
transcriptional and epigenetic regulation of human cd4+ t ...
Upload
independentCategory
view
4download
0
Gene 242 (2000) 407–418www.elsevier.com/locate/gene
Transcriptional regulation of the human DNAMethyltransferase (dnmt1) gene
Pascal Bigey, Shyam Ramchandani, Johanne Theberge, Felipe D. Araujo, Moshe Szyf *Department of Pharmacology and Therapeutics, McGill University, 3655 Drummond Street, Montreal, PQ, Canada, H3G 1Y6
Received 15 September 1999; accepted 8 November 1999Received by S.D.J. Pena
Abstract
DNA methylation is an important component of the epigenetic control of genome functions. Understanding the regulation ofthe DNA Methyltransferase (dnmt1) gene expression is critical for comprehending how DNA methylation is coordinated withother critical biological processes. In this paper, we investigate the transcriptional regulatory region of the human dnmt1 geneusing a combination of RACE, RNase protection analysis and CAT assays. We identified one major and three minor transcriptioninitiation sites in vivo (P1–P4), which are regulated by independent enhancers and promoter sequences. The minimal promoterelements of P1, P2 and P4 are mapped within 256 bp upstream of their respective transcription initiation sites. P1 is nested withina CG-rich area, similar to other housekeeping genes, whereas P2–P4 are found in CG-poor areas. Three c-Jun-dependent enhancersare located downstream to P1 and upstream to P2–P4, thus providing a molecular explanation for the responsiveness of dnmt1 tooncogenic signals that are mediated by the Ras-c-Jun oncogenic signaling pathway. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: DNA methylation; DNA methyltransferase; Enhancers; Promoters; Ras signaling pathway
1. Introduction 1987) or by attracting the binding of methylated-DNA-binding proteins that repress gene expression (Nan et al.,1997, 1998; Jones et al., 1998; Cross et al., 1997). It isMammalian DNA is modified by methylation of 60–
80% of the cytosines residing in the dinucleotide clear that understanding the mechanisms responsible forgenerating DNA methylation patterns is critical forsequence CpG (Razin and Szyf, 1984). DNA methyl-
ation is now being recognized as an important epigenetic elucidating the processes through which genomic expres-sion programs are generated.control over different genome functions including
differential gene expression (Razin and Cedar, 1991), The enzyme catalyzing the modification of DNA bymethylation is the DNA methyltransferase (Adamsallele-specific expression in parental imprinting
(Sapienza et al., 1987) and X inactivation (Zuccotti and et al., 1979). One dnmt1 cDNA has been cloned andMonk, 1995). Since vertebrate DNA is differentially extensively characterized to date in human and murinemethylated at CpG dinucleotide sequences, DNA meth- cells (Bestor et al., 1988; Cumaraswamy et al., 1992).ylation may serve as a signaling mechanism forming The 5∞ end of the murine and human dnmt1 RNA havesite-, gene-, and tissue-specific patterns of methylation recently been characterized, and it has been suggested(Razin and Szyf, 1984). Methylation of CpG dinucleo- that this is the only DNMT1 isoform expressed intides can inhibit gene expression either by directly somatic cells (Tucker et al., 1996; Yoder et al., 1996).interfering with the interaction of transcription factors However, splice variants have recently been charac-with their cognate recognition sequences (Becker et al., terized in germ cells (Mertineit et al., 1998) and somatic
cells (Deng and Szyf, 1998).It stands to reason that regulation of dnmt1 expres-
Abbreviations: AP-1, activator protein 1; DNMT1, DNA methyl- sion plays an important role in coordinating DNMT1transferase 1; EMSA, electrophoretic mobility shift assay; RACE, activity with different developmental and cellular pro-rapid amplification cDNA ends
grams (Szyf, 1994, 1998). In accordance with this* Corresponding author. Tel.: +1-514-398-7107;hypothesis, it has been previously shown that dnmt1 isfax: +1-514-398-6690.
E-mail address: [email protected] (M. Szyf ) post-transcriptionally regulated with the state of cell
0378-1119/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved.PII: S0378-1119 ( 99 ) 00501-6

408 P. Bigey et al. / Gene 242 (2000) 407–418
Fig. 1. RACE-PCR determination of initiation sites of dnmt1. (A) The genomic organization of dnmt1 is shown in the first diagram. The horizontalline represents introns and vertical lines indicate exons. The numbered arrows above the diagram indicate the position of the 3∞ amplimers used inthe RACE analysis. The 5∞ ends of dnmt1 mRNA in either human hippocampus or placenta Marathon cDNA library were determined by PCRamplification using as the 5∞ oligo the AP1 (adaptor primer Clontech) adaptor and as a 3∞ amplimer either oligos 161 encoding a sequence fromexon 6 or 145 encoding a sequence from exon 21. Ten picograms of marathon-ready cDNA were amplified using KlenTaq (Clontech). (B) TheRACE-PCR products were subcloned and sequenced. The start sites identified are shown as horizontal arrows and are indicated by letters a–i (theinitiation site corresponding to the letter a was previously characterized; Tucker et al., 1996; Yoder et al., 1996). Arrows below exon 3 indicateinitiations identified by RNase protection shown in Fig. 2 that were not identified in the cloned RACE products.
