
Methods of Endotoxin Removal from Biological Preparations: a Review
Upload
independentCategory
view
4download
0
Toxicology Letters. 49 (1989) I-13
Elsevier
1
TOXLET 022 11
Protein A protects mice from depletion of biotransformation enzymes and mortality induced by Salmonella typhimurium endotoxin
P.D. Dwivedi, A.S. Verma, Anil Mishra, K.P. Singh, A.K. Prasad, A.K. Saxena, K.K. Dutta, N. Mathur and P.K. Ray
Immunobiology Division. Industrial Toxicology Research Centre. P.O. Box No. 80, Mahatma Gandhi
Marg, Lucknow 226001 (India)
(Received 10 October 1988)
(Revision received 2 May 1989)
(Accepted 3 1 May 1989)
Ke.v words: Protein A; Biotransformation enzymes; Salmonella typhimurium endotoxin; Protection
SUMMARY
Changes in hepatic microsomal mixed-function oxidase enzyme levels (aniline hydroxylase, aminopyrine
demethylase, glutathione Stransferase), glutathione content, total sulphydryl content, and plasma enzyme
levels of aspartate transaminase, alanine transaminase and alkaline phosphatase were studied in male
Swiss albino mice exposed to Salmonella typhimurium endotoxin (S&l50 fig per mouse, LCsO 141.82 fig).
Animals exposed to the same dose of endotoxin but pretreated with protein A of Sfuphylococcus aureus
(5 pg/per mouse) protected the animals from both mortality and depletion of biotransformation enzymes.
INTRODUCTION
Endotoxins, such as that derived from SaImonella typhimurium, can cause a variety
of changes in the physiology of an animal. The administration of endotoxin to the
mouse increases its body temperature [l] and the levels of free fatty acids and triglyce-
rides. It decreases lipoprotein lipase [2], hepatic cytochrome P-450, aminopyrine de-
methylase and lipid peroxidase levels [3-51. Administration of bacterial endotoxin
(extracted from the cell wall of gram-negative bacteria) lowers mouse serum glucose
level and increases levels of serum pyruvate and acid phosphatase, whereas red blood
Address for correspondence: Dr. P.K. Ray, Director, Industrial Toxicology Research Centre, Post Box
No. 80, Mahatma Gandhi Marg, Lucknow 226001, India.
0378-4274/89/$3,50 @ 1989 Elsevier Science Publishers B.V. (Biomedical Division)

2
cells, haemoglobin and haematocrit values do not seem to change after endotoxin shock [6]. Endotoxin has been shown to be a potent activator for interleukin-1 pro- duction from human monocytes [7].
Protein A, a cell wall glycoprotein of Staphylococcus aweus Cowan I, has a multi- potent immunopotentiating property [8-lo]. Prior studies showed that protein A, when infused intravenously, can cause regression of primary rat mammary tumours [I I-1 31, dog tumours [14] and mouse tumours [IS]. In the present study, it has been investigated whether protein A can minimise or diminish the toxic effects of endo- toxins derived from S. typhimurium, with particular reference to its role in hepatic xenobiotic-metabolising enzymes. Endotoxins are known to decrease the levels of xenobiotic-metabolising enzymes, thereby proving that they are toxic to the liver 13, 4]. As protein A showed abrogation of endotoxin-induced mortality caused by endo- toxin in our preliminary studies, it was of interest to know whether protein A was responsible for the change in xenobiotic-metabolizing enzyme levels, thereby reduc- ing the toxicity of endotoxin.
MATERIALS AND METHODS
Animals Male outbred Swiss albino mice (20 &- 2 g) obtained from the ITRC animal breed-
ing colony and raised on a commercial pellet diet (Hindustan Lever, Bombay) and water ad libitum were used in the present study.
Chemicals Endotoxin extracted from S. typhimurium (LPS) was supplied by Difco Laborato-
ry, U.S.A. Protein A was purchased from Pharmacia Fine Chemicals, Sweden.
The animals were divided into 8 groups of 8 mice each and were treated as follows: Group I Normal saline 0.5 ml i.p. twice weekly for 2 weeks.
(Control) Group II Group III
Group IV
Group V
Group VI
Protein A (5 pg per mouse) i.p. twice weekly for 2 weeks. Protein A (5 fig per mouse) i.p. twice weekly for 2 weeks. Endotoxin 50 ,ug in 0.5 ml normal saline i-p. 48 h after the last injection of protein A. Protein A (5 pg per mouse) i.p. twice weekly for 2 weeks. Endotoxin 100 pg in 0.5 ml normal saline i.p. 48 h after the last injection of protein A. Protein A (5 ,ug per mouse) i.p. twice weekly for 2 weeks. Endotoxin 150 ,ug in 0.5 ml normal saline i.p. 48 h after the last injection of protein A. Endotoxin 50 ,ug in 0.5 ml normal saline per mouse once i.p.

