
Microtubules, but not actin microfilaments, regulate vacuole motility and morphology in hyphae of...
11
Microtubules, But Not Actin Microfilaments, Regulate Vacuole Motility and Morphology in Hyphae of Pisolithus tinctorius Geoffrey J. Hyde,* Danielle Davies, Lara Perasso, Louise Cole, and Anne E. Ashford School of Biological Science, University of New South Wales, Sydney, Australia While it is now recognised that transport within the endomembrane system may occur via membranous tubules, spatial regulation of this process is poorly understood. We have investigated the role of the cytoskeleton in regulating the motility and morphology of the motile vacuole system in hyphae of the fungus Pisolithus tinctorius by studying (1) the effects of anti-microtubule (oryzalin, nocodazole) and anti-actin drugs (cytochalasins, latrunculin) on vacuolar activity, monitored by fluorescence microscopy of living cells; and (2) the ultrastructural relationship of microtubules, actin microfilaments, and vacuoles in hyphae prepared by rapid-freezing and freeze-substitution. Anti-microtubule drugs re- duced the tubular component of the vacuole system in a dose-dependent and reversible manner, the extent of which correlated strongly with the degree of disruption of the microtubule network (monitored by immunofluorescence micros- copy). The highest doses of anti-microtubule drugs completely eliminated tubular vacuoles, and only spherical vacuoles were observed. In contrast, anti-actin drugs did not reduce the frequency of tubular vacuoles or the motility of these vacuoles, even though immunofluorescence microscopy confirmed perturbation of microfila- ment organisation. Electron microscopy showed that vacuoles were always accompanied by microtubules. Bundles of microtubules were found running in parallel along the length of tubular vacuoles and individual microtubules were often within one microtubule diameter of a vacuole membrane. Our results strongly support a role for microtubules, but not actin microfilaments, in the spatial regulation of vacuole motility and morphology in fungal hyphae. Cell Motil. Cytoskeleton 42:114–124, 1999. r 1999 Wiley-Liss, Inc. Key words: fungus; cytoskeleton; tubule motility; latrunculin; oryzalin; tubular vacuoles INTRODUCTION Over the last decade one of the previously accepted dogmas of cell biology, that intracellular membrane transport is mediated solely by vesicles, has been increas- ingly challenged by evidence of other transport mecha- nisms, most significantly membranous tubules [Cole and Lippincott-Schwartz, 1995; Mironov et al., 1997]. Work with fluorescent probes in animal, plant, fungal, and protistan endomembrane systems has established that their compartments are, in some cases at least, highly interconnected, with tubules enabling the bulk transfer of membrane, membrane-embedded proteins, and luminal contents [Ashford, 1997; Cole and Lippincott-Schwartz, 1995; Cooper et al., 1990; Hopkins et al., 1990; Lazzaro and Thomson, 1996; Mironov et al., 1997; Shepherd et al., 1993a,b; Tooze and Hollinshead, 1991]. Tubular *Correspondence to: Geoffrey J. Hyde, School of Biological Science, University of New South Wales, Kensington, NSW, 2052, Australia. E-mail: [email protected] Lara Perasso’s present address is Research School of Biological Sciences, Australian National University, Canberra, ACT, 2601, Austra- lia. Contract grant sponsor: Australian Research Council. Received 12 August 1998; accepted 23 November 1998 Cell Motility and the Cytoskeleton 42:114–124 (1999) r 1999 Wiley-Liss, Inc.
Transcript of Microtubules, but not actin microfilaments, regulate vacuole motility and morphology in hyphae of...

Microtubules, But Not Actin Microfilaments,Regulate Vacuole Motility and Morphology
in Hyphae of Pisolithus tinctorius
Geoffrey J. Hyde,* Danielle Davies, Lara Perasso, Louise Cole,and Anne E. Ashford
School of Biological Science, University of New South Wales, Sydney, Australia
While it is now recognised that transport within the endomembrane system mayoccur via membranous tubules, spatial regulation of this process is poorlyunderstood. We have investigated the role of the cytoskeleton in regulating themotility and morphology of the motile vacuole system in hyphae of the fungusPisolithus tinctoriusby studying (1) the effects of anti-microtubule (oryzalin,nocodazole) and anti-actin drugs (cytochalasins, latrunculin) on vacuolar activity,monitored by fluorescence microscopy of living cells; and (2) the ultrastructuralrelationship of microtubules, actin microfilaments, and vacuoles in hyphaeprepared by rapid-freezing and freeze-substitution. Anti-microtubule drugs re-duced the tubular component of the vacuole system in a dose-dependent andreversible manner, the extent of which correlated strongly with the degree ofdisruption of the microtubule network (monitored by immunofluorescence micros-copy). The highest doses of anti-microtubule drugs completely eliminated tubularvacuoles, and only spherical vacuoles were observed. In contrast, anti-actin drugsdid not reduce the frequency of tubular vacuoles or the motility of these vacuoles,even though immunofluorescence microscopy confirmed perturbation of microfila-ment organisation. Electron microscopy showed that vacuoles were alwaysaccompanied by microtubules. Bundles of microtubules were found running inparallel along the length of tubular vacuoles and individual microtubules wereoften within one microtubule diameter of a vacuole membrane. Our resultsstrongly support a role for microtubules, but not actin microfilaments, in the spatialregulation of vacuole motility and morphology in fungal hyphae. Cell Motil.Cytoskeleton 42:114–124, 1999.r 1999 Wiley-Liss, Inc.
Key words: fungus; cytoskeleton; tubule motility; latrunculin; oryzalin; tubular vacuoles
INTRODUCTION
Over the last decade one of the previously accepteddogmas of cell biology, that intracellular membranetransport is mediated solely by vesicles, has been increas-ingly challenged by evidence of other transport mecha-nisms, most significantly membranous tubules [Cole andLippincott-Schwartz, 1995; Mironov et al., 1997]. Workwith fluorescent probes in animal, plant, fungal, andprotistan endomembrane systems has established thattheir compartments are, in some cases at least, highlyinterconnected, with tubules enabling the bulk transfer ofmembrane, membrane-embedded proteins, and luminalcontents [Ashford, 1997; Cole and Lippincott-Schwartz,
1995; Cooper et al., 1990; Hopkins et al., 1990; Lazzaroand Thomson, 1996; Mironov et al., 1997; Shepherd etal., 1993a,b; Tooze and Hollinshead, 1991]. Tubular
*Correspondence to: Geoffrey J. Hyde, School of Biological Science,University of New South Wales, Kensington, NSW, 2052, Australia.E-mail: [email protected]
Lara Perasso’s present address is Research School of BiologicalSciences, Australian National University, Canberra, ACT, 2601, Austra-lia.
