
Research Article Dyeing Studies with Eucalyptus, Quercetin ...

RESEARCH New Phytol. (2000), 145, 333–346
Apoplasmic barriers and their significance
in the exodermis and sheath of Eucalyptus
pilularis–Pisolithus tinctorius
ectomycorrhizas
PETER A. VESK", ANNE E. ASHFORD#*, ANNE-LAURE
MARKOVINA" WILLIAM G. ALLAWAY"
"School of Biological Sciences, The University of Sydney, NSW 2006, Australia
#School of Biological Science, The University of New South Wales, Sydney, NSW 2052,
Australia
Received 17 June 1999; accepted 19 October 1999
The apoplasmic permeability of ectomycorrhizal roots of intact Eucalyptus pilularis seedlings infected with
Pisolithus tinctorius on aseptic agar plates was examined using the nonbinding fluorochrome 8-hydroxypyrene-
1,3,6-trisulphonate and lanthanum ions in conjunction with anhydrous freeze substitution and dry sectioning.
Most mycorrhizas formed in the air above the agar surface, and in these the sheath rapidly became nonwettable
and impermeable to the fluorochrome but was nevertheless permeable to lanthanum ions. In a few mycorrhizas
which developed in contact with the agar the sheath remained permeable to both tracers when fully developed.
This increased hydrophobicity of the sheath in mycorrhizas in the air above the agar surface might be explained
by deposition of hydrophobins, but nevertheless it still allows an apoplasmic pathway for radial movement of ions.
Regardless of their sheath permeation both apoplasmic tracers were always found throughout the Hartig net and
were arrested at the Casparian bands and suberin lamellae of the exodermis. It is concluded that the fluorochrome
must have moved longitudinally along the Hartig net which is a region of higher permeability than the sheath.
Casparian bands in the exodermis of ectomycorrhizal roots have similar properties to those in nonmycorrhizal
roots in excluding solutes and their exclusion of lanthanum ions indicates that they are not permeable to ions. The
data do not support the concept of a totally sealed apoplasmic exchange compartment, but the differential
permeability suggests that the sheath might allow radial transfer of ions but block loss of sugars and organic
molecules of similar size.
Key words: apoplasmic permeability, Casparian band, exodermis, ectomycorrhiza, freeze substitution,
hydrophobins, lanthanum, PTS.
Early physiological experiments tended to focus on
the ability of excised ectomycorrhizal roots to absorb
ions compared with nonmycorrhizal roots of the
same species (Harley, 1969). This approach treated
ectomycorrhizal roots essentially as uptake struc-
tures. However the ectomycorrhizal roots of many
associations are connected to an extraradical my-
celium which can grow out into the soil for
considerable distances and form structures of great
complexity (Read, 1991). This mycelium is now
*Author for correspondence (fax 61 2 9385 1558; e-mail
a.ashford!unsw.edu.au).
considered to play a crucial role in nutrient and
water acquisition and in mobilization of organic
resources. Nutrients acquired by the hyphal tips are
transported back to the mycorrhizal sheath which is
now viewed more as a storage and exchange
structure, or food base, than as a nutrient-absorbing
organ (Smith & Read, 1997). One of the many pieces
of evidence which agree with this view is the
discovery that the fluorochrome calcofluor would not
penetrate the sheath of two types of ectomycorrhiza,
those formed on Eucalyptus pilularis by Pisolithus
tinctorius and those found on Pisonia grandis,
suggesting that the sheath forms an impermeable
barrier to this fluorochrome (Ashford et al., 1988,
1989). Since the mycorrhizal interface in E. pilu-

334 RESEARCH P. A. Vesk et al.
laris–P. tinctorius mycorrhizas is sandwiched be-
tween an exodermis and an impermeable sheath, this
led to the hypothesis that there is a sealed apoplasmic
compartment at the interface between partners. The
benefits of the isolation of the exchange interface are
obvious, and were discussed in Ashford et al. (1989).
The low apoplasmic permeability of the sheath in
these two types of mycorrhizas led Ashford et al.
(1989) to suggest that ion transport across the sheath
is essentially symplasmic, and that ectomycorrhizal
roots are exchange rather than uptake structures.
The apoplasmic permeability of the sheath was
assessed in these earlier experiments by applying
calcofluor to excised roots with their cut ends sealed
with sticky wax, and determining the distribution of
calcofluor in hand sections following a short wash. It
was presumed that calcofluor was retained by
electrostatic binding in the plant and fungal walls to
which it had access, but there are problems in using
this approach because the results depend on dye
binding as well as permeability. Dye molecules
might be lost because they were not bound, they
might be redistributed and preferentially bound to
more electronegative sites, or bound dye might
impede the further progress of other dye molecules
through the walls (Canny, 1986; Canny & McCully,
1986). An approach which avoids these possibilities
is to allow low molecular weight fluorochromes that
do not bind to cell walls to permeate tissue, then to
retain them in situ by anhydrous freeze substitution.
Their distribution can then be determined by
fluorescence microscopy in dry-cut sections. This
approach works well for tracing pathways of rela-
tively high permeability in the apoplast of higher
plants (Canny, 1986, 1988; Canny & McCully,
1986). It has also been used in fungi to evaluate
changes in permeability of the rind during matu-
ration of sclerotia of Sclerotinia minor (Young &
Ashford, 1992, 1996). Behrmann & Heyser (1992)
evaluated the apoplasmic permeability of Pinus
sylvestris–Suillus bovinus ectomycorrhizas by the
latter approach. They treated the mycorrhizas of
intact seedlings with the anionic tracer sulpho-
rhodamine G for 20 h and examined the distribution
of sulphorhodamine in sections of anhydrously
processed tissue. They found, in contrast to the
results with E. pilularis–P. tinctorius and Pisonia,
that the sheath of their Pinus sylvestris–Suillus
bovinus mycorrhizas was permeable and that sulpho-
rhodamine G penetrated as far as the endodermis.
There are several possible explanations for the
differences between the two sets of results : that
calcofluor had been lost from the walls of the sheath
and cortex in the eucalypt mycorrhizas during the
wash period giving a false negative result ; that the
two types of mycorrhiza are permeable to different
degrees; or that experimental conditions and state of
the mycorrhizal roots differed. It is clearly important
to revisit the issue of the permeability of E.
pilularis–P. tinctorius mycorrhizas, using tracers that
do not bind to walls, and anhydrous processing
techniques. It is also desirable to apply fluoro-
chromes to undisturbed mycorrhizal root systems of
intact seedlings since leakiness due to damage is
minimized, and water and solutes might enter via
bulk flow rather than diffusion (see Moon et al.,
1986; Aloni et al., 1998).
Here we describe experiments where the fluores-
cent tracer 8-hydroxypyrene-1,3,6-trisulphonate
(PTS) has been added to otherwise undisturbed
whole mycorrhizal root systems of seedlings of E.
pilularis produced on agar and inoculated with P.
tinctorius for various lengths of time. Following
treatment, the ectomycorrhizas are anhydrously
freeze-substituted, dry-cut, and examined byfluores-
cence microscopy. Also described are parallel experi-
ments using lanthanum (La) as an apoplasmic tracer,
in conjunction with anhydrous freeze substitution,
to clarify further the permeability of the eucalypt–
Pisolithus sheath. Since La is believed to permeate
wall spaces of diameter as small as 2 nm (Evert et al.,
1985), the data better indicate where permeability
barriers to ions occur in the mycorrhizas. Problems
of La redistribution during tissue processing, which
would confound the results, are avoided by not using
chemical fixation and conventional embedding
methods. As a further check, frozen-hydrated speci-
mens were also analysed directly in the SEM after
planing in the cryoultramicrotome, eliminating all
other tissue preparation steps. The potential of
ectomycorrhizal roots (as distinct from the extra-
radical hyphae) for ion uptake, and whether they
have a direct role as uptake structures or are merely
structures for exchange, are considered. Our findings
are discussed in relation to the observed upregulation
of hydrophobin genes that occurs during sheath
proliferation.
