Autoinducer AI2 Is Involved in Regulating a Variety of Cellular Processes in Salmonella Typhimurium
7
FOODBORNE PATHOGENS AND DISEASE Volume 5, Number 2, 2008 © Mary Ann Liebert, Inc. DOl: 10.1 089/fpd.2007.0050 Autoinducer Al-2 Is Involved in Regulating a Variety of Cellular Processes in Salmonella Typhimurium Kamlesh A. Soni, 1 Palmy A. Jesudhasan, 1 Martha Cepeda, 1 Brad Williams, 3 Michael Hume, 1,2 William K. Russell , 3 Arul Jayaraman, 4 and Suresh D. Pillai1 Abstract Salmonella Typhimurium is known to exhibit LuxS/AI-2--mediated cell signaling. We investigated the role of LuxS/AI-2 system on Salmonella Typhimuriuni protein expression using a proteomics approach based on two-dimensional gel electrophoresis (2DGE)—MALDI-MS. The global protein expression profiles of the wild-type, a luxS mutant, and a luxS mutant strain supplemented with AI-2 were compared. Seven pro- teins were differentially expressed when comparing the wild-type and luxS mutant strains, whereas 13 proteins were differentially expressed when comparisons were made between loiS mutant strains with and without Al-2 supplementation. The seven proteins that were differentially expressed between the wild- type and the loiS mutant strain were also differentially expressed in the loiS mutant strain supplemented with AI-2. The level of PhoP, a virulence determinant, was higher in the presence of AI-2. Proteins associated with the carbohydrate metabolism (pflcA, gpml, and taiB) and ATP synthesis (I'tn gene product) were up-regulated by the presence of A1-2 molecules. These results provide experimental evidence that Al-2 molecules regulate a variety of cellular processes in Salmonella Typhirnurium. Introduction A UTOINDUCER (Al) MOLECULES such as Al-I, AI-2, and AI-3 are thought to influence a variety of bacterial processes including patho- genicity, biofilms, motility, and biolumines- cence (Surette and Bassler, 1998; Arevalo-Ferro et al., 2003; Henke and Bassler, 2004; Gonzalez etal., 2006; Pillai and Jesudhasan, 2007). Optimal levels of these cell signaling molecules and in- teraction of these molecules with regulatory proteins are thought to be key steps involved in the coordination of gene expression. Among these different autoiriducer molecules, AI-2 is synthesized by LuxS synthase. The IuxS gene has been identified in more than 55 species of gram-negative and gram-positive bacteria, in- cluding Salmonella Typh imurium (Xavier and Bassler, 2003). The loiS gene is involved in the synthesis of AI-2 molecules by catalyzing the reaction from S-ribosyl homocysteine to homo- cystein and 4,5-d ihydroxy-2,3-pentanedione (DPD). Reaction of these unstable DPD mole- cules with water leads to cyclation and conse- quent formation of furanones known as AI-2 molecules (Schauder et al., 2001). The AI-2 molecules are thought to exist in two different forms depending on the presence or absence of boron molecules (Chen et al., 2002; Miller et al., 2004). 'Food Safety & Environmental Microbiology Program, Departments of Poultry Science, and Nutrition and Food Science, Texas A&M University, College Station, Texas. 2 USDA, ARS, SPARC, Food and Feed Safety Research Unit, College Station, Texas. 'Laborator y for Biological Mass Spectrometry, Department of Chemistry, Texas A&M University, College Station, Texas. 4 Atrie McFerrin Department of Chemical Engineering, Texas A&M University, College Station, Texas. 147
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Autoinducer AI2 Is Involved in Regulating a Variety of Cellular Processes in Salmonella Typhimurium

FOODBORNE PATHOGENS AND DISEASEVolume 5, Number 2, 2008© Mary Ann Liebert, Inc.DOl: 10.1 089/fpd.2007.0050
Autoinducer Al-2 Is Involved in Regulating a Varietyof Cellular Processes in Salmonella Typhimurium
Kamlesh A. Soni, 1 Palmy A. Jesudhasan, 1 Martha Cepeda, 1 Brad Williams, 3 Michael Hume, 1,2
William K. Russell , 3 Arul Jayaraman, 4 and Suresh D. Pillai1
Abstract
Salmonella Typhimurium is known to exhibit LuxS/AI-2--mediated cell signaling. We investigated the roleof LuxS/AI-2 system on Salmonella Typhimuriuni protein expression using a proteomics approach based ontwo-dimensional gel electrophoresis (2DGE)—MALDI-MS. The global protein expression profiles of thewild-type, a luxS mutant, and a luxS mutant strain supplemented with AI-2 were compared. Seven pro-teins were differentially expressed when comparing the wild-type and luxS mutant strains, whereas 13proteins were differentially expressed when comparisons were made between loiS mutant strains with andwithout Al-2 supplementation. The seven proteins that were differentially expressed between the wild-type and the loiS mutant strain were also differentially expressed in the loiS mutant strain supplementedwith AI-2. The level of PhoP, a virulence determinant, was higher in the presence of AI-2. Proteinsassociated with the carbohydrate metabolism (pflcA, gpml, and taiB) and ATP synthesis (I'tn gene product)were up-regulated by the presence of A1-2 molecules. These results provide experimental evidence thatAl-2 molecules regulate a variety of cellular processes in Salmonella Typhirnurium.
