
Structural studies on enzymes from Salmonella typhimurium ...

© 2002 Blackwell Science Ltd
The invasion-associated type III secretion system ofSalmonella enterica serovar Typhimurium is necessaryfor intracellular proliferation and vacuole biogenesis inepithelial cells
mutant bacteria displayed abnormal maturation that was dependent on the mechanism of entry.Manipulation of Salmonella-containing vacuole (SCV)biogenesis by pharmacologically perturbing mem-brane trafficking in the host cell increased intra-cellular replication of wild-type but not mutant S. Typhimurium This demonstrates a previouslyunknown role for SPI-1 in vacuole biogenesis andintracellular survival in non-phagocytic cells.
Introduction
Following oral ingestion, Salmonella must cross theintestinal epithelium, and pathogenicity of these bacteriadepends on their ability to invade epithelial cells; (Galanand Curtiss, 1989; Clark et al., 1998). Our understandingof this process is largely from in vitro studies using cul-tured epithelial cells and Salmonella enterica serovarTyphimurium (S. Typhimurium), a common cause ofenteritis in humans. Such studies have shown that in non-phagocytic cells, Salmonella direct their own uptake byinducing dramatic membrane ruffles on the surface of thehost cell (Francis et al., 1993), a process that is distinctfrom receptor-mediated phagocytosis (Brumell et al.,1999). Induction of ruffling and bacterial invasion isabsolutely dependent upon a type III secretion system(TTSS) encoded by the Salmonella pathogenicity island-1 (SPI-1). A second TTSS, encoded on SPI-2, is inducedintracellularly (Cirillo et al., 1998; Pfeifer et al., 1999; Leeet al., 2000) and has been implicated in intracellular survival and replication in phagocytic cells (Ochman et al.,1996; Cirillo et al., 1998; Hensel et al., 1998).
Inside epithelial cells, Salmonella survive and replicatewithin a unique phagosome, the Salmonella-containingvacuole (SCV). Intracellular replication occurs after a lagof several hours and is coincident with the appearance ofdramatic tubular elements called Salmonella-induced fila-ments (Sifs), which protrude from the main body of theSCV (Garcia-del Portillo et al., 1993). SCV biogenesis isdependent on selective interactions with the endocyticpathway such that certain proteins characteristic of earlyendosomes, the recycling pathway and lysosomes areacquired sequentially (Garcia-del Portillo and Finlay,
Cellular Microbiology (2002) 4(1), 43–54
Olivia Steele-Mortimer,1* John H. Brumell,1
Leigh A. Knodler,1 Stephane Méresse,2 Ana Lopez1
and B. Brett Finlay1
1Biotechnology Laboratory, University of British Columbia, Vancouver BC V6T1Z3, Canada.2Centre d’Immunologie de Marseille-Luminy, INSERM-CNRS-Univ. Med., Campus de Luminy, Case 906, Marseille, France.
Summary
Type III secretion systems (TTSS) are used by Gram-negative pathogens to translocate proteins intoeukaryotic host cells. Salmonella enterica serovarTyphimurium (S. Typhimurium) has two of these spe-cialized systems, which are encoded on separate Sal-monella pathogenicity islands (SPI-1 and SPI-2) andtranslocate unique sets of effectors. The specificroles of these systems in Salmonella pathogenesisremain undefined, although SPI-1 is required for bacterial invasion of epithelial cells and SPI-2 for survival/replication in phagocytic cells. However,because SPI-1 TTSS mutants are invasion-incompetent, the role of this TTSS in post-invasionprocesses has not been investigated. In this study, wehave used two distinct methods to internalize a non-invasive SPI-1 TTSS mutant (invA) into culturedepithelial cells: (i) co-internalization with wild-type S.Typhimurium (SPI-1-dependent) and (ii) complemen-tation with the Yersinia pseudotuberculosis invasin(inv) gene (SPI-1-independent). In both cases, inter-nalized invA mutants were unable to replicate intra-cellularly, indicating that SPI-1 effectors are essentialfor this process and cannot be complemented bywild-type bacteria in the same cell. Analysis of thebiogenesis of SCVs showed that vacuoles containing
Received 5 January, 2001; revised 7 November, 2001; accepted 7 November, 2001. *For correspondence. E-mail [email protected]; Tel. (+1) 406 363 9292; Fax (+1) 406 363 9380.†Present address: Laboratory of Intracellular Parasites, NIAID, NIH,Rocky Mountain Laboratory, Hamilton, MT 59840, USA.

1995; Méresse et al., 1999a; Steele-Mortimer et al.,1999). Thus, biogenesis is characterized by the rapid lossof the early endocytic proteins, EEA1 and transferrinreceptor, and the concurrent acquisition of proteins asso-ciated with later stages of the endocytic pathway, such asthe vacuolar ATPase and lysosomal glycoproteins (lgp)(Garcia-del Portillo and Finlay, 1995; Méresse et al.,1999a; Steele-Mortimer et al., 1999). Delivery of lgp hasbeen shown to be dependent on the small GTP-bindingprotein rab7 and is not a result of direct fusion with preformed lysosomes (Garcia-del Portillo and Finlay,1995; Méresse et al., 1999a). In addition, the cation-independent mannose 6-phosphate receptor (CI-MPR) iseffectively excluded from the SCV (Garcia-del Portillo andFinlay, 1995; Méresse et al., 1999a; Steele-Mortimeret al., 1999). Hence, although SCV biogenesis has simi-larities with the endocytic and phagocytic pathways, it isclearly a distinct process. It appears that the intracellularbacteria are able to block specific membrane fusionevents thus determining the unique characteristics of theSCV.
The unique trafficking pattern of the SCV suggests amediatory role for bacterial effectors and indeed severalSPI-2 effectors have been implicated. The SPI-2-encodedSpiC prevents SCV fusion with lysosomes, apparently byinterfering with normal membrane trafficking in the hostcell (Uchiya et al., 1999). SifA, originally identified for itsrole in Sif formation, is suggested to promote fusion of the SCV with a novel endosomal compartment, and isrequired for maintenance of SCV membrane integrity(Beuzon et al., 2000; Brumell et al., 2001). Recently, twoother SPI-2-encoded putative effectors, SseF and SseG,have been shown to be required for Sif formation inepithelial cells (Guy et al., 2000). Although these findingsare consistent with a role for SPI-2 in modulating SCVmaturation, we have recently determined that mutants ofthis TTSS can replicate normally in some epithelial celllines (Brumell et al, 2001). This suggests that otherfactors, possibly SPI-1 effectors, are required for replica-tion in non-phagocytic cells. This idea is supported by twopublished findings: first, that some SPI-1 effectors may beinduced upon invasion of host cells (Pfeifer et al., 1999)and second, SopE (a SPI-1TTSS-translocated effector)interacts with the small GTP-binding protein rab5, to regu-late early endosome fusion (Mukherjee et al., 2000).
In this study, we have investigated the role of SPI-1 inintracellular survival/replication and SCV biogenesisusing a S. Typhimurium invA mutant, which is unable toinvade epithelial cells in vitro (Galan et al., 1992) and isattenuated for oral infection of mice (Galan and Curtiss,1989). InvA is a putative inner membrane component ofthe SPI-1TTSS apparatus. The mutant does not trans-locate SPI-1 effectors although they are synthesized(Collazo and Galan, 1997). We used two different mecha-
nisms to internalize the invA mutant into HeLa cells. Onemethod utilized an invA strain with a plasmid encoding the invasin (inv) gene of Yersinia pseudotuberculosis(Pace et al., 1993). Invasin-mediated uptake is a classicexample of the ‘zipper’ mechanism in which a bacterialsurface protein (Invasin) binds tightly to a receptor (b1-integrin) on the host cell surface (Isberg and Leong,1990). This binding leads to tight association of theplasma membrane with the bacteria and, eventually,engulfment in a close-fitting vacuole. In contrast, SPI-1-mediated invasion by Salmonella is characterized by dra-matic ruffles in the plasma membrane and uptake of thebacteria into a loose-fitting or spacious vacuole (Brumellet al., 1999). To internalize the invA mutant in a SPI-1-dependent mechanism, we took advantage of the abilityof Salmonella-induced ruffles to internalize non-invasivebacteria (Ginocchio et al., 1992) and co-internalized invAwith wild-type S. Typhimurium.
Our results show that the invA mutant internalized byeither method is replication incompetent, indicating thatSPI-1 is essential for intracellular replication. Further-more, effectors translocated by wild-type bacteria withinthe same host cell cannot rescue the invA mutant in trans.We also found that the biogenesis of vacuoles containingS. Typhimurium internalized via the Invasin-mediatedpathway were more similar to Y. pseudotuberculosisvacuoles than SCVs. This demonstrates that the methodof bacterial uptake into host cells has a determining rolein the biogenesis of bacteria-containing vacuoles.
Results
Internalization of S. Typhimurium SPI-1 mutants
To investigate whether there is a requirement for SPI-1effectors in intracellular survival of Salmonella, we useda well characterized invasion deficient SPI-1 mutant, invA.Two different techniques were used to promote inter-nalization of the invA mutant (Fig. 1A). The first methodinvolved transforming invA bacteria with a plasmid(pRI203) encoding the Invasin protein of Y. pseudotuber-culosis, which promotes bacterial uptake by binding to b1-integrin molecules on the host cell surface (Isberg and Leong, 1990). Alternatively, the invA mutant wasinternalized by SPI-1-dependent co-internalization withwild-type S. Typhimurium.
