
Cost-effectiveness of rotavirus vaccination in Bolivia from the state perspective
Upload
independentCategory
view
0download
0
R
OCa
b
a
ARRAA
KARNI
1
miftiemtt
teiawabf
m
0d
Vaccine 28 (2010) 3106–3111
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
otavirus enterotoxin NSP4 has mucosal adjuvant properties
wen V. Kavanagha, Nadim J. Ajamia,b, Elly Chenga, Max Ciarleta,1, Roberto A. Guerreroa,arl Q.-Y. Zenga, Sue E. Crawforda, Mary K. Estesa,∗
Department of Molecular Virology and Microbiology, Baylor College of Medicine, One Baylor Plaza, Houston, TX 77030, United StatesCenter for Infectious Disease, School of Public Heath, University of Texas Houston Health Science Center, 1200 Herman Pressler, Houston, TX 77030, United States
r t i c l e i n f o
rticle history:eceived 30 October 2009eceived in revised form 8 February 2010ccepted 15 February 2010vailable online 1 March 2010
a b s t r a c t
Rotavirus nonstructural protein 4 (NSP4) is a protein with pleiotropic properties. It functions in rotavirusmorphogenesis, pathogenesis, and is the first described viral enterotoxin. Since many bacterial toxinsfunction as potent mucosal adjuvants, we evaluated whether baculovirus-expressed recombinant simianrotavirus SA11 NSP4 possesses adjuvant activity by co-administering NSP4 with keyhole limpet hemo-cyanin (KLH), tetanus toxoid (TT) or ovalbumin (OVA) as model antigens in mice. Following intranasal
eywords:djuvantotavirusSP4
ntranasal
immunization, NSP4 significantly enhanced both systemic and mucosal immune responses to modelimmunogens, as compared to the control group, in an antigen-specific manner. Both full-length and acleavage product of SA11 NSP4 had adjuvant activity, localizing this activity to the C-terminus of the pro-tein. NSP4 forms from virulent and avirulent porcine rotavirus OSU strain, and SA11 NSP4 localized withina 2/6-virus-like particle (VLP) also exhibited adjuvant effects. These studies suggest that the rotavirusenterotoxin NSP4 can function as an adjuvant to enhance immune responses for a co-administered
antigen.. Introduction
Many infectious pathogens come into contact with the host atucosal surfaces. Conventional parenteral vaccines are generally
neffective at eliciting mucosal immunity [1–3]. Recent efforts haveocused on the development of mucosal vaccines in an attempto combat invading pathogens at the site of contact by efficientlynducing both mucosal and systemic immune responses. How-ver, one major drawback is the intrinsic low immunogenicity ofany protein antigens when administered mucosally. Therefore,
he need for mucosal adjuvants is pivotal for development of effec-ive and safe mucosal vaccines.
The most widely studied mucosal adjuvants are the choleraoxin (CT) from Vibrio cholerae, and its close relative, the heat-labilenterotoxin (LT) from Escherichia coli. Aside from their function-ng as enterotoxins, both CT and LT have been shown to functions potent adjuvants via binding to the ganglioside GM1 receptor,
hich results in cellular activation, expression of surface moleculesnd cytokine production [4]. However, intranasal delivery of theseacterial enterotoxins may induce neurotoxic effects [5,6]. Mutantorms of cholera (mCT) and heat-labile toxin (mLT), which lack tox-
∗ Corresponding author. Tel.: +1 713 798 3585; fax: +1 713 798 3586.E-mail address: [email protected] (M.K. Estes).
1 Present address: Infectious Diseases and Vaccines – Clinical Research Depart-ent, Merck Research Laboratories, North Wales, PA 19454, United States.
264-410X/$ – see front matter © 2010 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2010.02.063
© 2010 Elsevier Ltd. All rights reserved.
icity while retaining adjuvanticity, have been described [7]. Thedevelopment of a safe, non-toxic mucosal adjuvant that can bedelivered intranasally would be an attractive alternative to bac-terial toxins.
The first described viral enterotoxin is the rotavirus nonstruc-tural protein 4 (NSP4). NSP4 is capable of inducing dose- andage-dependent diarrhea in neonatal mice without causing histo-logical alterations [8]. A cleavage product, NSP4(112–175), foundin the supernatant of rotavirus-infected cell cultures [9] can causeCa2+ mobilization in vitro and induce dose- and age-dependentdiarrhea in vivo, just like the full-length protein. Since bacterialtoxins, such as CT and LT, are well established to function aspotent mucosal adjuvants, we asked if NSP4 also possesses adju-vant activity. In this study we tested the viral enterotoxin NSP4from several virus strains for adjuvant activity in mice followingintranasal administration of classical model protein antigens andevaluated the mucosal and systemic antibody responses.
2. Materials and methods
2.1. Animals
Six- to eight-week-old inbred BALB/c female mice wereobtained from Charles River Laboratories (Wilmington, MA). Allanimals were housed in microisolator cages throughout the studyperiod as previously described [10,11].

