
ON THE BEHAVIOUR OF WIREWORMS OF THE GENUS ...
13
[43] ON THE BEHAVIOUR OF WIREWORMS OF THE GENUS AGRIOTES ESCH. (COLEOPTERA, ELATERIDAE) I. REACTIONS TO HUMIDITY BY A. D. LEES, From the Zoological Laboratory, Cambridge {Received 14 July 1943) (With Six Text-figures) Humidity reactions have recently been demon- strated in a number of terrestrial arthropods. In several instances—for example in Locusta tmgratoria (Kennedy, 1937), Culex fatigans (Thomson, 1938), Blatta orientalis (Gunn & Cosway, 1938), Tenebrio molitor (Pielou & Gunn, 1940) and in Pediculus humanus corporis (Wigglesworth, 1941)—it has been shown that such responses lead to the avoidance of moist air, a type of behaviour which is not easily associated with any specific physiological require- ments. On the other hand, certain animals such as woodlice which are known to possess little capacity to resist desiccation, avoid dry, and aggregate in humid air (Gunn, 1937; Waloff, 1941); this type of behaviour is clearly of some adaptive significance. Wireworms also are unable to withstand prolonged desiccation. Thus according to the data of Subklew (1934a), large larvae weighing about 50 mg. lose up to half their body weight in 3 days when intro- duced into still, half-saturated air at 20 0 C. With this in mind, the humidity behaviour was tested in an alternative chamber. When it became clear that an intense response was shown to certain humidity gradients, the nature of the humidity receptors and the mechanisms displayed in orientating to humidity were also examined. The present paper deals with these aspects of the humidity behaviour and their relation to the humidity conditions prevailing in the normal environment, the soil. METHODS AND MATERIAL The apparatus used for testing the general humidity behaviour of wireworms was substantially the same as the 'alternative chamber' described by Gunn & Kennedy (1936). It consisted of a circular glass vessel, 22 cm. in diameter and 6 cm. deep, which was closed by an airtight glass roof (Fig. 1 A). In order to reduce slightly the area of contact between the two halves of the chamber, a vertical partition (a), 2 cm. deep, was attached to the roof. The desired humidities were maintained by means of sulphuric acid-water mixtures (Wilson, 1921) contained in dishes (6) on the floor. The arena was provided by a circular tray of wire gauze (c), supported from the floor, and attached to the sides of the chamber by a ring of plasticine. If wireworms are not provided with some form of contact, their movements may be abnormally rapid and sustained, and under these conditions the sharpness of the reactions at certain steep humidity gradients is considerably reduced; in order to remedy this defect, several layers of glass beads (average diameter about 0-25 cm.) were introduced into the gauze tray (d). This proved to be a very satisfactory d A 10 cm. k- C Fig. 1. Apparatus used for investigating the humidity behaviour of Agriotes. A, large, and B, small alternative chamber in section; C, plan and section of alternative 'burrow'. Further explanation in text. means of 'damping' their movements; wireworms placed on top of the beads immediately burrowed into them, and then crawled for the most part between the gauze and the beads. The humidities on the two sides of the chamber were recorded by means of Edney paper hygro- meters (e) placed on top of the beads. As it was impossible to view the woreworms con- veniently when they were crawling below the beads, the procedure adopted was to count the numbers of larvae on either side at the end of the experiment. Usually twenty-five larvae were used simultaneously in the chamber. If, however, the larvae were ad-
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of ON THE BEHAVIOUR OF WIREWORMS OF THE GENUS ...

[ 4 3 ]
ON THE BEHAVIOUR OF WIREWORMS OF THE GENUSAGRIOTES ESCH. (COLEOPTERA, ELATERIDAE)
I. REACTIONS TO HUMIDITY
BY A. D. LEES, From the Zoological Laboratory, Cambridge
{Received 14 July 1943)
(With Six Text-figures)
Humidity reactions have recently been demon-strated in a number of terrestrial arthropods. Inseveral instances—for example in Locusta tmgratoria(Kennedy, 1937), Culex fatigans (Thomson, 1938),Blatta orientalis (Gunn & Cosway, 1938), Tenebriomolitor (Pielou & Gunn, 1940) and in Pediculushumanus corporis (Wigglesworth, 1941)—it has beenshown that such responses lead to the avoidance ofmoist air, a type of behaviour which is not easilyassociated with any specific physiological require-ments. On the other hand, certain animals such aswoodlice which are known to possess little capacityto resist desiccation, avoid dry, and aggregate inhumid air (Gunn, 1937; Waloff, 1941); this typeof behaviour is clearly of some adaptive significance.
Wireworms also are unable to withstand prolongeddesiccation. Thus according to the data of Subklew(1934a), large larvae weighing about 50 mg. loseup to half their body weight in 3 days when intro-duced into still, half-saturated air at 200 C. With thisin mind, the humidity behaviour was tested in analternative chamber. When it became clear that anintense response was shown to certain humiditygradients, the nature of the humidity receptors andthe mechanisms displayed in orientating to humiditywere also examined. The present paper deals withthese aspects of the humidity behaviour and theirrelation to the humidity conditions prevailing in thenormal environment, the soil.
METHODS AND MATERIAL
The apparatus used for testing the general humiditybehaviour of wireworms was substantially the sameas the 'alternative chamber' described by Gunn &Kennedy (1936). It consisted of a circular glassvessel, 22 cm. in diameter and 6 cm. deep, whichwas closed by an airtight glass roof (Fig. 1 A). Inorder to reduce slightly the area of contact betweenthe two halves of the chamber, a vertical partition (a),2 cm. deep, was attached to the roof. The desiredhumidities were maintained by means of sulphuricacid-water mixtures (Wilson, 1921) contained indishes (6) on the floor. The arena was provided bya circular tray of wire gauze (c), supported from the
floor, and attached to the sides of the chamber bya ring of plasticine.
If wireworms are not provided with some formof contact, their movements may be abnormallyrapid and sustained, and under these conditions thesharpness of the reactions at certain steep humiditygradients is considerably reduced; in order to remedythis defect, several layers of glass beads (averagediameter about 0-25 cm.) were introduced into thegauze tray (d). This proved to be a very satisfactory
d A
10 cm.k-
C
Fig. 1. Apparatus used for investigating the humiditybehaviour of Agriotes. A, large, and B, small alternativechamber in section; C, plan and section of alternative'burrow'. Further explanation in text.
means of 'damping' their movements; wirewormsplaced on top of the beads immediately burrowedinto them, and then crawled for the most partbetween the gauze and the beads.
The humidities on the two sides of the chamberwere recorded by means of Edney paper hygro-meters (e) placed on top of the beads.
