
Hemokinin-1 is an important mediator of endotoxin-induced acute airway inflammation in the mouse
7
Peptides 64 (2015) 1–7 Contents lists available at ScienceDirect Peptides j ourna l h o mepa ge: www.elsevier.com/locate/peptides Hemokinin-1 is an important mediator of endotoxin-induced acute airway inflammation in the mouse Zsófia Hajna a,b , Éva Borbély a,b , Ágnes Kemény a,b , Bálint Botz a,b , László Kereskai c , János Szolcsányi a,e , Erika Pintér a,b,e , Christopher J. Paige d , Alexandra Berger d , Zsuzsanna Helyes a,b,e,f,∗ a Department of Pharmacology and Pharmacotherapy, Faculty of Medicine, University of Pécs, Pécs, Hungary b Szentágothai Research Centre, University of Pécs, Pécs, Hungary c Department of Pathology, Faculty of Medicine, University of Pécs, Pécs, Hungary d Ontario Cancer Institute, University Health Network, Toronto, Canada e PharmInVivo Ltd, Pécs, Hungary f MTA-PTE NAP B Pain Research Group a r t i c l e i n f o Article history: Received 24 October 2014 Received in revised form 12 December 2014 Accepted 12 December 2014 Available online 22 December 2014 Keywords: Acute pneumonitis Tachykinin Luminescence imaging Reactive oxygen species a b s t r a c t Objective: Hemokinin-1, the newest tachykinin encoded by the preprotachykinin C (Tac4) gene, is pre- dominatly produced by immune cells. Similarly to substance P, it has the greatest affinity to the tachykinin NK1 receptor, but has different binding site and signaling mechanisms. Furthermore, several recent data indicate the existence of a not yet identified own receptor and divergent non-NK1-mediated actions. Since there is no information on its functions in the airways, we investigated its role in endotoxin-induced pulmonary inflammation. Methods: Acute pneumonitis was induced in Tac4 gene-deleted (Tac4 −/− ) mice compared to C57Bl/6 wildtypes by intranasal E. coli lipopolysaccharide (LPS). Airway responsiveness to inhaled carbachol was measured with unrestrained whole body plethysmography 24 h later. Semiquantitative histopatho- logical scoring was performed; reactive oxygen species (ROS) production was measured with luminol bioluminescence, myeloperoxidase activity with spectrophotometry, and inflammatory cytokines with Luminex. Results: All inflammatory parameters, such as histopathological alterations (perivascular edema, neu- trophil/macrophage accumulation, goblet cell hyperplasia), myeloperoxidase activity, ROS production, as well as interleukin-1beta, interleukin-6, tumor necrosis factor alpha, monocyte chemoattractant protein- 1 and keratinocyte chemoattractant concentrations were significantly diminished in the lung of Tac4 −/− mice. However, bronchial hyperreactivity similarly developed in both groups. Interestingly, in LPS-treated Tac4 −/− mouse lungs, bronchus-associated, large, follicle-like lymphoid structures developed. Conclusions: We provide the first evidence that hemokinin-1 plays a crucial pro-inflammatory role in the lung by increasing inflammatory cell activities, and might also be a specific regulator of lymphocyte functions. © 2014 Elsevier Inc. All rights reserved. Introduction The preprotachykinin C (Tac4) gene was discovered in 2000 pro- viding the newest member of the tachykinin family [43]. The Tac4 gene encodes several related peptides in different species, such as ∗ Corresponding author at: Department of Pharmacology and Pharmacotherapy, Faculty of Medicine, University of Pécs, Szigeti út 12., H-7624 Pécs, Hungary. Tel.: +36 72 536 000/35097; fax: +36 72 536 218. E-mail address: [email protected] (Z. Helyes). hemokinin-1 (HK-1) in mice, endokinin-1 and -2 (EK-1, EK-2) in rabbits, as well as endokinins A-D (EKA, EKB, EKC, EKD) in humans [31]. The 11-amino-acid-long HK-1 shows structural similarities to substance P (SP) resulting in strong immunological crossreactivity with anti-SP antibodies [31] that might have led to several inade- quate conclusions in the literature. HK-1 is a highly selective and potent agonist at the neu- rokinin 1 (NK1) receptor [1,22] localized mainly on endothelial and immune cells besides neurones. The activation of NK1 receptors by HK-1 exerts recruitment and differentiation of inflammatory and http://dx.doi.org/10.1016/j.peptides.2014.12.002 0196-9781/© 2014 Elsevier Inc. All rights reserved.
-
Upload
medbiotech -
Category
Documents
-
view
0 -
download
0
Transcript of Hemokinin-1 is an important mediator of endotoxin-induced acute airway inflammation in the mouse

Ha
ZLAa
b
c
d
e
f
a
ARR1AA
KATLR
I
vg
FT
h0
Peptides 64 (2015) 1–7
Contents lists available at ScienceDirect
Peptides
j ourna l h o mepa ge: www.elsev ier .com/ locate /pept ides
emokinin-1 is an important mediator of endotoxin-induced acuteirway inflammation in the mouse
sófia Hajnaa,b, Éva Borbélya,b, Ágnes Keménya,b, Bálint Botza,b,ászló Kereskai c, János Szolcsányia,e, Erika Pintéra,b,e, Christopher J. Paiged,lexandra Bergerd, Zsuzsanna Helyesa,b,e,f,∗
Department of Pharmacology and Pharmacotherapy, Faculty of Medicine, University of Pécs, Pécs, HungarySzentágothai Research Centre, University of Pécs, Pécs, HungaryDepartment of Pathology, Faculty of Medicine, University of Pécs, Pécs, HungaryOntario Cancer Institute, University Health Network, Toronto, CanadaPharmInVivo Ltd, Pécs, HungaryMTA-PTE NAP B Pain Research Group
r t i c l e i n f o
rticle history:eceived 24 October 2014eceived in revised form2 December 2014ccepted 12 December 2014vailable online 22 December 2014
eywords:cute pneumonitisachykininuminescence imagingeactive oxygen species
a b s t r a c t
Objective: Hemokinin-1, the newest tachykinin encoded by the preprotachykinin C (Tac4) gene, is pre-dominatly produced by immune cells. Similarly to substance P, it has the greatest affinity to the tachykininNK1 receptor, but has different binding site and signaling mechanisms. Furthermore, several recent dataindicate the existence of a not yet identified own receptor and divergent non-NK1-mediated actions.Since there is no information on its functions in the airways, we investigated its role in endotoxin-inducedpulmonary inflammation.Methods: Acute pneumonitis was induced in Tac4 gene-deleted (Tac4−/−) mice compared to C57Bl/6wildtypes by intranasal E. coli lipopolysaccharide (LPS). Airway responsiveness to inhaled carbacholwas measured with unrestrained whole body plethysmography 24 h later. Semiquantitative histopatho-logical scoring was performed; reactive oxygen species (ROS) production was measured with luminolbioluminescence, myeloperoxidase activity with spectrophotometry, and inflammatory cytokines withLuminex.Results: All inflammatory parameters, such as histopathological alterations (perivascular edema, neu-trophil/macrophage accumulation, goblet cell hyperplasia), myeloperoxidase activity, ROS production, aswell as interleukin-1beta, interleukin-6, tumor necrosis factor alpha, monocyte chemoattractant protein-1 and keratinocyte chemoattractant concentrations were significantly diminished in the lung of Tac4−/−
mice. However, bronchial hyperreactivity similarly developed in both groups. Interestingly, in LPS-treatedTac4−/− mouse lungs, bronchus-associated, large, follicle-like lymphoid structures developed.Conclusions: We provide the first evidence that hemokinin-1 plays a crucial pro-inflammatory role inthe lung by increasing inflammatory cell activities, and might also be a specific regulator of lymphocytefunctions.
