
HT-29 cells: a new substrate for rotavirus growth
15
Arch Virol (1991) 116:159-173 _Archives Vi rology © Springer-Verlag 199i Printed in Austria HT-29 cells: a new substrate for rotavirus growth F. Superti, A. Tinari, L. Baldassarri, and G. Donelfi Laboratorio di Ultrastrutture, Istituto Superiore di Sanit/t, Roma, Italy Accepted July 16, 1990 Summary. Susceptibilities of a continuous rhesus monkey kidney cell line (MA- 104) and that of a human colon carcinoma cell line (HT-29) to infection by different human and animal rotavirus strains were compared. HT-29 cells appeared to be more sensitive to human rotavirus infection than MA-104 cells, whereas the latter cell line was more susceptible to animal ro- tavirus replication. The greater sensitivity to human rotavirus infection of HT- 29 cells was confirmed by the successful, direct isolation of these viruses from faecal specimens. Human rotavirus infection of HT-29 cells was also followed by transmission electron microscopy. In ultra-thin sections, unenveloped par- ticles of rotaviruses, representing infectious mature virions, were observed in large number. Moreover, many "double-shelled" particles were detected in negative-stained supernatants from infected cultures. Scanning electron mi- croscopy of uninfected HT-29 cells showed that in the presence of Ca + +, required for rotavirus growth, they are able to express some of the features of mature intestinal cells. In view of these results, HT-29 cells appear to be a useful in vitro model for the study of rotavirus infection. Introduction Rotaviruses are an important cause of acute diarrhoea in infants and young children in many areas of the world [13, 14, 15], including countries with high socio-economic levels and good sanitation standards [5, 9, 17, 29, 39]. Although many animal rotaviruses are readily grown in cell cultures, special conditions are required for the cultivation of human strains whose isolation from clinical specimens has, until recent years, proven to be very difficult. In 1981, Sato and co-workers [32] described the successful cultivation of three cytopathic strains of human rotaviruses from faecal specimens using roller cultures of a continuous line of rhesus monkey kidney cells (MA-104) without serial passages in animals. Specimens were pre-treated with trypsin and small amounts of this enzyme were also incorporated into the maintenance medium.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of HT-29 cells: a new substrate for rotavirus growth

Arch Virol (1991) 116:159-173 _Archives
Vi rology © Springer-Verlag 199i Printed in Austria
H T - 2 9 cells: a new substrate for rotavirus growth
F. Superti, A. Tinari, L. Baldassarri, and G. Donelfi
Laboratorio di Ultrastrutture, Istituto Superiore di Sanit/t, Roma, Italy
Accepted July 16, 1990
Summary. Susceptibilities of a continuous rhesus monkey kidney cell line (MA- 104) and that of a human colon carcinoma cell line (HT-29) to infection by different human and animal rotavirus strains were compared.
HT-29 cells appeared to be more sensitive to human rotavirus infection than MA-104 cells, whereas the latter cell line was more susceptible to animal ro- tavirus replication. The greater sensitivity to human rotavirus infection of HT- 29 cells was confirmed by the successful, direct isolation of these viruses from faecal specimens. Human rotavirus infection of HT-29 cells was also followed by transmission electron microscopy. In ultra-thin sections, unenveloped par- ticles of rotaviruses, representing infectious mature virions, were observed in large number. Moreover, many "double-shelled" particles were detected in negative-stained supernatants from infected cultures. Scanning electron mi- croscopy of uninfected HT-29 cells showed that in the presence of Ca + +, required for rotavirus growth, they are able to express some of the features of mature intestinal cells.
In view of these results, HT-29 cells appear to be a useful in vitro model for the study of rotavirus infection.
Introduction
Rotaviruses are an important cause of acute diarrhoea in infants and young children in many areas of the world [13, 14, 15], including countries with high socio-economic levels and good sanitation standards [5, 9, 17, 29, 39].
Although many animal rotaviruses are readily grown in cell cultures, special conditions are required for the cultivation of human strains whose isolation from clinical specimens has, until recent years, proven to be very difficult.
In 1981, Sato and co-workers [32] described the successful cultivation of three cytopathic strains of human rotaviruses from faecal specimens using roller cultures of a continuous line of rhesus monkey kidney cells (MA-104) without serial passages in animals. Specimens were pre-treated with trypsin and small amounts of this enzyme were also incorporated into the maintenance medium.

160 F. Superti et al.
This technique is widely used to cultivate rotavirus strains from diarrhoea1 stool [1, 2, 4, 11, 18, 21, 40, 42], but the isolation rate remains about 50% [2, 42]. In 1984, Ward and co-workers [41] demonstrated that primary cells from monkey kidney supported direct rotavirus growth from faecal specimens much more efficiently (14/14) than did continuous lines of rhesus (MA-104) or African green (CV-1) monkey kidney cells.
Although rotavirus isolation is an unsuitable diagnostic tool, time-consum- ing and costly, the ability to grow wild strains in cell cultures remains a critical step toward the identification of new variant viruses. A more complete antigenic characterization will be possible when a large number of isolates are cultivated in tissue culture.
In this paper, we analyze the susceptibilities to infection by human and animal rotavirus strains and the sensitivity for direct viral isolation of a human colon carcinoma cell line (HT-29) compared with a continuous cell line of rhesus monkey kidney (MA-104). The morphological characteristics of HT-29 cells and the viral growth were also studied by scanning and transmission electron microscopy.