Contract grant sponsor: Australian Research Council.
Received 12 August 1998; accepted 23 November 1998
Cell Motility and the Cytoskeleton 42:114–124 (1999)
r 1999 Wiley-Liss, Inc.

transfer could operate as a more rapid alternative tovesicular transport, especially where there is a require-ment for bulk flow [Ashford, 1997; Cooper et al., 1990;Mironov et al., 1997].
A tubule-based transport system, to be efficient,requires spatial control of its motility and morphology.From studies of the form and activity of endomembranecompartments in a range of animal cells, microtubules(MTs) have emerged as a major regulatory candidate, butmuch is still uncertain about their roles, the possiblecontribution of actin microfilaments (MFs), and the largerproblem of how directionality and specificity of transportare achieved [Cole and Lippincott-Schwartz, 1995; Miro-nov et al., 1997]. Owing to their small size, however,typical animal cells have presented a major technicalchallenge to fluorescence studies on the endomembranesystem in living cells. An alternative and promisingapproach is the study of endomembrane dynamics inelongated cells such as nerve axons or fungal hyphae.Indeed, studies to date have shown that the vacuolarsystem of fungal hyphae is one of the most elaborate andreadily examined of the tubule-utilising organelle sys-tems. The fungal vacuole system has two compartments:spherical or ovoid vacuoles, which typically becomelarger in cells distal from the hyphal apex, and are oftenpartially attached to the plasma membrane [Cole et al.,1998]; and tubules that intermittently interconnect thesespherical/ovoid vacuoles. The tubules exhibit extraordi-nary motility: At times their tips advance up to 60 µmalong the hypha at rates of up to 1.5 µm/s; they can weavethrough narrow pores in cell-separating septa; they ferrypartly-characterised content within their lumen; and theiractivity can be modulated by changes in the externalenvironment [Ashford, 1997; Hyde and Ashford, 1997;Shepherd et al., 1993a,b]. The highly developed nature ofthe fungal vacuolar system, and its widespread taxonomicdistribution [Rees et al., 1995], possibly reflect a require-ment for considerable longitudinal bulk transport alonghyphae. Until now, however, the role of the cytoskeletonin regulating tubule-mediated transport in fungal hyphaehas not been properly examined.
In this study, we have chemically perturbed the MTand MF networks of the basidiomycete fungus,Pisolithustinctorius, and monitored the effects on its vacuolarsystem with fluorescence microscopy of living cells. Wehave also used rapid-freezing and freeze-substitution withtransmission electron microscopy to examine the spatialrelationships of vacuoles and cytoskeletal elements. Aprevious immunofluorescence study ofP. tinctoriushasdemonstrated longitudinal MT bundles along the lengthof the hypha [Allaway, 1994]; and from electron micros-copy Shepherd et al. [1993b] reported both MTs and MFsadjacent to vacuolar elements. Given the likelihood thatP. tinctorius hyphae, like those of other fungi, have a
well-developed network of longitudinal MFs, there arepotentially two systems structured most suitably to co-ordinate the activity of a longitudinally-organised tubularvacuole system. Below we provide evidence that stronglysupports a role for MTs, but not MFs, in the regulation ofvacuolar motility and morphology in fungal hyphae.
MATERIALS AND METHODS
Reagents and Antibodies
Fluorescent probes, Oregon Green 488 carboxylicacid diacetate (carboxy-DFFDA, 10 mg/ml stock inDMSO, stored at -20°C) and 6-carboxyfluorescein diac-etate (CFDA, 10 mg/ml stock in acetone, stored at 4°C)were purchased from Molecular Probes (Eugene, OR).Anti-a-tubulin, lysing enzymes, cytochalasin B (10 mg/mlstock in DMSO, stored at -20°C), cytochalasin D (10mg/ml stock in DMSO, stored at -20°C), methyl-(5-[2-thienylcarbonyl]-1h-benzimidazol-2-yl)carbamate (Noco-dazole, 2 mg/ml stock in DMSO, stored at -20°C),2-[N-Morpholino]ethanesulfonic acid-free acid hydrate(MES), and phenylmethylsulfonyl fluoride (PMSF) wereall purchased from Sigma Chemical Co. Ltd. (St. Louis,MO). Sheep-anti-mouse Ig F(ab)2 fragment FITC conju-gate (SAM-FITC, stored at 4°C) was purchased fromSilenus Laboratories (Hawthorn, Victoria, Australia).Latrunculin B (2 mg/ml stock in DMSO stored at -20°C)and mouse anti-actin monoclonal (clone: C4) antibodywere obtained from ICN Biomedicals Inc. (Aurora, OH).Oryzalin was a gift from Dr Robyn Overall, School ofBiological Sciences, University of Sydney, Australia (10mM stock in acetone, used immediately).
Fungal Cultures
Pisolithus tinctorius(Pers.) Coker and Couch cul-tures, strain DI-15, isolated by Grenville et al. [1986],were grown on modified Melin-Norkrans (MMN) agar[Marx, 1969]. A modification of the cellophane sandwichtechnique of Campbell [1983] was used; the inoculumwas grown sandwiched between two discs of sterilecellophane placed on MMN agar according to Cole et al.[1997] for 9–14 days at 23°C in the dark.
Application and Monitoring of Anti-MTand Anti-Actin Drugs
Effects on vacuolar motility and morphology.Wedges of live mycelium were excised from the perim-eter of a colony, and incubated in a solution of reverseosmosis water (hereafter: H2O) containing a fluorescentvacuole probe, either CFDA, 20 µg/ml, or carboxy-DFFDA, 20 µg/ml [Cole et al., 1997], and either theappropriate control solvent (DMSO or acetone) or one ofvarious anti-MT (oryzalin or nocodazole) or anti-actin(cytochalasins B or D) drugs at a range of concentrations.
Vacuolar Motility in Fungal Hyphae 115

After 30 min, the wedges were rinsed for 15–45 min, andmounted on a glass slide under a coverslip in probe-freeH2O containing the same concentration of control solventor anti-cytoskeletal drug as was used during probe-loading. For latrunculin B treatments, the control solventor the drug itself was first applied at the probe rinsingstage (15 min). In some cases, wedges that had beensubjected to anti-cytoskeletal drug treatments and exam-ined microscopically, were subsequently re-rinsed in drugand solvent-free H2O (by perfusion under the coverslip)and re-examined after a recovery period of 30 min (forlatrunculin B) or 60 min (for oryzalin and nocodazole).