As it was impossible to view the woreworms con-veniently when they were crawling below the beads,the procedure adopted was to count the numbers oflarvae on either side at the end of the experiment.Usually twenty-five larvae were used simultaneouslyin the chamber. If, however, the larvae were ad-

44 A. D. LEES
mitted through a hole in the roof after the humiditygradient had become established, they never becameproperly dispersed and usually remained in a clusterin the centre of the arena. To avoid this difficulty,the larvae were scattered in the 'middle zone' (thiswas a narrow region extending across the centre ofthe chamber and corresponding to the steepest partof the gradient; animals recorded from this zonecould not properly be assigned to either side); thelid was then replaced and the gradient was allowedto develop with the larvae inside. Controls showedthat no appreciable bias was involved in this pro-cedure. With a humidity range within the chamberof 90-100% R.H. the length of each experiment was6 hr.; with lower humidities, 3 hr. This representeda compromise between the effects of the inactivityof the larvae at high humidities and the activity atlower humidities; the latter sometimes led to theclustering of the larvae if the experiment was tooprotracted. If such an aggregation did occur theexperiment was rejected. The different lengths ofthe experiments are unlikely to have any effect onthe intensity of the reaction for, once dispersal hasbeen achieved, there is evidently an equilibriumbetween the numbers on the two sides which ismaintained by the humidity reaction: increasing thelength of tHe experiment will not materially affectthis relationship. The humidity gradient was usuallyfully established within 2 hr.; it may therefore beassumed that the larvae were in fact reacting to thefinal humidities recorded by the hygrometers.
As an estimate of the intensity of the reaction the
excess percentage ratio 100 (——— J (Gunn &
Cosway, 1938) was employed. In this expressionW and D are the numbers of animals in the 'wet'and ' dry' sides respectively. The W/D ratio (Gunn,1937) has sometimes been calculated for purposes ofcomparison. In the first expression the theoreticalvalue for no reaction is o-o%, in the latter i-o.The small numbers of animals recorded from themiddle zone are omitted from the calculations. Eachexperiment was repeated eight times; altogether,therefore, 200 position records were obtained foreach pair of humidities tested. Controls in- whichthe humidity of the chamber was constant through-out were kept for each experiment.
For the analysis of the mechanisms of the humidityreaction single larvae were observed in a smalleralternative chamber (Fig. 1 B). This consisted of aglass ring (/) made from a ground-down Petri dish;it was covered by a glass lid and stood on a falsefloor of muslin stretched between a wire ring (g)(Wigglesworth, 1941). The required humidities weremaintained by means of filter papers soaked invarious saturated salt solutions (Spencer, 1926);these filter papers (h) were placed below the muslinarena in the cover of a Petri dish selected as havingthe same diameter as the glass ring. The muslinwas covered by only three layers of beads and the
apparatus was illuminated from above. Tracks ^individual larvae were recorded by tracing them iink,on the lid; these were subsequently transferredto paper.
In addition, observations were made on singlewireworms crawling in 'burrows' (Fig. 1 C) con-structed in the following manner. Hollow straws ofuniform diameter were split in two longitudinally,and the channels thus formed were closed by stripsof perforated cellophane (t) which were cemented tothe cut edges of the straw. Each ' burrow' could bepassed through holes in three waxed corks (j) whichformed equidistant partitions in a glass tube. Eachhalf of the burrow was then surrounded by a moreor less isolated jacket into which strips of filter paper(k), soaked in the required salt solutions, could beintroduced. A wireworm could then be inserted intoone of the open ends of the burrow and its behaviournoted, as it crossed the steep humidity gradient intothe opposite side.
The wireworms used in these studies were collectedin batches in the field, and were then kept in jars ofmoist sand provided with potato slices until requiredfor experiment. The most abundant species in thesecollections were Agriotes obscurus L. and A. lineatus L.,and these formed the material for this research; thelarvae of these species cannot be separated on mor-phological grounds; they were distinguished fromthe closely related species A. sputator L. by thediagnostic features given by Gu^niat (1934). Larvaeranging in length from 5 to 20 mm. were tested fortheir humidity behaviour, but no essential differenceswere noted between the various size categories whichmust in fact have included several instars. Inpractice, wireworms about 10-15 mm- m lengthwere most commonly used.
RESULTS
The direction, range and intensity of the reaction
The results of six typical experiments are shown inTable 1. The average mean excess percentage of allthe controls was i-8%, which closely approximatedthe theoretical value for no reaction. Twice thestandard deviation was 12-2%; intensities greaterthan this can then be taken as indicating a significantreaction.
An inspection of Table 1 shows that under certainconditions Agriotes reacts intensely to humidity; forexample, with alternative humidities of 99-5 and-92% R.H., 190 larvae were recovered from the wetside and none from the dry side; the remaining tenposition records were from the middle zone. Thiscomplete avoidance of the dry side gives, an excesspercentage on the wet side of ioo%, and a W/Dratio of infinity. Smaller humidity differences withinapproximately the same range yield decreasing in-tensities of reaction, but the response is still apparentwith differences too minute for the hygrometers torecord accurately: thus a significant response