To assay bacterial internalization, we used a gentamicinresistance assay (Finlay and Falkow, 1988). HeLa cellswere infected for 10 min with wild-type or mutant bacteriaand then incubated in the presence of gentamicin to killextracellular bacteria. Intracellular (gentamicin-resistant)bacteria were then estimated by solubilization of cells, followed by plating of appropriate dilutions on toLuria–Bertani (LB) agar. The results demonstrate that
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54
44 O. Steele-Mortimer et al.
Intracellular proliferation of S. typhimurium requires SPI-1 45
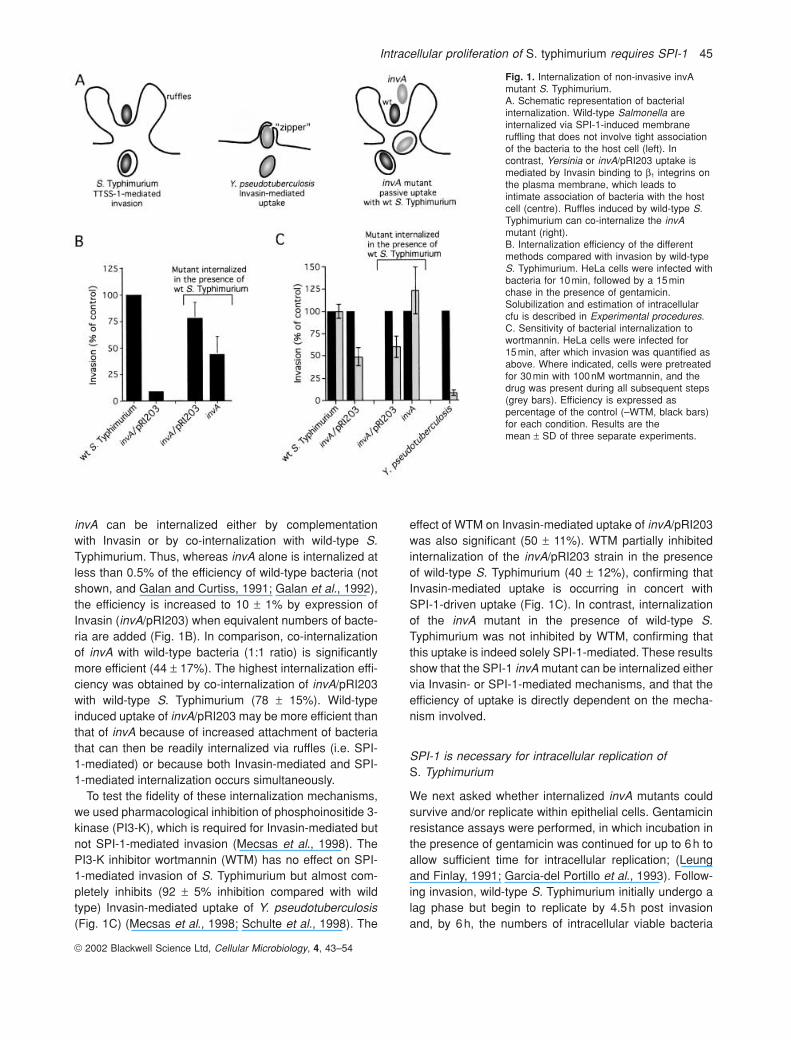
invA can be internalized either by complementation with Invasin or by co-internalization with wild-type S.Typhimurium. Thus, whereas invA alone is internalized atless than 0.5% of the efficiency of wild-type bacteria (notshown, and Galan and Curtiss, 1991; Galan et al., 1992),the efficiency is increased to 10 ± 1% by expression ofInvasin (invA/pRI203) when equivalent numbers of bacte-ria are added (Fig. 1B). In comparison, co-internalizationof invA with wild-type bacteria (1:1 ratio) is significantlymore efficient (44 ± 17%). The highest internalization effi-ciency was obtained by co-internalization of invA/pRI203with wild-type S. Typhimurium (78 ± 15%). Wild-typeinduced uptake of invA/pRI203 may be more efficient thanthat of invA because of increased attachment of bacteriathat can then be readily internalized via ruffles (i.e. SPI-1-mediated) or because both Invasin-mediated and SPI-1-mediated internalization occurs simultaneously.
To test the fidelity of these internalization mechanisms,we used pharmacological inhibition of phosphoinositide 3-kinase (PI3-K), which is required for Invasin-mediated butnot SPI-1-mediated invasion (Mecsas et al., 1998). ThePI3-K inhibitor wortmannin (WTM) has no effect on SPI-1-mediated invasion of S. Typhimurium but almost com-pletely inhibits (92 ± 5% inhibition compared with wildtype) Invasin-mediated uptake of Y. pseudotuberculosis(Fig. 1C) (Mecsas et al., 1998; Schulte et al., 1998). The
effect of WTM on Invasin-mediated uptake of invA/pRI203was also significant (50 ± 11%). WTM partially inhibitedinternalization of the invA/pRI203 strain in the presenceof wild-type S. Typhimurium (40 ± 12%), confirming thatInvasin-mediated uptake is occurring in concert with SPI-1-driven uptake (Fig. 1C). In contrast, internalizationof the invA mutant in the presence of wild-type S.Typhimurium was not inhibited by WTM, confirming thatthis uptake is indeed solely SPI-1-mediated. These resultsshow that the SPI-1 invA mutant can be internalized eithervia Invasin- or SPI-1-mediated mechanisms, and that theefficiency of uptake is directly dependent on the mecha-nism involved.
SPI-1 is necessary for intracellular replication of S. Typhimurium
We next asked whether internalized invA mutants couldsurvive and/or replicate within epithelial cells. Gentamicinresistance assays were performed, in which incubation inthe presence of gentamicin was continued for up to 6h toallow sufficient time for intracellular replication; (Leungand Finlay, 1991; Garcia-del Portillo et al., 1993). Follow-ing invasion, wild-type S. Typhimurium initially undergo alag phase but begin to replicate by 4.5h post invasionand, by 6h, the numbers of intracellular viable bacteria
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54
Fig. 1. Internalization of non-invasive invAmutant S. Typhimurium.A. Schematic representation of bacterialinternalization. Wild-type Salmonella areinternalized via SPI-1-induced membraneruffling that does not involve tight associationof the bacteria to the host cell (left). Incontrast, Yersinia or invA/pRI203 uptake ismediated by Invasin binding to b1 integrins onthe plasma membrane, which leads tointimate association of bacteria with the hostcell (centre). Ruffles induced by wild-type S.Typhimurium can co-internalize the invAmutant (right).B. Internalization efficiency of the differentmethods compared with invasion by wild-typeS. Typhimurium. HeLa cells were infected withbacteria for 10min, followed by a 15minchase in the presence of gentamicin.Solubilization and estimation of intracellularcfu is described in Experimental procedures.C. Sensitivity of bacterial internalization towortmannin. HeLa cells were infected for 15min, after which invasion was quantified asabove. Where indicated, cells were pretreatedfor 30min with 100nM wortmannin, and thedrug was present during all subsequent steps(grey bars). Efficiency is expressed aspercentage of the control (–WTM, black bars)for each condition. Results are themean ± SD of three separate experiments.
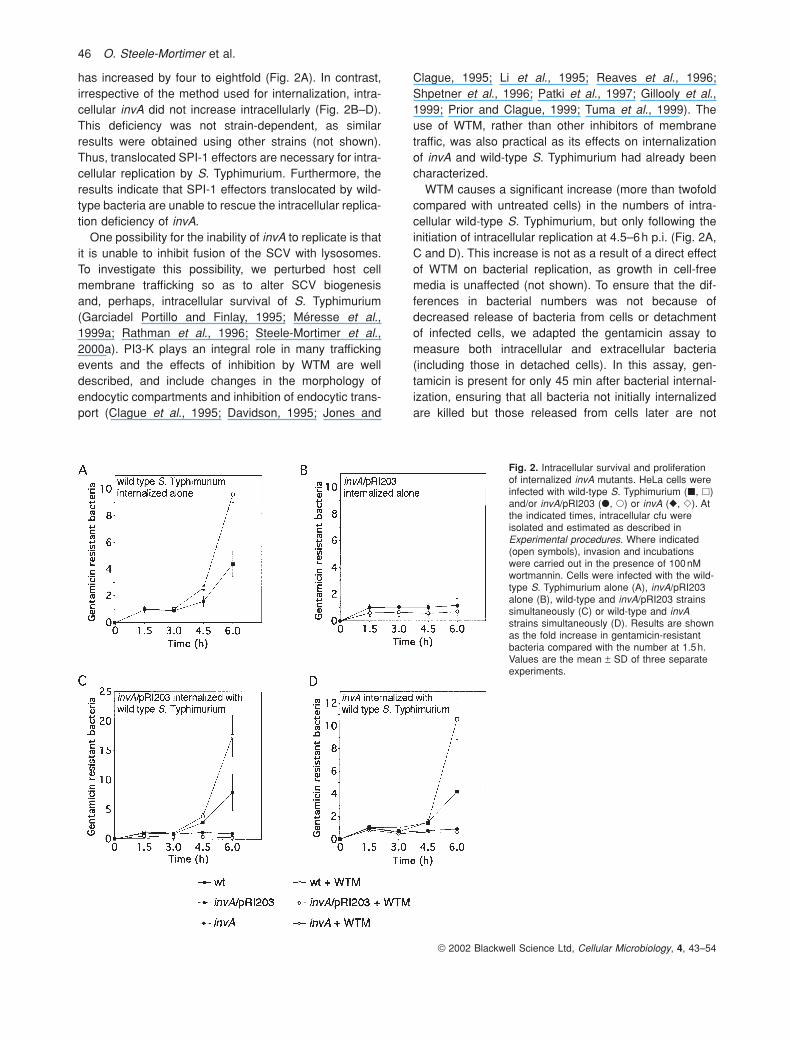
has increased by four to eightfold (Fig. 2A). In contrast,irrespective of the method used for internalization, intra-cellular invA did not increase intracellularly (Fig. 2B–D).This deficiency was not strain-dependent, as similarresults were obtained using other strains (not shown).Thus, translocated SPI-1 effectors are necessary for intra-cellular replication by S. Typhimurium. Furthermore, theresults indicate that SPI-1 effectors translocated by wild-type bacteria are unable to rescue the intracellular replica-tion deficiency of invA.