Vacuole motility and morphology were observedusing a Zeiss Axiophot (Sydney, Australia) microscopefitted with DIC and epifluorescence optics (filter combina-tion used; BP: 450–490 nm, FT:510 nm, and LP: 515–585nm), using a 40X, 0.75NA objective. Individual imagesor sequences of images were captured with a real-timedigital imaging set-up comprising an Image Point CCDcamera (Photometrics, Tucson, AZ), a PCI-compatibleLG3 framegrabber (Scion Corp, Fredrick, MD), and aScion version of NIH image (public domain imageanalysis software), on a Macintosh 9500 computer.Images were printed on an Epson Stylus 800 printer.
Vacuole motility and morphology were scored in avariety of ways, but in all cases mycelial wedges werescanned systematically around the perimeter from oneradial edge to the other, until either (1) a pre-determinednumber of hyphae had been scored (e.g., 50 or 100; or asnear to these numbers as possible) or (2) all hyphae in thepreparation had been scored. Using this procedure, mea-surements were typically made on the apical or penulti-mate cells of the hyphae. To minimise any errors due tovarying durations of exposure to drugs or control solu-tions, the initial time points of treatments and scoringwere varied appropriately.
Effects on the cytoskeleton.Mycelial wedges weretreated with oryzalin, latrunculin B, or control solvents asdescribed above but without fluorochrome loading andthen freeze-substituted for immunocytochemistry. Theywere plunge-frozen in liquid propane at below -185°C,and transferred to anhydrous ethanol at -90°C for 4 days,then gradually warmed to room temperature, at 5°C/h.Hyphae were then rehydrated in mixtures of ethanol and50 mM potassium phosphate buffer (pH 6.8, kept at 4°C)as follows: 95% ethanol for 30 min, 90% for 30 min, 80%for 30 min, 70% overnight, 60–0% by 10% steps at 15min each. They were next incubated for 15 min in MESbuffer (pH 5.5), and the walls digested with a lysingenzyme solution (10 mg/ml of lysing enzymes fromTrichoderma harzianum, having cellulase, protease, andchitinase activities), with 15 ml of 20 mM PMSF proteaseinhibitor and 1% BSA in MES buffer for 75 min at roomtemperature. Lysed hyphae were rinsed in MES buffer
(2x), phosphate buffered saline (PBS; 2x), and thenpermeabilized in 0.1% Triton X-100 in PBS for 15 min.Hyphae were rinsed again in PBS (3x), transferred toembryo cups, and incubated in anti-actin antibody (1:400dilution in PBS 1 1% BSA) or anti-a tubulin (1:400dilution in PBS1 1% BSA) for 60 min at 37°C. Theywere then rinsed in PBS buffer as before, and incubated inSAM-FITC (1:30 dilution in PBS1 1% BSA) for 60 min.Wedges were then mounted in Moviol [Wick and Duniec,1986] under a coverslip on a slide and kept at 4°Covernight before observation using the same microscope,filters, and imaging system as used for observations ofvacuole motility.
The effects of anti-cytoskeletal drugs and controlsolvents on MT and actin distributions were scored bysystematic scanning of mounted wedges, similar to thatdescribed above for scoring of vacuole motility andmorphology.
Cryofixation and Freeze-Substitutionfor Electron Microscopy
Cultures used for electron microscopy were 7–12days old. One day prior to fixation, the top layer ofcellophane was discarded and small triangles (approxi-mately 5 mm per side) of cellophane and mycelium werecut at the leading edge of the colony. The cellophaneextended about 2 mm beyond the hyphal tips. Thetriangles were then placed face down on a part of theremaining sheet of cellophane that had been cleared ofhyphae. The Petri dish was resealed and returned to theincubator. Next day, cultures were removed from theincubator about 15 min prior to fixation. Immediatelyprior to fixation, the triangles were picked up withreverse-action forceps, carefully dried with filter paper,and plunged into liquid propane at a temperature of lessthan -180°C. A propane plunge-freezing device based onthe designs of Hoch [1986] and Howard and O’Donnell[1987] was used. Samples were substituted in 2% OsO4 indry acetone in a Leica AFS machine for 3 days at -85°Cand then brought to room temperature at a rate of 2°C/h.The samples were rinsed in acetone and infiltrated withProcure-Araldite resin over 2 days. The cellophanetriangles with mycelium were polymerized between twoteflon-coated slides for 12 h at 70°C. The glass slideswere then separated and hyphae examined under a lightmicroscope. Well-fixed samples were selected with aZeiss diamond scriber, cut out and mounted on resinblocks for serial sectioning [see Hoch, 1986, and Howardand O’Donnell, 1987, for selection criteria]. Sectionswere stained for 7 min with saturated uranyl acetate in50% methanol, for 5 min in Reynolds [1963] lead citrate,and once again for 2 min in the uranyl acetate solution.Sections were viewed on a Hitachi 7100 TEM (Naka,Japan).
