
Putative-farnesoic acid O-methyltransferase (FAMeT) in medfly reproduction
Upload
independentCategory
view
0download
0
ELSEVIER Biochimica et Biophysica Acta 1251 (1995) 1-10
BB Biochl~Pic~a et Biophysica AEta
Cloning, expression and structure of catechol-O-methyltransferase
Kenneth Lundstri3m a,l,*, Jukka Tenhunen a Carola Tilgmann a, Tuula Karhunen b, Pertti Panula b, Ismo Ulmanen a
a Orion Corporation, Orion-Farmos, Orion Research, 02101 Espoo, Finland b Department of Biology, Abo Akademi University, 20520 Turku, Finland
Received 6 December 1994; revised 29 March 1995; accepted 3 April 1995
Keywords: cDNA; Gene expression; Recombinant protein; Transcription; Translation; Localization; Polymorphism; Atomic structure
C o n t e n t s
1. Introduction-multiple functions of the COMT enzyme . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2. S-and MB-COMTproteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
3. Cloning and ,;tructure of mammalian COMT cDNAs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
4. Cloning and structure of rat and human COMT genes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
5. Regulation of COMTexpression . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
6. Genetic polymorphism of COMT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
7. Localization of COMT in mammalian tissues . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
8. Intracellular localization of COMT proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
9. Expression of recombinant COMT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
9.1. Escherichia coli expression . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
9.2. Eukaryol:ic expression . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
10. Crystal structure of S-COMT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
11. Perspective . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
* Corresponding author. Present address: Department of Molecular Biology S.A., Glaxo Institute for Molecular Biology Geneva, 14. chemin des
Aulx-Case Postale 674, 1228 Plan-les-Ouates, Switzerland. Fax: + 41 22 7946965. i Present address: Department of Molecular Biology, Glaxo Institute for Molecular Biology, Geneva, Switzerland
0167-4838/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved
SSDI 01 6 7 - 4 8 3 8(95 ) 0 0 0 7 1 - 2
2 K. Lundstr6m et al. /Biochimica et Biophysica Acta 1251 (1995) 1-10
1. Introduct ion-mult iple functions of the C O M T en- zyme
Catechol-O-methyltransferase (COMT, EC 2.1.1.6) en- zyme catalyzes the transfer of a methyl group from S- adenosyl-L-methionine (AdoMet) to one of the hydroxyls (preferentially 3-hydroxyl) in a catechol substrate (Fig. 1). The substrates of COMT in mammals include catechol- amines with hormonal and neurotransmission activities (dopamine, norepinephrine, epinephrine, catecholestro- gens) and their metabolites, ascorbic acid [1-3], some indolic intermediates of melanin metabolism [4] and xeno- biotic catechols, like carcinogenic catechol-containing flavonoids [5]. Pharmacologically important is the methyla- tion and, consequently, inactivation of a multitude of drugs with a catechol structure [ 1,7].
Since the detection of COMT activity from mammals [8], the enzyme has been found in organisms at various phylogenic levels, from bacteria and yeast to man [1,9]. Activity determinations and immunohistochemical studies from different mammalian, mainly rat tissues, have clearly shown that COMT enzyme occurs in variable amounts in most tissues [1,2,10]. Thus it is not surprising that COMT has many different physiological roles [1,3,11] and has been suggested to participate in the development of vari- ous human disorders [1,12-14]. Except the preliminary report of Dunham et al. [15] suggesting a lesion in the chromosomal region of COMT in the velo-cardio-facial syndrome, so far no direct involvement of an aberrant COMT activity in any human disease has been demon- strated.
The different basic and applied aspects of COMT en- zyme have been recently extensively reviewed [ 1-3,6,9,16]. To avoid repetition, we decided here to concentrate on the progress achieved in the COMT research since the molecu- lar cloning of the mammalian COMT cDNAs and genes in 1990. This has resulted in a more elaborate understanding of the COMT polypeptides, their structure and function. Production of specific antisera using the recombinant anti- gens has allowed more detailed analysis of the localization of COMT proteins in tissues and cells. Further, with the aid of COMT clones, the regulation of COMT synthesis
HO CH 3 0
SAM SAH
Fig. 1. The O-rnethylation of the catechol substrate catalysed by COMT.
and the genetic aspects of COMT can now be experimen- tally studied.
2. S-and M B - C O M T proteins
Based on subcellular fractionation studies two major forms of the COMT enzyme, a soluble (S-COMT) and a membrane-bound form (MB-COMT), have been identified [16]. Protein purification has revealed some heterogeneity in COMT activity, stability and molecular weights [1]. Also multiple forms of COMT have been demonstrated in a variety of tissues and different animal species, supporting the hypothesis of isozymes [17,18].
In spite of several reports on S-COMT purification, primary structure data of the COMT polypeptide was not available until S-COMT was purified from rat liver [19], human placenta [20] and pig liver [21]. Tryptic peptide data from highly purified rat liver and human placental S-COMT [19,20] provided tools for the cloning and se- quencing of the COMT cDNAs. The predicted molecular mass is 24.7 kDa and 24,4 kDa for rat and human S-COMT, respectively. The apparent molecular weights in SDS- PAGE are 25 kDa for the purified rat liver and 26 kDa for human placental S-COMT proteins. The purification work also indicated that S-COMT is a monomeric enzyme [19,20], which later was confirmed by chemical cross-lin- king [22].
Protein and DNA data bank searches did not reveal any extensive homologies to any other reported sequences. However, a more refined computer-based comparison be- tween methyltransferases has revealed two consensus se- quences L D V / L GXGXG and LRPGGXL in mammalian
A. B.
kb 1 2 kb 3 4
2.4 - ~' 2 . 4 -
1 . 4 - -
k Da 1 2 3 4 kDa
- 30 ; MB...