One possibility for the inability of invA to replicate is thatit is unable to inhibit fusion of the SCV with lysosomes.To investigate this possibility, we perturbed host cell membrane trafficking so as to alter SCV biogenesis and, perhaps, intracellular survival of S. Typhimurium(Garciadel Portillo and Finlay, 1995; Méresse et al.,1999a; Rathman et al., 1996; Steele-Mortimer et al.,2000a). PI3-K plays an integral role in many traffickingevents and the effects of inhibition by WTM are welldescribed, and include changes in the morphology ofendocytic compartments and inhibition of endocytic trans-port (Clague et al., 1995; Davidson, 1995; Jones and
Clague, 1995; Li et al., 1995; Reaves et al., 1996; Shpetner et al., 1996; Patki et al., 1997; Gillooly et al.,1999; Prior and Clague, 1999; Tuma et al., 1999). Theuse of WTM, rather than other inhibitors of membranetraffic, was also practical as its effects on internalizationof invA and wild-type S. Typhimurium had already beencharacterized.
WTM causes a significant increase (more than twofoldcompared with untreated cells) in the numbers of intra-cellular wild-type S. Typhimurium, but only following theinitiation of intracellular replication at 4.5–6h p.i. (Fig. 2A,C and D). This increase is not as a result of a direct effectof WTM on bacterial replication, as growth in cell-freemedia is unaffected (not shown). To ensure that the dif-ferences in bacterial numbers was not because ofdecreased release of bacteria from cells or detachment of infected cells, we adapted the gentamicin assay tomeasure both intracellular and extracellular bacteria(including those in detached cells). In this assay, gen-tamicin is present for only 45 min after bacterial internal-ization, ensuring that all bacteria not initially internalizedare killed but those released from cells later are not
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54
46 O. Steele-Mortimer et al.
Fig. 2. Intracellular survival and proliferationof internalized invA mutants. HeLa cells wereinfected with wild-type S. Typhimurium (�, �)and/or invA/pRI203 (�, �) or invA (�, �). Atthe indicated times, intracellular cfu wereisolated and estimated as described inExperimental procedures. Where indicated(open symbols), invasion and incubationswere carried out in the presence of 100nMwortmannin. Cells were infected with the wild-type S. Typhimurium alone (A), invA/pRI203alone (B), wild-type and invA/pRI203 strainssimultaneously (C) or wild-type and invAstrains simultaneously (D). Results are shownas the fold increase in gentamicin-resistantbacteria compared with the number at 1.5h.Values are the mean ± SD of three separateexperiments.
Intracellular proliferation of S. typhimurium requires SPI-1 47
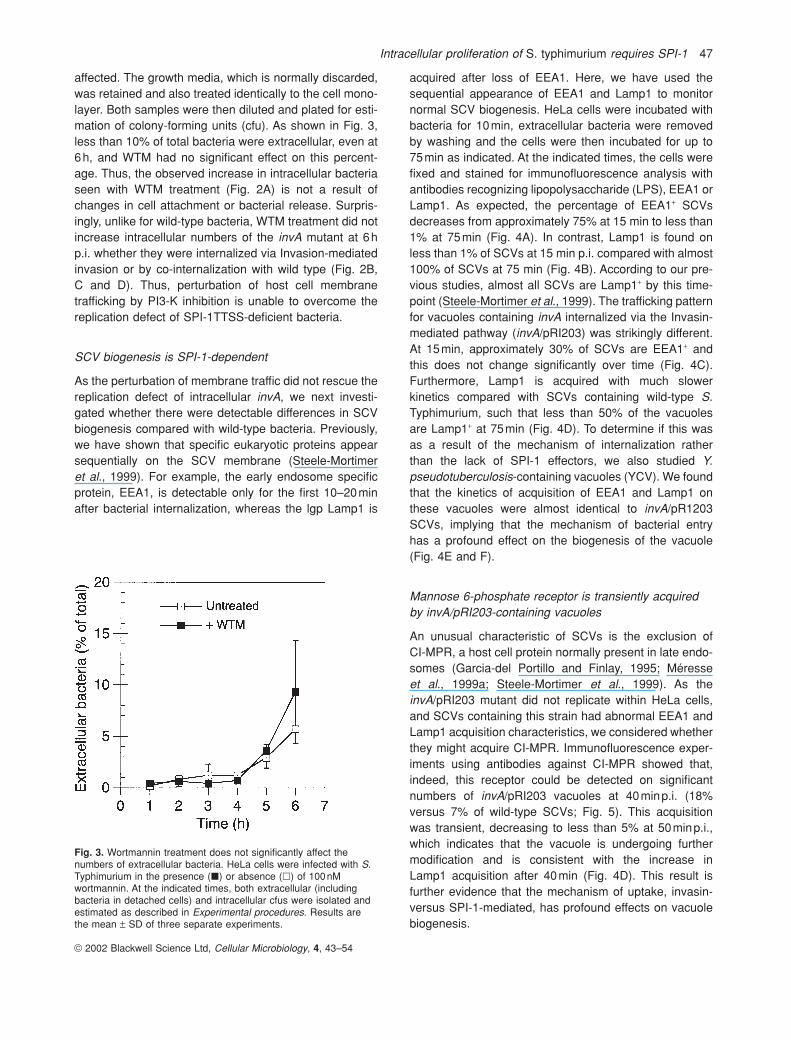
affected. The growth media, which is normally discarded,was retained and also treated identically to the cell mono-layer. Both samples were then diluted and plated for esti-mation of colony-forming units (cfu). As shown in Fig. 3,less than 10% of total bacteria were extracellular, even at6h, and WTM had no significant effect on this percent-age. Thus, the observed increase in intracellular bacteriaseen with WTM treatment (Fig. 2A) is not a result ofchanges in cell attachment or bacterial release. Surpris-ingly, unlike for wild-type bacteria, WTM treatment did notincrease intracellular numbers of the invA mutant at 6hp.i. whether they were internalized via Invasion-mediatedinvasion or by co-internalization with wild type (Fig. 2B, C and D). Thus, perturbation of host cell membrane trafficking by PI3-K inhibition is unable to overcome thereplication defect of SPI-1TTSS-deficient bacteria.