On the behaviour of vrireworms 45
rned with humidities of about ioo and 99-5 . %(the equilibrium humidities of the controlling
solutions were in this case 99 and 100 % R.H.). Theseresults are shown diagrammatically in Fig. 2.-
It is also clear that within this small humidityrange (90-100% R.H.), the intensity depends on theavailable humidity differences and not on anyparticular humidity within this range. Thus, in anexample included in Table 1, the excess percentage
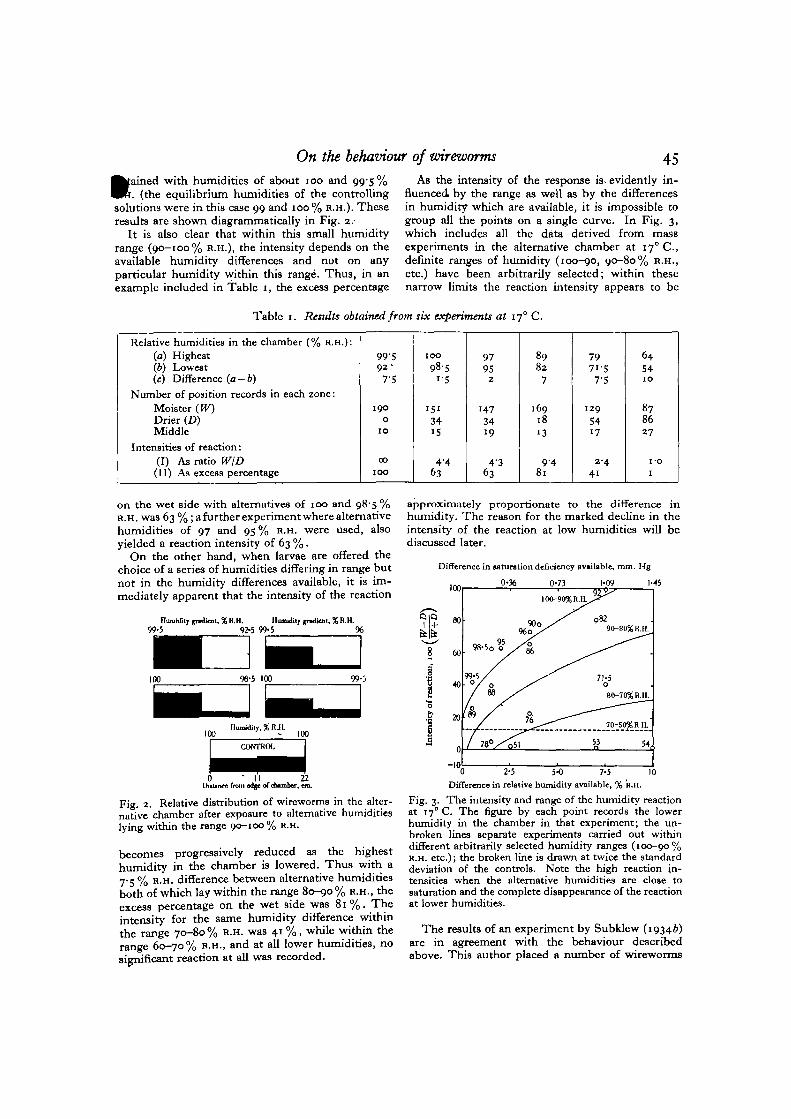
As the intensity of the response is. evidently in-fluenced by the range as well as by the differencesin humidity which are available, it is impossible togroup all the points on a single curve. In Fig. 3,which includes all the data derived from massexperiments in the alternative chamber at 170 C ,definite ranges of humidity (100-90, 90-80% R.H.,etc.) have been arbitrarily selected; within thesenarrow limits the reaction intensity appears to be
Table 1. Results obtained from six experiments at 170 C.
Relative humidities in the chamber (% R.H.):(a) Highest(6) Lowest(e) Difference (a-b)
Number of position records in each zone:Moister (W)Drier (D)Middle
Intensities of reaction:(I) As ratio W/D(II) As excesB percentage
99-59 2 -
7-5
1900
1 0
00
IOO
IOO
985I'S
I S 1
3415
4-463
9795
2
1473419
4'363
8982
7
16918
13
9'481
797i-S
7'5
129
5417
2 4
4 i
64541 0
878627
1 01
on the wet side with alternatives of 100 and 98-5%R.H. was 63 % ; a further experiment where alternativehumidities of 97 and 95 % R.H. were used, alsoyielded a reaction intensity of 63 %.
On the other hand, when larvae are offered thechoice of a series of humidities differing in range butnot in the humidity differences available, it is im-mediately apparent that the intensity of the reaction
Rurah% gradient, % R.H. Humidity gradient, % R.H.99-5 92-5 99-5 96
approximately proportionate to the difference inhumidity. The reason for the marked decline in theintensity of the reaction at low humidities will bediscussed later.
Difference in saturation deficiency available, mm. Hg
m 0 - 3 6 0-73 1-09 1-45
80
60
4099-5
0.
7
*5o
088
/
78°,
90o960/
° / 6 6
100-90% R . R ^ ^ ' ^
/ ^ o82/ 90-80% R.H.
" 71-5o80-70% R.EI.
' 70-50% RH.
5r? 54J
II 22o from edge of chamber, em.
Fig. 2. Relative distribution of wireworms in the alter-native chamber after exposure to alternative humiditieslying within the range 90-100% R.H.
becomes progressively reduced as the highesthumidity in the chamber is lowered. Thus with a7-5 % R.H. difference between alternative humiditiesboth of which lay within the range 80-90 % R.H., theexcess percentage on the wet side was 81%. Theintensity for the same humidity difference withinthe range 70-80% R.H. was 41 %, while within therange 60-70% R.H., and at all lower humidities, nosignificant reaction at all was recorded.
-100 2-5 5-0 7-5 10
Difference in relative humidity available, % R.H.
Fig. 3. The intensity and range of the humidity reactionat 170 C. The figure by each point records the lowerhumidity in the chamber in that experiment; the un-broken lines separate experiments carried out withindifferent arbitrarily selected humidity ranges (100-90%R.H. etc.); the broken line is drawn at twice the standarddeviation of the controls. Note the high reaction in-tensities when the alternative humidities are close tosaturation and the complete disappearance of the reactionat lower humidities.
The results of an experiment by Subklew (1934 ft)are in agreement with the behaviour describedabove. This author placed a number of wireworms

46 A. D. LEES
in an empty tube in which a humidity gradient wasmaintained by means of moist cotton-wooi at oneend and a drying agent at the other. After an intervalit was found that all the larvae had aggregated at thehumid end of the tube.
The effect of temperature on the reaction
The intensity of the humidity reaction at differenttemperatures has been investigated in Culex fatigans(Thomson, 1938) and in Tenebrio molitor (Pielou &Gunn, 1940). In both cases it was found that thereaction intensity was more closely related to therelative humidity than to the saturation deficiency,or evaporating power, of the air.
The behaviour of Agriotes was tested at threetemperatures, 11, 17 and 250 C, using the alter-native chamber in the same •manner as previously.Sulphuric acid solutions maintaining alternativerelative humidities of 995 and 92% R-H., 99'5 and96% R.H., and 100 and 98-5 % R.H. were employed.The hygrometer readings showed that the differencebetween any of the three pairs of alternativehumidities was the same at all three temperatures:on the other hand, the difference in saturationdeficiency was of course greatest at the highesttemperature.
In Fig. 4 the excess percentages given by theseexperiments are plotted against the available differ-
100
90
60: /
/o
'• B
• 110c •
0 i7»c
X 25°C
Difference in relative humidity Difference in saturation defi-available, % R.H. ciency available, mm. Hg
Fig. 4. The intensity of the reaction at different tem-peratures and humidities. The higher humidity of eachpair of alternativea used was nearly 100% R.H. In A,reaction intensities are plotted against the availabledifferences in relative humidity, and in B, against thedifferences in saturation deficiency.
ences in relative humidity (A), and against thecalculated differences in saturation deficiency (B).It will be clear that the scatter is less when saturationdeficiency is used as the measure of humidity. Theseobservations are then consistent with the view thatthe intensity of the humidity reaction in Agriotes isdependent on the evaporation of water from somesite on the body; the attempts to localize this regionare described in the next section.