growth (Szyf et al., 1991), that dnmt1 expression is obtained from Clontech. The 5∞ ends of dnmt1 mRNAinduced by the Ras-AP-1 signaling pathway (MacLeod were determined by RACE-PCR (Frohman et al., 1989)et al., 1995) and that an AP-1 regulatory element amplification using as the 5∞ oligo the AP1 (adaptorregulates the expression of the murine dnmt1 gene primer Clontech) adaptor and as a 3∞ amplimer either(Rouleau et al., 1995). We have therefore suggested that oligo 145 (ATCGAAGATCTGGTAGACCAGCT-induction of expression of dnmt1 is a downstream event TGGT) (Fig. 1) encoding a sequence from exon 21 orin oncogenic programs, such as the Ras/AP-1 signaling oligo 161 (TTTCCAAGTCTTTGAGCCTG) encodingpathway, and that it might play a causal role in a sequence from exon 6 (Fig. 1). Ten picograms ofcellular transformation (MacLeod and Szyf, 1995; marathon-ready cDNA were amplified using KlenTaqRamchandani et al., 1997). Recent data have provided (Clontech) under the following conditions: 94°C forfurther evidence to support the hypothesis that the 0.5 min, 55°C for 0.5 min, 68°C for 6 min for 40 cycles,Ras/AP-1 pathway regulates dnmt1 expression (Deng followed by 6 min incubation at 68°C. The amplificationet al., 1998; Bakin and Curran, 1999). products were resolved on an agarose gel. An aliquot
A putative promoter and enhancer have been iden- of the amplification reaction was ligated to pCR 2.1,tified previously in the mouse, upstream to exons 3 and and the resulting colonies were sequenced to determine4 (Rouleau et al., 1992), but it is clear that transcription the sequence of the 5∞ end of the human dnmt1.initiates upstream to the previously identified promoteras well (Tucker et al., 1996; Yoder et al., 1996). Thetranscriptional regulatory regions of this initiation site 2.2. Cell culture and RNase protectionare unknown, and it is not yet clear whether alternativetranscription initiation sites are utilized in vivo. The fact Human non-small cell lung carcinoma cell line A-549that the regulatory regions of dnmt1 have been unre- (ATCC CCL 185 ), human bladder carcinoma cell linesolved to date has limited our understanding of how its T-24 (ATCC HTB 4), human cervical carcinoma cellexpression is coordinated with other important biologi- line HeLa (ATCC CCL2) and mouse embryonal carci-cal and pathological processes. This paper identifies
noma cell line P-19 (ATCC CRL 1825) were culturedtranscription initiation sites of dnmt1 and genetic ele-in Dulbecco’s modified Eagle’s medium (with low glu-ments regulating them.cose for A-549 and high glucose for the other cell lines)supplemented with 10% (v/v) heat-inactivated fetal calfserum, 2 mM glutamine, and antibiotics. Total cellular2. Materials and methodsRNA was prepared by extraction with guanidine thiocy-anate (Chirgwin et al., 1979). Human mRNA samples2.1. RACE-PCR analysis of 5∞ ends of human brainwere purchased from Clontech.dnmt1 mRNA
Two probes were used for RNase protection bearingeither sequences encoding bases 168–611 (exons 1–4) orHuman hippocampus and placenta cDNA libraries
with a Marathon cDNA adaptor ligated to its ends were bases 396–740 (exons 3–5) in the antisense orientation

409P. Bigey et al. / Gene 242 (2000) 407–418
(see Fig. 2 for map). The antisense RNA probe was 2.3. Genomic cloning and construction of plasmidstranscribed using T7 RNA polymerase and[a-32P]UTP and was hybridized with either T-24 RNA, All plasmids were constructed using genomic sub-
clones of dnmt1 pTB and pFTR1 as previously describedA-549 RNA, human hippocampus mRNA or humanplacenta mRNA at 50°C overnight. The hybridized (Ramchandani et al., 1998). pTB (20 kb genomic insert)
was digested with either HindIII or XbaI. A 2 kb HindIIIRNA was treated with RNases A and T1, as describedpreviously ( Kreig and Melton, 1987), and the protected fragment containing exon 1, a 12 kb XbaI fragment
containing exon 1 and a 10 kb HindIII fragment contain-fragments were resolved on an 8 M urea sequencing gelnext to a century ladder (Ambion). ing exon 1b were isolated and inserted into a HindIII-
Fig. 2. Multiple 5∞ transcripts of dnmt1 detected by an RNase protection assay. RNA prepared from tumor cell lines and human placenta indicatedin the figure was subjected to an RNase protection assay using a riboprobe complementary to a cDNA sequence spanning either exons 1–4 of thehuman dnmt1 or a riboprobe complementary to exons 3–5, as described in Section 2 and separated on a sequencing gel next to an RNA molecularweight marker. Arrows indicate the observed fragment and their predicted size. The letters next to the arrows indicate the correspondingRACE-PCR product, whereas an 1 or 11 next to the arrow indicates a start site not identified by RACE-PCR. The indicated fragment cannot bea product of the three alternatively spliced mRNAs in this region since protection of these splice products would have resulted in fragments of 150or 346 nt. The physical structure of each probe is indicated under the respective autoradiogram. The striped box at the 5∞ of each probe representsleader sequence originating from the plasmid template. The size in nucleotides of each exon that is included in the probes is indicated on top ofthe lines. The first line indicates the expected fragment size if the mRNA is initiated at the previously reported site (Tucker et al., 1996; Yoderet al., 1996). Initiation downstream to the previously described initiation site results in shorter protected fragments, as indicated below thetop line.