30 - . S S
21.5 -
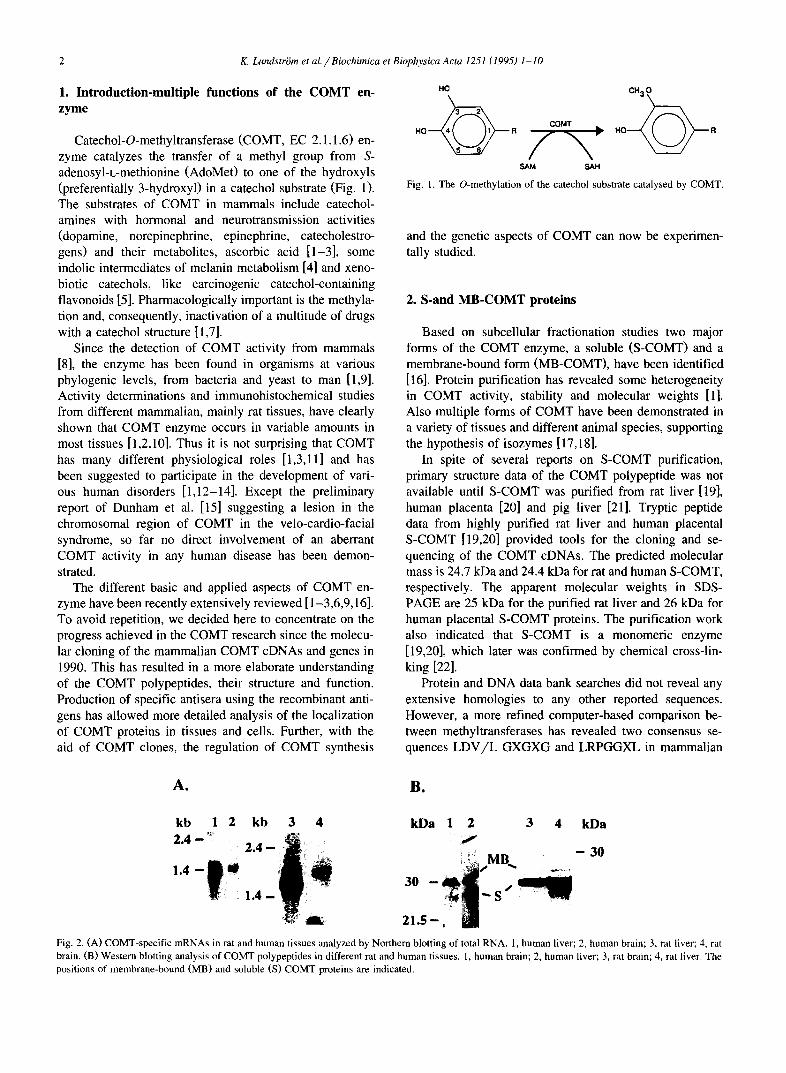
Fig. 2. (A) COMT-specific mRNAs in rat and human tissues analyzed by Northern blotting of total RNA. 1, human liver; 2, human brain; 3, rat liver; 4, rat brain. (B) Western blotting analysis of COMT polypeptides in different rat and human tissues. 1, human brain; 2, human liver; 3, rat brain; 4, rat liver. The positions of membrane-bound (MB) and soluble (S) COMT proteins are indicated.
K. LundstrUm et al. / Biochimica et Biophysica Acta 1251 (1995) 1-10 3
methyltransferases, corresponding to the amino acids LEL- GAYCG (containing the G-residue involved in AdoMet binding in COMT) and LRKGTVL in rat and human S-COMT [23-25].
The membrane associated COMT activity in different tissues consistently accounts for only a minor fraction of the total COMT activity [1,3]. Quantitative immuno- blotting analyses of rat arLd human tissue homogenates have shown that the S-COMT indeed predominates in rat, whereas in some human tissues, like brain, the MB-COMT protein is more abundant ('Fable 1 and Fig. 2). The bio- chemical characteristics of MB-COMT have been less extensively investigated than those of S-COMT. However, the MB-COMT enzyme has many properties in common with S-COMT [26-28], buLt differs from it in substrate specificity, kinetic propertie.s, subcellular localization and molecular weight [29-37]. Possibly the most distinct dif- ference between S-and MB-COMT is the higher affinity of MB-COMT for dopamine ~tnd some other substrates and the difference in the regioselectivity of methylation [38- 42].
Until the COMT cDNAs were cloned neither the exact molecular identity of MB-COMT nor the relationship be- tween S-and MB-COMT forms were established. Cloning of the human COMT [43,44] made it possible to express the recombinant MB-COM]" protein and characterize it biochemically. The amino terminal region of MB-COMT meets the criteria of a signal sequence with an extension of 43 or 50 amino acids, can'ying a stretch of 17 and 24
hydrophobic amino-acid residues in rat and man, respec- tively [43,44]. In vitro translation studies showed that the amino-terminus of this MB-COMT serves as a signal anchor sequence, directing the polypeptide co-translation- ally into the membrane [45]. The arrangement of positively charged amino acids spanning the putative anchor se- quence suggests that MB-COMT may be oriented towards the cytoplasmic side of the membrane, reminding thus membrane proteins of the class Ib type [46,47].
3. Cloning and structure of mammalian COMT cDNAs
Purification of COMT enzyme from rat [19] or porcine [21] liver allowed peptide sequencing and the design of COMT-specific oligonucleotides for hybridization screen- ings of cDNA libraries. It also enabled the production of COMT-specific polyclonal antibodies, which could be used for immunoscreenings of cDNA expression libraries. Im- munoscreenings yielded rat and porcine liver cDNA clones [21,48] and the nucleotide sequences could be verified to the tryptic peptide sequences of the purified COMT en- zymes. The rat clones contained an open reading frame of 663 nucleotides and a 739 nucleotides long 3' non-coding sequence carrying a putative polyadenylation site. The open reading frame codes for a 221 amino acid long S-COMT polypeptide. The deduced amino-acid sequence indicated that the S-COMT enzyme is a cytosolic protein with no hydrophobic signal sequence or transmembrane
S-AUG UGA l I
MB-AUG S-AUG UGA : :1 . . . .~" - -L '1_ . . .~ ~ " - - I _ ~ j I
MB-ATG S-ATG TGA
/'/ I I ,ooo I 268 I:)p 194 bp 132 I:q:) 937 bp
P2 P1
~p(A) 1.6 kb
ip(A) 1.9 kb
MB-ATG S-ATG TGA
I I,so , 400 4,00 , I I 110hi) 90bp 289bl) 194 I)p 132bp 4721)1)
gramme P2 Pl
MS-AUG S-AUG UGA
i l..,,J::3~.........J I , .... I ' ~ - 1 . . . ~ _ . . . . , . . . . . . . . I I ip(A) 1.5 kb
S-AUG UGA I I I i ~ r - - ' - l . . . . . . . ~ ,p(A) 1.3 kb
Fig. 3. The structure of rat (upper part) and human (lower part) COMT genes. Thin line represents introns and the boxes the exons. The black boxes indicate the protein coding regions. Two identified promoters, P1 and P2, are shown by black bars. COMT mRNA species expressed from the genes (1.6 and 1.9 kb in rat and 1.3 and 1.5 kb in human) are presented as white bars. The positions of translation initiation codons for MB-COMT polypeptide (MB-ATG, MB-AUG), S-COMT polypeptide (S-ATG, S-AUG) and for translation stop codons (TGA, UGA), and the sizes of exons and introns are also shown.