Pure cultures of Pisolithus tinctorius (Pers.) Coker &
Couch, (isolate 024, a culture reisolated by D. A.
Orlovich and J. W. G. Cairney from a mycorrhiza
that had been synthesized axenically with isolate DI-
15) were used in all experiments. Stock cultures were
maintained on Modified Melin-Norkrans (MMN)
agar medium (Marx, 1969) at 24°C, and subcultured
every 8 wk. Plugs of inoculum (8 mm) were cut from
the edges of actively growing colonies and trans-
ferred to fresh agar plates to produce 3-wk-old
mycelium for induction of ectomycorrhizas (Piche! &Fortin, 1982). Seeds of Eucalyptus pilularis Smith
from Royston Petrie Seeds, Kenthurst, Australia
were surface-sterilized by soaking in 70% ethanol
with 0.5% Teepol for 10 min followed by sodium
hypochlorite solution (4% available chlorine) for a
further 25 min with agitation. After three rinses in

RESEARCH Exodermis and sheath impermeability in ectomycorrhizas 335
sterile deionised water, seeds were germinated on
0.4% water agar at 24°C in the dark (Ashford et al.,
1989). Mycorrhizas were synthesized using a
modified version of ‘simple system’ plates
(Duddridge, 1986; Peterson & Chakravarty, 1991).
The rims of the bases and lids of sterile 90¬14 mm
plastic Petri dishes were notched with a hot needle
and the dishes filled with 15 ml of 0.8% water agar.
Seedlings 5–10 d old with 1–2 cm-long main roots
producing first order lateral roots were aseptically
transferred to these plates, their shoots protruding
through the notch and held in place by sealing the
notch with sterile Vaseline petroleum jelly. Two or
three plugs of inoculum were aseptically placed
around the root system. Each plate was watered with
1 ml of sterile nutrient solution before being sealed
with Parafilm. The nutrient solution contained the
following (quantities in millimoles per litre) : ammo-
nium 2.8, boron 1.7, calcium 0.33, chloride 4.3,
copper 0.033, ferric iron 8.7, magnesium 0.45,
molybdenum 0.014, manganese 0.33, potassium 6.0,
phosphate 4.1, sodium 0.30, sulphate 0.94, and zinc
0.13. The plates were incubated vertically inside a
Perspex chamber (Wong & Fortin, 1989) at approx.
23°C. Aluminium foil was placed over the bottom
half of the Petri dishes to reduce exposure of roots to
light. The seedlings received a photon flux density of
approx. 200 µmol m−# s−" in the visible range for 13 h
each day, and were watered as necessary with sterile
deionized water. The bottom and sides of the
Perspex chamber were lined with filter paper and
sprayed weekly with deionized water. The growth
chamber was cleaned routinely with 70% ethanol.
Fluorochrome experiments
Twenty-four dishes containing 3–10-wk-old healthy,
growing mycorrhizas were allocated randomly to
four fluorescent-tracer exposures: control, 15 min, 3
h and 24 h. The dishes were placed in a randomized
Latin-square arrangement with light at photon flux
density of c. 54 µmol m−# s−" from fluorescent tubes
for the whole of the short treatments and for 8 h of
the long treatment. Each treatment dish was gently
filled with 0.1% w}v 8-hydroxypyrene-1,3,6-trisul-
phonate (PTS) solution through a hole of approx. 6
mm diameter made with a soldering iron in the lid
about 1 cm from the edge of the dish and the seedling
stem. Typically, 35 ml of solution was required to
flood the root system completely. The hole was
sealed with Parafilm and the dish replaced under the
lights. After the appropriate time the shoots were cut
off outside the dish, the root systems left intact for 5
min to allow transpiration flow to cease and the
solution was gently removed. Mycorrhizas were
grasped with fine forceps, cut off with fine scissors
under the dissecting microscope, and plunge-frozen
in liquid propane cooled to near freezing point with
liquid N#, using a home-made free-fall plunging
device (G. C. Cox, pers. comm.). To avoid wounding
effects and pseudoreplication (Hurlbert, 1984), only
one mycorrhiza was taken from each plant. Samples
were freeze-substituted in tetrahydrofuran (Pa/ lsga/ rdet al., 1994) over molecular sieve for 3 d at ®80°C,
graduallywarmed to®30°Cand left for 24h,warmed
to 6°C and left for a further 1 h, then transferred to
a glove box flushed with dry N#
gas (Fitzgerald &
Allaway, 1991; Orlovich & Ashford, 1995). In the
dry box they were infiltrated over molecular sieve
with Spurr’s resin (Spurr, 1969) in 1-day steps of
30, 60, 90 and 100% at 23°C. Samples were flat-
embedded and polymerized at 60°C for 48 h and the
blocks were stored in a desiccator until sectioning.
Transverse sections 2.5 µm thick were cut with dry
glass knives in an air-conditioned room, placed on
dry glass slides pre-coatedwith chrome alum}gelatin,
and flattened by firm pressure using a pencil eraser
and cellophane (Canny & McCully, 1986). The
sections were mounted in nonfluorescent immersion
oil under a coverslip, sealed with nail polish and
stored in a desiccator in the dark until microscopy.
They were examined using a Zeiss Axiophot micro-
scope (Zeiss Goettingen, Germany) with bright-
field, Nomarski differential interference contrast,
epifluorescence using 365 nm excitation, 395 nm
dichroic mirror and 420 nm barrier filters, and
photographed on Kodak Select 100 or 400 ISO
colour-transparency film. Colours are described in
the text according to Kornerup & Wanscher (1978).
Laboratories were air-conditioned at 21–25°C.
Lanthanum treatment and X-ray microanalysis
Procedures were identical to those used for fluoro-
chrome tracing, with the following exception. Six
plates with healthy growing mycorrhizas 5–6 months
after inoculation were gently filled with 1% w}v
lanthanum nitrate solution (31 mM La(NO$)$) and
left for 24 h (of which 8 h were illuminated at c. 54
µmol m−# s−") at c. 24°C. After anhydrous freeze
substitution as described above, transverse sections
0.5 µm thick were cut with dry glass knives in an air-
conditioned room, placed on a parlodion- and
carbon-coated nickel slot grid, and covered with
another parlodion film. They were stored in a
desiccator until electron microscopy, which usually
took place within a few hours. These dry-sectioned
unstained specimens were examined at 120 kV in a
Philips (Eindhoven, The Netherlands) CM12 Scan-
ning Transmission Electron Microscope (STEM)
equipped with an Edax ultrathin-window energy-
dispersive X-ray detector and Edax DX4 analyser,
using a Be low background specimen holder. Images
were recorded in bright-field STEM mode in a
512¬400 pixel array. Energy-dispersive X-ray
microanalyses of individual areas of interest were
carried out with reduced raster sizes for 100 s. X-ray
dot maps were made by defining a 170 eV energy

336 RESEARCH P. A. Vesk et al.
window around the characteristic wavelength of each
element of interest and recording X-ray counts at
each point in a dwell time of 1000 ms in a 100¬128
pixel array, or 200 ms in a 200¬256 pixel array.
Background maps were collected from a window
between 4.09 and 4.33 keV.
Frozen-hydrated specimens
In a further experiment, frozen-hydrated specimens
were examined in SEM (after Romeo, 1996). Single
mycorrhizas (24-h La treatment and controls) were
placed on a brass block which was then immersed in
liquid N#for 45 s and the mycorrhiza fractured with
a cooled razor blade. Alternatively, mycorrhizas
were placed on aluminium pins, plunged in liquid N#
with hand-held forceps, planed with glass knives
with a Reichert-Jung FC-S cryoultramicrotome
(Leica, Vienna, Austria), then mounted in a brass
block under liquid N#. At all steps between oper-
ations the specimens, pins and brass blocks were
kept submerged in liquid N#in a covered, insulated
container. Each chilled block with specimen was
transferred as quickly as possible to the specimen
chamber (which was pumped down immediately) of
a Philips 505 SEM fitted with an Edax windowless
energy-dispersive X-ray detector and Edax DX4
analyser. The specimens remained frozen through-
out examination, which lasted up to 40 min, before
they started to freeze-dry rapidly and observation
halted. Images were recorded on Ilford FP4 roll
film. Energy dispersive X-ray microanalyses were
carried out by rastered 100-s scans of individual
areas with an accelerating voltage of 10 or 15 kV.
