
The rotavirus genus
16
~ Pergamon Comp. Immun. Microbiol. infect. Dis. Vol. 17, No. 3/4, pp. 305 320, 1994 Copyright© 1994Elsevier Science Ltd 0147-9571(94)E0010-Q Printed in Great Britain.All rights reserved 0147-9571/94 $7.00+ 0.00 THE ROTAVIRUS GENUS GIANFRANCO DONELLI a n d FABIANA SUPERTI Laboratorio di Ultrastrutture, Istituto Superiore di SanitY, Viale Regina Elena, 299-00161, Roma, Italy Abstract--Human rotaviruses, discovered nearly 20 years ago, have been proven to be major cause of paediatric diarrhoeal disease morbidity and mortality. The clinical significance of these viruses stimulated basic studies on their biology, molecular and antigenic properties and epidemiology. General features, clinical relevance, epidemiologic pattern and laboratory diagnosis of human rotavirus infections are here reviewed. Key words: Rotavirus, diarrhoea. R~sum~--Les rotaviruses humains, d6couverts depuis vingt ans, sont connus comme la principale cause de morbidit6 et de morbilit6 des maladies diarrheiques en p~diatrie. L'importance clinique de ces viruses a stimul~ des &udes de base sur leur biologie, leurs propriet~s mol+culaires et antig6niques et sur l'6pid~miologie. Dans cette revue sont present6es les caract6ristiques g6nb.rales, l'importance clinique, l'6pid~miologie et le diagnostic de laboratoire des infections par les rotaviruses. Mots-clefs: Rotavirus, diarrh6e. INTRODUCTION Rotaviruses are a genus belonging to the family Reoviridae. First identified by electron microscopy in 1973 [1], human rotaviruses are the main aetiologic agents of gastroenteritis in infants and young children worldwide and the major cause of infant mortality in undeveloped areas. STRUCTURE AND PHYSICOCHEMICAL PROPERTIES Rotaviruses, like other members of the Reoviridae family, have a double-shelled capsid composed of an outer layer, an inner capsid, and an icosahedral core (Table 1). Double- and single-shelled particles and core ghosts can be observed by electron microscopy (Fig. 1). The double-shelled particles show a distinctive morphology suggestive of a wheel. The virus name is derived from the Latin rota, meaning wheel. Intact particles, about 70 nm in dia, possess a core forming a hub, capsomeres radiating from the core as short spokes, and a precisely marginated outer layer forming the rim [2]. The single-shelled particles, measuring about 55 nm in dia, are often described as rough particles because they lack the smooth outer layer and their periphery shows projecting trimeric subunits of the inner capsid. Cores, about 37 nm in dia, are less frequently observed as ghost particles in that they usually lack genomic RNA [3]. The different particles have distinct biophysical and biological properties. Double- CIMID 17-3/4~V 305
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of The rotavirus genus

~ Pergamon Comp. Immun. Microbiol. infect. Dis. Vol. 17, No. 3/4, pp. 305 320, 1994 Copyright © 1994 Elsevier Science Ltd
0147-9571(94)E0010-Q Printed in Great Britain. All rights reserved 0147-9571/94 $7.00 + 0.00
THE ROTAVIRUS GENUS
GIANFRANCO DONELLI and FABIANA SUPERTI Laboratorio di Ultrastrutture, Istituto Superiore di SanitY, Viale Regina Elena, 299-00161, Roma,
Italy
Abstract--Human rotaviruses, discovered nearly 20 years ago, have been proven to be major cause of paediatric diarrhoeal disease morbidity and mortality. The clinical significance of these viruses stimulated basic studies on their biology, molecular and antigenic properties and epidemiology. General features, clinical relevance, epidemiologic pattern and laboratory diagnosis of human rotavirus infections are here reviewed.
Key words: Rotavirus, diarrhoea.
R~sum~--Les rotaviruses humains, d6couverts depuis vingt ans, sont connus comme la principale cause de morbidit6 et de morbilit6 des maladies diarrheiques en p~diatrie. L'importance clinique de ces viruses a stimul~ des &udes de base sur leur biologie, leurs propriet~s mol+culaires et antig6niques et sur l'6pid~miologie. Dans cette revue sont present6es les caract6ristiques g6nb.rales, l'importance clinique, l'6pid~miologie et le diagnostic de laboratoire des infections par les rotaviruses.
Mots-clefs: Rotavirus, diarrh6e.
INTRODUCTION
Rotav i ruses are a genus be longing to the family Reov i r idae . Firs t identif ied by electron mic roscopy in 1973 [1], h u m a n rotavi ruses are the ma in aet iologic agents o f gas t roenter i t i s in infants and young chi ldren wor ldwide and the ma jo r cause o f infant mor ta l i t y in undeve loped areas.
STRUCTURE AND PHYSICOCHEMICAL PROPERTIES
Rotavi ruses , like o ther members o f the R e o v i r i d a e family, have a double-shel led capsid c o m p o s e d o f an ou te r layer, an inner capsid, and an icosahedra l core (Table 1). Double - and single-shelled par t ic les and core ghosts can be observed by electron mic roscopy (Fig. 1). The double-she l led par t ic les show a dist inct ive m o r p h o l o g y suggestive o f a wheel. The virus name is der ived f rom the Lat in rota, meaning wheel. In tac t part icles, abou t 70 nm in dia, possess a core fo rming a hub, capsomeres rad ia t ing f rom the core as shor t spokes , and a precisely marg ina t ed outer layer forming the r im [2]. The single-shelled part icles , measur ing a b o u t 55 nm in dia, are often descr ibed as rough part icles because they lack the s m o o t h outer layer and their pe r iphery shows pro jec t ing tr imeric subuni ts o f the inner capsid. Cores , a b o u t 37 nm in dia, are less f requent ly observed as ghost part icles in that they usual ly lack genomic R N A [3].
The different par t ic les have dis t inct b iophys ica l and biological proper t ies . Doub le -
CIMID 17-3/4~V 3 0 5

306 GIANFRANCO DONELLI and FABIANA SUPERTI
shelled particles have a density of 1.36 g/cm 3 in CsC1 and a sedimentation coefficient of 520-530S; single-shelled particles have a density of 1.38 g/cm 3 and a sedimentation coefficient of 380-400S; cores have a density of 1.44 g/cm 3 and a sedimentation coefficient of 280S [4-6]. Only double-shelled particles are infectious [4] and treatment with calcium-chelating agents (e.g. EDTA or EGTA), causing the removal of the outer capsid, results in a loss of viral infectivity [7]. Cores can be obtained by caotropic agents, such as sodium thiocyanate [8] or highly concentrated calcium chloride, which degrade the inner capsid of the particles [6].
Rotavirus infectivity is not affected by fluorocarbon extraction or exposure to ether and deoxycholate [7], reflecting the absence of a lipid envelope. Moreover, viral particles are relatively resistant to repeated freeze-thawing, sonication or incubation at 37°C for I h or at room temperature (25°C) for 24h [7,9]. Chloroform treatment reduces infectivity slightly and destroys hemagglutinating activity [7]. Rotavirus infectivity is relatively stable within the pH range 3-9 [7, 10].
Treatment with proteolytic enzymes (trypsin, pancreatin or elastine) results in the enhancement of viral infectivity and is essential for successful cultivation of human rotaviruses [7, 11].
Viral infectivity can be inactivated by disinfectants such as ethanol, phenol, formalin, chlorine, and Lysol [12-14].