4 K. Lundstri3m et al. /Biochimica et Biophysica Acta 1251 (1995) 1-10
domains. Neither was any asparagine-linked N-glycosyla- tion sites found. Cloning of the rat COMT gene revealed an upstream open reading frame from the putative S-COMT ATG. This region coded for a stretch of 17 hydrophobic amino acids, which could act as a transmembrane domain responsible for the membrane association of the MB- COMT form. A putative initiation ATG for MB-COMT, located 129 nucleotides upstream from the S-COMT ATG, suggests that the two forms of COMT could be made from the same mRNA.
Cloning of the human COMT sequence from placental and Hep G2 cell line cDNA libraries using the rat or porcine cDNA probes, yielded clones with similar location of hydrophobic sequence as seen for the rat COMT: the deduced human MB-COMT polypeptide also contained a stretch of 24 hydrophobic amino acids at its amino termi- nus [43,44]. A putative ATG translation initiation codon for the MB-COMT was 150 nucleotides upstream from the S-COMT ATG. The coding region for the human S-COMT was 663 nucleotides giving a 221 amino acids long protein being thus of identical size as the rat S-COMT. The overall sequence similarity at the amino-acid level was 80% for the rat and human proteins and 83% between porcine and human proteins.
Isolation of cDNA clones from human liver and brain gave COMT clones which had identical sequences as the placental clones [49]. This supported the view that differ- ent human tissues, including the brain, contain identical COMT proteins.
4. Cloning and structure of rat and human COMT genes
The genes for rat [50] and human [49] COMT proteins have been cloned. The exon-intron organization of both genes was characterized by comparing the sequences of the known cDNA clones [43,44,48] and genomic regions, and by PCR analysis. The overall structure of the COMT gene is very similar in both species (Fig. 3). In rat the gene is composed of five exons, the first one, approx. 200 bp in length, being non-coding. The size of the 5' non-coding region in the human COMT gene is similar to that of rat, but is divided into two exons. The splice site for this 5' untranslated region to the first coding exon is identical in both genes. The translation initiation codons, MB-ATG (for MB-COMT polypeptide) and S-ATG (for S-COMT form) are located in the 2. and 3. exons in rat and human, respectively. The size of the 3' untranslated regions differs greatly, being 739 bp in rat and 274 bp in human. The sequence similarity of the coding region of COMT genes is 80% [43,44,48] and that of the 5' and 3' untranslated regions 68% and 64%, respectively [49].
The analysis of cell hybrids and in situ hybridization have shown that the human COMT gene is located in the chromosome 22 q 11.2 [51,52], confirming the earlier
biochemical analysis of cell hybrids [53]. Southern hy- bridization together with the in situ data suggest that in mammals there is only one locus for COMT [43,48,51].
The expression of the COMT gene is controlled by two distinctive promoter regions [49,50]. The 5' promoter (P2) regulates the synthesis of 1.9 kb and 1.5 kb mRNA species in rat and human, respectively. This mRNA has a capacity to code for both MB-and S-COMT proteins by using the leaky scanning mechanism of translation initiation [45,54]. The expression of the shorter transcript, 1.6 kb in rat and 1.3 kb in human, is regulated by the P1 promoter, which is located between the S-and MB-COMT ATG start codons and partly overlaps the MB-COMT coding sequence. MB- AUG translation initiation codon is not included in these transcripts and, hence, they are able to code for S-COMT polypeptide only (Fig. 2 and Fig. 3).
5. Regulation of COMT expression
Northern blot analysis of total RNAs from different rat and human tissues and cell lines revealed the presence of two COMT-specific transcripts (1.6 kb and 1.9 kb in rat and 1.3 kb and ! .5 kb in human) (Fig. 2 and Fig. 3). The larger mRNA form is present in all the samples studied showing that the upper promoter (P2) is constitutively expressed. In contrast, the lower promoter (P1) is regulated in a tissue-specific manner and, hence, the amount of the shorter transcript varies from one tissue to another. The highest amounts were found in the liver and kidney both in rat and human. Only the larger 1.9 kb mRNA is present in different regions of rat brain, but in human brain also a small amount of the shorter transcript can be found. Most of the cell lines studied by Northern analysis seem to contain only the larger mRNA species, the exception being human breast adenocarcinoma cell line MCF-7 where the shorter transcript is dominating.
The structures of the COMT mRNAs has been analysed in rat and human by conventional primer extension and nuclease protection methods and the size difference of the two COMT mRNA species has been located in the 5' end of the molecules [49,50]. The shorter transcript initiates at multiple sites in the region between the two translation initiation ATG codons in both species. In rat the initiation occurs approximately 17-18 nucleotides upstream of the S-ATG codon, in human the predominant site is located somewhat further upstream. In rat the region in front of the MB-AUG codon includes two alternative splicing acceptor sites 27 bp apart, which both are used when the non-cod- ing exon is spliced to the first coding exon (Fig. 3). These two alternative splicing products have different capacity for the translation of MB- and S-COMT polypeptides [54]. The presence of such alternative spliced COMT mRNAs has not been studied in human tissues. In both species the larger COMT mRNA also has several 5' ends.
The S-and MB-COMT protein expression has been

K. Lundstr6m et al. / Biochimica et Biophysica Acta 1251 (1995) 1-10 5
studied in different rat and human tissues by Western blotting (Table 1 and Fig. 2.). The S-COMT protein form dominates in most of the tissues studied. However, in brain tissues there is a clear difference in COMT protein patterns between rat and human. In rat the S-COMT form repre- sents approx. 70-80% of the total COMT proteins whereas in human brain only 30% of COMT proteins is in S-form. The difference in the expressed protein levels can be explained by different translation efficiency of COMT polypeptides from the larger COMT mRNA; in human the MB-AUG codon resides in a more favourable context compared to the rat counterpart. Furthermore, in rat the translation initiation context for the S-AUG codon is opti- mal, whereas in human it is sub-optimal according to the Kozak rules of translation initiation. In most of the other tissues the level of the shorter COMT mRNA, capable of expressing only the S-COMT polypeptide, exceeds that of the longer transcript. This raay explain the domination of the S-COMT form in these tissues. Mutational analysis of the translation initiation contexts around MB-and S-AUG codons also support the participation of the leaky scanning mechanism of translation initiation in the expression of COMT polypeptides [45,54]. The involvement of prote- olytic processing of the MB-COMT form to yield S-COMT has been suggested [41,44]. However, in vitro and in vivo studies do not support this model [45,54].
In summary, the regulation of the COMT gene expres- sion seems to occur at several levels: (i) at transcription initiation, (ii) at translation initiation and (iii) at mRNA splicing. The gene organization together with the complex regulation enables the expression of two different COMT proteins from a single gene in rat and human.