Materials and methods
Ce//s
MA-104 cells (a continuous line of foetal rhesus monkey kidney cells) were grown at 37 °C in Eagle's 199 medium containing 1.2 g/1 NaHCO3 and supplemented with 10% foetal calf serum (FCS), penicillin (100 IU/ml) and streptomycin (100 gg/ml).
HT-29 cells (a human colon adenocarcinoma celt line) were obtained fi'om American Type Culture Collection (ATCC; Rockville, MA) and maintained in RPMI 1640 medium for indefined number of passages. Cells were grown at 37°C in RPMI 1640 medium containing t.2g/1 NaHCO3 and supplemented with 10% FCS, glutamine (2raM), non- essential aminoacids, penicillin (100 IU/ml), and streptomycin (100 gg/ml).
Human and animal rotaviruses
The following human rotavirus strains were used: WA (serotype 1) and KUN (serotype 2) kindly provided by T. Konno (Sendai, Japan); DS-I (serotype 2), P (serotype 3), and St. Thomas 4 (serotype 4) obtained from A. Z. Kapikian (Bethesda, U.S.A.), and VA-70 (serotype 4) from G. Gerna (Pavia, Italy).
UK bovine rotavirus strain (serotype 6) and SA-11 simian rotavirus strain (serotype 3) were kindly provided by G. Castrucci (Perugia, Italy) and by F. Prevost (Paris, France), respectively.
Faecal specimens
Twenty stool specimens positive for rotavirus by both electron microscopy and enzyme- linked immunosorbent assay were selected for culture. They were from sporadic cases of infantile diarrhoea and showed, after polyacrylamide gel electrophoresis of the viral genome, different "long" RNA profiles.
All samples, collected from children hospitalized between 1986 and 1987, were kindly provided by R. Bisicchia of"Ospedale dei Bambini", Brescia (hospital A) and U. Scardellato of "Ospedale Multizonale", Treviso (hospital B). Stools, assessed by electron microscopy

Rotavirus infection in HT-29 cells 161
to contain approximately 108 to l01° intact double-shelled rotavirus particles per ml, were selected from 13 children in hospital A and from 7 in hospital B. Specimens were stored either as whole faeces or as 20% faecal homogenates in phosphate-buffered saline (PBS, pH 7.4) at - 70 °C for different periods.
At the time of the study, the specimens were thawed, vortexed and centrifuged at 3,000 x g for 10 min. Bacteria were removed from supernatants by passing through 0.22 Millipore membrane filters (Millipore Corp., Bedford, MA).
Viral cultivation
Passage of different rotavirus strains
Human and animal rotavirus strains, adapted to growth in MA-104 cells, were treated with 20 gg/ml trypsin (Sigma Chemical Company; type IX) for 30min at 37 °C.
100 gl of each virus sample were inoculated into six tissue culture tubes (16 x 125 ram, Falcon) containing either MA-104 or HT-29 cells (3 tubes per cell type), grown for 24h at 37 °C.
After 90 min adsorption at 37 °C, the inocula were removed from the culture tubes, the cells were washed three times with fresh medium and fed with 0.5 ml Eagle's 199 medium containing 10% tryptose phosphate broth (TPB) (Difco) and 1 ~tg/ml trypsin for MA-104 cells; or with 0.5 ml RPMI 1640 medium containing 10% TPB, 1 mM CaC12, and 1 gg/ml trypsin for HT-29 cells. The tubes were incubated at 37 °C on a roller apparatus for several days.
At different times (24, 48, 72 h) infection was stopped in two tubes of each cell line: cells from one tube were harvested for indirect immunofluorescence staining (IIF) and the content of the other tube was frozen. After three cycles of freezing and thawing, the supernatant from the centrifuged infected culture was harvested and designated as the first cell culture passage virus.
Subsequent passages were carried out in the same manner.
Direct isolation of human rotaviruses
For viral isolation, 0.1 ml of each rotavirus-containing, 10% stool suspension was filtered by a 0.22 g Millipore membrane and treated with 20 gg/ml trypsin. Cell inoculation and subsequent passages were carried out as described above.
Antiserum
Hyperimmune antiserum to inner rotavirus shell was produced in seronegative rabbits by using UK bovine rotavirus strain and the immunization schedule previously described by Bastardo and co-workers [3].
ImmunoJTuorescence
At 24, 48, and 72 h post-infection and at each passage, infected cells were harvested, washed, spotted onto glass slides, dried and then fixed in acetone for 10 rain at - 20 °C.
The slides were incubated with a hyperimmune rabbit anti-UK strain serum for 45 rain at 37 °C and stained with an anti-rabbit immunglobulin-fluorescein conjugate (Kallestad). After 45 min at 37 °C, cells were washed, mounted in glycerol and examined using a Dialux Leitz fluorescence microscope.
Transmission electron microscopy
HT-29 cells, grown in 4 tissue culture tubes, were infected with VA-70 strain at a m.o.i. of 20 p.f.u./cell for 90 rain at 37 °C as described above. After 24 h, two infected tubes were

162 F. Superti et al.
processed for ultra-thin section electron microscopy as previously described [38] and the others were further incubated for 24 h and negatively stained with 2% (w/v) sodium phos- phomngstate (pH 7.0). Samples were examined using a Philips 430 electron microscope at a magnification of x 40,000.