410 P. Bigey et al. / Gene 242 (2000) 407–418
digested pBluescript SK− plasmid or a XbaI-digested 2.4. CAT assayspBluescript SK− plasmid. The 2 kb HindIII insert was
P-19 cells or HeLa cells were plated at a density offully sequenced by the dideoxy chain termination105/well in a six-well tissue culture dish (Falcon) 24 hmethod using a T7 DNA sequencing kit (Pharmacia).prior to transfection. Fifteen micrograms of plasmidFusion CAT constructs containing different fragmentsDNA were incorporated into a calcium phosphate precip-from the 5∞-flanking dnmt1 gene were then generated.itate (Rouleau et al., 1992). For cotransfection, we usedThe following CAT vectors were used: pOCAT bore the15 mg of the CAT construct and 7.5 mg of either SK as abacterial chloramphenicol acetyl transferase gene withcontrol, RSV-c-Jun (Binetruy et al., 1991), ∂9 v-c-Junno eukaryotic promoter or enhancer; pCAT promoter(Lloyd et al., 1991) or pLRbRNL (ATCC R81 65003).bore the Herpes virus thymidine kinase promoter butTwo hundred microliters of the precipitate were addedno enhancer activity, and was used to determineper well containing 2 ml of the appropriate medium. Theenhancer activity (CAT constructs were purchased frommedium was replaced after 24 h, and cells were harvestedPromega). pMet-P1-CAT sense and antisense werefor CAT assays 48 h post-transfection. Cells were dis-obtained by ligating the 2 kb end-blunted HindIIIlodged by scraping in phosphate-buffered saline, andfragment into a XbaI-digested and end-bluntedextracts were prepared in 100 ml of 100 mM Tris–HClpOCAT. pMet-P1-PrCAT at either sense or antisense(pH 8) by five freeze–thaw cycles. CAT assays werewere obtained by ligating the 2 kb HindIII fragment,performed on 50 ml of extract (~10 mg of protein) usingrespectively, in a HindIII-digested pCAT Promoter[3H]acetyl coenzyme A (0.5 mCi/assay, 3.8 Ci/mmol )(Promega) and a HindIII-digested pCAT Enhanceressentially as described previously (Rouleau et al., 1992).(Promega).All transfection assays were performed in triplicate andThe 2 kb HindIII fragment described above waswere repeated using two different plasmid preparationsdigested with XbaI, and a 1.2 kb HindIII–XbaI fragmentand two different cultures of either P-19 or HeLa cells.(DHX ) and 0.8 kb XbaI–HindIII fragment containing
exon 1 (DXH) were isolated and end-blunted. pMet-2.5. Electromobility shift assayP1-DHX-CAT (in either orientation) and pMet-
P1-DHX-PrCAT (in either orientation) were generatedThe following oligonucleotide duplexes containingby inserting the 0.8 kb XbaI–HindIII fragment into an
putative AP-1 binding sites from promoters 1-3 wereend-blunted XbaI-digested site of pOCAT and pCATused: AP-1 consensus: 5∞-CGCTTGATGAGTCAGC-Promoter. pMet-P1-DXH-CAT (in either orientation),CGGAA (sense), 5∞-TTCCGGCTGACTCATCAAG-and pMet-P1-DXH-PrCAT (in either orientation) wereCG (antisense); 1: 5∞-GCTGGGATGACAGGCGTG
generated by inserting the 1.2 kb HindIII–XbaI into the(sense), 5∞-CACGCCTGTCATCCCAGC (antisense); 2:
same vectors. pFTR1 (20 kb) (Ramchandani et al., 5∞-ACAAGGTGACTCATGCCTG (sense), 5∞-CAGG-1998) was digested with XbaI. The reaction was run on CATGAGTCACCTTGA (antisense); 3: 5∞-TGCCAT-a gel, and all five fragments were purified and inserted TCACTCATTGTG (sense), 5∞-CACAATGAGTGA-into a XbaI-digested pBluescript SK plasmid. A 1.4 kb ATGGCA (antisense). The sense oligonucleotide wasXbaI fragment containing exons 2 and 3 and a 0.3 kb radiolabelled in a mixture containing 5 mM of oligo-XbaI fragment containing part of exon 4 were identified nucleotide, 50 mCi of (c-32P)-ATP (3000 Ci/mmol,by DNA sequencing. pMet-P4-CAT (in either orienta- Amersham), and 10 units of T4 polynucleotide kinasetion) and pMet-P4-PrCAT (in either orientation) were (Boehringer-Mannheim) in a final volume of 50 ml, andgenerated by ligating the 0.3 kb XbaI fragment into the the labeled oligonucleotide was then purified on aappropriate XbaI-digested plasmid. Sephadex G25 microcolumn (Pharmacia). The antisense
The 1.4 kb XbaI fragment was digested with NcoI, strand was then added (1 mg), and the duplex wasresulting in a 0.3 kb XbaI–NcoI fragment containing annealed following 5 min of boiling by gradual coolingexon 2 and a 1.1 kb NcoI–XbaI fragment containing of the reaction mixture to room temperature.exon 3. pMet-P2-CAT (in either orientation) and pMet- The DNA-binding reactions contained 20 mM HepesP2-PrCAT (in either orientation) were generated by pH=7, 12% glycerol, 1 mM EDTA, 4 mM DTT, 0.1%ligating the end-blunted 0.3 kb XbaI–NcoI fragment NP 40, 3 mM MgCl2, 5 mg of A549 nuclear extractcontaining exon 2 into the appropriate end-blunted prepared as previously described (Chirgwin et al., 1979),XbaI-digested plasmid. pMet-P3-CAT (in either orienta- 5 mg BSA, 2 mg poly dI–dC, and 10 ng of labeled duplextion) and pMet-P3-PrCAT (in either orientation) were in a 10 ml final volume. The mixtures were incubated forgenerated by ligating the end-blunted 1.1 kb NcoI–XbaI 20 min at room temperature, following which, a 300-foldfragment containing exon 3 into the appropriate end- excess of competitor was added, and the reaction wasblunted XbaI-digested CAT vector. The orientations of incubated for an additional 20 min at room temperature.all these constructs were determined by DNA The mixtures were then analyzed by electrophoresis
on a 4% non-denaturing polyacrylamide gel at 4°Csequencing.

411P. Bigey et al. / Gene 242 (2000) 407–418
for 3 h at 220 V. Gels were dried and exposed for order to exclude the possibility that a protected fragmentautoradiography. represents broken RNA. It would be highly improbable
that different preparations of RNA, randomly broken,will have the same 5∞ boundaries. We first used a
3. Results 32P-labeled riboprobe that is complementary to the firstpreviously described four exons of dnmt1 (Tucker et al.,
3.1. Multiple dnmt1 initiation sites and 5∞ splice-variants 1996; Yoder et al., 1996). To ensure complete synthesisdetermined by a RACE-PCR analysis of placenta and of the riboprobe, the synthesized probe was separatedhippocampus mRNA on a sequencing gel, and the full size probe was eluted.