6. Genetic polymorphism ~f COMT
The existence of a thermolabile low activity COMT e form and a thermostable high activity COMT r~ polymor- phic form has been reported [55-57]. The genetics of the COMT activity polymorphism has been reviewed earlier [58,59]. The allelic differences in the COMT activity seem to have shown some clinical significance in the responses of Parkinsonian patients to the levo-DOPA therapy [60]. The molecular difference between the COMT alleles has not been known, but recently it was reported that the COMT polymorphism in rrmn would be based on one amino-acid difference in the S-COMT position 108 (Val = high activity allele, Met = low activity allele) [56]. No- tably the two published cDNA sequences of human soluble COMT differ in this position, suggesting that they repre- sent copies of different alleles [43,44]. The recombinant allelic forms of human S-COMT produced in E. coli, indeed were found to reveal different thermal stabilities in vitro [42]. This confirmed that the amino acid 108 would be relevant for the COMT activity polymorphism.
7. Localization of COMT in mammalian tissues
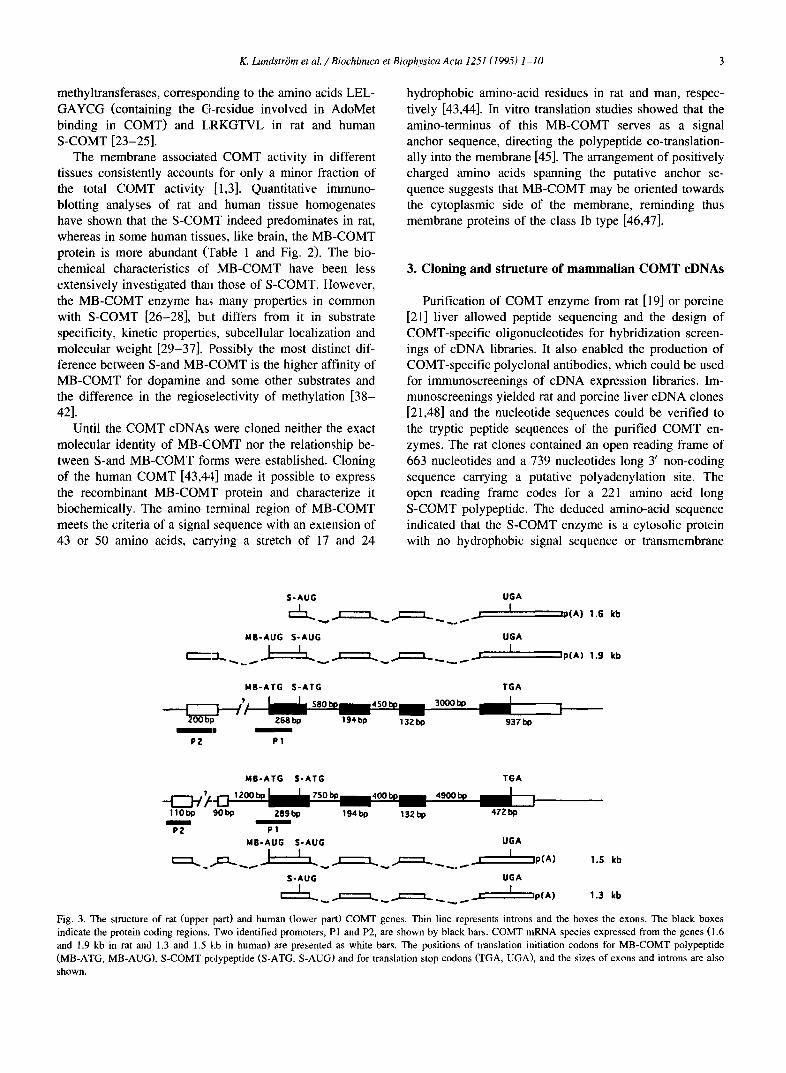
The immunohistochemical distribution of COMT in many rat tissues has been recently studied by using a COMT specific guinea pig antiserum raised against recom- binant rat COMT protein [9]. The COMT protein was found to occur in most organs and tissues and the distribu- tion of fluorescence was identical with that obtained with the antiserum raised against the native rat liver enzyme. An intense COMT immunoreaction was seen in hepato- cytes in the liver, tubuli and the collecting ducts of the kidney (Fig. 4a), perinodular ring in the spleen, somato- statin- and insulin-storing endocrine cells but not glucagon-storing cells of the pancreas, pituicytes in the posterior pituitary and in cleft cells of the intermediate lobe (Fig. 4d).
In rat, chinchilla and bovine brain, only non-neuronal structures have been found to display immunoreactivity for COMT [61-63]. However, COMT activity has been re- ported from fractions with enriched nerve terminals [64]. Intense COMT immunoreactivity in brain has been re- ported in neuroglial cells of the circumventricular organs [62], epithelial cells of the choroid plexus, cells in the pia and arachnoidea [63], ventricular ependyma, oligodendro- cytes and astrocytes including the Bergmann glia of the cerebellum [61]. Recent studies with antisera against the recombinant COMT enzyme shows clear evidence of both glial and neuronal COMT in the rat brain [9,65]. In the cerebral cortex COMT immunoreactivity resides in the neuropil (Fig. 4b). A strong immunoreaction is character- istic of the ventricular ependyma (Fig. 4c). COMT has also been localized in cultured cerebra[ endothelial cells and smooth muscle cells [66].
Immunoelectron microscopy suggests that the enzyme
Table 1 Quantitation of S-and MB-COMT polypeptides in rat and human tissues and two human cell lines expressed as % of total COMT in the im- munoblot
Tissue S-COMT MB-COMT
Rat Liver 93 7 Kidney 75 25 Heart 79 21 Cerebellum 86 14 Telencephalon 69 31
Human tissues and cells Liver 85 15 Kidney 77 23 Adrenal 74 26 Duodenum 89 11 Brain 30 70 HeLa-cells 35 65 MCF7-cells 92 8
The tissue homogenates were analysed by Western blotting and the amounts of S-and MB-COMT proteins were quantitated by densitometry (Tenhunen and Ulmanen [54]; Tenhunen et al. [49]).