Scanning electron microscopy
HT-29 cells were grown on glass coverslips in RPMI 1640 medium containing 10% FCS alone, 10% FCS plus 1 mM Ca + +, or 10% TPB plus 1 mM Ca + +. After 48 h at 37°C in 5% CO2, monolayers were processed for scanning electron microscopy as previously described [12]. Observations were performed with a Philips 515 scanning electron micro- scope.
Results
Cultivation of cell culture adapted rotavirus strains
The h u m a n ro tavi rus strains WA, K U N , DS-1, P, St. T h o m a s 4, and VA-70
and the animal ro tavi rus strains SA-11 and U K were utilized. All strains had
previously been adap ted to g rowth on MA-104 cells.
Table 1 shows the percent o f f luorescent cells ob ta ined at different times
Table 1. Growth of different rotavirus strains in MA-104 and HT-29 cells
Strain Serotype Cell line Immunofluorescence (%)
24 h 48 h 72 h
WA a 1 MA- 104 - 10 HT-29 - 40
KUN a 2 MA-104 10 l0 HT-29 80 90
DS-1 a 2 MA-104 10 20 HT-29 60 100
p a 3 MA-104 10 10 HT-29 80 100
St. Thomas 4 a 4 MA-104 10 10 HT-29 20 90
VA-70 ~ 4 MA- 104 30 60 HT-29 80 100
SA-11 b 3 MA-104 50 70 HT-29 50 4O
UK b 6 MA-104 100 CPE 100% ° HT-29 5 15
20 60
60 90
50 100
20 100
20 90
70 80
CPE 100% ° CPE 100% °
CPE 100% ° 15
a Human rotavirus strain b Animal rotavirus strain
Cytopathic effect in 100% of cells

Rotavirus infection in HT-29 cells
Table 2. Direct isolation of rotaviruses from 13 children with acute diarrhoea (Hospital A)
163
Specimen no. (year of collec- tion)
Cell line Immunofluorescence (%)*
Original Passage no. sample
1 2 3 4 5
1 (1986) MA-104 40 10 i0 10 20 10 HT-29 50 40 30 30 30 30
2 (1986) MA-104 . . . . . . HT-29 100 40 20 30 30 40
3 (1986) MA-104 . . . . . . HT-29 50 30 30 20 30 30
4 (1986) MA-104 . . . . . . HT-29 50 20 30 30 30 40
5 (1986) MA-104 . . . . . . HT-29 80 30 30 40 40 40
6 (1986) MA-104 . . . . . . HT-29 50 30 20 15 15 20
7 (1987) MA-104 . . . . . . HT-29 20 30 20 15 15 20
8 (1987) MA-104 10 10 . . . . HT-29 10 10 10 I0 15 30
9 (1987) MA-104 10 10 . . . . HT-29 20 l0 10 10 10 20
10 (1987) MA-104 10 10 10 10 10 10 HT-29 20 10 10 10 10 10
11 (1987) MA-104 10 10 10 10 10 10 HT-29 20 10 20 20 20 20
12 (1987) MA-104 10 10 . . . . HT-29 20 10 10 10 10 20
13 (1987) MA-104 10 10 . . . . HT-29 30 10 10 10 10 10
* Immunofluorescence staining of infected cells was carried out at 48 h p.i.
after infection in bo th cell lines. The highest levels o f h u m a n rotavirus ant igen synthesis were obta ined in HT-29 cells. In this line, the most substant ial increase o f f luorescent cells was generally observed 48 h p.i., while in MA-104 cells similar
behaviour was observed at 72 h p.i. Viral yield was assessed by serial passages o f superna tants f rom infected cells which revealed a product ive infection, as

164 F. Superti et al.
mon i to red by indirect immunof luorescence (data not shown). With regard to simian rotavirus (SA-11), the two cell lines tested were comparab le in their sensitivity to the infection. W h e n M A - I 0 4 and HT-29 cells were infected with bovine rotavirus (UK), only a poor viral ant igen synthesis (15% of f luorescent cells) was achieved in HT-29 cells, while MA-104 monolayers were heavily infected and 48 h p.i. cytopathic effect was observed in 100% o f cells.
Direct isolation of rotaviruses
Since HT-29 cells appeared to be more susceptible than MA- 104 cells to infection by h u m a n rotavirus strains, we investigated the suitability o f this cell line for direct isolation o f h u m a n rotaviruses f rom faecal specimens.
Table 3. Direct isolation of rotaviruses from 13 children with acute diarrhoea (Hospital B)
Specimen no. (year of collec- tion)
Cell line Immunofluorescence (%)*
Original Passage no. sample
1 2 3 4 5
14 (1986) MA-104 10 20 10 10 10 10 HT-29 20 10 20 10 20 30
15 (1986) MA-104 . . . . . . HT-29 50 10 10 20 20 20
16 (1986) MA-104 . . . . . . HT-29 30 10 10 10 10 20
17 (1986) MA-104 . . . . . . HT-29 10 10 30 20 10 10
18 (1986) MA-104 . . . . . . HT-29 40 10 20 10 10 20
19 (1986) MA-104 10 10 . . . . HT-29 30 10 10 10 10 10
20 (1986) MA-104 . . . . . . HT-29 20 10 30 10 10 20
* Immunofluorescence staining of infected cells was carried out at 48 h p.i.
