
Bilirubin From Heme Oxygenase1 Attenuates Vascular Endothelial Activation and Dysfunction
Upload
independentCategory
view
3download
0
Journal of Neuroimmunology 225 (2010) 13–21
Contents lists available at ScienceDirect
Journal of Neuroimmunology
j ourna l homepage: www.e lsev ie r.com/ locate / jneuro im
A murine model of stress controllability attenuates Th2-dominant airwayinflammatory responses
Aniket Deshmukh a, Byung-Jin Kim a, Xavier Gonzales a, James Caffrey b,Jamboor Vishwanatha a, Harlan P. Jones a,⁎a Department of Molecular Biology and Immunology, University of North Texas Health Science Center, Fort Worth, Texas 3500 Camp Bowie Boulevard, Fort Worth, Texas 76107, United Statesb Department of Integrative Physiology, University of North Texas Health Science Center, Fort Worth, Texas 3500 Camp Bowie Boulevard, Fort Worth, Texas 76107, United States
⁎ Corresponding author. Tel.: +817 735 2448; fax: +E-mail address: [email protected] (H.P. Jones).
0165-5728/$ – see front matter © 2010 Elsevier B.V. Aldoi:10.1016/j.jneuroim.2010.03.010
a b s t r a c t
a r t i c l e i n f oArticle history:Received 22 September 2009Received in revised form 8 March 2010Accepted 15 March 2010
Keywords:StressControllabilityAsthmaImmunity
Epidemiological and experimental studies suggest a positive correlation between chronic respiratoryinflammatory disease and the ability to cope with adverse stress. Interactions between neuroendocrine andimmune systems are believed to provide insight toward the biological mechanisms of action. The utility of anexperimental murine model was employed to investigate the immunological consequences of stress-controllability and ovalbumin-induced airway inflammation. Pre-conditioned uncontrollable stress exacerbatedOVA-induced lunghistopathological changes thatwere typical of Th2-predominant inflammatory response alongrespiratory tissues. Importantly, mice given the ability to exert control over aversive stress attenuatedinflammatory responses and reduced lung pathology. This model represents a means of investigating the neuro-immune axis in defining mechanisms of stress and respiratory disease.
817 735 2133.
l rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The relationship betweenperceived anduncontrollable stress eventsand propensity of disease has been an accepted idea for decades, butuncovering the mechanistic links has only recently become a focus ofepidemiological and experimental research. Meta-analysis studies onthe role of psychosocial factors and disease have begun to quantifymeasures that support such associations (Steptoe et al., 2008). Inparticular, a majority of data has been collected on populations withatopic disorders (e.g. allergic rhitnitis, dermatitis and asthma), whichsuggest that psychosocial interventions in conjunction with typicalpharmacologic treatments can facilitate useful treatments in combat ofatopic diseases (Chida and Hamer, 2008). Furthermore, researchershave demonstrated apositive correlationbetweenmental disorders andthe incidence of disease (Hartmann et al, 2010; Li et al., 2010; Giardinoet al., 2010).
Stress disorders continue to escalate each year and are believed tocontribute to the increase in mal-adaptive behavioral and physiolog-ical responses, resulting in pathological conditions along the respira-tory airways including asthma, chronic obstructive pulmonary diseaseand respiratory infection (Sandberg et al., 2000; Chen and Miller,2007; Wright, 2005). In a study by Jones et al., a positive correlationwas found between posttraumatic stress disorder (PTSD) and COPD(Jones et al., 2009). A recent study also documented an increase risk
for contracting SARS among healthcares was related to chronic stressamong healthcare workers (McAlonan et al., 2007). Furthermore,there is a growing appreciation for the association between stress andasthma as reported by in the National Survey of Children Health(NSCH), which documented psychosocial stressors as important riskfactors (Subranmanian and Kennedy, 2009). On the basis of emergingepidemiological evidence in support of stress and health status,further experimental data is needed to reveal the underlyingmechanisms relating stress and disease pathogenesis.
Immune status is believed to be amajor physiological focal point inexplaining the interaction between stress and disease. In this regard, ithas previously been shown that stress-induced alterations in neuro-peptide activity have a profound impact on immune-mediatedinflammatory processes. Mediators of immune function play a keyrole in both the protective and pathogenic responses along therespiratory airways. Notably, altered CD4+ T cell responses including,T helper cell-type 2 (Th2) mediated by the increased secretion of IL-4,IL-5, and IL-13 (Larche, et al., 2003; Wills-Karp, 1999), IFN-γproduction by Th1 cells, IL-10 and/or TGF-β by CD4+ T regulatory(Tregs) cells and IL-17 by Th17 subsets can all participate in both theprogress and limitation of respiratory disease (Wakashin et al. 2008;Maddox and Schwartz, 2002). Overactive Th2 cellular activation isknown to exacerbate allergic airway disease (Marshall and Agarwal,2000), and has been implicated in stress-induced immunopathoglo-gical responses. Likewise, studies have demonstrated an associationbetween stress-induced impaired Th1 cellular responses and in-creased susceptibility to viral and bacterial pneumonias (Gonzales etal., 2008; Mays et al., 2010). Also, recent experimental studies have

Fig. 1. Stress paradigm and allergen sensitization and challenge. Prior to stress exposure,mice were sensitized with OVA and aluminum hydroxide adjuvant on day 0 and day 7.Groups of mice were exposed to escapable or inescapable stress on days 8, 10, 12, 13, 14and 15. Escapable, inescapable and non-stressed groups (Home cage and Sham) of micewere exposed to either OVA or PBS by intranasal route on days 13, 14 and 15. On day 17,mice were euthanized.
14 A. Deshmukh et al. / Journal of Neuroimmunology 225 (2010) 13–21
documented the significant role of Th17 as well as T regulatory cellactivation in modulation of chronic inflammatory conditions (Waka-shin et al., 2008; Wright, 2007; Joachim et al., 2006; Larche et al.,2003), and their role in neuro-immune regulation of respiratorydisease. Therefore, the immunoregulatorymechanisms responsible forthe pathogenesis of respiratory disease are believed to involvecomplex interactions among multiple cellular immune populations.
The interaction between stress and disease has led to the emergingfield of psychoneuroimmunology research (PNIR), aimed towardunderstanding the relationship between central nervous and immunesystem functioning. In this regard, animal models of stress are useful tostudy neuro-immune interactions and disease. Previous studies haveestablished unpredictable stressors as valid instruments for producingfear, depression and anxiety all indicative ofmal-adaptive physiologicaland behavioral responses (Anisman andMatheson, 2005) (Mikics et al.,2008). (Chourbaji et al., 2005b; Chourbaji et al., 2005a). (Rybnikovaet al., 2007). While these studies in mice have focused on behavioralpsychopathologies, few have documented alterations in diseasesusceptibility. For example, exposure to aversive shock increasedtumor progression (Yee et al., 2008; Vegas et al., 2009) and viralinfection (Ashcraft et al., 2008). Importantly, experimentalmodels havedemonstrated stress-associated influencesonCD4+Tcell responses andrespiratory disease (Marshall and Agarwal, 2000, Marshall, 2004a,b).Utilizing a murine model of stress-controllability and allergen-inducedpulmonary inflammation, we implement for the first time, a model ofallergic airway inflammation a model of learned helpless behavior inmice to demonstrate that the extent of perceived stress and the ability toexert control translates into a concomitant diminished Th2-dominantinflammatory response.
2. Methods
2.1. Mice
The Institutional Animal Care and Use Committee approved allprotocols in compliance with the NIH guide for the Care and Use ofLaboratory Animals (National Institutes of Health Publications No. 85-23,revised 1996). Adult (6–8 weeks of age) female Balbc/J mice (JacksonLaboratories, PA) were used in all studies. Mice were fed ad libitum andmaintained in a 12 h light–dark cycle under specific pathogen-freeconditions. Mice were housed 4 per cage under conditions of optimaltemperature and humidity. All animals were allowed to acclimate to thesurrounding environment two weeks prior to experimentation.