Preparations of mRNA from two cancer lines (A-549In order to identify regulatory elements of dnmt1 and T-24) and human placenta were hybridized with
gene expression, it is critical to know the transcription the labeled riboprobe and subjected to RNase treatment,initiation sites utilized in vivo. We performed a RACE and the protected fragments were size-fractionated on aanalysis on human hippocampus and placenta mRNA sequencing gel. If dnmt1 mRNA initiates exclusively atin order to delineate the initiation sites of dnmt1 in vivo. the first exon, as has previously been suggested (TuckerThis technique allows the 5∞ ends of mRNA transcribed et al., 1996; Yoder et al., 1996), the full probe would bein a given cell or tissue to be identified (Frohman et al., protected with no additional fragments to be observed.1989). Double-stranded cDNA prepared from both However, if dnmt1 is transcriptionally initiated also attissues, ligated at their ends to a Marathon adaptor, proximal sites, shorter fragments corresponding to thewere amplified with either a 3∞ antisense oligonucleotide, shorter messages would be protected, including thosewhich is complementary to a 27 bp sequence from exon identified in the RACE analysis. The size of the protected21 (oligo 145), or a 20 bp oligo from exon 6 (oligo 161) fragments corresponds to the distance between the initia-and the Marathon anchor primer, as described in tion site and the start point of the RNA probe (Fig. 2).Section 2. The size of the protected fragments does not correspond
In order to precisely delineate the initiation sites to what would be expected from protection of spliceobtained by our RACE analysis, the RACE amplifica- products (Fig. 2).tion products were subcloned into pCR 2.1 vectors and Three fragments, which correspond to messages initi-subjected to dideoxy chain termination sequencing. The ated in exon 1, are observed in the both placenta andresults of this analysis (Fig. 1A and B) show four distinct cancer cell lines (corresponding to the RACE productsclusters of 5∞ ends of dnmt1 mRNA (indicated by letters b, c, d and f in Figs. 1 and 3). There is, however, aa–i in Fig. 1B; the initiation site indicated by the letter
different relative use of these sites in the different tissues.‘a’ was previously characterized (Tucker et al., 1996;
Fragments corresponding to initiations upstream toYoder et al., 1996), but it did not appear in our results)exon 2 (corresponding to the RACE product f in Figs. 1being expressed in vivo at previously defined exons 1,and 3) and in exon 3 (corresponding to RACE product2, 3 and 4. Our analysis also revealed four alternativelyh as well as a 5∞ boundary not identified by RACEspliced new exons that are in frame with the rest of theindicated as an asterisk in Fig. 2) are identified in thecoding sequence of dnmt1. Three of these alternativelycancer cells.spliced sites initiate at exon 1 (unpublished data).
In order to determine whether additional initiationsites are utilized in exon 4, we utilized a riboprobe3.2. RNase protection analysis of the 5∞ ends of dnmt1encoding exons 3, 4 and 5. If transcription is exclusivelyinitiated upstream to exon 3, the entire riboprobe shouldSince both the hippocampus and placenta RACEbe protected (345 nt). Downstream initiation withinanalyses revealed transcripts initiated at the 5∞ exonexon 3 and 4, as predicted from the RACE analysiscorresponding to the fully known transcript (Fig. 1),(products h and i in Figs. 1 and 3), would result inthis suggested to us that the Marathon libraries used inshorter protected fragments, as indicated in the sche-this assay bear full-length cDNAs. However, it is pos-matic diagram (Fig. 2). The result (Fig. 2) indicates thatsible that the downstream 5∞ boundaries identified inboth placenta and cancer cells express dnmt1 mRNAthis study reflect broken RNAs or incomplete primerinitiated within exon 3 (340 nt indicated by a doubleextension by the reverse-transcriptase. In order to verifyasterisk in Fig. 2 and 315 nt corresponding to RACEthat the 5∞ boundaries identified in the RACE analysisproduct h) and at exon 4 (258 nt corresponding toare in fact a result of transcription initiation in vivo, weRACE product i in Figs. 1 and 2). Thus, the RNasehave resorted to an independent method for identifyingprotection analysis is consistent with the initiation clus-5∞ boundaries of RNA, RNase protection assay ( Kreigters identified by the RACE analysis; the result (Fig. 2)and Melton, 1987). We analyzed RNA samples fromalso indicates that initiations at exon 1 are the mostplacenta and two other cell lines (a bladder carcinoma
line, T-24, and a human lung carcinoma, A-549) in abundant.

412 P. Bigey et al. / Gene 242 (2000) 407–418

413P. Bigey et al. / Gene 242 (2000) 407–418
3.3. Nucleotide sequence of 5∞ region of the human dnmt1 with the cell cycle (Szyf et al., 1991), we tested whetherthe independent promoter elements are responsive to
In order to determine whether the 5∞ region of dnmt1 either ectopic c-Jun or tumor suppressor retinoblastomabears multiple promoters and regulatory regions, as the (Rb). In order to verify that the CAT activities observedresults above suggest, we subcloned and sequenced reflect true differences in promoter strength rather thangenomic regions flanking the previously defined exons irregular differences in the quality of the plasmid prepa-1–4 from our previously described genomic dnmt1 ration or transfection efficiency, two independentphages p TB and p FTR (Ramchandani et al., 1998). plasmids and three independent transfections were per-The precise 5∞ boundaries of the RACE products are formed per construct. The AP-1 transactivationindicated by arrows above the sequence of the 5∞ region activity of the c-Jun expression vector was confirmedof dnmt1 (Fig. 3). An analysis of the sequences residing by testing both its ability to transactivate aupstream to the transcription initiation sites identified murine pMetCAT vector bearing consensus AP-1 sitesby the RACE experiment reveals the absence of canoni- and its inability to do so with a construct that does notcal TATAA or CAAT sequences, which are characteristic bear AP-1 sites (data not shown).of many housekeeping genes (Seiser et al., 1989). A Since an XbaI fragment bearing 0.256 kb upstreamcommon trait of TATAA-less housekeeping gene pro- to the first exon (pMet-P1-DHX-CAT ) can direct tran-moters is that they are nested within a CG-rich area scription of a promoter-less CAT in the sense, but not(Bird, 1986). The upstream transcription initiation the antisense, orientation (Fig. 4A, II ), a promoterregion in exon 1 is CG-rich, whereas the downstream element should reside within 250 bp upstream of theinitiation sites are nested within a relatively CG-poor first transcription initiation cluster identified in vivoarea. This setting is similar to the situation described (P1). No additional elements required for P1 expressionfor the mouse homologue (Rouleau et al., 1992) and is are present in the 1.2 kb upstream to the P1 promoterconsistent with the hypothesis that there is differential (pMet-P1-CAT ) since inclusion of this sequence doesregulation of the upstream initiation sites versus the not enhance, but rather represses, the activity of thedownstream initiation sites.