ISOLATION AND CULTIVATION
Although the various animal rotaviruses, such as simian rotavirus SA-11, multiply easily in cell cultures, group A human strains have only recently been successfully cultivated in vitro [15-17].
The cultivation of human rotaviruses requires preliminary treatment of the virus with trypsin (10 #g/mi), the presence of the enzyme in the culture medium, the use of rotating cell cultures and incubation at 37°C. Before the establishment of these culture conditions, abortive in vitro infections were often observed, in which, despite the production of viral antigens, infectious particles were not released in the supernatant. Using the cultivation procedures described above, human rotaviruses extracted from stool samples can be
Table 1. General features of rotaviruses
Morphology lcosahedral 65-75 nm particles Two outer capsid shells and a core
Physicochemical properties Resistant to ether (non enveloped) Infectivity stable at pH 3.0 and relatively heat-stable
Nucleic acid 11 dsRNA segments Each segment codes for at least one protein
Proteins Six structural proteins (for group A): VPI, VP2, VP3 (inner core), VP6 (inner capsid), VP4 and VP7 (outer capsid)
Antigenic properties Six VP6-serogroups (A-F) described Two VP6-subgroups recognized within group A Group A serotypes: I1 VP7- and nine VP4-types (G and P, respectively)
Replication Penetration depends on VP4 after specific cleavage by proteases Cytoplasmic replication Inclusion body formation Final assembly occurs within cisternae of ER

The rotavirus genus 307
isolated in MA-104 (simian foetal kidney) cell cultures with an about 70% efficiency. Nevertheless, an efficiency rate of approx. 100% can be achieved in the direct virus isolation from clinical samples when primary cultures of simian kidney cells or a cell line derived from a human colon adenocarcinoma (HT-29) are used [18]. The cultivation of the non-A group rotavirus presents considerable difficulties and, until now, only a group C strain has been isolated in cell cultures.
VIRUS-CELL INTERACTIONS
The identification of the rotavirus protein which mediates viral binding to the host cell has been uncertain. The two virus outer capsid proteins VP7 and VP4 have been considered [19, 20]. The chemical nature of the cellular receptor for rotaviruses has not been completely elucidated. Different glycoconjugates have been shown to bind the virus and to prevent cell attachment [20-26]. The morphogenesis of rotaviruses is unique. Viral particles assemble near the periphery of viroplasm and subsequently bud into the endoplasmic reticulum (ER) cisternae acquiring membrane. Mature non enveloped particles derive from enveloped particles by the loss or transformation of the outer rim and these infective virions are released by lysis from infected cells (Fig. 2).
GENOME
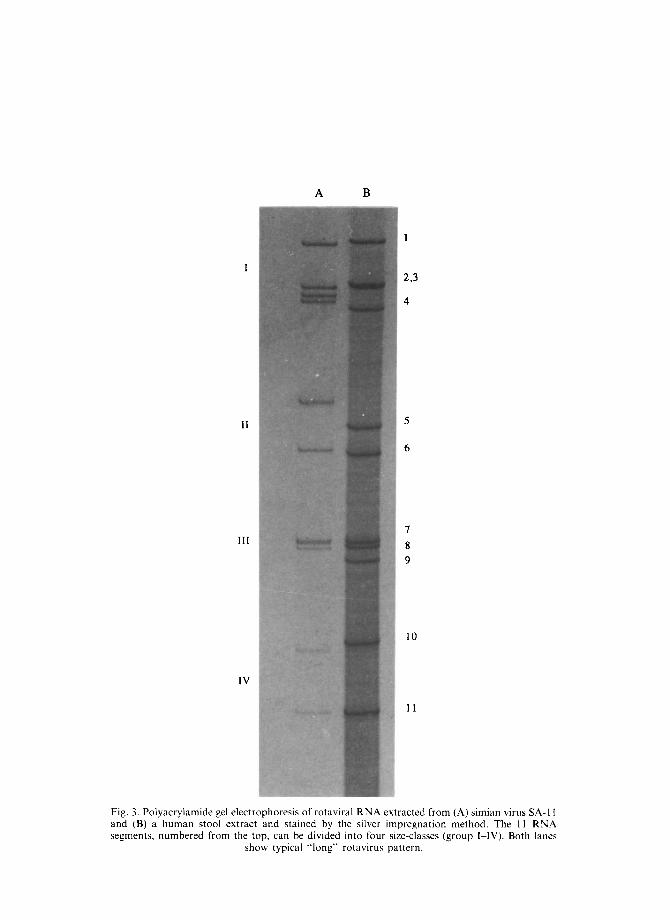
The rotavirus genome contains 11 segments of double strand RNA which range in molecular weight from 2 x 105 to 2.2 x 106 Da [27]. Analysis of genome by polyacrylamide gel electrophoresis (PAGE) is a relatively easy and rapid technique for virus detection and epidemiological studies. In most cases, the electrophoretic pattern is comprised of four high molecular weight dsRNA segments, two middle-sized segments, a distinctive triplet of small segments and two smaller segments (Fig. 3). On the basis of the different electrophoretic mobility of the last two genomic segments (10 and 11), electrophoretic patterns referred to as "long", "short" or "super short" have been identified. However, since distinct RNA patterns can arise by different mechanisms, (shift, drift, rearrange- ments), these profiles cannot be used as the only criteria for classification of a virus strain. Protein products encoded in each of the 11 genome segments is shown in Table 2.
STRUCTURAL PROTEINS
The virion is made up of four major structural proteins: VP4, VP7, VP6 and VP2 [28-29]. VP4 and VP7 are the two surface proteins on the outer capsid. VP4, the product of gene 4, is the viral hemagglutinin [30], forms spike-like structures of about 10 nm [31-33], and is an important determinant of virulence [34]. The cleavage of VP4 into two subunit proteins (VP5* and VP8*) by trypsin is essential for viral penetration into host cells [35-36]. VP7, the product of gene 9, makes up over 90% of the viral surface, is a glycoprotein of the high mannose type and is primarily responsible for determining the viral serotype [37]. VP7 is encoded by genome segment 9 of the SA-11 simian rotavirus, genome segment 8 of the UK bovine virus and genome segment 7 of rhesus rotavirus. The inner capsid of rotavirus is composed of the major structural protein VP6 (the product of gene 6). VP6 makes up 50% of the virion by weight and is primarily responsible for group and subgroup antigenic reactivity [38, 39]. Rotaviruses comprise six distinct groups

308 GIANFRANCO DONELLI and FABIANA SUPERTI
Table 2. Rotavirus genome segments and related proteins (Adapted from Estes and Cohen [28] and Estes [3])
Nascent Location Protein polypeptide in virus Main
Segment product (mol. wt) particles functions
1 VPI 125.000 Inner core Slightly basic 2 VP2 102.000 Inner core RNA binding 3 VP3 98.000 Inner core Basic 4 VP4
(VP5* + VP8*) 88.000 Outer capsid
5 NS53 58.600 Nonstructural 6 VP6 44.800 Inner capsid
7 NS34 34.600 Nonstructural 8 NS35 36.600 Nonstructural 9 VP7 34.000 Outer capsid
10 NS20 20.300 Nonstructu ral
11 NS26 21.700 Nonstructural
Hemagglutinin, protease-enhanced in- fectivity, neutralization antigen, vir- ulence, putative fusion region
Slightly basic Hydrophobic, trimer, group-subgroup
antigen required for transcription Slightly acidic, RNA binding Basic, RNA binding Rough endoplasmic reticulum (RER)
integral membrane protein, neutral- ization antigen, glycoprotein
RER transmembrane glycoprotein, role in morphogenesis
Slightly basic
(A-F): A, B and C groups are found in both humans and animals, whereas D, E and F groups have been found only in animals to date [3, 28]. The inner shell surrounds the core, which encompasses the genome and is composed predominantly of VP2 (the product of gene 2) as well as minor amounts of VP1 and VP3 [33].