3
Group VII ~ndotoxin 100 ,ug in 0.5 ml normal saline per mouse once i.p. Group VIII Endotoxin 150 I;tg in 0.5 ml normal saline per mouse once i.p.
Preparation of enzyme source material from mouse liver
The animals were killed 48 hours after the last treatment by cervical dislocation. Livers were removed, blotted free of blood, and washed in ice-cold saline. Tissue ho- mogenates were made in 10% of 0.25 M sucrose. Homogenates were centrifuged at 9000 x g for 20 minutes at 4°C. The resulting supernatant (postmitochondrial super- natant) was used for the measurement of cytochrome P-450 dependent enzymes and glutathione S-transferase (GST) activity. Total sulphydryl (-SH) and glutathione (GSH) contents were measured in tissue homogenate.
Co~Iectio~ of blood and separation of plasma
Blood was collected in heparinised test tubes from the retro-orbital plexus of the animals; plasma was separated by centrifugation of blood and was used for estima- tion of alkaline phosphatase (ALK-P), alanine transaminase (ALT) and aspartate transaminase (AST).
Enzyme assays
Aminopyrine demethylase (APD) activity was assayed according to Cochin and Axelrod [ 161. The specific activity was expressed as nanomoles. of formaldehyde fo~ed/min/mg protein. Aniline hydroxylase (AH) activity was assayed by measur- ing the formation of p-aminophenol according to Kato and Gillette [17] and the ac- tivity was expressed as nanomoles of p-aminophenol fo~ed/min/mg protein. GST activity was determined according to the method of Habig et al. [18] using l-chloro- 2,4_dinitrobenzene (CDNB) as substrate.
Total sulphydryl and GSH contents were assayed according to Jollow et al. [19]. AST and ALT in plasma were measured according to the method of Reitman and Frankel [20]. ALK-P in plasma was measured by the method of Wotten [21] using dinitrophenyl phosphate as substrate. Protein content was determined according to Lowry et al. [22], using bovine serum albumin as a reference standard.
statistical analysis
The LCsO value of endotoxin was determined by Probit analysis [23]. The effect of different doses of protein A on the toxicity of endotoxin was analysed
by using two-way classification with an unequal number of animals in different groups. The model thus used was:
Xijk=p++i+Bj+lij+eijk,
where x@ denotes mortality at the ith level of endotoxin and jth level of protein A, and Li and Bi denote the endotoxin and protein A effects, respectively. & represents the interaction between the two factors, ,U the overall mean effect; eijk denotes the independent random error, which is normally distributed at mean zero variance.

4
TABLE I
DETERMINATION OF LETHAL AND SUBLETHAL DOSES OF ENDOTOXIN
Dose of endotoxin &g) S/T (24 h) S/T (48 h)
0 lo/lo lo/lo
50 lo/lo IO/l0
100 s/10 S/IO
200 s/10 2/10
400 2/10 O/IO
S/T = Surviving animals/total animals; LCso = 141.82 pg per mouse; 95% lower confidence = 110.71;
95% upper confidence = 18 1.62.
The values presented for enzyme estimation are means f standard errors. Stu-
dent’s r-test was used for determination of statistical significance. Differences that re-
sulted in probability values smaller than 0.05 were considered to be significant.
RESULTS
Determination of lethal and sublethal doses of S. typhimurium endotoxin
Fifty mice were divided into 5 equal groups and were given 0, 50, 100, 200 and
400 ,ug of endotoxin per mouse respectively by the intraperitoneal route in 0.5 ml
sterile saline (Table I). The control group received only saline. The LC5,, value of en-
q Untreated Control Gr.
61 Gr with pretreatment of lug PA.
q Gr. with pretreatment of 2 up PA
0 Gr. with pretreatment of 5 ,ug PA. 100
150 200
fig Endotoxin /mouse
Fig. 1. Effect of varying doses (lOWlO pg per mouse) of S. typhimurium endotoxin on mortality of mice
pretreated with protein A (PA) (I,2 and 5 fig doses, respectively).

5
dotoxin (48 h) was determined to be 141.82 pg per mouse with a lower confidence
limit (95%) of 110.71 and an upper confidence limit of 181.62.