It should be noted that the activity of the ^was not noticeably reduced at i i° C.; low reactiointensities at this temperature cannot then be dueto the inactivity of the wireworms.
The humidity ' receptors'In finding the position of the humidity receptors,
advantage was taken of the intense reaction shownby larvae to alternative humidities of 92 and 99-5 %R.H. In a preliminary series of experiments the pitson the 9th abdominal segment were covered withshellac; twenty-five larvae with the pits thus coveredwere used simultaneously in the alternative chamber,ico position records obtained with these wirewormsgave an excess percentage on the wet side of 94 %.From this it -would appear that these structures, forwhich various possible functions have been proposed(Langenbuch, 1932), are not concerned with theresponse to humidity.
Further possible sites of the receptors includesome or all of the head appendages (antennae,maxillary and labial palps), or the whole integu-mental surface. These possibilities were tested withtwo batches of twenty-five larvae; in one batch theabdomen of each larva was smeared with vaseline;in the other, the head and mouthparts were similarlytreated. This did not noticeably interfere with normallocomotion, for the elongated shape of the larvamade possible the thorough covering of the abdomen,comprising the greater part of the body surface,without entangling the legs. Both batches were thentested four times in the alternative chamber.
The larvae with the abdomen covered gave anexcess percentage on the wet side of 83 %, whilethose with the head covered yielded a reaction in-tensity of only 22 %. The response then, if noteliminated, was at least considerably reduced bygreasing the mouthparts, while covering the greaterpart of the integumental surface had comparativelylittle effect. These results seemed to indicate thatthe receptors lay on the head, and that loss of waterfrom the whole integument was not an essentialfactor in the evocation of the reaction.
In order to localize the receptors more precisely,groups of wireworms, deprived of some or all of theirhead appendages, were tested in the same humiditygradient. It has been recognized that amputationmay affect the behaviour in other ways than byremoving the specific receptors which are beingtested (Pielou, 1940). Thus, the removal of theantenna (Wigglesworth & Gillett, 1934) may giverise to a state of lowered activity which will naturallyreduce the intensity of the response to a particularstimulus. In Agriotes apparently normal activitywas retained, even after the removal of all the headappendages. In addition, such operations mayimpair the general health of the insect. In thisrespect it may perhaps be mentioned that wirewormsdeprived of all the appendages survived in a healthystate for several months; in the course of the.

On the behaviour of zoireworms 47eriments one larva moulted and regenerated trie organs, which were removed bilaterally, were am-: organs. putated in the following combinations, each in
The relevant mouthparts of Agriotes are shown in batches of twenty-five larvae: (i) antennae, (2) maxil-Fig. 5. The basal components of the maxilla andlabium are partially fused, and form a single
lary palps, (3) labial palps, (4) antennae and maxillarypalps, (5) antennae and labial palps, (6) maxillary
functional unit which can be protracted and retracted and labial palps, and (7) antennae, maxillary and
Fig. 5. The head appendages concerned with the response to humidity in Agriotes. A, maxilla and labium;B, antenna. The figures indicate the sites of the various amputations. Note the campaniform organs (shown ascircles) and the unsclerotized regions.
Table 2. 27K; intensity of the humidity reaction in larvae toith various combinationsof bilaterally amputated appendages
Organs amputatedExcess percentageOrgans amputatedExcess percentageOrgans amputated
Excess percentageOrgans amputated
Excess percentage
Antennae79 ± 4 2Antennae and maxillary
palps38 ± 6 0
None1 0 0
Maxillary palps92 ±2-3Antennae and labial
palps65±4'iAntennae, maxillary
and labial palps— 4± I2'I
Labial palpsoo±3-oMaxillary and labial
palps9i±3"i
by muscles inserted on the dorsal surface. The labialpalps are borne by the stout ligula. The fleshy,two-segmented galeae arise near the insertion of themaxillary palps, while the lacinia is plate-like and isclosely associated with the 'brush'. The diminutiveantennae are also two-segmented structures.
The removal of whole organs was accomplishedby inserting a finely pointed needle through thebasal intersegmental membrane. Before the opera-tion the wireworms were lightly etherized. Whole
labial palps. For the exact sites of the operationsreference may be made to Fig. 5; in all cases thegalea was removed with the maxillary palps and theligula with the labial palps. After a few days hadbeen allowed for recovery, each batch was testedeight times in the 92-99-5 % R.H. gradient at 170 C ,so that 200 position records were obtained fromlarvae of each category. The results are shown inTable 2, where reaction intensities are again ex-pressed as excess percentages on the wet side.

A. D. LEES
An inspection of Table 2 shows that the bilateralamputation of either antennae, maxillary palps orlabial palps only reduces the intensity by a smallfraction; no one pair of these organs is then solelyconcerned with the response. Removal of theantennae has more effect in reducing the intensitythan the removal of the other appendages. On thewhole, however, in each of these cases avoidance ofthe lower humidity is clearly shown; indeed, itappears to be almost uninfluenced by the absenceof maxillary or labial palps. If two pairs of appen-dages are removed, the intensity of the reactiondiminishes more markedly: thus when the antennaealone were left intact there was still a very intenseresponse (91 %), but when, on the other hand, themaxillary palps and the labial palps were the soleremaining organs, excess percentages of only 65 and38 were recorded. These experiments show that eachof the three pairs of appendages is concerned withthe humidity reaction, though not to the same extent.Finally, when all the organs are amputated simul-taneously the reaction is entirely eliminated (excesspercentage of — 4). Evidently then these appendageseither bear the only sensilla concerned with thehumidity reaction, or in themselves constitute thesole ' receptors'.
The temperature experiments have shown that theintensity of the reaction can be more closely relatedto the saturation deficiency of the air than to itsrelative humidity: that is to say, the reaction probablydepends on water loss rather than on any hygrometermechanism. If this is so, and the site of evaporationis on the head appendages, it might be possible tocorrelate the surface areas exposed by the appen-dages with their respective contributions to thereaction intensity. The maxillary palps and antennaeare, in fact, roughly equal in volume and surfacearea, and both are considerably larger than the labialpalps. This can perhaps be partly correlated with thepresent results, for the labial palps alone give a muchless intense response than the maxillary palps orantennae alone. On the other hand, the relativelygreater importance of the antennae than themaxillary palps cannot be explained along these lines.
