the compound,decreased to a much lower percentage in HT-29 cells stimu-lated with adrenaline (1.25 mmol/L) than in the untreatedcontrols (37.4 6 5.4% vs. 96.2 6 2.8%; P ! 0.01) after incu-bation for 10 minutes in Rh123-free culture media (Fig. 3).An even higher disparity (13.8 6 3.7% vs. 78.6 6 1.9%;P ! 0.01) was observed after incubation for 60 minutes (the
0
0.5
1
1.5
2
2.5
3
3.5
Control Adr Adr+Pra Adr+Yoh Adr+Pro
Treatments
AB
CB
1 m
RN
A/G
AP
DH
m
RN
A
P<0.001
Fig. 2. Blocking the a2-adrenergic receptors reverses adrenaline-induced
ABCB1 (mdr1) expression. Error bars represent the standard deviation of
the mean. All experiments were performed in triplicate. HT-29 colon adeno-
carcinoma cells were treated with adrenaline (1.25 mmol/L) plus various
antagonists of adrenergic receptors as indicated. Total RNA was extracted
from the cells, and real-time reverse transcriptaseepolymerase chain reac-
tion was performed for ABCB1 mRNA. Addition of yohimbine (1 mmol/
L), an a2 adrenergic receptor antagonist, instead of propranolol (10 mmol/
L), a b-adrenergic receptor blocker, or prazosin (10 mmol/L), an a1-adren-
ergic receptor blocker, abrogates the adrenaline-induced ABCB1 expression.
Abbreviations: Adr, adrenaline; Pra, prazosin; Pro, propranolol.
end time point of the test). Hence, adrenaline (1.25 mmol/L)stimulation resulted in a fairly rapid decrease of intracellularRh123 in HT-29 cells, whereas the untreated controls re-tained the dye. In addition, adrenaline-induced reduction ofRh123 retention was reversed by the a2-adrenergic receptorblocker (yohimbine; 1 mmol/L) (Fig. 3). Thus, the enhancingeffect of adrenaline on functional levels of P-glycoproteinwas mediated by a2-adrenergic receptors.
3.3. Influence of adrenaline on chemosensitivity ofHT-29 cells
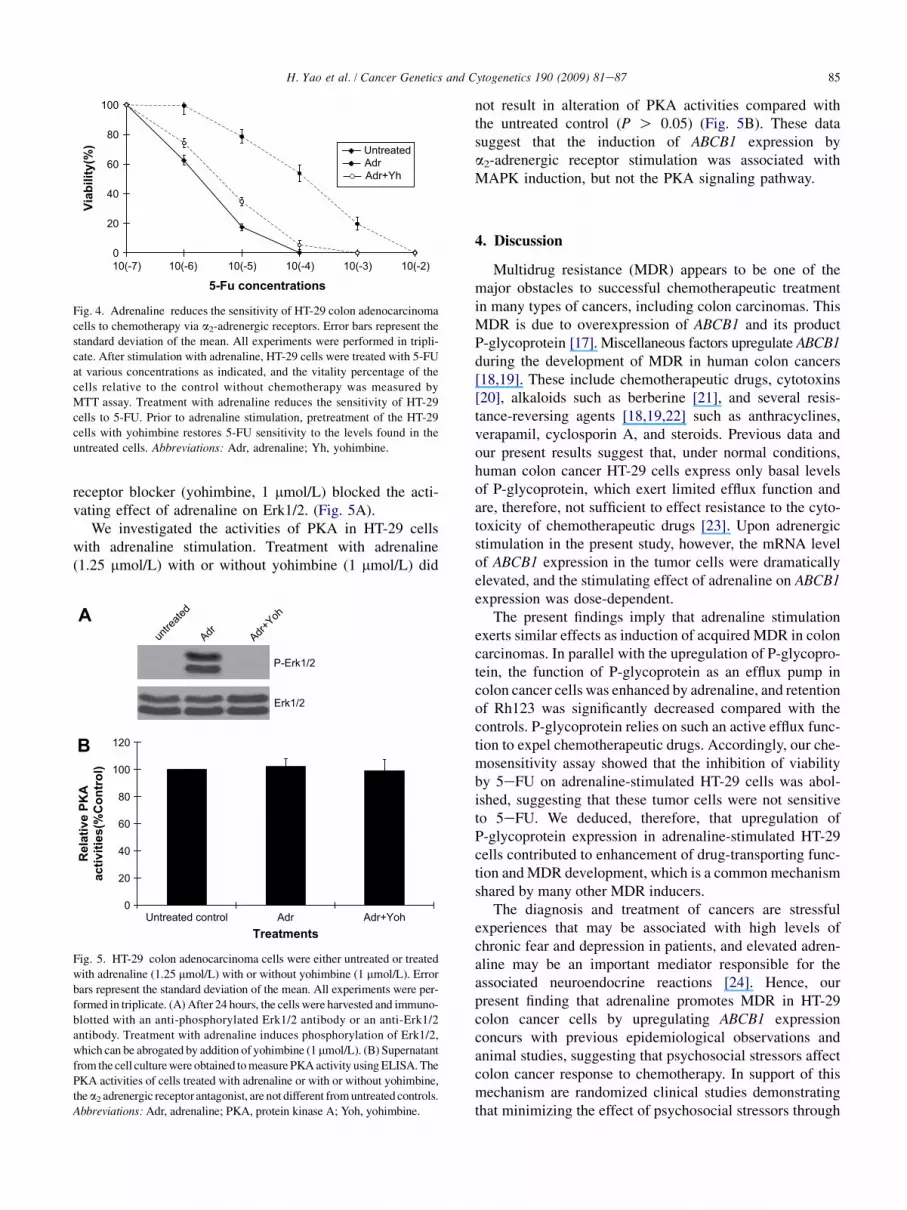
In order to test whether the enhanced expression andefflux of P-glycoprotein induced by adrenaline correlate withdrug resistance in HT-29 cells, we further investigated theeffect of adrenaline stimulation on chemosensitivity of thetumor cells by evaluating growth inhibition induced by 5-FU. Stimulation with adrenaline (1.25 mmol/L) resulted inresistance of HT-29 cells to the growth inhibiting activityof 5-FU (Fig. 4). The 5-FU was approximately 130-fold lesseffective in killing adrenaline-pretreated HT-29 cells. Thesensitizing effect of adrenaline was significantly reversedby pretreating the cells with the a2-adrenergic receptorblocker, yohimbine, at a concentration of 1 mmol/L.
3.4. MAPK Erk1/2 activation induced by adrenaline