Determination of the EDJO dose of protein A
The extent of protection provided by protein A (PA) in mice against endotoxin
lethality was estimated by using the latter in doses of 100, 150, 200 and 400 ,ug per
mouse respectively in PA-pretreated mice. Mice were divided into 4 groups, viz. con-
trol (without pretreatment) and those exposed to 1 .O, 2.0 and 5.0 pg PA, respectively
(Fig. 1). Survival was recorded for 7 days, but there was no death in any group after
2 days of endotoxin exposure. When 100 pg of endotoxin (En) was administered, the
mortality in the untreated group (control) was 25%, whereas in 1 ,ug PA-pretreated
(PA + En) animals the mortality was 16.7 % and with 2 pg PA (PA + En) it was 8.3 %.
No mortality was observed in animals treated with 5 pg PA (PA + En).
With 150 pg of endotoxin equal protection was observed in 2 and 5 ,ug PA-treated
groups (8.3%) which is significantly lower (P~0.05) than the mortality in the un-
treated group (33.3 96); 1 pg PA did not give any protection in this group (PA + En).
With 200 pug of endotoxin, the mortality was 73%. The mortalities with 1, 2 and
LJ Untrratw! Control En (lOO,ug )
•m pA El PA +En (100&g 1
61 En(SOfig) /x-J En (15Omg)
PA+En(50mg) I”
PA +En (15Obg)
120 I
NS PA 50 100 150 bg Endotoxin /mouse
Fig. 2. Abrogation of S. r_@irvlurium endotoxin-induced depression of hepatic aniline hydroxylase by pro-
tein A in mice. Note the significant difference between the enzyme levels estimated after treatment with
endotoxin (En) alone (50, 100, 150 fig) as compared with mice pretreated with protein A (PA) (5 pg).
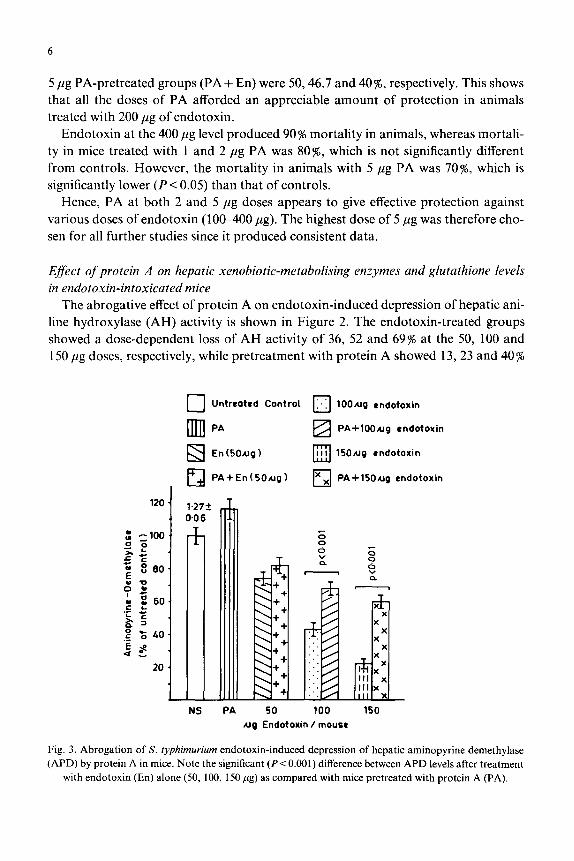
6
5 pg PA-pretreated groups (PA + En) were 50,46.7 and 40 %, respectively. This shows
that all the doses of PA afforded an appreciable amount of protection in animals
treated with 200 pg of endotoxin.
Endotoxin at the 400 ,ug level produced 90% mortality in animals, whereas mortali-
ty in mice treated with 1 and 2 pg PA was 80%, which is not significantly different
from controls. However, the mortality in animals with 5 pg PA was 70%, which is
significantly lower (P < 0.05) than that of controls.
Hence, PA at both 2 and 5 yg doses appears to give effective protection against
various doses of endotoxin (1 O&400 pug). The highest dose of 5 pg was therefore cho-
sen for all further studies since it produced consistent data.
Effect of protein A on hepatic xenobiotic-metabolising enzymes and glutathione levels
in endotoxin-intoxicated mice
The abrogative effect of protein A on endotoxin-induced depression of hepatic ani-
line hydroxylase (AH) activity is shown in Figure 2. The endotoxin-treated groups
showed a dose-dependent loss of AH activity of 36, 52 and 69% at the 50, 100 and
150 pg doses, respectively, while pretreatment with protein A showed 13, 23 and 40 %
cl Untreotrd Control q lOOmu l ndotoxin
ml PA El PA+lOONg l ndotoxin
Ezi En (50&g 1 Q III 15Oug ondotoxin
PA+En(5Oag) PA+15Otig endotoxin
NS PA 50 100 150
ag Endotoxin / mouse
Fig. 3. Abrogation of S. typhimurium endotoxin-induced depression of hepatic aminopyrine demethylase
(APD) by protein A in mice. Note the significant (P-C 0.001) difference between APD levels after treatment
with endotoxin (En) alone (50, 100, 150 pg) as compared with mice pretreated with protein A (PA).