6 K. LundstriSm et al. /Biochimica et Biophysica Acta 1251 (1995) 1-10
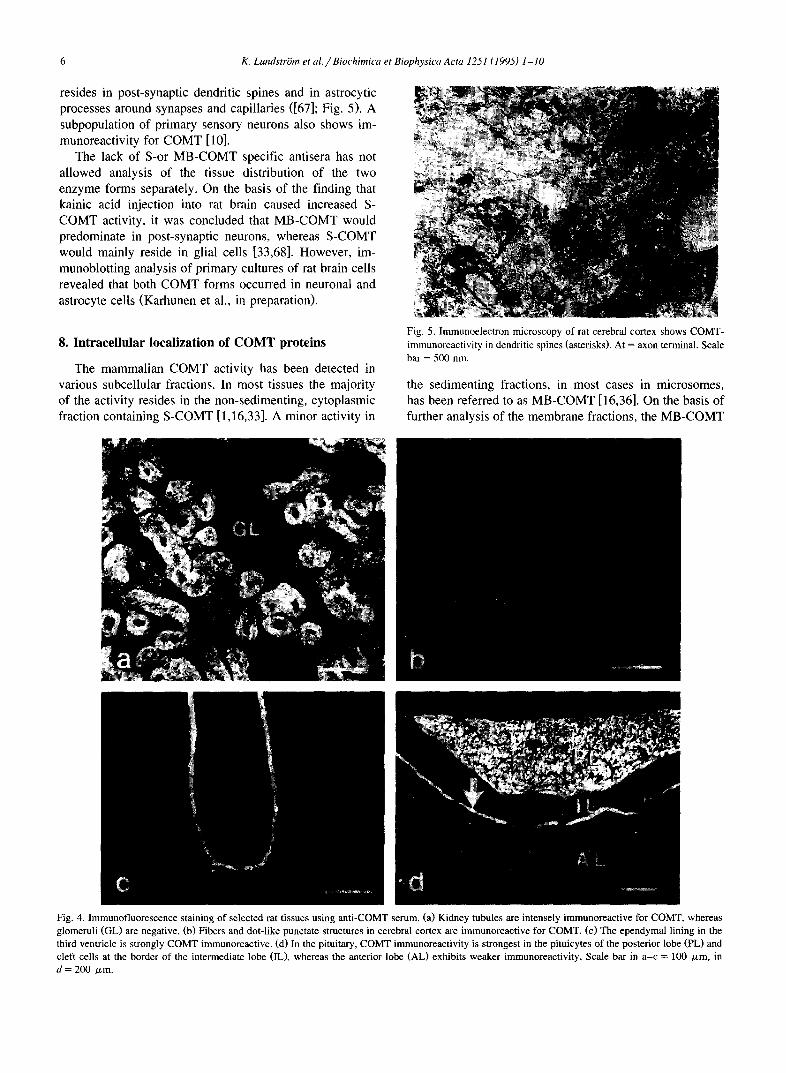
resides in post-synaptic dendritic spines and in astrocytic processes around synapses and capillaries ([67]; Fig. 5). A subpopulation of primary sensory neurons also shows im- munoreactivity for COMT [10].
The lack of S-or MB-COMT specific antisera has not allowed analysis of the tissue distribution of the two enzyme forms separately. On the basis of the finding that kainic acid injection into rat brain caused increased S-
COMT activity, it was concluded that MB-COMT would predominate in post-synaptic neurons, whereas S-COMT would mainly reside in glial cells [33,68]. However, im-
munoblotting analysis of primary cultures of rat brain cells revealed that both COMT forms occurred in neuronal and astrocyte cells (Karhunen et al., in preparation).
8. Intracellular localization of COMT proteins
The mammalian COMT activity has been detected in
various subceUular fractions. In most tissues the majority of the activity resides in the non-sedimenting, cytoplasmic fraction containing S-COMT [1,16,33]. A minor activity in
Fig. 5. Immunoelectron microscopy of rat cerebral cortex shows COMT- immunoreactivity in dendritic spines (asterisks). At = axon terminal. Scale bar = 500 nm.
the sedimenting fractions, in most cases in microsomes, has been referred to as MB-COMT [16,36]. On the basis of further analysis of the membrane fractions, the MB-COMT
Fig. 4. Immunofluorescence staining of selected rat tissues using anti-COMT serum. (a) Kidney tubules are intensely immunoreactive for COMT, whereas glomeruli (GL) are negative. (b) Fibers and dot-like punctate structures in cerebral cortex are immunoreactive for COMT, (c) The ependymal lining in the third ventricle is strongly COMT immunoreactive. (d) In the pituitary, COMT immunoreactivity is strongest in the pituicytes of the posterior lobe (PL) and cleft cells at the border of the intermediate lobe (IL), whereas the anterior lobe (AL) exhibits weaker immunoreactivity, Scale bar in a-c = 100 /zm, in d = 200 /xm.
K. Lundstr6m et al. / Biochimica et Biophysica Acta 1251 (1995) 1-10 7
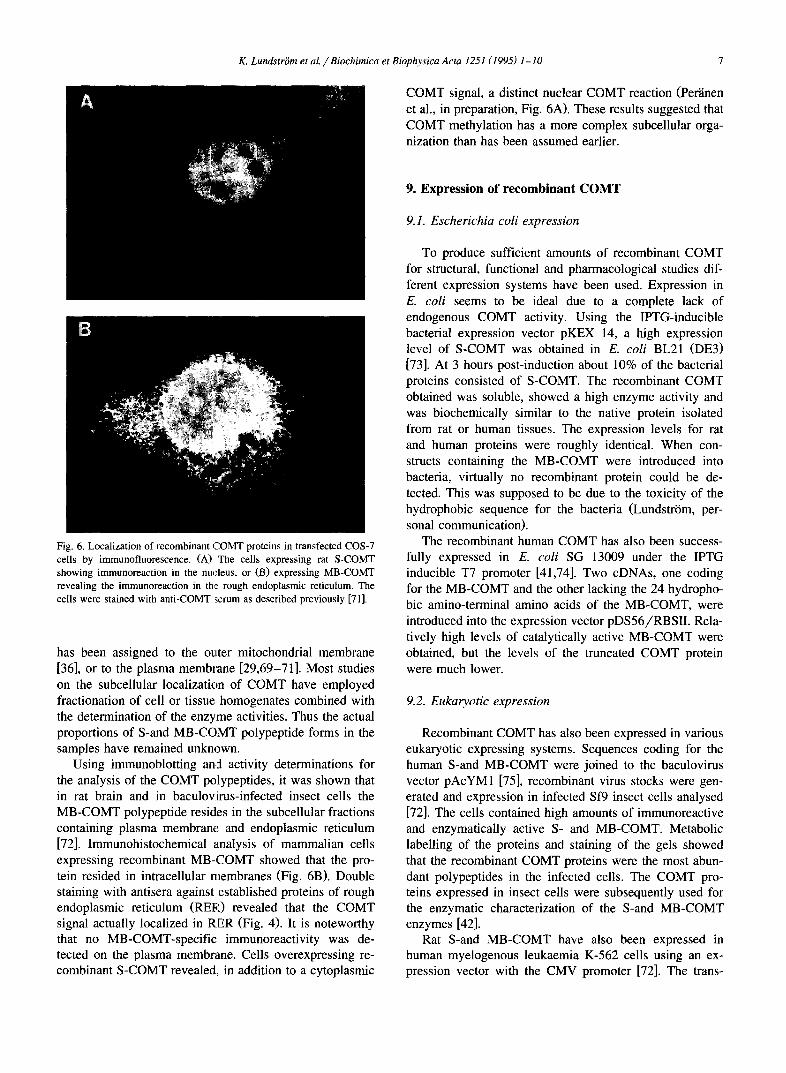
COMT signal, a distinct nuclear COMT reaction (Per'dnen et al., in preparation, Fig. 6A). These results suggested that COMT methylation has a more complex subcellular orga- nization than has been assumed earlier.