accine
2
uvwwBtsgawteHpfiTTs
22
psw(ttctodfit
2p
fTassXt
2
st5Tp
2
5a(wv
O.V. Kavanagh et al. / V
.2. Expression and purification of recombinant NSP4
Recombinant NSP4 including the full-length and cleavage prod-ct (112–175) from simian SA11 C13, full-length NSP4 fromirulent and attenuated porcine rotavirus strains OSU-v and OSU-aere purified from Spodoptera frugiperda (Sf9) insect cells infectedith baculovirus recombinants as described previously [9,12,13].riefly, NSP4-encoding rotavirus gene 10 sequences were cloned inhe TOPO TA vector (Invitrogen Life Technologies, Chicago, IL) andubcloned into the baculovirus transfer vector pFastBAC1 (Invitro-en). Recombinant baculoviruses expressing NSP4 were generateds described by the manufacturer, and recombinant virus stocksere plaque purified. NSP4 was first semi-purified by fast pro-
ein liquid chromatography using a quaternary methylamine anionxchange column pre-equilibrated with buffer (20 mM Glycine-Cl, pH 8.1). The NSP4-rich fractions were pooled and furtherurified using an agarose immunoaffinity column onto which puri-ed anti-NSP4 (114–135) rabbit IgG had been immobilized [8].he bound NSP4 was eluted with 0.1 M Tris–HCl buffer at pH 2.8.he eluate was dialyzed against 50 mM NH4HCO3, lyophilized, andtored at 4 ◦C. Prior to use, NSP4 proteins were reconstituted in PBS.
.3. Expression and purification of NSP4-containing/6-virus-like particle (VLP)
Rotavirus 2/6-virus-like particles were expressed using com-lementary DNA sequences (cDNA) for simian rotavirus SAl1 geneegment 2, which codes VP2, and gene segment 6, which codes VP6ere made from mRNA and subcloned into pCRII TOPO TA vectors
Invitrogen). The rotavirus genes were inserted into a baculovirusransfer vector capable of co-expressing up to four different pro-eins (see below). The plasmid, pBAC4X (Novagen, San Diego, CA),ontains two polyhedron promoters and two p10 promoters withhe homologous promoters orientated in opposite directions, onef each in the left-hand direction, and the others, in the right-handirection. Each newly inserted sequence was subsequently con-rmed by restriction digestion and the cloned gene was sequencedo confirm its integrity.
.3.1. Insertion of VP6 into the pBAC4X baculovirus transferlasmid (pB4X/VP6)
The VP6 gene segment was PCR amplified from theull-length clone pSP65/SA11–6 using the sense primer 5′-CTAGAGGCCGGCCTTTTAAACG (XbaI restriction site underlined)nd the antisense primer 5′-AGGCCTGGTGAATCCTCTCAC-3′ (StuIite underlined). Cohesive ends were generated by digesting theequence with XbaI and StuI and the gene was inserted intobaI/StuI linearized baculovirus transfer plasmid pBAC4X behind
he left-hand polyhedron promoter.
.3.2. Insertion of VP2 into a cloning vector (pVP�2)A truncated form of the SA11 VP2 gene lacking the protease-
ensitive region encoding amino acid residues from the N-terminuso residue 92 (VP�2) [14] was amplified using the sense primer′-ATGGGAGGCGGAGGCGCTAACAAAACTATCC-3′ and antisense 5′-TAGGTCATATCTCCACAATGG-3′ and cloned into the TOPO TA pCRIIlasmid (pVP�2).
.3.3. Fusion of NSP4 to VP2 (pNSP4-�2)NSP4(112–175) was PCR-amplified using the 5′-ended primer
′-CCATGGTTGACAAATTGAC-3′ (NcoI restriction site underlined)nd 3′-ended primer 5′-GCTAGCTCCTCCTCCCATTGCTGCAGT-3′
NheI site underlined). The amplified DNA fragment was digestedith NcoI and NheI and ligated into NcoI/NheI linearized pVP�2
ector.
28 (2010) 3106–3111 3107
2.3.4. Consruction of baculovirus transfer plasmid encoding SA11VP6 and NSP4-VP2 (pB4X/NSP4-�2/6)
The pNSP4-�2 was digested with NotI and AvrII restrictionenzymes to remove the gene encoding the fusion protein NSP4-�2and inserted behind the second, right-hand, polyhedron promoterby ligation into pB4X/VP6 linearized by NotI and SpeI restrictionenzymes.
A recombinant baculovirus encoding the three rotavirus recom-binant proteins was generated as described by the manufacturer,and virus stocks were plaque purified. VLPs containing the SA11rotavirus proteins VP6 and fusion protein NSP4-VP2 (NSP4-2/6 VLP)were purified using CsCl2 gradients and characterized as previouslydescribed [15]. The endotoxin level in each 2/6-VLP prepara-tion was quantitated (<0.05 U/dose) using the Limulus amebocyteassay (Associates of Cape Cod, Inc., Woods Hole, MA). Electronmicroscopy was performed on each of the VLP preparations justprior to inoculation to confirm the integrity of the VLPs.
2.4. Inoculation of animals
Groups of five BALB/c mice were used to test each antigen.All experiments included a group of mice co-administered 10 �gof the mucosal adjuvant, mutant E. coli heat-labile enterotoxin[LT(R192G)] (mLT) as a immunostimulatory control [16]. Theanimals were anaesthetized by intraperitoneal administration ofketamine (3.75 mg/mouse), xylazine (0.19 mg/mouse), and acepro-mazine (0.037 mg/mouse) [10] before immunization. Two doses ofintranasal immunization of 100 �g of KLH or OVA alone or withfull-length NSP4 (6 �g) or the truncated NSP4(112–175) (10 or20 �g) were carried out three weeks apart. Tetanus toxoid used forimmunization was kindly provided by Dr. Jerry McGhee (Univer-sity of Alabama, Birmingham) or from the Statens Serum Institute(Copenhagen, Denmark). Animals were immunized intranasallywith 10 �g of TT alone or co-administered with 10 �g of eitherfull-length NSP4 or NSP4 internalized in VLPs (NSP4-2/6 VLP) threetimes, two weeks apart.