Introduction
A
UTOINDUCER (Al) MOLECULES such as Al-I,AI-2, and AI-3 are thought to influence a
variety of bacterial processes including patho-genicity, biofilms, motility, and biolumines-cence (Surette and Bassler, 1998; Arevalo-Ferroet al., 2003; Henke and Bassler, 2004; Gonzalezetal., 2006; Pillai and Jesudhasan, 2007). Optimallevels of these cell signaling molecules and in-teraction of these molecules with regulatoryproteins are thought to be key steps involved inthe coordination of gene expression. Amongthese different autoiriducer molecules, AI-2 issynthesized by LuxS synthase. The IuxS gene
has been identified in more than 55 species ofgram-negative and gram-positive bacteria, in-cluding Salmonella Typh imurium (Xavier andBassler, 2003). The loiS gene is involved in thesynthesis of AI-2 molecules by catalyzing thereaction from S-ribosyl homocysteine to homo-cystein and 4,5-d ihydroxy-2,3-pentanedione(DPD). Reaction of these unstable DPD mole-cules with water leads to cyclation and conse-quent formation of furanones known as AI-2molecules (Schauder et al., 2001). The AI-2molecules are thought to exist in two differentforms depending on the presence or absence ofboron molecules (Chen et al., 2002; Miller et al.,2004).
'Food Safety & Environmental Microbiology Program, Departments of Poultry Science, and Nutrition and Food Science, TexasA&M University, College Station, Texas.
2USDA, ARS, SPARC, Food and Feed Safety Research Unit, College Station, Texas.'Laboratory for Biological Mass Spectrometry, Department of Chemistry, Texas A&M University, College Station, Texas.4 Atrie McFerrin Department of Chemical Engineering, Texas A&M University, College Station, Texas.
147

148 SONI ET AL.
Salmonella Typhimurium is a key foodbomepathogen (Gomez et al., 1997). While the role ofLuxS/Ai-2 system in different bacterial specieshas been associated with genes responsible forvirulence, motility, and biofilm formation, theonly known role of AI-2 in Salmonella Typhi-murium is the regulation of the lsr operon (Tagaet al., 2003). The objective of this work was toidentify the proteins that are influenced byLuxS/AI-2 in Salmonella Typhimurium usinga two-dimensional gel electrophoresis (2DGE)-based proteomic approach. Transcriptome anal-ysis with a DNA microarray can be used toidentify genes influenced by the autoinducermolecules. However, a microarray approachprovides information at the mRNA level onlyand not at the functional protein level (Kim et al.,2007). To the best of our knowledge this is thefirst report correlating the presence of AI-2 withprotein expression patterns and their relation-ship to cellular processes in Salmonella.