Fig. 6. Localization of recombinant COMT proteins in transfected COS-7 cells by immunofluorescence. (A) The cells expressing rat S-COMT showing immunoreaction in the nucleus, or (B) expressing MB-COMT revealing the immunoreaction in the rough endoplasmic reticulum. The cells were stained with anti-COMT serum as described previously [71].
has been assigned to the outer mitochondrial membrane [36], or to the plasma membrane [29,69-71]. Most studies on the subcellular localization of COMT have employed fractionation of cell or tissue homogenates combined with the determination of the enzyme activities. Thus the actual proportions of S-and MB-C(3,MT polypeptide forms in the samples have remained unknown.
Using immunoblotting and activity determinations for the analysis of the COMT polypeptides, it was shown that in rat brain and in baculovirus-infected insect cells the MB-COMT polypeptide resides in the subcellular fractions containing plasma membrane and endoplasmic reticulum [72]. Immunohistochemical analysis of mammalian cells expressing recombinant MB-COMT showed that the pro- tein resided in intracellular membranes (Fig. 6B). Double staining with antisera against established proteins of rough endoplasmic reticulum (RER) revealed that the COMT signal actually localized in RER (Fig. 4). It is noteworthy that no MB-COMT-specific immunoreactivity was de- tected on the plasma membrane. Cells overexpressing re- combinant S-COMT revealed, in addition to a cytoplasmic
9. Expression of recombinant COMT
9.1. Escherichia coli expression
To produce sufficient amounts of recombinant COMT for structural, functional and pharmacological studies dif- ferent expression systems have been used. Expression in E. coli seems to be ideal due to a complete lack of endogenous COMT activity. Using the IPTG-inducible bacterial expression vector pKEX 14, a high expression level of S-COMT was obtained in E. coli BL21 (DE3) [73]. At 3 hours post-induction about 10% of the bacterial proteins consisted of S-COMT. The recombinant COMT obtained was soluble, showed a high enzyme activity and was biochemically similar to the native protein isolated from rat or human tissues. The expression levels for rat and human proteins were roughly identical. When con- structs containing the MB-COMT were introduced into bacteria, virtually no recombinant protein could be de- tected. This was supposed to be due to the toxicity of the hydrophobic sequence for the bacteria (LundstrSm, per- sonal communication).
The recombinant human COMT has also been success- fully expressed in E. coli SG 13009 under the IPTG inducible T7 promoter [41,74]. Two cDNAs, one coding for the MB-COMT and the other lacking the 24 hydropho- bic amino-terminal amino acids of the MB-COMT, were introduced into the expression vector pDS56/RBSII. Rela- tively high levels of catalytically active MB-COMT were obtained, but the levels of the truncated COMT protein were much lower.
9.2. Eukaryotic expression
Recombinant COMT has also been expressed in various eukaryotic expressing systems. Sequences coding for the human S-and MB-COMT were joined to the baculovirus vector pAcYM1 [75], recombinant virus stocks were gen- erated and expression in infected Sf9 insect cells analysed [72]. The cells contained high amounts of immunoreactive and enzymatically active S- and MB-COMT. Metabolic labelling of the proteins and staining of the gels showed that the recombinant COMT proteins were the most abun- dant polypeptides in the infected cells. The COMT pro- teins expressed in insect cells were subsequently used for the enzymatic characterization of the S-and MB-COMT enzymes [42].
Rat S-and MB-COMT have also been expressed in human myelogenous leukaemia K-562 cells using an ex- pression vector with the CMV promoter [72]. The trans-

8 K. LundstriSm et aL/Biochimica et Biophysica Acta 1251 (1995) 1-10
fected stable cell lines showed a 5-to 10-fold increase in COMT activity but the amount of the immunoreactive recombinant proteins was relatively low. The immuno- reactive and active human MB-COMT has also been ex- pressed in transfected human embryonic kidney fibroblast 293 cell lines under the CMV promoter [41,44]. Cell fractionation indicated that the recombinant COMT was mainly associated with the microsomal membranes. Re- cently, expression of rat S-and MB-COMT has been per- formed using the transient expression in monkey COS-7 cells as well as the vaccinia virus expression system in human HeLa-and hamster BHK-cells ([54], Per~inen et al., in preparation). Introduction of the MB-COMT constructs into mammalian cells always yielded both MB-and S- COMT polypeptides. This supported the conclusion based on the in vitro translation experiments [45] that the two COMT proteins could be produced from the same mRNA. Pulse-chase experiments in transfected and native cells excluded the possibility that S-COMT would have been processed from the MB-COMT protein by proteolysis [54]. In order to produce only recombinant MB-COMT in mam- malian cells for the immunocytochemical analysis (see above), the S-COMT initiation codon in the MB-COMT sequence was mutated into an alanine codon. Cells trans- fected with this mutant construct produced only recombi- nant MB-COMT with no detectable S-COMT (Per~inen et al., in preparation).
10. Crystal structure of S-COMT
The recombinant rat S-COMT protein has been puri- fied, crystallized and its atomic structure recently deter- mined at 2.0 A resolution [23,73,76]. The conditions of the crystallization in the presence of magnesium, co-enzyme AdoMet and inhibitor 1,5-dinitrocatechol have been de- scribed in detail by Vidgren et al. [76]. The COMT structure has an a//3-fold containing eight alpha-helices and seven beta-strands. The AdoMet-binding motif has a close similarity with the NAD-binding structure of alco- holdehydrogenase. The active site of the enzyme is formed in the vicinity of the Mg 2÷ by residues from the amino terminal helices, AdoMet-binding fold (Rossman fold) and the loop in the C-terminal /3-strands. The inhibitor and probably also the substrates are bound to a shallow groove on the surface of the enzyme. Modelling of dopamine to the active site of COMT suggests that both hydroxyls of the substrate are coordinated to the Mg 2÷ ion. One hy- droxyl group is tetrahedrally surrounded by the Mg 2+ ion, the amino group of Lys-144, and the methyl group of the coenzyme. The other hydroxyl of the catechol ring forms a hydrogen bond to Glu-199. Trp-38 also contributes to the specificity of COMT for aromatic dihydroxy compounds. The structure and site-directed mutagenesis experiments [2,77] strongly suggest that the four cysteine residues of rat S-COMT are not important for the catalysis. AdoMet has
no interaction with magnesium, and kinetic experiments as well as the structure clearly indicate that AdoMet binds to the enzyme prior to Mg 2+ [42]. On the basis of the atomic structure the catalytic mechanism, differences of the sub- strate methylation as well as the inhibitor binding can be explained [23].