PATHOGENESIS
Rotaviruses infect the mature enterocytes of the upper small intestine. Infection usually spreads from the proximal small bowel to the ileum over a period of 1-2 days. Histopathologic studies have revealed villous shortening, reticular cell enlargement, mitochondrial swelling, lymphocyte infiltration of the villous lamina propria and irregular microvilli. Infection generally lasts about one week; however, the period may be substantially longer in immunocompromised patients [40].
Recent investigations on mice with combined immunodeficiency as well as on immuno- compromised children indicate that rotaviruses can also infect the liver under some circumstances [41, 42].
The mechanism of rotavirus-induced diarrhoea is not completely known. The major mechanism appears to be a decreased absorption of salt and water related to selective infection of the absorptive intestinal villous cells, resulting in net fluid secretion. Impaired D-xylose absorption has been observed [43], and some patients have depressed levels of disaccharidases [1] resulting in carbohydrate malabsorption with osmotic diarrhoea.
EPIDEM1OLOGY
Group A rotaviruses have clearly been established as causing severe dehydrating gastroenteritis in young children (under the age of two) in both industrialized and developing countries [44, 45].
Group B rotaviruses have been associated with very large epidemics of diarrhoeal disease primarily in adults in China and have only rarely been identified elsewhere [46].
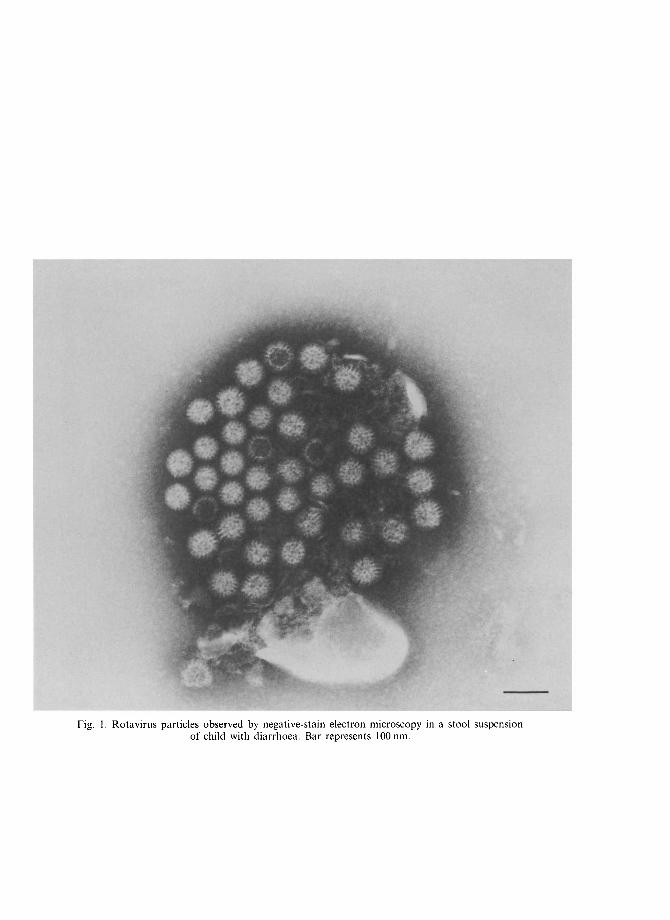
Fig. 1. Rotavirus particles observed by negative-stain electron microscopy in a stool suspension of child with diarrhoea. Bar represents 100 rim.
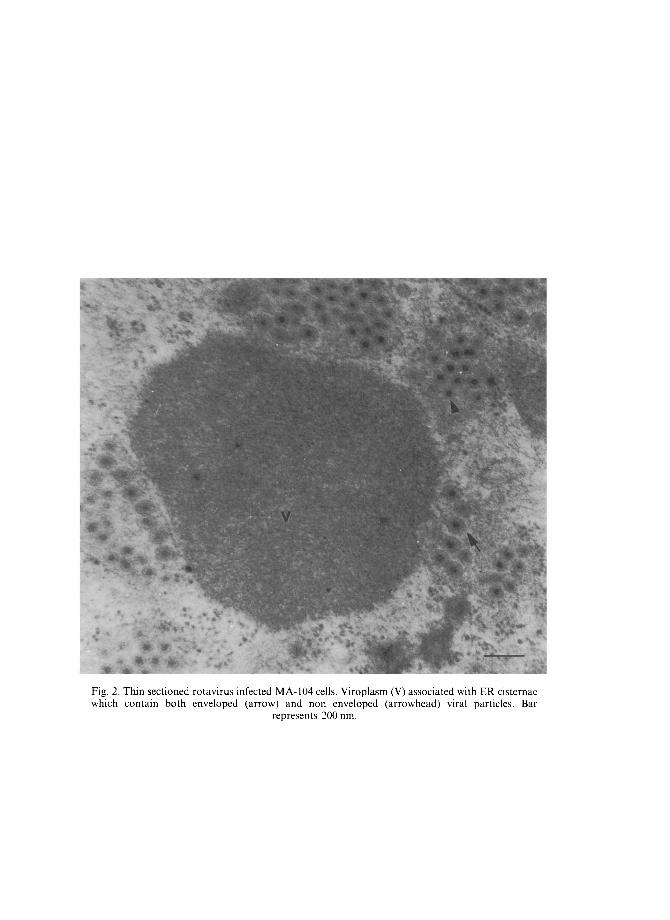
Fig. 2. Thin sectioned rotavirus infected MA-104 cells. Viroplasm (V) associated with ER cistcrnae which contain both enveloped (arrow) and non enveloped (arrowhead) viral particles. Bar
represents 200 rim.
I
II
III
IV
5
6
Fig. 3. PO lyacrylan lide gel and (B) a I human stool e segr nents, numbere :d from
elect .rophoresis of rotaviral RNA extr acted xtraq :t and stained by the silver imr lregna the top. can be divided into four siz e-clas
Sl how typical “long” rotavirus pat tern.
from (A) simian 1 virus SA-I I ltion method. Th e I1 RNA
ses (E group I-IV). Botl h lanes


The ro tav i rus genus 313
Group C viruses have been sporadically reported to cause diarrhoeal disease in children in several areas of the world [47] (Table 3).
In temperate climates, group A rotavirus infection may be responsible for Close to half of the hospitalizations of infants during the winter months, whereas in tropical climates, infections tend to occur year around. Transmission of the virus is thought to be primarily by the fecal-oral route, although transmission through food and water has been suggested. Virus specific IgA has been detected in pharingeal secretions of patients with rotavirus gastroenteritis, suggesting the possibility of transmission via a respiratory route or by ingestion of virus aerosols [48]. Moreover, rotavirus infection of the oropharynx and respiratory tract has been recently reported [49].