We further investigated the signaling pathway of adren-aline-induced ABCB1 expression by examining the Erk1/2phosphorylation and PKA activity. Using immunoblotswith antibodies against the Erk1/2 protein (in general) orthe phosphorylated protein, we observed that incubationof HT-29 cells with 1.25 mmol/L adrenaline for 15 minutesresulted in activation of Erk1/2, as evidenced by increasedlevels of the phosphorylated form of Erk1/2 compared tothe control (Fig. 5A). Furthermore, the a2-adrenergic
0
20
40
60
80
100
10(-7) 10(-6) 10(-5) 10(-4) 10(-3) 10(-2)
5-Fu concentrations
Via
bility
(%
) UntreatedAdrAdr+Yh
Fig. 4. Adrenaline reduces the sensitivity of HT-29 colon adenocarcinoma
cells to chemotherapy via a2-adrenergic receptors. Error bars represent the
standard deviation of the mean. All experiments were performed in tripli-
cate. After stimulation with adrenaline, HT-29 cells were treated with 5-FU
at various concentrations as indicated, and the vitality percentage of the
cells relative to the control without chemotherapy was measured by
MTT assay. Treatment with adrenaline reduces the sensitivity of HT-29
cells to 5-FU. Prior to adrenaline stimulation, pretreatment of the HT-29
cells with yohimbine restores 5-FU sensitivity to the levels found in the
untreated cells. Abbreviations: Adr, adrenaline; Yh, yohimbine.
85H. Yao et al. / Cancer Genetics and Cytogenetics 190 (2009) 81e87
receptor blocker (yohimbine, 1 mmol/L) blocked the acti-vating effect of adrenaline on Erk1/2. (Fig. 5A).
We investigated the activities of PKA in HT-29 cellswith adrenaline stimulation. Treatment with adrenaline(1.25 mmol/L) with or without yohimbine (1 mmol/L) did
0
20
40
60
80
100
120
Untreated control Adr Adr+YohTreatments
Relative
P
KA
activities(%
Co
ntro
l)
A
B
Erk1/2
P-Erk1/2
Adr+Yoh
untre
ated
Adr
Fig. 5. HT-29 colon adenocarcinoma cells were either untreated or treated
with adrenaline (1.25 mmol/L) with or without yohimbine (1 mmol/L). Error
bars represent the standard deviation of the mean. All experiments were per-
formed in triplicate. (A) After 24 hours, the cells were harvested and immuno-
blotted with an anti-phosphorylated Erk1/2 antibody or an anti-Erk1/2
antibody. Treatment with adrenaline induces phosphorylation of Erk1/2,
which can be abrogated by addition of yohimbine (1 mmol/L). (B) Supernatant
from the cell culture were obtained to measure PKA activity using ELISA. The
PKA activities of cells treated with adrenaline or with or without yohimbine,
the a2 adrenergic receptor antagonist, are not different from untreated controls.