SCV biogenesis is SPI-1-dependent
As the perturbation of membrane traffic did not rescue thereplication defect of intracellular invA, we next investi-gated whether there were detectable differences in SCVbiogenesis compared with wild-type bacteria. Previously,we have shown that specific eukaryotic proteins appearsequentially on the SCV membrane (Steele-Mortimeret al., 1999). For example, the early endosome specificprotein, EEA1, is detectable only for the first 10–20minafter bacterial internalization, whereas the lgp Lamp1 is
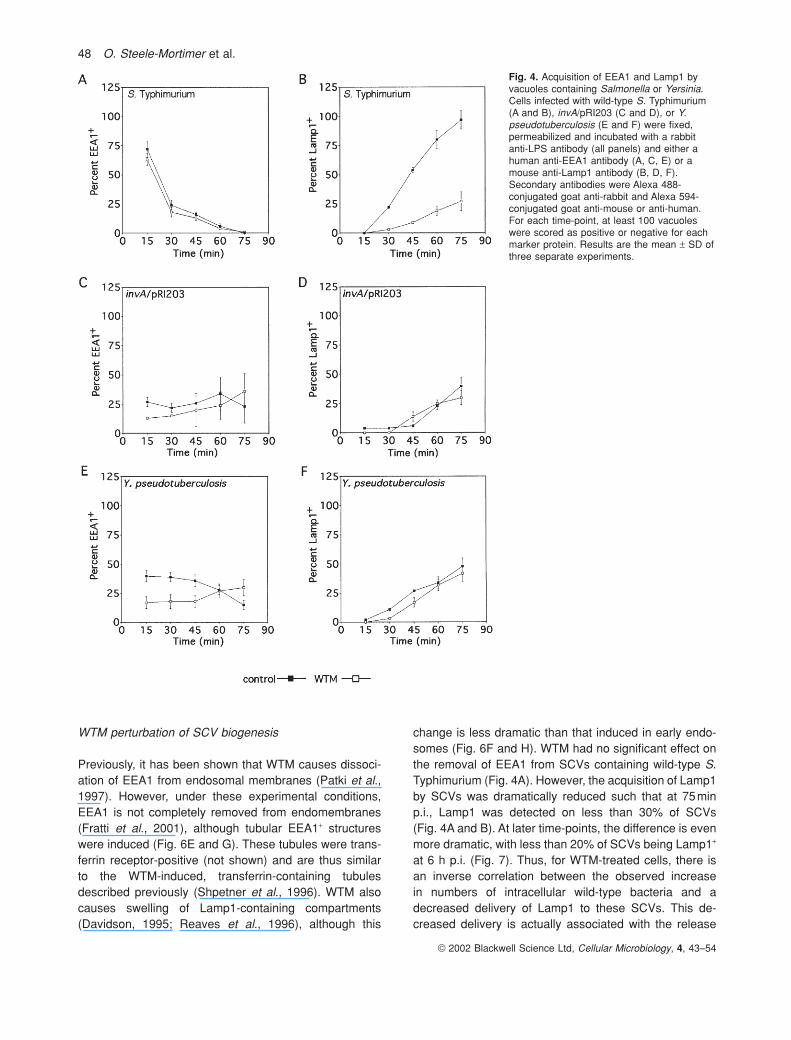
acquired after loss of EEA1. Here, we have used thesequential appearance of EEA1 and Lamp1 to monitornormal SCV biogenesis. HeLa cells were incubated withbacteria for 10min, extracellular bacteria were removedby washing and the cells were then incubated for up to75min as indicated. At the indicated times, the cells werefixed and stained for immunofluorescence analysis withantibodies recognizing lipopolysaccharide (LPS), EEA1 orLamp1. As expected, the percentage of EEA1+ SCVsdecreases from approximately 75% at 15 min to less than1% at 75min (Fig. 4A). In contrast, Lamp1 is found onless than 1% of SCVs at 15 min p.i. compared with almost100% of SCVs at 75 min (Fig. 4B). According to our pre-vious studies, almost all SCVs are Lamp1+ by this time-point (Steele-Mortimer et al., 1999). The trafficking patternfor vacuoles containing invA internalized via the Invasin-mediated pathway (invA/pRI203) was strikingly different.At 15min, approximately 30% of SCVs are EEA1+ and this does not change significantly over time (Fig. 4C). Furthermore, Lamp1 is acquired with much slower kinetics compared with SCVs containing wild-type S.Typhimurium, such that less than 50% of the vacuoles are Lamp1+ at 75min (Fig. 4D). To determine if this wasas a result of the mechanism of internalization rather than the lack of SPI-1 effectors, we also studied Y.pseudotuberculosis-containing vacuoles (YCV). We foundthat the kinetics of acquisition of EEA1 and Lamp1 onthese vacuoles were almost identical to invA/pR1203SCVs, implying that the mechanism of bacterial entry has a profound effect on the biogenesis of the vacuole(Fig. 4E and F).
Mannose 6-phosphate receptor is transiently acquiredby invA/pRI203-containing vacuoles
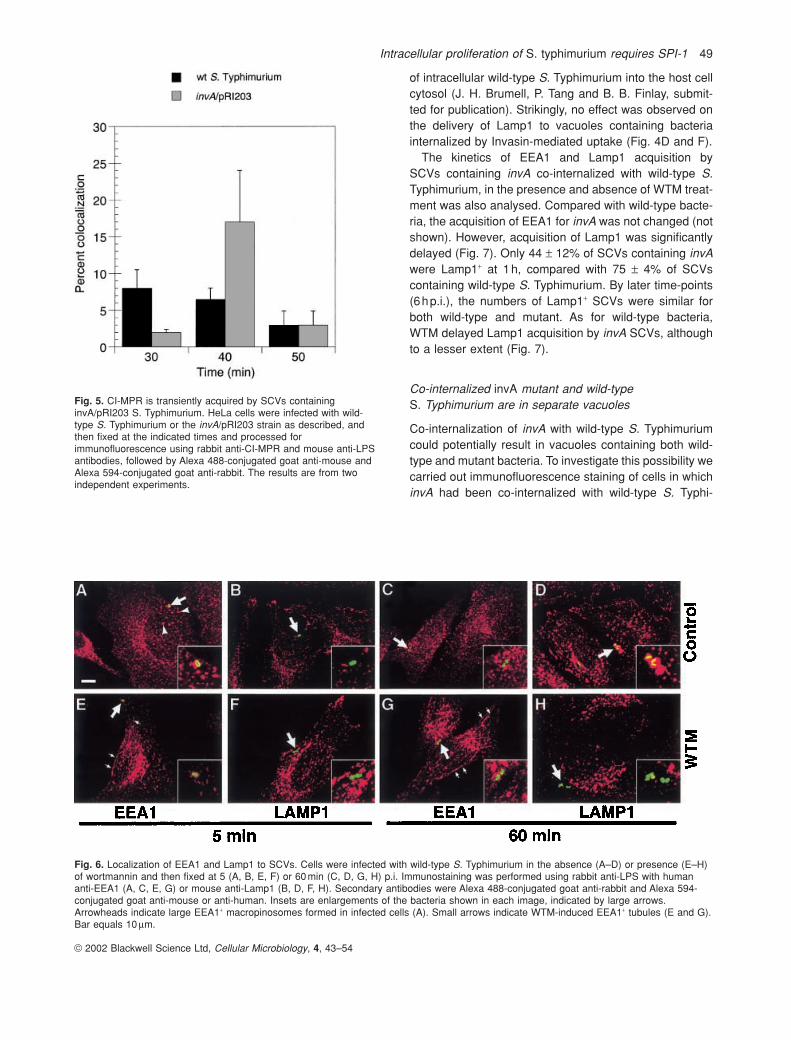
An unusual characteristic of SCVs is the exclusion of CI-MPR, a host cell protein normally present in late endo-somes (Garcia-del Portillo and Finlay, 1995; Méresseet al., 1999a; Steele-Mortimer et al., 1999). As theinvA/pRI203 mutant did not replicate within HeLa cells,and SCVs containing this strain had abnormal EEA1 andLamp1 acquisition characteristics, we considered whetherthey might acquire CI-MPR. Immunofluorescence exper-iments using antibodies against CI-MPR showed that,indeed, this receptor could be detected on significantnumbers of invA/pRI203 vacuoles at 40minp.i. (18%versus 7% of wild-type SCVs; Fig. 5). This acquisition was transient, decreasing to less than 5% at 50minp.i.,which indicates that the vacuole is undergoing furthermodification and is consistent with the increase in Lamp1 acquisition after 40min (Fig. 4D). This result isfurther evidence that the mechanism of uptake, invasin-versus SPI-1-mediated, has profound effects on vacuolebiogenesis.
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54
Fig. 3. Wortmannin treatment does not significantly affect thenumbers of extracellular bacteria. HeLa cells were infected with S.Typhimurium in the presence (�) or absence (�) of 100nMwortmannin. At the indicated times, both extracellular (includingbacteria in detached cells) and intracellular cfus were isolated andestimated as described in Experimental procedures. Results arethe mean ± SD of three separate experiments.