Fluorochrome localizations
Mycorrhizal roots showed a structure typical of this
association, with a well developed sheath and a
Hartig net restricted to the epidermis. There was a
distinct pattern of autofluorescence, and careful
comparison of treatments with controls was necess-
ary to differentiate this from PTS fluorescence. Fig.
1a shows a control section. The sheath and Hartig
net autofluoresced a high-intensity greyish yellow
while most of the root cell walls showed light-blue
autofluorescence, highest in intensity in the xylem
(not illustrated) and also seen in exodermal and other
cortical cell walls in this section. Autofluorescence of
the epidermal cell walls was a low-intensity greyish
yellow and so autofluorescence in the Hartig net was
dominated by that of the fungus, and similar to the
sheath. In the exodermis, the outer tangential walls
and outer part of the radial walls autofluoresced
yellowish white. As a result there was a distinct
difference in autofluorescence between the Hartig
net and outer tangential walls of the exodermis (Fig.
1a).
All 15-min treatments showed PTS fluorescence
only in the walls of free external hyphae and of
hyphae at the surface of the sheath (Fig. 1b). No
samples showed deeper localization of the fluoro-
chrome. The PTS fluorescence (high-intensity vivid
blue) was readily distinguished from the greyish-
yellow autofluorescence in the sheath, and also from
the light-blue and yellowish-white autofluorescence
of the root cells (Fig. 1b).
Mycorrhizas exposed to PTS solution for 3 or 24
h showed variable fluorochrome localization in the
sheath. This variability was not correlated with the
duration of treatment, but differed according to
the state of development of the mycorrhiza and to
whether or not it was surrounded by agar. In most of
the mycorrhizas exposed to PTS solution for 3 or 24
h, PTS fluorescence (high-intensity vivid blue) was
detected in the outermost sheath hyphae and in the
Hartig net and innermost sheath, but not throughout
the intervening sheath region (Fig. 1c). These
mycorrhizaswere fully developed andhad a cemented
sheath which autofluoresced yellow. Fluorochrome
in the Hartig net region was clearly restricted to the
apoplast (Fig. 1c,d) and often extended along the
interface between plant and fungal cells in the radial
walls of the epidermis and the outer tangential wall
of the exodermis (Fig. 1d). There was a sharp cut-off
of PTS fluorescence at the radial walls of the
exodermis, which show whitish autofluorescence
(Fig. 1c,d).
Fluorochrome was not detected inside the exo-
dermal Casparian band in mature regions of the
mycorrhizas (Fig. 1c,d). However, in specimens
sectioned close to the mycorrhizal tip, fluorochrome
was observed in walls of the layer of cortical cells
internal to the exodermis, suggesting that the
Casparian band had not fully developed in this
region (Fig. 1e). This PTS fluorescence was of lesser
intensity than elsewhere in the section. In some cases
there was patchy PTS fluorescence in the sheath
(Fig. 1e,f). The sheath shown in Fig. 1e was thin,
with loosely arranged but cemented hyphae. Never-
theless, only the yellow autofluorescence was present
in the middle part of the sheath, and the vivid blue
PTS fluorescence was restricted to the outer sheath,
Hartig net and innermost sheath. This pattern was
typical, and rarely was PTS fluorescence seen
spanning the sheath to connect the inner and outer
regions as in Fig. 1f. In the latter case the cemented
region of the sheath was quite thin and Hartig net
development was not complete.
In a few cases, PTS fluorescence in the mycorrhizal
sheath was more extensive. This was much rarer
than the severely restricted permeation as already
described. Fig. 1g shows one example (3-h treatment)
of a mycorrhiza from the agar surface in which PTS
fluorescence was present throughout the entire
sheath and Hartig net as far as the exodermis. The
PTS fluorescence was located in the cell walls of the

RESEARCH Exodermis and sheath impermeability in ectomycorrhizas 337
(a)
s
ep
ex
c
(b)
ep
s
(e)
ep
ex
c
s
(d)
ep
ex
s
(c)
s
ep
ex
(f)
s
ep
(g)
epex
c
s
✻
Fig. 1. Fluorescence microscopy of transverse sections of mycorrhizas : ep, epidermis; ex, exodermis; c, the
next layer of cortex; s, sheath. (a) Autofluorescence in a control specimen. Bar, 20 µm. (b) PTS fluorescence
(vivid blue) after a 15-min treatment is restricted to the walls of the outermost sheath hyphae. Bar, 20 µm. (c)
24-h PTS treatment. Vivid-blue PTS fluorescence is seen in the outer sheath hyphal walls, Hartig net and
innermost sheath, but the intervening sheath region shows only the yellow autofluorescence. Bar, 20 µm. (d)
24-h PTS treatment. Hartig net region at higher magnification. Bar, 10 µm. (e) 3-h PTS treatment. Specimen
sectioned close to the mycorrhizal tip. There are small areas of PTS fluorescence at the inner angles of
exodermal cells (arrows), and in the intervening inner walls of the exodermis. Bar, 20 µm. (f) 3-h PTS
treatment. Patches of alternating PTS fluorescence and autofluorescence in the sheath of a specimen sectioned
close to the mycorrhizal tip. Bar, 10 µm. (g) 3-h PTS treatment. Mycorrhiza of unusually high permeability,
from the agar surface. Walls of all the sheath hyphae and extra-radical hyphae show vivid-blue PTS
fluorescence. Adhering agar is also strongly PTS-fluorescent (*). Bar, 20 µm.
fungal sheath and to a lesser extent in the in-
tercellular spaces but not in the lumen of the fungal
cells. PTS fluorescence was also detected throughout
the adhering agar (Fig. 1g). From the exodermis
inwards, the walls showed autofluorescence identical
with that in the controls (Fig. 1g).
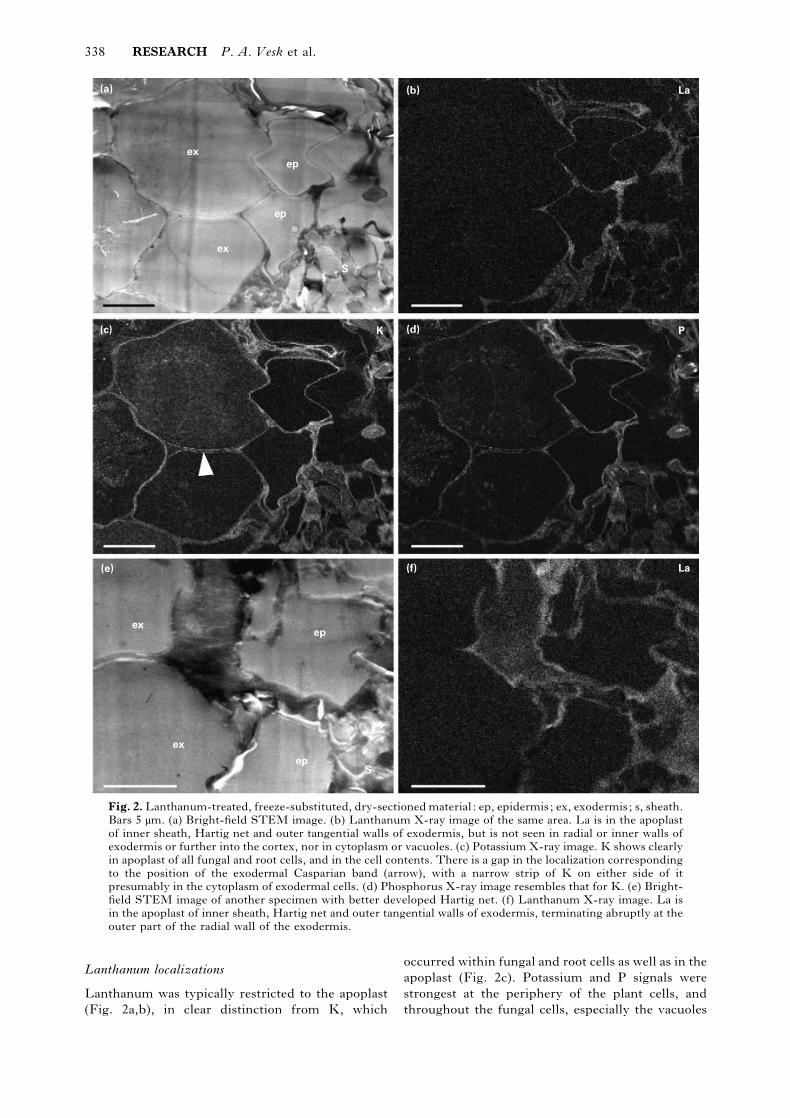
338 RESEARCH P. A. Vesk et al.
(a)
ex
ex
ep
ep
(b) La
(d) P(c) K
(f) La(e)
ex
ex
ep
ep
S
S
Fig. 2. Lanthanum-treated, freeze-substituted, dry-sectioned material : ep, epidermis; ex, exodermis; s, sheath.