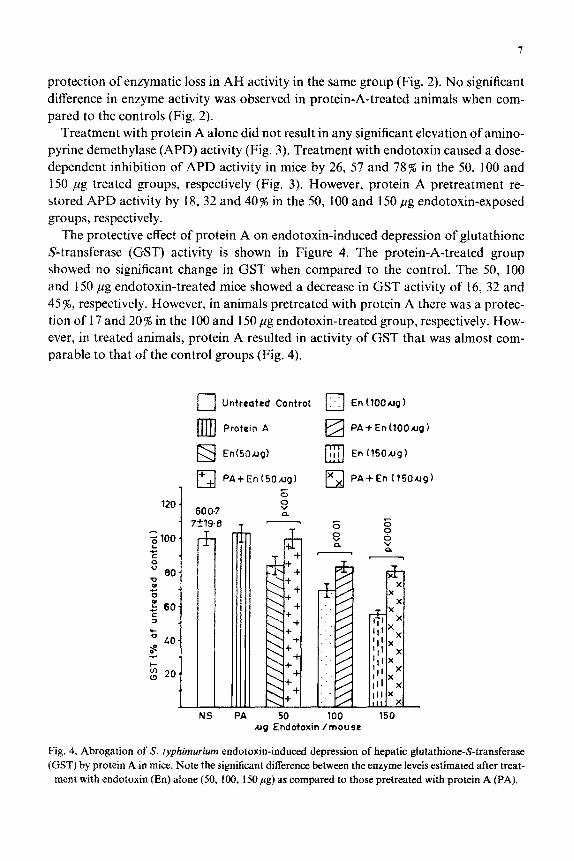
protection of enzymatic loss in AH activity in the same group (Fig. 2). No significant difference in enzyme activity was observed in protein-A-treated animals when com- pared to the controls (Fig. 2).
Treatment with protein A alone did not result in any significant elevation of amino- pyrine demethylase (APD) activity (Fig. 3). Treatment with endotoxin caused a dose- dependent inhibition of APD activity in mice by 26, 57 and 78% in the 50, 100 and 150 fig treated groups, respectively (Fig. 3). However, protein A pretreatment re- stored APD activity by 18, 32 and 40% in the 50, 100 and 150 pg endotoxin-exposed groups, respectively.
The protective effect of protein A on endotoxin-induced depression of glutathione S-transferase (GST) activity is shown in Figure 4. The protein-A-treated group showed no significant change in GST when compared to the control. The 50, 100 and 150 fig endotoxin-treated mice showed a decrease in GST activity of 16, 32 and 45 %> respectively. However, in animals pretreated with protein A there was a protec- tion of 17 and 20% in the 100 and 150 fig endotoxin-treated group, respectively. How- ever, in treated animals, protein A resulted in activity of GST that was almost com- parable to that of the control groups (Fig. 4).
El Untreated Control q En (~00~9)
•n Protein A KI PAiEnUOOugf
&I En&Oug) /?z-J En (15Ofig)
Ll PA+En (5Otig) PA+En (15049) E;
NS PA
60 O-7 $ a
Ng Endotoxm /mouse
Fig. 4. Abrogation of S. ~y~~~uri~ endotoxin-indu~d depression of hepatic glutathione-S-transferase (GST) by protein A in mice. Note the si~ifi~nt difference between the enzyme levels estimated after treat-
ment with endotoxin (En) alone (50, 100,150 pg) as compared to those pretreated with protein A (PA).

il Untrcatrd Control q En(lOOag)
i PA
q En (50.44 1
, El PA+En( 5O~lg)
PA+En (100~~1
Q ‘I’ En (15Ougl
Ll PA+En (15O.ag)
NS PA 50 ag Endotoxin lmouso
Fig. 5. Effect of protein A challenge an hepatic glutathione (GSH) levels in S. f~phi~ur~~~ endotoxin
(En) (50, 100, 1.50 ,~lgf treated mice. Note that protein A (5 p(g) significantly increased (P~0.02) GSH levels
in animals showing depletion of GSH levels following endotoxin inoculation.
Protein A treatment alone did not result in any significant increase in GSH content
(Fig. 5). The endotoxin-treated group showed a reduction of 10, 21 and 41% with
increasing doses of endotoxin. However, protein A pretreatment restored the GSH
content by bringing it back to the normal level in the 50 pug endotoxin group but
it was 8 and 14% less than control in the 100 and 150 pig endotoxin groups, respective-
Protein A treatment alone showed a margina increase of 996 in total sulphydryl
(-SH) content. Endotoxin treatment caused a 12, 6 and 12% decrease in total SH
content in 50, 100 and 150 ,ug exposed animals. In this case protein A pretreatment
did not markedly affect the total -SH content (Fig. 6).