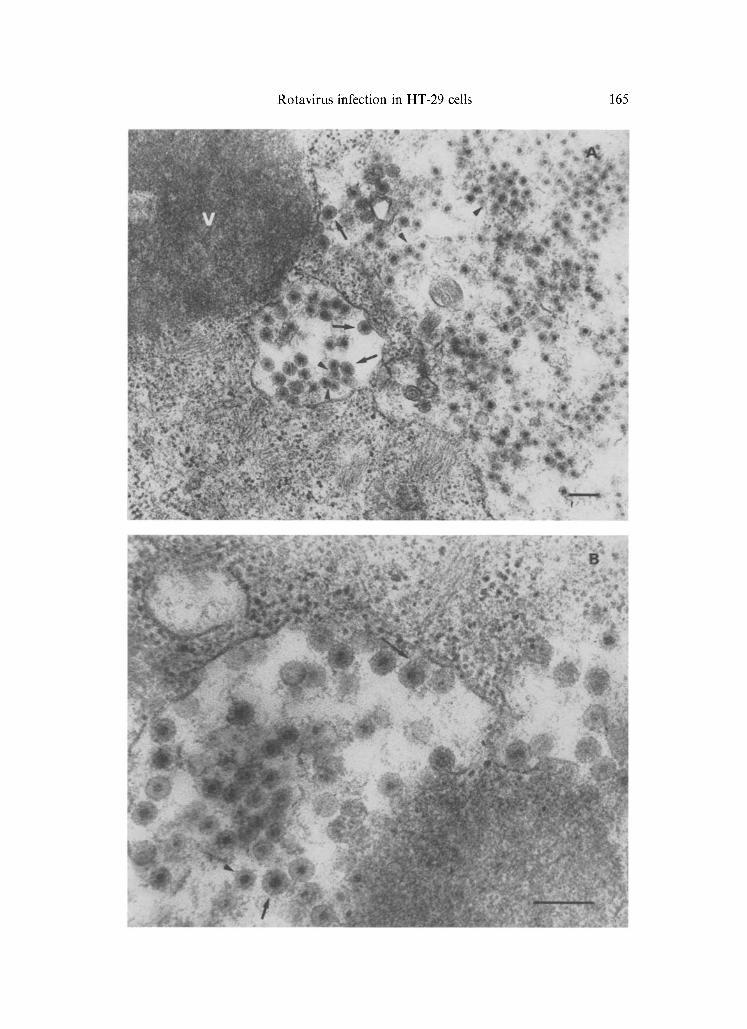
Fig. 1. Thin sectioned rotavirus-infected HT-29 cells. A Viroplasm (V) associated with RER cisternae which contain primarily 65-70 nm virus particles (Chasey's type III) (see arrow heads). A few 80-90 nm particles (Chasey's type II) (see short arrows) are present [7]. B Rotavirus Chasey's type II (short arrow) and III (arrowhead) within dilated cisternae of rough endoplasmic reticulum. Some virus particles appear to be budding through the RER
membrane (long arrow). Bars: 0.2 btm
Rotavirus infection in HT-29 cells 165
166 F. Superti et al.
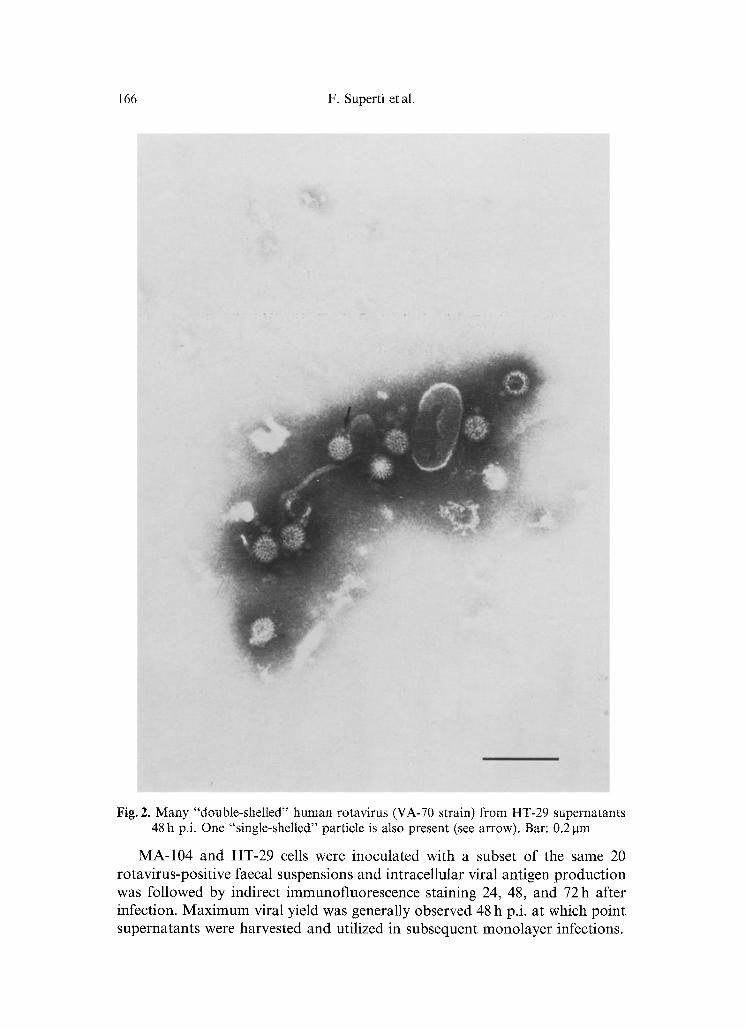
Fig. 2. Many "double-shelled" human rotavirus (VA-70 strain) from HT-29 supernatants 48 h p.i. One "single-shelled" particle is also present (see arrow). Bar: 0.2 p,m
MA-104 and HT-29 cells were inoculated with a subset of the same 20 rotavirus-positive faecal suspensions and intracellular viral antigen production was followed by indirect immunofluorescence staining 24, 48, and 72 h after infection. Maximum viral yield was generally observed 48 h p.i. at which point supernatants were harvested and utilized in subsequent monolayer infections.
