
Misguided Transcriptional Elongation Causes Mixed Lineage Leukemia
Upload
independentCategory
view
7download
0
Immunity, Vol. 18, 801–811, June, 2003, Copyright 2003 by Cell Press
Transcriptional Regulation of Th2 Differentiationby Inducible Costimulator
IFN� and lymphotoxin (LT) to mediate cellular immunity.Th2 cells, on the other hand, express IL-4, -5, and -13,whose genes are localized in the same chromosome
Roza I. Nurieva,1 Julie Duong,1 Hiroko Kishikawa,2
Umberto Dianzani,3 Jose M. Rojo,4 I-cheng Ho,5
Richard A. Flavell,6 and Chen Dong1,*region. They regulate humoral immunity and, in patho-1Department of Immunologylogical conditions, asthma and allergy. Th activation andUniversity of Washington School of Medicinedifferentiation is an essential step of T-dependent im-Seattle, Washington 98195mune responses. Work in the past decade has revealed2 Department of Immunology and Infectious Diseasescomplex regulation of the Th differentiation program,Harvard School of Public Healthwhich includes T cell receptor (TcR) and costimulatoryBoston, Massachusetts 02115receptor signals, polarizing cytokines and lineage-spe-3 Interdisciplinary Research Centercific transcriptional factors, and signal transductionof Autoimmune Diseases andpathways (Constant and Bottomly, 1997; Dong and Fla-Department of Medical Sciencesvell, 2000a, 2000b; Glimcher and Murphy, 2000).A.Avogadro University of Eastern Piedmont
IL-4, produced by activated T cells and perhaps some28100 Novaraother cell types in vivo, is the determining factor for Th2Italydifferentiation. In this regard, IL-4 autoregulates its own4 Immunology Departmentexpression during Th differentiation. The IL-4 receptorCentro de Investigaciones Biologicassignals through Janus kinases (JAK) to induce phos-Consejo Superior de Investigaciones Cientı́ficasphorylation and nuclear translocation of signal trans-Velazquez, 144ducer and activator of transcription 6 (STAT6). STAT6-E-28006 Madriddeficient T cells are impaired in Th2 differentiation andSpaincytokine production (Kaplan et al., 1996; Shimoda et al.,5 Division of Rheumatology, Immunology, and Allergy1996; Takeda et al., 1996). The mechanism of STAT6Department of Medicineautoregulation of IL-4 expression has not been well es-Brigham and Women’s Hospitaltablished, since until now a fully functional STAT6 bind-Boston, Massachusetts 02115ing site in the Th2 locus has not been well defined.6 Division of ImmunobiologyOn the other hand, Kurata et al. showed that activatedYale University School of MedicineSTAT6 induced expression of Th2-specific lineage fac-Howard Hughes Medical Institutetors GATA3 and c-Maf (Kurata et al., 1999), suggestingNew Haven, Connecticut 06520that STAT6 action is indirect. GATA3 is a master regula-tor of Th2 differentiation—it is both necessary and suffi-cient for Th2 development (Zhang et al., 1997, 1999;SummaryZheng and Flavell, 1997). GATA3 is expressed by naiveCD4� T cells, and this expression is sustained in Th2Helper T (Th) cell differentiation is accompanied bycells but repressed in Th1 by IL-12 (Ouyang et al., 1998;complex transcriptional changes. Although costimula-Zheng and Flavell, 1997). GATA3 binds to the IL-5 buttory receptors are important in Th differentiation, thenot the IL-4 proximal promoter (Lee et al., 2001; Zhangunderlying mechanisms are poorly understood. Hereet al., 1997), and it is widely believed to modulate chro-we examine the transcriptional mechanisms by whichmatin accessibility of the entire Th2 locus (Lee et al.,ICOS regulates Th2 differentiation and selective IL-42000, 2001; Ouyang et al., 2000; Takemoto et al., 2000).expression by effector T cells. We found impaired ex-Furthermore, GATA3 also inhibits IFN� expression (Ou-
pression of c-Maf transcription factor functionally as-yang et al., 1998), probably through downregulation of
sociated with the IL-4 defect in ICOS�/� cells. c-Maf STAT4 expression (Usui et al., 2003). c-Maf was identi-expression in effector cells was regulated by IL-4 lev- fied as the first Th2-specific transcription factor thatels during Th differentiation. ICOS costimulation po- binds to the IL-4 proximal promoter (Ho et al., 1996).tentiated the T cell receptor (TcR)-mediated initial IL-4 Studies of c-Maf knockout mice and overexpressionproduction, possibly through the enhancement of transgenic mice indicate that it selectively regulates IL-4NFATc1 expression. These data indicate that ICOS, expression (Ho et al., 1996; Kim et al., 1999). c-Mafby enhancing TcR signals at an early stage of T cell probably acts in synergy with the nuclear factor of ac-activation, regulates IL-4 transcription and T cell func- tivated T cells (NFAT) and activating protein (AP)-1 pro-tion in effector cells. teins (Ho et al., 1996; Li et al., 1999), which are down-
stream of TcR signaling. A member of the NFAT family,Introduction NFATc1 (also called NFAT2 or NFATc), although not
expressed in a Th2-specific fashion, plays an importantAfter activation, CD4 helper T (Th) cells differentiate into role in Th2 differentiation and cytokine expressionTh1 or Th2 effector cells, which specialize in producing (Ranger et al., 1998; Yoshida et al., 1998). NFATc1, asdistinct cytokines to mediate different types of immune well as NFATc2, 3, and 4 proteins, is regulated by phos-responses (Paul and Seder, 1994). Th1 cells produce phorylation-dependent subcellular localization (Crabtree
and Olson, 2002; Rao et al., 1997). TcR-activated phos-phatase calcineurin dephosphorylates NFATc proteins,*Correspondence: [email protected]

Immunity802
which leads to their nuclear localization. Blockade of fector cells, expression of the c-Maf transcription factorcalcineurin action resulted in selective Th2 defects (Ya- is selectively impaired. ICOS, by enhancing NFATc1 ex-mashita et al., 2000). On the other hand, loss of JNK1, pression and initial IL-4 production during early T cella specific kinase of NFATc1, exacerbated Th2 differenti- differentiation, regulates c-Maf expression in the ef-ation and cytokine expression (Chow et al., 2000; Dong fector phase. These data provide a connection betweenet al., 1998). Th2 differentiation is therefore regulated a costimulatory receptor and the transcriptional machin-by transcription factors downstream of TcR and IL-4, ery mediating Th2 differentiation and effector IL-4 ex-the interplay of which is not well defined. pression.