In order to see whether this hypothesis could bereconciled with their morphology, the mouthpartswere examined in whole mounts and sections, as wellas in the living animal under a binocular microscope.In the living animal the antennae, maxillary andlabial palps present a rather uniform appearance:each is made up of a small number of annuli ofrelatively thin integument separated from one anotherby areas of colourless membrane. Particularly con-spicuous is the broad cushion of unsclerotizedcuticle on which the base of each appendage isinserted. It is possible that, these regions in par-ticular may be associated with rapid water loss. Inlife the haemolymph in these organs is kept undera slight positive pressure by the contraction of thethoracic musculature, and amputation is usually
accompanied,by a gush of blood; inanimal when the musculature is relaxed thismore rarely.
While it is not intended to describe in detail thesensilla on the head appendages, a brief considerationof the main types and their distribution is necessaryin order that they may be evaluated as possiblehygro-receptors. The sensilla are as follows: (a) Cam-paruform organs. These are distributed on the dorsalsurfaces of the antenna, galea, ligula, maxillary andlabial palps; a few are found more basally (Fig. 5).In unstained whole mounts these sensilla appear asconspicuous circular areas, and were described byHawkins (1936) as 'round pit-like sensory pores'.(b) Peg organs. A few of these sensilla, seldom more.than a dozen, occur on the ventral surfaces of thelabial and maxillary palps and the galea; they areabsent from the antennae, (c) Tactile bristles. Theseare found on all the head appendages; they are forthe most part thick-walled although their structureis somewhat variable. Certain other structures,possibly semsory in function, include the following:(rf) the numerous minute projections borne by thedistal segments of the labial and maxillary palps,and («) the cup-shaped and apparently perforatestructure carried by the distal joint of the antenna.
It will be noted that apart from thick-walledbristles, the only organs found on antennae,maxillary and labial palps are the campaniftjrmsensilla, and these are unlikely to respond to anyother than mechanical stimuli (Pringle, 1938). Inview of the probable importance of the peg organsas hygro-receptors in Tenebrio (Pieloa, 1940), it is ofinterest that in Agriotes these sensilla cannot functionin a similar manner, for, as has been shown above,there is still an intense residual response when onlythe antennae, which do not bear peg organs, are leftintact. Since, also, (d) is confined to the maxillaryand labial palps, and (e) to the antenna, thesestructures may also be ruled out as possible humidityreceptors.
The uniformity in the appearance and structureof the head appendages as well as the negativeevidence of the lack of any specific sensilla, lendssupport to the hypothesis that the loss of water fromthe head appendages initiates the humidity reaction.The sites of evaporation cannot be precisely de-limited, but they are likely to include the wholesurface of the appendages and perhaps, in particular,the unsclerotized intersegmental regions.
The mechanisms of orientationIt has been shown that wireworms avoid dry, and
aggregate in moist air, but it remains to decide thenature of orientation mechanisms which enable thisresult to be achieved. This was investigated byobserving single animals in the small alternativechamber described above. Four typical tracks arereproduced in Fig. 6: each 30 sec. interval is indi-cated by a cross-bar on the track; definite halts for

On the behaviour of toirewormsperiods are shown by numbers which record
alf-minutes which elapsed before locomotionbegan again.
If larvae are exposed to uniformly saturated airtheir movements are usually slow and spasmodic(Fig. 6 A); there may be frequent halts, often ofconsiderable duration, when the larva is immobilesave for slight movements of the head. If, on theother hand, the humidity within the chamber is lowerthan 100% R.H., the rate of locomotion is greatly
49behaviour of several1 arthropods—for instance ofTenebrio (Gunn & Pielou, 1940) and PorcelUo(Gunn, 1937).
Ortho-kinesis can also be demonstrated in Agrioteiif alternative humidities are used in the chamber, thehigher humidity being saturated air and the alter-*native some lower humidity. If, as occasionallyhappens, a larva crosses the humidity gradient fromthe moist side of the chamber to the dry, the rate ofmovement, as may be judged from an inspection of
34%R.H. 100% R.H. 34% R.H. D 100% R.H.
Fig. 6. Tracks followed by single wireworms in the small alternative chamber; each track was completed in 30 min.;successive half-minute intervals are indicated by cross-bars. A, behaviour in uniformly saturated air; B-D,examples of the mechanisms of orientation to dry air; in B, the activity is seen to be greater on the dry side;in B, C and D, several instances of directed reaction* are shown; the larva enters the dry half of the arena, hesitates,and then recoils backwards into the wet half.
increased, and the animals rarely, if ever, becomeinactive. There is no evidence of adaptation to dryair; on the contrary, in unsaturated atmosphereslarvae remain in a state of violent activity until theydie from desiccation. At all humidities the rate ofchange of direction appears to be entirely at random.This type of reaction which involves a differentialrate of movement or a difference in the amount ofactivity (number of 'rests ' , etc.) when the animal isexposed to two intensities of a stimulus, has beentermed an ortho-kinesis (Gunn et al. 1937): thismechanism forma an integral part of the humidity
JBB.ao, I
the tracks, instantly increases (Fig. 6 B). In the typeof apparatus used here the animal is then rapidlybrought back to the moister half of the chamber.On re-entering saturated air, the rate of locomotionusually decreases within a few minutes, the changein rate, however, seeming to be less rapid than whenthe larva emerges from moist into dry air.
There is also a further very typical orientationmechanism which is usually clearly exhibited if thealternative humidity to saturated air is rather low(e.g. 34% K.H., a saturated solution of.MgCl! beingused). On approaching the humidity boundary the

A. D. LEES
larva hesitates, moves its head from side to side, andfinally recoils backwards into the moist side.Fig. 6 C, D shows several examples of this type ofbehaviour; the larva has sometimes made shortexcursions into the lower humidity but has theninvariably retreated along the same track, afterwardsmoving off in a different direction. This strikingreaction can be seen to even better advantage in theartificial 'burrow' described on p. 44. If a wire-worm, selected so that its diameter is just accom-modated by the burrow, is inserted into the moisthalf, it will gradually move up to the junction withthe dry half; it then stops, manipulates its headappendages, and then retreats backwards into thehumid section of the burrow. The distance of recoilmay be very considerable, and wireworms frequentlyejected themselves with some violence from the endof the tube (a distance of 10 cm. in the burrowsused). This peculiar type of locomotion, no doubta characteristic response to unfavourable stimuli ingeneral, has frequently been observed in wirewormsmoving about in self-constructed burrows in sandor soil, particularly if they had migrated into a regionwhere the medium had obviously become rather dry.The recoil was usually sufficient to carry the larvaeout of this region.