Materials and Methods
Preparation and quantification of Al-2 molecules
In vitro synthesized AI-2 was prepared andquantified as described previously (Sperandioet al., 2003). Briefly, His-tagged LuxS and Pfs(proteins required for the formation of Al-2)were isolated and purified using a nickel resincolumn (Qiagen Inc., Valencia, CA). The puri-fied enzymes were incubated with 1 mM S-adenosyl homocysteirie (Sigma-Aldrich, St.Louis, MO) for 1 hour at 37"C, and AI-2 wasfurther separated from the enzymes using theBiomax-5 (Millipore, Billerica, MA) column andthe AI-2 concentration was determined. Theconcentration of in vitro synthesized Al-2 was125 tM, and 20% (25 tM) of this AI-2 prepara-tion was used in experiments that involved theaddition of AI-2 molecules.
Bacterial strains and growth condition
Salmonella Typhimurium (Accession no. 87-26254, National Veterinary Service Laboratory,Ames, IA) and its isogenic luxS mutant (PJ 002)were used in this study. The luxS mutant strainwas created in our laboratory as described pre-viously (Widmer et al., 2007). Salmonella Ty-phimurium cells (wild-type and luxS mutant)
were cultured into Luria-Bertani (LB) mediumsupplemented with 0.5% glucose at 37C withmoderate agitation (100 rpm) until the late-log phase of growth was reached (optical den-sity [OD] 1.2). In vitro synthesized AI-2(25 jiM) was added to the LB broth from thebeginning of growth period. Vihrio linrveiistrain BB170 (luxN::Tn5 sensor 1-, sensor 2+),a reporter strain used to determine AI-2 ac-tivity, was grown at 30CC with aeration inautoinducer bioassay (AB) medium (Lu et al.,2004).
Al-2 bioassay to verify the inability of the IuxSmutant to produce Al-2
Overnight cultures of Salmonella Typhimur-ium (wild-type and luxS mutant) were inocu-lated (1:100) in fresh LB broth (0.5 0% glucose)and cultured at 37C with shaking (100 rpm).Optical densities (OD()nm) of these cultureswere noted and the cell-free supernatants (CFS)were collected by centrifugation (10,000 g for2 minutes) at different time points. The super-natants were then passed through 0.22-jim sy-ringe filters (Corning ® Inc., Corning, NY) andstored at —20'C until used in the AI-2 bioassay.The V. harveii reporter strain BBI70, whichproduces luminescence in the presence of AI-2molecules, was used as a bioassay reporterstrain to verify the lack of AI-2 production bythe luxS mutant strain. The AI-2 bioassay wasperformed as described earlier (Lu et al., 2004).Briefly, 90 jiL of the freshly diluted (1:5000)culture of the reporter strain in AB medium wasmixed with 10 jiL of test samples (CFS) or 10 j.iof AB media (negative control) in a 96-well plate.The plates were incubated at 30CC with moder-ate shaking (100 rpm) and the luminescence re-sponse of the reporter strains was monitoredusing a Wallac 1420 plate reader (PerkinElmer,Shelton, CT). The in vitro synthesized AI-2molecules were also verified using the similarbioassay to confirm their ability to induce theluminescence response.
Sample preparation
Late log-phase cultures of the wild-type(no. 87-26254), luxS mutant, and the luxS mu-tant grown in the presence of Al-2 were ob-tained. The cells were centrifuged (2 minutes at

PROTEOMIC ANALYSIS IN S. TYPI-IIMURIUM 149
12,000 g), and the pellets were resuspended inBPerx bacterial protein extraction reagents(Pierce, Rockford, IL) to collect the soluble pro-tein fractions. The Ready IM 2-D cleanup-kit (Bio-Rad, Hercules, CA) was used to removeionic contaminants and the resulting proteinpreparations were dissolved in 100 iL of rehy-dration buffer (9.5 M urea, 2% [w/v] CHAPS,18 mM DTT, 0.5% [v/v] amphyolytes, and onetablet of protease inhibitor [Roche Diagnostic,Mannheim, Germany]). The insoluble proteinsand cell debris were removed by centrifugation(2 minutes at 12,000 g).