T cell activation and differentiation requires two sig-nals: the first derives from the TcR and the second from Resultscostimulatory receptors, the most important beingCD28. When bound by its ligands CD80 and CD86, which c-Maf Expression Is Selectively Impairedare expressed on activated antigen-presenting cells in ICOS�/� Effector T Cells(APC), CD28 plays an essential role in the early phase ICOS�/� effector T cells exhibited a selective IL-4 deficitof T cell activation, including expression of IL-2 and in vitro and in vivo. In the current study, we address thebcl-xL upregulation (Alegre et al., 2001; Carreno and molecular mechanisms that account for this IL-4 defect.Collins, 2002; Sharpe and Freeman, 2002). CD28 costim- In our previous study, we differentiated CD4� naiveulation also favors Th2 differentiation (Lenschow et al., T cells with anti-CD3 and irradiated wild-type splenic1996), but the underlying transcriptional mechanism is APC (Dong et al., 2001a). Exogenous IL-2 was added tonot clear. The function of CD28 at the effector phase is compensate for the proliferative defect by the ICOS-less dominant. This is probably due to the induction of deficient cells. Under these differentiation conditions,its antagonist, CTLA-4, on activated T cells, which binds costimulation was provided by APC. After 4 days ofthe same ligands with much higher affinity and functions differentiation, T cells were extensively washed and re-to inhibit T cell proliferation and cytokine expression stimulated with anti-CD3. ICOS�/� cells failed to pro-(Chambers and Allison, 1999). ICOS is the third member duce IL-4 when restimulated. This result suggests thatof the CD28 family, and its expression is induced after ICOS functions during the 4 day Th differentiation pro-T cell activation and is highly dependent on CD28 stimu- cess by somehow reprogramming the IL-4 inductionlation (Hutloff et al., 1999; McAdam et al., 2000; Yoshi- machinery downstream of TcR. We also found that re-naga et al., 1999). In the effector phase, ICOS is ex- stimulation of differentiated cells in the presence of APCpressed at high levels by Th2 cells and at low levels by did not alter the cytokine expression profiles in this sys-Th1 cells in mice (Coyle et al., 2000). The ligand for ICOS, tem (C.D., unpublished data), supporting a general ideaB7h, has been identified (Swallow et al., 1999; Yoshinaga that the function of effector cells is relatively costimula-et al., 1999). It is constitutively expressed on B cells and tion independent. Since the expression of IL-5 and IL-induced in nonlymphoid tissues by the inflammatory 10 Th2 cytokines was not severely affected by ICOScytokine TNF� (Swallow et al., 1999). gene targeting, we hypothesized that ICOS may act on
Recently, we and other groups constructed and ana-some IL-4-specific transcription machinery; for in-
lyzed mice deficient in the ICOS gene (Dong et al., 2001a;stance, on IL-4 promoter activity instead of global chro-
McAdam et al., 2001; Tafuri et al., 2001). ICOS-deficientmatin remodeling of the Th2 cytokine locus. Although
mice exhibited impaired humoral immunity and germinalboth the IL-4 and IL-5 promoters have functional AP-center reactions (Dong et al., 2001a, 2001b; McAdam1 elements, the IL-4 promoter is bound and stronglyet al., 2001; Tafuri et al., 2001). On the other hand, weactivated by c-Maf and NFAT (Glimcher and Murphy,found that ICOS�/� mice were extremely sensitive to2000; Ho et al., 1996); IL-5, by GATA3 (Zhang et al.,experimental autoimmune encephalomyelitis (EAE)1997).(Dong et al., 2001a). These defects can be attributable
We initiated our study by addressing what accountedto the observed Th2 cytokine defects by ICOS�/� T cellsfor the IL-4 defect by the ICOS�/� effector T cells. We(Dong et al., 2001a; Tafuri et al., 2001). We found thatdifferentiated wild-type and knockout naive T cells forICOS�/� T cells were selectively impaired in IL-4 expres-4 days as described above under neutral condition assion after in vitro differentiation or in vivo priming bywell as those favoring Th1 (IL-12 plus anti-IL-4) or Th2protein antigen in complete Freund’s adjuvant or Alum,(IL-4 plus anti-IFN�) differentiation (Dong et al., 2001a).yet capable of secreting IL-5 (Dong et al., 2001a). Con-The differentiated T cells were extensively washed andsistent with this in a murine asthma model, we foundrestimulated for 4 hr with anti-CD3 (Figure 1), and nu-ICOS knockout mice with normal lung eosinophil infiltra-clear extracts were prepared from them or naive T cells.tion, which is mediated by IL-5, and that they were defi-The expression of several key Th2-acting transcriptioncient in IL-4-dependent IgE production (Dong et al.,factors was analyzed by Western blots, including2001a). Therefore, ICOS does not appear to regulate theGATA3, c-Maf, NFATc1, and JunB (Figure 1). GATA-3 isglobal Th2 differentiation program; instead, it is requiredexpressed by naive T cells as well as effector T cellsfor expression of IL-4 at the effector phase. The role ofdifferentiated under neutral (Th0) or Th2 conditions, butICOS is to fine-tune antibody production, and eosinophilnot by Th1 cells (Figure 1). T-bet is not expressed bygeneration and tissue infiltration, both of which are me-naive T or Th2 cells but by effector Th0 or Th1 cellsdiated by Th2 cells, however with distinct biological/(Figure 1). We found no significant differences betweenpathological function.wild-type and knockout T cells in expression of theseIn this study, we address the transcriptional mecha-two lineage-determining factors (Figure 1) or STAT6nisms by which ICOS regulates IL-4 gene expression
and Th2 differentiation. We found that, in ICOS�/� ef- (data not shown), in keeping with our earlier results that

ICOS Regulation of IL-4 Transcription803
expression by ICOS�/� effector T cells (Figure 2B). Inaddition, it completely restored the IL-4 defect byICOS�/� effector T cells (Figure 2B). All these resultsclearly support that the IL-4 defect by ICOS�/� effectorT cells is functionally associated with a c-Maf deficiency.
c-Maf Expression Is Regulated by IL-4 Levelsduring Th DifferentiationThe above analysis established a linkage between acostimulatory receptor, ICOS, and a Th2-specific tran-scription factor, c-Maf, which we have not seen reportedbefore. ICOS can regulate c-Maf directly or indirectly.We have previously noticed that the IL-4 defect ofICOS�/� effector cells could be rescued when these cellswere differentiated with high levels of exogenous IL-4(Dong et al., 2001a). We therefore examined whetherIL-4 could restore c-Maf expression. Indeed, we foundFigure 1. Defective Expression of c-Maf by ICOS�/� Cellsthat, when ICOS�/� cells were differentiated under Th2-Naive wild-type or ICOS knockout Th cells (1 � 106/ml) were acti-polarizing conditions, they expressed c-Maf at the samevated with plate-bound anti-CD3, irradiated wild-type APC (1 � 106/
ml), and IL-2 (30 units/ml) for 4 days with or without Th1 (IL-12 level as the wild-type cells (Figure 1). Thus, restorationand anti-IL-4) or Th2 (IL-4 plus anti-IFN�) polarizing agents. After of c-Maf and IL-4 expression in ICOS�/� cells correlate.extensive washing, the differentiated cells were restimulated with The above result not only supports the data presentedanti-CD3 for 4 hr, and nuclear extraction was prepared. The expres- above that ICOS regulates IL-4 through c-Maf but alsosion of transcription factors (GATA3, c-Maf, NFATc1, and JunB) was
suggests that c-Maf expression is regulated by theanalyzed by Western blot analysis. JNK and �-actin antibodies wereamount of IL-4 during Th differentiation. To assess this,used to indicate equal protein loading.we purified naive CD4� T cells from IL-4 knockout miceand differentiated them with different amounts of exoge-nous IL-4. Since IL-4 is not produced endogenously, theICOS is not essential for global Th1 or Th2 differentiationamount of IL-4 during Th differentiation is thus precisely(Dong et al., 2001a). The only factor we found consis-predetermined. When we examined c-Maf expressiontently and significantly reduced in expression in ICOS�/�
in cells differentiated with different amounts of IL-4, weeffector Th0 cells was c-Maf (Figure 1). Interestingly,found that the levels of c-Maf expression by effectorgene targeting and overexpressing transgenic studiesT cells correlated very well with the amount of IL-4 addedindicate that c-Maf is an IL-4-specific transcription fac-(Figure 3). High c-Maf expression was achieved in ef-tor (Ho et al., 1996; Kim et al., 1999), and c-Maf knockoutfector cells when there were high IL-4 levels during Thmice appeared to share a similar phenotype as ICOS-differentiation. On the other hand, expression of GATA3deficient mice (Kim et al., 1999). Our result, therefore,did not appear to be as sensitive to the IL-4 levels duringsuggests that ICOS may regulate IL-4 productionTh differentiation, as determined by Western blot analy-through c-Maf expression.sis (Figure 3) or real-time PCR (data not shown). GATA3expression by naive cells is sustained during Th2 differ-c-Maf Overexpression Restores Normal IL-4entiation (Zheng and Flavell, 1997); the termination ofExpression in ICOS�/� Effector T CellsGATA3 in Th1 cells is mediated by IL-12 (Ouyang etTo assess whether c-Maf is the target transcription fac-al., 1998). In our neutral differentiation system, sincetor regulated by ICOS, we overexpressed c-Maf retrovi-exogenous IL-12 is not provided, GATA3 expression isrally in wild-type and knockout T cells. Naive T cellslikely to be maintained and not significantly responsivewere stimulated with anti-CD3, APC, and IL-2 for 24–36to changes in IL-4 concentration during Th differentia-hr, infected with GFP-only or c-Maf-GFP virus, and fur-tion. It is also possible that IL-13 plays a compensatoryther differentiated for 2–3 days with IL-2. GFP-express-role in maintaining GATA-3 expression in this experi-ing cells were FACS-sorted and restimulated with anti-ment. These data indicate that the amount of IL-4 pro-CD3 for 24 hr. IL-4 expression was measured by ELISA.duced by differentiating Th cells can regulate c-Maf ex-Wild-type cells infected with GFP-only retroviruses pro-pression in effector cells. It also correlates well withduced 10-fold more IL-4 than knockout counterpartsan early report in which active STAT6 induced c-Maf(Figure 2A). c-Maf overexpression in wild-type cellsexpression (Kurata et al., 1999).moderately increased the levels of IL-4 expression (Fig-
ure 2A). On the other hand, knockout cells infected withthe c-Maf-GFP virus produced IL-4 at a similar level with ICOS Regulates the Initial IL-4 Expression
during Th Differentiationthe wild-type cells treated in the same manner (Figure 2A).To further substantiate the above result, we crossed The data described above indicate a role for IL-4, pro-
duced by differentiating T cells, to regulate c-Maf andc-Maf transgenic mice with ICOS knockout mice. NaiveCD4� T cells from ICOS�/� and ICOS�/� mice with or hence IL-4 expression levels in the effector phase. They
also suggest that ICOS may regulate c-Maf expressionwithout c-Maf transgene were differentiated under theconditions stated above for 4 days followed by restimu- indirectly by mediating IL-4 production by activated
T cells during differentiation. To test this, we differenti-lation. As measured by ELISA, c-Maf transgene signifi-cantly increased IL-4 expression and decreased IFN� ated wild-type and ICOS�/� naive Th cells as above and

Immunity804
Figure 2. c-Maf Overexpression Restores IL-4 Expression in ICOS�/� Cells
(A) Naive wild-type or ICOS knockout Th cells were activated with plate-bound anti-CD3, irradiated wild-type APC, and IL-2 for 36 hr, infectedwith GFP-only or c-Maf-GFP virus, and further differentiated for 2 days with IL-2. GFP-expressing cells were FACS sorted and restimulatedwith anti-CD3 for 24 hr, and IL-4 production was assayed by ELISA.(B) Naive T cells from ICOS�/� and ICOS�/� mice with or without a c-Maf transgene were differentiated, and effector cytokine expression wasmeasured as in (A).
harvested cells 1 day after activation for measurement When we used a combination of anti-CD3, CD28, andICOS, we found neither enhancement of T cell prolifera-of IL-4 and IFN� cytokines. Since IL-4 and IFN� can
be consumed by differentiating cells, we measured the tion nor IFN� production as compared to cells treatedonly with anti-CD3 and CD28 (Figure 4C). This suggestscytokine messenger RNA directly by real-time RT-PCR
analysis. We found that, at this early phase of Th differ- that maximum CD28 costimulation can achieve maxi-mum T cell proliferation and Th1 cytokine expression.entiation, ICOS�/� cells produced roughly similar
amounts of IFN� as wild-type; their IL-4 production was On the other hand, IL-4 production by activated T cellswas approximately doubled with ICOS costimulationdetectable but significantly reduced (Figure 4A). A re-
duction of IL-4 expression by differentiating ICOS�/� Th (Figure 4C). This analysis indicates that ICOS costimula-tion could further increase IL-4 expression by activatedcells was also observed 2 and 3 days after Th differentia-
tion (data not shown). IL-4 expression by cells after 1 T cells, despite a lack of enhancement of proliferationand Th1 cytokine expression.day differentiation was not dependent on IL-4 signaling,
as presence or absence of an anti-IL-4-blocking anti-body in the Th cell culture gave a similar result for both ICOS Regulates NFATc1 Expressionwild-type and ICOS�/� cells (Figure 4B). This result indi- during Th Differentiationcates that, in the absence of ICOS, the IL-4-independent The above observation indicates that ICOS costimula-initial IL-4 production by differentiating T cells is defec- tion regulates IL-4-independent initial IL-4 productiontive, which could result in impaired expression of c-Maf during Th differentiation, which could determine theat the effector phase.
To substantiate the above finding, we further testedthe ability of ICOS costimulation to induce IL-4. C398-4A is a monoclonal antibody against ICOS (Buonfiglioet al., 2000; Redoglia et al., 1996); its specificity wasfurther confirmed by us previously based on its lackof binding to ICOS�/� cells (Dong et al., 2001b). Westimulated naive Th cells from C57BL/6 mice with differ-ent combinations of anti-CD3, CD28 (or its agonist,B7.1-Ig), and ICOS. CD28 costimulation greatly en-hanced anti-CD3-initiated T cell proliferation (Figure 4C)
Figure 3. c-Maf Expression in Effector Th Cells Is Regulated by theand IL-2 production (data not shown), while anti-ICOSAmount of IL-4 during Th Differentiationdid not have a significant effect (Figure 4C). It has beenNaive CD4� cells from IL-4 knockout mice were activated with differ-previously shown that efficient ICOS expression de-ent amounts of exogenous IL-4 (0, 50, 500, and 5000 pg/ml). Afterpends on CD28 costimulation (McAdam et al., 2000).extensive wash, the differentiated cells were restimulated with plate-
Therefore, during the above-mentioned stimulation of bound anti-CD3 for 4 hr, and nuclear extraction was prepared. Thehighly purified naive T cells, we may not obtain signifi- amount of c-Maf was examined by Western blot analysis. Anti-JNK
was used to indicate equal protein loading.cant ICOS expression without CD28 costimulation.
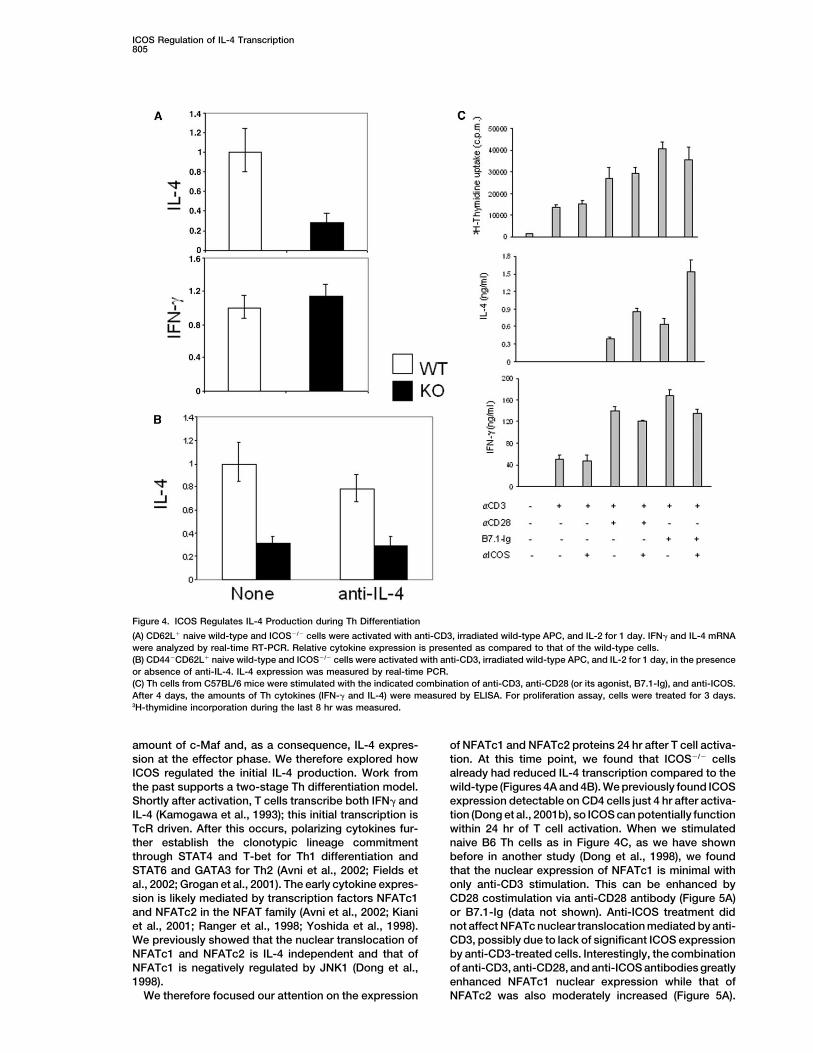
ICOS Regulation of IL-4 Transcription805
Figure 4. ICOS Regulates IL-4 Production during Th Differentiation
(A) CD62L� naive wild-type and ICOS�/� cells were activated with anti-CD3, irradiated wild-type APC, and IL-2 for 1 day. IFN� and IL-4 mRNAwere analyzed by real-time RT-PCR. Relative cytokine expression is presented as compared to that of the wild-type cells.(B) CD44�CD62L� naive wild-type and ICOS�/� cells were activated with anti-CD3, irradiated wild-type APC, and IL-2 for 1 day, in the presenceor absence of anti-IL-4. IL-4 expression was measured by real-time PCR.(C) Th cells from C57BL/6 mice were stimulated with the indicated combination of anti-CD3, anti-CD28 (or its agonist, B7.1-Ig), and anti-ICOS.After 4 days, the amounts of Th cytokines (IFN-� and IL-4) were measured by ELISA. For proliferation assay, cells were treated for 3 days.3H-thymidine incorporation during the last 8 hr was measured.