Infant mortality due to diarrhoeal diseases is high in developing countries, and rotavirus is thought to be of major importance. Chronic disease is not usually found, but may be seen in immunodeficient or immunosuppressed patients [40, 50-52]. Rotavirus infection may also be asymptomatic, especially in newborn children [53-55] and in adults. Outbreaks of rotavirus gastroenteritis among premature infants have been reported [56].
The peak incidence of illness occurs between 6 and 24 months of life, but considerable morbidity is seen throughout the second year. During the first 2 or 3 months of life, infants appear to be relatively resistant to rotaviral illness, most likely due to transplacentai transfer of maternal antibodies [57]. After 3 years of age, children rarely have severe rotaviral illness. Group A rotaviruses are usually not considered common pathogens in adults, but they can cause illness under a variety of circumstances [58]. Nosocomial infections in hospital wards are a known problem and appropriate infection control measures should be undertaken when patients with suspected viral gastroenteritis are admitted [59~50].
The epidemiology of rotavirus infections can be followed by monitoring a variety of serologic markers or by determining the electrophoretic pattern of viral genomic RNA (the electropherotype) in faecal specimens. Several serotypes of human group A rotavirus have been identified, although the great majority of infections appear to be caused by only the 1-4 serotypes [61-63].
IMMUNITY
Seroconversion occurs following infection in both animals and man. Serological surveys carried out in different areas of the world have demonstrated that over 80% of the population has antibodies from the age of about 3 years [64-65]. Immunity may wane among the elderly, rendering them susceptible again to severe disease.
Genetic studies with specific viral reassortants and passive-transfer studies with mono- clonal antibodies directed at specific rotaviral proteins have demonstrated that, in general,
Table 3. Clinical relevance and epidemiology characteristics of human rotaviruses
Epidemiologic Clinical Group features symptoms
Major cause of severe dehydrating gastroenteritis in infants and young children worldwide (in winter months in temperate climates) Large epidemics of diarrhoeal disease primarily in adults in China Sporadically reported to cause diarrhoeal disease in children in many areas of the world
Dehydrating diarrhoea for 3-8 days, fever and vomiting frequent
Severe dehydrating diarrhoea for 3-5 days
Dehydrating diarrhoea for 3-8 days, fever and vomiting frequent
314 GIANFRANCO DONELLI and FABIANA SUPERTI
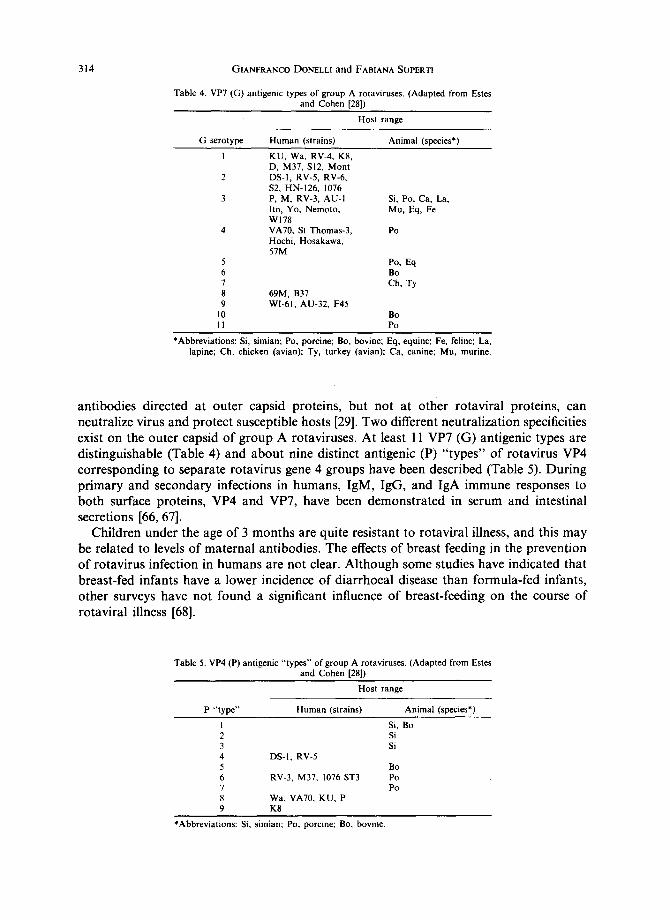
Table 4. VP7 (G) antigenic types of group A rotaviruses. (Adapted from Estes and Cohen [28])
G serotype Human (strains)
Host range
Animal (species*)
I KU, Wa, RV-4, K8, D, M37, SI2, Mont
2 DS-I, RV-5, RV-6, $2, HN-126, 1076
3 P, M, RV-3, AU-I lto, Yo, Nemoto, W178
4 VA70, St Thomas-3, Hochi, Hosakawa, 57M
Si, Po, Ca, La, Mu, Eq, Fe
Po
5 Po, Eq 6 Bo 7 Ch, Ty 8 69M, B37 9 Wl-61, AU-32, F45
10 Bo I 1 Po
*Abbreviations: Si, simian; Po, porcine; Bo, bovine; Eq, equine; Fe, feline; La, lapine; Ch, chicken (avian); Ty, turkey (avian); Ca, canine; Mu, murine.
antibodies directed at outer capsid proteins, but not at other rotaviral proteins, can neutralize virus and protect susceptible hosts [29]. Two different neutralization specificities exist on the outer capsid of group A rotaviruses. At least 11 VP7 (G) antigenic types are distinguishable (Table 4) and about nine distinct antigenic (P) "types" of rotavirus VP4 corresponding to separate rotavirus gene 4 groups have been described (Table 5). During primary and secondary infections in humans, IgM, IgG, and IgA immune responses to both surface proteins, VP4 and VP7, have been demonstrated in serum and intestinal secretions [66, 67].
Children under the age of 3 months are quite resistant to rotaviral illness, and this may be related to levels of maternal antibodies. The effects of breast feeding in the prevention of rotavirus infection in humans are not clear. Although some studies have indicated that breast-fed infants have a lower incidence of diarrhoeal disease than formula-fed infants, other surveys have not found a significant influence of breast-feeding on the course of rotaviral illness [68].
Table 5. VP4 (P) antigenic "types" of group A rotaviruses. (Adapted from Estes and Cohen [281)
Host range
P "type" Human (strains) Animal (species*)
Si, Bo Si Si
DS-I, RV-5 Bo
RV-3, M37, 1076 ST3 Po Po
Wa, VA70, KU, P K8
*Abbreviations: Si, simian; Po, porcine; Bo, bovine.

The rotavirus genus 315
CLINICAL DISEASE
The clinical spectrum of rotavirus infections varies from asymptomatic to severe, with occasional fatal episodes. The commonest clinical findings usually include vomiting (which arise early in the illness), abdominal pain, watery diarrhoea, mild to moderate dehydration with normal serum sodium concentration and mild acidosis [69]. Fever and vomiting frequently precede diarrhoea. Associated clinical findings may include irritability and pharingeal or tympanic membrane erythema. Stools usually do not contain blood or leukocytes. The incubation period is approx. 2 days and the course of the disease is generally 3-8 days. When required, due to severe dehydration, mean hospitalization time is 4 days.
Numerous case reports have associated rotavirus infection with a variety of conditions such as intususception, Reye syndrome, hemolytic uremic syndrome, and Kawasaki syndrome. Evidence for an association with rotavirus has been reported for some cases of neonatal necrotizing enterocolitis [70].
Deaths, rare in developed countries, are frequent in developing countries and typically due to electrolite imbalance.