2.2. Tail shock apparatus and stress paradigm
Following acclimation to the colony environment, mice wererandomly assigned to the following experimental conditions: escap-able stress, inescapable stress and home cage environment. Mice inthe escapable conditionwere able to extinguish tail shock by turning awheel placed in reach of its front legs. When the mouse terminatedthe shock it also terminated the tail shock for a parallel mouse in theinescapable condition, allowing both subjects to receive shocks for thesame duration. Selectedmice were also placed in the shock apparatus,but were not exposed to shock (sham). An additional group of miceremained in the home cage environment and were handled similarlyto escapable, inescapable, and sham shock conditions, but were neverexposed to the shock apparatus.
Mice were exposed to escapable and inescapable conditions ondays 8, 10 and 12 of the experimental protocol (Fig. 1). On experimentalstress days, mice were individually transported to shock apparatus(Med Associates, St. Albans, VT) housed in a separate location. Micewere placed in their respective plexiglass boxes divided by an opaquesound-proof wall to inhibit visual and audible responses. The electricalprobe that delivered the electrical shockwas secured to the tail, ¾ fromthe end, including the sham-treated mice. Aversive shocks were
administered for exactly 15 min on the day of the stress paradigmbetween thehours of 2:00 PMto3:00 PM.MED-PC®Version IV software(Med associates, INC.) controlled the administration and timing of a0.2 mA current over a random 30 s intervals lasting 15 min. Thecomputer software recorded duration of shock and wheel turns todetermine the number of wheel and latency frequency between shockand wheel turn. Each chamber was thoroughly cleansed between trials.
2.3. Allergen sensitization and challenge
On day 1 and 7, mice were administered 25 mg ovalbumim i.p.(Sigma-Aldrich, St. Louis, Missouri) with aluminum hydroxide gel(13 mg/ml) adjuvant (Sigma-Aldrich) diluted in the PBS (MediatechInc., Herndon, VA). On day 13, 14, 15 mice were anesthetized withisoflurane and challenged intranasally with 20 μl OVA (10 μg/ml) inPBS or PBS alone. On day 17 mice were sacrificed after anaesthetizingwith xylazine+ketamine.
2.4. Cell isolation
Bronchiolalveolar lavage fluid (BALF) containing cells was aspi-rated by perfusion of 2 ml of wash media using a blunt 25 guageneedle. The volume of the fluid retrieved was recorded and stored forfurther analysis.
Single cell suspensions from lung tissue were prepared aspreviously described (Jones et al., 2002). Briefly lungs were infusedwith sterile PBS to eliminate contaminating RBCs. Lung tissue wasseparated into single lobes, pooled, minced and digested for 1 h indigestion medium containing 300 units/ml collagenase type II(Worthington, Lakewood, NJ.) and 0.01% DNAse (Sigma-Aldrich, St.Louis, MO). After digestion lung supernatants were passed through anylonmesh filter (LabPak, Depew, NY.) into sterile 15 ml conical tubesand washed 2× with RPMI (Hyclone, Logan, UT.) wash mediasupplemented with 1% antibiotic cocktail. Lymphocytes were pre-pared by ficoll-hypaque (Lympholyte M, Cedarlane Laboratories ltd.,Ontario, CA.) centrifugation. Contaminating RBCswere removed usingammonium chloride potassium (ACK) lysis buffer. The total viable cellcount was determined with a haemocytometer (Hausser Scientific,Horsham, PA).
2.5. Histopathology and immunohistochemistry
Dissected lungs were perfused by instillation of 2 ml of 2%paraformaldehyde (Sigma Chemical, Co., St. Louis, MO) intratracheallyfollowed byfixation in 2% paraformaldehyde at 4 °C for 4 h and overnightincubation in 30% solution of sucrose at 4 °C. On the next day eachindividual lobes of the lung were frozen using OCT (Optimum CuttingTemperature) embedding medium (Sakkura Finetechnical, Torance, CA).

15A. Deshmukh et al. / Journal of Neuroimmunology 225 (2010) 13–21
Thin tissue sections (10 µm) were prepared on SuperFrost/plus micro-scope slides (Fisher Scientific) using Ultrapro 5000 cryostat (vibratome,St. Louis, MO). The tissue sections were stained with Hematoxylin andEosin (H&E) (FisherBrand). Additional tissue sections were stained usingPeriodic Acid Schiff's Base Staining (PAS) for further confirmation.Analysis was performed using Image Pro software (Olympus, Centerval-ley, PA). The Images were captured on Olympus AX70 fluorescentmicroscope (Olympus, Centervalley, PA).
2.6. Cytokine detection in bronchiolar lavage
Enzyme-Linked Immunosorbent Assays (ELISAs) were performedusing OptEIA sets for mouse interferon-gamma (IFN-γ), interleukin-4(IL-4), interleukin-5 (IL-5), interleukin-10 (IL-10) from BD Biosciences(Franklin Lakes, NJ) and interleukin-13 (IL-13) from eBiosciences (SanDiego, CA). Briefly flat bottom 96 micro-well plates were coated with100 µl (per well) capture antibody (OptEIA anti-mouse monoclonalantibodies for different cytokines IL-4, IL-5, IL-10, IL-13, and IFN-γdiluted1:250 in coating buffer (0.1 M carbonate or 0.1 M phosphate). The platewas sealed and incubated overnight at 4 °C. The platewas thenwashed 5times with ≥200 µl/well wash buffer (PBS with 0.05% Tween-20). Afterthe last wash the plate was inverted and blotted on a paper towel toremove residual buffer. This wash procedure was used throughout therest of the protocol unless otherwise noted. After washing, the plate wasblocked with ≥200 µl/well assay diluent (PBS with 10% FBS) andincubated overnight at 4 °C. The plate was washed, and then samples(with suitable dilutions) and standards (OptEIA serially diluted recom-binant mouse IL-4, IL-5, IL-10, IL-13, IFN) were pipetted into theappropriate wells of the plate (100 µl/well of sample and standard).The plate was sealed and incubated overnight at 4 °C. After washprocedure, 100 µl/well Detection Antibody (OptEIA biotinylated anti-mouse IL-4, IL-5, IL-10, IL-13, IFN-γmonoclonal antibody) diluted 1:250in assay diluentwas added, and the platewas sealed and incubated for anhour at RT (room temperature). After washing, 100 µl/well EnzymeReagent (OptEIAavidin-horseradishperoxide conjugate)diluted1:250 inassay diluent was added, and the plate was sealed and incubated for30 minatRT. For thefinalplatewash, plateswere rinsedwithwashbuffer7 times, letting the buffer sit in the wells for≥30 s per rinse. 100 µl/wellTMB (Tetramethylbenzidine, BD Pharmingen) was added and the platewas incubated for approximately 20 min (normally) unsealed at RT indark. 50 µl/well stop solution (0.25 MHCL)wasadded tohalt the reactionand the absorbance (at 450 nm) of the plate was read on an ELISA readerusing the computer program Gen5 (Bio-Tek Instruments).
Fig. 2. Provocation of airway hypersensitivity diminishes avoidance response to aversivetailshock. The total number of wheel turns during each session of tailshock was recordedfor each subject and plotted as a function of stress days during the duration of the stressparadigm. Data represents mean (n=15)±std. error in the number of wheel turns at agiven time point. (*) and (#) designates significant p≤0.05) difference between eachexperimental group at each time point. OVA; ovalbumin-treated, PBS; Phosphate bufferedsaline-treated, ES; escapable stress and IS; inescapable stress, and Sham.