amount of c-Maf and, as a consequence, IL-4 expres- of NFATc1 and NFATc2 proteins 24 hr after T cell activa-tion. At this time point, we found that ICOS�/� cellssion at the effector phase. We therefore explored how
ICOS regulated the initial IL-4 production. Work from already had reduced IL-4 transcription compared to thewild-type (Figures 4A and 4B). We previously found ICOSthe past supports a two-stage Th differentiation model.
Shortly after activation, T cells transcribe both IFN� and expression detectable on CD4 cells just 4 hr after activa-tion (Dong et al., 2001b), so ICOS can potentially functionIL-4 (Kamogawa et al., 1993); this initial transcription is
TcR driven. After this occurs, polarizing cytokines fur- within 24 hr of T cell activation. When we stimulatednaive B6 Th cells as in Figure 4C, as we have shownther establish the clonotypic lineage commitment
through STAT4 and T-bet for Th1 differentiation and before in another study (Dong et al., 1998), we foundthat the nuclear expression of NFATc1 is minimal withSTAT6 and GATA3 for Th2 (Avni et al., 2002; Fields et
al., 2002; Grogan et al., 2001). The early cytokine expres- only anti-CD3 stimulation. This can be enhanced byCD28 costimulation via anti-CD28 antibody (Figure 5A)sion is likely mediated by transcription factors NFATc1
and NFATc2 in the NFAT family (Avni et al., 2002; Kiani or B7.1-Ig (data not shown). Anti-ICOS treatment didnot affect NFATc nuclear translocation mediated by anti-et al., 2001; Ranger et al., 1998; Yoshida et al., 1998).
We previously showed that the nuclear translocation of CD3, possibly due to lack of significant ICOS expressionby anti-CD3-treated cells. Interestingly, the combinationNFATc1 and NFATc2 is IL-4 independent and that of
NFATc1 is negatively regulated by JNK1 (Dong et al., of anti-CD3, anti-CD28, and anti-ICOS antibodies greatlyenhanced NFATc1 nuclear expression while that of1998).
We therefore focused our attention on the expression NFATc2 was also moderately increased (Figure 5A).

Immunity806
Figure 5. ICOS Regulates NFATc Nuclear Lo-calization
(A) Naive B6 Th cells were stimulated as inFigure 4C for 1 day, and nuclear factors wereisolated. The expression of nuclear factorswas examined by Western blot analysis.(B) Naive wild-type and ICOS�/� Th cells werestimulated as in Figure 4A for 24 hr. The nu-clear fraction of these cells was prepared,and the amount of nuclear factors (NFATc1,NFATc2, GATA3, and JNK) was analyzed byWestern blot analysis.
Although NFATc2-deficient cells have exaggerated Th2 antibodies in the absence or presence of anti-ICOS.NFATc1 expression was determined by Western blottingresponses, their nuclear localization could result in IL-4
expression (Monticelli and Rao, 2002). In addition, cal- on days 0, 1, 2, and 3 after treatment (Figure 6D). Anti-ICOS treatment greatly enhances NFATc1 expressioncineurin signaling generally favors Th2 differentiation
(Yamashita et al., 2000). This result suggests that the on day 1 but not on subsequent days of T cell activation.This result indicates that ICOS has a very early effectincrease in IL-4 expression by ICOS costimulation is
likely mediated by enhanced NFATc nuclear expression. on NFATc1 induction, and it may function to accelerateNFATc1 and IL-4 induction. This early costimulation byWhen we examined wild-type and ICOS�/� Th cells
24 hr after in vitro differentiation, we found that ICOS�/� ICOS possibly acts through c-Maf to impact IL-4 expres-sion at the effector phase.T cells had less NFATc1 and NFATc2 in the nucleus
(Figure 5B). In view of the fact that NFATc proteins,especially NFATc1, are critical for IL-4 expression andTh2 differentiation (Ranger et al., 1998; Yoshida et al., Discussion1998), ICOS may synergize with the TcR and CD28 path-ways to mediate the initial IL-4 expression probably Th cell differentiation, a crucial step in immune re-
sponses, is the result of complex cellular and molecularthrough enhancing the amount of NFATc in the nucleusof activated T cells. Consistent with this notion, we regulation. Recent years have seen progress in under-
standing the transcriptional mechanisms of Th linagepreviously found that enhanced NFATc1 nuclear local-ization by Jnk1 gene deletion led to exacerbated IL-4 commitment and effector cytokine expression. How-
ever, how these T cell-intrinsic mechanisms are regu-expression (Chow et al., 2000; Dong et al., 1998).NFATc1, unlike NFATc2, is induced after T cell activa- lated by environmental stimuli, such as costimulation
by APC, needs further definition. CD28 costimulation oftion. To assess whether ICOS regulates NFATc1 totalexpression or merely its nuclear localization, we also T cells is important for their full activation. In addition,
Th2 differentiation is more dependent on the CD28 sig-measured the amount of NFATc1 in the total lysates ofTh cells either costimulated with or without anti-ICOS, naling than Th1 (Lenschow et al., 1996). The basis for
this differential dependency is unclear. We showed ear-or differentiating Th cells from wild-type or knockoutmice. We found that the expression of NFATc1 is regu- lier that CD28 costimulation could enhance nuclear lo-
calization of NFATc1 (Dong et al., 1998) (Figure 5A),lated by ICOS. Th cells costimulated with anti-ICOS haddramatically increased NFATc1 expression compared suggesting that it synergizes with TcR signals in NFAT
regulation.to those stimulated with only anti-CD3 and anti-CD28(Figure 6A) or B7.1-Ig (data not shown). This action of We have recently shown that ICOS is required for
humoral immune responses and for immune toleranceICOS can be blocked by cyclosporine A, an inhibitor ofcalcineurin (Figure 6B) but independent of IL-4 signaling, against inflammatory EAE diseases (Dong et al., 2001a,
2001b). Interestingly, in this study ICOS is required foras STAT6-deficient cells exhibited the same amount ofNFATc1 expression as the wild-type after treatment with selective cytokine expression in the effector phase,
most importantly IL-4, which could account for severalanti-CD3, CD28, and ICOS antibodies (data not shown).On the other hand, cells lacking ICOS exhibited reduced key immune defects in the knockout mice. In the current
paper, we describe the molecular mechanisms by whichamounts of NFATc1 compared to the wild-type 24 hrafter T cell activation (Figure 6C). ICOS therefore en- ICOS regulates IL-4 expression at the effector phase.