ntroduction
The preprotachykinin C (Tac4) gene was discovered in 2000 pro-iding the newest member of the tachykinin family [43]. The Tac4ene encodes several related peptides in different species, such as
∗ Corresponding author at: Department of Pharmacology and Pharmacotherapy,aculty of Medicine, University of Pécs, Szigeti út 12., H-7624 Pécs, Hungary.el.: +36 72 536 000/35097; fax: +36 72 536 218.
E-mail address: [email protected] (Z. Helyes).
ttp://dx.doi.org/10.1016/j.peptides.2014.12.002196-9781/© 2014 Elsevier Inc. All rights reserved.
© 2014 Elsevier Inc. All rights reserved.
hemokinin-1 (HK-1) in mice, endokinin-1 and -2 (EK-1, EK-2) inrabbits, as well as endokinins A-D (EKA, EKB, EKC, EKD) in humans[31].
The 11-amino-acid-long HK-1 shows structural similarities tosubstance P (SP) resulting in strong immunological crossreactivitywith anti-SP antibodies [31] that might have led to several inade-quate conclusions in the literature.
HK-1 is a highly selective and potent agonist at the neu-rokinin 1 (NK1) receptor [1,22] localized mainly on endothelial andimmune cells besides neurones. The activation of NK1 receptors byHK-1 exerts recruitment and differentiation of inflammatory and

2 eptide
iem[fp
esp[piaadnp
ccl1hb
ntn[bg2[trmt
cevaH
M
E
As(mo
A
m(dmwm
Z. Hajna et al. / P
mmune cells. However, as compared to SP, HK-1 might have differ-nt binding site on NK1 receptors; moreover, its receptor activationechanism or signal transduction pathway might also be different
22]. However, based on several data suggesting effects differentrom that of NK1 receptor activation, the existence of an undefined,resumably own HK-1 receptor has also been proposed [4,11].
In contrast to classical tachykinins, HK-1 is predominantlyxpressed in non-neuronal tissues. Although Tac4 is expressed inome brain regions, its expression level is significantly higher ineripheral tissues compared to the preprotachykinin A (Tac1) gene9] in various inflammatory and immune cells, such as T and B lym-hocytes, macrophages and dendritic cells [27,43]. Therefore, HK-1
s suggested to participate in inflammatory processes and to playn important role in immune regulation. Tac4 gene expression haslso been detected in human bronchi [14]. Moreover recent clinicalata suggest that there might be a correlation between Tac4 (butot Tac1) polymorphisms and inflammatory processes in asthmaatients [21].
HK-1 has been described to be involved in several biologi-al actions. Its role in the hematopoietic system has been wellharacterized, such as in the development of B cells [2,13] and Tymphopoesis [7]. Furthermore, intrathecal administration of HK-
is able to evoke nociceptive responses [11,41], but not thermalyperalgesia [38] and it also decreases blood pressure and causeradycardia [1].
We have previously established that besides anti-inflammatoryeuropeptides, such as somatostatin [18], pro-inflammatoryachykinins of predominantly sensory neural origin, such as SP andeurokinin A (NKA) play an important role in airway inflammation16]. SP and NKA, encoded by the Tac1 gene and expressed mainlyy capsaicin-sensitive primary sensory neurones of the dorsal rootanglia, exert these effects via the activation of NK1 and neurokinin
(NK2) receptors, to which they show greatest affinity, respectively23]. The recent discovery of Tac4-derived tachykinins and the facthat they have been shown to be expressed in the airways [14]aised the possibility that this group of hemokinins and endokininsay be the missing link in the pathophysiological mechanism of
achykinin-mediated pro-inflammatory actions [33].Due to the similarities of SP and HK-1, data regarding their spe-
ific functions are often confusing. To reveal the roles of HK-1 inndotoxin-induced airway inflammation in comparison with pre-iously obtained results with SP [16], we designed and performed
complex examination with the help of mice specifically lackingK-1.
aterials and methods
thical approval
All experiments were performed according to the 1998/XXVIIIct of the Hungarian Parliament on Animal Protection and Con-ideration Decree of Scientific Procedures of Animal Experiments243/1988). They were approved by the Ethics Committee on Ani-
al Research of University of Pécs according to the Ethical Codexf Animal Experiments (licence No.: BA02/2000-5/2011).
nimals
Experiments were performed on female Tac4 gene-deficientice (Tac4−/−) compared to C57Bl/6 wildtype (WT) controls
20–25 g, 8–10 weeks). The original breeding pairs of Tac4−/− gene-
eficient animals were donated by Berger et al. [2], while C57Bl/6ice were purchased from Charles-River Ltd. (Hungary). Animalsere bred and kept in the Laboratory Animal House of the Depart-ent of Pharmacology and Pharmacotherapy, University of Pécss 64 (2015) 1–7
at 24–25 ◦C, provided with standard chow and water ad libitum,maintained under 12-h light–dark cycle.
Experimental protocol
Acute airway inflammation was elicited by intranasal adminis-tration of Escherichia coli (serotype: 083) endotoxin (lipopolysac-charide, LPS; 60 �l, 167 �g/ml) inducing an interstitial pneumonitiswith a well-defined mechanism. LPS stimulates Toll-like recep-tor 4 (TLR4)-CD14 glycoprotein complex on immune cells andbronchial epithelia [15] resulting in their activation, accumulationand inflammatory cytokine release. These cells release leukotriens,prostaglandins, bradykinin, etc., directly activating sensory nerves[17]. LPS was administered under light ether anesthesia, non-inflamed intact mice received the same volume of sterile PBS asdescribed [16].