2.5. Sample collection
Serum and fecal samples were collected before vaccination (0DPI) and at 14 days post second or third immunization. Blood sam-ples were collected by tail bleed for separation of serum. Fecalsamples were collected with a fecal collection cage as previouslydescribed [17] and processed to make 20% (w/v) suspensions instool diluent as described previously [11,18]. All samples werestored at −80◦ until assayed.
2.6. Sample analysis
(i) ELISA to measure KLH- or OVA-specific serum antibodyresponses. All ELISAs were performed on 96-well polyvinylchloride microtiter plates (Dynatech, McLean, VA). Plateswere coated with 100 �l of KLH or OVA (10 �g/ml) incarbonate–bicarbonate buffer (pH 9.6) and incubated for 4 hat room temperature. Non-specific protein binding sites wereblocked with 5% BLOTTO. Following each step after the block,the plates were washed three times with 0.05% Tween 20 inPBS with an Ultrawasher Plus Platewasher (Dynatech). Serumsamples from individual animals were serially diluted two-folddown the plate in 5% BLOTTO. Samples were then incubatedfor 2 h at 37 ◦C. Horseradish peroxidase-conjugated goat anti-
mouse IgG antibody (Sigma) diluted 1:7500 in 2.5% BLOTTOwas then added to all wells and incubated for 1 h at roomtemperature. All reactions were detected using TMB MicrowellELISA substrate (Kirkegaard and Perry Laboratories, Gaithers-burg, Md.). The substrate was allowed to react for 10 min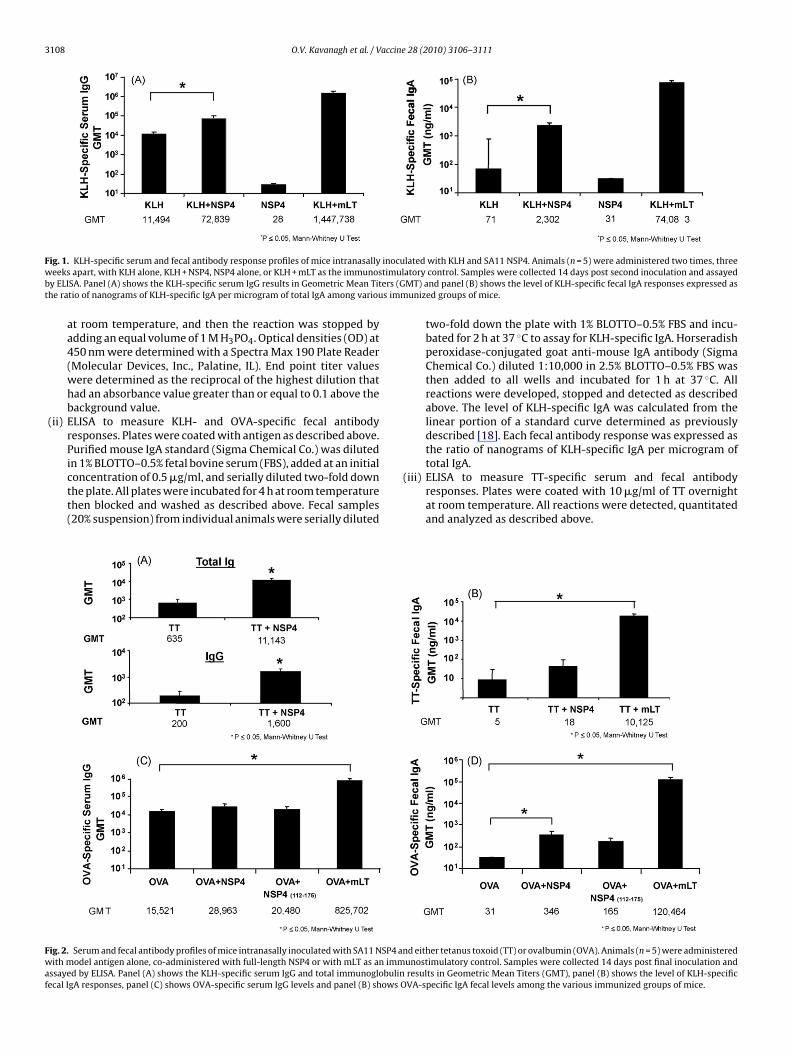
3108 O.V. Kavanagh et al. / Vaccine 28 (2010) 3106–3111
F inocuw timulb ers (Gt s imm
Fwaf
ig. 1. KLH-specific serum and fecal antibody response profiles of mice intranasallyeeks apart, with KLH alone, KLH + NSP4, NSP4 alone, or KLH + mLT as the immunos
y ELISA. Panel (A) shows the KLH-specific serum IgG results in Geometric Mean Tithe ratio of nanograms of KLH-specific IgA per microgram of total IgA among variou
at room temperature, and then the reaction was stopped byadding an equal volume of 1 M H3PO4. Optical densities (OD) at450 nm were determined with a Spectra Max 190 Plate Reader(Molecular Devices, Inc., Palatine, IL). End point titer valueswere determined as the reciprocal of the highest dilution thathad an absorbance value greater than or equal to 0.1 above thebackground value.