Abbreviations: Adr, adrenaline; PKA, protein kinase A; Yoh, yohimbine.
not result in alteration of PKA activities compared withthe untreated control (P O 0.05) (Fig. 5B). These datasuggest that the induction of ABCB1 expression bya2-adrenergic receptor stimulation was associated withMAPK induction, but not the PKA signaling pathway.
4. Discussion
Multidrug resistance (MDR) appears to be one of themajor obstacles to successful chemotherapeutic treatmentin many types of cancers, including colon carcinomas. ThisMDR is due to overexpression of ABCB1 and its productP-glycoprotein [17]. Miscellaneous factors upregulate ABCB1during the development of MDR in human colon cancers[18,19]. These include chemotherapeutic drugs, cytotoxins[20], alkaloids such as berberine [21], and several resis-tance-reversing agents [18,19,22] such as anthracyclines,verapamil, cyclosporin A, and steroids. Previous data andour present results suggest that, under normal conditions,human colon cancer HT-29 cells express only basal levelsof P-glycoprotein, which exert limited efflux function andare, therefore, not sufficient to effect resistance to the cyto-toxicity of chemotherapeutic drugs [23]. Upon adrenergicstimulation in the present study, however, the mRNA levelof ABCB1 expression in the tumor cells were dramaticallyelevated, and the stimulating effect of adrenaline on ABCB1expression was dose-dependent.
The present findings imply that adrenaline stimulationexerts similar effects as induction of acquired MDR in coloncarcinomas. In parallel with the upregulation of P-glycopro-tein, the function of P-glycoprotein as an efflux pump incolon cancer cells was enhanced by adrenaline, and retentionof Rh123 was significantly decreased compared with thecontrols. P-glycoprotein relies on such an active efflux func-tion to expel chemotherapeutic drugs. Accordingly, our che-mosensitivity assay showed that the inhibition of viabilityby 5eFU on adrenaline-stimulated HT-29 cells was abol-ished, suggesting that these tumor cells were not sensitiveto 5eFU. We deduced, therefore, that upregulation ofP-glycoprotein expression in adrenaline-stimulated HT-29cells contributed to enhancement of drug-transporting func-tion and MDR development, which is a common mechanismshared by many other MDR inducers.
The diagnosis and treatment of cancers are stressfulexperiences that may be associated with high levels ofchronic fear and depression in patients, and elevated adren-aline may be an important mediator responsible for theassociated neuroendocrine reactions [24]. Hence, ourpresent finding that adrenaline promotes MDR in HT-29colon cancer cells by upregulating ABCB1 expressionconcurs with previous epidemiological observations andanimal studies, suggesting that psychosocial stressors affectcolon cancer response to chemotherapy. In support of thismechanism are randomized clinical studies demonstratingthat minimizing the effect of psychosocial stressors through

86 H. Yao et al. / Cancer Genetics and Cytogenetics 190 (2009) 81e87
psychosocial intervention (e.g., relaxation therapy) reducedadrenaline levels, and thus was positively correlated withresponse to chemotherapy [25].
The effects of the neuroendocrine hormone adrenaline arecaused by the interaction of the catecholamine with specifichigh-affinity receptor sites on the external surface of targetcells. The HT-29 colon carcinoma cell line has been foundto express both a- and b-adrenergic receptors, and theirexpression can be modulated by environmental stimulus[26,27]. Hence, we further characterized the appropriatereceptors mediating the MDR-induction effect of adrenalinein the tumor cells. We found that only a2-receptors wereresponsible for MDR induction in HT-29 cells, as adrena-line-induced upregulation of P-glycoprotein expression andfunction was only blocked by the a2 adrenergic receptorblocker, yohimbine. This is in agreement with previous find-ings that most of the biochemical effects of catecholamineson HT-29 cells were mediated via a2 adrenergic receptors.
The a2-adrenergic receptors are linked to inhibition ofcAMP-dependent PKA [28] and activation of MAPK indepen-dently [29]. These kinase pathways play an important role inthe signal transduction of gene regulation, and might beinvolved in the regulation of ABCB1 expression under psycho-logical stress. In agreement with Ding et al. [30], we found thata2-adrenergic stimulation activates a MAPK family member,Erk1/2, suggesting the association of MAPK pathway withadrenaline-induced upregulation of ABCB1. On the otherhand, we found that adrenaline treatment with or without thea2-adrenergic blocker did not influence the basal activity ofPKA in HT-29 cells. Collectively, the a2-adrenergic stimula-tion of ABCB1 expression is probably mediated via the MAPKkinase pathway, rather than the PKA pathway.