116 Hyde et al.
RESULTS
Effects of Anti-MT Drugs on Vacuoleorphology and Motility
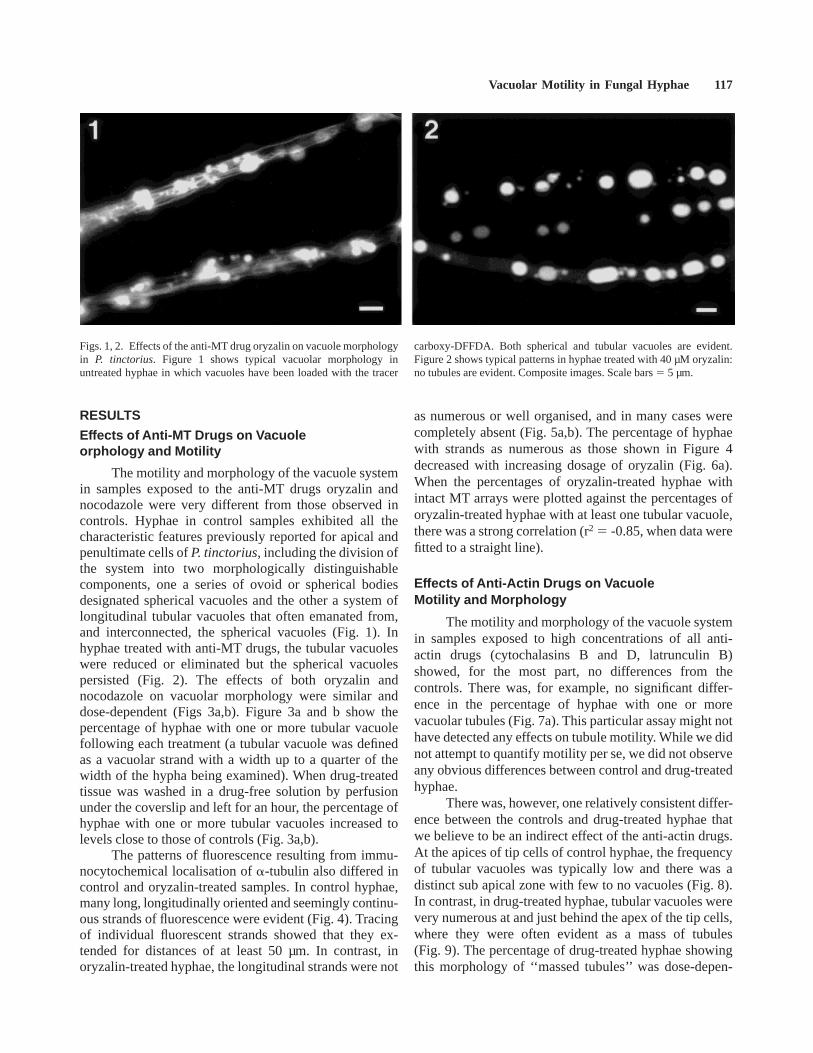
The motility and morphology of the vacuole systemin samples exposed to the anti-MT drugs oryzalin andnocodazole were very different from those observed incontrols. Hyphae in control samples exhibited all thecharacteristic features previously reported for apical andpenultimate cells ofP. tinctorius, including the division ofthe system into two morphologically distinguishablecomponents, one a series of ovoid or spherical bodiesdesignated spherical vacuoles and the other a system oflongitudinal tubular vacuoles that often emanated from,and interconnected, the spherical vacuoles (Fig. 1). Inhyphae treated with anti-MT drugs, the tubular vacuoleswere reduced or eliminated but the spherical vacuolespersisted (Fig. 2). The effects of both oryzalin andnocodazole on vacuolar morphology were similar anddose-dependent (Figs 3a,b). Figure 3a and b show thepercentage of hyphae with one or more tubular vacuolefollowing each treatment (a tubular vacuole was definedas a vacuolar strand with a width up to a quarter of thewidth of the hypha being examined). When drug-treatedtissue was washed in a drug-free solution by perfusionunder the coverslip and left for an hour, the percentage ofhyphae with one or more tubular vacuoles increased tolevels close to those of controls (Fig. 3a,b).
The patterns of fluorescence resulting from immu-nocytochemical localisation ofa-tubulin also differed incontrol and oryzalin-treated samples. In control hyphae,many long, longitudinally oriented and seemingly continu-ous strands of fluorescence were evident (Fig. 4). Tracingof individual fluorescent strands showed that they ex-tended for distances of at least 50 µm. In contrast, inoryzalin-treated hyphae, the longitudinal strands were not
as numerous or well organised, and in many cases werecompletely absent (Fig. 5a,b). The percentage of hyphaewith strands as numerous as those shown in Figure 4decreased with increasing dosage of oryzalin (Fig. 6a).When the percentages of oryzalin-treated hyphae withintact MT arrays were plotted against the percentages oforyzalin-treated hyphae with at least one tubular vacuole,there was a strong correlation (r2 5 -0.85, when data werefitted to a straight line).
Effects of Anti-Actin Drugs on VacuoleMotility and Morphology
The motility and morphology of the vacuole systemin samples exposed to high concentrations of all anti-actin drugs (cytochalasins B and D, latrunculin B)showed, for the most part, no differences from thecontrols. There was, for example, no significant differ-ence in the percentage of hyphae with one or morevacuolar tubules (Fig. 7a). This particular assay might nothave detected any effects on tubule motility. While we didnot attempt to quantify motility per se, we did not observeany obvious differences between control and drug-treatedhyphae.
There was, however, one relatively consistent differ-ence between the controls and drug-treated hyphae thatwe believe to be an indirect effect of the anti-actin drugs.At the apices of tip cells of control hyphae, the frequencyof tubular vacuoles was typically low and there was adistinct sub apical zone with few to no vacuoles (Fig. 8).In contrast, in drug-treated hyphae, tubular vacuoles werevery numerous at and just behind the apex of the tip cells,where they were often evident as a mass of tubules(Fig. 9). The percentage of drug-treated hyphae showingthis morphology of ‘‘massed tubules’’ was dose-depen-
Figs. 1, 2. Effects of the anti-MT drug oryzalin on vacuole morphologyin P. tinctorius. Figure 1 shows typical vacuolar morphology inuntreated hyphae in which vacuoles have been loaded with the tracer
carboxy-DFFDA. Both spherical and tubular vacuoles are evident.Figure 2 shows typical patterns in hyphae treated with 40 µM oryzalin:no tubules are evident. Composite images. Scale bars5 5 µm.
Vacuolar Motility in Fungal Hyphae 117

Fig. 3. a,b: Effects of anti-MT drugs oryzalin and nocodazole onfrequency of tubular vacuoles in hyphae loaded with CFDA. Both thesedrugs caused a dose-dependent reduction in the frequency of hyphae inwhich at least one tubular vacuole was observed. The effects of the
highest concentrations used were reversed by washing out the drugsand leaving hyphae to recover for 1 h. Results are the means of threeexperiments, with standard errors shown. n5 100 for all experiments.
Figs. 4, 5. Effects of oryzalin on MT organisation. Figure 4 shows thetypical pattern of fluorescence seen in a control hypha that had beenrapidly-frozen, freeze-substituted, rehydrated, and then labelled withanti-a-tubulin and SAM-FITC antibodies. The hypha shown in Figure
5a,b has been processed similarly, except for a pre-treatment with 40µM oryzalin.a: No strands of fluorescence are evident.b: DIC imageof the same hypha, illustrating that the cytoplasm in the MT-free areaappears well preserved. Scale bars5 5 µm.