promoter (Fig. 4A, I ). This 1.2 kb upstream fragmentA computer analysis of the 5∞ sequences for putative
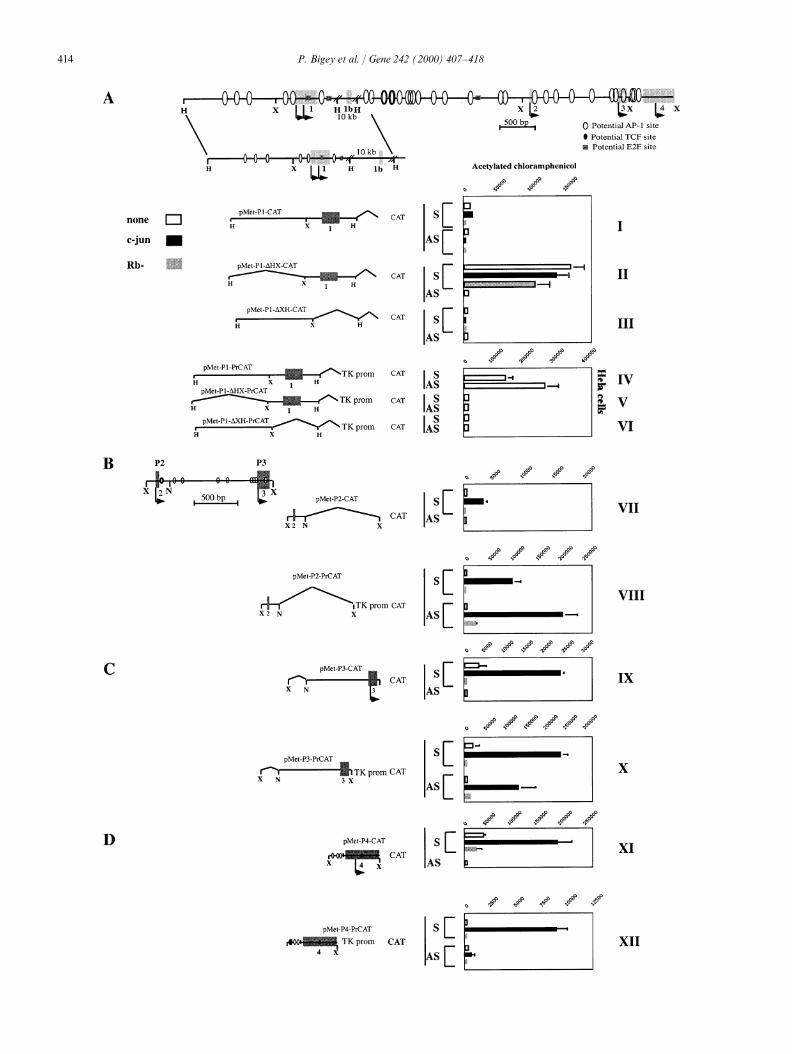
cannot induce reporter activity by itself (pMet-P1-DXH-recognition sequences for known transcription factorsCAT, Fig. 4A, III ). The P1 promoter is not responsivereveals multiple AP-1 recognition sites (Binetruy et al.,to ectopic expression of c-Jun but is repressed by coex-1991), an E2F recognition site (Bagchi et al., 1991) aspression of Rb. This result is expected since there is anwell as multiple TCF binding sites ( Korinek et al.,E2F recognition sequence downstream to the promoter1997). Identification of putative recognition sites does(see physical map).not prove that they are functional, but it raises the
An enhancer resides in the region flanking the XbaIpossibility that dnmt1 is regulated by central cellularsite (250 bp upstream to exon 1), which is expressed insignaling pathways (Szyf, 1998).HeLa cells, but in P-19 cells, it acts as a repressor(Fig. 4A, I and IV ). This is indicated by the ability of3.4. Identification of the regulatory regions controllingthe 2 kb HIII fragment to enhance the transcription ofthe initiation sites of dnmt1an enhancer-less vector bearing the TK promoter (pMet-P1-PrCAT, Fig. 4A, IV ) in both the sense and antisenseVerification of the four transcription initiation clus-orientations. This activity is lost once either the upstreamters identified above and demonstration of independent1.2 kb (pMet-P1-DXH-PrCAT) or 0.256 kb down-active promoter and enhancer sequences in their proxim-stream fragments (pMet-P1-DHX-PrCAT) are removedity was done using standard heterologous reporter(Fig. 4A, V and VI ). The addition of 10 kb of down-expression assays. P-19 cells were used here because theystream sequence does not affect the P1 promoter activity,bear a low background of AP-1 (de Groot et al., 1990)suggesting that all the elements required for P1 activityand Rb (Slack et al., 1993) activity. Since expression ofare contained within the minimal promoter 256 bpmurine dnmt1 has been shown to be induced by activa-upstream of the first transcription initiation site (datation of the Ras-c-Jun signaling pathway (MacLeod
et al., 1995; Rouleau et al., 1995) and to be regulated not shown). HeLa cells already have high levels of c-Jun
Fig. 3. Nucleotide sequence of 5∞-region of human dnmt1 and the position of the 5∞ boundaries of the sequenced RACE-PCR products. Bases−1454 to 317 include exon 1 and 1.5 kb 5∞ of exon 1. Bases 347 to 3122 represent a 2775 nt sequence containing exons 2, 3 and 4. Exon 1b isincluded without base numbering. Restriction enzymes utilized in the construction of different CAT constructs are indicated: XbaI and NcoI.Transcription initiation sites determined by RACE are indicated by arrowheads. Putative AP-1 binding sites are boxed with solid lines, putativeTCF-1 sites are boxed with dashed lines, the putative E2F site is underlined, and putative SP1 sites are shaded. Exons (1, 1b, 2, 3 and 4) are shownas bold letters.
414 P. Bigey et al. / Gene 242 (2000) 407–418

415P. Bigey et al. / Gene 242 (2000) 407–418
and Rb, so the addition of ectopic expression was not The three downstream promoters are either dependentor inducible by c-Jun, which is consistent with previousnecessary.