Efect qf protein A on plasma enzymes, e.g. alkaline phosphatase (ALK-P) , aspartate transaminase ( AST) and alanine transaminase (ALT) , in endotosin-intoxicated mice
Protein A treatment alone did not affect the plasma enzyme levels of ALK-P, AST
and ALT as compared to the enzyme levels among controls treated with normal sa-
line (Table II). Treatment of mice with endotoxin resulted in a significant elevation
of plasma ALK-P (2~70~), AST (3462%) and ALT (103-169%) in a dose-depen-

9
c) Untreated Control
ml PA
fzI En (5ONg)
lzl PA+En(50q)
120 30.98
El 1,‘. En (?OO~g)
Ezl PA+En (lOO&gf
q III En(l50tig)
X
Q PA+ En ( 150 ug 1
NS PA 50 100 ug Endotoxin /moos8
Fig. 6. Effect of protein A (PA) challenge on hepatic total sulphydryl (SH) levels in S. ~yp~~~uri~~ en&-
toxin (En) (50, 100, 150 pg) treated mice.
dent manner. Pretreatment with protein A resulted in substantial lowering of ALK-P (9-21%), AST (I 3-19s) and ALT (19-24s) values (Table II). Although protein A pretreatment did not totally normalise the enhanced plasma enzyme levels caused by endotoxin exposure, it did reduce to some extent the enhanced levels of plasma en- zymes.
DISCUSSION
The xenobiotic-metabolising enzyme system is responsible for the biotransfo~a- tion of many drugs, xenobiotics, pesticides and certain endogenous substances in- cluding steroids, fatty acids and prostagtandins. The activity of the drug-metabolising enzyme system is necessary for inactivation of many therapeutic agents as well as for the activation of several prodrugs.
Toxicity induced by endotoxin, such as early mortality of test animals and en- hancement of ALK-P and AST/ALT, has been shown to be significantly abrogated by pretreatment of test mice with protein A. It is well known that endotoxin de- presses the xenobiotic-metabolizing enzyme system [24-271, but there is no definite information about their exact mode of action. Endotoxins may act in two or more

10
TABLE 11
EFFECT OF VARYING DOSES (SO-150 fig) OF ENDOTOXIN (En) CHALLENGE GIVEN TO
MICE, PRETREATED WITH 5 .ug PROTEIN A (PA) ON PLASMA ACTIVITY OF ALKALINE
PHOSPHATASE (ALK-P), ASPARTATE TRANSAMINASE (AST) AND ALANINE TRANSAMIN-
ASE (ALT) ACTIVITIES
Treatment ALK-P Increase (s) AST (GOT) Increase (96) ALT (GPT) Increase (%)
Control (NS)
Protein A
alone (5 fig)
58.3 + 1.44 0.0
60.6 f 1.83 4.0
Endotoxin (50 pg) 73.28 & 3.24 26.0
PA + En (50 ,ug) 66.9 +5.49 15(9)
Endotoxin (100
pg) 87.7 k3.17 51.0
PA+En(lOO~g) 76.58*4.8 32(13)
Endotoxin (150
fig) 99.22k6.7 70.0
PA+En (15Opg) 78.85+5.1 35(21)
125.28 + 4.2 0.0
127.22k4.5 0.0
167.73 f 5.8 34 136.0 k6.1 9(9)
188.7 +7.0 51
164.3 kg.12 31(13)
202.3 k7.7 62
174.58k8.94 40(14)
45.0 *3.1 0.0
45.5 +3.1 0.0
92.41+ 5.7 103
73.6 *2.85 62(20)
105.8 +6.7 132 85.7 +3.5 88(19)
122.28+5.13 169 92.9 +3.86 155(24)
Each value is mean k SE of 6 animals. Activities are expressed in pmoles product formed/min/l plasma.
PA = 5 pg. Values in parentheses show percent protection given to respective endotoxin-treated groups.
ways, first by causing depression or activation of the reticuloendothelial system and
secondly by inducing the formation of interferon, both of which appear to be impli-
cated in reducing the hepatic metabolism of drugs.
Since cytochrome P-450 is composed of an apoprotein and haem embedded in a
lipophilic environment, interaction of the reactive intermediates of either of these
components may lead to inactivation. Treatment with endotoxin induces haem oxy-
genase [28]. which can degrade haem from cytochrome P-450 in hepatic reticuloen-
dothelial cells.