2.7. Flow cytometry
Two and three color flow cytometric staining was performedusing fluorescently-labeled antibodies conjugated to the followingflourochromes: FITC (Fluroscein isothiocyanate), R-PE (R-phycoerythrin)and cy-chrome. One million lymphocytes out of lung single cellsuspensions were washed with washing buffer (2% FBS in 1X PBS with2 mM EDTA) and incubated with an optimal concentration of anti-FcBlock Antibody (clone 2.4G2) (BD Pharmingen, SanDiego, CA) to blocknon-specific binding to FcRs for 30 min. Multicoloured staining wasperformed by incubating cells at 4 °C with compatible combinationsof pre-tittered concentrations of following antibodies: anti-GR1+FITC,anti-CD8+FITC, anti-CD3+PE, anti-CD4+PE-Cy7, anti-B220+FITC, anti-MHC II+PE, anti-CD19+PE-Cy5, anti-CD11c+Alexa Flour 488, anti-F/480+PEcy7. Unstained lymphocytes served as negative controls. Antibodieswere obtained from BD Pharmingen, San Deigo, CA. Gating oflymphocytes was identified by forward-scatter/ side-scatter profile.Data acquisition was done on FC500 flow cytometer (Beckman Coulter,Miami, FL). Further analysis was done on CXP software (BeckmanCoulter).
2.8. Statistical analysis
Statistical analysis was performed using GraphPad prismVersion 4.0for Windows (GraphPad Software, San Diego, USA). For multi-experimental group analysis, datawere subjected to analysis of variance(univariant ANOVA) followed Post Hoc tests (Tukey) for groupdifferences. For analysis of two-group differences, student's t test wasemployed, followed by Post Hoc analysis. All data are expressed as themean±SEMs. The two-tailed level of significancewas set up at p≤0.05.
3. Results
3.1. Induction of airway inflammation diminishes avoidance response toaversive tailshock and attenuates weight gain
Prior to experimentation, mice exposed to the stress paradigm weretested to assure equivalent wheel turn performance. OVA sensitizedmicewere subjected to four sessions (days 8,10,12, and 14) of escapableor inescapable tail shock. Sham-shocked and home cage groups of miceserved as non-shocked controls. On days 12 and 14, the stress paradigmwas preceded by an intranasal OVA or PBS challenge. The wheel turnfrequency was recorded as a measure of learned avoidance proficiency.Fig. 2 illustrates a similar number of wheel turn responses to aversiveshock on the first day (Day 8) between all mice regardless of theirpriming status (OVA vs PBS). On the second stress day (Day 10), highernumbers (p≤0.05) of wheel turn responses were observed among OVAand PBS-primed escapable mice as compared to OVA and PBS-primedinescapable and sham-treated mice. By the third stress exposure (Day12), escapable mice increased their wheel response by more than 300%(p≤0.05) compared to their inescapable mates and sham-treated mice.Acute OVA challenge restrained this response by half among theescapable PBS counterparts (p≤0.05). Both escapable groups were farmore active than their inescapable counterparts. No significantdifference in wheel turn response was observed between OVA andPBS-challenged inescapable-treated and sham-treated mice. On day 14,PBS-treated escapable mice continued to increase their wheel turnresponses while those of the other three groups reached a plateau orbegan todecline. These results confirm theprogressive learningability ofmice to escape aversive stimuli. Interestingly, OVAprimingwasnot itselfa discriminating factor but the subsequent acute airway challenge didreduce response the wheel turn responses in both escapable andinescapable groups as compared to their PBS experimental counterparts.
Upon initiation of the stress paradigm, altered wheel turn profi-ciency given uncontrolled aversive stress impacted growth rate.Thoughmore variable, those in the escapable stress groupmaintained
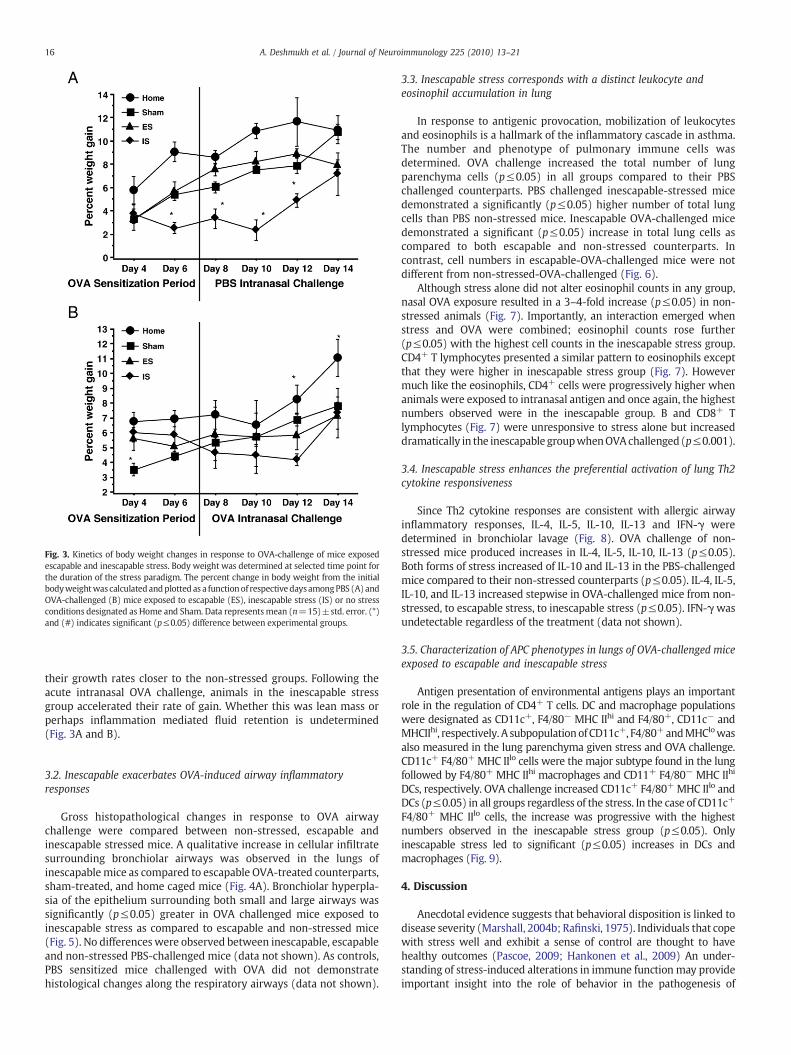
Fig. 3. Kinetics of body weight changes in response to OVA-challenge of mice exposedescapable and inescapable stress. Body weight was determined at selected time point forthe duration of the stress paradigm. The percent change in body weight from the initialbodyweightwas calculatedandplotted as a functionof respectivedaysamongPBS (A) andOVA-challenged (B) mice exposed to escapable (ES), inescapable stress (IS) or no stressconditions designated as Home and Sham. Data representsmean (n=15)±std. error. (*)and (#) indicates significant (p≤0.05) difference between experimental groups.
16 A. Deshmukh et al. / Journal of Neuroimmunology 225 (2010) 13–21
their growth rates closer to the non-stressed groups. Following theacute intranasal OVA challenge, animals in the inescapable stressgroup accelerated their rate of gain. Whether this was lean mass orperhaps inflammation mediated fluid retention is undetermined(Fig. 3A and B).
3.2. Inescapable exacerbates OVA-induced airway inflammatoryresponses
Gross histopathological changes in response to OVA airwaychallenge were compared between non-stressed, escapable andinescapable stressed mice. A qualitative increase in cellular infiltratesurrounding bronchiolar airways was observed in the lungs ofinescapablemice as compared to escapable OVA-treated counterparts,sham-treated, and home caged mice (Fig. 4A). Bronchiolar hyperpla-sia of the epithelium surrounding both small and large airways wassignificantly (p≤0.05) greater in OVA challenged mice exposed toinescapable stress as compared to escapable and non-stressed mice(Fig. 5). No differenceswere observed between inescapable, escapableand non-stressed PBS-challenged mice (data not shown). As controls,PBS sensitized mice challenged with OVA did not demonstratehistological changes along the respiratory airways (data not shown).