In summary, adrenaline stimulation via a2-adrenergicreceptors contributes to upregulation of ABCB1 expressionin colon carcinoma cells via the MAPK signaling pathway,and is responsible for expelling and inducing MDR tochemotherapeutic agents in cancer therapy. These findingsprovide a molecular basis to explain the adverse influenceof psychological stress on the outcome of chemotherapyin colon cancers. The present study is limited, however,in providing insights into catecholamine stimulation ofMDR of colon cancer cells only in an artificial in vitrosystem. Given that MDR is a complex in vivo event,affected by neuroendocrine and immunological factors,several questions remain unaddressed. Thus, further in vivostudies are needed to elucidate the mechanisms by whichpsychological stress influences MDR.
Acknowledgments
This work was supported by research grants from theFoundation of the Department of Health of Guangdong Prov-ince (A2005226), the Science and Technology Foundation ofGuangdong Province (2006B36002010), and the NaturalScience Foundation of Guangdong Province (7001592).
References
[1] Carlson LE, Angen M, Cullum J, Goodey E, Koopmans J, Lamont L,
MacRae JH, Martin M, Pelletier G, Robinson J, Simpson JS,
Speca M, Tillotson L, Bultz BD. High levels of untreated distress
and fatigue in cancer patients. Br J Cancer 2004;90:2297e304.
[2] Herschbach P, Keller M, Knight L, Brandl T, Huber B, Henrich G,
Marten-Mittag B. Psychological problems of cancer patients: a cancer
distress screening with a cancer-specific questionnaire. Br J Cancer
2004;91:504e11.
[3] Montgomery GH, McClary KA, Bovbjerg DH. Adjuvant therapy for
breast cancer and psychological distress. Ann Oncol 1996;7:977e8.
[4] Sabbioni ME, Bovbjerg DH, Jacobsen PB, Manne SL, Redd WH.
Treatment related psychological distress during adjuvant chemo-
therapy as a conditioned response. Ann Oncol 1992;3:393e8.
[5] Rawl SM, Given BA, Given CW, Champion VL, Kozachik SL,
Barton D, Emsley CL, Williams SD. Intervention to improve psycho-
logical functioning for newly diagnosed patients with cancer. Oncol
Nurs Forum 2002;29:967e75.
[6] Kornblith AB, Herndon JE 2nd, Weiss RB, Zhang C, Zuckerman EL,
Rosenberg S, Mertz M, Payne D, Jane Massie M, Holland JF,
Wingate P, Norton L, Holland JC. Long-term adjustment of survivors
of early-stage breast carcinoma, 20 years after adjuvant chemo-
therapy. Cancer 2003;98:679e89.
[7] Su F, Ouyang N, Zhu P, Jia W, Gong C, Ma X, Xu H, Song E.
Psychological stress induces chemoresistance in breast cancer by up-
regulating mdr1. Biochem Biophys Res Commun 2005;329:888e97.
[8] Kerr LR, Hundal R, Silva WA, Emerman JT, Weinberg J. Effects of social
housing condition on chemotherapeutic efficacy in a Shionogi carcinoma
(SC115) mouse tumor model: influences of temporal factors, tumor size,
and tumor growth rate. Psychosom Med 2001;63:973e84.
[9] Strange KS, Kerr LR, Andrews HN, Emerman JT, Weinberg J. Psycho-
social stressors and mammary tumor growth: an animal model. Neuro-
toxicol Teratol 2000;22:89e102.
[10] Wu W, Murata J, Hayashi K, Yamaura T, Mitani N, Saiki I. Social
isolation stress impairs the resistance of mice to experimental liver
metastasis of murine colon 26-L5 carcinoma cells. Biol Pharm Bull
2001;24:772e6.
[11] Wu W, Yamaura T, Murakami K, Murata J, Matsumoto K,
Watanabe H, Saiki I. Social isolation stress enhanced liver metastasis
of murine colon 26-L5 carcinoma cells by suppressing immune
responses in mice. Life Sci 2000;66:1827e38.
[12] Weems WA, Szurszewski JH. An intracellular analysis of some
intrinsic factors controlling neural output from inferior mesenteric
ganglion of guinea pigs. J Neurophysiol 1978;41:305e21.