There is no evidencenhat the turning reactions ofAgriotes are affected by humidity, nor indeed wouldthis be expected in view of the normal habitat of thelarva. This mechanism is not then an example ofklino-kinesis even in its extreme form (see Gunn &Walshe, 1941; Wigglesworth, 1941). The orientationis in fact a directed one, the interesting feature inthis case being that under natural conditions it isnecessarily so since the direction of movement isrestricted by the limits of the burrow. Two types ofdirected orientation (taxes) have been distinguished.In the first place, an animal behaving klino-tactically' compares' in successive intervals of time theintensity of the stimulus to which it is reacting;secondly, an animal exhibiting tropo-taxis comparesthe two intensities of the stimulus simultaneously bymeans of bilaterally placed sense organs (Fraenkel& Gunn, 1940). As a mechanism of reacting to thediffuse stimulus of humidity, tropo-taxis is at leasttheoretically possible although it has not yet beendemonstrated in any insect. One means of dis-tinguishing between these two mechanisms is toeliminate the receptors on one side; many animalsbehaving tropo-tactically will then show circusmovements.
In order to examine this point, the head appen-dages which, as has been shown above, constitutethe humidity 'receptors' were eliminated on oneside. In \ batch of twenty-five larvae the left antennaand left maxillary palp were amputated. These wire-worms were tested individually in the small alter-native chamber and showed no tendency to performcircus movements. They were also subjected to massexperiments in the large alternative chamber where
alternative humidities of 92 and 99 % R.H.maintained. The unilateral removal of the rhad no effect on the intensity of the reaction, for outof a total of seventy-five position records seventy-onewere from the moist side while only one larva wasrecovered from the dry side. Had circus movementsbeen involved they would probably have beenrevealed by an increased number of animals on thedry side. It may be concluded therefore that thisorientation is not tropo-tactic but is probably klino-tactic. It may also be pointed out in this connexionthat in Agriotes, as in all wireworms, the headappendages are extremely short, and it is difficultto believe that they could be affected bilaterally, butdifferentially, by any gradient of humidity howeversteep.
This analysis of the mechanisms of orientationthrows some light on the results obtained from themass experiments in the large alternative chamber,although it is unfortunately impossible to decidewith any precision the relative contribution made byeach type of mechanism to the general humiditybehaviour. One of the most striking features ofthese results was the very complete reaction at theextreme moist end of the humidity range (100% ofthe larvae avoiding 92 % R.H. and aggregating in99'S % R.H.). In the light of the mechanismsdescribed above it seems evident that the clear-cutseparation brought about by these humidities is to agreat extent caused by the operation of the klino-tactic mechanism. This reaction is no doubt veryeffective in causing the avoidance of the lowerhumidity if the larva happens to cross the boundaryfrom the high into the low humidity. At the sametime it seems probable that the low hygro-kinesiswhich results in the partial immobilization of thelarvae on the moist side ('virtual immobility'; Gunn& Pielou, 1940) is a very important contributoryfactor, and this was certainly the impression gainedwhen the larvae were being removed from the moisthalf of the chamber at the end of an experiment.
Fig. 3 also indicates, however, that as the humidityrange in the alternative chamber is lowered, theintensity of the response to similar differences inhumidity declines, until within the general humidityrange of 50-70% R.H., for instance, there is nosignificant reaction to differences as large as 15%R.H. The reason for this is probably quite simple.At low humidities the larvae are always intenselyactive, and as a result, those which are crossing thearena are rapidly carried over the steepest part ofthe humidity gradient into the lower, thoughrelatively uniform, humidity in the opposite side ofthe chamber. This has the effect of virtuallyeliminating the avoiding mechanism which is so wellshown at higher humidities. It may be added as acorollary that the rate of movement is an importantfactor in the normal operation of this klino-tacticmechanism. Lack of contact, which also leads to anincreased rate of movement, exercises a similar

On the behaviour of wirewormseffect on the reaction: but the complete
of the reaction at low humidities mustalso indicate that the rate of movement is not in anysimple way proportionate to the relative humidity orsaturation deficiency; that is to say, it is probablethat the differences in the rates of movementbetween larvae in uniform humidities of ioo and90% R.H. is much greater than that betweenhumidities of for instance, 60 and 70% R.H. Thispoint has not been examined experimentally.
DISCUSSION
It has been shown that wireworms react strongly tocertain gradients of humidity. If some allowance ismade for the slightly different form and mode ofoperation of the apparatus used to test this be-haviour, it would appear that Agriotes avoids dryair with an intensity roughly equal to the intensityof avoidance of moist air by Tenebrio (Pielou & Gunn,1940) or by Culex (Thomson, 1938). Over the rangeof humidities where the intense reaction is given,differences in saturation deficiency of 1 mm. Hg, orin relative humidity of 7-5% R.H. at 170 C. over a.22 cm. gradient, are sufficient to bring about a suc-cessful avoiding reaction by nearly every individual,while statistically significant reactions are given bymuch smaller humidity differences. But in Agriotesas in Tenebrio and Culex the intense reaction onlyoperates over a small part of the whole humidityrange: in the first insect, if one alternative is saturatedair, the reaction leads to the avoidance of anyhumidity which falls short of saturation, while inTenebrio, Culex and Pediculus (Wigglesworth, 1941),only high humidities are avoided, the insects ap-pearing more or less indifferent to lower humidities.In a sense it would seem that Agriotes also is in-different to large humidity differences if the valuesfor the alternative humidities are under 70% R.H.It has been suggested, however, that the failure ofthe reaction is due rather to the elimination of anintegral part of the orientation mechanism underunfavourable physiological conditions rather than toany real' indifference' to humidity as a factor capableof influencing behaviour. In some respects thegeneral behaviour of Agriotes most closely resemblesthat of Porcellio. Thus Gunn (1937) has shown thatwoodlice avoid dry air; at the same time his workindicated that the region of maximum sensitivity tosmall humidity differences lay in the region of50% R.H. Waloff (1941) has, however, demon-strated that, under certain conditions at least,woodlice exhibit an intense reaction at the extrememo.ist end of the humidity scale, for instance, withalternatives of 86 and 100% R.H.