3.3. Inescapable stress corresponds with a distinct leukocyte andeosinophil accumulation in lung
In response to antigenic provocation, mobilization of leukocytesand eosinophils is a hallmark of the inflammatory cascade in asthma.The number and phenotype of pulmonary immune cells wasdetermined. OVA challenge increased the total number of lungparenchyma cells (p≤0.05) in all groups compared to their PBSchallenged counterparts. PBS challenged inescapable-stressed micedemonstrated a significantly (p≤0.05) higher number of total lungcells than PBS non-stressed mice. Inescapable OVA-challenged micedemonstrated a significant (p≤0.05) increase in total lung cells ascompared to both escapable and non-stressed counterparts. Incontrast, cell numbers in escapable-OVA-challenged mice were notdifferent from non-stressed-OVA-challenged (Fig. 6).
Although stress alone did not alter eosinophil counts in any group,nasal OVA exposure resulted in a 3–4-fold increase (p≤0.05) in non-stressed animals (Fig. 7). Importantly, an interaction emerged whenstress and OVA were combined; eosinophil counts rose further(p≤0.05) with the highest cell counts in the inescapable stress group.CD4+ T lymphocytes presented a similar pattern to eosinophils exceptthat they were higher in inescapable stress group (Fig. 7). Howevermuch like the eosinophils, CD4+ cells were progressively higher whenanimals were exposed to intranasal antigen and once again, the highestnumbers observed were in the inescapable group. B and CD8+ Tlymphocytes (Fig. 7) were unresponsive to stress alone but increaseddramatically in the inescapable groupwhenOVAchallenged (p≤0.001).
3.4. Inescapable stress enhances the preferential activation of lung Th2cytokine responsiveness
Since Th2 cytokine responses are consistent with allergic airwayinflammatory responses, IL-4, IL-5, IL-10, IL-13 and IFN-γ weredetermined in bronchiolar lavage (Fig. 8). OVA challenge of non-stressed mice produced increases in IL-4, IL-5, IL-10, IL-13 (p≤0.05).Both forms of stress increased of IL-10 and IL-13 in the PBS-challengedmice compared to their non-stressed counterparts (p≤0.05). IL-4, IL-5,IL-10, and IL-13 increased stepwise in OVA-challenged mice from non-stressed, to escapable stress, to inescapable stress (p≤0.05). IFN-γwasundetectable regardless of the treatment (data not shown).
3.5. Characterization of APC phenotypes in lungs of OVA-challenged miceexposed to escapable and inescapable stress
Antigen presentation of environmental antigens plays an importantrole in the regulation of CD4+ T cells. DC and macrophage populationswere designated as CD11c+, F4/80− MHC IIhi and F4/80+, CD11c− andMHCIIhi, respectively. A subpopulationof CD11c+, F4/80+andMHClowasalso measured in the lung parenchyma given stress and OVA challenge.CD11c+ F4/80+ MHC IIlo cells were the major subtype found in the lungfollowed by F4/80+ MHC IIhi macrophages and CD11+ F4/80− MHC IIhi
DCs, respectively. OVA challenge increased CD11c+ F4/80+ MHC IIlo andDCs (p≤0.05) in all groups regardless of the stress. In the case of CD11c+
F4/80+ MHC IIlo cells, the increase was progressive with the highestnumbers observed in the inescapable stress group (p≤0.05). Onlyinescapable stress led to significant (p≤0.05) increases in DCs andmacrophages (Fig. 9).
4. Discussion
Anecdotal evidence suggests that behavioral disposition is linked todisease severity (Marshall, 2004b; Rafinski, 1975). Individuals that copewith stress well and exhibit a sense of control are thought to havehealthy outcomes (Pascoe, 2009; Hankonen et al., 2009) An under-standing of stress-induced alterations in immune function may provideimportant insight into the role of behavior in the pathogenesis of
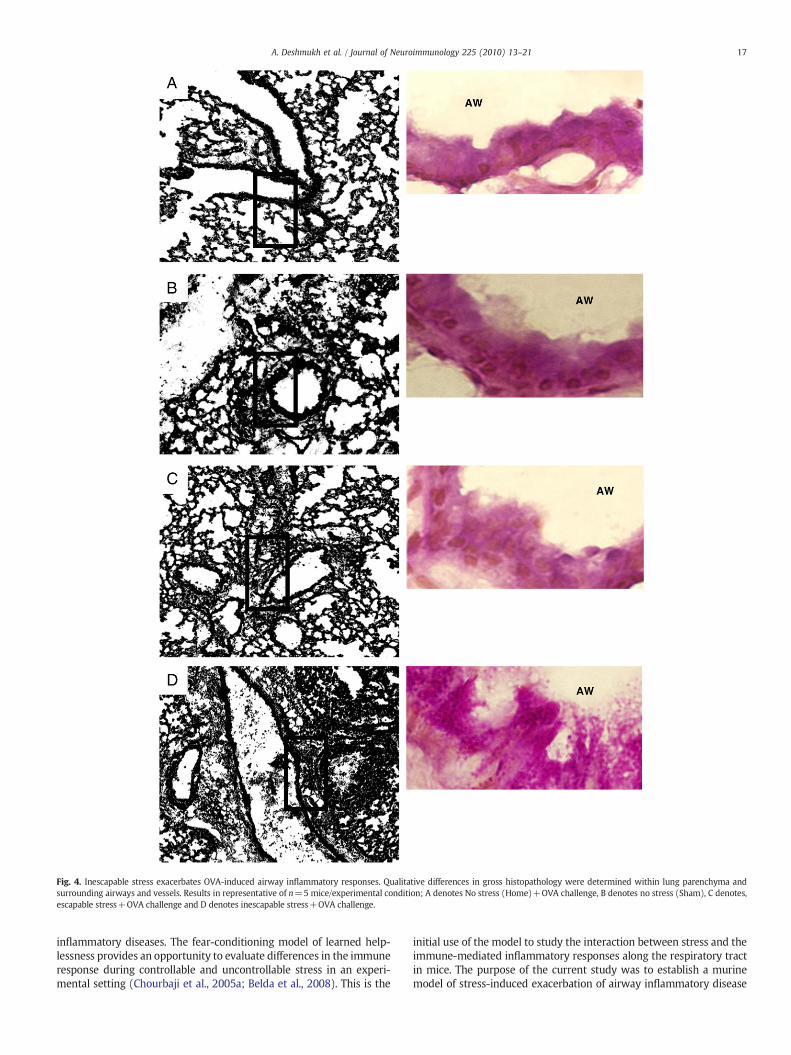
Fig. 4. Inescapable stress exacerbates OVA-induced airway inflammatory responses. Qualitative differences in gross histopathology were determined within lung parenchyma andsurrounding airways and vessels. Results in representative of n=5 mice/experimental condition; A denotes No stress (Home)+OVA challenge, B denotes no stress (Sham), C denotes,escapable stress+OVA challenge and D denotes inescapable stress+OVA challenge.