In adults the infections are usually asymptomatic and accompanied by seroconversion to the virus. Symptomatic infections in adults are usually mild, but severe illness has been reported both in elderly patients [71] and in apparently normal young adults [72]. Infections due to non-group A rotaviruses in China have been reported to cause epidemics of gastroenteritis with severe symptoms in adults [73].
LABORATORY DIAGNOSIS
As clinical manifestations are not sufficiently distinctive, diagnosis requires detection of virus (or viral antigens, or viral nucleic acid) and/or demonstration of a serologic response. The epidemiologic pattern may suggest the diagnosis, but laboratory confirmation is required.
Commercial rotavirus antigen-detection kits are widely available, inexpensive, and allow rapid diagnosis. These include the enzyme immunoassays EIA or ELISA and the latex particles agglutination tests [74-77]. Only small amounts of stools are required for the tests, and samples may be frozen before testing. Since virus shedding is generally maximal during diarrhoea, faecal samples should be obtained as close to illness as possible (within 4 days).
Other methods for virus detection include hybridization of radiolabeled nucleic acid probes to viral RNA [78], or PAGE analysis of viral RNA [77, 79, 80]. Polymerase chain reaction (PCR) techniques have also been developed in the amplification and typing of rotaviral nucleic acid from stool specimens [81, 82]. Direct electron microscopy remains a useful technique for viral detection, in particular for non-group-A rotaviruses, and immune-electron microscopy (IEM) is frequently used as reference method with respect to other assays [83, 84].
Viruses can also be identified by direct isolation from stool samples in cell cultures with the aid of proteolitic enzymes [15-18].
To measure serologic response to infection, there are many techniques such as ELISA, IEM, complement fixation, immunofluorescence, immune-adherence hemagglutination, radio-immunoassay and neutralization tests [85-91]. The complement fixation, initially widely used for the detection of rotavirus antibodies, is efficient for revealing an antibody

316 GIANFRANCO DONELLI and FABIANA SUPERTI
response only in patients between 6 and 24 months o f age [85]. An E L I S A IgA has been particularly effective in demonstra t ing serologic responses in young infants who possess passively acquired maternal IgG antibodies, since IgA does not cross the placenta [92]. This assay may also be used to examine stools for coproant ibodies [93].
Virus detection and/or demonst ra t ion o f a serologic response in an individual patient does not necessarily establish an aetiologic association with patient 's illness, especially in newborns and adults who commonly undergo inapparent infection.
PREVENTION, CONTROL AND TREATMENT OF ROTAVIRUS ILLNESS
Extraordinary progress toward the development o f a rotaviral vaccine has been made [94-95] and a number o f trials in developed and less-developed countries have been carried out [96-98]. Current strategies for development o f a rotavirus vaccine range from cell culture cultivation o f human or animal strains to the application o f molecular biology techniques. The most developed candidate strains have been derived from animal rotaviruses (bovine and simian) or are reassortants containing 10 animal rotavirus gene segments and the human rotavirus gene 9 (encoding VP7 in humans) [98-101]. Another approach is based on the use o f "na tura l ly" at tenuated neonatal human strains [98].
Other possible vaccines under study include the use o f synthetic viral proteins (VP7 or VP4) or viral proteins expressed in prokaryot ic or eukaryotic cell systems by cloned rotavirus c D N A [102 104].
Al though passive immunizat ion has shown promising results [105, 106], it is not practical for protect ion against rotavirus illness, with the exception o f immunocompromised individuals, requiring repeated treatments.
Treatment remains mainly symptomatic . The replacement o f fluid and electrolytes lost by vomiting and diarrhoea, preferably with use o f oral rehydrat ion solutions, is highly effective in the great majori ty o f cases [107]. Of course, if the oral rehydrat ion regimen is not able to correct the fluid and electrolyte loss or if the patient is severely dehydrated or in shock, intravenous fluids must be given. Normal and immunodeficient children have been fed with gammaglobul ins [108] or human milk containing rotavirus antibodies [50], respectively, but these therapies are still under study.
REFERENCES
I, Bishop R. F., Davidson G. P., Holmes |. H. and Ruck B. J. Virus particles in epithelial cells of duodenal mucosa from children with viral gastroenteritis. Lancet 2, 1281-1283 (1973).
2. Flewett T. H., Bryden A. S., Davies H., Woode G. N., Bridger J. C. and Derrick J. M. Relationship between virus from acute gastroenteritis of children and newborn calves. Lancet 2, 61-63 (1974).
3. Estes M. K. Rotaviruses and their replication. In Virology (Edited by Fields B. N. and Knipe D. M.) 2nd Edn, vol. 2, pp. 1329-1352. Raven Press, New York (1990).
4. Bridger J. C. and Woode G. N. Characterization of two particle types of calf rotavirus. J. Gen. Virol. 31, 245 250 (1976).
5. Elias M. M. Separation and infectivity of two particle types of human rotavirus. J. Gen. Virol. 37, 191 194 (1977).
6. Bican P., Cohen J., Charpilienne A. and Scherrer R. Purification and characterization of bovine rotavirus cores. J. Virol. 43, 1113-1117 (1982).
7. Estes M. K., Graham D. Y., Smith E. M. and Gerba C. P. Rotavirus stability and inactivation. J. Gen. Virol. 43, 403~,09 (1979).
8. Almeida J. D., Bradburne A. F. and Wreghitt T. G. The effect of sodium thiocyanate on virus structure. J. Med. Virol. 4, 269 277 (1979).

The rotavirus genus 317
9. Ansari S. A., Springthorpe V. S. and Sattar S. A. Survival and vehicular spread of human rotaviruses: possible relation to seasonality of outbreaks. Rev. Infect. Dis. 13, 448-461 (1991).
10. Weiss C. and Clark H. F. Rapid inactivation of rotaviruses by exposure to acid buffer or acidic gastric juice. J. Gen. Virol. 66, 2725-2730 (1985).
11. Sato K., Inaba Y., Shinozaki T., Fujii R. and Matumoto M. Isolation of human rotavirus in cell cultures. ,4rchs Virol. 69, 155-160 (1981).
12. Tan J. A. and Schnagl R. D. Inactivation of a rotavirus by disinfectants. Med. J. Aust. 1, 19-23 (1981). 13. Sattar S. A., Raphael R. A., Lochnan H. and Springthorpe V. S. Rotavirus inactivation by chemical
disinfectants and antiseptics used in hospitals. Can. J. Microbiol. 29, 1464-1469 (1983). 14. Ward R. L., Bernstein D. I., Knowlton D. R., Sherwood J. R., Young E. C., Cusack T. M., Rubino J. R.
and Schiff G. M. Prevention of surface-to-human transmission of rotaviruses by treatment with disinfectant spray. J. Clin. Microbiol. 29, 1991 1996 (1991).
15. Hasegawa A., Matsuno S., Inouye S., Kono R., Tsurukubo Y., Mukoyama A. and Saito Y. Isolation of human rotaviruses in primary cultures of monkey kidney cells. J. Clin. Microbiol. 16, 387-390 (1982).
16. Wyatt R. G., James H. D. Jr, Pittman A. L., Hoshino Y., Greenberg H. B., Kalica A. R., Flores J. and Kapikian A. Z. Direct isolation in cell culture of human rotaviruses and their characterization into four serotypes. J. Clin. Microb'iol. 18, 310-317 (1983).