A c-Jun-dependent promoter activity resides within reports in the murine system that dnmt1 is regulated byc-Jun (MacLeod et al., 1995; Rouleau et al., 1995).160 bp upstream of the second transcription initiation
cluster (P2) identified in vivo (Fig. 4B, VII ). This is Three downstream enhancer elements that are inducibleby c-Jun were also identified. Since these elements areindicated by the ability of the XbaI–NcoI fragment
spanning 160 bp upstream and 40 bp downstream to the bidirectional, they can act in concert to activate theentire dnmt1 locus, including the 5∞ initiation sites. Thesesecond initiation cluster (pMet-P2-CAT) to direct CAT
activity when inserted into a promoter-less CAT vector multiple elements, regulated by c-Jun, may explain theinduction of dnmt1 by the Ras-AP-1 signaling pathwayin the sense orientation only in the presence of ectopic
c-Jun (Fig. 4B, VII ). A c-Jun-dependent enhancer activ- (Szyf, 1994).ity resides in the same region as demonstrated by itsability to enhance expression of CAT in both orienta-tions in a c-Jun dependent manner when inserted into a 3.5. Putative AP-1 sites form specific DNA–protein
complexesvector bearing the TK promoter (pMet-P2-PrCAT,Fig. 4B, VIII ). A single AP-1 recognition sequence isfound downstream to exon 2, which may explain the Consistent with the hypothesis that dnmt1 is transacti-
vated by c-Jun, we show that double-stranded oligonu-regulation of P2 by c-Jun. No other promoter orenhancer elements reside in the 2.4 kb upstream to the cleotides bearing either the putative AP-1 sequence
located upstream to P2 or upstream to P3 form a specificXbaI site (data not shown).c-Jun activation of both promoter and enhancer DNA-protein complex that comigrates with the complex
formed with a consensus AP-1 sequence in an EMSAelements is found in the 1.2 kb of sequence upstream ofthe third initiation cluster observed in vivo (P3), as assay (Fig. 5, oligonucleotides 2 and 3). The DNA–
protein complexes formed with the putative AP-1judged by its ability to direct CAT activity from apromoter-less CAT construct in the sense orientation sequences located upstream of P2 and P3 are competed
out by an AP-1 consensus oligonucleotide but not by a(pMet-P3-CAT, Fig. 4C, IX ) and to enhance the TKpromoter activity in both sense and antisense orienta- mismatched AP-1 sequence (Fig. 5). The putative AP-1
binding site corresponding to oligonucleotide 1 did nottions (pMet-P3-PrCAT, Fig. 4C, X ). The promoteractivity is induced by c-Jun and repressed by Rb show any relevant binding in our experiments, which is
consistent with the fact that there is no c-Jun transactiva-(Fig. 4C, IX ). The enhancer activity is augmented byc-Jun in both orientations and is repressed by Rb only tion on the region upstream to exon 1 (Fig 4A, I ).in the sense orientation (Fig. 4C, X ).
A fourth promoter resides within 170 bp upstream ofthe fourth cluster of transcription initiation identified in 3.6. Schematic diagram of transcriptional regulation of
dnmt1vivo (P4) (pMet-P4-CAT, Fig. 4D, XI). This promoteris also induced by c-Jun. The 263 bp fragment bearingP4 also contains a bidirectional enhancer (pMet- Whereas the 5∞ transcription initiation site is nested
within a CG-rich area (Fig. 6) similar to many otherP4-PrCAT, Fig. 4D, XII) that is dependent on expres-sion of ectopic c-Jun. housekeeping promoters, the downstream sites are
located within CG-poor areas. P1 has a high basalIn summary, in accordance with the hypothesis thatfour independent transcription initiation sites of dnmt1 activity and is not induced by ectopic expression of
c-Jun but is repressed by the tumor suppressor Rb. P2–are found in vivo (Figs. 1–3), we have identified fourpromoter elements upstream to each of the sites iden- P4 have a low basal activity and are significantly induced
by c-Jun (Fig. 6). A more detailed discussion of thetified in vivo. Three promoters were delineated withinless than 256 bp from the transcription initiation site. results is presented below.
Fig. 4. Analysis of dnmt1 promoter and enhancer functions. The physical map of dnmt1 5∞-CAT constructs are shown relative to the dnmt1 5∞region. Exons and transcription initiation sites are indicated. The predicted transcription initiation clusters are indicated as P1–P4. Fragmentsdeleted in each construct are indicated by a broken line. Sense (S) and antisense (AS) indicate the orientation of the genomic dnmt1 insert relativeto the CAT gene. Either HeLa or P19 cells were transfected with the indicated plasmids, and CAT activity was measured as described in Section 2.To determine whether transcription activity is controlled by either c-Jun or Rb, the cells were cotransfected as indicated with either control SKplasmid (indicated as an empty box), RSV-c-Jun (Binetruy et al., 1991) indicated as a dark box, or pLRbRNL (ATCC RB1 65003) (indicated bya hatched box). Each value is presented as the mean±S.D. of triplicate independent transfections. Similar results were observed using two differentplasmid preparations and cell line cultures. For each transfection, the values obtained for the empty vectors pOCAT or pPr-CAT were subtractedas background. (A) Promoter and enhancer functions of the P1 region. (B) Promoter and enhancer function of the P2 region. (C ) Promoter andenhancer function of the P3 region. (D) Promoter and enhancer function of the P4 region.

416 P. Bigey et al. / Gene 242 (2000) 407–418
Fig. 5. Binding of the AP-1 transcription complex to sequences in the dnmt1 5∞ upstream region. 32P-labeled double-stranded oligomers bearingeither the consensus AP-1 recognition sequence, a putative AP-1 in P1 (1): a putative AP-1 site in P2 (2) and a putative AP-1 site in P3 (3) wereincubated for 20 min at room temperature, following which, a 300-fold excess of either a consensus AP-1 or a mismatch AP-1 double-strandedoligomer competitor was added (as indicated), and the reaction was incubated for an additional 20 min. The mixtures were then analyzed byelectrophoresis on a 4% non-denaturing polyacrylamide gel. The schematic diagram below the EMSA indicates the position of the sequences testedfor ability to form an AP-1-specific complex with nuclear extracts prepared from human lung carcinoma H446 cells.
Fig. 6. Schematic diagram of transcriptional regulation of dnmt1. The first line shows the CG density of the 5∞ of the dnmt1 gene relative to thephysical map shown in the second line. Boxes (darkly shaded) indicate exons and horizontal arrows indicate transcription initiation sites. Theposition of putative binding sites for AP-1 (open circle) and E2F ( lightly shaded box) are indicated. Boundaries of the promoters as studied in thisreport (patterned boxes) and enhancers (trapezoids) are indicated. Enhancers that are dependent on c-Jun for activity are indicated. c-Jun activationis indicated by an arrow, and Rb repression is indicated by a blunted line. The region upstream to exon 1 bears an enhancer that is active in HeLacells and a repressor that is active in P19 cells.