Rotavirus infection in HT-29 cells 167
For each specimen, five serial passages were carried out; the percentage of infected cells, monitored by immunofluorescence 48 h p.i., is shown in Tables 2 and 3. In MA-104 cells 9 out of 20 strains were isolated on the first passage and four of them underwent successful subsequent passages.
In HT-29 cells all strains tested grew on first and subsequent passages, even though the percentages of infected cells remained rather low.
Transmission electron microscopy
To obtain further information about the ability of HT-29 cells to permit human rotavirus replication, infection with the VA-70 strain was followed by electron microscopy.
Cells contained electron-dense viroplasm and a large amount of viral particles which were associated with rough endoplasmic reticulum (RER) cisternae (Fig. 1 A), or were budding through the RER membrane (Fig. 1 B). Both en- veloped and unenveloped particles, measuring 80-90 nm and 65-7'0 nm in di- ameter, respectively, were detected.
Moreover, many double-shelled particles were observed in negatively-stained supernatants (Fig. 2).
Scanning electron microscopy of HT-29 cells
HT-29 cells were examined using scanning electron microscopy to evaluate the potential relationship between their ability to allow human rotavirus growth
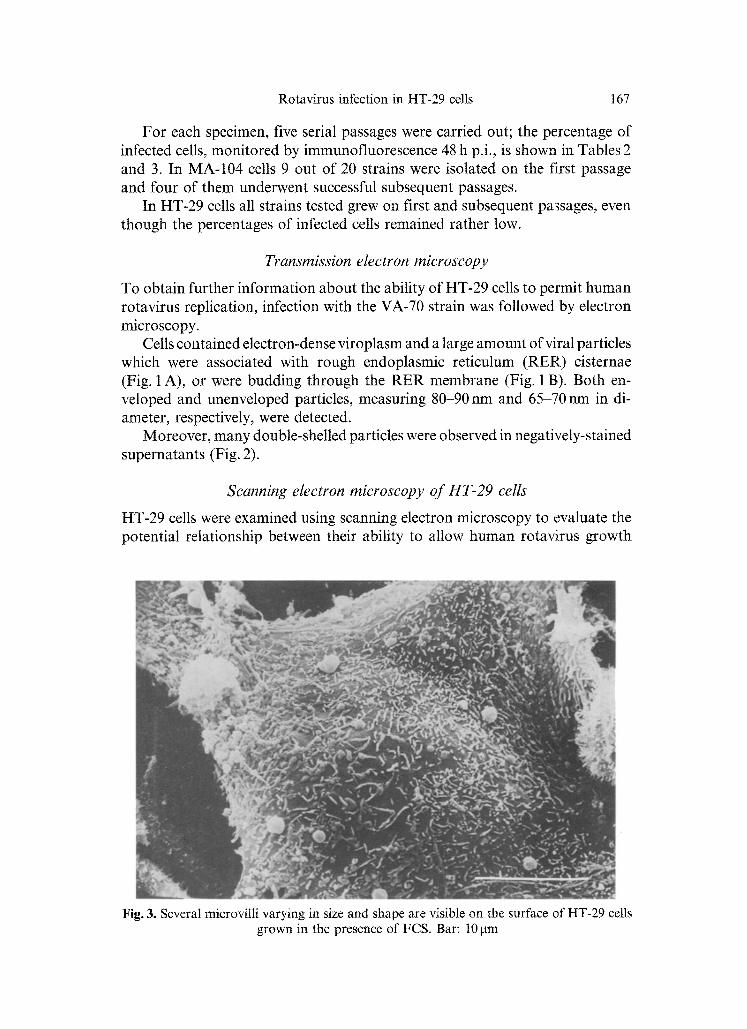
Fig. 3. Several microvilti varying in size and shape are visible on the surface of HT-29 cells grown in the presence of FCS. Bar: 10 gm

168 F. Superti et al.
and a different iat ion of cells s t imulated by culture condit ions used for viral infection.