2D gel electrophoresis
The Bradford assay (A580 nm) was used tomeasure the protein concentration (Pierce). Pre-liminary studies using immobilized pH gradi-ent (IPG) strips in the range of pH 3-10indicated that the majority of the soluble pro-teins were in the pH 4-7 range. Hence for sub-sequent analysis, IPG strips in the pH 4-7 rangewere employed. The protein samples were ad-justed to 35 tg in 125 tL of rehydration bufferfor the rehydration of 7-cm IPG strips (Bio-Rad).The isoelectric focusing of the 7-cm IPG stripswas performed at a linear voltage gradient withapproximately 15,000 final Vnhours in a ProteanIEF Cell (Bio-Rad). For spot excision, 13-cm IPGstrips (pH 4-7) with a total protein load of 800tg in a final volume of 250 iL (rehydration
buffer) were used. Equilibration of the IIGstrips was performed as previously mentioned(Arevalo-Ferro et al., 2003). The second dimen-sion gel electrophoresis was performed using10% (w/v) SDS-PAGE gel at 150V constantvoltage. The protein spots were visualized bystaining the gels with Sypro Ruby fluorescencestain (Molecular Probe, Eugene, OR).
Data analysis
Three independent 2DGE analyses (i.e., threebiological replications) were performed for eachof the three experimental treatments; namely,the wild-type strain, the luxS mutant strain, andthe luxS mutant strain grown in the presence ofAI-2. The 2D gels were scanned using Gel DocImage System (Bio-Rad) and the raw imageswere analyzed using the PDQuest' 2-D gelanalysis software version-8 (Bio-Rad). The data
was analyzed to achieve two different compar-isons, namely 1) protein profile of the wild-typecompared to protein profile of the !uxS mutant,and 2) protein profile of the liixS mutant com-pared to the protein profile of the !uxS mutantgrown in the presence of AI-2. Only those pro-tein spots that showed significant differences inintensities (i.e., ±1.5-fold change) were identi-fied using MALDI-MS (Arevalo-Ferro et al.,2005).
Protein identification using MALDI-MS
The differentially expressed protein spotswere manually excised (approximately 1 mm insize) and placed in a 96-well microtiter plate forproteolytic digestion. The proteolytic digestionswere performed overnight at 37C using trypsin(20 g/mL in 25 mM ammonium bicarbonate)(Promega, Madison, WI). The digests were spot-ted onto MALDI targets using a Pr 0MSTM robotcapable of sample cleanup prior to MALDI-MSanalysis (Genomic Solutions, Ann Arbor, MI).The MALDI-MS experiments were performedin a 4700 Proteomics Analyzer MALDI-TOF/TOF (Applied Biosystem, Foster City, CA).Twenty tandem MS spectra per spot were ac-quired. To confirm the generated MALDI-MSdata, five spots were reanalyzed.
Results
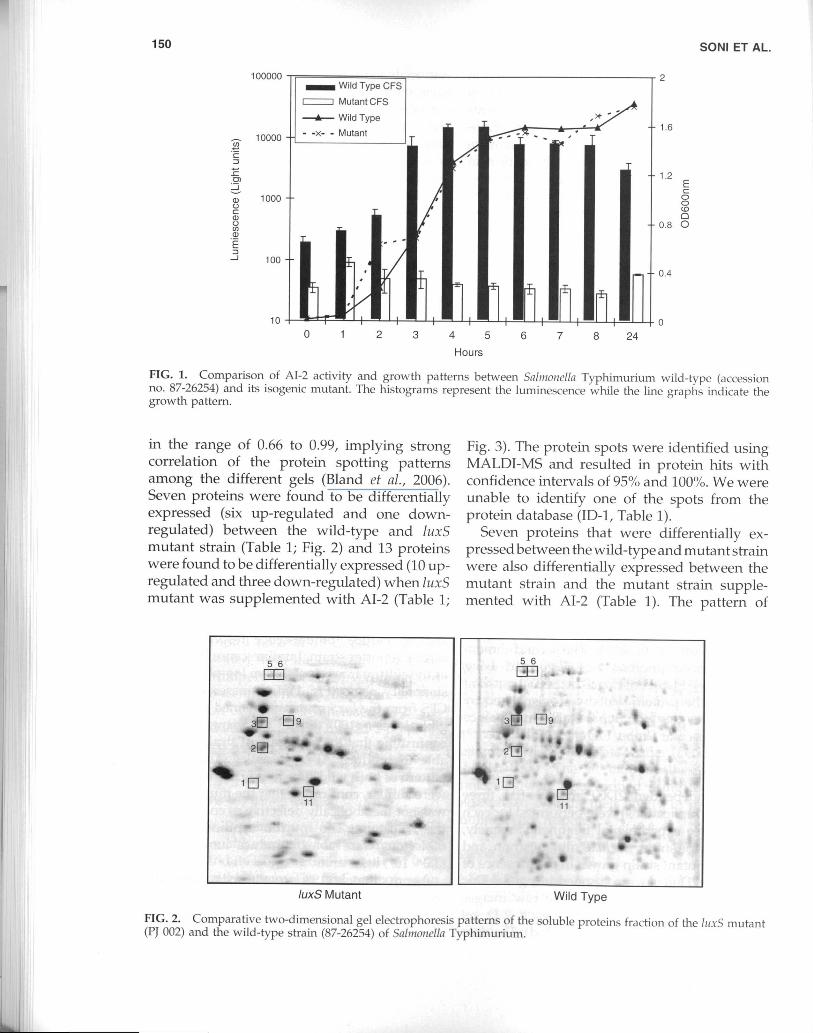