Functional investigation of the airway responsiveness
Airway responsiveness was determined by unrestrained wholebody plethysmography (Buxco Europe Ltd, Winchester, UK) inconscious, spontaneously breathing animals 24 h after PBS/LPSadministration. Bronchoconstriction was induced by increasingconcentrations (5.5, 11 and 22 mM) of aerosolized carbachol (car-bamoylcholine; Sigma, St. Louis, MO, USA; dissolved in saline, 50 �lper mouse) following baseline measurements performed withaerosolized saline. We determined the breathing frequency andenhanced pause (Penh) value, as an indicator of bronchoconstric-tion. Penh is a calculated parameter ((expiratory time/relaxationtime) − 1)/(max. expiratory flow/max. inspiratory flow), corre-lating with airway resistance measured by traditional invasivetechniques using ventilated animals [18].
Histological examination
Following functional airway measurements mice were anaes-thesized with ketamine (100 mg/kg i.p.; Calypsol; Richter-GedeonPlc., Budapest, Hungary) and xylazine (10 mg/kg i.m.; Xylavet;Phylaxia-Sanofi, Veterinary Biology Co. Ltd., Budapest, Hungary),sacrificed by cervical dislocation and their lungs were excised [18].Left lungs were fixed in 4% formaldehyde for 8 h, embedded inparaffin, sectioned with a microtome (5–7 �m) and stained withhematoxylin (Sigma, St. Louis, MO, USA) and eosin (Molar Chem-icals Ltd., Budapest, Hungary). One section of each sample wasstained with periodic acid-Schiff reaction for the visualization ofmucus producing goblet cells. Semiquantitative histopathologicalscoring of the inflammatory changes was performed by an expertpathologist blinded from the experimental design. Scoring param-eters were determined on the basis of the extent of perivascularedema (0–3), perivascular/peribronchial neutrophil accumulation(0–3), infiltration of activated macrophages/mononuclear cells inthe alveolar spaces (0–2) and goblet cell hyperplasia of the bron-chioles (0–2), as previously described [16,42]. Score values given forthese individual histopathological features were added to generatea composite inflammatory score ranging between 0 and 10.
Determination of the myeloperoxidase (MPO) activity in the lungwith spectrophotometry
The endotoxin-induced inflammatory reaction of the pul-monary tissues results in the recruitment of neutrophil granulo-cytes, monocytes and macrophages of myeloid origin expressing
MPO enzyme [29,34]. The activity of this enzyme correlates withthe amount of neutrophils and macrophages and is a widely usedbiochemical marker for the evaluation of the inflammatory reactionintensity.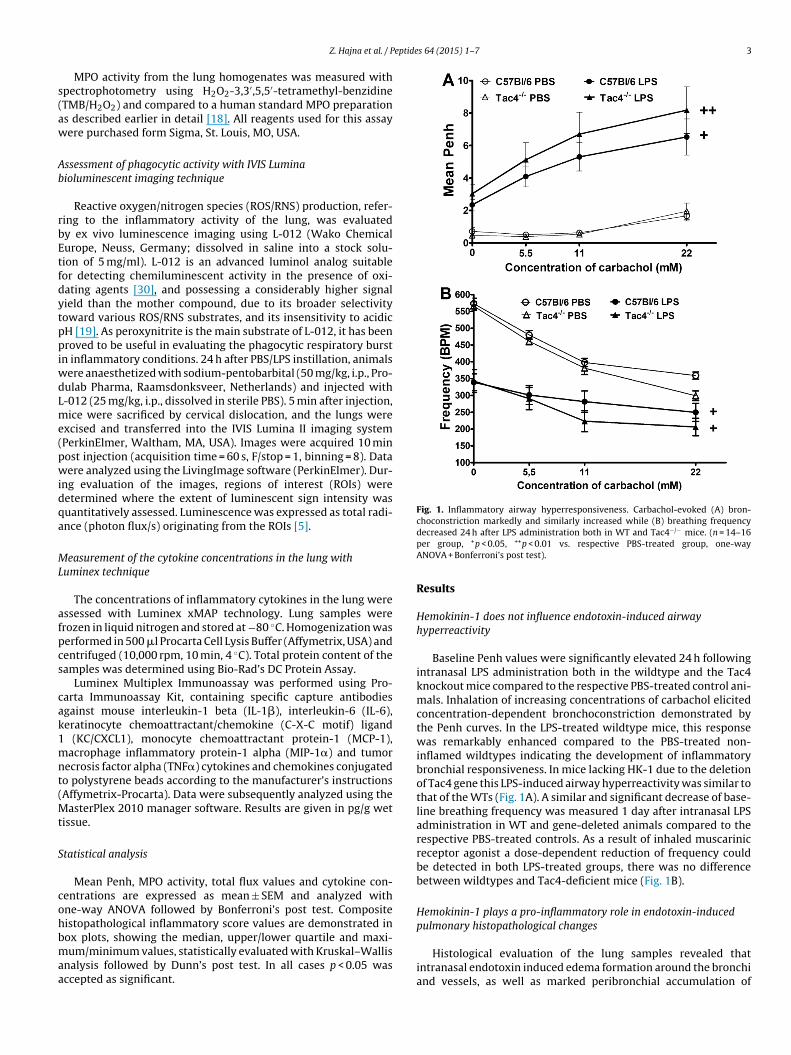
eptides 64 (2015) 1–7 3
s(aw
Ab
rbEtfdytppiwdLme(pwidqa
ML
afpcs
cak1mnt(Mt
S
cohbmaa
Fig. 1. Inflammatory airway hyperresponsiveness. Carbachol-evoked (A) bron-choconstriction markedly and similarly increased while (B) breathing frequency
−/−
Z. Hajna et al. / P
MPO activity from the lung homogenates was measured withpectrophotometry using H2O2-3,3′,5,5′-tetramethyl-benzidineTMB/H2O2) and compared to a human standard MPO preparations described earlier in detail [18]. All reagents used for this assayere purchased form Sigma, St. Louis, MO, USA.