Bars 5 µm. (a) Bright-field STEM image. (b) Lanthanum X-ray image of the same area. La is in the apoplast
of inner sheath, Hartig net and outer tangential walls of exodermis, but is not seen in radial or inner walls of
exodermis or further into the cortex, nor in cytoplasm or vacuoles. (c) Potassium X-ray image. K shows clearly
in apoplast of all fungal and root cells, and in the cell contents. There is a gap in the localization corresponding
to the position of the exodermal Casparian band (arrow), with a narrow strip of K on either side of it
presumably in the cytoplasm of exodermal cells. (d) Phosphorus X-ray image resembles that for K. (e) Bright-
field STEM image of another specimen with better developed Hartig net. (f) Lanthanum X-ray image. La is
in the apoplast of inner sheath, Hartig net and outer tangential walls of exodermis, terminating abruptly at the
outer part of the radial wall of the exodermis.
Lanthanum localizations
Lanthanum was typically restricted to the apoplast
(Fig. 2a,b), in clear distinction from K, which
occurred within fungal and root cells as well as in the
apoplast (Fig. 2c). Potassium and P signals were
strongest at the periphery of the plant cells, and
throughout the fungal cells, especially the vacuoles
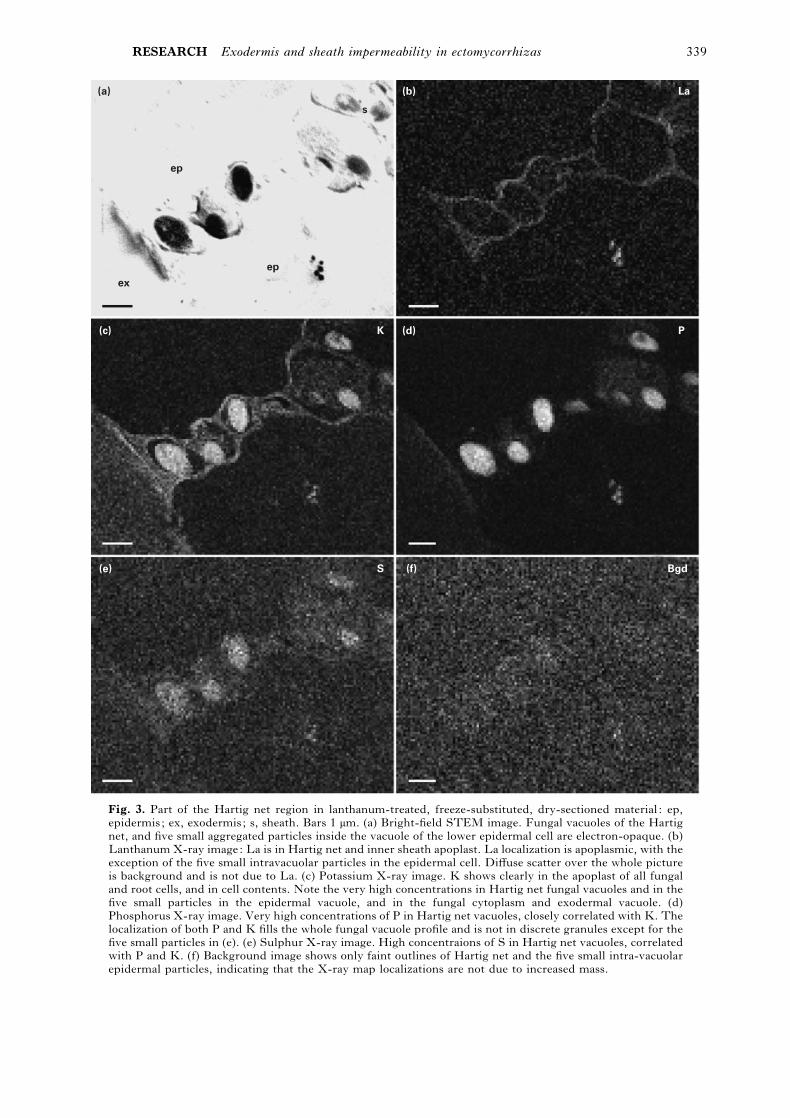
RESEARCH Exodermis and sheath impermeability in ectomycorrhizas 339
(a)
ep
ep
ex
(b) La
(c) (d) PK
(e) (f) BgdS
s
Fig. 3. Part of the Hartig net region in lanthanum-treated, freeze-substituted, dry-sectioned material : ep,
epidermis; ex, exodermis; s, sheath. Bars 1 µm. (a) Bright-field STEM image. Fungal vacuoles of the Hartig
net, and five small aggregated particles inside the vacuole of the lower epidermal cell are electron-opaque. (b)
Lanthanum X-ray image: La is in Hartig net and inner sheath apoplast. La localization is apoplasmic, with the
exception of the five small intravacuolar particles in the epidermal cell. Diffuse scatter over the whole picture
is background and is not due to La. (c) Potassium X-ray image. K shows clearly in the apoplast of all fungal
and root cells, and in cell contents. Note the very high concentrations in Hartig net fungal vacuoles and in the
five small particles in the epidermal vacuole, and in the fungal cytoplasm and exodermal vacuole. (d)
Phosphorus X-ray image. Very high concentrations of P in Hartig net vacuoles, closely correlated with K. The
localization of both P and K fills the whole fungal vacuole profile and is not in discrete granules except for the
five small particles in (e). (e) Sulphur X-ray image. High concentraions of S in Hartig net vacuoles, correlated
with P and K. (f) Background image shows only faint outlines of Hartig net and the five small intra-vacuolar
epidermal particles, indicating that the X-ray map localizations are not due to increased mass.