We show that the expression of the c-Maf transcriptionhances TcR and CD28 signaling in upregulation ofNFATc1 expression. However, despite this early effect, factor is selectively impaired in ICOS�/� Th cells. More-
over, c-Maf expression in effector cells is regulated bywe did not find NFATc1 nuclear (Figure 1) or total (datanot shown) expression to be significantly different be- the amount of IL-4 present during Th differentiation.
ICOS, possibly by enhancement of NFATc1 expressiontween wild-type and ICOS�/� effector T cells. To exam-ine whether ICOS costimulation is only required in the at a very early stage of T cell activation, enhances initial
IL-4 expression to regulate the c-Maf and IL-4 expres-early phase of T cell activation for NFATc1 induction,we activated naive Th cells with anti-CD3 and anti-CD28 sion in the effector phase. This study provides an insight

ICOS Regulation of IL-4 Transcription807
Figure 6. ICOS Regulates NFATc1 Expression
(A and B) Naive B6 Th cells were stimulated as in Figure 4C for 1 day (A) or in the presence/absence of cyclosporine A (B).(C) Naive wild-type and ICOS�/� Th cells were stimulated as in Figure 4A for 24 hr. Total cell extract was prepared and the amount of NFATc1was examined by Western blot analysis.(D) Naive Th cells were stimulated with anti-CD3 and anti-CD28 in the absence or presence of anti-ICOS. NFATc1 expression was examinedat days 0, 1, 2, and 3 after activation.
into how Th2 differentiation is mechanistically regulated entiation culture (Figure 3). IL-4 regulation of c-Maf isfurther supported by a previous report that a constitu-by a costimulatory receptor.
c-Maf is the first transcription factor identified to be tively active form of STAT6 induces c-Maf expression(Kurata et al., 1999). In view of the fact that c-Maf isrestrictively expressed in Th2 cells (Ho et al., 1996). It
binds to the IL-4 proximal promoter and activates it in required for effector IL-4 production, this work also sug-gests that c-Maf is involved in IL-4 autoregulation, butsynergy with NFAT and AP-1 factors (Ho et al., 1996; Li
et al., 1999). Earlier studies revealed that c-Maf serves how IL-4/STAT6 regulate c-Maf remains to be resolved.GATA3 may be a candidate downstream of STAT6, sinceas an IL-4-specific transcription factor. c-Maf-deficient
T cells were impaired in IL-4 expression at the effector Ouyang et al. (2000) showed that STAT6-deficient Th1cells had more reduced c-Maf expression than the wild-phase, while the expression of other Th2 cytokines were
either moderately reduced or not affected (Kim et al., type, and that GATA3 overexpression led to c-Maf ex-pression independent of STAT6 (Ouyang et al., 2000).1999). In the current study, we indicate that ICOS medi-
ates effector IL-4 production through regulating c-Maf Furthermore, we found that ICOS regulates the initialIL-4 production by newly activated T cells. ICOS�/� Thexpression in effector Th cells. We first found that
ICOS�/� effector Th0 cells, which are impaired in IL-4 cells expressed less IL-4 than the wild-type cells 24 hrafter activation with anti-CD3, APC, and IL-2 (Figureexpression, had a drastic reduction in c-Maf expression
(Figure 1). Second, Th2-skewing differentiation rescued 4A), indicating that ICOS is required for optimal IL-4expression early during Th differentiation. This early IL-4the IL-4 as well as c-Maf defects in ICOS�/� effector Th
cells (Dong et al., 2001a; Figure 1). Last, overexpression expression is independent of IL-4 signaling as additionof an anti-IL-4 antibody had no effect on IL-4 expressionof c-Maf alone in ICOS�/� cells, retrovirally or via a trans-
gene, restored the IL-4 expression to the same level as by wild-type or ICOS-deficient T cells (Figure 4B). Onthe other hand, ICOS costimulation of primary Th cellsthat in wild-type cells (Figure 2). These results support
c-Maf to be functionally associated with ICOS costimu- treated with anti-CD3 plus anti-CD28 or B7.1-Ig led toa greater amount of IL-4 secretion, although it did notlation during Th differentiation.
Although c-Maf has been identified as a crucial regula- greatly enhance T cell proliferation or IFN� production(Figure 4C). This result indicates an additive role for ICOStor in Th2 cells, how it is regulated has not been fully
understood. Our results provide a linkage between this in IL-4 expression, enhancing initial IL-4 production. Itis noteworthy that IL-4 expression during Th differentia-transcriptional factor and a costimulatory receptor,
ICOS. We went on further to investigate how ICOS regu- tion is significantly reduced but not absent in ICOS�/�
cells (Figures 4A and 4B). Other factors such as TcR,lates c-Maf expression. The fact that exogenous IL-4restored c-Maf and IL-4 expression in ICOS�/� effector CD28, and IL-6 may account for this remaining IL-4 ex-
pression (Diehl et al., 2002; Dong and Flavell, 2000a;cells led us to hypothesize that ICOS may regulate c-Mafexpression indirectly through initial IL-4 expression dur- Lenschow et al., 1996). However, the IL-4 reduction in
activated ICOS�/� Th cells may be sufficient to lead toing Th differentiation. In support of this hypothesis, wefound that c-Maf expression levels in the effector phase impaired c-Maf expression but not to global Th2 defects.
In other words, c-Maf may be more sensitive than otherare determined by the amount of IL-4 added to the differ-

Immunity808
Figure 7. Diagram of ICOS Function in Th2Differentiation
transcription factors to IL-4 regulation. In support of this NFATc1 regulation has been extensively studied,mostly at the nuclear import/export level. Calcineurin, inassumption, we found GATA3 expression to be little
affected within the IL-4 dose range we used (Figure 3). response to cellular calcium signals, dephosphorylatesfour NFATs (c1, c2, c3, and c4), which leads to theirGATA3 is expressed by naive CD4� T cells, and this
expression is sustained in Th2 cells but repressed in nuclear translocation. T cells carrying a dominant-nega-tive form of calcineurin have defective Th2 differentiationTh1 by IL-12 (Ouyang et al., 1998; Zheng and Flavell,
1997). In our differentiation system, since there is no IL- (Yamashita et al., 2000). NFATc1 cytoplasmic localiza-tion is mediated by serine/threonine kinases, including12 present, GATA3 expression is thus not very respon-
sive to IL-4 (Figure 3). Therefore, ICOS, through the regu- GSK-3 and MAP kinases (Kiani et al., 2000). While thephysiological function of GSK-3 in lymphocytes has notlation of IL-4 expression during Th cell differentiation,
regulates c-Maf expression in the effector phase, which yet been clearly elucidated, we previously reported thatNFATc1 nuclear localization in activated T cells is nega-in turn determines effector IL-4 production.