17. Ward R. L., Knowlton D. R. and Pierce M. J. Efficiency of human rotavirus propagation in cell culture. J. Clin. Microbiol. 19, 748-753 (1984).
18. Superti F., Tinari A., Baldassarri L. and Donelli G. HT-29 cells: a new substrate for rotavirus growth. Archs Virol. 116, 159-173 (1991).
19. Fukuhara N., Yoshie O., Kitaoka S. and Konno T. Role of VP3 in human rotavirus internalization after target cell attachment via VP7. J. Virol. 62, 2209-2218 (1988).
20. Bass D. M., Mackow E. R. and Greenberg H. B. Identification and partial characterization of a rhesus rotavirus binding glycoprotein on murine enterocytes. Virology 183, 602~10 (1991).
21. Superti F. and Donelli G. Gangliosides as binding sites in SA-I 1 rotavirus infection of LLC-MK2 cells. J. Gen. Virol. 72, 2467-2474 (1991).
22. Superti F., Marchetti M., Seganti L., Conte M. P. and Orsi N. Human serum non-antibody inhibitors towards SA-l 1 rotavirus hemagglutination. Microbiologica 14, 25-30 (1991).
23. Yolken R. H., Peterson J. A., Vonderfecht S. L., Fouts E. T., Midthun K. and Newburg D. S. Human milk mucin inhibits rotavirus replication and prevents experimental gastroenteritis. J. Clin. Invest. 90, 1984-1991 (1992).
24. Srnka C. A., Tiemeyer M., Gilbert J. H., Moreland M., Schweingruber H., De Lappe B. W., James P. G., Gant T., Willoughby R. E., Yolken R. H., Nashed M. A., Abbas S. A. and Laine R. A. Cell surface ligands for rotavirus: mouse intestinal glycolipids and synthetic carbohydrate analogs. Virology 190, 794-805 (1992).
25. Superti F., Seganti L., Marchetti M., Marziano M. L. and Orsi N. SA-11 rotavirus binding to human serum lipoproteins. Med. Microbiol. Immunol. 181, 77-86 (1992).
26. Superti F., Marziano M. L., Tinari A. and Donelli G. Effect of polyions on the infectivity of SA-I 1 rotavirus in LLC-MK2 cells. Comp. hnmun. Microbiol. Infect. Dis. 16, 55~2 (1993).
27. Kalica A. R., Sereno M. M., Wyatt R. G., Mebus C. A., Chanock R. M. and Kapikian A. Z. Comparison of human and animal rotavirus strains by gel electrophoresis of RNA. Virology 87, 247-255 (1978).
28. Estes M. K. and Cohen J. Rotavirus gene structure and function. Microbiol. Rev. 53, 410-449 (1989). 29. Blacklow N. R. and Greenberg H. B. Viral gastroenteritis. N. Eng. J. Med. 325, 252-264 (1991). 30. Mackow E. R., B~trnett J., Chan H. and Greenberg H. B. The rhesus rotavirus outer capsid protein VP4
functions as a hemagglutinin and is antigenically conserved when expressed by a baculovirus recombinant. J. Virol. 63, 1661-1668 (1989).
31. Yeager M., Dryden K. A., Olson N. H., Greenberg H. B. and Baker T. S. Three-dimensional structure of rhesus rotavirus by cryoelectron microscopy and image reconstruction. J. Cell Biol. 110, 2133-2144 (1990).
• 32. Prasad B. V. V., Burns J. W., Marietta E., Estes M. K. and Chiu W. Localization of VP4 neutralization sites in rotavirus by three-dimensional cryo-electron microscopy. Nature 343, 476-479 (1990).
33. Shaw A. L., Rothnagel R., Chert D., Ramig R. F., Chiu W. and Prasad B. V. V. Three-dimensional visualization of the rotavirus hemagglutinin structure. Cell 74, 693-701 (1993).
34. OflSt P. A., Blavat G., Greenberg H. B. and Clark H. F. Molecular basis of rotavirus virulence: role of gene segment 4. J. Virol. 57, 46-49 (1986).
35. Kaljot K. T., Shaw R. D., Rubin D. H. and Greenberg H. B. Infectious rotavirus enters the cells by direct membrane penetration, not by endocytosis. J. Virol. 62, 1136-1144 (1988).
36. Konno T., Suzuki H., Kitaoka S., Sato T., Fukuhara N., Yoshie O., Fukudome K. and Numazaki Y. Proteolytic enhancement of human rotavirus infectivity. Clin. Infect. Dis. 16, 92-97 (1993).
37. Both G. W., Mattick J. S. and Bellamy A. R. Serotype-specific glycoprotein of simian 11 rotavirus: coding assignment and gene sequence. Proc. hath. Acad. Sci. U.S.A. 80, 3091-3095 (1982).
38. Gary G. W. Jr, Black D. R. and Palmer E. Monoclonal IgG to the inner capsid of human rotavirus. Archs Virol. 72, 223-227 (1982).

318 GIANFRANCO DONELLI and FABIANA SUPERTI
39. Singh N., Sereno M., Flores J. and Kapikian A. Z. Monoclonal antibodies to subgroup 1 rotavirus. Infect. Immun. 42, 835-837 (1983).
40. Oishi I., Kimura T., Murakami T., Haruki K., Seto Y., Minekawa Y. and Funamoto H. Serial observations of chronic rotavirus infection in an immunodeficient child. Microbiol. ImmunoL 35, 953 961 (1991).
41. Riepenhoff-Talty M., Dharakul T., Kowalski E., Michalak S. and Ogra P. L. Persistent rotavirus infection in mice with severe combined immunodeficiency. J. Virol. 61, 3345-3348 (1987).
42. Grunow J. E., Dunton S. F. and Waner J. L. Human rotavirus-like particles in a hepatic abscess. J. Pediatr. 106, 73-76 (1985).
43. Mavromichalis J., Evans N., McNeish A. S., Bryden A. S., Davies H. A. and Flewett H. A. Intestinal damage in rotavirus and adenovirus gastroenteritis assessed by o-xylose malabsorption. Arch. Dis. Child. 52, 589-591 (1977).
44. Kapikian A. Z. and Chanock R. M. Rotaviruses. In Virology (Edited by Fields B. N. and Knipe D. M.) 2nd Edn, vol. 2, pp. 1353 1404. Raven Press, New York (1990).
45. Cook S. M., Glass R. I., LeBaron C. W. and Ho M. S. Global seasonality of rotavirus infections. Bull. Worm Health Organ. 68, 171--177 (1990).
46. Penaranda M. E., Ho M., Fang Z., Dong H., Bai X., Duan S., Ye W. W., Estes M. K., Echeverria P., Hung T. and Glass R. I. Seroepidemiology of adult diarrhea rotavirus in China. 1977 1987. J. Clin. Microbiol. 27, 2180~2183 (1989).
47. Von Bonsdorf C. H. and Svensson L. Human serogroup C rotavirus in Finland. Scand. J. Infect. Dis. 20, 475-478 (1988).
48. Stals F., Walther F. 3. and Bruggeman C. A. Faecal and pharyngeal shedding of rotavirus and rotavirus IgA in children with diarrhoea. J. Med. Virol. 14, 333 339 (1984).
49. Zheng B. J., Chang R. X., Ma G. Z., Xie J. M., Liu Q., Liang X. R. and Ng M. H. Rotavirus infection of the oropharynx and respiratory tract in young children. J. Med. Virol. 34, 29--37 (1991).