17A. Deshmukh et al. / Journal of Neuroimmunology 225 (2010) 13–21
inflammatory diseases. The fear-conditioning model of learned help-lessness provides an opportunity to evaluate differences in the immuneresponse during controllable and uncontrollable stress in an experi-mental setting (Chourbaji et al., 2005a; Belda et al., 2008). This is the
initial use of the model to study the interaction between stress and theimmune-mediated inflammatory responses along the respiratory tractin mice. The purpose of the current study was to establish a murinemodel of stress-induced exacerbation of airway inflammatory disease
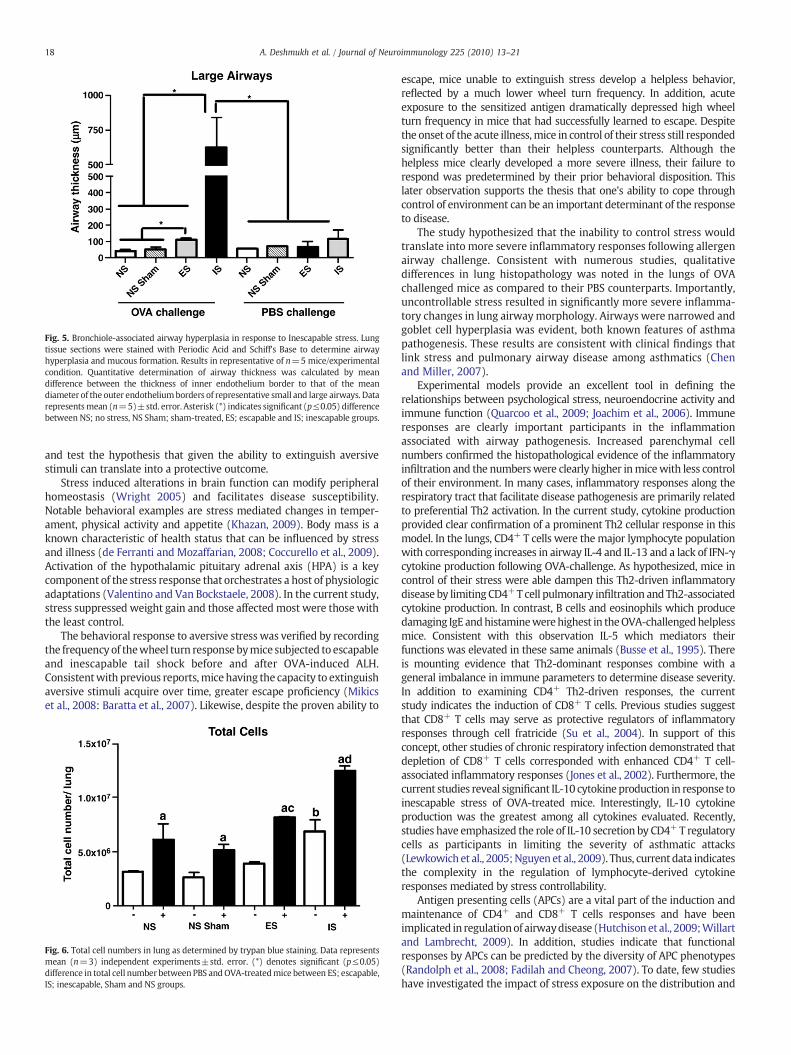
Fig. 5. Bronchiole-associated airway hyperplasia in response to Inescapable stress. Lungtissue sections were stained with Periodic Acid and Schiff's Base to determine airwayhyperplasia and mucous formation. Results in representative of n=5 mice/experimentalcondition. Quantitative determination of airway thickness was calculated by meandifference between the thickness of inner endothelium border to that of the meandiameter of the outer endotheliumborders of representative small and large airways. Datarepresentsmean (n=5)±std. error. Asterisk (*) indicates significant (p≤0.05) differencebetween NS; no stress, NS Sham; sham-treated, ES; escapable and IS; inescapable groups.
18 A. Deshmukh et al. / Journal of Neuroimmunology 225 (2010) 13–21
and test the hypothesis that given the ability to extinguish aversivestimuli can translate into a protective outcome.
Stress induced alterations in brain function can modify peripheralhomeostasis (Wright 2005) and facilitates disease susceptibility.Notable behavioral examples are stress mediated changes in temper-ament, physical activity and appetite (Khazan, 2009). Body mass is aknown characteristic of health status that can be influenced by stressand illness (de Ferranti and Mozaffarian, 2008; Coccurello et al., 2009).Activation of the hypothalamic pituitary adrenal axis (HPA) is a keycomponent of the stress response that orchestrates a host of physiologicadaptations (Valentino and Van Bockstaele, 2008). In the current study,stress suppressed weight gain and those affected most were those withthe least control.
The behavioral response to aversive stress was verified by recordingthe frequencyof thewheel turn response bymice subjected to escapableand inescapable tail shock before and after OVA-induced ALH.Consistentwith previous reports,mice having the capacity to extinguishaversive stimuli acquire over time, greater escape proficiency (Mikicset al., 2008: Baratta et al., 2007). Likewise, despite the proven ability to
Fig. 6. Total cell numbers in lung as determined by trypan blue staining. Data representsmean (n=3) independent experiments±std. error. (*) denotes significant (p≤0.05)difference in total cell number between PBS andOVA-treatedmice between ES; escapable,IS; inescapable, Sham and NS groups.
escape, mice unable to extinguish stress develop a helpless behavior,reflected by a much lower wheel turn frequency. In addition, acuteexposure to the sensitized antigen dramatically depressed high wheelturn frequency in mice that had successfully learned to escape. Despitethe onset of the acute illness,mice in control of their stress still respondedsignificantly better than their helpless counterparts. Although thehelpless mice clearly developed a more severe illness, their failure torespond was predetermined by their prior behavioral disposition. Thislater observation supports the thesis that one's ability to cope throughcontrol of environment can be an important determinant of the responseto disease.
The study hypothesized that the inability to control stress wouldtranslate into more severe inflammatory responses following allergenairway challenge. Consistent with numerous studies, qualitativedifferences in lung histopathology was noted in the lungs of OVAchallenged mice as compared to their PBS counterparts. Importantly,uncontrollable stress resulted in significantly more severe inflamma-tory changes in lung airwaymorphology. Airways were narrowed andgoblet cell hyperplasia was evident, both known features of asthmapathogenesis. These results are consistent with clinical findings thatlink stress and pulmonary airway disease among asthmatics (Chenand Miller, 2007).
Experimental models provide an excellent tool in defining therelationships between psychological stress, neuroendocrine activity andimmune function (Quarcoo et al., 2009; Joachim et al., 2006). Immuneresponses are clearly important participants in the inflammationassociated with airway pathogenesis. Increased parenchymal cellnumbers confirmed the histopathological evidence of the inflammatoryinfiltration and the numberswere clearly higher inmicewith less controlof their environment. In many cases, inflammatory responses along therespiratory tract that facilitate disease pathogenesis are primarily relatedto preferential Th2 activation. In the current study, cytokine productionprovided clear confirmation of a prominent Th2 cellular response in thismodel. In the lungs, CD4+ T cells were themajor lymphocyte populationwith corresponding increases in airway IL-4 and IL-13 and a lack of IFN-γcytokine production following OVA-challenge. As hypothesized, mice incontrol of their stress were able dampen this Th2-driven inflammatorydisease by limiting CD4+T cell pulmonary infiltration and Th2-associatedcytokine production. In contrast, B cells and eosinophils which producedamaging IgE andhistaminewerehighest in theOVA-challengedhelplessmice. Consistent with this observation IL-5 which mediators theirfunctions was elevated in these same animals (Busse et al., 1995). Thereis mounting evidence that Th2-dominant responses combine with ageneral imbalance in immune parameters to determine disease severity.In addition to examining CD4+ Th2-driven responses, the currentstudy indicates the induction of CD8+ T cells. Previous studies suggestthat CD8+ T cells may serve as protective regulators of inflammatoryresponses through cell fratricide (Su et al., 2004). In support of thisconcept, other studies of chronic respiratory infection demonstrated thatdepletion of CD8+ T cells corresponded with enhanced CD4+ T cell-associated inflammatory responses (Jones et al., 2002). Furthermore, thecurrent studies reveal significant IL-10 cytokine production in response toinescapable stress of OVA-treated mice. Interestingly, IL-10 cytokineproduction was the greatest among all cytokines evaluated. Recently,studies have emphasized the role of IL-10 secretion by CD4+ T regulatorycells as participants in limiting the severity of asthmatic attacks(Lewkowich et al., 2005;Nguyen et al., 2009). Thus, current data indicatesthe complexity in the regulation of lymphocyte-derived cytokineresponses mediated by stress controllability.