Interferon inducers [29] as well as purified preparations of human interferon [30]
are known to depress hepatic cytochrome P-450 dependent mono-oxygenase system
enzymes. Endotoxin does not affect hepatic drug metabolism in isolated parenchymal
cells [31, 321, suggesting a role of certain endogenously released factors. The macro-
phage, a well-known target for endotoxin, releases a number of secretory products
upon endotoxin exposure of which interleukin-1 [33] has been reported to depress
liver drug metabolism in endotoxin-treated mice. On the other hand, we have re-
ported recently that protein A stimulates macrophages in both normal and tumour-
bearing hosts [9].
Our studies have shown that a purified protein (protein A) from S. aureu.s
Cowan I can save animals from dying due to toxicity associated with endotoxin (Fig.

11
1). Animals treated with purified protein A did not show any mortality or loss of body weight indicating that protein A itself is not toxic at this dose (data not shown).
Pretreatment of animals with protein A, which were subsequently exposed to endo- toxin, protects the xenobiotic-metabolizing apparatus from endotoxin toxicity. It may exert its effect by modifying reticuloendothelial function [34], by abrogating this interferon-inducing ability of cells or by inhibiting interleukin-1 producing activity by macrophages, or it may directly act with the endotoxin. Interestingly, we have ob- served significant effects of protein A in increasing both macrophage numbers and function in normal as well as tumour-bearing hosts [9]. Protein A itself has been re- ported to have the ability to induce interferon activity [8]. Thus, abrogation of endo- toxin activity by protein A could rest& from its competitive binding on similar types of receptors on reticuloendothelial cells as of endotoxin. The latter being too lethal, it exerts its toxicity too rapidly. However, if the endotoxin-binding sites are partly blocked by protein A, lethality can be expected to be reduced. More research, howev- er, is needed to prove or disprove such a hypothesis.
One of the known effects of endotoxin on the liver in vivo is hepatocellular necrosis [35]. E. coli endotoxin is known to cause liver injury and thereby an increase in aspar- tate transaminase (AST) [3.5]. Bacterial endotoxins are also known to be an impor- tant cause of cholestasis [36], which increases alkaline phosphatase activity in plasma. The studies mentioned above fully corroborate our results, showing liver injury endo- toxin-treated mice and thereby an increase in AST, ALT and alkaline phosphatase activity. Pretreatment with protein A reduces levels of plasma enzymes, thus indicat- ing a lesser magnitude of liver injury.
The observations made in this study corroborate our earlier findings with cyclo- phosphamide [38, 391 and CC4 [40-42]. It has been shown earlier that toxicity in- duced following exposure to a large dose of cyclophosphamide could be abrogated by protein A pretreatment [38]. Protein-A-pretreated animals showed an excellent re- covery, accelerated regeneration of the damage, and repletion of blood cells and en- zymes of the hepatic mixed-function oxidase (MFO) system [38].
In conclusion, it may be said that because of the fast recovery of MFO levels this system may provide an excellent model for the investigation of xenobiotic-induced liver damage and its regeneration. Additionally, the present observations appear to indicate a unique interaction between protein A activity and hepatic metabolic activi- ty, thus supporting our previous observations [40]. However, detailed studies are needed to understand the exact mechanism of the action of protein A in protecting the MFO system from the endotoxin toxicity reported here as well as from the cyclo- phosphamide and CC14 toxicity reported earlier [39,40].
ACKNOWLEDGEMENTS
We are grateful to Dr. Mukul Das and Dr. B.M. Gupta for critical evaluation of the manuscript and for their fruitful suggestions. Thanks are also due to Mr. Chedi

12
La1 for sustained help and to Mr. L.K. Goswami for secretarial assistance. Financial assistance to one of the authors (P.D.D.) from the Council of Scientific and Indus- trial Research (CSIR), India, is gratefully acknowledged.
REFERENCES
1 Heikkill, J.J. and Brown, I.R. (1977) Hyperthermia and disaggregation of brain polysomes induced
by bacterial pyrogen. Life Sci. 25,347-352.
2 Sakaguchi, S.. Sakaguchi. 0. and Hus, C.C. (1979) Metabolic disorders of lipids in mice administered
with endotoxin. Jpn J. Med. Sci. Biol. 32.95-98.
3 Green. S. and Dorbjansky. A. (1979) Corynebacterium parvum effects on the biochemistry of mouse
serum and liver. J. Natl. Cancer Inst. 63.407-502.
4 Egawa, K. and Kasai, N. (1979) Endotoxin glycohpid as a potent depressor of the hepatic drug meta-
bolizing enzyme systems in mice. Microbial. Immunol. 23.89-94.
5 Sanewane, B.R., Yaffe, S.J. and Witmer, C.M. (1982) Effects of endotoxin upon rat hepatic microso-
ma1 drug metabolism in vivo and in vitro. Xenobiotica 12.3033312.