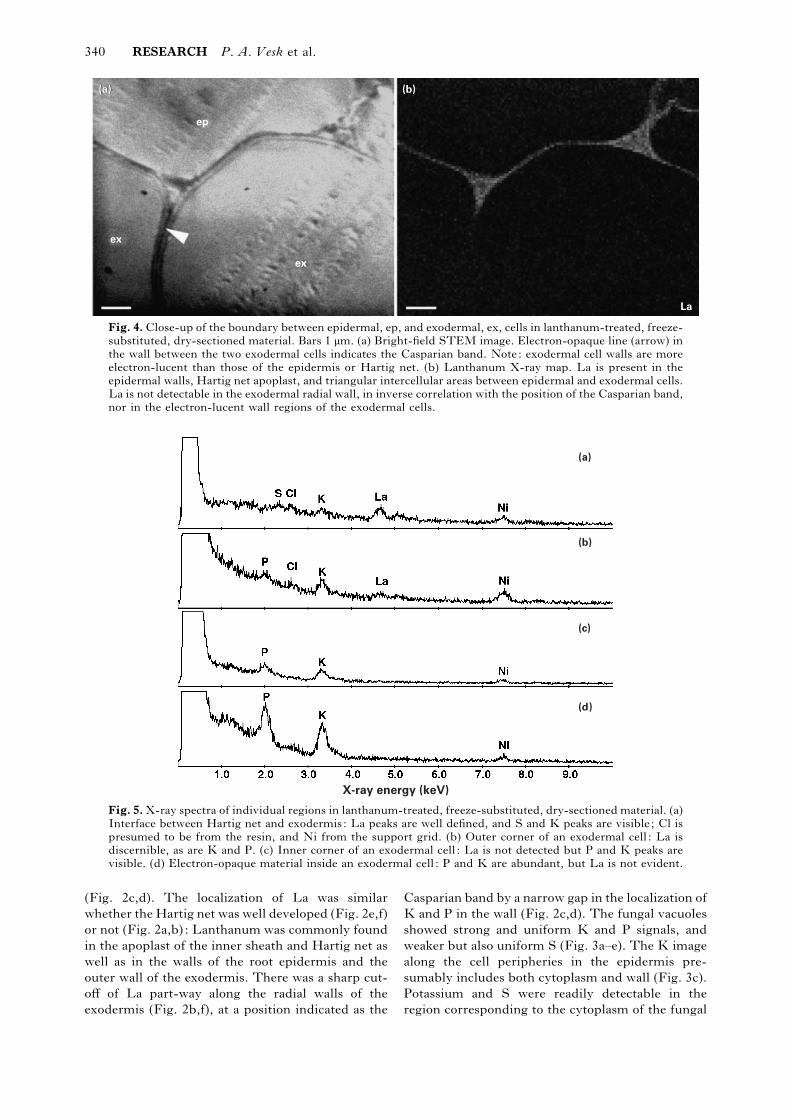
340 RESEARCH P. A. Vesk et al.
La
(b)(a)(a)
ep
ex
ex
Fig. 4. Close-up of the boundary between epidermal, ep, and exodermal, ex, cells in lanthanum-treated, freeze-
substituted, dry-sectioned material. Bars 1 µm. (a) Bright-field STEM image. Electron-opaque line (arrow) in
the wall between the two exodermal cells indicates the Casparian band. Note: exodermal cell walls are more
electron-lucent than those of the epidermis or Hartig net. (b) Lanthanum X-ray map. La is present in the
epidermal walls, Hartig net apoplast, and triangular intercellular areas between epidermal and exodermal cells.
La is not detectable in the exodermal radial wall, in inverse correlation with the position of the Casparian band,
nor in the electron-lucent wall regions of the exodermal cells.
(a)
(b)
(c)
(d)
X-ray energy (keV)
Fig. 5. X-ray spectra of individual regions in lanthanum-treated, freeze-substituted, dry-sectioned material. (a)
Interface between Hartig net and exodermis: La peaks are well defined, and S and K peaks are visible; Cl is
presumed to be from the resin, and Ni from the support grid. (b) Outer corner of an exodermal cell : La is
discernible, as are K and P. (c) Inner corner of an exodermal cell : La is not detected but P and K peaks are
visible. (d) Electron-opaque material inside an exodermal cell : P and K are abundant, but La is not evident.
(Fig. 2c,d). The localization of La was similar
whether the Hartig net was well developed (Fig. 2e,f)
or not (Fig. 2a,b) : Lanthanum was commonly found
in the apoplast of the inner sheath and Hartig net as
well as in the walls of the root epidermis and the
outer wall of the exodermis. There was a sharp cut-
off of La part-way along the radial walls of the
exodermis (Fig. 2b,f), at a position indicated as the
Casparian band by a narrow gap in the localization of
K and P in the wall (Fig. 2c,d). The fungal vacuoles
showed strong and uniform K and P signals, and
weaker but also uniform S (Fig. 3a–e). The K image
along the cell peripheries in the epidermis pre-
sumably includes both cytoplasm and wall (Fig. 3c).
Potassium and S were readily detectable in the
region corresponding to the cytoplasm of the fungal

RESEARCH Exodermis and sheath impermeability in ectomycorrhizas 341
(a) (b)
ep
s
La
Fig. 6. Sheath region of lanthanum-treated, freeze-substituted, dry-sectioned material : ep, epidermis; s,
sheath. Bars 5 µm. (a) Bright-field STEM image. (b) Lanthanum X-ray image. La occurs throughout the
apoplast of the sheath, in extramatrical hyphae, other extramatrical material, the Hartig net apoplast, in
intercellular spaces in the sheath, as well as in the fungal cell walls.
(a) (b)
s
exep
c
s
h
s
s
h
m
ex
ep
Fig. 7. Cryo-SEM images of frozen-hydrated, lanthanum-treated mycorrhiza. Specimens have been planed in
the cryoultramicrotome before being put into the microscope. (a) Low-magnification image of a slightly oblique
longitudinally planed mycorrhiza shows extraradical hyphae, h; well formed sheath, s ; root apical meristem
region, m; exodermal cells, ex; and epidermal cells, ep. Bar, 100 µm. (b) Higher magnification shows sheath,
s ; epidermis, ep; well developed Hartig net, exodermis, ex; and the next layer of cortex, c. Bar, 25 µm.
cells, and there was also a distinct but weak signal for
P in this region (Fig. 3c–e). In two specimens, a few
small granules inside a plant cell showed La and
these also gave signals for K, P, S (Fig. 3a–f) and Cl.
In no other case was there any detectable La inside
the plasma membrane of any cell, either fungal or
plant. The background signal was almost uniform,
with only slightly increased intensity in areas of high
tissue density (Fig. 3f). Close examination of the
exodermis showed the presence of La in the outer
part of the outer-tangential wall but not in the inner
part of this wall (Fig. 4a,b), and in triangular
intercellular areas between epidermal and exodermal
cells (Fig. 4b) but not in the exodermal radial wall.

342 RESEARCH P. A. Vesk et al.
(a)
(b)
(c)
(d)
(e)
X-ray energy (keV)
Fig. 8. X-ray spectra of individual regions in planed lanthanum-treated, frozen-hydrated material. (a) Outer
part of the sheath: P and K are the most prominent peaks, with smaller peaks for Na, S and Cl, and clearly
defined La peaks. Cu and Fe are presumed to be in the block on which the specimen was mounted. (b) Root
epidermal wall : La is detectable, and Na, Mg, P, S, Cl and K are prominent. (c) Contents of an exodermal cell
(probably vacuole) : La is barely discernible. (d) Cortical cell inside the exodermis: typically, La is not detected.
(e) Cortical cell inside the exodermis near the mycorrhizal tip showing a rare occurrence of La in the cortex.
The bright-field image shows an electron-opaque
region in the radial walls where the La signal was not
detected (Fig. 4a), corresponding to the location of
the Casparian band. The electron-lucence of the
exodermal walls (Fig. 4a), and the absence of La
from them (Fig. 4a,b), are consistent with suberi-
zation in these walls.
Spectra of individual regions showed well defined
La peaks in the interface between the Hartig net and
exodermis, and S and K peaks could also be
discerned (Fig. 5a). It is presumed that Cl is from the
resin, which was not chloride-free, and Ni from the
support grid. At the outer corner of an exodermal
cell, La, K and P were present (Fig. 5b). At an inner
corner of an exodermal cell, however, La was not
detected, although the P and K peaks remained (Fig.
5c). Electron-opaque material inside an exodermal
cell did not show La, but P and K peaks were clearly
present (Fig. 5d): this was the typical situation inside
the cells. In view of the impermeability to PTS of
the outer and middle regions of the sheath we
examined these regions very carefully for La. All
samples, including aerial ectomycorrhizas and re-
gardless of sheath development, showed La through-
out the sheath (Fig. 6a,b).
Localizations in frozen-hydrated specimens
Frozen-hydrated specimens showed the structure of
the mycorrhizas clearly, with extraradical hyphae,
sheath and Hartig net plainly discernible (Fig. 7a,b).