WTM perturbation of SCV biogenesis
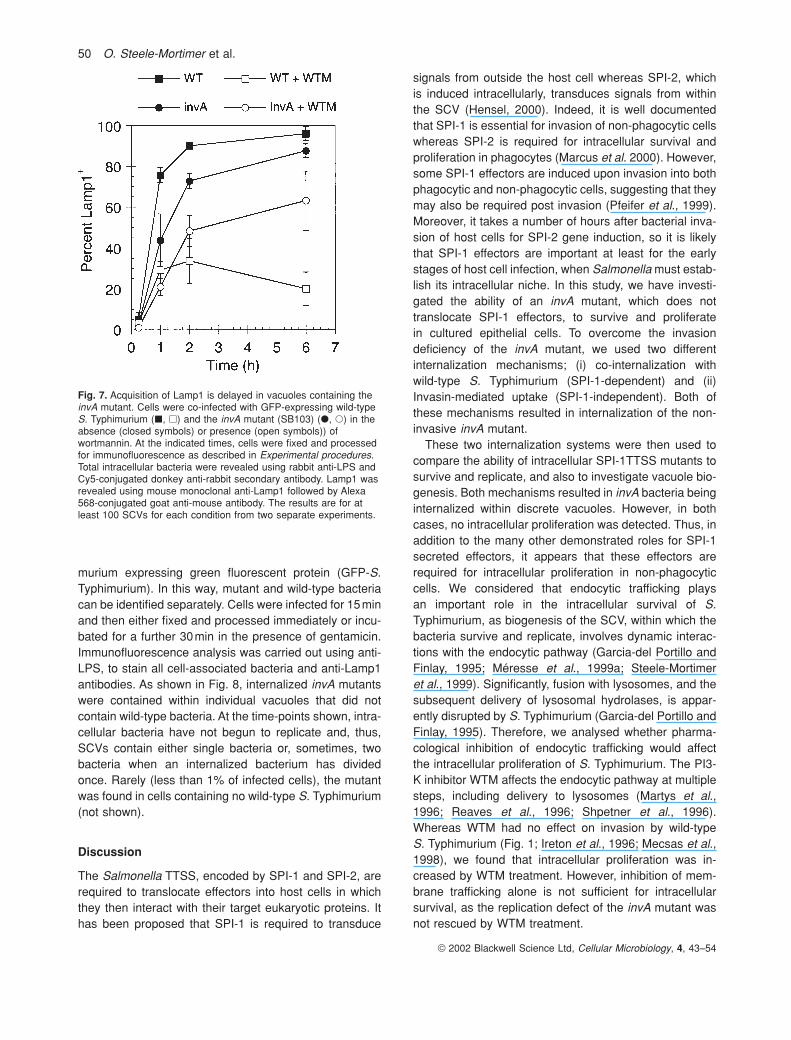
Previously, it has been shown that WTM causes dissoci-ation of EEA1 from endosomal membranes (Patki et al.,1997). However, under these experimental conditions,EEA1 is not completely removed from endomembranes(Fratti et al., 2001), although tubular EEA1+ structureswere induced (Fig. 6E and G). These tubules were trans-ferrin receptor-positive (not shown) and are thus similarto the WTM-induced, transferrin-containing tubulesdescribed previously (Shpetner et al., 1996). WTM alsocauses swelling of Lamp1-containing compartments(Davidson, 1995; Reaves et al., 1996), although this
change is less dramatic than that induced in early endo-somes (Fig. 6F and H). WTM had no significant effect onthe removal of EEA1 from SCVs containing wild-type S.Typhimurium (Fig. 4A). However, the acquisition of Lamp1by SCVs was dramatically reduced such that at 75minp.i., Lamp1 was detected on less than 30% of SCVs(Fig. 4A and B). At later time-points, the difference is evenmore dramatic, with less than 20% of SCVs being Lamp1+
at 6 h p.i. (Fig. 7). Thus, for WTM-treated cells, there isan inverse correlation between the observed increase in numbers of intracellular wild-type bacteria and adecreased delivery of Lamp1 to these SCVs. This de-creased delivery is actually associated with the release
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54
48 O. Steele-Mortimer et al.
Fig. 4. Acquisition of EEA1 and Lamp1 byvacuoles containing Salmonella or Yersinia.Cells infected with wild-type S. Typhimurium(A and B), invA/pRI203 (C and D), or Y.pseudotuberculosis (E and F) were fixed,permeabilized and incubated with a rabbitanti-LPS antibody (all panels) and either ahuman anti-EEA1 antibody (A, C, E) or amouse anti-Lamp1 antibody (B, D, F).Secondary antibodies were Alexa 488-conjugated goat anti-rabbit and Alexa 594-conjugated goat anti-mouse or anti-human.For each time-point, at least 100 vacuoleswere scored as positive or negative for eachmarker protein. Results are the mean ± SD ofthree separate experiments.
Intracellular proliferation of S. typhimurium requires SPI-1 49
of intracellular wild-type S. Typhimurium into the host cellcytosol (J. H. Brumell, P. Tang and B. B. Finlay, submit-ted for publication). Strikingly, no effect was observed onthe delivery of Lamp1 to vacuoles containing bacteriainternalized by Invasin-mediated uptake (Fig. 4D and F).
The kinetics of EEA1 and Lamp1 acquisition by SCVs containing invA co-internalized with wild-type S.Typhimurium, in the presence and absence of WTM treat-ment was also analysed. Compared with wild-type bacte-ria, the acquisition of EEA1 for invA was not changed (notshown). However, acquisition of Lamp1 was significantlydelayed (Fig. 7). Only 44 ± 12% of SCVs containing invAwere Lamp1+ at 1h, compared with 75 ± 4% of SCVs containing wild-type S. Typhimurium. By later time-points(6hp.i.), the numbers of Lamp1+ SCVs were similar forboth wild-type and mutant. As for wild-type bacteria, WTM delayed Lamp1 acquisition by invA SCVs, althoughto a lesser extent (Fig. 7).
Co-internalized invA mutant and wild-type S. Typhimurium are in separate vacuoles
Co-internalization of invA with wild-type S. Typhimuriumcould potentially result in vacuoles containing both wild-type and mutant bacteria. To investigate this possibility wecarried out immunofluorescence staining of cells in whichinvA had been co-internalized with wild-type S. Typhi-
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54
Fig. 5. CI-MPR is transiently acquired by SCVs containinginvA/pRI203 S. Typhimurium. HeLa cells were infected with wild-type S. Typhimurium or the invA/pRI203 strain as described, andthen fixed at the indicated times and processed forimmunofluorescence using rabbit anti-CI-MPR and mouse anti-LPSantibodies, followed by Alexa 488-conjugated goat anti-mouse andAlexa 594-conjugated goat anti-rabbit. The results are from twoindependent experiments.
Fig. 6. Localization of EEA1 and Lamp1 to SCVs. Cells were infected with wild-type S. Typhimurium in the absence (A–D) or presence (E–H)of wortmannin and then fixed at 5 (A, B, E, F) or 60 min (C, D, G, H) p.i. Immunostaining was performed using rabbit anti-LPS with humananti-EEA1 (A, C, E, G) or mouse anti-Lamp1 (B, D, F, H). Secondary antibodies were Alexa 488-conjugated goat anti-rabbit and Alexa 594-conjugated goat anti-mouse or anti-human. Insets are enlargements of the bacteria shown in each image, indicated by large arrows.Arrowheads indicate large EEA1+ macropinosomes formed in infected cells (A). Small arrows indicate WTM-induced EEA1+ tubules (E and G).Bar equals 10 mm.
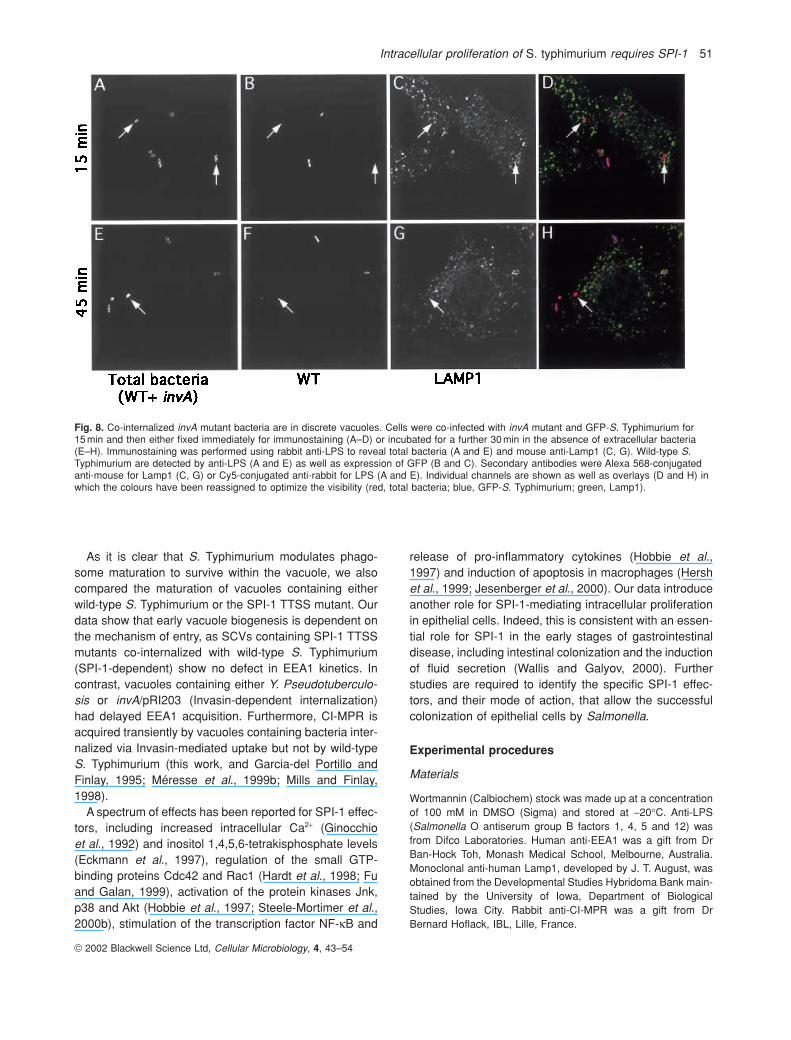
murium expressing green fluorescent protein (GFP-S.Typhimurium). In this way, mutant and wild-type bacteriacan be identified separately. Cells were infected for 15minand then either fixed and processed immediately or incu-bated for a further 30min in the presence of gentamicin.Immunofluorescence analysis was carried out using anti-LPS, to stain all cell-associated bacteria and anti-Lamp1antibodies. As shown in Fig. 8, internalized invA mutantswere contained within individual vacuoles that did notcontain wild-type bacteria. At the time-points shown, intra-cellular bacteria have not begun to replicate and, thus,SCVs contain either single bacteria or, sometimes, twobacteria when an internalized bacterium has dividedonce. Rarely (less than 1% of infected cells), the mutantwas found in cells containing no wild-type S. Typhimurium(not shown).