ssessment of phagocytic activity with IVIS Luminaioluminescent imaging technique
Reactive oxygen/nitrogen species (ROS/RNS) production, refer-ing to the inflammatory activity of the lung, was evaluatedy ex vivo luminescence imaging using L-012 (Wako Chemicalurope, Neuss, Germany; dissolved in saline into a stock solu-ion of 5 mg/ml). L-012 is an advanced luminol analog suitableor detecting chemiluminescent activity in the presence of oxi-ating agents [30], and possessing a considerably higher signalield than the mother compound, due to its broader selectivityoward various ROS/RNS substrates, and its insensitivity to acidicH [19]. As peroxynitrite is the main substrate of L-012, it has beenroved to be useful in evaluating the phagocytic respiratory burst
n inflammatory conditions. 24 h after PBS/LPS instillation, animalsere anaesthetized with sodium-pentobarbital (50 mg/kg, i.p., Pro-ulab Pharma, Raamsdonksveer, Netherlands) and injected with-012 (25 mg/kg, i.p., dissolved in sterile PBS). 5 min after injection,ice were sacrificed by cervical dislocation, and the lungs were
xcised and transferred into the IVIS Lumina II imaging systemPerkinElmer, Waltham, MA, USA). Images were acquired 10 minost injection (acquisition time = 60 s, F/stop = 1, binning = 8). Dataere analyzed using the LivingImage software (PerkinElmer). Dur-
ng evaluation of the images, regions of interest (ROIs) wereetermined where the extent of luminescent sign intensity wasuantitatively assessed. Luminescence was expressed as total radi-nce (photon flux/s) originating from the ROIs [5].
easurement of the cytokine concentrations in the lung withuminex technique
The concentrations of inflammatory cytokines in the lung weressessed with Luminex xMAP technology. Lung samples wererozen in liquid nitrogen and stored at −80 ◦C. Homogenization waserformed in 500 �l Procarta Cell Lysis Buffer (Affymetrix, USA) andentrifuged (10,000 rpm, 10 min, 4 ◦C). Total protein content of theamples was determined using Bio-Rad’s DC Protein Assay.
Luminex Multiplex Immunoassay was performed using Pro-arta Immunoassay Kit, containing specific capture antibodiesgainst mouse interleukin-1 beta (IL-1�), interleukin-6 (IL-6),eratinocyte chemoattractant/chemokine (C-X-C motif) ligand
(KC/CXCL1), monocyte chemoattractant protein-1 (MCP-1),acrophage inflammatory protein-1 alpha (MIP-1�) and tumor
ecrosis factor alpha (TNF�) cytokines and chemokines conjugatedo polystyrene beads according to the manufacturer’s instructionsAffymetrix-Procarta). Data were subsequently analyzed using the
asterPlex 2010 manager software. Results are given in pg/g wetissue.
tatistical analysis
Mean Penh, MPO activity, total flux values and cytokine con-entrations are expressed as mean ± SEM and analyzed withne-way ANOVA followed by Bonferroni’s post test. Compositeistopathological inflammatory score values are demonstrated in
ox plots, showing the median, upper/lower quartile and maxi-um/minimum values, statistically evaluated with Kruskal–Wallisnalysis followed by Dunn’s post test. In all cases p < 0.05 wasccepted as significant.
decreased 24 h after LPS administration both in WT and Tac4 mice. (n = 14–16per group, +p < 0.05, ++p < 0.01 vs. respective PBS-treated group, one-wayANOVA + Bonferroni’s post test).
Results
Hemokinin-1 does not influence endotoxin-induced airwayhyperreactivity
Baseline Penh values were significantly elevated 24 h followingintranasal LPS administration both in the wildtype and the Tac4knockout mice compared to the respective PBS-treated control ani-mals. Inhalation of increasing concentrations of carbachol elicitedconcentration-dependent bronchoconstriction demonstrated bythe Penh curves. In the LPS-treated wildtype mice, this responsewas remarkably enhanced compared to the PBS-treated non-inflamed wildtypes indicating the development of inflammatorybronchial responsiveness. In mice lacking HK-1 due to the deletionof Tac4 gene this LPS-induced airway hyperreactivity was similar tothat of the WTs (Fig. 1A). A similar and significant decrease of base-line breathing frequency was measured 1 day after intranasal LPSadministration in WT and gene-deleted animals compared to therespective PBS-treated controls. As a result of inhaled muscarinicreceptor agonist a dose-dependent reduction of frequency couldbe detected in both LPS-treated groups, there was no differencebetween wildtypes and Tac4-deficient mice (Fig. 1B).
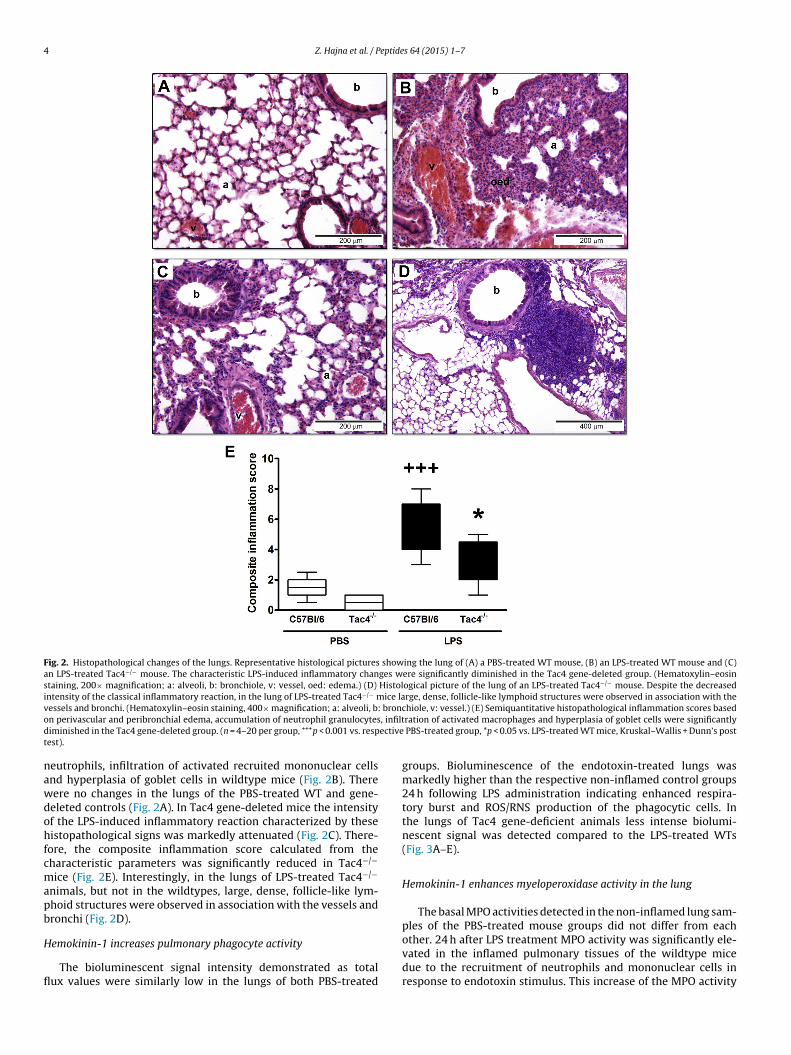
Hemokinin-1 plays a pro-inflammatory role in endotoxin-inducedpulmonary histopathological changes
Histological evaluation of the lung samples revealed thatintranasal endotoxin induced edema formation around the bronchiand vessels, as well as marked peribronchial accumulation of
4 Z. Hajna et al. / Peptides 64 (2015) 1–7
Fig. 2. Histopathological changes of the lungs. Representative histological pictures showing the lung of (A) a PBS-treated WT mouse, (B) an LPS-treated WT mouse and (C)an LPS-treated Tac4−/− mouse. The characteristic LPS-induced inflammatory changes were significantly diminished in the Tac4 gene-deleted group. (Hematoxylin–eosinstaining, 200× magnification; a: alveoli, b: bronchiole, v: vessel, oed: edema.) (D) Histological picture of the lung of an LPS-treated Tac4−/− mouse. Despite the decreasedintensity of the classical inflammatory reaction, in the lung of LPS-treated Tac4−/− mice large, dense, follicle-like lymphoid structures were observed in association with thev brono , infiltd ectivet
nawdohfcmapb
H
fl
essels and bronchi. (Hematoxylin–eosin staining, 400× magnification; a: alveoli, b:n perivascular and peribronchial edema, accumulation of neutrophil granulocytesiminished in the Tac4 gene-deleted group. (n = 4–20 per group, +++p < 0.001 vs. respest).