Antigen presenting cells (APCs) are a vital part of the induction andmaintenance of CD4+ and CD8+ T cells responses and have beenimplicated in regulationof airwaydisease (Hutchisonet al., 2009;Willartand Lambrecht, 2009). In addition, studies indicate that functionalresponses by APCs can be predicted by the diversity of APC phenotypes(Randolph et al., 2008; Fadilah and Cheong, 2007). To date, few studieshave investigated the impact of stress exposure on the distribution and
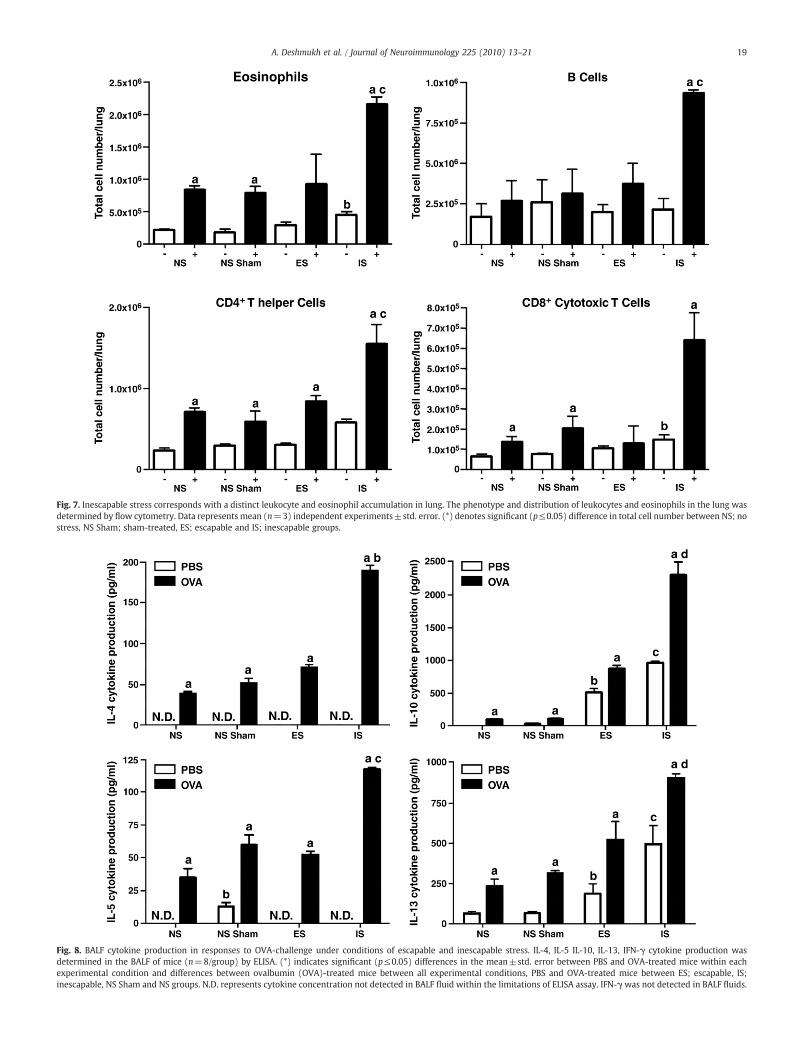
Fig. 7. Inescapable stress corresponds with a distinct leukocyte and eosinophil accumulation in lung. The phenotype and distribution of leukocytes and eosinophils in the lung wasdetermined by flow cytometry. Data represents mean (n=3) independent experiments±std. error. (*) denotes significant (p≤0.05) difference in total cell number between NS; nostress, NS Sham; sham-treated, ES; escapable and IS; inescapable groups.
Fig. 8. BALF cytokine production in responses to OVA-challenge under conditions of escapable and inescapable stress. IL-4, IL-5 IL-10, IL-13, IFN-γ cytokine production wasdetermined in the BALF of mice (n=8/group) by ELISA. (*) indicates significant (p≤0.05) differences in the mean±std. error between PBS and OVA-treated mice within eachexperimental condition and differences between ovalbumin (OVA)-treated mice between all experimental conditions, PBS and OVA-treated mice between ES; escapable, IS;inescapable, NS Sham and NS groups. N.D. represents cytokine concentration not detected in BALF fluid within the limitations of ELISA assay. IFN-γ was not detected in BALF fluids.
19A. Deshmukh et al. / Journal of Neuroimmunology 225 (2010) 13–21
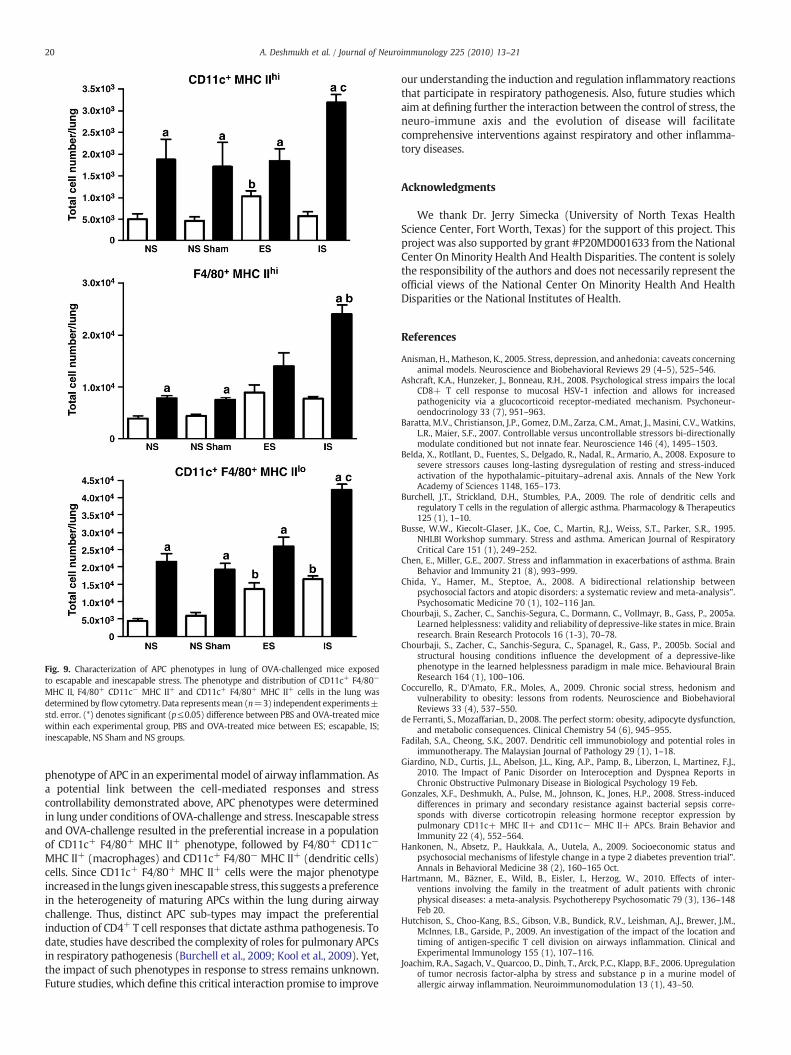
Fig. 9. Characterization of APC phenotypes in lung of OVA-challenged mice exposedto escapable and inescapable stress. The phenotype and distribution of CD11c+ F4/80−
MHC II, F4/80+ CD11c− MHC II+ and CD11c+ F4/80+ MHC II+ cells in the lung wasdetermined by flow cytometry. Data representsmean (n=3) independent experiments±std. error. (*) denotes significant (p≤0.05) difference between PBS and OVA-treated micewithin each experimental group, PBS and OVA-treated mice between ES; escapable, IS;inescapable, NS Sham and NS groups.