The A1-2 bioassay verified that CFS fromwild-type Salmonella Tvphimurium cells wereable to induce luminescence response in V.Jiarvc'ii reporter strain. Luminescence decreasedas the culture transitioned from log-phase to thestationary phase (Fig. 1). Luminescence by theCFS from the !uxS mutant remained at a negli-gible level (<100) during the growth period,confirming that the luxS mutant strain was un-able to produce AI-2 molecules. The growthpattern of both wild-type and !uxS mutant cellswere similar, confirming that the mutant strainwas not metabolically defective as compared tothe wild-type. The in vitro synthesized AI-2molecules were able to induce luminescence(3.0 x 10) light units.
Approximately 198 protein spots were de-tectable in the experimental gels (Figs. 2 and 3).The similarity in spotting patterns between in-dividual gels resulted in correlation coefficients
100000
10000Co=
-c0)
—J
10000
ci)0U)
E=—J100
10 0
2
1.6
1.2E=CCCDa
0.8 0
0.4
150
SONI ET AL.
I
01234567824Hours
FIG. 1. Comparison of AI-2 activity and growth patterns between Salmonella Typhimurium wild-type (accessionno. 87-26254) and its isogenic mutant. The histograms represent the luminescence while the line graphs indicate thegrowth pattern.
in the range of 0.66 to 0.99, implying strong Fig. 3). The protein spots were identified usingcorrelation of the protein spotting patterns MALDJ-MS and resulted in protein hits withamong the different gels (Bland et al., 2006). confidence intervals of 95% and 100%. We wereSeven proteins were found to be differentially unable to identify one of the spots from theexpressed (six up-regulated and one down- protein database (ID-1, Table 1).regulated) between the wild-type and luxS Seven proteins that were differentially ex-mutant strain (Table 1; Fig. 2) and 13 proteins pressed between the wild-type and mutant strainwere found to be differentially expressed (10 up- were also differentially expressed between theregulated and three down-regulated) when luxS mutant strain and the mutant strain supple-mutant was supplemented with AI-2 (Table 1; mented with AI-2 (Table 1). The pattern of
56 56ifi
3E1L19 S 31 E-1 9 S
2a2
1LEl 1L1
Ll
S
I
. SS....
S
IuxS Mutant Wild Type
FIG. 2. Comparative two-dimensional gel electrophoresis patterns of the soluble proteins fraction of the luxS mutant(PJ 002) and the wild-type strain (87-26254) of Salmonella Typhimurium.

*
13
56rn43W F19• 0
2E! L1
710
•ii
4,3LTh
4V 02 •&•..
0 ED 40
+1.53—4.34
+2.69
+2.86
+3.49
+2.15+1.81
+4.69—2.17+3.02
+5.72
+2.26
—2.7
+5.85
—3.03
+2.84+2.24+2.53
pf kA
riggroLvaeT
plioP
ta/B
+8.01
O#n1,F
gpnzl
,J.'OA
t,vBptaruM
+4.04
1Unidentified protein spots26-phosphofructokinase isozyme I
3Trigger factor (TF)460 kDa chaperonin5Outer membrane protein
precursor6Transcriptional regulatory
protein phoP7Transadolase B
8Outer membrane proteinF precursor
92,3-hiphosphoglycerate mutase
10DNA directed RNA polymerasealpha chain
11Thioredoxin reductase12Phosphate acetyl transferase13Hypothetical protein yiiM
'ID refers to spot shown in Figs. 1 and 2.