The resolution of analysis on these bulk specimens
did not permit a restricted location of analysis, in
particular precluding distinction between extra-
cellular material and cytoplasm. Monte Carlo mod-
elling, assuming solid oxygen (a conservative ap-
proximation of ice) and density 1 g cm−$, suggested
that at 15 kV, as in Fig. 8, La X-rays could be
generated within a volume of up to 2.5 µm depth and
2 µm lateral radius, and that the P interaction volume
would be slightly larger. When the raster area for
analysis was on the outer part of the sheath, P and K

RESEARCH Exodermis and sheath impermeability in ectomycorrhizas 343
were the most prominent peaks, with smaller peaks
for Na, S and Cl, and clearly defined La peaks (Fig.
8a). Copper and Fe were presumed to be in the block
on which the specimen was mounted; La was also
clearly detectable in analyses with the raster area on
the root epidermal wall (Fig. 8b), but was only
marginally discernible when the raster was on the
contents (probably vacuole) of an exodermal cell
(Fig. 8c). In a cell of the cortex inside the exodermis,
La was not detected (Fig. 8d), although in two
specimens regions near the mycorrhizal tip did show
La in the cortex (Fig. 8e).
The results are consistent with our earlier obser-
vations for Eucalyptus and Pisonia mycorrhizas that
the fungal sheath of mature mycorrhizas is im-
permeable to fluorochromes. PTS is a much better
tracer than calcofluor in that it does not rely on
attraction to cell walls for tissue retention, but
requires anhydrous processing for maintenance at
the sites to which it has permeated. There is now a
body of evidence indicating that the anhydrous
freeze-substitution techniques employed here pre-
vent loss and redistribution of water-soluble fluoro-
chromes including PTS (Canny & McCully, 1986;
Fitzgerald & Allaway, 1991). The general agreement
of the localizations in frozen-hydrated material
presented here, although necessarily at lower res-
olution because of the several µm$ interaction
volume, is consistent with this.
One cannot assume that walls impermeable to
PTS are also impermeable to inorganic ions and
water. Lanthanum is a much better indicator of
permeability to ions and has been used as an ion
tracer in a number of investigations. However, most
studies have followed application of lanthanum
nitrate (La(NO$)$) with chemical fixation and}or
processing techniques where conditions were not
anhydrous. In these cases La might only have been
maintained in situ by electrostatic binding and could
have been lost, displaced or redistributed during
processing and sectioning: therefore the results have
little credibility in determining barriers to ion
permeability. The use of anhydrous techniques
avoids these problems and there is now good
evidence that ions are retained in situ by the
procedure (Pa/ lsga/ rd et al., 1994; Orlovich & Ash-
ford, 1995). To our knowledge ours is the only study
in which anhydrous processing rather than a chemi-
cal fixation has been used with La to trace apoplasmic
pathways. Use of PTS, which is anionic, and La,
which is cationic, in conjunction with freeze sub-
stitution, allows us to exclude electrostatic binding
as the primary reason for the distribution obtained in
the apoplast of these mycorrhizas.
The use of intact mycorrhizal seedlings minimizes
root disturbance and damage, thereby reducing the
potential for transient leakages (Moon et al., 1986).
The plants were capable of transpiration throughout
the experimental treatments, with potential for flow
of solution in the apoplast as well as for diffusion
(Canny, 1995; Aloni et al., 1998). In this respect
these experiments resemble a field situation more
closely than previous ones using excised roots with
sticky-wax-sealed cut ends. Restriction of sampling
to one mycorrhiza per plant also ensures that there is
no leakage into cut surfaces which might affect
results if a series of samples were taken from the
same plant. Lanthanum is known to be toxic and
might have physiological effects even at the low
concentration used here (Lazzaro & Thomson,
1992). This toxicity might account for the only two
instances of intracellular La observed in the plant
cells. The data, however, do not support extensive
‘endocytosis ’ of the tracer as commonly reported in
studies where chemical fixation has been used
(Robards & Robb, 1974; Evert et al., 1985) but are
more consistent with the occasional presence of
damaged cells than with uptake by endocytosis. It
should also be noted that the effects of La on ion
transporters might have influenced the distribution
of other ions (Lewis & Spalding, 1998).
The presence of an exodermis with Casparian
bands and suberized lamellae is well established for
eucalypt mycorrhizas, but it has been assumed that
these are barriers to apoplasmic continuity only by
extrapolation to results with nonmycorrhizal euca-
lypt roots (Ashford et al., 1989). Most evidence from
the literature suggests that the Casparian bands in
the exodermis of nonmycorrhizal roots are im-
permeable to ions, although views differ on their
permeability to water (Peterson, 1987; Peterson et
al., 1993; Varney et al., 1993; Canny & Huang,
1994; Peterson & Cholewa, 1998; Steudle & Peter-
son, 1998; Zimmermann & Steudle, 1998). It has
been shown here that the exodermal Casparian
bands are a barrier to penetration of both tracers into
the cortex of all the mycorrhizas, regardless of the
degree of permeation of PTS or La into the sheath.
The exclusion of La indicates that any pores in the
Casparian bands in the exodermis of the mycorrhizas
are small enough to exclude ions. Similarly, ex-
clusion is observed for the suberin lamellae, La
being restricted to the outer region of the outer
tangential wall of the exodermis. Only in very young
parts of the root, where there is reason to expect that
Casparian bands would not yet be fully developed,
did the apoplasmic tracers penetrate more deeply.
This supports the literature and establishes that the
exodermal Casparian bands in mycorrhizal roots
have a similarly low permeability to those of
nonmycorrhizal root systems and to the tight
junctions of animal cells (Madara, 1988). The finding
that the Casparian bands of the exodermis are a
barrier to both PTS and La$+, contrasts with the
findings of Zimmermann & Steudle (1998) that the

344 RESEARCH P. A. Vesk et al.
exodermis of corn roots allows PTS leakage. It is
consistent with their suggestion that the leakage of
PTS into the stele which they observed might be via
sites of emergence of lateral roots which breach the
exodermis. In ectomycorrhizas the apical meristem
is small and cells differentiate much closer to the
apex than in nonmycorrhizal roots. This is also true
of the exodermis, suggesting that the exodermal
apoplasmic barrier might operate much closer to the
apex in mycorrhizas than in nonmycorrhizal roots.
Mycorrhizas in contact with the wet agar surface
remained permeable to PTS while those developed
in air excluded this fluorochrome even at an early
stage. Fluorochrome was excluded from the in-
tercellular spaces and cementing material and from
the cell walls of individual hyphae in the outer sheath
whether cemented or not. The walls of these outer
sheath hyphae not only show reduced permeability,
but also tend to exclude solution from the sheath as
a whole, presumably by effects of surface tension.
Thus, mycorrhizas developed in air rapidly become
hydrophobic and nonwettable well before sheath
development is complete and the sheath is cemented,
whereas those submerged in the agar or touching its
wet surface do not. The most likely explanation for
this lies in the deposition of hydrophobins in the
walls of the sheath hyphae, providing a reason for the
upregulation of hydrophobin genes known to occur
early in sheath development and during sheath
proliferation in Eucalyptus globulus–Pisolithus tinc-
torius mycorrhizas (Martin & Tagu, 1995; Tagu &
Martin, 1995, 1996; Tagu et al., 1996, 1998).
Hydrophobins would be expected to form a hydro-
phobic layer on walls in contact with air, but to
diffuse away as monomers where walls are in contact
with water or, in this case, agar (Wessels, 1993, 1994,
1996, 1999). The higher concentrations of S found in
the vacuoles of the Hartig net hyphae, than in
vacuoles of free-living P. tinctorius mycelium
(Orlovich & Ashford, 1993; Cole et al., 1998) would
provide the S necessary for the large amounts of
cysteine required in hydrophobin production.
Thus, the observation that the aerial mycorrhizas
impermeable to PTS are permeable to La is
fascinating. The data imply that the sheath apoplast
is not totally impermeable as we formerly thought,
but that there are limits on the size of molecules
which can move across. The outer sheath is less
permeable than unmodified hyphal or plant cell
walls, but more permeable than the Casparian bands.