20 A. Deshmukh et al. / Journal of Neuroimmunology 225 (2010) 13–21
phenotype of APC in an experimental model of airway inflammation. Asa potential link between the cell-mediated responses and stresscontrollability demonstrated above, APC phenotypes were determinedin lung under conditions of OVA-challenge and stress. Inescapable stressand OVA-challenge resulted in the preferential increase in a populationof CD11c+ F4/80+ MHC II+ phenotype, followed by F4/80+ CD11c−
MHC II+ (macrophages) and CD11c+ F4/80− MHC II+ (dendritic cells)cells. Since CD11c+ F4/80+ MHC II+ cells were the major phenotypeincreased in the lungsgiven inescapable stress, this suggests apreferencein the heterogeneity of maturing APCs within the lung during airwaychallenge. Thus, distinct APC sub-types may impact the preferentialinduction of CD4+ T cell responses that dictate asthma pathogenesis. Todate, studies have described the complexity of roles for pulmonary APCsin respiratory pathogenesis (Burchell et al., 2009; Kool et al., 2009). Yet,the impact of such phenotypes in response to stress remains unknown.Future studies, which define this critical interaction promise to improve
our understanding the induction and regulation inflammatory reactionsthat participate in respiratory pathogenesis. Also, future studies whichaim at defining further the interaction between the control of stress, theneuro-immune axis and the evolution of disease will facilitatecomprehensive interventions against respiratory and other inflamma-tory diseases.
Acknowledgments
We thank Dr. Jerry Simecka (University of North Texas HealthScience Center, Fort Worth, Texas) for the support of this project. Thisproject was also supported by grant #P20MD001633 from the NationalCenter OnMinority Health And Health Disparities. The content is solelythe responsibility of the authors and does not necessarily represent theofficial views of the National Center On Minority Health And HealthDisparities or the National Institutes of Health.
References
Anisman, H., Matheson, K., 2005. Stress, depression, and anhedonia: caveats concerninganimal models. Neuroscience and Biobehavioral Reviews 29 (4–5), 525–546.
Ashcraft, K.A., Hunzeker, J., Bonneau, R.H., 2008. Psychological stress impairs the localCD8+ T cell response to mucosal HSV-1 infection and allows for increasedpathogenicity via a glucocorticoid receptor-mediated mechanism. Psychoneur-oendocrinology 33 (7), 951–963.
Baratta, M.V., Christianson, J.P., Gomez, D.M., Zarza, C.M., Amat, J., Masini, C.V., Watkins,L.R., Maier, S.F., 2007. Controllable versus uncontrollable stressors bi-directionallymodulate conditioned but not innate fear. Neuroscience 146 (4), 1495–1503.
Belda, X., Rotllant, D., Fuentes, S., Delgado, R., Nadal, R., Armario, A., 2008. Exposure tosevere stressors causes long-lasting dysregulation of resting and stress-inducedactivation of the hypothalamic–pituitary–adrenal axis. Annals of the New YorkAcademy of Sciences 1148, 165–173.
Burchell, J.T., Strickland, D.H., Stumbles, P.A., 2009. The role of dendritic cells andregulatory T cells in the regulation of allergic asthma. Pharmacology & Therapeutics125 (1), 1–10.
Busse, W.W., Kiecolt-Glaser, J.K., Coe, C., Martin, R.J., Weiss, S.T., Parker, S.R., 1995.NHLBI Workshop summary. Stress and asthma. American Journal of RespiratoryCritical Care 151 (1), 249–252.
Chen, E., Miller, G.E., 2007. Stress and inflammation in exacerbations of asthma. BrainBehavior and Immunity 21 (8), 993–999.
Chida, Y., Hamer, M., Steptoe, A., 2008. A bidirectional relationship betweenpsychosocial factors and atopic disorders: a systematic review and meta-analysis”.Psychosomatic Medicine 70 (1), 102–116 Jan.
Chourbaji, S., Zacher, C., Sanchis-Segura, C., Dormann, C., Vollmayr, B., Gass, P., 2005a.Learned helplessness: validity and reliability of depressive-like states inmice. Brainresearch. Brain Research Protocols 16 (1-3), 70–78.
Chourbaji, S., Zacher, C., Sanchis-Segura, C., Spanagel, R., Gass, P., 2005b. Social andstructural housing conditions influence the development of a depressive-likephenotype in the learned helplessness paradigm in male mice. Behavioural BrainResearch 164 (1), 100–106.
Coccurello, R., D'Amato, F.R., Moles, A., 2009. Chronic social stress, hedonism andvulnerability to obesity: lessons from rodents. Neuroscience and BiobehavioralReviews 33 (4), 537–550.
de Ferranti, S., Mozaffarian, D., 2008. The perfect storm: obesity, adipocyte dysfunction,and metabolic consequences. Clinical Chemistry 54 (6), 945–955.
Fadilah, S.A., Cheong, S.K., 2007. Dendritic cell immunobiology and potential roles inimmunotherapy. The Malaysian Journal of Pathology 29 (1), 1–18.
Giardino, N.D., Curtis, J.L., Abelson, J.L., King, A.P., Pamp, B., Liberzon, I., Martinez, F.J.,2010. The Impact of Panic Disorder on Interoception and Dyspnea Reports inChronic Obstructive Pulmonary Disease in Biological Psychology 19 Feb.
Gonzales, X.F., Deshmukh, A., Pulse, M., Johnson, K., Jones, H.P., 2008. Stress-induceddifferences in primary and secondary resistance against bacterial sepsis corre-sponds with diverse corticotropin releasing hormone receptor expression bypulmonary CD11c+ MHC II+ and CD11c− MHC II+ APCs. Brain Behavior andImmunity 22 (4), 552–564.
Hankonen, N., Absetz, P., Haukkala, A., Uutela, A., 2009. Socioeconomic status andpsychosocial mechanisms of lifestyle change in a type 2 diabetes prevention trial".Annals in Behavioral Medicine 38 (2), 160–165 Oct.
Hartmann, M., Bäzner, E., Wild, B., Eisler, I., Herzog, W., 2010. Effects of inter-ventions involving the family in the treatment of adult patients with chronicphysical diseases: a meta-analysis. Psychotherepy Psychosomatic 79 (3), 136–148Feb 20.
Hutchison, S., Choo-Kang, B.S., Gibson, V.B., Bundick, R.V., Leishman, A.J., Brewer, J.M.,McInnes, I.B., Garside, P., 2009. An investigation of the impact of the location andtiming of antigen-specific T cell division on airways inflammation. Clinical andExperimental Immunology 155 (1), 107–116.
Joachim, R.A., Sagach, V., Quarcoo, D., Dinh, T., Arck, P.C., Klapp, B.F., 2006. Upregulationof tumor necrosis factor-alpha by stress and substance p in a murine model ofallergic airway inflammation. Neuroimmunomodulation 13 (1), 43–50.

21A. Deshmukh et al. / Journal of Neuroimmunology 225 (2010) 13–21
Jones, H.P., Tabor, L., Sun, X., Woolard, M.D., Simecka, J.W., 2002. Depletion of CD8+ T cellsexacerbates CD4+ Th cell-associated inflammatory lesions duringmurine mycoplasmarespiratory disease. Journal of immunology 168 (7), 3493–3501 Baltimore, Md.: 1950.
Jones, R.C., Donaldson, G.C., Chavannes, N.H., Kida, L., Dickson-Spillmann, M., Harding, S.,Wedzicha, J.A., Price, D., Hyland, M.E., 2009. Derivation and validation of a compositeindex of severity in chronic obstructive pulmonary disease: theDOSE Index. AmericanJournal of Respiratory and Critical Care Medicine 180 (12), 1189–1195.