An interesting feature of the humidity reactionsof terrestrial arthropods is their association with twofundamentally different types of receptor apparatus.These might be described as ' hygrometer' and'evaporimeter' receptors respectively. The first
5 1
depends on the presence of specific sensilla capableof functioning hygrometrically; the second type of' receptor' appears to depend in the first instance onthe evaporation - of water either from a localizedregion of the body or from the whole integument.Apart from verifying the presence or absence of therelevant sensilla, the only experimental methodavailable for deciding between these alternativesinvolves the examination of the reaction intensityat different temperatures and humidities—a methodwhich assumes that the intensity remains approxi-mately the same at the various uniform temperaturesemployed. In Agriotes, experiments at three tem-peratures have indicated that the intensity of thereaction is related more closely to the differences insaturation deficiency than to the differences inrelative humidity offered in the alternative chamber.This is suggestive of the 'evaporimeter' receptor.On the other hand, similar experiments have shownthat in Culex (Thomson, 1938) and Tenebrio (Pielou& Gunn, 1940) the reverse is true.
With'regard to the actual 'receptors', it appearsthat in Agriotes the response is dependent on theintegrity of the head appendages, and it has provedimpossible to associate it with any particular type of8ensillum. In Porcellio, Gunn was also unable tolocalize the position of the hygro-receptors, andWaloff has suggested that loss of water from theintegument and the consequent concentration ofthe body fluids may initiate the reaction. The latterauthor also proposes an alternative explanation,namely, that a secondary proprioceptive mechanismmay be involved. Although in Agriotes all the headappendages concerned with the reaction are wellsupplied with campanifbrm sensilla, it is difficult tobelieve that evaporation could stimulate these organsin any way, for even extreme desiccation causes novisible shrinkage. The rapidity with which thereaction develops suggests that local changes inosmotic pressure may perhaps be sufficient tostimulate the peripheral sensory nerves directly.
This type of 'evaporimeter' receptor contrastswith the 'hygrometer' receptor occurring, forexample, in Tenebrio (Pielou, 1940), where thenumerous peg and pit peg organs are the sensillaconcerned with the response. In mosquitoes asimilar function has been proposed for the fine hairsof the antenna (Necheles, 1925), while in Pediculusthe reaction has been shown to depend on thepresence of the ' tuft organs'—sensilla whosestructure also suggests that they may functionhygroscopically (Wigglesworth, 1941).
It may be noted that while the 'hygrometer'receptor might conceivably be associated' withreactions in either direction, the ' evaporimeter' typeis only likely to function successfully if, by theoperation of the receptor, dry air is avoided. It isnot surprising then that Tenebrio, Culex andPediculus, all of which avoid moist air, appear topossess the former type of receptor.
4-i

A. D. LEES
The nature of the humidity receptors may alsoinfluence the mechanisms displayed in orientatingto humidity. Thus in klino-kinesis, which involvesthe random turning movements of the animal,sensory adaptation is an essential adjunct; but, asWaloff (1941) has pointed-out, adaptation to a lowhumidity is unlikely to take place in an animal whichis consistently losing water, or from which appro-priate sensilla are absent. In Agriotes klino-kinesisis in fact absent, although the burrowing habit ofthe larva would perhaps exclude this mechanism inany case. In Porcellio also typical klino-kinesis seemsto be lacking. This is in direct contrast to theimportance of this mechanism in the humiditybehaviour of Tenebrio (Gunn & Pielou, 1940) andPediculus (Wigglesworth, 1941).
If, as seems probable, the humidity responses ofAgriotes are set in train by the evaporation of waterfrom the head appendages, some suggestions can beoffered as to the factors leading to the operation ofthe orientation mechanisms. It has been noted abovethat wireworms in unsaturated air are always active.It seems reasonable to suppose that this kinesis ismaintained by a relatively constant rate of evapora-tion from the appendages. A larva crossing ahumidity gradient into drier air will, on the otherhand, be subjected to a rapid increase in the rate ofwater loss. An abrupt change of this sort may serveto evoke the second type of mechanism described,namely, the sudden recoil from dry air. As, however,desiccation has no effect on the intensity of theresponse, both mechanisms probably 4ePend on therates of evaporation rather than on the level ofconcentration, whether local or general, of the bodyBuid.
The recoiling mechanism may be classified as aklino-taxi8 for it seems to involve a comparison ofintensities (or rates of evaporation) at successiveintervals of time; also, the head appendages con-stitute collectively what is in effect a single receptor.At the same time, the response is more simple thanthat shown by, for instance, a maggot orientating tolight—a typical example of klino-taxis (Fraenkel& Gunn, 1940)—where, in order to achieve orienta-tion, a succession of such comparisons is made. InAgriotes the whole reaction comes into play after acertain threshold of stimulation is exceeded. For thismechanism the term 'shock reaction' (Mast, 1938),if unsatisfactory in some respects, is certainly apt.
Humidity behaviour and the normal environmentThe inability of wireworms to withstand desicca-
tion has already been mentioned. This has b£enshown by Subklew (1934a) to be due to the excep-tional permeability of the cuticle to water. The rapidpassage of water-soluble arsenicals through theintegument (Woodworth, 1938) also points to thesame conclusion.
Now according to the recent data of Evans (1943),the osmotic pressure of the haemolymph of Agriotes
is approximately equivalent to that of a 0-3solution. As the vapour-pressure depression ofa solution is o-6% and the relative humidity about99-4% R.H., wireworms exposed to humidities lowerthan this will lose water by evaporation, and—'-whatis important in this context—the rate of water lossthrough the integument will be relatively high incomparison with the rate of loss in insects possessingless permeable cuticles.
It is necessary therefore to inquire whether thereare- reasons for doubting the common assumptionthat the soil atmosphere is under all conditionsuniformly saturated. If so, the risk of desiccationmay be real, and the humidity behaviour may havean important, if indirect, bearing on the maintenanceof the normal water balance. Actually, substancesin solution reduce the relative humidity to someslight extent. Of greater importance, however, arethe hygroscopic properties of soils. Thus, for thewireworm, a critical state will be reached when thesoil moisture corresponds to a vapour-pressure de-pression of o-6 % , a value equivalent to a ' suction'pressure on the pF scale of 3-9 (see Evans, 1943).Such conditions are of course most likely to occurduring the summer drying out of the surface layersof the soil.
The absorbtion isotherms of the various types ofsoils given by Puri, Crowther & Keen (1925) areinstructive from several points of view. These curvesshow the typical S-shape displayed bycnany hygro-scopic materials; they indicate that if a soil is dried,comparatively large amounts of water have to beremoved to shift the equilibrium humidity from, forinstance, 99 to 90% R.H., while with further waterloss, small differences in moisture content cause arapid drop in humidity.* It follows that the saturationdeficiencies which may be expected to occur mostfrequently, during, for example, periods when thesoil is moderately dry, will be small and thereforedifficult to detect experimentally. The values givenby Puri et al. leave little doubt, however, that suchdeficiencies must often occur, although their in-cidence will clearly depend, among other factors, onclimatic conditions and the soil type. It is of interestto note in this connexion that Lebedeff (1928), usinga specially designed hair hygrometer, was able todemonstrate in the field that high ground tem-peratures were usually accompanied by measurablesaturation deficiencies, although this was only trueof the surface 10 cm. of soil.