Discussion
The Salmonella TTSS, encoded by SPI-1 and SPI-2, arerequired to translocate effectors into host cells in whichthey then interact with their target eukaryotic proteins. Ithas been proposed that SPI-1 is required to transduce
signals from outside the host cell whereas SPI-2, whichis induced intracellularly, transduces signals from withinthe SCV (Hensel, 2000). Indeed, it is well documentedthat SPI-1 is essential for invasion of non-phagocytic cellswhereas SPI-2 is required for intracellular survival andproliferation in phagocytes (Marcus et al. 2000). However,some SPI-1 effectors are induced upon invasion into bothphagocytic and non-phagocytic cells, suggesting that theymay also be required post invasion (Pfeifer et al., 1999).Moreover, it takes a number of hours after bacterial inva-sion of host cells for SPI-2 gene induction, so it is likelythat SPI-1 effectors are important at least for the earlystages of host cell infection, when Salmonella must estab-lish its intracellular niche. In this study, we have investi-gated the ability of an invA mutant, which does nottranslocate SPI-1 effectors, to survive and proliferate in cultured epithelial cells. To overcome the invasion deficiency of the invA mutant, we used two different internalization mechanisms; (i) co-internalization withwild-type S. Typhimurium (SPI-1-dependent) and (ii)Invasin-mediated uptake (SPI-1-independent). Both ofthese mechanisms resulted in internalization of the non-invasive invA mutant.
These two internalization systems were then used tocompare the ability of intracellular SPI-1TTSS mutants tosurvive and replicate, and also to investigate vacuole bio-genesis. Both mechanisms resulted in invA bacteria beinginternalized within discrete vacuoles. However, in bothcases, no intracellular proliferation was detected. Thus, inaddition to the many other demonstrated roles for SPI-1secreted effectors, it appears that these effectors arerequired for intracellular proliferation in non-phagocyticcells. We considered that endocytic trafficking plays an important role in the intracellular survival of S.Typhimurium, as biogenesis of the SCV, within which thebacteria survive and replicate, involves dynamic interac-tions with the endocytic pathway (Garcia-del Portillo andFinlay, 1995; Méresse et al., 1999a; Steele-Mortimeret al., 1999). Significantly, fusion with lysosomes, and thesubsequent delivery of lysosomal hydrolases, is appar-ently disrupted by S. Typhimurium (Garcia-del Portillo andFinlay, 1995). Therefore, we analysed whether pharma-cological inhibition of endocytic trafficking would affect the intracellular proliferation of S. Typhimurium. The PI3-K inhibitor WTM affects the endocytic pathway at multiplesteps, including delivery to lysosomes (Martys et al.,1996; Reaves et al., 1996; Shpetner et al., 1996).Whereas WTM had no effect on invasion by wild-type S. Typhimurium (Fig. 1; Ireton et al., 1996; Mecsas et al.,1998), we found that intracellular proliferation was in-creased by WTM treatment. However, inhibition of mem-brane trafficking alone is not sufficient for intracellularsurvival, as the replication defect of the invA mutant wasnot rescued by WTM treatment.
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54
50 O. Steele-Mortimer et al.
Fig. 7. Acquisition of Lamp1 is delayed in vacuoles containing theinvA mutant. Cells were co-infected with GFP-expressing wild-typeS. Typhimurium (�, �) and the invA mutant (SB103) (�, �) in theabsence (closed symbols) or presence (open symbols)) ofwortmannin. At the indicated times, cells were fixed and processedfor immunofluorescence as described in Experimental procedures.Total intracellular bacteria were revealed using rabbit anti-LPS andCy5-conjugated donkey anti-rabbit secondary antibody. Lamp1 wasrevealed using mouse monoclonal anti-Lamp1 followed by Alexa568-conjugated goat anti-mouse antibody. The results are for atleast 100 SCVs for each condition from two separate experiments.
Intracellular proliferation of S. typhimurium requires SPI-1 51
As it is clear that S. Typhimurium modulates phago-some maturation to survive within the vacuole, we alsocompared the maturation of vacuoles containing eitherwild-type S. Typhimurium or the SPI-1 TTSS mutant. Ourdata show that early vacuole biogenesis is dependent onthe mechanism of entry, as SCVs containing SPI-1 TTSSmutants co-internalized with wild-type S. Typhimurium(SPI-1-dependent) show no defect in EEA1 kinetics. Incontrast, vacuoles containing either Y. Pseudotuberculo-sis or invA/pRI203 (Invasin-dependent internalization)had delayed EEA1 acquisition. Furthermore, CI-MPR isacquired transiently by vacuoles containing bacteria inter-nalized via Invasin-mediated uptake but not by wild-typeS. Typhimurium (this work, and Garcia-del Portillo andFinlay, 1995; Méresse et al., 1999b; Mills and Finlay,1998).
A spectrum of effects has been reported for SPI-1 effec-tors, including increased intracellular Ca2+ (Ginocchioet al., 1992) and inositol 1,4,5,6-tetrakisphosphate levels(Eckmann et al., 1997), regulation of the small GTP-binding proteins Cdc42 and Rac1 (Hardt et al., 1998; Fuand Galan, 1999), activation of the protein kinases Jnk,p38 and Akt (Hobbie et al., 1997; Steele-Mortimer et al.,2000b), stimulation of the transcription factor NF-kB and
release of pro-inflammatory cytokines (Hobbie et al.,1997) and induction of apoptosis in macrophages (Hershet al., 1999; Jesenberger et al., 2000). Our data introduceanother role for SPI-1-mediating intracellular proliferationin epithelial cells. Indeed, this is consistent with an essen-tial role for SPI-1 in the early stages of gastrointestinaldisease, including intestinal colonization and the inductionof fluid secretion (Wallis and Galyov, 2000). Furtherstudies are required to identify the specific SPI-1 effec-tors, and their mode of action, that allow the successfulcolonization of epithelial cells by Salmonella.
Experimental procedures
Materials
Wortmannin (Calbiochem) stock was made up at a concentrationof 100 mM in DMSO (Sigma) and stored at -20∞C. Anti-LPS (Salmonella O antiserum group B factors 1, 4, 5 and 12) wasfrom Difco Laboratories. Human anti-EEA1 was a gift from DrBan-Hock Toh, Monash Medical School, Melbourne, Australia.Monoclonal anti-human Lamp1, developed by J. T. August, wasobtained from the Developmental Studies Hybridoma Bank main-tained by the University of Iowa, Department of BiologicalStudies, Iowa City. Rabbit anti-CI-MPR was a gift from DrBernard Hoflack, IBL, Lille, France.
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54
Fig. 8. Co-internalized invA mutant bacteria are in discrete vacuoles. Cells were co-infected with invA mutant and GFP-S. Typhimurium for 15 min and then either fixed immediately for immunostaining (A–D) or incubated for a further 30 min in the absence of extracellular bacteria(E–H). Immunostaining was performed using rabbit anti-LPS to reveal total bacteria (A and E) and mouse anti-Lamp1 (C, G). Wild-type S.Typhimurium are detected by anti-LPS (A and E) as well as expression of GFP (B and C). Secondary antibodies were Alexa 568-conjugatedanti-mouse for Lamp1 (C, G) or Cy5-conjugated anti-rabbit for LPS (A and E). Individual channels are shown as well as overlays (D and H) inwhich the colours have been reassigned to optimize the visibility (red, total bacteria; blue, GFP-S. Typhimurium; green, Lamp1).

Cell culture and bacterial strains
HeLa (human cervical adenocarcinoma cells; ATCC CCL2) werecultured in growth media (GM), consisting of Eagle’s Minimalessential medium (MEM) (Gibco BRL) supplemented with 10%heat-inactivated fetal calf serum (FCS) (Gibco BRL), at 37∞C in5% CO2. Cells were passaged no more than 20 times.
Salmonella Typhimurium SL1344, invA (SB111), invA (SB103)and invA/pRI203, and Y. pseudotuberculosis YIII (plasmid cured)strains have been described previously (Hoiseth and Stocker,1981; Pace et al., 1993; Garcia-del Portillo et al., 1994; Steele-Mortimer et al., 2000b). Bacteria were grown in Luria–Bertani(LB) broth or on LB agar plates. When appropriate, kanamy-cin (50mgml-1) or ampicillin (100mgml-1) was added. GFP-expressing bacteria were as described (Méresse et al., 1999a;2001).