(ii) ELISA to measure KLH- and OVA-specific fecal antibodyresponses. Plates were coated with antigen as described above.Purified mouse IgA standard (Sigma Chemical Co.) was dilutedin 1% BLOTTO–0.5% fetal bovine serum (FBS), added at an initial
concentration of 0.5 �g/ml, and serially diluted two-fold downthe plate. All plates were incubated for 4 h at room temperaturethen blocked and washed as described above. Fecal samples(20% suspension) from individual animals were serially dilutedig. 2. Serum and fecal antibody profiles of mice intranasally inoculated with SA11 NSP4 anith model antigen alone, co-administered with full-length NSP4 or with mLT as an imm
ssayed by ELISA. Panel (A) shows the KLH-specific serum IgG and total immunoglobulinecal IgA responses, panel (C) shows OVA-specific serum IgG levels and panel (B) shows O
lated with KLH and SA11 NSP4. Animals (n = 5) were administered two times, threeatory control. Samples were collected 14 days post second inoculation and assayedMT) and panel (B) shows the level of KLH-specific fecal IgA responses expressed asunized groups of mice.
two-fold down the plate with 1% BLOTTO–0.5% FBS and incu-bated for 2 h at 37 ◦C to assay for KLH-specific IgA. Horseradishperoxidase-conjugated goat anti-mouse IgA antibody (SigmaChemical Co.) diluted 1:10,000 in 2.5% BLOTTO–0.5% FBS wasthen added to all wells and incubated for 1 h at 37 ◦C. Allreactions were developed, stopped and detected as describedabove. The level of KLH-specific IgA was calculated from thelinear portion of a standard curve determined as previouslydescribed [18]. Each fecal antibody response was expressed asthe ratio of nanograms of KLH-specific IgA per microgram oftotal IgA.
(iii) ELISA to measure TT-specific serum and fecal antibodyresponses. Plates were coated with 10 �g/ml of TT overnightat room temperature. All reactions were detected, quantitatedand analyzed as described above.
d either tetanus toxoid (TT) or ovalbumin (OVA). Animals (n = 5) were administeredunostimulatory control. Samples were collected 14 days post final inoculation andresults in Geometric Mean Titers (GMT), panel (B) shows the level of KLH-specificVA-specific IgA fecal levels among the various immunized groups of mice.
O.V. Kavanagh et al. / Vaccine 28 (2010) 3106–3111 3109
F y inocc art ann ed gr
3
d(wwCwM
4
4
diip(m(faI3aI
s(w
FOfi
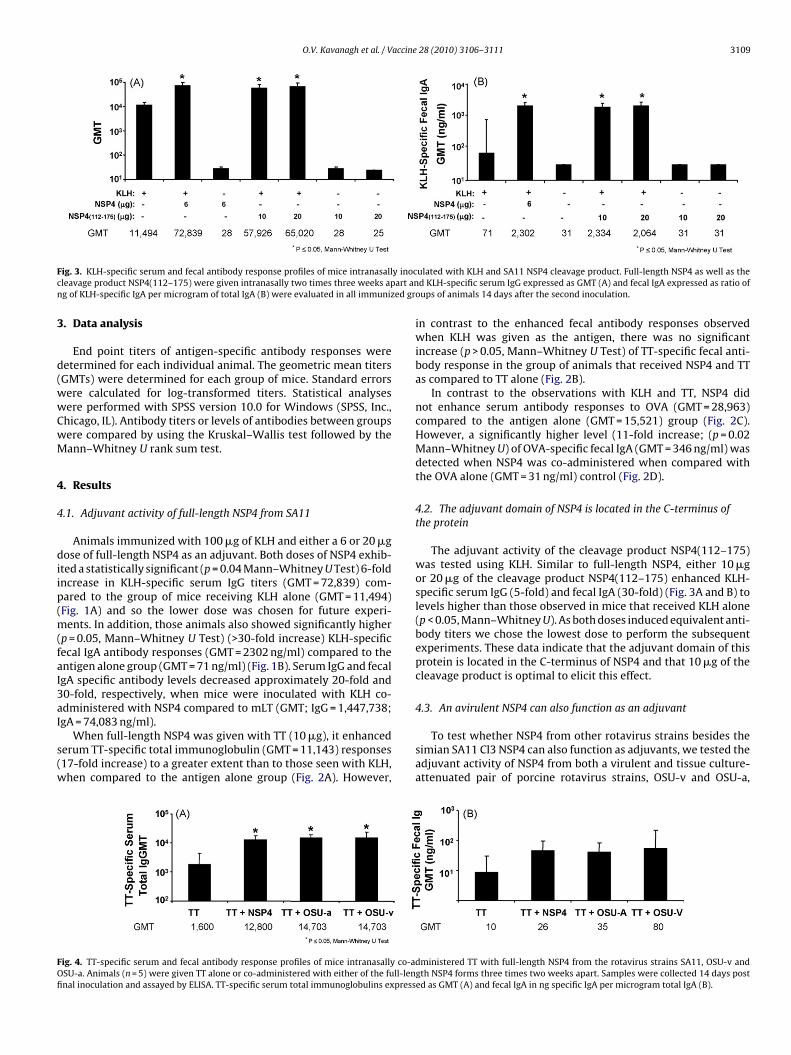
ig. 3. KLH-specific serum and fecal antibody response profiles of mice intranasallleavage product NSP4(112–175) were given intranasally two times three weeks apg of KLH-specific IgA per microgram of total IgA (B) were evaluated in all immuniz
. Data analysis
End point titers of antigen-specific antibody responses wereetermined for each individual animal. The geometric mean titersGMTs) were determined for each group of mice. Standard errorsere calculated for log-transformed titers. Statistical analysesere performed with SPSS version 10.0 for Windows (SPSS, Inc.,hicago, IL). Antibody titers or levels of antibodies between groupsere compared by using the Kruskal–Wallis test followed by theann–Whitney U rank sum test.