The humidity behaviour of wireworms is evidentlycorrelated with these environmental conditions. The' preferendum' is 100% R.H., for in the presence ofsaturated air all other humidities are avoided; but itis also significant that humidities close to saturation,such as are expected to prevail in the soil, representjust that range over which the reactions are mostsensitive; indeed, the whole mechanism of theresponse is only capable of functioning successfullyin the presence of a high humidity.

JOURNAL OF EXPERIMENTAL BIOLOGY, 2o, PLATE I
20 cm.
NO IO LARVAE
LARVAE PER SLICE
10cm.
L
•%
I. 10 LARVAE PER PAPER
SATURATION PAPERS + PAPERS -f-
OF SAND WATER CARROT
-80
2.
LEES—ON THE BEHAVIOUR OF WIREWORMS OF THE GENUS AGRIOTES ESCH. (pp. 43~S3)


On the behaviour of wireworms
^ ^ t, of course, be stated with certainty thathuWRity is a limiting environmental factor, but thenumerous references in the literature to the migrationor wireworms out of the surface layers of the soilduring periods of drought (e.g. Miwa & Yanagihara,1929) are perhaps suggestive of this; doubtless otherfactors, including the effects of soil moisture (whichare discussed in the next paper (Lees, 1943)), con-tribute to this behaviour, but it is probable that theresponse to humidity plays an integral part.
SUMMARY
1. The general humidity behaviour, the humidityreceptors and the orientation mechanisms have beenstudied in the larvae of Agriotes.
2. Wireworms avoid dry air, the intensity ofavoidance being greatest when the alternatives areclose to saturation. Within this humidity range adifference of 7-5 % R.H. in the alternative chamber (at17° C.) is sufficient to ensure the successful avoidanceof the lower humidity by nearly every individual,while statistically significant reactions are obtainedwith differences as small as 0-5% R.H. At lowerranges of humidity the same differences yield pro-gressively less intense reactions; at low humiditiesthe response is entirely eliminated.
3. The intensity of the reaction is in better accordwith the humidity differences when these are ex-pressed as saturation deficiencies rather than asrelative humidities. This suggests that the reactionis initiated by the evaporation of water ('evapori-
53meter' receptor) and not by the operation of re-ceptors which function hygrometrically ('hygro-meter' receptors).
4. Trie humidity 'receptors' lie on the head.Amputation experiments indicate that the relevantsites of evaporation are distributed between theantennae, maxillary and labial'palps; the structureof these appendages is compatible with such afunction;-on the other hand, sensilla which mightconceivably have a hygroscopic function areabsent.
5. Orientation is achieved by the operation of twomechanisms. First, larvae are more active in drythan in moist air (low hygro-kinesis); secondly, larvaeshow a directed response to low humidities (klino-'taxis); this is displayed as a powerful backwardrecoil if the wireworm crosses a steep gradient frommoist into dry air. No tropo-tactic component isinvolved. The failure of the reaction at low humiditiesis explained by the great activity of the larvae whichinterferes with - the operation of the klino-tacticmechanism. It is suggested that the kinesis ismaintained by a relatively constant rate of water lossfrom the head appendages, while the klino-taxis tsinitiated by any sudden increase in the rate ofevaporation.
6. The behaviour can readily be correlated withthe humidity conditions prevailing in the soil; thenecessity for such a sensitive response is possiblydictated by the permeability of die cuticle whichrenders wireworms peculiarly liable to water loss inunsaturated atmospheres.
REFERENCESEVANS, A. C. (1943). Nature, Lond., 153, 31.FRAENKEL, G. & GUNN, D. L. (1940). The Orientation
of Anhnals. Oxford.GUENIAT, E. (1934). Mitt, tchweiz. ent. Get. 16, 167.GUNN, D. L. (1937). J. Exp. Biol. 14, 178.GUNN, D. L. & COSWAY, C. A. (1938). J. Exp. Biol. 15,
555-GUNN, D. L. & KENNEDY, J. S. (1936). J. Exp. Biol. 13,
450.GUNN, D. L., KENNEDY, J. S. & PIKLOU, D. P. (1937).
Nature, Lond., 140, 1064.GUNN, D. L. & PIELOU, D. P. (1940). J. Exp. Biol. 17,
3°7-GUNN, D. L. & WALSHE, B. M. (1941). Nature, Lond.,
148. 564-HAWKINS, J.H. (1936). Bull. Me. Agric. Exp. Sta. no. 381.KENNEDY, J. S. (1937)- 7- Exp- Biol. 14, 187.LANGENBUCH, R. (1932). Z. angew. Ent. 19, 278.LEBEDEFF, A. F. (1928). Proc. itt Int. Congr. Soil Set.,
Washington.LEES, A. D. (1943)- J- Exp. Biol. 20, 54.MAST, S. O. (1938). Biol. Rmi. 13, 186.
MIWA, Y. & YANAOIHARA, M. (1929). J. Trop. Agric.,Taihoku, 1, 275.
NECHELES, H. (1925). Arch. Schiffs- u. Tropenkyg. 39,288.
PTELOU, D. P. (1940). J. Exp. Biol. 17, 295.FIELOU, D. P. & GWNN, D. L. (1940). J. Exp. Biol. 17,
286.PRINGLE, J. W. S. (1938). J. Exp. Biol. 15, 114.PURI, A. N., CROWTHER, E. M. & KEEN, B. A. (1925).
J. Agric. Set. 15, 68.SPENCER, H. M. (1926). international Critical Tablet,
1, 67.SUBKLEW, W. (1934 a). Z. Morph. Okol. Tiere, 38, 184.SUBKLEW, Wi (19346). Z. vergl^Physiol. 31, 157.THOMSON, R. C. M. (1938). Bull. Ent. Res. 39, 125.WALOFF, N. (1941). J. Exp. Biol. 18, 115.WIGGLESWORTH, V. B. (1941).- Porasitology, 33, 67.WlGGLESWORTH, V. B. & GlLLBTT, J. D. (1934). J. Exp.
Biol. I I , 120.WILSON, R. E. (1921). J. Industr. Engng Chem. 13,
326.WOODWORTH, C. E. (1938). J. Agric. Res. 57, 229.




