417P. Bigey et al. / Gene 242 (2000) 407–418
4. Discussion signaling through c-Jun and Rb. The fact that the dnmt1gene is structured in this way and that this frameworkof genomic regulatory elements is conserved in theThis manuscript examines the basic modes of regula-
tion of the dnmt1 gene. Several lines of evidence point mouse, rat (unpublished data) and human pointstowards the significance of coordinating expression oftowards the conclusion that dnmt1 transcription is initi-
ated from at least four independent transcription initia- dnmt1 with other cell-cycle events (Szyf, 1998).Accumulating evidence strongly suggests that multipletion clusters. First, RACE-PCR analysis defines four
distinct 5∞ boundaries of dnmt1 transcripts in vivo mechanisms are in place to tune expression of dnmt1 toother cell cycle events such as mRNA stability (Szyf(Fig. 1). Second, RNase protection assays identify the
corresponding protected fragments (Fig. 2). Third, clon- et al., 1991) and protein–protein interactions (Chuanget al., 1997). The fact that dnmt1 bears the capacity toing and DNA sequencing of the RACE-PCR products,
recognized within dnmt1 putative binding sites for pro- respond to both proto-oncogene and tumor suppressorpathways is consistent with the hypothesis that dnmt1teins of the central cellular signaling pathways (Szyf,
1998) (Fig. 3). Fourth, promoter and enhancer elements plays an important role in oncogenic programs triggeredby either inhibition of tumor suppressors or activationwere identified in proximity to the transcription initia-
tion sites identified in vivo (within ~250 bp to P1, 2 of proto-oncogenes.In summary, our study has delineated the transcrip-and 4, and within ~1.4 kb of P3, Fig. 4). These promot-
ers were shown to be responsive to both c-Jun and Rb. tional start sites of dnmt1, the regulatory elementscontrolling its expression and their capacity to respondLastly, AP-1 putative binding sites located upstream of
the promoter elements P2 and P3, are shown to be to different cellular signals. The identification of theseelements will make it possible to establish the molecularfunctional in an EMSA assay (Fig. 5).
The putative transcription initiation sites are basis of the coordination of dnmt1 expression with othercritical biological processes.differentially utilized in vivo. The most upstream initia-
tion sites upstream to exon 1 (P1) are the most abundantin all the tissues studied here, as determined by bothRACE (26 out of 36 sequenced clones) and RNase
Acknowledgementsprotection analysis. This is consistent with previouspublished data (Tucker et al., 1996; Yoder et al., 1996).
This work was supported by a grant from theThe downstream and upstream transcription initiation
National Institute of Cancer (Canada) and a contractregions are separated by ~14 kb of genomic sequence.
with MethylGene Inc. Montreal.Whereas the 5∞ transcription initiation site is nestedwithin a CG-rich area (Fig. 6), similar to many otherhousekeeping promoters, the downstream sites arelocated within CG poor areas, as previously described Referencesin the mouse homologue (Rouleau et al., 1992). Noconsensus TATAA sequences were identified upstream Adams, R.L., McKay, E.L., Craig, L.M., Burdon, R.H., 1979. Mouse
DNA methylase: methylation of native DNA. Biochim. Biophys.to either P1 or P2–P4. Thus, the 5∞ region of dnmt1Acta 561, 345–357.bears typical TATAA-less CG-rich promoter and typical
Bagchi, S., Weinman, R., Raychaudhuri, P., 1991. RetinoblastomaTATAA-less CG-poor promoters ~14 kb downstream. protein copurifies with E2F-1, an E1A-regulated inhibitor of theP1 has a high basal activity and is not induced by transcription factor E2F. Cell 65, 1063–1072.
Bakin, A.V., Curran, T., 1999. Role of DNA 5-methylcytosine trans-ectopic expression of c-Jun (Fig. 4) but is repressed byferase in cell transformation by fos. Science 283, 387–390.the tumor suppressor Rb. P2–P4 have a low basal
Becker, P.B., Ruppert, S., Schutz, G., 1987. Genomic footprintingactivity and are significantly induced by c-Jun (Fig. 4).reveals cell type-specific DNA binding of ubiquitous factors. Cell
In addition to the four promoters, the dnmt1 5∞ region 51, 435–443.bears three enhancers that are either dependent on Bestor, T.H., Laudano, A., Mattaliano, R., Ingram, V., 1988. Cloning
and sequencing of a cDNA encoding DNA methyltransferase ofectopic expression of c-Jun or are strongly induced bymouse cells. The carboxyl-terminal domain of the mammalianit. Since enhancers act bidirectionally, they might act inenzymes is related to bacterial restriction methyltransferases.combination to induce transcription from all dnmt1J. Mol. Biol. 203, 971–983.
promoters. These multiple AP-1 response elements can Binetruy, B., Smeal, T., Karin, M., 1991. Ha-Ras augments c-Junexplain the hyperresponsiveness of dnmt1 to induction activity and stimulates phosphorylation of its activation domain.
Nature 351, 122–127.of the Ras-AP-1 signaling pathway in mouse (MacLeodBird, A.P., 1986. CpG-rich islands and the function of DNA methyla-et al., 1995; Rouleau et al., 1995) and human cells
tion. Nature 321, 209–213.(Deng et al., 1998) and the hyperactivation of dnmt1Chirgwin, J.M., Przybyla, A.E., MacDonald, R.J., Rutter, W.J., 1979.
during tumorigenesis (el-Deiry et al., 1991). Isolation of biologically active ribonucleic acid from sourcesOur analysis reveals that the dnmt1 has the capacity enriched in ribonuclease. Biochemistry 18, 5294–5299.
Chuang, L.S.H., Ian, H.I., Koh, T.W., Ngg, H.H., Xu, G., Li, B.F.L.,to respond to two nodal cellular regulatory programs

418 P. Bigey et al. / Gene 242 (2000) 407–418
1997. Human DNA-(cytosine-5) methyltransferase-PCNA com- Nan, X., Campoy, F.J., Bird, A., 1997. MeCP2 is a transcriptionalrepressor with abundant binding sites in genomic chromatin. Cellplex as a target for p21WAF1. Science 277, 1996–2000.88, 471–481.Cross, S.H., Meehan, R., Nan, X., Bird, A., 1997. A component of
Nan, X., Ng, H.-H., Jonson, C.A., Laherty, C.D., Turner, B.M., Eisen-the transcriptional repressor MeCP1 shares a motif with DNAman, R.N., Bird, A., 1998. Transcriptional repression by themethyltransferase and HRX proteins. Nat. Genet. 16, 256–259.methyl-CpG-binding protein MeCP2 involves a histone deacetylaseCumaraswamy, G., Lennon, G., Trask, B.J., Celano, P., Baylin, S.B.,complex. Nature 393, 386–389.1992. Isolation and characterization of the cDNA encoding human
Ramchandani, S., MacLeod, A.R., Pinard, M., von Hofe, E., Szyf,DNA methyltransferase. Nucleic Acids Res. 20, 2287–2291.M., 1997. Inhibition of tumorigenesis by a cytosine-DNA methyl-de Groot, R.P., Kruyt, F.A., van der Saag, P.T., Kruijer, W., 1990.transferase antisense oligodeoxynucleotide. Proc. Natl. Acad. Sci.Ectopic expression of c-Jun leads to differentiation of P19 embryo-USA 94, 684–689.nal carcinoma cells. EMBO J. 9, 1831–1837.