. Results
.1. Adjuvant activity of full-length NSP4 from SA11
Animals immunized with 100 �g of KLH and either a 6 or 20 �gose of full-length NSP4 as an adjuvant. Both doses of NSP4 exhib-
ted a statistically significant (p = 0.04 Mann–Whitney U Test) 6-foldncrease in KLH-specific serum IgG titers (GMT = 72,839) com-ared to the group of mice receiving KLH alone (GMT = 11,494)Fig. 1A) and so the lower dose was chosen for future experi-
ents. In addition, those animals also showed significantly higherp = 0.05, Mann–Whitney U Test) (>30-fold increase) KLH-specificecal IgA antibody responses (GMT = 2302 ng/ml) compared to thentigen alone group (GMT = 71 ng/ml) (Fig. 1B). Serum IgG and fecalgA specific antibody levels decreased approximately 20-fold and0-fold, respectively, when mice were inoculated with KLH co-dministered with NSP4 compared to mLT (GMT; IgG = 1,447,738;
gA = 74,083 ng/ml).When full-length NSP4 was given with TT (10 �g), it enhancederum TT-specific total immunoglobulin (GMT = 11,143) responses17-fold increase) to a greater extent than to those seen with KLH,hen compared to the antigen alone group (Fig. 2A). However,
ig. 4. TT-specific serum and fecal antibody response profiles of mice intranasally co-aSU-a. Animals (n = 5) were given TT alone or co-administered with either of the full-lennal inoculation and assayed by ELISA. TT-specific serum total immunoglobulins express
ulated with KLH and SA11 NSP4 cleavage product. Full-length NSP4 as well as thed KLH-specific serum IgG expressed as GMT (A) and fecal IgA expressed as ratio of
oups of animals 14 days after the second inoculation.
in contrast to the enhanced fecal antibody responses observedwhen KLH was given as the antigen, there was no significantincrease (p > 0.05, Mann–Whitney U Test) of TT-specific fecal anti-body response in the group of animals that received NSP4 and TTas compared to TT alone (Fig. 2B).
In contrast to the observations with KLH and TT, NSP4 didnot enhance serum antibody responses to OVA (GMT = 28,963)compared to the antigen alone (GMT = 15,521) group (Fig. 2C).However, a significantly higher level (11-fold increase; (p = 0.02Mann–Whitney U) of OVA-specific fecal IgA (GMT = 346 ng/ml) wasdetected when NSP4 was co-administered when compared withthe OVA alone (GMT = 31 ng/ml) control (Fig. 2D).
4.2. The adjuvant domain of NSP4 is located in the C-terminus ofthe protein
The adjuvant activity of the cleavage product NSP4(112–175)was tested using KLH. Similar to full-length NSP4, either 10 �gor 20 �g of the cleavage product NSP4(112–175) enhanced KLH-specific serum IgG (5-fold) and fecal IgA (30-fold) (Fig. 3A and B) tolevels higher than those observed in mice that received KLH alone(p < 0.05, Mann–Whitney U). As both doses induced equivalent anti-body titers we chose the lowest dose to perform the subsequentexperiments. These data indicate that the adjuvant domain of thisprotein is located in the C-terminus of NSP4 and that 10 �g of thecleavage product is optimal to elicit this effect.
4.3. An avirulent NSP4 can also function as an adjuvant
To test whether NSP4 from other rotavirus strains besides thesimian SA11 Cl3 NSP4 can also function as adjuvants, we tested theadjuvant activity of NSP4 from both a virulent and tissue culture-attenuated pair of porcine rotavirus strains, OSU-v and OSU-a,
dministered TT with full-length NSP4 from the rotavirus strains SA11, OSU-v andgth NSP4 forms three times two weeks apart. Samples were collected 14 days posted as GMT (A) and fecal IgA in ng specific IgA per microgram total IgA (B).