[13] Shepherd AP, Riedel GL, Keeton TK. Perfused rat intestine for study
of norepinephrine release. Am J Physiol 1988;254:H1187e93.
[14] Agrawal DK, Wildrick DM, Boman BM. Characteristics of a-adreno-
ceptors in two human colorectal cancer cell lines. Biochem Biophys
Res Commun 1992;185:176e84.
[15] Cantiello HF, Lanier SM. a2-Adrenergic receptors and the Naþ/Hþ
exchanger in the intestinal epithelial cell line, HT-29. J Biol Chem
1989;264:16000e7.
[16] Paris H, Bouscarel B, Cortinovis C, Murat JC. Growth-related varia-
tion of a2-adrenergic receptivity in the HT-29 adenocarcinoma cell-
line from human colon. FEBS Lett 1985;184:82e6.
[17] Meijer GA, Schroeijers AB, Flens MJ, Meuwissen SG, van der
Valk P, Baak JP, Scheper RJ. Increased expression of multidrug resis-
tance related proteins Pgp, MRP1, and LRP/MVP occurs early in
colorectal carcinogenesis. J Clin Pathol 1999;52:450e4.
[18] Kohno K, Sato S, Takano H, Matsuo K, Kuwano M. The direct acti-
vation of human multidrug resistance gene (MDR1) by anticancer
agents. Biochem Biophys Res Commun 1989;165:1415e21.
[19] Scotto KW. Transcriptional regulation of ABC drug transporters.
Oncogene 2003;22:7496e511.
[20] Guichard SM, Macpherson JS, Thurston DE, Jodrell DI. Influence of
P-glycoprotein expression on in vitro cytotoxicity and in vivo

87H. Yao et al. / Cancer Genetics and Cytogenetics 190 (2009) 81e87
antitumour activity of the novel pyrrolobenzodiazepine dimer SJG-
136. Eur J Cancer 2005;41:1811e8.
[21] Lin HL, Liu TY, Wu CW, Chi CW. Berberine modulates expression
of mdr1 gene product and the responses of digestive track cancer
cells to paclitaxel. Br J Cancer 1999;81:416e22.
[22] Goldstein LJ. Clinical reversal of drug resistance. Curr Probl Cancer
1995;19:65e124.
[23] Kunzelmann K, Slotki IN, Klein P, Koslowsky T, Ausiello DA,
Greger R, Cabantchik ZI. Effects of P-glycoprotein expression on
cyclic AMP and volume-activated ion fluxes and conductances in
HT-29 colon adenocarcinoma cells. J Cell Physiol 1994;161:
393e406.
[24] Giraldi T, Zorzet S, Perissin L, Rapozzi V. Stress and chemotherapy:
combined effects on tumor progression and immunity in animal
models. Ann N Y Acad Sci 2000;917:549e59.
[25] Walker LG, Walker MB, Ogston K, Heys SD, Ah-See AK, Miller ID,
Hutcheon AW, Sarkar TK, Eremin O. Psychological, clinical and
pathological effects of relaxation training and guided imagery during
primary chemotherapy. Br J Cancer 1999;80:262e8.
[26] Heck DA, Bylund DB. Mechanism of down-regulation of a-2 adren-
ergic receptor subtypes. J Pharmacol Exp Ther 1997;282:1219e27.
[27] Nylund G, Nordgren S, Delbro DS. Demonstration of functional
receptors for noradrenaline and adenosine-5’-triphosphate, but not
for prostaglandin E2, in HT-29 human colon cancer cell line. Auton
Autacoid Pharmacol 2003;23:193e9.
[28] Bylund DB, Blaxall HS, Iversen LJ, Caron MG, Lefkowitz RJ,
Lomasney JW. Pharmacological characteristics of a2-adrenergic
receptors: comparison of pharmacologically defined subtypes with
subtypes identified by molecular cloning. Mol Pharmacol 1992;42:
1e5.
[29] Kribben A, Herget-Rosenthal S, Lange B, Erdbrugger W, Philipp T,
Michel MC. a2-Adrenoceptors in opossum kidney cells couple to
stimulation of mitogen-activated protein kinase independently of ad-
enylyl cyclase inhibition. Naunyn Schmiedebergs Arch Pharmacol
1997;356:225e32.
[30] Ding S, Chamberlain M, McLaren A, Goh L, Duncan I, Wolf CR.
Cross-talk between signalling pathways and the multidrug resistant
protein MDR-1. Br J Cancer 2001;85:1175e84.




