HT-29 cells g rown for 48 h in m e d i u m containing 10% FCS showed several microvilli, varying in size and shape, r andomly distr ibuted over the surface m e m b r a n e (Fig. 3). W h e n cells were grown either with FCS plus Ca + + (Fig. 4 A)
Fig. 4. HT-29 cells grown in the presence of either FCS + Ca + + (A) or TPB + Ca + + (B) show a local organization of microvilli (insert) resembling the brush border of the intestinal mucosa. Mucus-like material is also visible (arrows). Bars: A and B, 10 gin; insert, 100 rim)

Rotavirus infection in HT-29 cells 169
or TPB plus Ca + + (Fig. 4 B), they appeared enlarged or fused to one another, with short straight microvilli locally organized in such a way as to resemble the brush border of the intestinal mucosa. Amorphous, fuzzy, mucus-like ma- terial was also present.
Discussion
In this study we analyzed the susceptibility to rotavirus infection of a human colon carcinoma cell line (HT-29). These cells appear to be more sensitive to human rotavirus replication as compared with MA-104, a monkey kidney cell line known to be a substrate of choice for rotavirus growth [40].
When the infection was performed with animal rotaviruses, different be- haviour was observed. HT-29 and MA-104 cells were comparable in their sensitivity to simian rotavirus infection, whereas only the latter were heavily infected by bovine rotavirus. It is interesting to point out that the human cell line preferentially supports human rotavirus infection while MA-104 cells, of animal origin, seem to be more permissive to animal rotaviruses.
To further investigate the sensitivity of HT-29 cells to human rotavirus infection, we tested faecal extracts obtained from 20 children hospitalized with acute diarrhoea for direct isolation.
In HT-29 cells all the screened specimens grew on first and subsequent passages, whereas 9 out of 20 strains were isolated in MA-104 cells and only four of them successfully underwent subsequent passages. It was previously reported that MA-104 cells supported human rotavirus growth directly from faecal specimens less efficiently than did primary cells from monkey kidney even though passage of virus in the latter appeared to adapt it for consistent growth in continuous cell lines [41].
The results presented here clearly indicate the human colon carcinoma cell line to be a better medium than MA-104 cells for human rotavirus isolation. Our results are in agreement with a previous study of Patel et al. [22] dem- onstrating that two gut tumour-derived cell lines, HT-29 and SKCO-1, have a markedly wider susceptibility with comparable or wider sensitivity for entero- viruses than primary monkey kidney cells (PMK) and rhabdomyosarcoma cells (RD).
In addition to its suitability as a substrate for enterovirus and human ro- tavirus growth, the ease in handling of the HT-29 cell line indicates it as a very useful tool for the study of these infectious agents.
For a better understanding of the rotavirus/HT-29 cell interaction, and for a morphological characterization of this cell line under our experimental con- ditions, electron microscopy studies were performed. Enveloped and unenvel- oped particles measuring 8090 nm and 65-70 nm in diameter, respectively, and electron-dense viroplasms were detected in ultra-thin sections of infected cells.
The viroplasm is probably the site of synthesis of the "single-shelled" particles that contain RNA [25]. Viruses assemble near the periphery of these inclusions

170 F. Superti et al.
and subsequently bud into the rough endoplasmic reticutum cisternae acquiring membrane. The two distinct viral forms, observed within dilated cisternae of the RER, represent different steps in the rotavirus replication cycle. McNulty etal. [20] first suggested, in bovine rotavirus infected MBDK cells, that the enveloped and unenveloped forms correspond to the negatively-stained "dou- ble" and "single"-shelled particles, respectively.
An alternative hypothesis was proposed by Chasey [7]: unenveloped, Chas- ey's type III, particles represent the "double" capsid structures seen in negative- stain, while enveloped, Chasey's type II, particles are transient forms acquiring membrane by budding through the RER. These suggestions were subsequently confirmed by other authors [10, 23, 24]. The loss or transformation of the outer rim would thus be an integral process in the development of infective particles.
Infectivity has been shown to be associated with the "double" capsid particles [6] and the Chasey's type III forms, seen in large number in our system, could represent the final stage in virus maturation. In fact, a large number of "double- shelled" particles were observed in negatively-stained infected HT-29 cell su- pernatants.
Furthermore, we have never observed the Chasey's type IV particles which represent the smaller (54-65nm in diameter), non-infective, "single" capsid forms. These ultrastructural observations fit well with the results on the pro- ductive infection of HT-29 cells. Supernatants from this cell line were more infective than those from MA-104 cells at the same time (data not shown).
Since the human colon adenocarcinoma cell line (HT-29) is able to differ- entiate, similarly to the parental cells, when the medium carbon sources are changed [26, 43], we analyzed cell morphological alterations under our nutri- tional conditions, i.e., cell growth or viral growth.
Scanning electron microscopy showed that cells grown in standard culture medium, supplemented with FCS, exhibited an undifferentiated morphology. On the contrary, enteric features where observed when calcium, required for "double-shell" and infectivity stabilization [36], was added to the medium containing either FCS or TPB. It is well known that rotavirus maturation is a calcium-dependent process: in absence of calcium, virus yield is decreased and viral particles are found to be exclusively "single-shelled" [33, 34].
For viral cultivation in HT-29 cells, 1 mM CaC12 was added to the medium since we observed an increase of viral yield under these conditions (data not shown).