3110 O.V. Kavanagh et al. / Vaccine 28 (2010) 3106–3111
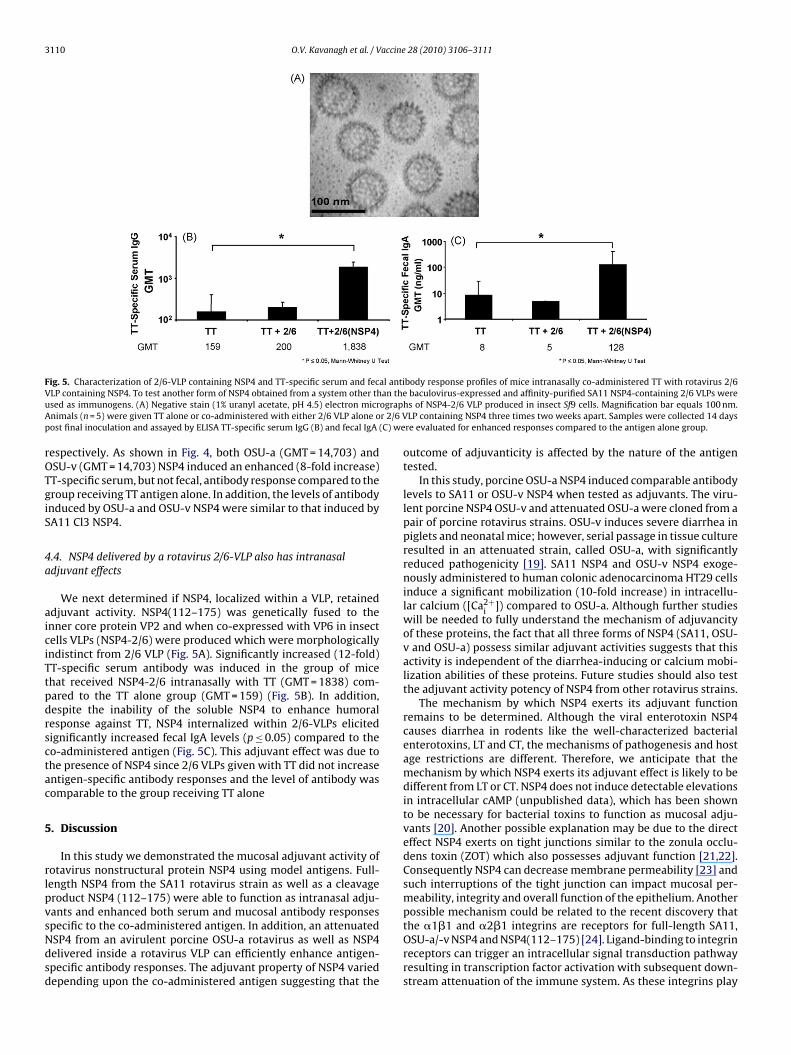
Fig. 5. Characterization of 2/6-VLP containing NSP4 and TT-specific serum and fecal antibody response profiles of mice intranasally co-administered TT with rotavirus 2/6V an theu ograpA r 2/6 Vp (C) we
rOTgiS
4a
aiciTtpdrsctac
5
rlpvsNdsd
LP containing NSP4. To test another form of NSP4 obtained from a system other thsed as immunogens. (A) Negative stain (1% uranyl acetate, pH 4.5) electron micrnimals (n = 5) were given TT alone or co-administered with either 2/6 VLP alone oost final inoculation and assayed by ELISA TT-specific serum IgG (B) and fecal IgA
espectively. As shown in Fig. 4, both OSU-a (GMT = 14,703) andSU-v (GMT = 14,703) NSP4 induced an enhanced (8-fold increase)T-specific serum, but not fecal, antibody response compared to theroup receiving TT antigen alone. In addition, the levels of antibodynduced by OSU-a and OSU-v NSP4 were similar to that induced byA11 Cl3 NSP4.
.4. NSP4 delivered by a rotavirus 2/6-VLP also has intranasaldjuvant effects
We next determined if NSP4, localized within a VLP, retaineddjuvant activity. NSP4(112–175) was genetically fused to thenner core protein VP2 and when co-expressed with VP6 in insectells VLPs (NSP4-2/6) were produced which were morphologicallyndistinct from 2/6 VLP (Fig. 5A). Significantly increased (12-fold)T-specific serum antibody was induced in the group of micehat received NSP4-2/6 intranasally with TT (GMT = 1838) com-ared to the TT alone group (GMT = 159) (Fig. 5B). In addition,espite the inability of the soluble NSP4 to enhance humoralesponse against TT, NSP4 internalized within 2/6-VLPs elicitedignificantly increased fecal IgA levels (p ≤ 0.05) compared to theo-administered antigen (Fig. 5C). This adjuvant effect was due tohe presence of NSP4 since 2/6 VLPs given with TT did not increasentigen-specific antibody responses and the level of antibody wasomparable to the group receiving TT alone
. Discussion
In this study we demonstrated the mucosal adjuvant activity ofotavirus nonstructural protein NSP4 using model antigens. Full-ength NSP4 from the SA11 rotavirus strain as well as a cleavageroduct NSP4 (112–175) were able to function as intranasal adju-ants and enhanced both serum and mucosal antibody responses
pecific to the co-administered antigen. In addition, an attenuatedSP4 from an avirulent porcine OSU-a rotavirus as well as NSP4elivered inside a rotavirus VLP can efficiently enhance antigen-pecific antibody responses. The adjuvant property of NSP4 variedepending upon the co-administered antigen suggesting that thebaculovirus-expressed and affinity-purified SA11 NSP4-containing 2/6 VLPs werehs of NSP4-2/6 VLP produced in insect Sf9 cells. Magnification bar equals 100 nm.
LP containing NSP4 three times two weeks apart. Samples were collected 14 daysre evaluated for enhanced responses compared to the antigen alone group.
outcome of adjuvanticity is affected by the nature of the antigentested.
In this study, porcine OSU-a NSP4 induced comparable antibodylevels to SA11 or OSU-v NSP4 when tested as adjuvants. The viru-lent porcine NSP4 OSU-v and attenuated OSU-a were cloned from apair of porcine rotavirus strains. OSU-v induces severe diarrhea inpiglets and neonatal mice; however, serial passage in tissue cultureresulted in an attenuated strain, called OSU-a, with significantlyreduced pathogenicity [19]. SA11 NSP4 and OSU-v NSP4 exoge-nously administered to human colonic adenocarcinoma HT29 cellsinduce a significant mobilization (10-fold increase) in intracellu-lar calcium ([Ca2+
i ]) compared to OSU-a. Although further studieswill be needed to fully understand the mechanism of adjuvancityof these proteins, the fact that all three forms of NSP4 (SA11, OSU-v and OSU-a) possess similar adjuvant activities suggests that thisactivity is independent of the diarrhea-inducing or calcium mobi-lization abilities of these proteins. Future studies should also testthe adjuvant activity potency of NSP4 from other rotavirus strains.