This should not come as a surprise. It has long been
known that tight junctions, which occlude the
apoplasmic spaces in animal systems, are selectively
permeable, and their permeability to small molecules
and La ions appears to differ greatly between
different tissues (Madara, 1988; Anderson & Van
Itallie, 1995; Chau & Lu, 1996). Although the two
are not equated in a structural sense, there is a
parallel with the sheath apoplasmic barrier which
might allow ions to move across but restrict larger
organic molecules such as those exchanged across
the interface, so that they remain trapped. A
mycorrhizal root submerged in a watery medium for
some time might therefore still be able to take up
ions via the apoplast, although it is generally
nonwettable and not very permeable.
Our long-term observations suggest that eucalypt
ectomycorrhizas tend to form best and most fre-
quently in the air above the agar. In growth pouches
they do not form in any liquid in the bottom of the
pouch. They are often found in air pockets in soil in
the wild, or at the edge of pots in the glasshouse, an
observation also made for other ectomycorrhizas in
the field and in microcosms (Francis & Read, 1994).
Such mycorrhizas formed in air are invariably
nonwettable and water-repellent, and when placed in
a liquid they float and trap air. Attention has been
drawn to the extensiveness of the extraradical
mycelium and its importance in nutrient acquisition
for a range of ectomycorrhizal associations, including
those with European isolates of P. tinctorius (Read,
1984, 1991, 1993; Smith & Read, 1997). Read has
proposed that individual ectomycorrhizal roots have
little role in nutrient uptake and considers that they
act as a food base for mycelial outgrowth and also as
structures of storage and exchange. Taking this view
further, eucalypt mycorrhizal roots, though sub-
terranean, can be viewed as aerial structures that
have more in common with fungal fruiting bodies
than with plant roots, primarily structures of storage
and exchange where the interface between partners,
supplied via uptake and symplasmic transport from
the connected extraradical mycelium (and the plant),
must be protected by a relatively impervious sheath.
Indeed, the large amount of closely packed fungal
tissue in the mycorrhizal sheath suggests a high
demand for oxygen which would not be easily
satisfied in a submerged structure with water-filled
intercellular spaces. A high requirement for oxygen
is to be anticipated during sheath expansion, for
active uptake of solutes into hyphae (Harley, 1969)
and for efficient nutrient transfer at the symbiont
interface. Retention of air-filled intercellular space
and the general nonwettability of the structure as a
whole would greatly enhance the capacity for gas
exchange, providing a clear role for hydrophobins
early in ectomycorrhizal root development, similar
to that seen in fruiting body development (Wessels,
1992; Wessels et al., 1995; Lugones et al., 1999).
However, hydrophobins are not the only substances
which might modify permeability of walls and the
extracellular matrix (Kershaw & Talbot, 1998;
Wo$ sten et al., 1999), and the possibility that other
substances could contribute to the impermeability of
the sheath in mycorrhizas should not be discounted.
It is important to emphasize that the isolate of P.
tinctorius used in this study is strongly hydrophobic.
Differences in hydrophobicity between mycorrhizas

RESEARCH Exodermis and sheath impermeability in ectomycorrhizas 345
formed with either hydrophilic or hydrophobic fungi
could explain the discrepancy in results between
different groups on permeability of different mycor-
rhizas (Behrmann & Heyser, 1992) and on expression
of ectomycorrhizins during mycorrhizal synthesis
(Guttenberger, 1994). Unestam (1991) found that
mycorrhizal mantles formed by 14 of 20 fungal
species were highly hydrophobic, although the other
six were hydrophilic. Extramatrical hyphae and
cords of fungi which produce a hydrophobic mantle
were themselves hydrophobic, except for hyphal tips
in contact with a soaked substrate (Unestam, 1991;
Unestam & Sun, 1995). Mycorrhizas produced by
the group of hydrophilic fungi might not have
upregulated expression of hydrophobin genes, could
produce lower amounts of hydrophobins, might be
more permeable, and might exhibit more direct
uptake from the environment. This is an avenue for
further study.
The permeability of the Hartig net contrasts with
that of the sheath. One explanation for the frequency
of cases where the Hartig net and inner sheath
contain significant amounts of PTS, whereas the
bulk of the sheath contains little or none, could be
that there is significant radial flow with exclusion of
this tracer, but not of water, at the exodermis. This
could concentrate tracer in the apoplast of the Hartig
net region as a sump (Canny, 1990; Varney &
Canny, 1993). We think this an unlikely explanation
here, since those sheaths that are permeable show no
obvious gradients, whereas most sheaths show
discontinuities in PTS distribution. In the latter, the
pattern of fluorescence often suggests that PTS
moves outwards from the Hartig net into some inner
parts of the sheath, whereas the material of the outer
and middle sheath is impermeable. Our data are
better explained by movement of PTS longitudinally
within the apoplast of the Hartig net from more
permeable regions, such as young or wet parts of
mycorrhizas, to hydrophobic regions. This is more
likely to be important in mycorrhizal roots of whole
transpiring plants, where mass flow of solution as
well as diffusion might occur. In the sealed excised
roots used previously (Ashford et al., 1988, 1989)
movement would certainly be restricted to diffusion.
The Casparian bands in the radial walls of the
exodermis are clearly impermeable to ions, but the
sheath has variable and differential permeability. We
can therefore now rephrase the hypothesis presented
by Ashford et al. (1989): there is an isolated shared
apoplasmic compartment at the interface between
partners, although it is not as tightly sealed on the
outside as we previously thought.
We thank the Electron Microscope Unit at The University
of Sydney for facilities, Malcolm Ricketts for assistance,
and the Australian Research Council for research grants.
W. G. Allaway thanks Professor A. R. Hardham and the
Plant Cell Biology Group, Research School of Biological
Sciences, Australian National University, and the School
of Biological Science and Life Sciences Imaging Unit, The
University of New South Wales, for facilities during study
leave.
Aloni R, Enstone DE, Peterson CA. 1998. Indirect evidence
for bulk water flow in root cortical cell walls of three
dicotyledonous species. Planta 207 : 1–7.
Anderson JM, Van Itallie CM. 1995. Tight junctions and the
molecular basis for regulation of paracellular permeability.
American Journal of Physiology 269 : G467–G475.
Ashford AE, Allaway WG, Peterson CA, Cairney JWG. 1989.Nutrient transfer and the fungus–root interface. Australian
Journal of Plant Physiology 16 : 85–97.
Ashford AE, Peterson CA, Carpenter JL, Cairney JWG,Allaway WG. 1988. Structure and permeability of the fungal
sheath in the Pisonia mycorrhiza. Protoplasma 147 : 149–161.
Behrmann P, Heyser W. 1992. Apoplastic transport through
the fungal sheath of Pinus sylvestris}Suillus bovinus ectomy-
corrhizae. Botanica Acta 105 : 427–434.
Canny MJ. 1986. Water pathways in wheat leaves. III. The
passage of the mestome sheath and the function of the suberised
lamellae. Physiologia Plantarum 66 : 637–647.
Canny MJ. 1988. Water pathways in wheat leaves. IV. The
interpretation of images of a fluorescent apoplastic tracer.
Australian Journal of Plant Physiology 15 : 541–555.
Canny MJ. 1990. Fine veins of dicotyledon leaves as sites for
enrichment of solutes of the xylem sap. New Phytologist 115 :
511–516.
Canny MJ. 1995. Apoplastic water and solute movement: new
rules for an old space. Annual Review of Plant Physiology and
Plant Molecular Biology 46 : 215–236.
Canny MJ, Huang CX. 1994. Rates of diffusion into roots of
maize. New Phytologist 126 : 11–19.
Canny MJ, McCully ME. 1986. Locating water-soluble vital
stains in plant tissues by freeze-substitution and resin-
embedding. Journal of Microscopy 142 : 63–70.
Chau Y-P, Lu K-S. 1996. Differential permeability of blood
microvasculatures in various sympathetic ganglia of rodents.
Anatomy and Embryology 194 : 259–269.
Cole L, Orlovich DA, Ashford AE. 1998. Structure, function,
and motility of vacuoles in filamentous fungi. Fungal Genetics
and Biology 24 : 86–100.