Gentamicin protection assays
Invasion assays were carried out essentially as described previ-ously (Steele-Mortimer et al., 1999). Where indicated, cells weretreated with WTM (100nM) for 30min before the addition of bacteria and continued throughout the experiment. Briefly, S.Typhimurium were grown in LB broth overnight at 37∞C withshaking, and then subcultured (300ml in 10ml of LB) for a further3h. The culture was centrifuged at 10000 g for 2min at roomtemperature, and then directly resuspended in phosphate-buffered saline (PBS). Yersinia pseudotuberculosis were grownovernight in 2ml of brain–heart infusion (BHI) media at 26∞C. Thissuspension was diluted 1:100 in growth medium and used forinvasion. Invasion was initiated by the addition of diluted bacte-ria directly to subconfluent HeLa cells grown in 24-well plates,and proceeded at 37∞C, in 5% CO2, for 5 or 10min Extracellularbacteria were then removed by washing with PBS and the cellswere incubated in GM. In experiments requiring longer infectiontimes, GM was supplemented with 50mgml-1 of gentamicin(Sigma), at 20min post infection, to kill extracellular bacteria. Atthe indicated times, monolayers were washed twice with PBSand solubilized in 1ml of 1% Triton X-100, 0.1% SDS in PBS for5 min at room temperature. The solute was serially diluted in PBSand plated on to duplicate LB agar plates. LB plates were sup-plemented with 25mgml-1 of kanamycin for the invA mutant S.Typhimurium, and with 25mgml-1 of kanamycin and 25mgml-1
chloramphenicol for invA/pRI203. Plates were incubated at 37∞Cand the colonies were counted the next day.
To assay extracellular bacteria, gentamicin was present for thefirst 45min only. Thereafter, the monolayers were incubated inantibiotic free media. At the indicated times the supernatant washarvested and brought to 1% Triton X-100, 0.1% SDS by theaddition of 10¥ stock solution. Incubation and plating was thencarried out as above.
Immunofluorescence
HeLa cells on glass coverslips were infected with bacteria, fixedand processed for immunofluorescence as described previously(Steele-Mortimer et al., 1999). Fixation was in 2.5% formalde-hyde followed by permeabilization in PBS containing 10% (v/v)normal goat serum (Gibco BRL) and 0.1% (w/v) Saponin (SS-PBS) for 10min. For CI-MPR labelling, an additional extraction
with 0.1% Triton X-100 for 4 min was performed. Primary anti-bodies were diluted in SS-PBS as follows; anti-LPS at 1:300,anti-CI-MPR at 1:250, anti-EEA at 1:1000 and anti-Lamp1 at1:100. Secondary antibodies, Cy5-conjugated donkey anti-rabbitIgG, Alexa 488-conjugated goat anti-rabbit IgG, Alexa 594-conjugated goat anti-mouse or goat anti-human IgG were diluted1:600 in SS-PBS. After each antibody incubation (30min each),the monolayers were washed extensively with PBS containing0.05% Saponin. Coverslips were mounted on to glass slidesusing Mowiol (Calbiochem). Cells were examined using a ZeissAxiophot microscope using a 63¥ oil immersion objective. Foreach time-point, at least 50 intracellular bacteria were scored forco-localization with Lamp1, EEA1 or CI-MPR. Micrographs weretaken using TMAX 400 film (Eastman Kodak) at ASA 1600.Images were processed using ADOBE PHOTOSHOP.
Acknowledgements
O.S.-M. was the recipient of a postdoctoral fellowship from theEuropean Molecular Biology Organization (EMBO). J.H.B. is sup-ported by a Medical Research Council of Canada postdoctoralfellowship and is an honorary fellow of the Izaac Walter KillamMemorial foundation. This work was supported by operatinggrants from the Medical Research Council of Canada (B.B.F) anda Howard Hughes International Research Scholar Award(B.B.F.). B.B.F. is a CIHR Distinguished Investigator.
References
Beuzon, C.R., Méresse, S., Unsworth, K.E., Ruiz-Albert, J.,Garvis, S., Waterman, S.R., et al. (2000) Salmonella maintainsthe integrity of its intracellular vacuole through the action ofSifA. EMBO J 19: 3235–3249.
Brumell, J.H., Steele-Mortimer, O., and Finlay, B.B. (1999) Bac-terial invasion: force feeding by Salmonella. Curr Biol 9:277–280.
Brumell, J.H., Rosenberger, C.M., Gotto, G.T., Marcus, S.L., andFinlay, B.B. (2001) SifA permits survival and replication of Sal-monella typhimurium in murine macrophages. Cell Microbiol 3:75–84.
Cirillo, D.M., Valdivia, R.H., Monack, D.M., and Falkow, S. (1998)Macrophage-dependent induction of the Salmonella patho-genicity island 2 type III secretion system and its role in intra-cellular survival. Mol Microbiol 30: 175–188.
Clague, M.J., Thorpe, C., and Jones, A.T. (1995) Phosphatidyli-nositol 3-kinase regulation of fluid phase endocytosis. FEBSLett 367: 272–274.
Clark, M.A., Hirst, B.H., and Jepson, M.A. (1998) Inoculum com-position and Salmonella pathogenicity island 1 regulate M-cellinvasion and epithelial destruction by Salmonella typhimurium.Infect Immun 66: 724–731.
Collazo, C.M., and Galan, J.E. (1997) The invasion-associatedtype III system of Salmonella typhimurium directs the trans-location of Sip proteins into the host cell. Mol Microbiol 24:747–756.
Davidson, H.W. (1995) Wortmannin causes mistargeting of pro-cathepsin D. evidence for the involvement of a phosphatidyli-nositol 3-kinase in vesicular transport to lysosomes. J Cell Biol130: 797–805.
Eckmann, L., Rudolf, M.T., Ptasznik, A., Schultz, C., Jiang, T.,Wolfson, N., et al. (1997) D-myo-Inositol 1,4,5,6-tetrakisphos-
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54
52 O. Steele-Mortimer et al.

Intracellular proliferation of S. typhimurium requires SPI-1 53
phate produced in human intestinal epithelial cells in responseto Salmonella invasion inhibits phosphoinositide 3-kinase sig-naling pathways. Proc Natl Acad Sci USA 94: 14456–14460.
Finlay, B.B., and Falkow, S. (1988) Comparison of the invasionstrategies used by Salmonella cholerae-suis, Shigella flexneriand Yersinia enterocolitica to enter cultured animal cells: endo-some acidification is not required for bacterial invasion or intra-cellular replication. Biochimie 70: 1089–1099.
Francis, C.L., Ryan, T.A., Jones, B.D., Smith, S.J., and Falkow,S. (1993) Ruffles induced by Salmonella and other stimulidirect macropinocytosis of bacteria. Nature 364: 639–642.
Fratti, R.A., Backer, J.M., Gruenberg, J., Corvera, S., andDeretic, V. (2001) Role of phosphatidylinositol 3-kinase andRab5 effectors in phagosomal biogenesis and mycobacterialphagosome maturation arrest. J Cell Biol 154: 631–644.
Fu, Y., and Galan, J.E. (1999) A Salmonella protein antagonizesRac-1 and Cdc42 to mediate host-cell recovery after bacterialinvasion. Nature 401: 293–297.
Galan, J.E., and Curtiss, R., III. (1989) Cloning and molecularcharacterization of genes whose products allow Salmonellatyphimurium to penetrate tissue culture cells. Proc Natl AcadSci USA 86: 6383–6387.
Galan, J.E., Curtiss, R., III. (1991) Distribution of the invA, -B, -C and -D genes of Salmonella typhimurium among other Salmonella serovars: invA mutants of Salmonella typhi aredeficient for entry into mammalian cells. Infect Immun 59:2901–2908.
Galan, J.E., Ginocchio, C., and Costeas, P. (1992) Molecular andfunctional characterization of the Salmonella invasion geneinvA: homology of InvA to members of a new protein family. J Bacteriol 174: 4338–4349.
Garcia-del Portillo, F., Zwick, M.B., Leung, K.Y., and Finlay, B.B.(1993) Salmonella induces the formation of filamentous struc-tures containing lysosomal membrane glycoproteins in epithe-lial cells. Proc Natl Acad Sci USA 90: 10544–10548.
Garcia-del Portillo, F., Pucciarelli, M.G., Jefferies, W.A., andFinlay, B.B. (1994) Salmonella typhimurium induces selectiveaggregation and internalization of host cell surface proteinsduring invasion of epithelial cells. J Cell Sci 107: 2005–2020.
Garcia-del Portillo, F., and Finlay, B.B. (1995) Targeting of Sal-monella typhimurium to vesicles containing lysosomal mem-brane glycoproteins bypasses compartments with mannose6-phosphate receptors. J Cell Biol 129: 81–97.
Gillooly, D.J., Melendez, A.J., Hockaday, A.R., Harnett, M.M., and Allen, J.M. (1999) Endocytosis and vesicular trafficking of immune complexes and activation of phospholipase D by the human high-affinity IgG receptor requires distinct phosphoinositide 3-kinase activities. Biochem J 344 Part 2:605–611.
Ginocchio, C., Pace, J., and Galan, J.E. (1992) Identification andmolecular characterization of a Salmonella typhimurium geneinvolved in triggering the internalization of Salmonellae intocultured epithelial cells. Proc Natl Acad Sci USA 89:5976–5980.
Guy, R.L., Gonias, L.A., and Stein, M.A. (2000) Aggregation ofhost endosomes by Salmonella requires SPI2 translocation ofSseFG and involves SpvR and the fms-aroE intragenic region[In Process Citation]. Mol Microbiol 37: 1417–1435.