118 Hyde et al.
dent, and the effect was reversed after hyphae were rinsedwith drug-free solution and left to recover (Fig. 7b).
While anti-actin drugs did not affect the vacuolarsystem in the dramatic fashion of the anti-MT drugs,immunofluorescence microscopy demonstrated that latrun-culin B, at least, was efficacious in disturbing MForganisation. The patterns of anti-actin fluorescence werevery different in control and drug-treated samples. Incontrol hyphae, the immunolocalisation procedure dem-onstrated apical actin caps and ‘‘plaques’’ along thelength of 84% of the hyphae (Figs. 10, 12), but attemptsto localise the peripheral longitudinal actin componentwere not successful. In contrast in samples treated with10 or 25 µM latrunculin B, actin caps and plaques werenot found (Figs. 11, 12).
Electron Microscopy of Hyphae: Vacuolesand the Cytoskeleton
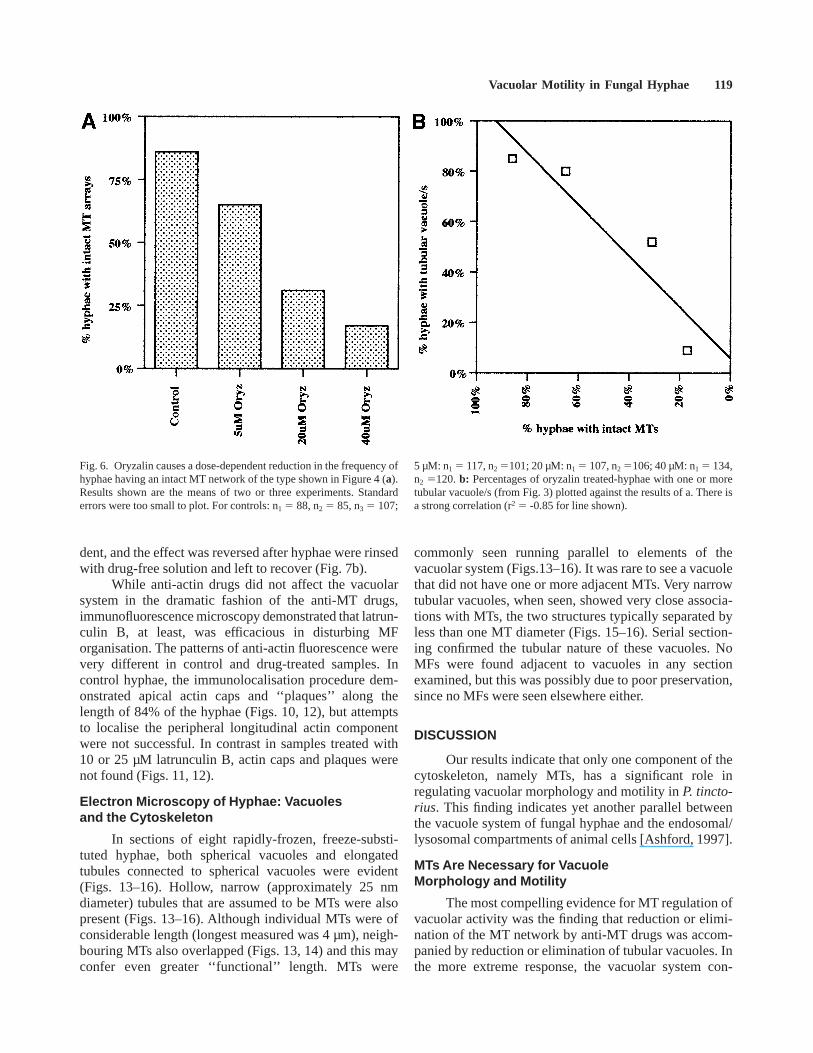
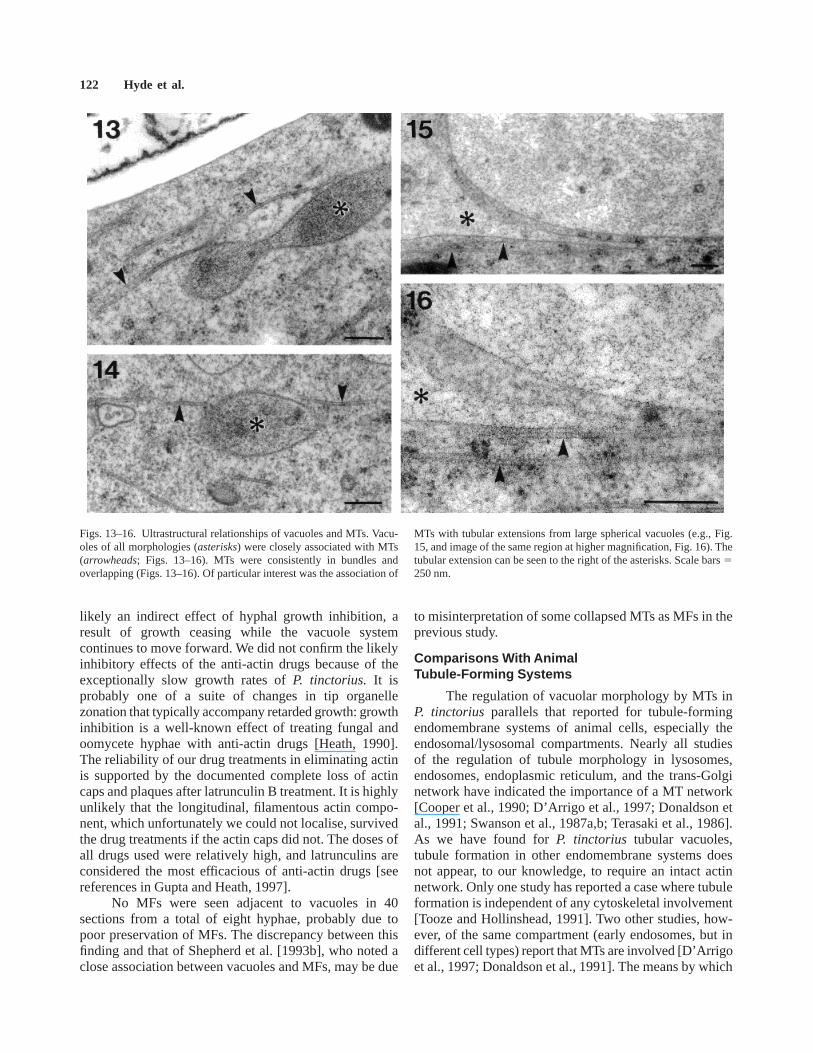
In sections of eight rapidly-frozen, freeze-substi-tuted hyphae, both spherical vacuoles and elongatedtubules connected to spherical vacuoles were evident(Figs. 13–16). Hollow, narrow (approximately 25 nmdiameter) tubules that are assumed to be MTs were alsopresent (Figs. 13–16). Although individual MTs were ofconsiderable length (longest measured was 4 µm), neigh-bouring MTs also overlapped (Figs. 13, 14) and this mayconfer even greater ‘‘functional’’ length. MTs were
commonly seen running parallel to elements of thevacuolar system (Figs.13–16). It was rare to see a vacuolethat did not have one or more adjacent MTs. Very narrowtubular vacuoles, when seen, showed very close associa-tions with MTs, the two structures typically separated byless than one MT diameter (Figs. 15–16). Serial section-ing confirmed the tubular nature of these vacuoles. NoMFs were found adjacent to vacuoles in any sectionexamined, but this was possibly due to poor preservation,since no MFs were seen elsewhere either.