The role of Ca + + in the early stages of rotavirus infection has been exten- sively studied. The use of calcium ionophore A 23187 to increase the intracellular Ca + + concentration was found to block viral uncoating, suggesting that low Ca + + concentrations in the intracellular microenvironment may be responsible for this process [19].
Calcium seems to be the critical factor stimulating cellular differentiation: a relationship between the observed organization of microvilli and a regulatory

Rotavirus infection in HT-29 cells 171
activity for Ca + + could be speculated. In particular, calcium may act on villin, a Ca + +-regulated actin-binding protein known to be specifically associated with the cytoskeleton of the brush border microvilli [31], which has been demonst ra ted in HT-29 cells [30].
It is no tewor thy that, in spite of their colon origin, differentiated HT-29 cells mimic the situation found in the small intestine [31], which is the site o f rotavirus replication [8, 16, 35, 37]. As the few cell lines derived f rom the small intestinal mucosa did not exhibit the morphological features of differentiated enterocytes [27, 28], HT-29 cells may represent a valuable in vitro tool for a better unders tanding of h u m a n rotavirus cell interactions.
References
1. Albert MJ, Bishop RF (1984) Cultivation of human rotaviruses in cell culture. J Med Virol 13:377-383
2. Albert M J, Unicomb LE, Barnes GL, Bishop RF (1987) Cultivation and characteri- zation of rotavirus strains infecting newborn babies in Melbourne, Australia, from 1975 to 1979. J Clin Microbiol 25:1635-1640
3. Bastardo JW, McKimm-Breschkin JL, Sonza S, Mercer LD, Holmes IH (1981) Prep- aration and characterization of antisera to electrophoretically purified SA-11 virus polypeptides. Infect Immun 34:641-647
4. Birch CJ, Rodger SM, Marshall JA, Gust ID (1983) Replication of human rotavirus in cell culture. J Med Virol 11:241-250
5. Brandt CD, Kim HW, Rodriguez WJ, Arrobio JO, Jeffries BC, Stallings EP, Lewis C, Miles A J, Chanock RM, Kapikian AZ, Parrott RH (1983) Pediatric viral gastroen- teritis during eight years of study. J Clin Microbiol 18:7t-78
6. Bridger JC, Woode GN (1976) Characterization of two particle types of calf rotavirus. J Gen Virol 31:245-250
7. Chasey D (1977) Different particle types in tissue culture and intestinal epithelium infected with rotavirus. J <3en Virol 37:443-451
8. Davidson <3P, Gall DG, Petric M, Butler DG, Hamilton JR (1977) Human rotavirus enteritis induced in conventional piglets: intestinal structure and transport. J Clin Invest 60:1402-1409
9. Donelti G, Ruggeri FM, Tinari A, Marziano ML, Menichella D, Caione D, Concato C, Rocchi (3, Vella S (1988) A three-year diagnostic and epidemiological study on viral infantile diarrhoea in Rome. Epidemiol Infect 100:311-320
10. Estes MK (1990) Rotaviruses and their replication. In: Fields BN (ed) Virology, vol 2, 2nd edn. Raven, New York, pp 1329-1352
11. Hasegawa A, Matsuno S, Inouye S, Kono R, Tsurukubo Y, Mukoyama A, Saito Y (1982) Isolation of human rotaviruses in primary cultures of monkey kidney cells. J Clin Microbiol I6:387-390
12. Huet C, Sahuquillo-Merino C, Coudrier E, Louvard D (1987) Absorptive and mucus- secreting subclones isolated from a multipotent intestinal cell line (HT-29) provide new models for cell polarity and terminal differentiation. J Cell Biol 105:345-357
13. Kapikian AZ, Wyatt RG, <3reenberg HB, Kalica AR, Kim HW, Brandt CD, Rodriguez WJ, Parrott RH, Chanock RM (1980) Approaches to immunization of infants and young children against gastroenteritis due to rotaviruses. Rev Infect Dis 2:459-469
14. Kapikian AZ, Greenberg HB, Wyatt R<3, Kalica AR, Kim HW, Brandt CD, Rodriguez WJ, Parrott RH, Chanock RM (1982) Viral gastroenteritis. In: Evans AS (ed) Viral infections of humans, 2nd edn. Plenum, New York, pp 283-236