Carbohydrate transport andmetabolism
Cellular processes and signalingCellular processes and signalingCellular processes and signaling
Cellular processes and signaling
Carbohydrate transport andmetabolism
Cellular processes and signaling
Carbohydrate transport andmetabolism
Information storage and processing
Cellular processes and signalingMetabolismPutative hypothetical protein
PROTEOMIC ANALYSIS IN S. TYPHIMURIUM 151
.Vall -
luxS Mutant luxS Mutant + AI-2
FIG. 3. Comparative two-dimensional gel electrophoresis patterns of soluble proteins fraction of the luxS mutant(P1 002) and the luxS mutant supplemented with Al-2.
up-regulation of six of these proteins (TiG, YaeT, ference in the expression of six proteins (GroL,PhoP, GpmI, TrxB, and one unidentified TalB, OmpF, RpoA, Pta, and YiiM) between thespot) were similar; however, the protein iden- wild-type and IuxS mutant strain. However, intified as 6-phosphofructokinase isozyme I the presence of AI-2, three of these proteins(PfkA) was down-regulated in the wild-type (Ta1B, Pta, and YiiM) were up-regulated andand up-regulated in the mutant strain supple- three (GroL, OmpF, and RpoA) were down-mented with AI-2 (Table 1). There was no dif- regulated.
TABLE 1. IDENTIFIED QUORUM SENSING REGULATED PROTEINS USING MALDJ-MS.FOLD CHANGE SHOWN FOR BOTH WILD-TYPE AND LUXS MUTANT SUPPLEMENTED
WITH Al-2 ARE BASED ON COMPARISON WITH LUXS MUTANT
Clusters of ortliologoiis1O Pmti'i,, name Gene nameWild-typeA1-2 (COG) categories

152 SONI ET AL.
III
Discussion
The Salmonella Typhimurium wild-type cellsproduced maximal amount of AI-2 lumines-cence between mid- and early log phases ofgrowth (Fig. 1). The AI-2 level decreased as theculture entered into stationary phase. This de-creased level of AI-2 in stationary phase couldbe due to degradation or uptake of AI-2 mole-cules by bacterial cells (Xavier and Bassler,2005). The two component regulatory systemPhoP/PhoQ is a key transcriptional regulatorysystem that controls Salmonella pathogenicity(Aguirre et al., 2006; Nishino et al., 2006; Tu et al.,2006). We observed that PhoP was up-regulatedin the presence of AI-2 molecules compared toits absence, suggesting that AI-2 is involved inmodulating PhoP expression in Salmonella Ty-phimurium (Table 1). The two-component reg-ulatory system OmpR/EnvZ regulates the poringenes oinpF and ompC in response to osmolaritychanges in Salmonella (Mills et al., 1998). In thisstudy, the OmpF protein was down-regulatedin the luxS mutant supplemented with Al-2. Renet cii. (2004), however, reported that in E. co/i,onipi was down-regulated as a function of sig-naling molecules other than AI-2 implying thatthe Salmonella Tvphimurium and E. co/i may usean alternate signaling system to modulate OmpFexpression.
Proteins associated with the carbohydratetransport and metabolism also appear to beinfluenced by the presence of AI-2 molecules.The gene products of pJkA, gpnzl, and ta/B,which catalyze reactions in glycolysis, were up-regulated in the mutant strain supplementedwith AI-2 molecules. Compared to the -4.34fold down-regulation of PfkA protein in thewild-type (Table 1), we observed a 1.81-fold up-regulation of PfkA protein in the luxS mutantsupplemented with AI-2 molecules (Table 1).This difference in PfkA expression levels raisesthe possibility that there may be autoinducermolecules other than AI-2 (in the wild-typestrain) which are involved in modulating ex-pression of PfkA. Sperandio et al. (2003) havereported that the LuxS gene controls anotherautoinducer molecule (AI-3) in addition to AI-2in E. coli 0157:H7. In this study, there was a2.24-fold up-regulation of the Pta protein in thepresence of AI-2 molecules. Pta gene product is
involved in the phosphotransacetylase-ATP-acetate phosphotransferase (Pta-AckA) path-way to generate ATP molecules (Hardie et al.,2003). The observed up-regulation of the Ptaprotein by Al-2 implies that AI-2 may havesome role in Salmonella Typhimurium metabo-lism. Furthermore, consistent with our obser-vation, recent studies have also suggested thatthe LuxS/AI-2 system in bacterial cells mayhave an important metabolic role (Winzer et al.,2002a, 2002b; Walters and Sperandio, 2006).