We then examined how ICOS regulates the initial CD4 tively regulated by the JNK MAP kinase (Chow et al.,2000; Dong et al., 1998). A mutant NFATc1 that cannotT cell IL-4 production. We found that ICOS regulates
NFATc1 expression. ICOS costimulation led to greater be phosphorylated by JNK is found constitutively in thenucleus and can activate IL-4 promoter activity (ChowNFATc1 expression at 24 hr after T cell activation (Figure
6), which could then result in increased NFATc1 in the et al., 2000). On the other hand, NFATc1 itself is inducedafter T cell activation, although the mechanism wherebynucleus of activated T cells (Figure 5). Nuclear transloca-
tion of NFATc2 was also moderately enhanced (Figure this occurs is not very clear. Chuvpilo et al. (2002) re-cently showed that transcription of multiple forms is5). Considering that NFATc1 has been shown to be im-
portant for Th2 differentiation (Ranger et al., 1998; Yo- regulated differently. The short isoform, NFATc1/A, isautoregulated by NFATs and potentiates effector func-shida et al., 1998) and that calcineurin generally regu-
lates Th2 differentiation (Yamashita et al., 2000), ICOS tion but not apoptosis of T cells. NFATc1/B�C isoformsare not regulated by this means. The precise mechanismis likely to mediate early IL-4 expression through an
NFATc-dependent mechanism. We should point out by which ICOS regulates NFATc1 expression apparentlywill require further, more detailed investigation. The factthat, although NFATc1 nuclear localization was found
reduced in ICOS�/� T cells activated at the presence that CsA but not STAT6 defiency inhibited NFATc1 in-duction indicates a role of ICOS in synergy with TcRof APC during early T cell differentiation (Figure 5), no
significant deficiency was observed in ICOS�/� effector and CD28, independent of IL-4 action.How can we incorporate this study into the currentT cells treated with only anti-CD3 (Figure 1). Therefore,
NFATc1 nuclear localization appears to be differentially scheme of Th2 differentiation regulation? We summarizeour model in Figure 7. A two-step Th2 differentiationregulated in recently activated and effector T cells. To
support this notion, we did find that ICOS costimulation process is receiving more and more support in the litera-ture. TcR and maybe CD28 signaling activates naiveonly has an enhancement effect on NFATc1 expression
at 24 hr after T cell activation, but not at later days Th cells and induces both IL-4 and IFN� production(Kamogawa et al., 1993). This is accompanied by chro-(Figure 6D). NFATc1 is a key regulator of initial IL-4
production downstream of TcR and costimulatory re- matin remodeling of both loci, likely driven by NFATc1and NFATc2 factors (Avni et al., 2002; Fields et al., 2002).ceptors. TcR potency regulates NFATc1 binding to the
target sequences relative to that of NFATc2 (Brogdon et Thus, the newly activated T cells are highly proliferatingand transcribing both IFN� and IL-4 at low levels. Naiveal., 2002). CD28 enhances NFATc1 nuclear localization
(Figure 5A; Dong et al., 1998), potentially through sus- T cell activation and cytokine expression is driven byantigen but not by polarizing cytokines. ICOS acts ontained calcium flux and calcineurin activity. ICOS, on
the other hand, appears to act differently by strongly this stage by enhancing NFATc1 expression, leading togreater amounts of IL-4 expression. Several pieces ofenhancing total NFATc1 expression.

ICOS Regulation of IL-4 Transcription809
Retroviral Infectiondata shown in this paper support a role for ICOS inAn �2.3 kb EcoRI restriction fragment, containing the coding regionenhancing TcR- and CD28-mediated signaling events.of murine c-Maf (�1 to �2321), was cloned into the GFP-RV vectorFirst, ICOS-deficient Th cells exhibited reduced IL-4 ex-(a gift of Ken Murphy at Washington University), which has been
pression 24 hr after activation (Figure 4A); this initial IL-4 partially digested with EcoRI, to generate the c-Maf-RV. The Phenix-expression is not dependent on IL-4 signaling (Figure Eco ecotropic packaging cell line was transfected with retroviral
vectors GFP-RV or c-Maf-RV, and the resulting retrovirus was4B). Second, ICOS enhancement of NFATc1 expressionharvested following procedures published by Nolan et al. (http://was dependent on calcineurin (Figure 6B) but not STAT6www.stanford.edu/group/nolan/protocols/pro_helper_free.html).activity (data not shown). The second step of Th differen-Naive wild-type or ICOS�/� CD4� T cells (1 � 106/ml) were stimulatedtiation is from activated T cells to Th1 or Th2. This iswith plate-bound anti-CD3, irradiated wild-type APC (1 � 106/ml) and
mediated by lineage-specific factors such as T-bet and IL-2 (30 units/ml) for 24–36 hr and spin-infected by centrifugation atGATA3, which are regulated by cytokine products of 2700 rpm for 1.5 hr at room temperature with viral supernatant
containing 6 �g/ml Lipofectamin 2000 (Invitrogen). The viral super-newly activated T cells—IFN� and IL-4, respectively.natant was removed 24 hr after the spin infection and replaced withT-bet and GATA3 stabilize the chromatin structure ofmedium containing IL-2 (30 units/ml). GFP-positive cells were FACSthe cytokine gene loci and allow the accessibility ofsorted and restimulated with anti-CD3 for 24 hr, and IL-4 productiongene-specific transcription factors, such as c-Maf, towas assayed by ELISA.
regulate the expression of each cytokine gene at theeffector phase. In this paper, we described that IL-4 Real-Time PCR Analysis
cDNA (1 �l each) samples, prepared from total RNA extracted usingproduction mediated by c-Maf at the effector phase isTRIzol (Invitrogen) and reverse-transcribed using oligo(dT) (In-predetermined by the amount of initial IL-4 during Thvitrogen), were used in a 25 �l reaction and analyzed by Taqmandifferentiation. ICOS, by enhancing IL-4 expression atPCR with the Brilliant QPCR kit (Stratagene) for target and referencethe early activation stage, regulates c-Maf expressiongene (�-actin) expression. Cycling conditions were 2 min at 50�C
at the effector phase and hence effector IL-4 production. and 10 min at 95�C followed by 40 repeats of 15 s at 95�C and 1Th cell function is determined by its interaction with min at 60�C. Sequence-specific fluorescent signal was detected by
an ABI Prism 7700 Sequence Detector, and CT values above athe microenvironment. Costimulation provides a meansset threshold were defined using the sequence detection softwareby which T cells sense the environmental needs. Wesupplied with machine. mRNA data was normalized relative toand others reported that ICOS is involved in T-B cell�-actin and then used to calculate expression levels according to thecollaboration (Dong et al., 2001a, 2001b; McAdam et al.,Ct method (Pfaffl, 2001). Subsequently, all data was expressed
2001; Tafuri et al., 2001). The fact that B cells constitu- relative to the sample with the lowest expression which was set astively express B7h makes it possible for these cells to one. IL-4 primer (IL-4 forward, 5-AGATCATCGGCATTTTGAACG-3;
IL-4 reverse, 5-TTTGGCACATCCATCTCCG-3) and probe (5-6-instruct T cells through ICOS to regulate IL-4 expressionFAM TCACAGGAGAAGGGACGCCATGC TAMRA-3) sequencesin order to get enough “help” for their proliferation andwere according to a previous report (Grogan et al., 2001). Otherimmunoglobulin gene somatic mutation. The Th2 regula-primer and probe sequences are as follows: �-actin (�-actin forward,tory mechanisms elucidated in this paper are therefore5-TCCTTCGTTGCCGGTCCAC-3; �-actin reverse, 5-ACCAGC
functionally significant in humoral immunity. GCAGCGATATCGTC-3; �-actin probe, 5-6-FAM CCGCCAGTTCGCCATG TAMRA-3), IFN-� (IFN-� forward, 5-GATGCATTCAT
Experimental Procedures GAGTATTGCCAAGT-3; IFN-� reverse, 5-GTGGACCACTCGGATGAGCTC-3; IFN-� probe, 5-6-FAM TGAGGTCAACAACCCACA
Mice GGTCCAGCG TAMRA-3).ICOS knockout mice were previously analyzed on B6x129 F1 back-ground and subsequently backcrossed six generations onto Nuclear Extract Preparation and Immunoblot AnalysisC57BL/6. ICOS mice on the B6 F6 background were used in most Nuclear fraction of Th cells was prepared as described (Schreiberof the experiments, in which B6 mice were used as controls. In et al., 1989). The amounts of protein were determined by Bio-Radsome experiments, wild-type and knockout B6x129 F1 mice were protein assay to ensure equal protein loading for Western blot analy-used and later confirmed with mice on the B6 background. IL-4 sis with antibodies to NFATc1, NFATc2, GATA3, STAT6, Jun B,gene-deficient mice on B6 background were generous gifts of Dr. �-actin (Santa Cruz Biotechnology), JNK (PharMingen), T-bet, andMark Bix at the University of Washington. c-Maf (generous gifts of L. Glimcher).