The mechanism by which NSP4 exerts its adjuvant functionremains to be determined. Although the viral enterotoxin NSP4causes diarrhea in rodents like the well-characterized bacterialenterotoxins, LT and CT, the mechanisms of pathogenesis and hostage restrictions are different. Therefore, we anticipate that themechanism by which NSP4 exerts its adjuvant effect is likely to bedifferent from LT or CT. NSP4 does not induce detectable elevationsin intracellular cAMP (unpublished data), which has been shownto be necessary for bacterial toxins to function as mucosal adju-vants [20]. Another possible explanation may be due to the directeffect NSP4 exerts on tight junctions similar to the zonula occlu-dens toxin (ZOT) which also possesses adjuvant function [21,22].Consequently NSP4 can decrease membrane permeability [23] andsuch interruptions of the tight junction can impact mucosal per-meability, integrity and overall function of the epithelium. Anotherpossible mechanism could be related to the recent discovery that
the �1�1 and �2�1 integrins are receptors for full-length SA11,OSU-a/-v NSP4 and NSP4(112–175) [24]. Ligand-binding to integrinreceptors can trigger an intracellular signal transduction pathwayresulting in transcription factor activation with subsequent down-stream attenuation of the immune system. As these integrins play
accine
ait
ehmwtwmpftaNptb
dpablptia
iwrfiNwTin
eafa
A
SNDgN
a
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
1996;26(9):2023–9.[27] Rubio MA, Sotillos M, Jochems G, Alvarez V, Corbi AL. Monocyte activation:
O.V. Kavanagh et al. / V
role in modulating the immune system [25–27] it will be interest-ng to determine if NSP4 exerts its adjuvant effect through bindingo these receptors.
Even though other mucosal adjuvants have been exploredxtensively in the past, to date, none have been approved foruman use to be given by mucosal routes. Our studies showedLT induced significantly higher fecal and serum antibody titershen co-administered with TT and KLH model antigens, a result
hat confirms previous studies with this bacterial enterotoxin orith CT as well as its mutants in extensive testing in various ani-al models in combination with a variety of viral, bacterial, and
arasitic pathogens; however, safety issues still remain a concernor these bacterial enterotoxin adjuvants [5]. Therefore, an effec-ive, safe and practical mucosal adjuvant remains to be identifiednd characterized for the development of mucosal vaccines. SinceSP4 does not bind to GM1 receptors like CT or LT [13] it may notossess neurotoxic side effects. However future preclinical, safetyrials will need to be undertaken to ensure NSP4 does not enter therain or possess other toxicity.
Furthermore, we observed differences in adjuvant responseepending upon the nature of the co-administered antigen. Theresence of NSP4 induced a stronger immune response to the co-dministered antigen compared to the immune response elicitedy administering the same antigen alone. This finding corre-
ates with the fact that inclusion of specific adjuvants in vaccinereparations can modify the presentation modality of antigenso the immune system and/or improve the induction of themmune response over that induced by the same antigen givenlone [28].
Virus-like particles as an alternative vaccine strategy is anmportant area in the field of rotavirus vaccinology. In this study
e explored the ability of NSP4 to act as an adjuvant for non-eplicating rotavirus VLP vaccines developed in our laboratory. Weound that NSP4 retained its adjuvant properties even when admin-stered within a NSP4-2/6 VLP. The observed adjuvant effect ofSP4-2/6 was due to the presence of NSP4 since 2/6 VLPs givenith antigen did not increase antigen-specific antibody responses.
he addition of NSP4 to 2/6 VLPs could increase the adjuvantic-ty and immunogenicity of rotaviral vaccines and may alleviate theeed for co-administered adjuvants.
Future experiments will examine any adjuvant effect NSP4xerts on the cellular arm of the immune system against co-dministered antigen, elucidate the mechanism by which NSP4unctions as an adjuvant and also determine if NSP4 also possessesdjuvant properties when administered by alternative routes.
cknowledgements
This work was supported by funding from the U.S. Public Healthervice, The Enteric Pathogens Research Unit, NIAID contract01-A165299 and from the National Institutes of Health (grantsK30144, DK56338, AI080656), and E.C. was funded by a pediatricastroenterology training fellowship (grant T32 DK07664) from theational Institutes of Health.
We thank Dr. Jerry R. McGhee for providing the tetanus toxoidnd Dr. John D. Clements for providing the mutant LT (LT-R192G).
eferences
[1] Hasegawa H, Ichinohe T, Ainai A, Tamura S, Kurata T. Development of mucosaladjuvants for intranasal vaccine for H5N1 influenza viruses. Ther Clin RiskManag 2009;5(1):125–32.
[
28 (2010) 3106–3111 3111
[2] Stanley M, Gissmann L, Nardelli-Haefliger D. Immunobiology of humanpapillomavirus infection and vaccination—implications for second generationvaccines. Vaccine 2008;26(Suppl. 10):K62–7.
[3] Lehner T, Bergmeier L, Wang Y, Tao L, Mitchell E. A rational basis for mucosalvaccination against HIV infection. Immunol Rev 1999;170:183–96.
[4] Schnitzler AC, Burke JM, Wetzler LM. Induction of cell signaling eventsby the cholera toxin B subunit in antigen-presenting cells. Infect Immun2007;75(6):3150–9.