eutrophils, infiltration of activated recruited mononuclear cellsnd hyperplasia of goblet cells in wildtype mice (Fig. 2B). Thereere no changes in the lungs of the PBS-treated WT and gene-eleted controls (Fig. 2A). In Tac4 gene-deleted mice the intensityf the LPS-induced inflammatory reaction characterized by theseistopathological signs was markedly attenuated (Fig. 2C). There-
ore, the composite inflammation score calculated from theharacteristic parameters was significantly reduced in Tac4−/−
ice (Fig. 2E). Interestingly, in the lungs of LPS-treated Tac4−/−
nimals, but not in the wildtypes, large, dense, follicle-like lym-hoid structures were observed in association with the vessels andronchi (Fig. 2D).
emokinin-1 increases pulmonary phagocyte activity
The bioluminescent signal intensity demonstrated as totalux values were similarly low in the lungs of both PBS-treated
chiole, v: vessel.) (E) Semiquantitative histopathological inflammation scores basedration of activated macrophages and hyperplasia of goblet cells were significantly
PBS-treated group, *p < 0.05 vs. LPS-treated WT mice, Kruskal–Wallis + Dunn’s post
groups. Bioluminescence of the endotoxin-treated lungs wasmarkedly higher than the respective non-inflamed control groups24 h following LPS administration indicating enhanced respira-tory burst and ROS/RNS production of the phagocytic cells. Inthe lungs of Tac4 gene-deficient animals less intense biolumi-nescent signal was detected compared to the LPS-treated WTs(Fig. 3A–E).
Hemokinin-1 enhances myeloperoxidase activity in the lung
The basal MPO activities detected in the non-inflamed lung sam-ples of the PBS-treated mouse groups did not differ from each
other. 24 h after LPS treatment MPO activity was significantly ele-vated in the inflamed pulmonary tissues of the wildtype micedue to the recruitment of neutrophils and mononuclear cells inresponse to endotoxin stimulus. This increase of the MPO activity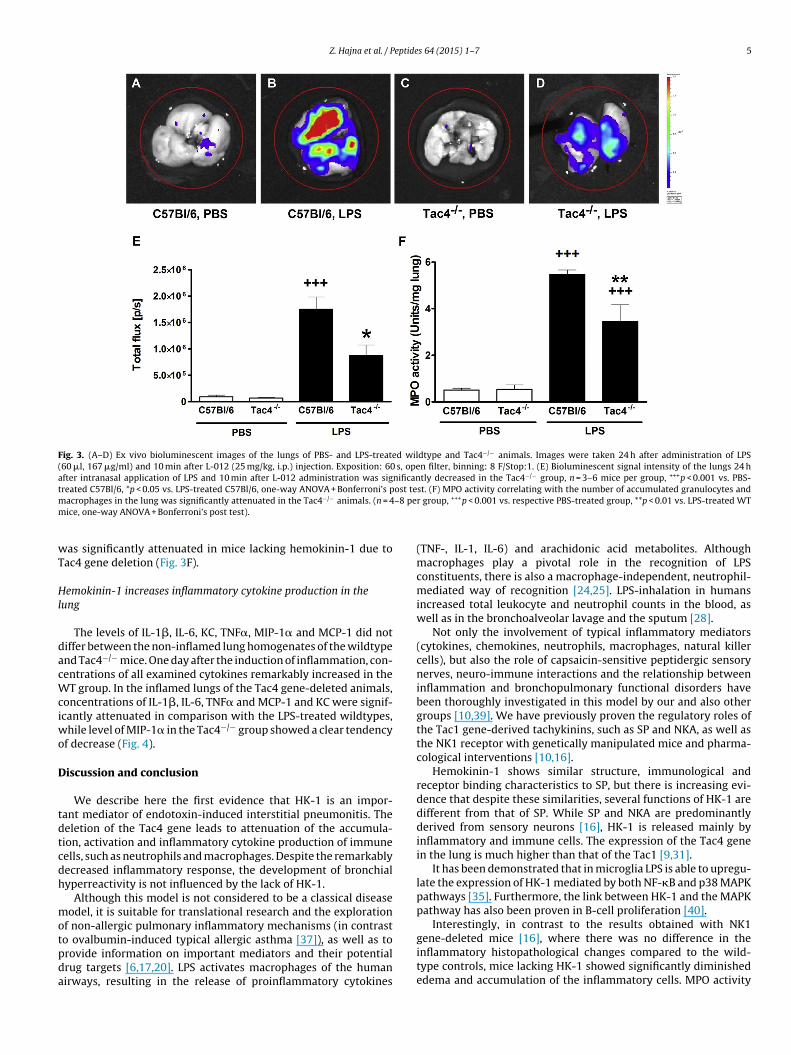
Z. Hajna et al. / Peptides 64 (2015) 1–7 5
Fig. 3. (A–D) Ex vivo bioluminescent images of the lungs of PBS- and LPS-treated wildtype and Tac4−/− animals. Images were taken 24 h after administration of LPS(60 �l, 167 �g/ml) and 10 min after L-012 (25 mg/kg, i.p.) injection. Exposition: 60 s, open filter, binning: 8 F/Stop:1. (E) Bioluminescent signal intensity of the lungs 24 hafter intranasal application of LPS and 10 min after L-012 administration was significantly decreased in the Tac4−/− group, n = 3–6 mice per group, +++p < 0.001 vs. PBS-treated C57Bl/6, *p < 0.05 vs. LPS-treated C57Bl/6, one-way ANOVA + Bonferroni’s post test. (F) MPO activity correlating with the number of accumulated granulocytes andm –8 pem
wT
Hl
dacWciwo
D
tdtcdh
motpda
acrophages in the lung was significantly attenuated in the Tac4−/− animals. (n = 4ice, one-way ANOVA + Bonferroni’s post test).