Hardt, W.D., Chen, L.M., Schuebel, K.E., Bustelo, X.R., andGalan, J.E. (1998) S. typhimurium encodes an activator of RhoGTPases that induces membrane ruffling and nuclearresponses in host cells. Cell 93: 815–826.
Hensel, M. (2000) Salmonella pathogenicity island 2. Mol Micro-biol 36: 1015–1023.
Hensel, M., Shea, J.E., Waterman, S.R., Mundy, R., Nikolaus, T.,Banks, G., et al. (1998) Genes encoding putative effector proteins of the type III secretion system of Salmonella patho-genicity island 2 are required for bacterial virulence and prolif-eration in macrophages. Mol Microbiol 30: 163–174.
Hersh, D., Monack, D.M., Smith, M.R., Ghori, N., Falkow, S., andZychlinsky, A. (1999) The Salmonella invasin SipB inducesmacrophage apoptosis by binding to caspase-1. Proc NatlAcad Sci USA 96: 2396–2401.
Hobbie, S., Chen, L.M., Davis, R.J., and Galan, J.E. (1997)Involvement of mitogen-activated protein kinase pathways inthe nuclear responses and cytokine production induced bySalmonella typhimurium in cultured intestinal epithelial cells. J Immunol 159: 5550–5559.
Hoiseth, S.K., and Stocker, B.A. (1981) Aromatic-dependent Salmonella typhimurium are non-virulent and effective as livevaccines. Nature 291: 238–239.
Ireton, K., Payrastre, B., Chap, H., Ogawa, W., Sakaue, H.,Kasuga, M., and Cossart, P. (1996) A role for phosphoinositide3-kinase in bacterial invasion. Science 274: 780–782.
Isberg, R.R., and Leong, J.M. (1990) Multiple beta 1 chain integrins are receptors for invasin, a protein that promotes bacterial penetration into mammalian cells. Cell 60: 861–871.
Jesenberger, V., Procyk, K.J., Yuan, J., Reipert, S., and Bac-carini, M. (2000) Salmonella-induced caspase-2 activation in macrophages: a novel mechanism in pathogen-mediatedapoptosis. J Exp Med 192: 1035–1046.
Jones, A.T., and Clague, M.J. (1995) Phosphatidylinositol 3-kinase activity is required for early endosome fusion. BiochemJ 311: 31–34.
Lee, A.K., Detweiler, C.S., and Falkow, S. (2000) OmpR regu-lates the two-component system SsrA-ssrB in Salmonellapathogenicity island 2. J Bacteriol 182: 771–781.
Leung, K.Y., and Finlay, B.B. (1991) Intracellular replication isessential for the virulence of Salmonella typhimurium. ProcNatl Acad Sci USA 88: 11470–11474.
Li, G., D’Souza-Schorey, C., Barbieri, M.A., Roberts, R.L.,Klippel, A., Williams, L.T., and Stahl, P.D. (1995) Evidence forphosphatidylinositol 3-kinase as a regulator of endocytosis via activation of Rab5. Proc Natl Acad Sci USA 92: 10207–10211.
Marcus, S.L., Brumell, J.H., Pfeifer, C.G., and Finlay, B.B. (2000)Salmonella pathogenicity islands: big virulence in small pack-ages. Microbes Infect 2: 145–156.
Martys, J.L., Wjasow, C., Gangi, D.M., Kielian, M.C., McGraw,T.E., and Backer, J.M. (1996) Wortmannin-sensitive traffickingpathways in Chinese hamster ovary cells. Differential effectson endocytosis and lysosomal sorting. J Biol Chem 271:10953–10962.
Mecsas, J., Raupach, B., and Falkow, S. (1998) The YersiniaYops inhibit invasion of Listeria, Shigella and Edwardsiella but not Salmonella into epithelial cells. Mol Microbiol 28:1269–1281.
Méresse, S., Steele-Mortimer, O., Finlay, B.B., and Gorvel, J.P.(1999a) The rab7 GTPase controls the maturation of Salmo-nella typhimurium- containing vacuoles in HeLa cells. EMBOJ 18: 4394–4403.
Méresse, S., Steele-Mortimer, O., Moreno, E., Desjardins, M.,Finlay, B., and Gorvel, J.P. (1999b) Controlling the maturationof pathogen-containing vacuoles: a matter of life and death.Nat Cell Biol 1: 183–188.
Méresse, S., Unsworth, K.E., Habermann, A., Griffiths, G., Fang,F., Martinez-Lorenzo, M.J., et al. (2001) Remodelling of the
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54

actin cytoskeleton is essential for replication of intravacuolarSalmonella. Cell Microbiol 3: 567–577.
Mills, S.D., and Finlay, B.B. (1998) Isolation and characterizationof Salmonella typhimurium and Yersinia pseudotuberculosis-containing phagosomes from infected mouse macrophages:Y. pseudotuberculosis traffics to terminal lysosomes wherethey are degraded. Eur J Cell Biol 77: 35–47.
Mukherjee, K., Siddiqi, S.A., Hashim, S., Raje, M., Basu, S.K.,and Mukhopadhyay, A. (2000) Live Salmonella recruits N-ethylmaleimide-sensitive fusion protein on phagosomal membrane and promotes fusion with early endosome. J CellBiol 148: 741–753.
Ochman, H., Soncini, F.C., Solomon, F., and Groisman, E.A.(1996) Identification of a pathogenicity island required for Salmonella survival in host cells. Proc Natl Acad Sci USA 93:7800–7804.
Pace, J., Hayman, M.J., and Galan, J.E. (1993) Signal trans-duction and invasion of epithelial cells by S. typhimurium. Cell72: 505–514.
Patki, V., Virbasius, J., Lane, W.S., Toh, B.H., Shpetner, H.S., andCorvera, S. (1997) Identification of an early endosomal proteinregulated by phosphatidylinositol 3-kinase. Proc Natl Acad SciUSA 94: 7326–7330.
Pfeifer, C.G., Marcus, S.L., Steele-Mortimer, O., Knodler, L.A.,and Finlay, B.B. (1999) Salmonella typhimurium virulencegenes are induced upon bacterial invasion into phagocytic andnonphagocytic cells. Infect Immun 67: 5690–5698.
Prior, I.A., and Clague, M.J. (1999) Localization of a class II phosphatidylinositol 3-kinase, PI3KC2alpha, to clathrin-coatedvesicles. Mol Cell Biol Res Commun 1: 162–166.
Rathman, M., Sjaastad, M.D., and Falkow, S. (1996) Acidificationof phagosomes containing Salmonella typhimurium in murinemacrophages. Infect Immun 64: 2765–2773.
Reaves, B.J., Bright, N.A., Mullock, B.M., and Luzio, J.P. (1996)The effect of wortmannin on the localisation of lysosomal type
I integral membrane glycoproteins suggests a role for phos-phoinositide 3-kinase activity in regulating membrane trafficlate in the endocytic pathway. J Cell Sci 109: 749–762.
Schulte, R., Zumbihl, R., Kampik, D., Fauconnier, A., and Autenrieth, I.B. (1998) Wortmannin blocks Yersinia invasin-triggered internalization, but not interleukin-8 production by epithelial cells. Med Microbiol Immunol (Berl) 187: 53–60.
Shpetner, H., Joly, M., Hartley, D., and Corvera, S. (1996) Poten-tial sites of PI-3 kinase function in the endocytic pathwayrevealed by the PI-3 kinase inhibitor, wortmannin. J Cell Biol132: 595–605.
Steele-Mortimer, O., Méresse, S., Gorvel, J.-P., Toh, B.-H., andFinlay, B.B. (1999) Biogenesis of Salmonella typhimurium-containing vacuoles in epithelial cells involves interactions withthe early endocytic pathway. Cell Micro 1: 33–49.
Steele-Mortimer, O., St-Louis, M., Olivier, M., and Finlay, B.B.(2000a) Vacuole acidification is not required for survival of salmonella enterica serovar typhimurium within culturedmacrophages and epithelial cells. Infect Immun 68: 5401–5404.
Steele-Mortimer, O., Knodler, L.A., Marcus, S., Scheid, M.P.,Pfeifer, C.G., Goh, B., et al. (2000b) Activation of Akt/proteinkinase B by the Salmonella typhimurium effector SigD. J BiolChem 275: 37718–37724.
Tuma, P.L., Finnegan, C.M., Yi, J.H., and Hubbard, A.L. (1999)Evidence for apical endocytosis in polarized hepatic cells:phosphoinositide 3-kinase inhibitors lead to the lysosomalaccumulation of resident apical plasma membrane proteins. J Cell Biol 145: 1089–1102.
Uchiya, K., Barbieri, M.A., Funato, K., Shah, A.H., Stahl, P.D.,and Groisman, E.A. (1999) A Salmonella virulence protein thatinhibits cellular trafficking. EMBO J 18: 3924–3933.
Wallis, T.S., and Galyov, E.E. (2000) Molecular basis of Salmonella-induced enteritis. Mol Microbiol 36: 997–1005.
© 2002 Blackwell Science Ltd, Cellular Microbiology, 4, 43–54
54 O. Steele-Mortimer et al.
Copyright © 2022 FDOKUMEN




