[5] van Ginkel FW, Jackson RJ, Yuki Y, McGhee JR. Cutting edge: the mucosal adju-vant cholera toxin redirects vaccine proteins into olfactory tissues. J Immunol2000;165(9):4778–82.
[6] Couch RB. Nasal vaccination, Escherichia coli enterotoxin, and Bell’s palsy. NEngl J Med 2004;350(9):860–1.
[7] Hagiwara Y, Kawamura YI, Kataoka K, Rahima B, Jackson RJ, Komase K, et al. Asecond generation of double mutant cholera toxin adjuvants: enhanced immu-nity without intracellular trafficking. J Immunol 2006;177(5):3045–54.
[8] Ball JM, Tian P, Zeng CQ, Morris AP, Estes MK. Age-dependent diarrhea inducedby a rotaviral nonstructural glycoprotein. Science 1996;272(5258):101–4.
[9] Zhang M, Zeng CQ, Morris AP, Estes MK. A functional NSP4 enterotoxin peptidesecreted from rotavirus-infected cells. J Virol 2000;74(24):11663–70.
10] Guerrero RA, Ball JM, Krater SS, Pacheco SE, Clements JD, Estes MK.Recombinant Norwalk virus-like particles administered intranasally to miceinduce systemic and mucosal (fecal and vaginal) immune responses. J Virol2001;75(20):9713–22.
11] O’Neal CM, Crawford SE, Estes MK, Conner ME. Rotavirus virus-likeparticles administered mucosally induce protective immunity. J Virol1997;71(11):8707–17.
12] Zhang M, Zeng CQ, Dong Y, Ball JM, Saif LJ, Morris AP, et al. Mutations in rotavirusnonstructural glycoprotein NSP4 are associated with altered virus virulence. JVirol 1998;72(5):3666–72.
13] Dong Y, Zeng CQ, Ball JM, Estes MK, Morris AP. The rotavirus enterotoxin NSP4mobilizes intracellular calcium in human intestinal cells by stimulating phos-pholipase C-mediated inositol 1,4,5-trisphosphate production. Proc Natl AcadSci USA 1997;94(8):3960–5.
14] Zeng CQ, Estes MK, Charpilienne A, Cohen J. The N terminus of rotavirus VP2 isnecessary for encapsidation of VP1 and VP3. J Virol 1998;72(1):201–8.
15] Bertolotti-Ciarlet A, Ciarlet M, Crawford SE, Conner ME, Estes MK. Immuno-genicity and protective efficacy of rotavirus 2/6-virus-like particles producedby a dual baculovirus expression vector and administered intramuscularly,intranasally, or orally to mice. Vaccine 2003;21(25–26):3885–900.
16] Dickinson BL, Clements JD. Dissociation of Escherichia coli heat-labileenterotoxin adjuvanticity from ADP-ribosyltransferase activity. Infect Immun1995;63(5):1617–23.
17] Ball JM, Hardy ME, Atmar RL, Conner ME, Estes MK. Oral immunization withrecombinant Norwalk virus-like particles induces a systemic and mucosalimmune response in mice. J Virol 1998;72(2):1345–53.
18] Franco MA, Greenberg HB. Role of B cells and cytotoxic T lymphocytesin clearance of and immunity to rotavirus infection in mice. J Virol1995;69(12):7800–6.
19] Bohl EH, Theil KW, Saif LJ. Isolation and serotyping of porcine rotavirusesand antigenic comparison with other rotaviruses. J Clin Microbiol1984;19(2):105–11.
20] Cheng E, Cardenas-Freytag L, Clements JD. The role of cAMP in mucosaladjuvanticity of Escherichia coli heat-labile enterotoxin (LT). Vaccine1999;18(1–2):38–49.
21] Marinaro M, Fasano A, De Magistris MT. Zonula occludens toxin acts as anadjuvant through different mucosal routes and induces protective immuneresponses. Infect Immun 2003;71(4):1897–902.
22] Marinaro M, Di Tommaso A, Uzzau S, Fasano A, De Magistris MT. Zonula occlu-dens toxin is a powerful mucosal adjuvant for intranasally delivered antigens.Infect Immun 1999;67(3):1287–91.
23] Tafazoli F, Zeng CQ, Estes MK, Magnusson KE, Svensson L. NSP4 enterotoxinof rotavirus induces paracellular leakage in polarized epithelial cells. J Virol2001;75(3):1540–6.
24] Seo NS, Zeng CQ, Hyser JM, Utama B, Crawford SE, Kim KJ, et al. Inauguralarticle: integrins alpha1beta1 and alpha2beta1 are receptors for the rotavirusenterotoxin. Proc Natl Acad Sci USA 2008;105(26):8811–8.
25] Dietl J, Ruck P, Marzusch K, Horny HP, Kaiserling E, Handgretinger R. Uter-ine granular lymphocytes are activated natural killer cells expressing VLA-1.Immunol Today 1992;13(6):p236.
26] Perez-Villar JJ, Melero I, Gismondi A, Santoni A, Lopez-Botet M. Functional anal-ysis of alpha 1 beta 1 integrin in human natural killer cells. Eur J Immunol
rapid induction of alpha 1/beta 1 (VLA-1) integrin expression by lipopolysac-charide and interferon-gamma. Eur J Immunol 1995;25(9):2701–5.
28] Buonaguro FM, Tornesello ML, Buonaguro L. Virus-like particle vaccines andadjuvants: the HPV paradigm. Expert Rev Vaccines 2009;8(10):1379–98.
Copyright © 2022 FDOKUMEN




