T Cell Isolation and Stimulation AcknowledgmentsCD4� T cells from lymph nodes and spleens of 6- to 8-week-oldmice were isolated and stimulated as we previously reported (Dong The authors dedicate this work to the memory of Dr. Charles Jane-et al., 2000, 2001a). In brief, the CD4� T cells were first enriched by way. We thank Dr. Laurie Glimcher for her generous help, support,immunomagnetic negative selection using antibodies against CD8, and provision of key reagents, and Drs. Karen Makar and Hong YuMHC class II, and NK1.1 cells and magnetic beads conjugated with for their advice/help on the real-time PCR and retroviral transductiongoat anti-mouse and anti-rat Ig (Polysciences, Inc). The CD62L� experiments, respectively. We are also grateful to Drs. Heon Parknaive cells were further purified by an AutoMACS sorter (Miltenyi and Thang Nguyen, Eileen Yoshida, and Caroline Bishop for theirBiotec, Auburn, CA). In the experiment shown in Figures 1, 4B, and critical reading of the manuscript and to the entire Dong lab for their6D, CD44� cells were also depleted from this naive T cell population. help and discussion. This work is supported in part by a start-Splenic APC were prepared by complement-mediated lysis of Thy1� up fund from the Department of Immunology at the University ofT cells. For Th differentiation, naive T cells (1 � 106/ml) were treated Washington and by grants from the National Institutes of Health (towith plate-bound anti-CD3, irradiated wild-type APC (1�106/ml), and C.D. and R.A.F.). R.A.F. is an investigator of the Howard HughesIL-2 (30 units/ml, a kind gift of Dr. Linda Burkly, Biogen) for 4 days. Medical Institute and C.D. is a recipient of an Arthritis InvestigatorFor primary T cell activation, purified naive CD4� T cells were stimu- award of the Arthritis Foundation and a Basil O’Connor Starterlated in triplicate with plate-bound anti-CD3 in the presence of anti- Scholar award from March of Dimes Foundation.CD28 or B7.1-Ig (R&D) and/or anti-ICOS. Proliferation was assayedafter 3 days of treatment by adding 3H-thymidine to the culture for Received: August 15, 2002the last 8 hr. At day 4, the supernatant of stimulated cells was Revised: March 31, 2003removed, and Th cytokine production was measured by ELISA Accepted: April 17, 2003
Published: June 17, 2003(PharMingen).

Immunity810
References proto-oncogene c-maf is responsible for tissue-specific expressionof interleukin-4. Cell 85, 973–983.
Alegre, M.L., Frauwirth, K.A., and Thompson, C.B. (2001). T-cell Hutloff, A., Dittrich, A.M., Beier, K.C., Eljaschewitsch, B., Kraft, R.,regulation by CD28 and CTLA-4. Nat. Rev. Immunol. 1, 220–228. Anagnostopoulos, I., and Krocsek, R.A. (1999). ICOS is an inducible
T-cell co-stimulator structurally and functionally related to CD28.Avni, O., Lee, D., Macian, F., Szabo, S.J., Glimcher, L.H., and Rao,Nature 397, 263–266.A. (2002). T(H) cell differentiation is accompanied by dynamic
changes in histone acetylation of cytokine genes. Nat. Immunol. 3, Kamogawa, Y., Minasi, L.A., Carding, S.R., Bottomly, K., and Flavell,643–651. R.A. (1993). The relationship of IL-4- and IFN �-producing T cells
studied by lineage ablation of IL-4-producing cells. Cell 75, 985–995.Brogdon, J.L., Leitenberg, D., and Bottomly, K. (2002). The potencyof TCR signaling differentially regulates NFATc/p activity and early Kaplan, M.H., Schindler, U., Smiley, S.T., and Grusby, N.J. (1996).IL-4 transcription in naive CD4� T cells. J. Immunol. 168, 3825–3832. Stat6 is required for mediating responses to IL-4 and for develop-
ment of Th2 cells. Immunity 4, 313–319.Buonfiglio, D., Bragardo, M., Redoglia, V., Vaschetto, R., Bottarel,F., Bonissoni, S., Bensi, T., Mezzatesta, C., Janeway, C.A., Jr., and Kiani, A., Rao, A., and Aramburu, J. (2000). Manipulating immuneDianzani, U. (2000). The T cell activation molecule H4 and the CD28- responses with immunosuppressive agents that target NFAT. Immu-like molecule ICOS are identical. Eur. J. Immunol. 30, 3463–3467. nity 12, 359–372.
Carreno, B.M., and Collins, M. (2002). The B7 family of ligands and its Kiani, A., Garcia-Cozar, F.J., Habermann, I., Laforsch, S., Aebischer,receptors: new pathways for costimulation and inhibition of immune T., Ehninger, G., and Rao, A. (2001). Regulation of interferon-gammaresponses. Annu. Rev. Immunol. 20, 29–53. gene expression by nuclear factor of activated T cells. Blood 98,
1480–1488.Chambers, C.A., and Allison, J.P. (1999). Costimulatory regulationof T cell function. Curr. Opin. Cell Biol. 11, 203–210. Kim, J.I., Ho, I.C., Grusby, M.J., and Glimcher, L.H. (1999). The tran-
scription factor c-Maf controls the production of interleukin-4 butChow, C.-W., Dong, C., Flavell, R., and Davis, R. (2000). JNK inhibitsnot other Th2 cytokines. Immunity 10, 745–751.targeting of protein phosphatase calcineurin to NFATc1. Mol. Cell.Kurata, H., Lee, H.J., O’Garra, A., and Arai, N. (1999). Ectopic expres-Biol. 20, 5227–5234.sion of activated Stat6 induces the expression of Th2-specific cyto-Chuvpilo, S., Jankevics, E., Tyrsin, D., Akimzhanov, A., Moroz, D.,kines and transcription factors in developing Th1 cells. ImmunityJha, M.K., Schulze-Luehrmann, J., Santner-Nanan, B., Feoktistova,11, 677–688.E., Konig, T., et al. (2002). Autoregulation of NFATc1/A expressionLee, G.R., Fields, P.E., and Flavell, R.A. (2001). Regulation of IL-4facilitates effector T cells to escape from rapid apoptosis. Immunitygene expression by distal regulatory elements and GATA-3 at the16, 881–895.chromatin level. Immunity 14, 447–459.Constant, S.L., and Bottomly, K. (1997). Induction of Th1 and Th2Lee, H.J., Takemoto, N., Kurata, H., Kamogawa, Y., Miyatake, S.,CD4� T cell responses: the alternative approaches. Annu. Rev.O’Garra, A., and Arai, N. (2000). GATA-3 induces T helper cell typeImmunol. 15, 297–322.2 (Th2) cytokine expression and chromatin remodeling in committedCoyle, A.J., Lehar, S., Lloyd, C., Tian, J., Delaney, T., Manning, S.,Th1 cells. J. Exp. Med. 192, 105–115.Nguyen, T., Burwell, T., Schneider, H., Gonzalo, J.A., et al. (2000).Lenschow, D.J., Walunas, T.L., and Bluestone, J.A. (1996). CD28/The CD28-related molecule ICOS is required for effective T cell-B7 system of T cell costimulation. Annu. Rev. Immunol. 14, 233–258.dependent immune responses. Immunity 13, 95–105.Li, B., Tournier, C., Davis, R.J., and Flavell, R.A. (1999). RegulationCrabtree, G.R., and Olson, E.N. (2002). NFAT signaling: choreo-of T helper cell differentiation by the transcription factor JunB. EMBOgraphing the social lives of cells. Cell Suppl. 109, S67–S79.J. 19, 420–432.Diehl, S., Chow, C.W., Weiss, L., Palmetshofer, A., Twardzik, T.,McAdam, A.J., Chang, T.T., Lumelsky, A.E., Greenfield, E.A., Boussi-Rounds, L., Serfling, E., Davis, R.J., Anguita, J., and Rincon, M.otis, V.A., Duke-Cohan, J.S., Chernova, T., Malenkovich, N., Jabs, C.,(2002). Induction of NFATc2 expression by interleukin 6 promotesKuchroo, V.K., et al. (2000). Mouse inducible costimulatory moleculeT helper type 2 differentiation. J. Exp. Med. 196, 39–49.(ICOS) expression is enhanced by CD28 costimulation and regulatesDong, C., and Flavell, R.A. (2000a). Cell fate decision: T-helper 1differentiation of CD4(�) T cells. J. Immunol. 165, 5035–5040.and 2 subsets in immune responses. Arthritis Res. 2, 179–188.McAdam, A.J., Greenwald, R.J., Levin, M.A., Chernova, T., Malen-
Dong, C., and Flavell, R.A. (2000b). Control of T helper cell differenti-kovich, N., Ling, V., Freeman, G.J., and Sharpe, A.H. (2001). ICOS
ation—in search of master genes. Sci STKE 2000, E1.is critical for CD40-mediated antibody class switching. Nature 409,
Dong, C., Yang, D., Tournier, C., Whitmarsh, A., Xu, J., Davis, R., 102–105.and Flavell, R. (2000). JNK is required for effector T-cell function
Monticelli, S., and Rao, A. (2002). NFAT1 and NFAT2 are positivebut not for T-cell activation. Nature 405, 91–94.