DISCUSSION
Our results indicate that only one component of thecytoskeleton, namely MTs, has a significant role inregulating vacuolar morphology and motility inP. tincto-rius. This finding indicates yet another parallel betweenthe vacuole system of fungal hyphae and the endosomal/lysosomal compartments of animal cells [Ashford, 1997].
MTs Are Necessary for VacuoleMorphology and Motility
The most compelling evidence for MT regulation ofvacuolar activity was the finding that reduction or elimi-nation of the MT network by anti-MT drugs was accom-panied by reduction or elimination of tubular vacuoles. Inthe more extreme response, the vacuolar system con-
Fig. 6. Oryzalin causes a dose-dependent reduction in the frequency ofhyphae having an intact MT network of the type shown in Figure 4 (a).Results shown are the means of two or three experiments. Standarderrors were too small to plot. For controls: n1 5 88, n2 5 85, n3 5 107;
5 µM: n1 5 117, n2 5101; 20 µM: n1 5 107, n2 5106; 40 µM: n1 5 134,n2 5120.b: Percentages of oryzalin treated-hyphae with one or moretubular vacuole/s (from Fig. 3) plotted against the results of a. There isa strong correlation (r2 5 -0.85 for line shown).
Vacuolar Motility in Fungal Hyphae 119

verted entirely to a series of spherical vacuoles. Whenoryzalin was applied over a range of doses, there was astrong correlation between the degree of MT disruptionand the extent of disruption of tubular vacuoles. The rapidrecovery of typical vacuolar morphology after washingout the oryzalin indicates that the depression of vacuolaractivity is largely a direct result of changes to the MTnetwork and is not due to cell death.
In addition to the evidence above, the form anddistribution of theP. tinctoriusMT network, as revealedby both immunofluorescence and electron microscopy,are consistent with a role in supporting the morphologyand motility of the longitudinally oriented fungal tubularvacuole system. Localisation ofa-tubulin by immunoflu-orescence indicated the existence of a more complex MTnetwork than that previously reported forP. tinctorius[Allaway, 1994], probably due to better preservation bythe rapid-freezing and freeze-substitution techniques usedhere. The lengths of the strands of fluorescence arecommensurate with the distances over which individualvacuolar tubules extend. Previous electron microscopy of
oomycete hyphae indicates, however, that individual MTsare much shorter than strands of fluorescence seen withlight microscopy [Heath and Kaminskyj, 1989]; andcorrelation of fluorescent images with electron micros-copy suggests that, inP. tinctorius, each fluorescentstrand is probably a series of shorter MTs whose overlap-ping configuration may allow them to function as a muchlonger single unit.
The close spatial relationship seen between MTsand vacuoles further supports a role for MTs in regulatingvacuolar motility, as first speculated forP. tinctoriusbyShepherd et al. [1993b]. In the present study, longitudi-nally oriented tubular vacuoles were nearly always accom-panied by longitudinally oriented MTs. In many in-stances, the distance between the vacuolar membranesurface and an adjacent MT was less than the diameter ofa MT. Of particular interest were our observations ofassociations between MTs and the most narrow tubularvacuoles. In living cells, such narrow tubules are seen tocarry out the most dramatic motile behaviour of thesystem; acting in ‘‘pioneer’’ fashion, single tubules
Fig. 7. Effects of anti-actin drugs on the frequency of hyphae withtubular vacuoles loaded with carboxy-DFFDA. Neither cytochalasin B,cytochalasin D, nor latrunculin B had a significant effect on thefrequency of hyphae with one or more tubular vacuole/s (a). Results aremeans of three experiments. Some standard errors were too small toplot. n5 100 for all experiments. Anti-actin drugs did however cause achange in the distribution of tubular vacuoles at the hyphal apex.Latrunculin increased the frequency of hyphal apices having a ‘‘massedtubules’’ configuration of the type shown in Figure 9 (b). This effectwas dose-dependent and reversible after the drug was washed out and
recovery was allowed to take place. Results shown are the means ofthree experiments. Standard errors were too small to plot. For control(a) and control recovery (b): n1a, n1b 5 133, 140; n2a, n2b 5 126, 118;n3a, n3b 5 112, 122; for 0.2% DMSO (a) and recovery (b): n1a, n1b 5135, 135; n2a, n2b 5 108, 120; n3a, n3b 5 121, 114; for 0.5% DMSO (a)and recovery (b): n1a, n1b 5 137, 132; n2a, n2b 5 123, 129; n3a, n3b 5118, 107; for 10 µM LatB (a) and recovery (b): n1a, n1b 5 151, 145; n2a,n2b 5 105, 100; n3a, n3b 5 99, 102; for 25 µM LatB (a) and recovery (b):n1a, n1b 5 139, 130; n2a, n2b 5 116, 112; n3a, n3b 5 132, 129.
120 Hyde et al.

regularly and rapidly extend through the cytoplasm fordistances of 60 µm or more, before fusing with a distantvacuole [Shepherd et al., 1993b]. No other vacuolaractivity suggests the existence of intracellular scaffoldingmore than the forays of these delicate tubules. Theindication from our electron microscopy that narrowtubular vacuoles are most frequent in the peripheralcytoplasm is consistent with the long-acknowledgedgreater density of MTs in this region of fungal andoomycete hyphae [Derksen and Emons, 1990; Heath andKaminskyj, 1989]. Allaway et al [1997] have also docu-mented an associtaion between MTs and vacuoles in thehyphal organism,Saprolegnia ferax.