Ramchandani, S., Bigey, P., Szyf, M., 1998. Genomic structure of theDeng, C., Yang, J., Scott, J., Hanash, S., Richardson, B.C., 1998. Rolehuman DNA methyltransferase gene. Biol. Chem. 379, 535–540.of the ras-MAPK signaling pathway in the DNA methyltransferase
Razin, A., Cedar, H., 1991. DNA methylation and gene expression.response to DNA hypomethylation. Biol. Chem. 379, 1113–1120.Microbiol. Rev. 55, 451–458.Deng, J., Szyf, M., 1998. Multiple isoforms of DNA methyltransferase
Razin, A., Szyf, M., 1984. DNA methylation patterns. Formation andare encoded by the vertebrate cytosine DNA Methyltransferase.function. Biochim. Biophys. Acta 782, 331–342.J. Biol. Chem. 273, 22869–22872.
Rouleau, J., Tanigawa, G., Szyf, M., 1992. The mouse DNA methyl-el-Deiry, W.S., Nelkin, B.D., Celano, P., Yen, R.W., Falco, J.P., Ham-transferase 5∞-region. A unique housekeeping gene promoter.ilton, S.R., Baylin, S.B., 1991. High expression of the DNA methyl-J. Biol. Chem. 267, 7368–7377.transferase gene characterizes human neoplastic cells and
Rouleau, J., MacLeod, A.R., Szyf, M., 1995. Regulation of the DNAprogression stages of colon cancer. Proc. Natl. Acad. Sci. USA 88,methyltransferase by the Ras-AP-1 signaling pathway. J. Biol.3470–3474.Chem. 270, 1595–1601.Frohman, M.A., Downs, T.R., Chomczynski, P., Frohman, L.A.,
Sapienza, C., Peterson, A.C., Rossant, J., Balling, R., 1987. Degree of1989. Cloning and characterization of mouse growth hormone-methylation of transgenes is dependent on gamete of origin. Nature
releasing hormone (GRH) complementary DNA: increased GRH328, 251–254.
messenger RNA levels in the growth hormone-deficient lit/litSeiser, C., Knofler, M., Rudelstorfer, I., Haas, R., Wintersherger, E.,
mouse. Mol. Endocrinol. 3, 1529–1536.1989. Mouse thymidine kinase: the promoter sequence and the gene
Jones, P.L., Veenstra, G.J.C., Wade, P.A., Vermaak, D., Kass, S.U., and pseudogene structures in normal cells and in thymidine kinaseLandsberger, N., Strouboulis, J., Wolffe, A.P., 1998. Methylated deficient mutants. Nucleic Acids Res. 17, 185–195.DNA and MeCP2 recruit histone deacetylase to repress transcrip- Slack, R.S., Hamel, P.A., Bladon, T.S., Gill, R.M., McBurney, M.W.,tion. Nat. Genet. 19, 187–191. 1993. Regulated expression of the retinoblastoma gene in
Korinek, V., Barker, N., Morin, P.J., van Wichen, D., de Weger, R., differentiating embryonal carcinoma cells. Oncogene 8, 1585–1591.Kinzler, K.W., Vogelstein, B., Clevers, H., 1997. Constitutive tran- Szyf, M., 1994. DNA methylation properties: consequences for phar-scriptional activation by a beta-catenin-Tcf complex in APC−/− macology. Trends Pharmacol. Sci. 7, 233–238.colon carcinoma. Science 275, 1784–1787. Szyf, M., 1998. Targeting DNA methyltransferase in cancer. Cancer
Kreig, P.A., Melton, D.A., 1987. In vitro RNA synthesis with SP6 Metastasis Rev. 17, 219–231.RNA polymerase. Meth. Enzymol. 155, 397–415. Szyf, M., Bozovic, V., Tanigawa, G., 1991. Growth regulation of
Lloyd, A., Anchovy, N., Wasylik, B., 1991. Transformation suppressor mouse DNA methyltransferase gene expression. J. Biol. Chem. 266,activity of a c-Jun transcription factor lacking its activation 10027–10030.domain. Nature 352, 635–638. Tucker, K.L., Talbot, D., Lee, M.A., Leonhardt, H., Jaenisch, R.,
MacLeod, A.R., Szyf, M., 1995. Expression of an antisense to the 1996. Complementation of methylation deficiency in embryonicDNA methyltransferase mRNA induces DNA demethylation and stem cells by a DNA methyltransferase minigene. Proc. Natl. Acad.inhibits tumorigenesis. J. Biol. Chem. 270, 8037–8043. Sci. USA 93, 12920–12925.
MacLeod, A.R., Rouleau, J., Szyf, M., 1995. Regulation of DNA Yoder, J.A, Yen, R.W.C., Vertino, P.M., Bestor, T.H., Baylin, S.B.,methylation by the Ras signaling pathway. J. Biol. Chem. 270, 1996. New 5∞ regions of the murine and human genes for DNA11327–11337. (cytosine-5)-methyltransferase. J. Biol. Chem. 271, 31092–31097.
Mertineit, C., Yoder, J.A., Taketo, T., Laird, D.W., Trasler, J.M., Zuccotti, M., Monk, M., 1995. Methylation of the mouse Xist gene inBestor, T.H., 1998. Sex-specific exons control DNA methyltransfer- sperm and eggs correlates with imprinted Xist expression and pater-
nal X-inactivation. Nat. Genet. 9 (316), 320–1995.ase in mammalian germ cells. Development 125, 889–897.
Copyright © 2022 FDOKUMEN




