50. Saulsbury F. T., Winkelstein J. A. and Yolken R. H. Chronic rotavirus infection in immunodeficiency. J. Pediatr. 97, 61~55 (1980).
51. Yolken R. H., Bishop C. A., Townsend T. R., Bolyard E. A., Bartlett J., Santos G. W. and Saral R. Infectious gastroenteritis in bone-marrow transplant recipients. N. Eng. J. Med. 306, 1009 1012 (1982).
52. Wood D. J., David T. J., Chrystie I. L. and Totterdell I. L. Chronic enteric virus infection in two T-cell immunodeficient children. J. Med. Virol. 24, 435-444 (1988).
53. Haffejee 1. E. Neonatal rotavirus infections. Rev. lnfect. Dis. 13, 957 962 (1991). 54. Steele A. D., van Niekerk M. C., Geyer A., Bos P. and Alexander J. J. Further characterisation of human
rotavirus isolated from asymptomatically infected neonates in South Africa. J. Med. Virol. 38, 22 26 (1992). 55. Sukumaran M., Gowda K., Maiya P. P., Srinivas T. P., Kumar M. S., Aijiaz S., Reddy R. R., Padilla L.,
Greenberg H. B. and Rao C. D. Exclusive asymptomatic neonatal infections by human rotavirus strains having subgroup 1 specifcity and "long" RNA electropherotype. Archs ViroL 126, 239-251 (1992).
56. Rocchi G., Vella S., Resta S., Cochi D., Donelli G., Tangucci F., Menichella D., Varveri A., lnglese R. Outbreak of rotavirus gastroenteritis among premature infants. Br. Med. J. 283, 886 (1981).
57. Tufvesson B., Polberger S., Svanberg L. and Sveger T. A prospective study of rotavirus infections in neonatal and maternity wards. Acta Paediatr. Stand. 75, 211 215 (1986).
58. Hardy D. B. Epidemiology of rotaviral infection in adults Rev. Infect. Dis. 9, 461-469 (1987). 59. Flewett T. H., Rotavirus in the home and hospital nursery. Br. Med. J. 287, 568 569 (1983). 60. Rodriguez W. J., Kim H. W., Brandt C. D., Gardner M. K. and Parrott R. H. Use of electrophoresis of
RNA from human rotavirus to establish the identity of strains involved in outbreaks in a tertiary care nursery. J. Infect. Dis. 148, 34-40 (1993).
61. Hoshino Y., Wyatt R. G., Greenberg H. B., Flores J. and Kapikian A. Z. Serotypic similarity and diversity of rotaviruses of mammalian and avian origin as studied by plaque-reduction neutralization. J. lnfecc Dis. 149, 694-702 (1984).
62. Arista S., Giovannelli L., Pistoia D., Cascio A., Parea M. and Gerna G. Electropherotypes, subgroups and serotypes of human rotavirus strains causing gastroenteritis in infants and young children in Palermo, Italy, from 1985 to 1989. Res. Virol. 141,435-448 (1990).
63. Woods P. A., Gentsch J., Gouvea V., Mata L., Santosham M., Bai Z. S., Urasawa S. and Glass R. I. Distribution ofserotypes of human rotavirus in different populations. J. Clin. Microbiol. 30, 781 785 (1992).
64. Bishop R. F., Barnes G. L., Cipriani E. and Lund J. S. Clinical immunity after rotavirus infection: a prospective longitudinal study in young children. N. Eng. J. Med. 309, 72-76 (1983).
65. Riepenhoff-Talty M. Local and systemic antibody response to rotavirus infection. J. Pediatr. Gastroenterol. Nutr. 5, 167-172 (1986).
66. Coulson B. S., Grimwood K., Masendycz P. J., Lund J. S., Mermelstein N., Bishop R. F. and Barnes G. L. Comparison of rotavirus immunoglobulin A coproconversion with other indices of rotavirus infection in a longitudinal study in childhood. J. Clin. Microbiol. 28, 1367 1374 (1990).
67. Richardson S. C. and Bishop R. F. Homotypic serum antibody responses to rotavirus proteins following primary infection of young children with serotype 1 rotavirus. J. Clin. Microbiol. 28, 1891 1897 (1990).

The rotavirus genus 319
68. Glass R. I., Stoll B. J., Wyatt R. G., Hoshino Y., Banu H. and Kapikian A. Z. Observations questioning a protective role for breast-feeding in severe rotavirus diarrhea. Acta Pediatr. Scand. 75, 713-718 (1986).
69. Isolauri E., Jalonen T. and Maki M. Acute gastroenteritis. Acta Pediatr. Scand. 78, 685~591 (1989). 70. Rotbart H. A., Nelson W. L., Glode M. P., Triffon T. C., Kogut S. J. H., Yolken R. H., Hernandez J. A.
and Levin M. J. Neonatal rotavirus associated necrotizing enterocolitis: case control study and prospective surveillance during an outbreak. J. Pediatr. 112, 87-93 (1988).
71. Marrie T. J., Lee S. H. S., Faulkner R. S., Ethier J., Young C. H. Rotavirus infections in a geriatric population. Arch. Intern. Med. 142, 313-316 (1982).
72. Echeverria P., Blacklow N. R., Cukor G. G., Vibulbandhitkit S., Changchawalit S. and Boonthai P. Rotavirus as cause of severe gastroenteritis in adults. J. Clin. Microbiol. 18, 663--667 (1983).
73. Hung T., Chen G., Wang C., Yao H., Fang Z., Chao T., Chou Z., Ye W., Chang X., Den S., Liong X. and Chang W. Waterborne outbreak of rotavirus diarrhoea in adults in China caused by a novel rotavirus. Lancet 1, 1139-1142 (1984).
74. Thomas E. E., Puterman M. L., Kawano E. and Curran M. Evaluation of seven immunoassays for detection of rotavirus in pediatric stool samples. J. Clin. Microbiol. 26, 1189 1193 (1988).
75. Dennehy P. H., Gauntlett D. R. and Tente W. E. Comparison of nine commercial immunoassays for the detection of rotavirus in fecal specimens. J. Clin. Microbiol. 26, 1630-1634 (1988).
76. Donelli G., Ruggeri F. M., Tinari A., Marziano M. L., Menichella D., Caione D., Concato C., Rocchi G. and Vella S. A three year diagnostic and epidemiological study on viral infantile diarrhoea in Rome. Epidemiol. Infect. 11~, 311-320 (1988).
77. Donelli G., Superti F., Tinari A., Marziano M. L., Caione D., Concato C. and Menichella D. Viral childhood diarrhoea in Rome: a diagnostic and epidemiological study. Microbiologica 16, 215-224 (1993).
78. Dimitrov D. H., Graham D. Y. and Estes M. K. Detection of rotaviruses by nucleic acid hybridization with cloned DNA of simian rotavirus SA11 genes. J. Infect. Dis. 162, 293-300 (1985).
79. Pacini D. L., Brady M. T., Budde C. T., Connell M. J., Hamparian V. V. and Hughes J. H. Polyacrylamide gel electrophoresis of RNA compared with polyclonal- and monoclonal-antibody-based enzyme im- munoassays for rotavirus. J. Clin. Microbiol. 26, 194 197 (1988).
80. Ruggeri F. M., Marziano M. L., Tinari A., Salvatori E. and Donelli G. Four-year study of rotavirus electropherotypes from cases of infantile diarrhea in Rome. J. Clin. Microbiol. 27, 1522 1526 (1989).