as significantly attenuated in mice lacking hemokinin-1 due toac4 gene deletion (Fig. 3F).
emokinin-1 increases inflammatory cytokine production in theung
The levels of IL-1�, IL-6, KC, TNF�, MIP-1� and MCP-1 did notiffer between the non-inflamed lung homogenates of the wildtypend Tac4−/− mice. One day after the induction of inflammation, con-entrations of all examined cytokines remarkably increased in the
T group. In the inflamed lungs of the Tac4 gene-deleted animals,oncentrations of IL-1�, IL-6, TNF� and MCP-1 and KC were signif-cantly attenuated in comparison with the LPS-treated wildtypes,
hile level of MIP-1� in the Tac4−/− group showed a clear tendencyf decrease (Fig. 4).
iscussion and conclusion
We describe here the first evidence that HK-1 is an impor-ant mediator of endotoxin-induced interstitial pneumonitis. Theeletion of the Tac4 gene leads to attenuation of the accumula-ion, activation and inflammatory cytokine production of immuneells, such as neutrophils and macrophages. Despite the remarkablyecreased inflammatory response, the development of bronchialyperreactivity is not influenced by the lack of HK-1.
Although this model is not considered to be a classical diseaseodel, it is suitable for translational research and the exploration
f non-allergic pulmonary inflammatory mechanisms (in contrast
o ovalbumin-induced typical allergic asthma [37]), as well as torovide information on important mediators and their potentialrug targets [6,17,20]. LPS activates macrophages of the humanirways, resulting in the release of proinflammatory cytokinesr group, +++p < 0.001 vs. respective PBS-treated group, **p < 0.01 vs. LPS-treated WT
(TNF-, IL-1, IL-6) and arachidonic acid metabolites. Althoughmacrophages play a pivotal role in the recognition of LPSconstituents, there is also a macrophage-independent, neutrophil-mediated way of recognition [24,25]. LPS-inhalation in humansincreased total leukocyte and neutrophil counts in the blood, aswell as in the bronchoalveolar lavage and the sputum [28].
Not only the involvement of typical inflammatory mediators(cytokines, chemokines, neutrophils, macrophages, natural killercells), but also the role of capsaicin-sensitive peptidergic sensorynerves, neuro-immune interactions and the relationship betweeninflammation and bronchopulmonary functional disorders havebeen thoroughly investigated in this model by our and also othergroups [10,39]. We have previously proven the regulatory roles ofthe Tac1 gene-derived tachykinins, such as SP and NKA, as well asthe NK1 receptor with genetically manipulated mice and pharma-cological interventions [10,16].
Hemokinin-1 shows similar structure, immunological andreceptor binding characteristics to SP, but there is increasing evi-dence that despite these similarities, several functions of HK-1 aredifferent from that of SP. While SP and NKA are predominantlyderived from sensory neurons [16], HK-1 is released mainly byinflammatory and immune cells. The expression of the Tac4 genein the lung is much higher than that of the Tac1 [9,31].
It has been demonstrated that in microglia LPS is able to upregu-late the expression of HK-1 mediated by both NF-�B and p38 MAPKpathways [35]. Furthermore, the link between HK-1 and the MAPKpathway has also been proven in B-cell proliferation [40].
Interestingly, in contrast to the results obtained with NK1
gene-deleted mice [16], where there was no difference in theinflammatory histopathological changes compared to the wild-type controls, mice lacking HK-1 showed significantly diminishededema and accumulation of the inflammatory cells. MPO activity
6 Z. Hajna et al. / Peptides 64 (2015) 1–7
F centraL differ+ one-w
dmodMmr
meriei
tiirdb[
nitivas
istpIniTtbts
ig. 4. Concentrations of the inflammatory cytokines in the lung homogenates. ConPS-treated Tac4 gene-deficient mice compared to the WTs. MIP-1� levels did not
++p < 0.001 vs. PBS-treated C57Bl/6, **p < 0.01, ***p < 0.001 vs. LPS-treated C57Bl/6,
ue to the activation of neutrophils and macrophages was alsoarkedly attenuated in the Tac4 knockout animals, whereas
ur previous results revealed no difference between NK1 gene-eficient and wildtypes [16]. The parallel outcomes of histology andPO activity indicate, that in the LPS-induced pulmonary inflam-atory reaction HK-1 plays a crucial role, but not through the NK1
eceptor.In accordance with these results, in the CFA-induced murine
odel of chronic arthritis, our recent study showed that the periph-ral inflammatory processes in the joints are independent of NK1eceptor activation, while the consequent mechanical hyperalgesias NK1 receptor-mediated [4]. These results strongly support thexistence of another, specific receptor for HK-1 that mediatesnflammatory actions at the periphery [4,11].
Interestingly, the formation of large, dense, follicle-like struc-ures associated to the vessels and bronchi were only observedn the lungs of LPS-treated Tac4−/− animals. This suggests thatn endotoxin-induced pneumonitis HK-1 also contributes to theegulated activation of lymphocytes under pathophysiological con-itions. This first in vivo observation in the airways is supportedy the earlier described B-lymphopoesis promoting effect of HK-12,43].
During endotoxin-induced lung injury mainly neutrophil gra-ulocytes are recruited besides macrophages [36]. Therefore, it
s important to consider that not only the accumulation of neu-rophils and macrophages, but also their activation was attenuatedn Tac4−/− animals, as shown by MPO activity measurement and exivo bioluminescent imaging of ROS. These latter parameters show
perfect correlation in our model, therefore, the bioluminescentignal in the lung is likely to be derived from the MPO activity.
The LPS-induced inflammatory reaction is classically character-zed by the production and release of proinflammatory cytokines,uch as TNF�, IL-1�, IL-6 and KC [17]. The cytokines examined inhis study (IL-1�, Il-6, KC, TNF�, MIP-1� and MCP-1) are mainlyroduced by activated macrophages and monocytes. Besides, IL-1,
L-6, KC and MIP-1� are described to be involved in the activation ofeutrophils [8,26]. Moreover, upon endotoxin stimulation, MIP-1�
ncreases proinflammatory cytokine synthesis, such as IL-1, Il-6 andNF� [12], leading to the augmentation of the reaction. The interac-
ion between HK-1 and the cytokine system (IL-1� and TNF�) haseen suggested earlier [3]. In accordance with these data we foundhat the LPS-induced elevation of the inflammatory cytokines wasignificantly lower in Tac4−/− mice. These results further indicatetions of IL-1�, IL-6, KC, TNF� and MCP-1 were significantly reduced in the lungs of in the two examined groups of animals. (n = 4 mice per group, +p < 0.05, ++p < 0.01,ay ANOVA + Bonferroni’s post test).
that HK-1 plays a crucial role in cytokine release and consequentimmune cell activation during pulmonary inflammation.