172 F. Superti et at.
15. Kapikian AZ, Chanock RM (1990) Rotaviruses. In: Fields BN (ed) Virology, vo12, 2nd edn. Raven, New York, pp 1353-1404
16. Kerzner B, Kelly MH, Gall DG, Butler DG, Hamilton JR (t977) Transmissible gas- troenteritis: sodium transport in the intestinal epithelium during the course of viral enteritis. Gastroenterology 72:457-461
17. Konno T, Suzuki H, Imai A, Kutsuzawa T, Ishida N, Katsushima N, Sakamoto M, Kitaoka S, Tsuboi R, Adachi M (1978) A long-term survey of rotavirus infection in Japanese children with acute gastroenteritis. J Infect Dis 138:569-576
18. Kutsuzawa T, Konno T, Suzuki H, Kapikian AZ, Ebina T, Ishida N (t982) Isolation of human rotavirus subgroups 1 and 2 in cell cultures. J Clin Microbiol 16:727-730
19. Ludert JE, Michelangeli F, Gil F, Liprandi F, Esparza J (1987) Penetration and uncoating of rotaviruses in cultured cells. Intervirology 27:95-101
20. McNulty MS (1976) The rnorphogenesis of a cytopathic bovine rotavirus in Madin- Darby bovine kidney cells. J Gen Virol 33:503-508
21. Naguib T, Wyatt RG, Mohieldin MS, Zaki AM, Imam IZ, DuPont HL (1984) Cul- tivation and subgroup determination of human rotaviruses from Egyptian infants and young children. J Clin Microbiol 19:210-212
22. Patel JR, Daniel J, Mathan VI (1985) A comparison of the susceptibility of three human gut tumour-derived differentiated epithelial cell lines, primary monkey kidney cells and human rhabdomyosarcoma cell line to 66 prototype strains of human enterovirus. J Virol Methods 12:209-216
23. Petrie BL, Graham DY, Estes MK (1981) Identification of rotavirus particle types. Intervirology 16:20-28
24. Petrie BL, Graham DY, Haussen H, Estes MK (1982) Localization ofrotavirus antigens in infected cells by ultrastructural immunocytochemistry. J Gen Virol 63:457-467
25. Petrie BL, Greenberg HB, Graham DY, Estes MK (1984) Ultrastructural localization of rotavirus antigens using colloidal gold. Virus Res 1:133-152
26. Pinto M, Appay MD, Simon-Assmann P, Chevalier G, Dracopoli N, Fogh J, Zweibaum A (1982) Enterocytic differentiation of cultured human colon cancer cells by replace- ment of glucose by galactose in the medium. Biol Cell 44:193-196
27. Quaroni A, May RJ (1980) Establishment and characterization of intestinal epithelial cell cultures. In: Harris GC, Trump BF, Stoner GD (eds) Methods in cell biology, vo121 B. Academic Press, New York, pp 403-427
28. Quaroni A (1985) Development of fetal rat intestine in organ and monolayer culture. J Cell Biol 100:1601-1610
29. Riepenhoff-Talty M, Saif L J, Barrett H J, Suzuki H, Ogra PL (1983) Potential spectrum of etiological agents of viral gastroenteritis in hospitalized infants. J Clin Microbiol 17:352-356
30. Robine S, Huet C, Moll R, Sahuquillo-Merino C, Coudrier E, Zweibaum A, Louvard D (1985) Can villin be used to identify malignant and undifferentiated normal digestive epithelial cells? Proc Natl Acad Sci USA 82:8488 8492
31. Rousset M (1986) The human colon carcinoma cell lines HT-29 and Caco-2: two in vitro models for the study of intestinal differentiation. Biochimie 68:1035-1040
32. Sato K, Inaba Y, Shinozaki T, Fujii R, Matumotu M (1981) Isolation of human rotavirus in cell cultures. Arch Virol 69:155--t60
33. Shahrabadi MS, Lee PWK (1986) Bovine rotavirus maturation is a calcium-dependent process. Virology 152:298-307
34. Shahrabadi MS, Babink LA, Lee PWK (1986) Further analysis of the role of calcium in rotavirus morphogenesis. Virology 158:103-111
35. Shepherd RW, Gall DG, Butler DG, Hamilton JR (1979) Determinants of diarrhea in viral enteritis: the role of ion transport and epithelial changes in ileum in trasmissible gastroenteritis in piglets. Gastroenterology 76:20-24

Rotavirus infection in HT-29 cells 173
36. Shirley JA, Beards GM, Thouless ME, Flewett TH (1981) The influence of divalent cations on the stability of human rotavirus. Arch Virol 67:1-9
37. Snodgrass DR, Angus KW, Gray EW (1977) Rotavirus infection in lambs: pathogenesis and pathology. Arch Virol 55:263-274
38. Tinari A, Ruggeri FM, Divizia M, Panfi A, Donelli G (1989) Morphological changes in HAV-infected Frp/3 cells and immunolocalization of HAAg. Arch Viro1104: 209-224
39. Tufvesson B, Johnsson T (1976) Occurrence of reo-like viruses in young children with acute gastroenteritis. Acta Pathol Microbiol Scand 84:22-28
40. Urasawa T, Urasawa S, Taniguchi K (1981) Sequential passages of human rotavirus in MA-104 cells. Microbiol Immunol 25:1025-1035
41. Ward RL, Knowlton DR, Pierce MJ (1984) Efficiency of human rotavirus propagation in ceil culture. J Clin Microbiol 19:748-753
42. Wyatt RG, James HD jr, Pittman AL, Hoshino Y, Greenberg HB, Kalica AR, Flores J, Kapikian AZ (1983) Direct isolation in cell culture of human rotaviruses and their characterization into four serotypes. J Clin Microbiol 18:310-317
43. Zweibaum A, Pinto M, Chevalier G, Dussaulx E, Triadou N, Sacroix B, Haffen K, Brun JL, Rousset M (1985) Enterocytic differentiation of a subpoputation of the human colon turnout cell line HT-29 selected for growth in sugar-free medium and its inhibition by glucose. J Cell Physiol 122:21-29
Authors' address: Dr. F. Superti, Laboratorio di Ultrastrutture, Istituto Superiore di SanitY, Viale Regina Elena 299, 1-00161 Rome, Italy.
Received May 16, 1990




