The lsr operon, consisting of seven genes(1srACDBFGE) has been suggested to be invol-ved in the cellular uptake of Al-2 molecules inSalmonella Typhimurium (Taga et al., 2003). How-ever, Xavier and Bassler (2005) report that ifglucose is present in the medium, the lsr operonis not influenced by the AI-2 molecules. Theyhypothesized that catabolite repression occur-ring in the presence of glucose, prevents regu-lation of the Isr operon by AI-2 molecules. Thiscould explain the results we observed when wesupplemented LB media with 0.5% glucose anddid not observe the differential expression ofthe isr operon-related proteins (Table 1).
Our study focused only on the soluble proteinfraction and we detected approximately 198protein spots. The possibility that the LuxS/AI-2-mediated system in Salmonella Typhimuriummay control expression of some other (non-soluble) proteins cannot be ruled out. These re-sults confirm the importance of AI-2 moleculesas a key metabolic intermediate controlling avariety of proteins related to metabolism andvirulence. Thus it can be expected that any al-teration in metabolic pathways involving AI-2will be manifested in the pathogen's cellularprocesses.
Acknowledgments
This work was supported by Hatch grantH0858. We would like to thank Dr. Larry Dan-gott of the Protein Chemistry Laboratory, TexasA&M University for his valuable suggestions.The advice by Dr. Sperandio regarding AI-2synthesis is greatly appreciated.
References
Aguirre A, Cabeza ML, Spinelli SV, McClelland M, GarciaVescovi E, and Soncini FC. PhoP-induced genes within

PROTEOMIC ANALYSIS IN S. TYPHIMURIUM 153
Salmonella pathogenicity island 1. J . Bacteriol. 2006;188:6889-6898.
Arevalo-Ferro C, Hentzer M, and Roil C. Identification ofquorum-sensing regulated proteins in the opportunisticpathogen Pseudoinonas aeniginosa by proteomics. En-viron. Microhiol. 2003;5:1350-1369.
Arevalo-Ferro C, Red G, Gorg A, Ebert L, and Riedel K.Biofilm formation of Pseudomonas putida IsoF: the role ofquorum sensing as assessed by proteomics. Syst. Appl.Microhiol. 2005;28:87-114.
Bland AM, D'Eugcnio LR, Dugan MA, Janech MG,Almeida JS, Zile MR, and Arthur JM. Comparison ofvariability associated with sample preparation in two-dimensional gel electrophoresis of cardiac tissue. J . Bio-mol. Tech. 2006;17:195-199.
Chen X, Schauder 5, Potier N, Van Dorsselaer A, Pelczer 1,Hassler BL, and Hughson FM. Structural identification ofa bacterial quorum-sensing signal containing boron.Nature 2002;415:545-549.
Gomez TM, Motarjemi Y, Miyagawa 5, Kaferstein FK, andStohr K. Foodhorne salmonellosis. World Health Stat. Q.1997;50:81-89.
Gonzalez AF, Zuo R, Hashimoto Y, Yang L, Bentley WE,and Wood TK. Autoinducer 2 controls biofilm formationin Lsclierichia coli through a novel motility quorum-sensing regulator. J . Bacteriol. 2006;188:305-316.
Hardie KR, Cooksley C, Green AD, and Winzer K. Auto-inducer 2 activity in Escherichia co/i culture supernatantscan be actively reduced despite maintenance of an activesynthase, LuxS. Microbiology 2003;149:715-728.
Henke JM and Hassler BL. Bacterial social engagements.Trends Cell Biol. 2004;14:648-656.