In contrast to SP and NKA, HK-1 had no influence over eitherbreathing frequency or bronchial hyperreactivity. Although somedata show that hHK-1 evokes bronchoconstriction [14], in thismouse model we could not confirm a similar effect for HK-1. Oneexplanation for this contradiction is that the human HK-like pep-tides make up a probably more complicated system than it is inmice. The human TAC4 gene has 4 different splice variants, andtheir sequences are different from its murine counterparts. Theyshow different expression characteristics: � and �TAC4 are consti-tutively, while �TAC4 and �TAC4 are only ubiquitously expressed indifferent tissues [14]. Not only the structures, but also the receptoraffinities are different in cases of the mouse and human peptides,EKC and EKD have low affinities to all presently known NK receptors[32]. Another explanation is that as our previous results suggested[10,16], predominantly the NK2 receptor is responsible for inflam-matory bronchoconstriction in mice and HK-1 has a markedly loweraffinity to this than to the NK1 receptor [1].
In summary, we demonstrate here the first data for an impor-tant mediator role of HK-1 in airway inflammation. It is involved inLPS-evoked perivascular edema formation, neutrophil accumula-tion and activation, as well as macrophage influx into the lung, butit does not influence bronchial hyperresponsiveness. Furthermore,HK-1 seems to have a specific regulatory role on lymphocyte func-tions in this type of inflammatory process, its precise mechanismsneed to be further investigated.
Acknowledgements
The present scientific contribution is dedicated to the 650thanniversary of the foundation of the University of Pécs, Hungary.
This study was supported by Developing Competitiveness ofUniversities in the South Transdanubian Region “SROP-4.2.2.A-11/1/KONV-2012-0024”.
The authors are grateful to Anikó Perkecz for her expert techni-cal help in histological processing.
References
[1] Bellucci F, Carini F, Catalani C, Cucchi P, Lecci A, Meini S, et al. Pharmacolog-ical profile of the novel mammalian tachykinin, hemokinin 1. Br J Pharmacol2002;135:266–74.

eptide
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
deficient mice exposed to bacterial lipopolysaccharide. Am J Respir Cell Mol
Z. Hajna et al. / P
[2] Berger A, Benveniste P, Corfe SA, Tran AH, Barbara M, Wakeham A, et al. Tar-geted deletion of the tachykinin 4 gene (TAC4−/−) influences the early stagesof B lymphocyte development. Blood 2010;116:3792–801.
[3] Berger A, Tran AH, Paige CJ. Co-regulated decrease of neurokinin-1 receptor andHemokinin-1 gene expression in monocytes and macrophages after activationwith pro-inflammatory cytokines. J Neuroimmunol 2007;187:83–93.
[4] Borbély E, Hajna Z, Sándor K, Kereskai L, Tóth I, Pintér E, et al. Role oftachykinin 1 and 4 gene-derived neuropeptides and the neurokinin 1 receptorin adjuvant-induced chronic arthritis of the mouse. PLOS ONE 2013;8:e61684,http://dx.doi.org/10.1371/journal.pone.0061684.
[5] Botz B, Bölcskei K, Kereskai L, Kovács M, Németh T, Szigeti K, et al.Differential regulatory role of pituitary adenylate-cyclase activating polypep-tide in the serum-transfer-induced arthritis model. Arthritis Rheumatol2014;66:2739–50, http://dx.doi.org/10.1002/art.38772.
[6] Chen H, Bai C, Wang X. The value of the lipopolysaccharide-induced acute lunginjury model in respiratory medicine. Expert Rev Respir Med 2010;4:773–83.
[7] Cunin P, Caillon A, Corvaisier M, Garo E, Scotet M, Blanchard S, et al. Thetachykinins substance P and hemokinin-1 favor the generation of humanmemory Th17 cells by inducing IL-1�, IL-23, and TNF-like 1A expression bymonocytes. J Immunol 2011;186:4175–82.
[8] Deshmane SL, Kremlev S, Amini S, Sawaya BE. Monocyte chemoattractantprotein-1 (MCP-1): an overview. J Interferon Cytokine Res 2009;29:313–26.
[9] Duffy RA, Hedrick JA, Randolph G, Morgan CA, Cohen-Williams ME, VassilevaG, et al. Centrally administered hemokinin-1 (HK-1), a neurokinin NK1 recep-tor agonist, produces substance P-like behavioral effects in mice and gerbils.Neuropharmacology 2003;45:242–50.
10] Elekes K, Helyes Z, Németh J, Sándor K, Pozsgai G, Kereskai L, et al. Role ofcapsaicin-sensitive afferents and sensory neuropeptides in endotoxin-inducedairway inflammation and consequent bronchial hyperreactivity in the mouse.Regul Pept 2007;141:44–54.
11] Endo D, Ikeda T, Ishida Y, Yoshioka D, Nishimori T. Effect of intrathecal admin-istration of hemokinin-1 on the withdrawal response to noxious thermalstimulation of the rat hind paw. Neurosci Lett 2006;392:114–7.
12] Fahey 3rd TJ, Tracey KJ, Tekamp-Olson P, Cousens LS, Jones WG, Shires GT,et al. Macrophage inflammatory protein 1 modulates macrophage function. JImmunol 1992;148:2764–9.
13] Grassin-Delyle S, Buenestado A, Vallat L, Naline E, Marx S, Decocq J, et al.Expression and proliferative effect of hemokinin-1 in human B-cells. Peptides2011;32:1027–34, http://dx.doi.org/10.1016/j.peptides.2011.02.012.
14] Grassin-Delyle S, Naline E, Buenestado A, Risse PA, Sage E, Advenier C, et al.Expression and function of human hemokinin-1 in human and guinea pig air-ways. Respir Res 2010;11:139.
15] Haddad EB, Birrell M, McCluskie K, Ling A, Webber SE, Foster ML, et al. Role ofp38 MAP kinase in LPS-induced airway inflammation in the rat. Br J Pharmacol2001;132:1715–24.
16] Helyes Z, Elekes K, Sándor K, Szitter I, Kereskai L, Pintér E, et al. Involve-ment of preprotachykinin A gene-encoded peptides and the neurokinin 1receptor in endotoxin-induced murine airway inflammation. Neuropeptides2010;44:399–406, http://dx.doi.org/10.1016/j.npep.2010.05.004.