regulators of IL-4 gene transcription. Eur. J. Immunol. 32, 2971–2978.Dong, C., Juedes, A.E., Temann, U.-A., Shresta, S., Allison, J.P., Ouyang, W., Ranganath, S.H., Weindel, K., Bhattacharya, D., Mur-Ruddle, N.H., and Flavell, R.A. (2001a). ICOS co-stimulatory receptor phy, T.L., Sha, W.C., and Murphy, K.M. (1998). Inhibition of Th1is essential for T-cell activation and function. Nature 409, 97–102. development mediated by GATA-3 through an IL-4-independentDong, C., Temann, U.A., and Flavell, R.A. (2001b). Cutting edge: mechanism. Immunity 9, 745–755.critical role of inducible costimulator in germinal center reactions. Ouyang, W., Lohning, M., Gao, Z., Assenmacher, M., Ranganath, S.,J. Immunol. 166, 3659–3662. Radbruch, A., and Murphy, K.M. (2000). Stat6-independent GATA-3Dong, C., Yang, D.D., Wysk, M., Whitmarsh, A.J., Davis, R.J., and autoactivation directs IL-4-independent Th2 development and com-Flavell, R.A. (1998). Defective T cell differentiation in the absence mitment. Immunity 12, 27–37.of Jnk1. Science 282, 2092–2095. Paul, W.E., and Seder, R.A. (1994). Lymphocyte response and cyto-Fields, P.E., Kim, S.T., and Flavell, R.A. (2002). Cutting edge: kines. Cell 76, 241–251.changes in histone acetylation at the IL-4 and IFN-gamma loci ac- Pfaffl, M.W. (2001). A new mathematical model for relative quantifi-company Th1/Th2 differentiation. J. Immunol. 169, 647–650. cation in real-time RT-PCR. Nucleic Acids Res. 29, e45.Glimcher, L.H., and Murphy, K.M. (2000). Lineage commitment in Ranger, A.M., Hodge, M.R., Gravallese, E.M., Oukka, M., Davidson,the immune system: the T helper lymphocyte grows up. Genes Dev. L., Alt, F.W., de la Brousse, F.C., Hoey, T., Grusby, M., and Glimcher,14, 1693–1711. L.H. (1998). Delayed lymphoid repopulation with defects in IL-4-Grogan, J.L., Mohrs, M., Harmon, B., Lacy, D.A., Sedat, J.W., and driven responses produced by inactivation of NF-ATc. Immunity 8,Locksley, R.M. (2001). Early transcription and silencing of cytokine 125–134.genes underlie polarization of T helper cell subsets. Immunity 14, Rao, A., Luo, C., and Hogan, P.G. (1997). Transcription factors of205–215. the NFAT family: regulation and function. Annu. Rev. Immunol. 15,
707–747.Ho, I.C., Hodge, M.R., Rooney, J.W., and Glimcher, L.H. (1996). The

ICOS Regulation of IL-4 Transcription811
Redoglia, V., Dianzani, U., Rojo, J.M., Portoles, P., Bragardo, M.,Wolff, H., Buonfiglio, D., Bonissoni, S., and Janeway, C.A., Jr. (1996).Characterization of H4: a mouse T lymphocyte activation moleculefunctionally associated with the CD3/T cell receptor. Eur. J. Immu-nol. 26, 2781–2789.
Schreiber, E., Matthias, P., Muller, M.M., and Schaffner, W. (1989).Rapid detection of octamer binding proteins with ‘mini-extracts’,prepared from a small number of cells. Nucleic Acids Res. 17, 6419.
Sharpe, A.H., and Freeman, G.J. (2002). The B7–CD28 superfamily.Nat. Rev. Immunol. 2, 116–126.
Shimoda, K., van Deursen, J., Sangster, M.Y., Srawar, S.R., Carson,R.T., Tripp, R.A., Chu, C., Quelle, F.W., Nosaka, T., Vignali, D.A., etal. (1996). Lack of IL4-induced Th2 response and IgE class switchingin mice with disrupted Stat6 gene. Nature 380, 630–633.
Swallow, M.M., Wallin, J.J., and Sha, W.C. (1999). B7h, a novelcostimulatory homolog of B7.1 and B7.2, is induced by TNFa. Immu-nity 11, 423–432.
Tafuri, A., Shahinian, A., Bladt, F., Yoshinaga, S.K., Jordana, M.,Wakeham, A., Boucher, L.-M., Bouchard, D., Chan, V.S.F., Duncan,G., et al. (2001). ICOS is essential for effective T-helper-cell re-sponses. Nature 409, 105–109.
Takeda, K., Tanaka, T., Shi, W., Matsumoto, M., Minami, M., Kashi-wamura, S.-I., Nakanishi, K., Yoshida, N., Kishimoto, T., and Akira, S.(1996). Essential role of Stat6 in IL-4 signaling. Nature 380, 627–630.
Takemoto, N., Kamogawa, Y., Jun Lee, H., Kurata, H., Arai, K.I.,O’Garra, A., Arai, N., and Miyatake, S. (2000). Cutting edge: chroma-tin remodeling at the IL-4/IL-13 intergenic regulatory region for Th2-specific cytokine gene cluster. J. Immunol. 165, 6687–6691.
Usui, T., Nishikomori, R., Kitani, A., and Strober, W. (2003). GATA-3suppresses Th1 development by downregulation of Stat4 and notthrough effects on IL-12Rb2 chain or T-bet. Immunity 18, 415–428.
Yamashita, M., Katsumata, M., Iwashima, M., Kimura, M., Shimizu,C., Kamata, T., Shin, T., Seki, N., Suzuki, S., Taniguchi, M., andNakayama, T. (2000). T cell receptor-induced calcineurin activationregulates T helper type 2 cell development by modifying the interleu-kin 4 receptor signaling complex. J. Exp. Med. 191, 1869–1879.
Yoshida, H., Nishina, H., Takimoto, H., Marengere, L.E., Wakeham,A.C., Bouchard, D., Kong, Y.Y., Ohteki, T., Shahinian, A., Bachmann,M., et al. (1998). The transcription factor NF-ATc1 regulates lympho-cyte proliferation and Th2 cytokine production. Immunity 8, 115–124.
Yoshinaga, S.K., Whoriskey, J.S., Khare, S.D., Sarmiento, U., Guo,J., Horan, T., Shih, G., Zhang, M., Coccia, M.A., Kohno, T., et al.(1999). T-cell co-stimulation through B7RP-1 and ICOS. Nature 402,827–832.
Zhang, D.H., Cohn, L., Ray, P., Bottomly, K., and Ray, A. (1997).Transcriptional factor GATA-3 is differentially expressed in murineTh1 and Th2 cells and controls Th2-specific expression of the in-terleukin-5 gene. J. Biol. Chem. 272, 21597–21603.
Zhang, D.H., Yang, L., Cohn, L., Parkyn, L., Homer, R., Ray, P., andRay, A. (1999). Inhibition of allergic inflammation in a murine modelof asthma by expression of a dominant-negative mutant of GATA-3.Immunity 11, 473–482.
Zheng, W., and Flavell, R.A. (1997). The transcription factor GATA-3is necessary and sufficient for Th2 cytokine gene expression in CD4T cells. Cell 89, 587–596.
Copyright © 2022 FDOKUMEN




