6 Mohammed, T.. Rahman Abdul. M. and Parmer, N.S. (1986) Decrease by naloxone of some electro-
cardiographic and biochemical changes following endotoxin induced shock in rats. Toxicon 24,
101-103.
7 Newton, R.C. (1986) Human monocyte production of interleukin-I: parameters of the induction of
interleukin-I secretion by lipopolysaccharide. J. Leukocyte, Biol. 39.299-312.
8 Catalona. W.J.. Ratliff, T.L. and McCool, R.E. (1981) Interferon induced by S. aureu.r protein A aug-
ments natural killing and ADCC. Nature 291, 77-79.
9 Prasad, A.K., Singh, K.P.. Saxena, A.K., Mathur, N. and Ray, P.K. (1987) Increased macrophage
activity in protein A treated tumor regressed animals. Immunopharmacol. Immunotoxicol. 9, %-%I.
IO Ray, P.K., Rayc~dudhuri, S. and Allen, P. (1982) Mechanism of mammary adenocarcinomas in rats
following plasma adsorption over protein A containing ~~u~h.~~oc~c~us aweus. Cancer Res. 42,
49704974.
11 Ray. P.K. and Bandyopadhyay, SK. (1983) Inhibition of rat mammary tumor growth by purified pro-
tein A: a potential anti-tumor agent. Immunol. Commun. 12,4533464.
12 Ray, P.K., Bandyopadhyay. S., Dohadwala. M., Canchanapan, P. and Mobini, J. (1984) Antitumor
activity with nontoxic doses of protein A. Cancer Immunol. Immunother. 18, 29-34.
13 Ray. P.K., McLaughlin, D., Allen. P., Bandyopadhyay, S., Idiculla, A.. Clarke, L., Mark, R., Rhoads
Jr., J.E., Basset. J.J. and Cooper, D.R. (1984) Adsorption of plasma from tumor bearing host over
protein A containing non viable Staphylococcus aweus Cowan I: possible mechanism of antitumor re-
action. J. Biol. Res. Modif. 3, 293-302.
14 Ray, P.K., McLaughlin, D., Mohammed, J.. Idiculla. A., Rhoads Jr., J.E., Mark, R., Basset, J.G. and
Cooper. D.R. (1981) Ex vivo immunoadsorption of IgG or its complexes ~ a new modality of cancer
treatment. In: B. Serroui and C. Rosenfeld (Eds.), Inlmune Complexes and Plasma Exchanges in Can-
cer Patients. ~Isevier~North-Holland Biomedical Press, pp. 197-207.
15 Saha, S.. Prdsad, A.K. and Ray, P.K. (1987) Changes and correlation of plasma immunoglobulins with
tumor growth in fibrosarcoma bearing mice. Ind. J. Exp. Biol. 25.514518.
16 Cochin, J. and Axelrod. J. (1959) Biochemical and pharmacological changes in the rat following chron-
ic administration of morphine, nalorphine and normorphine. J. Pharmacol. Exp. Ther. 125. 105-l 10.
17 Kato, R. and Gillette, J.R. (1965) Effect of starvation of NADPH dependent enzymes in liver micro-
somes of male and female rats. J. Pharmacol. Exp. Ther. 150, 2799284.
18 Habig, W.H.. Pabst, M.J. and Jakoby, W.B. (1974) Glutathione-S-transferase: the first enzymatic step
in mercapturic acid formation. J. Biol. Chem. 249, 713~-7139.
19 Jollow. D.J.. Mitchell. J.R., Zampaglione, N. and Gillette, J.R. (1974) Bromobenzene induced liver
necrcsis: protective role of glutathione and evidence for 3,4_bromobenzene oxide as the hepatotoxic
intermediate. Pharmacology 11, 151-169.

13
20 Reitman, S. and Frankel, S. (1957) A eolorimetric method for the dete~ination of serum glutamic
oxalacetic and glutamic pyruvate transaminases. Am. J. Clin. Pathol. 28, 5663.
21 Wotten. I.D.P. (1964) In: Microanalysis in Medical Biochemistry, 4th edn. J.A. Churchill Ltd., Lon-
don, pa 101.
22 Lowry, O.H.. Rosebrough. N.J., Farr, A.L. and Randall, R.J. (1951) Protein measurement with the
Folin phenol reagent. J. Biol. Chem. 193,265-275.
23 Finney. D.J. (1978) In: Statistical Methods in Biological Assay, 3rd edn., Charles Griffin and Company
Ltd., London and High Wycombe, pp. 3722385.