Lack of Evidence for Involvement of Mfs inRegulating Vacuole Morphology and Motility
The results offer no support for any involvement ofactin MFs in the regulation of vacuolar morphology inP.tinctoriushyphae. Prolonged exposure of hyphae to threeanti-actin drugs neither reduced the frequency of tubularvacuoles, nor their motility. In fact, the only obviouseffect of anti-actin drugs was to increase the number oftubules near the hyphal apex. We interpret this to be most
Fig. 12. Quantification of the inhibitory effect of latrunculin B on thefrequency of actin caps shown pictorially in Figures 10 and 11. Forcontrol: n5 186; for 10 µM: n5 231; for 25 µM: n5 186.
Figs. 8–11. Effect of latrunculin B on distribution of tubular vacuolesin the hyphal apex. In the control hypha, the frequency of tubularvacuoles is typically low at the apex (arrowhead, Fig. 8). In latrunculin-treated hyphae, however, tubular vacuoles are massed near the apex(arrowheads, Fig. 9). Figure 9 is a composite image. Vacuoles were
loaded with carboxy-DFFDA. Scale bars in Figures 8 and 95 5 µm.Immunolocalisation experiments with anti-actin monoclonal antibod-ies showed the presence of actin caps and plaques in healthy untreatedhyphal tips (Fig. 10), and the absence of these structures in hyphae treatedwith 10 µM latrunculin B (Fig. 11). Scale bars in Figures 10 and 115 5 µm.
Vacuolar Motility in Fungal Hyphae 121
likely an indirect effect of hyphal growth inhibition, aresult of growth ceasing while the vacuole systemcontinues to move forward. We did not confirm the likelyinhibitory effects of the anti-actin drugs because of theexceptionally slow growth rates ofP. tinctorius. It isprobably one of a suite of changes in tip organellezonation that typically accompany retarded growth: growthinhibition is a well-known effect of treating fungal andoomycete hyphae with anti-actin drugs [Heath, 1990].The reliability of our drug treatments in eliminating actinis supported by the documented complete loss of actincaps and plaques after latrunculin B treatment. It is highlyunlikely that the longitudinal, filamentous actin compo-nent, which unfortunately we could not localise, survivedthe drug treatments if the actin caps did not. The doses ofall drugs used were relatively high, and latrunculins areconsidered the most efficacious of anti-actin drugs [seereferences in Gupta and Heath, 1997].
No MFs were seen adjacent to vacuoles in 40sections from a total of eight hyphae, probably due topoor preservation of MFs. The discrepancy between thisfinding and that of Shepherd et al. [1993b], who noted aclose association between vacuoles and MFs, may be due
to misinterpretation of some collapsed MTs as MFs in theprevious study.
Comparisons With AnimalTubule-Forming Systems
The regulation of vacuolar morphology by MTs inP. tinctorius parallels that reported for tubule-formingendomembrane systems of animal cells, especially theendosomal/lysosomal compartments. Nearly all studiesof the regulation of tubule morphology in lysosomes,endosomes, endoplasmic reticulum, and the trans-Golginetwork have indicated the importance of a MT network[Cooper et al., 1990; D’Arrigo et al., 1997; Donaldson etal., 1991; Swanson et al., 1987a,b; Terasaki et al., 1986].As we have found forP. tinctorius tubular vacuoles,tubule formation in other endomembrane systems doesnot appear, to our knowledge, to require an intact actinnetwork. Only one study has reported a case where tubuleformation is independent of any cytoskeletal involvement[Tooze and Hollinshead, 1991]. Two other studies, how-ever, of the same compartment (early endosomes, but indifferent cell types) report that MTs are involved [D’Arrigoet al., 1997; Donaldson et al., 1991]. The means by which
Figs. 13–16. Ultrastructural relationships of vacuoles and MTs. Vacu-oles of all morphologies (asterisks) were closely associated with MTs(arrowheads; Figs. 13–16). MTs were consistently in bundles andoverlapping (Figs. 13–16). Of particular interest was the association of
MTs with tubular extensions from large spherical vacuoles (e.g., Fig.15, and image of the same region at higher magnification, Fig. 16). Thetubular extension can be seen to the right of the asterisks. Scale bars5250 nm.
122 Hyde et al.

MTs regulate tubule activity in animal cells is uncertainbut several studies have indicated the involvement of aMT motor-protein, kinesin [Hollenbeck and Swanson,1990; Swanson et al., 1992; Steinberg et al., 1998, andwork referenced therein]. The bidirectional nature ofvacuolar transport in hyphae suggests that either two ormore motor proteins are involved, or adjacent MTs are ofdiffering polarity.
CONCLUSIONS
The involvement of MTs in the regulation of bothfungal vacuoles and animal endosomes/lysosomes addsto a long list of previously reported similarities betweenthe two systems, including their characteristic morphol-ogy and motility, the passage of material within tubules,their internal acidity, and the sensitivity of the system’smotility to changes in the external environment [Hydeand Ashford, 1997; Rees et al., 1995; Rost et al., 1995;Shepherd et al., 1993b]. These similarities all point to theappropriateness of considering the vacuoles of theserelatively large fungal cells as an amenable model systemfor the study of endosomal/lysosomal tubule formation ineukaryotic cells generally.
ACKNOWLEDGMENTS
We thank Xiaomei Niu, Jim Pearse, and CandidaBriggs for assistance. Lara Perasso thanks Sally Stowefor use of the electron microscope facilities at theAustralian National University and to Adrienne Hardhamof the ANU for use of the Plant Cell Biology Groupfacilities. This work was supported by a AustralianResearch Council Postdoctoral Fellowship to G.J.H. andan ARC Large Grant to A.E.A.
REFERENCES
Allaway WG. 1994. Microtubules and the cytoplasmic motile tubularvacuole system inPisolithus tinctoriushyphae. In StephensonAG, Kao T-H, editors. Pollen-pistil interactions and pollen tubegrowth. Rockville MD: American Society of Plant Physiolo-gists, p 265–267.
Allaway WG, Ashford AE, Heath IB, Hardham AR. 1997. Vacuolarreticulum in oomycete hyphal tips - an additional component ofthe Ca21 regulatory system. Fung Genet Biol 22:209–220.