11. Perspective
The cloning of rat and human COMT has provided tools to address many of the open questions concerning the structure, functions and expression of this important en- zyme.
We can now quite safely conclude that the S-and MB- COMT polypeptides deduced from the cDNA clones and detected from mammalian tissues with the specific antisera are responsible for the enzymatic COMT activity. Any indications of other isoforms of COMT do not exist. The intracellular localization of the two COMT proteins needs further investigations. Of particular importance will be application of immunoelectron microscopy to unravel the localization of COMT in the mammalian central nervous system. The present data suggests that the MB-COMT is located on the rough endoplasmic reticulum and oriented towards cytoplasm. If this holds true for neurons, it is unlikely that COMT would be the methylating activity on the post-synaptic neural membrane.
Isolation of the rat and human COMT gene and analysis of COMT mRNAs suggested that the regulation of COMT synthesis may take place at several levels of expression: transcription initiation, splicing and translation initiation. Analysis of the DNA sequences responsible for the activity of the two COMT promoters will possibly reveal some regulation mechanisms. Future studies applying the COMT specific molecular probes can be directed to the expression of COMT during mammalian development and under vari- ous physiological conditions. We have now also better possibilities to find out whether abnormal expression of COMT is connected with some human disorders.
The determination of the atomic structure of rat S- COMT will be valuable for the understanding of the catalytic mechanism of the enzyme. The predictions based on the structure and molecular modelling can now be tested by using site-directed mutagenesis and expression of recombinant enzymes. The structure can further be utilized in order to develop new COMT targetted pharmacological compounds.
Acknowledgements
We thank Dr. Timo Lotta, Orion-Farmos, for com- ments. This work was supported by grants from the Medi- cal Research Council of Academy of Finland, The Sigrid

K. Lundstr6m et aL / Biochimica et Biophysica Acta 1251 (1995) 1-10 9
Juselius Foundation and Research and Science Foundation of Farmos (to T.K. and P.P.).
References
[1] Guldberg, H.C. and Marsden, C.A. (1975) Pharmacol. Rev. 27, 135 - 206.
[2] M[innisto, P., Ulmanen, I., LundstriSm, K., Taskinen, J., Tilgmann, C. and Kaakkola, S. (1992) in Progress in Drug Research (Jucker, E., ed.), pp. 291-350, Birkhauserg, Basel.
[3] Kopin, I. (1985) Pharmacol. Rev. 37, 334-364. [4] Smit, N., Pavel, S., Kammeyer, A. and Westerhof, W. (1990) Anal.
Biochem. 190, 286-291. [5] Zhu, B.T. and Liehr, J.G. (1994)Toxicol. Appl. Pharmacol. 125,
149-158. [6] Kaakkola, S., Gordin, A. and M~innisto, P. (1994) Gen. Pharmac. 25,
813-824. [7] Alberici, M., Rodriguez de Lores Arnaiz, G. and De Robertis, E.
(1965) Life Sci. 4. 1951-196,9. [8] Axelrod, J. (1957) Science 17.6, 400-401. [9] M~innisto, P., Ulmanen, I., Taskinen, J. and Kaakkola, S. (1994) in
Design of Enzyme lnhibitors as Drugs (Sandier, M. and Smith, H., eds.), pp. 621-647, Oxford University Press, Oxford.
[10] Karhunen, T., Tilgmann, C., Julkunen, I., Ulmanen, I. and Panula, P. (1994) J. Histochem. Cytochem. 42, 1079-1090.
[11] Meister, B., Bean, A.J. and Aperia, A. (1993) Kidney Int. 44, 726-733.
[12] Karege, F., Bovier, P., Gaillard, J.-M. and Tissot, R. (1987) Acta Psychiatr. Scand. 76, 303-308.
[13] Mathew, R.J., Ho, B.T. Kralik, P., Taylor, D. and Claghorn, J.L. (1980) J. Clin. Psychiatr. 41 (12, Sec. 2), 25-27.
[14] Philippu, G., Hoo, J.J., Mile(zh, U., Argawall, D.P., Schrappe, O. and Goedde, H.W. (1981) Ps:ychiatr. Res. 4, 139-146.
[15] Dunham, I., Collins, J., Wadey, R. and Scambler, P. (1992) The Lancet 340, 1361-1362.
[16] Roth, J. (1992) Rev. Physiol. Biochem. Pharmacol. 120, 1-29. [17] Assicot, B. and Bohuon, C. (1971) Biochimie 53, 871-874. [18] Huh, M. and Friedhoff, A. (1979) J. Biol. Chem. 254, 299-308. [19] Tilgmann, C. and Kalkkinen, N. (1990) FEBS Lett. 264, 95-99. [20] Tilgmann, C. and Kalkkinen, N. (1991) Biochem. Biophys. Res.
Commun. 174, 995-1002. [21] Bertocci, B., Garotta, G., DaF'rada, M., Lahm, H.-W., Zurcher, G.,
Virgalita, G. and Miggiano, V. (1991) Biochim. Biophys. Acta 1080, 103-109.
[22] Melen, K., Ronni, T., Broni, B., Krug, R., Bonsdorff, C.-H. and Julkunen, I. (1992) J. Biol, Chem. 267, 25898-25907.
[23] Vidgren, J., Svensson, A. and Liljas, A. (1994) Nature 368, 354-358. [24] Gomi, T., Tanihara, K., Date,, T. and Fujioka, M. (1992) Int. J.
Biochem. 24, 1639-1649. [25] Fujioka, M. (1992) Int. J. Bioc, hem. 24, 1917-1924. [26] Borchardt, R., Cheng, C-F., Cooke, P. and Creveling, C. (1974) Life
Sci. 14, 1089-1100. [27] Rivett, A. and Roth, J. (1982) Biochemistry 21, 1740-1742. [28] Jeffery, D. and Roth, J. (1984) J. Neurochem. 42, 826-832. [29] Aprille, J. and Malamud, D. (1975) Biochem. Biophys. Res. Com-
mun. 64, 1293-1302. [30] Roffman, M., Reigle, T., Orsulak, P. and Schildkraut, J. (1976)
Biochem, Pharmacol. 25, 208-209. [31] Tong, J. and D'Iorio, A. (1977) Can. J. Biochem. 55, 1108-1113. [32] Roth, J. (1980) Biochem. Pharmacol. 29, 3119-3122. [33] Rivett, A., Francis, A. and Poth, J. (1983a) J. Neurochem. 40,
215-219.
[34] Rivett, A., Francis, A. and Roth, J. (1983b) J. Neurochem. 40, 1494-1496.
[35] Trendelenburg, U. (1980) Rev. Physiol. Biochem. Pharmacol. 87, 33-115.