81. Gouvea V., Glass R. I., Woods P., Taniguchi K., Clark H. F., Forrester B. and Fang Z. Polymerase chain reaction amplification and typing of rotavirus nucleic acid from stool specimens. J. Clin. Microbiol. 28, 276-282 (1990).
82. Ushijima H., Koike H., Mukoyama A., Hasegawa A., Nishimura S. and Gentsch J. Detection and serotyping of rotaviruses in stool specimens by using reverse transcription and polymerase chain reaction amplification. J. Med. Virol. 38, 292-297 (1992).
83. Gerna G., Sarasini A., Coulson B. S., Parea M., Torsellini M., Arbustini E. and Battaglia M. Comparative sensitivities of solid-phase immune electron microscopy and enzyme-linked immunosorbent assay for serotyping of human rotavirus strains with neutralizing monoclonal antibodies. J. Clin. Microbiol, 26, 1383-1387 (1988).
84. Ruggeri F. M., Marziano M. L., Salvatori E., Bisicchia R., Scardellato V., Scagnelli M., Modolo M. L., Santini G. and Donelli G. Laboratory diagnosis of rotavirus infection in diarrhoeal patients by immunoen- zymatic and latex-agglutination assay. Microbiologica 15, 249-258 (1992).
85. Gust I. D., Pringle R. C., Barnes G. L., Davidson G. P. and Bishop R. F. Complement-fixing antibody response to rotavirus infection. J. Clin. Microbiol. 5, 125-130 (1977).
86. Watanabe H. and Holmes I. H. Filter paper solid-phase radioimmunoassay for human rotavirus surface immunoglobulins. J. Clin. Microbiol. 6, 319-324 (1977).
87. Martin M. L., Gary G. W. Jr and Palmer E. L. Comparison of hemagglutination-inhibition, complement- fixation and enzyme-linked immunosorbent assay for quantitation of human rotavirus antibodies. Archs Virol. 62, 131-136 (1979).
88. Matsuno S., Inouye S., Hasegawa A. and Kono R. Assay of human rotavirus antibody by immune adherence hemagglutination with a cultivable human rotavirus as antigen. J. Clin. Microbiol. 15, 163-165 (1982).
89. Grimwood K., Lund J. C. S., Coulson B. S., Hudson I. L., Bishop R. F. and Barnes G. L. Comparison of serum and mucosal antibody responses following severe acute rotavirus gastroenteritis in young children. J. Clin. MicrobioL 26, 732-739 (1988).
90. Knowlton D. R., Spector D. M. and Ward R. L. Development of an improved method for measuring neutralizing antibody to rotavirus. J. Virol. Meth. 33, 127-134 (1991).
91. Tsunemitsu H., Jiang B. and Saif L. J. Detection of group C rotavirus antigens and antibodies in animals and humans by enzyme-linked immunosorbent assays. J. Clin. Mierobiol. 30, 2129-2134 (1992).
92. Losonsky G. A., Rennels M. B., Lim Y., Krall G., Kapikian A. Z. and Levine M. M. Systemic and mucosal immune responses to rhesus rotavirus vaccine MMU 18006. Pediatr. Infect. Dis. J. 7, 388-393 (1988).

320 GIANFRANCO DONELLI and FABIANA SUPERTI
93. Coulson B. S., Grimwood K., Hudson I. L., Barnes G. L. and Bishop R. F. Role of coproantibody in clinical protection of children during reinfection with rotavirus. J. Clin. Microbiol. 30, 1678 1684 (1992).
94. Flores J. and Kapikian A. Z. Rotavirus vaccines. Biotechnology 20, 255-258 (1992). 95. Bishop R. F. Development of candidate rotavirus vaccines. Vaccine 11, 247-254 (1993). 96. Clark H. F., Borian F. E. and Plotkin S. A. Immune protection of infants against rotavirus gastroenteritis
by serotype 1 reassortant of bovine rotavirus WC3. Infect. Dis. 161, 1099-1104 (1990). 97. Perez-Schael I., Garcia D., Gonzales M., Daoud N., Perez M., Cunto W., Kapikian A. Z. and Flores J.
Prospective study of diarrhoeal diseases in Venezuelan children to evaluate the efficacy of rhesus rotavirus vaccine. J. Med. Virol. 30, 219-229 (1990).
98. Flores J., Perez-Schael I., Blanco M., White L., Garcia D., Vilar M., Cunto W., Gonzalez R., Urbina C., Boher J., Mendez M. and Kapikian A. Z. Comparison of reactogenicity and antigenicity of M37 rotavirus vaccine and rhesus-rotavirus-based quadrivalent vaccine. Lancet 2, 330-334 (1990).
99. Reennels M. B., Losonsky G. A., Young A. E., Schindledecker C. L., Kapikian A. Z. and Levine M. M. An efficacy trial for the rhesus rotavirus vaccine in Maryland. Am. J. Dis. Child. 144, 601-604 (1990).
I00. Bernstein D. I., Smith V. E., Sander D. S., Pax K. A., Schiff G. M. and Ward R. L. Evaluation of WC3 rotavirus vaccine and correlates of protection in healthy infants. J. Infect. Dis. 162, 1055-1062 (1990).
101. Koboyashi M., Araki K., Thompson J., Tollefson S. J. and Wright P. F. Sequential administration of two human-rhesus rotavirus reassortant strains of VP7 serotype I and 2 specificity of infants and young children. J. Infect. Dis. 167, 748-752 (1993).
102. Streckert H. J., Brussow H. and Werchau H. Synthetic peptide corresponding to the cleavage region of VP3 from rotavirus SA-11 induces neutralizing antibodies. J. Virol. 62, 4265-4269 (1988).
103. Andrew M. E., Boyle D. B., Coupar B. E., Withfeld P. L., Both G. W. and Bellamy A. R. Vaccinia virus recombinants expressing the SA11 rotavirus VP7 glycoprotein gene induce serotype-specific neutralizing antibodies. J. Virol. 61, 1054-1060 (1987).
104. Redmond M. J., ljaz M. K., Parker M. D., Sabara M. I., Dent D., Gibbons E. and Babiuk L. A. Assembly of recombinant rotavirus proteins into virus-like particles and assessment of vaccine potential. Vaccine 11, 273-281 (1993).
105. Barnes G. L., Doyle L. W., Hewson P. H., Knoches A. M. L., McLellan J. A. and Kitchen W. H. A randomised trial of oral gamma globulin in low-birth-weight infants infected with rotavirus. Lancet 1, 1371-1373 (1982).
106. Turner R. B. and Douglas K. K. Passive immunization for prevention of rotavirus illness in healthy infants. Pediatr. Infect. Dis. J. 12, 718-722 (1993).
107. Santosham M., Burns B., Nadkarni V., Foster S., Garrett S., Croll L., O'Donovan J. C., Pathak R. and Sack R. B. Oral rehydration therapy for acute diarrhea in ambulatory children in the United States: a double-blind comparison of four different solutions. Pediatrics 76, 159-166 (1985).
108. Guarino A., Berni Canani R., Russo S., Albano F., Berni Canani M., Ruggeri F. M., Donelli G. and Rubino A. Oral immunoglobulins for treatment of acute rotaviral gastroenteritis. Pediatrics, 93, 12-16 (1994).




