17] Helyes Z, Hajna Z. Endotoxin-induced airway inflammation and asthma models.In: Szallasi A, Bíró T, editors. TRP channels in drug discovery. Methods in phar-macology and toxicology, vol. 1. 2012., ISBN 978-1-62703-077-9 p. 301–42,http://dx.doi.org/10.1007/978-1-62703-077-9 16.
18] Helyes Z, Pintér E, Sándor K, Elekes K, Bánvölgyi A, Keszthelyi D, et al. Impaireddefense mechanism against inflammation, hyperalgesia, and airway hyperre-activity in somatostatin 4 receptor gene-deleted mice. Proc Natl Acad Sci U S A2009;106:13088–93, http://dx.doi.org/10.1073/pnas.0900681106.
19] Kielland A, Blom T, Nandakumar KS, Holmdahl R, Blomhoff R, Carlsen H. In vivoimaging of reactive oxygen and nitrogen species in inflammation using theluminescent probe L-012. Free Radic Biol Med 2009;47:760–6.
20] Kips JC, Anderson GP, Fredberg JJ, Herz U, Inman MD, Jordana M, et al. Murine
models of asthma. Eur Respir J 2003;22:374–82.21] Klassert TE, Sánchez JJ, Almeida TA, Candenas L, Pinto F, Acosta O, et al.Common variants of the neuropeptide expressing tachykinin genes and sus-ceptibility to asthma: a case–control study. J Neuroimmunol 2010;227:202–7,http://dx.doi.org/10.1016/j.jneuroim.2010.05.044.
[
s 64 (2015) 1–7 7
22] Kurtz MM, Wang R, Clements MK, Cascieri MA, Austin CP, Cunningham BR, et al.Identification, localization and receptor characterization of novel mammaliansubstance P-like peptides. Gene 2002;296:205–12.
23] Maggi CA. Tachykinins and calcitonin gene-related peptide (CGRP) as cotrans-mitters released from peripheral endings of sensory nerves. Prog Neurobiol1995;45:1–98.
24] Marasco WA, Phan SH, Krutzsch H, Showell HJ, Feltner DE, Nairn R, et al.Purification and identification of formyl-methionyl-leucyl-phenylalanine asthe major peptide neutrophil chemotactic factor produced by Escherichia coli.J Biol Chem 1984;259:5430–9.
25] Martin TR. Recognition of bacterial endotoxin in the lungs. Am J Respir Cell MolBiol 2000;23:128–32.
26] Menten P, Wuyts A, Van Damme J. Macrophage inflammatory protein-1.Cytokine Growth Factor Rev 2002;13:455–81.
27] Metwali A, Blum AM, Elliott DE, Setiawan T, Weinstock JV. Cutting edge:hemokinin has substance P-like function and expression in inflammation. JImmunol 2004;172:6528–32.
28] Michel O. Systemic and local airways inflammatory response to endotoxin.Toxicology 2000;152:25–30.
29] Nichols BA, Bainton DF. Differentiation of human monocytes in bone mar-row and blood. Sequential formation of two granule populations. Lab Invest1973;29:27–40.
30] Nishinaka Y, Aramaki Y, Yoshida H, Masuya H, Sugawara T, Ichimori Y. A newsensitive chemiluminescence probe, L-012, for measuring the production ofsuperoxide anion by cells. Biochem Biophys Res Commun 1993;193:554–9.
31] Page NM. Characterization of the gene structures, precursor processing andpharmacology of the endokinin peptides. Vasc Pharmacol 2006;45:200–8.
32] Page NM. New challenges in the study of the mammalian tachykinins. Peptides2005;26:1356–68.
33] Pintér E, Pozsgai G, Hajna Z, Helyes Z, Szolcsányi J. Neuropeptide receptors aspotential drug targets in the treatment of inflammatory conditions. Br J ClinPharmacol 2014;77:5–20.
34] Rodrigues MR, Rodriguez D, Russo M, Campa A. Macrophage activation includeshigh intracellular myeloperoxidase activity. Biochem Biophys Res Commun2002;292:869–73.
35] Sakai A, Takasu K, Sawada M, Suzuki H. Hemokinin-1 gene expres-sion is upregulated in microglia activated by lipopolysaccharide throughNF-�B and p38 MAPK signaling pathways. PLoS ONE 2012;7:e32268,http://dx.doi.org/10.1371/journal.pone.0032268.
36] Savov JD, Gavett SH, Brass DM, Costa DL, Schwartz DA. Neutrophils play a criticalrole in development of LPS-induced airway disease. Am J Physiol Lung Cell MolPhysiol 2002;283:952–62.
37] Schymeinsky J, Mayer H, Tomsic C, Tilp C, Schuetz JD, Cui Y, et al. The absence ofmrp4 has no effect on the recruitment of neutrophils and eosinophils into thelung after LPS, cigarette smoke or allergen challenge. PLoS ONE 2013;8:e61193,http://dx.doi.org/10.1371/journal.pone.0061193.
38] Sunakawa N, Naono R, Ikeda T, Matsushima O, Sakoda S, NishimoriT. The amino-terminal region of hemokinin-1 regulates the induc-tion of thermal hyperalgesia in rats. Neuropeptides 2010;44:273–8,http://dx.doi.org/10.1016/j.npep.2010.01.003.
39] Takeda M, Miyake M, Muto T, Kamijima M, Sakamoto T. Proliferation of sensoryC-fibers and subsequent neurogenic inflammation in rat airway induced byinhaled lipopolysaccharide. Neurotoxicology 2011;32:954–62.
40] Wang W, Li Q, Zhang J, Wu H, Yin Y, Ge Q, et al. Hemokinin-1 activates theMAPK pathway and enhances B cell proliferation and antibody production. JImmunol 2010;184:3590–7.
41] Watanabe C, Mizoguchi H, Yonezawa A, Sakurada S. Characterization ofintrathecally administered hemokinin-1-induced nociceptive behaviors inmice. Peptides 2010;31:1613–6, http://dx.doi.org/10.1016/j.peptides.
42] Zeldin DC, Wohlford-Lenane C, Chulada P, Bradbury JA, Scarborough PE, RoggliV, et al. Airway inflammation and responsiveness in prostaglandin H synthase-
Biol 2001;25:457–65.43] Zhang Y, Lu L, Furlonger C, Wu GE, Paige CJ. Hemokinin is a hematopoietic-
specific tachykinin that regulates B lymphopoiesis. Nat Immunol2000;1:392–7.




