Kim Y, Oh 5, Ahn EY, 1mm JY, Oh S, Park 5, and Kim SH.Proteome analysis of virulence factor regulated by au-toinducer-2-like activity in Esc/iericlua co/i 0157:F17.J . Food Prot. 2007; 70:300-307.
Lu L, Hume ME, and Pillai SD. Autoinducer-2-like activityassociated with foods and its interaction with food ad-ditives. J . Food Prot. 2004;67:1457-1462.
Miller ST, Xavier KB, Campagna SR, Taga ME, Semmel-hack MF, Hassler BL, and l-Iughson FM. Salmonella hi-p/iimuriuni recognizes a chemically distinct form of thebacterial quorum-sensing signal AI-2. Mot Cell 2004;15:677-687.
Mills SID, Ruschkowski SR, Stein MA, and Finlay BB.Trafficking of porin-deficient Salmonella typinmuriunimutants inside HeLa cells: ompR and envZ mutants aredefective for the formation of Salmonella-induced fila-ments. Infect. Immun. 1998;66:1806-1811.
Nishino K, Latifi T, and Groisman EA. Virulence and drugresistance roles of multid rug efflux systems of Salmonellaenferica serovar Typhimurium. Mot. Microhiol. 2006;59:126-141.
Pillai SD and Jesudhasan PR. Quorum sensing: how bac-teria communicate. Food Technol. 2007;60:42-50.
Ren D, Bedzyk LA, Ye RW, Thomas SM, and WoodTK. Stationary-phase quorum-sensing signals affectautoinducer-2 and gene expression in Esc/ieric/ua co/i.AppI. Environ. Microbiol. 2004;70:2038-2043.
Schauder S. Shokat K, Surette MG, and Hassler BL. TheLuxS family of bacterial autoinducers: biosynthesis ofa novel quorum-sensing signal molecule. Mot. Micro-biol. 2001;41:463-476.
Sperandio V. Torres AG, Jarvis B, Nataro JP, and KaperB. Bacteria-host communication: the language of hor-
mones. Proc. Natl. Acad. Sci. 2003;100:8951-8956.Surette MG and Hassler BL. Quorum sensing in Eschericlmia
co/i and Salmonella tijphinmurium. Proc. Natl. Acad. Sci.1998;95:7046-7050.
Taga ME, Miller ST. and Hassler BL. Lsr-mediated trans-port and processing of AI-2 in Salmonella h/p/unnhrwm.Mot. Microbiol. 2003;50:1411-1427.
Tu X, Latifi T, Bougdour A, Gottesman 5, and GroismanEA. The PhoF/PhoQ two-component system stabilizesthe alternative sigma factor RpoS in Salmonella emmtL'rica.Proc. Nati. Acad. Sci. 2006;103:13503-13508.
Walters M and Sperandio V. Quorum sensing in Esc/merichiaco/i and Salmonella. hit. J . Med. Microhiol. 2006;296:125-131.
Widmer KW, Jesudhasan PR, Dowd SE, and I'illai SD.Differential expression of virulence-related genes in aSalmonella enterica serotype Typhimurium luxS mutant inresponse to autoiriducer Al-2 and poultry meat-derivedAI-2 inhibitor. Foodhorne Pathog. Dis. 2007;4:5-15.
Winzer K, Hardie KR, and Burgess N. LuxS: its rolein central metabolism and the in vitro synthesis of 4-hydroxy-5-mcthyl-3(2H)-furanoflc. Microbiology 2002a;148:909-922.
Winzer K, Hardie KR, and Williams I'. Bacterial cell-to-cellcommunication: sorry, can't talk now-gone to lunch!Curr. Opin. Microbiol. 2002b;5:216-222.
Xavier KB and Hassler BL. LuxS quorum sensing: morethan just a numbers game. Curr. Opin. Microbiol. 2003;6:191-197.
Xavier KB and Hassler BL. Regulation of uptake and pro-cessing of the quorum-sensing autoinducer AT-2 inEsclmcric/ua co/i. J . Bacteriol. 2005;187:238-248.
Address reprint requests to:Suresh D. Pillai, Ph.D.
418D Kleberg CenterMS 2472, Texas A&M University
College Station, TX 77843
E-mail: [email protected]