24 Gorodisher, R.. Krasner. J.. McDevitt, J.J., Nolan, J.P. and Yaffe, S.J. (1976) Hepatic microsomal
drug metabolism after administration of endotoxin in rats. Biochem. Pharmacol. 25, 351-352.
25 Abernathy, C.O. and Zimmerman, H.J. (1980) Effect of endotoxin tolerance on in vivo drug metabo-
lism in mice. Res. Commun. Chem. Pathol. Pharmacol. 29. 1933196.
26 Williams, J.F. and Szentivanyi, A. (1973) Investigation of adrenergic and prosta~andin inff~~ences in
the endotoxin alteration of hepatic heme oxygenase, microsomal mixed function oxidase and glucocor-
ticoid-induced tryptophan oxygenase activities. Immunopha~acology 675-86.
27 Williams, J.F.. Low&t, S. and Szentivanyi, A. (1980) Endotoxin depression of hepatic mixed function
oxidase system in C3H/HeJ and C3H/HeN mice. Immunopharmacology 2,2855291.
28 Gemesa. D., Woo. C.H., Fudenberg, H. and Schmide, R. (1974) Stimulation of heme oxygenase in
macrophage and liver by endotoxin. J. Clin. Invest. $3. 647651.
29 Renton, K.W. and Mennering. G.J. (1976) Depression of hepatic cytochrome P-450 dependent mono-
xygenase system with administrated interferon inducing agent. Biochem. Biophys. Res. Commun. 73,
343-348.
30 Single, G.. Renton, K.W. and Stebbing, N. (1981) Effect of interferon on hepatic cytochrome P-450
drug metabolizing systems. Pharmacologist 23, 283-287.
31 Berry, L.J. (1977) Bacterial toxins. CRC Crit. Rev. Toxicol. 5239318.
32 Filkins, J.P. and Buchanan, B.J. (1977) In vivo vs. in vitro effects of endotoxin on glycogenolysis, glu-
coneogenesis and glucose utilization. Proc. Sot. Exp. Biol. Med. I55.216218.
33 Ghezzi, P.. Saccardo, B., Villa. P., Rossi, U., Bianchi, M. and Dinerello, CA. (1986) Role of inter-
leukin 1 in the depression of liver drug metabolism by endotoxin. Infect. Immun. 54,837-840.
34 Singh, K.P., Saxena. A.K., Prasad, A.K.. Dwivedi, P.D., Zaidi, S.I.A. and Ray, P.K. (1987) Effect
of protein A on mast cell numbers and macrophage phagocytic activity. Immunopharmacol. Immuno-
toxicol. 283, 281-~298.
35 Lee, D.W., Kim, C.S.. Lee. Y .B. and Kim, D.S. (1973) A study of hepatic injury induced by endotoxin
in rats. Yonsei Med. J. 19, 19.
36 Curtis, M.J., Jenkins, H.G. and Butler, E.J. (1980) The effect of E. coli endotoxin and adenocortical
hormone on plasma enzyme activities in the domestic fowl. Res. Vet. Sci. 28,445O.
37 Minuk, G.Y., Rascanin, N., Sarjeant. E.S. and Pai, C.H. (1986) Sepsis and cholestasis: the in vitro
effects of bacterial products in C-14 teusocholete uptake by isolated rat hepatocytes. Liver 6, 199-204.
38 Ray, P.K., Dohad~dla, M., Bandyopadhyay, SK. and McLaughlin, D. (1985) Rescue of rats from
large dose cyclophosphamide toxicity using protein A. Cancer Chemother. Pharmacol. 14,59-62.
39 Dohadwala. M. and Ray, P.K. (1985) In vivo protection by protein A of hepatic microsomal mixed
function oxygenase system of cyclophosphamide treated rats. Cancer Chemother. Pharmacol. 14,
135-138.
40 Srivastava S.P., Singh. K.P., Saxena, A.K., Seth, P.K. and Ray, P.K. (1987) In vivo protection by
protein A of hepatic microsomal mixed function oxidase system of CCL administered rats. Biochem.
Pharmacol. 36.405554058.
41 Singh, K.P., Saxena, A.K., Zaidi, S.I.A., Dwivedi. P.D., Srivastava, S.P., Seth, P.K. and Ray, P.K.
(1988) Protection against carbon tetrachloride induced hepatotoxicity in rats by protein A. J. Appl.
Toxicol. 8, 407410.
42 Singh, K.P., Saxena. A.K.. Srivastava, S.P., Seth. P.K., Zaidi, S.I.A., Dwivedi, P.D. and Ray, PK.
(1988) Protection against carbon tetrachloride induced lymphoid organotoxicity in rats by protein A.
Toxicol. Lett. (submitt~).
Copyright © 2022 FDOKUMEN




