Khazan, I., 2009. Psychophysiological stress assessment using biofeedback. Journal ofVisualized Experiments : JoVE 29 (29) 1443, 10.3791-1443.
Kool, M., van Nimwegen, M., Willart, M.A., Muskens, F., Boon, L., Smit, J.J., Coyle, A.,Clausen, B.E., Hoogsteden, H.C., Lambrecht, B.N., Hammad, H., 2009. An anti-inflammatory role for plasmacytoid dendritic cells in allergic airway inflammation.Journal of Immunology 183 (2), 1074–1082 Baltimore, Md.: 1950.
Larche,M., Robinson, D.S., Kay, A.B., 2003. The role of T lymphocytes in the pathogenesis ofasthma. The Journal of Allergy and Clinical Immunology 111 (3), 450–463 quiz 464.
Lewkowich, I.P., Herman, N.S., Schleifer, K.W., Dance, M.P., Chen, B.L., Dienger, K.M.,Sproles, A.A., Shah, J.S., Kohl, J., Belkaid, Y., Wills-Karp, M., 2005. CD4+ CD25+ T cellsprotect against experimentally induced asthma and alter pulmonary dendritic cellphenotype and function. The Journal of experimental medicine 202 (11), 1549–1561.
Li, C., Ford, E.S., Zhao, G., Balluz, L.S., Berry, J.T., Mokdad, A.H., 2010. Undertreatment ofmental health problems in adults with diagnosed diabetes and serious psychologicaldistress: the behavioral risk factor surveillance system, 2007. Diabetes Care 25 Feb.
Maddox, L., Schwartz, D.A., 2002. The pathophysiology of asthma. Annual Review ofMedicine 53, 477–498.
Marshall, G.D., 2004a. Internal and external environmental influences in allergicdiseases. Journal of American Osteopathic Association 104 (5 Suppl 5), S1–S6.
Marshall, G.D., 2004b. Neuroendocrine mechanisms of immune dysregulation: applica-tions to allergy and asthma. Annals of Allergy Asthma Immunology 93 (2 Suppl 1),S11-7.
Marshall Jr, G.D., Agarwal, S.K., 2000. Stress, immune regulation, and immunity:applications for asthma. Allergy and Asthma Proceedings: The Official Journal ofRegional and State Allergy Societies 21 (4), 241–246.
Mays, J.W., Bailey, M.T., Hunzeker, J.T., Powell, N.D., Papenfuss, T., Karlsson, E.A.,Padgett, D.A., Sheridan, J.F., 2010. Influenza virus-specific immunological memoryis enhanced by repeated social defeat. Journal of Immunology 184 (4), 2014–2025Feb 15.
McAlonan, G.M., Lee, A.M., Cheung, V., Cheung, C., Tsang, L.W., Sham, P.C., Chua, S.E.,Wong,J.G., 2007. Immediate and sustained psychological impact of an emerging infectiousdisease on health care workers. Canadian Journal of Psychiatry 52 (4), 241–247.
Mikics, E., Baranyi, J., Haller, J., 2008. Rats exposed to traumatic stress bury unfamiliarobjects—a novel measure of hyper-vigilance in PTSD models? Physiology &Behavior 94 (3), 341–348.
Nguyen, K.D., Vanichsarn, C., Nadeau, K.C., 2009. Impaired IL-10-dependent induction oftolerogenic dendritic cells by CD4 + CD25hiCD127lo/- natural regulatory t cells inhuman allergic asthma". American journal of respiratory and critical care medicine180 (9), 823–833 Nov 1.
Pascoe, E.A., 2009. Perceived discrimination and health: a meta-analytic review”.Psychology Bulletins 135 (4), 531–554 Jul.
Quarcoo, D., Pavlovic, S., Joachim, R.A., 2009. Stress and airway reactivity in a murinemodel of allergic airway inflammation. Neuroimmunomodulation 16 (5), 318–324.
Rafinski, T., 1975. Mental aspects of allergic diseases. Polski Tygodnik Lekarski 30 (5),217-9.
Randolph, G.J., Jakubzick, C., Qu, C., 2008. Antigen presentation by monocytes andmonocyte-derived cells. Current Opinion in Immunology 20 (1), 52–60.
Rybnikova, E., Mironova, V., Pivina, S., Tulkova, E., Ordyan, N., Vataeva, L., Vershinina, E.,Abritalin, E., Kolchev, A., Nalivaeva, N., Turner, A.J., Samoilov, M., 2007.Antidepressant-like effects of mild hypoxia preconditioning in the learnedhelplessness model in rats. Neuroscience Letters 417 (3), 234–239.
Sandberg, S., Paton, J.Y., Ahola, S., McCann, D.C., McGuinness, D., Hillary, C.R., Oja, H.,2000. Stress increases the risk of asthmatic attacks in children. DuodecimLaaketieteellinen Aikakauskirja 116 (20), 2305–2306.
Steptoe, A., O'Donnell, K., Marmot, M., Wardle, J., 2008. Positive affect and psychosocialprocesses related to health. British Journal of Psychology 99 (Pt 2), 211–227 May.
Su, M.W., Pyarajan, S., Chang, J.H., Yu, C.L., Jin, Y.J., Stierhof, Y.D., Walden, P., Burakoff, S.J.,2004. Fratricide of CD8+ cytotoxic T lymphocytes is dependent on cellular activationand perforin-mediated killing. European Journal of Immunology 34 (9), 2459–2470.
Subranmanian, S.V., Kennedy,M.H., 2009. Perception of neighborhood safety and reportedchildhood lifetime asthma in the United States (U.S.): a study based on a nationalsurvey. PLoS ONE 4 (6), e6091.
Valentino, R.J., Van Bockstaele, E., 2008. Convergent regulation of locus coeruleus activity asan adaptive response to stress. European Journal of Pharmacology 583 (2–3), 194–203.
Vegas, O., Garmendia, L., Arregi, A., Beitia, G., Azpiroz, A., 2009. Effects of antalarmin andnadolol on the relationship between social stress and pulmonary metastasisdevelopment in male OF1 mice. Behavioural Brain Research 205 (1), 200–206.
Wakashin, H., Hirose, K., Maezawa, Y., Kagami, S., Suto, A., Watanabe, N., Saito, Y., Hatano,M., Tokuhisa, T., Iwakura, Y., Puccetti, P., Iwamoto, I., Nakajima, H., 2008. IL-23 andTh17 cells enhance Th2-cell-mediated eosinophilic airway inflammation in mice.American journal of Respiratory and Critical Care Medicine 178 (10), 1023–1032.
Willart, M.A., Lambrecht, B.N., 2009. The danger within: endogenous danger signals,atopy and asthma. Clinical and experimental allergy: Journal of the British Societyfor Allergy and Clinical Immunology 39 (1), 12–19.
Wills-Karp, M., 1999. Immunologic basis of antigen-induced airway hyperresponsive-ness. Annual Review of Immunology 17, 255–281.
Wright, R.J., 2005. Stress and atopic disorders. The Journal of Allergy and ClinicalImmunology 116 (6), 1301–1306.
Wright, K., 2007. Examining racial and ethnic disparities and predictors of medicationuse among California's African-American, Latino, and White children with asthma.Journal of National Black Nurses' Association : JNBNA 18 (2), 1–15.
Yee, J.R., Cavigelli, S.A., Delgado, B., McClintock, M.K., 2008. Reciprocal affiliation amongadolescent rats during a mild group stressor predicts mammary tumors andlifespan. Psychosomatic Medicine 70 (9), 1050–1059.
Copyright © 2022 FDOKUMEN




