[36] Grossman, M., Creveling, C., Rybczynski, R., Braverman, N., Iser- sky, C. Breakefield, X. (1985) J. Neurochem. 44, 421-432,
[37] Trendelenburg, U. (1988) Exp. Pharmacol. 90, 279-319. [38] Creveling, C., Morris, N., Schimizu, H., Ong, H. and Daly, J. (1972)
Mol. Pharmacol. 8, 398-409. [39] Nissinen, E. (1984) Biochem. Pharmacol. 33, 3105-3108. [40] Jeffery, D. and Roth, J. (1987) Biochemistry 26, 2955-2958. [41] Malherbe, P., Bertocci, B., Caspers, P., Zurcher, G. and DaPrada,
M. (1992) J. Neurochem. 58, 1782-1789. [42] Lotta, T., Vidgren, J., Tilgmann, C., Ulmanen, I., Melen, K.,
Julkunen, I. and Taskinen, J. (1995) Biochemistry, 34, 4202-4210. [43] LundstrBm, K., Salminen, M., Jalanko, A., Savolainen, R. and
Ulmanen, I. (1991) DNA Cell Biol. 10, 181-189. [44] Bertocci, B., Miggiano, V., DaPrada, M., Dembic, Z., Lahm, H.-W.
and Malherbe, P. (1991) Proc. Natl. Acad. Sci. USA 88, 1416-1420. [45] Ulmanen, I. and LundstrBm, K. (1991) Eur. J. Biochem. 202,
1013-1020. [46] Von Heijne, G. and Gavel, Y. (1988) Eur. J. Biochem. 174, 671-678. [47] Singer, S.J. (1990) Annu. Rev. Cell Biol. 6, 247-296. [48] Salminen, M., LundstrBm, K., Tilgmann, C., Savolainen, R., Kalkki-
nen, N. and Ulmanen, I, (1990) Gene 93, 241-247. [49] Tenhunen, J., Salminen, M., LundstrBm, K., Kiviluoto, T.,
Savolainen, R. and Ulmanen, I. (1994) Eur. J. Biochem. 223, 1049 - 1059.
[50] Tenhunen, J., Salminen, M., Jalanko, A., Ukkonen, S. and Ulmanen, I. (1993) DNA Cell Biol. 12, 253-263.
[51] Winqvist, R., Lundstr~Sm, K., Salminen, M., Laatikainen, M. and Ulmanen, I. (1992) Cytogenet. Cell Genet. 59, 253-257.
[52] Grossman, M. H., Emanuel, B.S. and Budart, M.L. (1992) Genomics 12, 822-825.
[53] Brahe, C., Bannetta, B., Meera, K., Arwert, F. and Serra, A. (1986) Human Genet. 74, 230-234.
[54] Tenhunen, J. and Ulmanen, I. (1993) Biochem. J. 296, 595-600. [55] Boudikova, B., Szumlanski, C., Maidak, B. and Weishilboum, R.
(1990) Clin. Pharmacol. Ther. 48, 381-389. [56] Grossman, M.H., Littrell, J.B., Weinstein, R., Szumlanski, C.L. and
Weischilboum, R.M. (1992) Soc. Neurosci. 18, 70. [57] Klemetsdal, B., Straume, B., Giverhaug, T. and Aarbakke, J. (1994)
Eur. J. Pharmacol. 46, 231-235. [58] Weischilboum, R. (1988) Clin. Biochem. 21 ,201-210 [59] Evans, D.A.P. (1993) Genetic Factors in Drug Therapy, Clinical and
Molecular Pharmacogenetics, pp. 338-341, Cambridge Univ. Press, Cambridge.
[60] Rivera-Calimlim, L. and Reilly, D.K. (1984) Clin. Pharmacol. Ther. 28, 278-286.
[61] Kaplan, G.P., Hartman, B.K. and Creveling, C.R. (1979) Brain Res. 167, 241-250.
[62] Kaplan, G.P., Hartman, B.K. and Creveling, C.R. (1981)Brain Res. 204, 353-360.
[63] Kaplan, G.P., Hartman, B.K. and Creveling, C.R. (1981) Brain Res. 229, 323-335.
[64] Alberici, M., Rodriguez de Lores Arnaiz, G. and De Robertis, E. (1965) Life Sci. 4, 1951-1960,
[65] Karhunen, T., Tilgmann, C., Per'~inen, J., Ulmanen, I. and Panula, P. (1993) Soc. Neurosci. Abstr. 19 (Part 1), 739.
[66] Spatz, M., Kaneda, N., Sumi, C., Nagatsu, I., Creveling, C.R. and Nagatsu, T. (1986) Brain Res. 381,363-367.
[67] Karhunen, T., Tilgmann, C., Per'~inen, J., Ulmanen, I. and Panula, P. (1995) Neurosci. Lett. 187, 1-4.
[68] Kaakkola, S., M~innisto, P.T. and Nissinen, E. (1987) J. Neural Transm. 69, 221-228.
[69] Lowe, M.C. and Creveling, C.R. (1979) in Catecholamines: Basic

10 K. Lundstri~m et al. / Biochimica et Biophysica Acta 1251 (1995) 1-10
and Clinical Frontiers (Usdin, E., ed.), pp. 219-221, Pergamon Press, Elmsford, NY.
[70] Raxworthy, M.J., Gulliver, P.A. and Hughes, P.J. (1982) Naunyn- Schmiedeberg's Arch. Pharmacol. 320, 182-188.
[71] Head, R.J., Irvice, R.J., Barone, S., Stitzel, R.E. and De la Lande, T.S. (1985) J. Pharmacol. Exp. Ther. 234, 184-189.
[72] Tilgmann, C., Melen, K., Lundstr~Sm, K., Jalanko, A., Julkunen, I., Kalkkinen, N. and Ulmanen, I. (1992) Eur. J. Biochem. 207, 813- 821.
[73] Lundstr~Sm, K. Tilgmann, C., Per'~inen, J., Kalkkinen, N. and Ulma- nen, I. (1992)Biochim. Biophys. Acta 1129, 149-154.
[74] Vilbois, F., Caspers, P., DaPrada, M., Lang, G., Karrer, C., Lahm, H.-W. and Cesura, A.M. (1994) Eur. J. Biochem. 222, 377-386.
[75] Matsuura, Y., Possee, R.D., Overton, H.A. and Bishop, D.H.L. (1987) J. Gen. Virol. 68, 1233-1250.
[76] Vidgren, J., Tilgmann, C., LundstriSm, K. and Liljas, A. (1991) Proteins 11,233-236.
[77] LundstrSm, K., Ahti, H. and Ulmanen, I. (1991) in Site-Directed Mutagenesis and Protein Engineering (EI-Gewely, M.R., ed.), pp. 119-122, Elsevier, Amsterdam.
Copyright © 2022 FDOKUMEN




