Ashford AE. 1997. Dynamic pleiomorphic vacuole systems: Are theyendosomes and transport compartments in fungal hyphae? AdvBot Res 28:119–159.
Campbell IM. 1983. Fungal secondary metabolism research: past,present and future. J Nat Products 46:60–70.
Cole L, Hyde GJ, Ashford AE. 1997. Uptake and compartmentalisationof fluorescent probes byPisolithus tinctoriushyphae: evidencefor an anion transport mechanism at the tonoplast but not forfluid-phase endocytosis. Protoplasma 199:18–29.
Cole L, Orlovich DA, Ashford AE. 1998. Structure, function and
motility of vacuoles in filamentous fungi. Fung Genet Biol24:86–100.
Cole NB, Lippincott-Schwartz J. 1995. Organisation of organelles andmembrane traffic by microtubules. Curr Opin Cell Biol 7:55–64.
Cooper MS, Cornell-Bell AH, Chernjavsky A, Dani JW, Smith SJ.1990. Tubulovesicular processes emerge fromtrans-Golgi cis-ternae, extend along microtubules, and interlink adjacenttrans-Golgi elements into a reticulum. Cell 61:135–145.
D’Arrigo A, Bucci C, Toh B-H, Stenmark H. 1997. Microtubules areinvolved in bafilomycin A1-induced tubulation and Rab5-dependent vacuolation of early endosomes. Eur J Cell Biol72:95–103.
Derksen J, Emons AM. 1990. Microtubules in tip growth systems. InHeath IB, editor. Tip growth of plant and fungal cells. NewYork: Academic Press, p 148–181.
Donaldson JG, Kahn RA, Lippincott-Schwartz J, Klausner RD. 1991.Binding of ARF and beta-COP to Golgi membranes: possibleregulation by a trimeric G protein. Science 254:1197–1199.
Grenville DJ, Peterson RL, Ashford AE. 1986. Synthesis in growthpouches of mycorrhizae betweenEucalyptus pilularis andseveral strains ofPisolithus tinctorius. Aust J Bot 34:95–102.
Gupta GD, Heath IB. 1997. Actin disruption by latrunculin B causesturgor-related changes in tip growth ofSaprolegnia feraxhyphae. Fung Genet Biol 21:64–75.
Heath IB. 1990. The roles of actin in tip growth in fungi. Int Rev Cytol123:95–127.
Heath IB, Kaminskyj SGW. 1989. The organization of tip-growth-related organelles and microtubules revealed by quantitativeanalysis of freeze-substituted oomycete hyphae. J Cell Sci93:41–52.
Hoch HC. 1986. Freeze substitution of fungi. In Aldrich HC, Todd WJ,editors. Ultrastructure techniques for microorganisms. NewYork: Plenum Publishing, p 183–212.
Hollenbeck PJ, Swanson JA. 1990. Radial extension of macrophagetubular lysosomes supported by kinesin. Nature 346:864–866.
Hopkins CR, Gibson A, Shipman M, Miller K. 1990. Movement ofinternalized ligand-receptor complexes along a continuousendosomal reticulum. Nature 346:335–339.
Howard RJ, O’Donnell KL. 1987. Freeze substitution of fungi forcytological analysis. Exp Mycol 11:250–269.
Hyde GJ, Ashford AE. 1997. Vacuole motility and tubule-formingactivity in Pisolithus tinctoriushyphae are modified by environ-mental conditions. Protoplasma 198:85–92.
Lazzaro MD, Thomson WW. 1996. The vacuolar-tubular continuum inliving trichomes of chickpea (Cicer arietinum) provides a rapidmeans of solute delivery from base to tip. Protoplasma 193:181–190.
Marx DH. 1969. The influence of ectotrophic mycorrhizal fungi on theresistance of pine roots to pathogenic infections. I. Antagonismof mycorrhizal fungi to root pathogenic fungi and soil bacteria.Phytopathology 59:159–163.
Mironov AA, Weidman P, Luini A. 1997. Variations on the intracellulartransport theme: maturing cisternae and trafficking tubules. JCell Biol 138:481–484.
Rees BJ, Shepherd VA, Ashford VA. 1995. Presence of a motilevacuolar system in different phyla of fungi. Mycol Res 99:549.
Reynolds ES. 1963. The use of lead citrate at high pH as an electronopaque stain in electron microscopy. J Cell Biol 17:208–212.
Rost FWD, Shepherd VA, Ashford AE. 1995. Estimation of vacuolarpH in actively growing hyphae of the fungusPisolithustinctorius. Mycol Res 99:549–553.
Shepherd VA, Orlovich DA, Ashford AE. 1993a. Cell-to-cell transportvia motile tubules in growing hyphae of a fungus. J Cell Sci105:1171–1178.
Vacuolar Motility in Fungal Hyphae 123

Shepherd VA, Orlovich DA, Ashford AE. 1993b. A dynamic continuumof pleiomorphic tubules and vacuoles in growing hyphae of afungus. J Cell Sci 104:495–507.
Steinberg G, Schliwa M, Lehmler C, Bolker M, Kahmann R, McintoshJR. 1998. Kinesin from the plant pathogenic fungusUstilagomaydis is involved in vacuole formation and cytoplasmicmigration. J Cell Sci 111:2235–2246.
Swanson JA, Locke A, Ansel P, Hollenbeck PJ. 1992. Radial move-ment of lysosomes along microtubules in permeabilized macro-phages. J Cell Sci 103:201–209.
Swanson J, Burke E, Silverstein SC. 1987a. Tubular lysosomesaccompany stimulated pinocytosis in macrophages. J Cell Biol104:1217–1222.
Swanson J, Bushnell A, Silverstein SC. 1987b. Tubular lysosomemorphology and distribution within macrophages depend on theintegrity of cytoplasmic microtubules. Proc Natl Acad Sci USA84:1921–1925.
Terasaki M, Chen LB, Fujiwara K. 1986. Microtubules and theendoplasmic reticulum are highly interdependent structures. JCell Biol 103:1557–1568.
Tooze J, Hollinshead M. 1991. Tubular early endosomal networks inAtT20 and other cells. J Cell Biol 115:635–653.
Wick SM, Duniec J. 1986. Effects of various fixatives on the reactivityof plant cell tubulin and calmodulin in immunofluorescencemicroscopy. Protoplasma 133:1–18.
124 Hyde et al.