Duddridge JA. 1986. The development and ultrastructure of
ectomycorrhizas. III. Compatible and incompatible inter-
actions between Suillus grevillei (Klotzsch) Sing. and 11 species
of ectomycorrhizal hosts in vitro in the absence of exogenous
carbohydrate. New Phytologist 103 : 457–464.
Evert RF, Botha CEJ, Mierzwa RJ. 1985. Free-space marker
studies on the leaf of Zea mays L. Protoplasma 126 : 62–73.
Fitzgerald MA, Allaway WG. 1991. Apoplastic and symplastic
pathways in the leaf of the grey mangrove Avicennia marina
(Forsk.) Vierh. New Phytologist 119 : 217–226.
Francis R, Read DJ. 1994. The contributions of mycorrhizal
fungi to the determination of plant community structure. Plant
and Soil 159 : 11–25.
Guttenberger M. 1994. The protein complement of ectomy-
corrhizas. In: Varma A, Hoch B, eds. Mycorrhiza structure,
function, molecular biology and biotechnology. Berlin, Germany:
Springer Verlag, 59–77.
Harley JL. 1969. The biology of mycorrhiza, 2nd edn. London,
UK: Leonard Hill.
Hurlbert SH. 1984. Pseudoreplication and the design of eco-
logical field experiments. Ecological Monographs 54 : 187–211.
Kershaw MJ, Talbot NJ. 1998. Hydrophobins and repellents :
proteins with fundamental roles in fungal morphogenesis.
Fungal Genetics and Biology 23 : 18–33.
Kornerup A, Wanscher JH. 1978. Methuen handbook of colour,
3rd edn. London, UK: Methuen.
Lazzaro MD, Thomson WW. 1992. Endocytosis of lanthanum
nitrate in the organic acid-secreting trichomes of chickpea
(Cicer arietinum L.). American Journal of Botany 79 : 1113–1118.

346 RESEARCH P. A. Vesk et al.
Lewis BD, Spalding EP. 1998. Nonselective block by La$+ of
Arabidopsis ion channels involved in signal transduction. The
Journal of Membrane Biology 162 : 81–90.
Lugones LG, Wo$ sten HAB, Birkenkamp KU, Sjollema KA,
Zagers J, Wessels JGH. 1999. Hydrophobins line air channels
in fruiting bodies of Schizophyllum commune and Agaricus
bisporus. Mycological Research 103 : 635–640.
Madara JL. 1988. Tight junction dynamics: is paracellular
transport regulated? Cell 53 : 497–498.
Martin F, Tagu D. 1995. Ectomycorrhiza development: a
molecular perspective. In: Varma A, Hoch B, eds. Mycorrhiza
structure, function, molecular biology and biotechnology. Berlin,
Germany: Springer Verlag, 29–58.
Marx DH. 1969. The influence of ectotrophic mycorrhizal fungi
on the resistance of pine roots to pathogenic infections. I.
Antagonism of mycorrhizal fungi to root pathogenic fungi and
soil bacteria. Phytopathology 59 : 153–163.
Moon GJ, Clough BF, Peterson CA, Allaway WG. 1986.
Apoplastic and symplastic pathways in Avicennia marina
(Forsk.) Vierh. roots revealed by fluorescent tracer dyes.
Australian Journal of Plant Physiology 13 : 637–648.
Orlovich DA, Ashford AE. 1993. Polyphosphate granules are an
artefact of specimen preparation in the ectomycorrhizal fungus
Pisolithus tinctorius. Protoplasma 173 : 91–102.
Orlovich DA, Ashford AE. 1995. X-ray microanalysis of ion
distribution in frozen salt}dextran droplets after freeze-sub-
stitution and embedding in anhydrous conditions. Journal of
Microscopy 180 : 117–126.
Pa/ lsga/ rd E, Lindh U, Roomans GM. 1994. Comparative study
of freeze-substitution techniques for X-ray microanalysis of
biological tissue. Microscopy Research and Technique 28 :
254–258.
Peterson CA. 1987. The exodermal Casparian band of onion
roots blocks the apoplasmic movement of sulphate ions. Journal
of Experimental Botany 38 : 2068–2081.
Peterson CA, Cholewa E. 1998. Structural modifications of the
apoplast and their potential impact on ion uptake. Zeitschrift fuX rPflanzenernaX hrung und Bodenkunde 161 : 521–531.
Peterson CA, Murrmann M, Steudle E. 1993. Location of the
major barriers to water and ion movement in young roots of Zea
mays L. Planta 190 : 127–136.
Peterson RL, Chakravarty P. 1991. Techniques in synthesizing
ectomycorrhiza. Methods in Microbiology 23 : 75–106.
Piche! Y, Fortin JA. 1982. Development of mycorrhizae, extra-
matrical mycelium and sclerotia on Pinus strobus seedlings. New
Phytologist 91 : 211–220.
Read DJ. 1984. The structure and function of the vegetative
mycelium of mycorrhizal roots. In: Jennings DH, Rayner
ADM, eds. The ecology and physiology of the fungal mycelium.
Cambridge, UK: Cambridge University Press, 215–240.
Read DJ. 1991. Mycorrhizas in ecosystems. Experientia 47 :
376–391.
Read DJ. 1993. Mycorrhiza in plant communities. Advances in
Plant Pathology 9 : 1–31.
Robards AW, Robb ME. 1974. The entry of ions and molecules
into roots : an investigation using electron-opaque tracers.
Planta 120 : 1–12.
Romeo T. 1996. A simple cold-block procedure for the SEM.
Australian EM Newsletter 52 : 16–18.
Smith SE, Read DJ. 1997. Mycorrhizal symbiosis, 2nd edn. San
Diego, CA, USA: Academic Press.
Spurr AR. 1969. A low-viscosity epoxy resin embedding medium
for electron microscopy. Journal of Ultrastructural Research 26 :
31–43.
Steudle E, Peterson CA. 1998. How does water get through
roots? Journal of Experimental Botany 49 : 775–788.
Tagu D, Kottke I, Martin F. 1998. Hydrophobins in ecto-
mycorrhizal symbiosis : hypothesis. Symbiosis 25 : 5–18.
Tagu D, Martin F. 1995. Expressed sequence tags of randomly
selected cDNA clones from Eucalyptus globulus–Pisolithus
tinctorius ectomycorrhiza. Molecular Plant Microbe Interactions
8 : 781–783.
Tagu D, Martin F. 1996. Molecular analysis of cell wall proteins
expressed during the early steps of ectomycorrhiza devel-
opment. New Phytologist 133 : 73–85.
Tagu D, Nasse B, Martin F. 1996. Cloning and characterization
of hydrophobins-encoding cDNAs from the ectomycorrhizal
Basidiomycete Pisolithus tinctorius. Gene 168 : 93–97.
Unestam T. 1991. Water repellency, mat formation, and leaf-
stimulated growth of some ectomycorrhizal fungi. Mycorrhiza
1 : 13–20.
Unestam T, Sun Y-P. 1995. Extramatrical structures of hydro-
phobic and hydrophilic ectomycorrhizal fungi. Mycorrhiza 5 :
301–311.
Varney GT, Canny MJ. 1993. Rates of water uptake into the
mature root system of maize plants. New Phytologist 123 :
775–786.
Varney GT, McCully ME, Canny MJ. 1993. Sites of entry of
water into the symplast of maize roots. New Phytologist 125 :
733–741.
Wessels JGH. 1992. Gene expression during fruiting in Schizo-
phyllum commune. Mycological Research 96 : 609–620.
Wessels JGH. 1993. Wall growth, protein excretion and mor-
phogenesis in fungi. New Phytologist 123 : 397–413.
Wessels JGH. 1994. Developmental regulation of fungal cell wall
formation. Annual Review of Phytopathology 32 : 413–437.
Wessels JGH. 1996. Fungal hydrophobins: proteins that function
at an interface. Trends in Plant Science 1 : 9–15.
Wessels JGH. 1999. Fungi in their own right. Fungal Genetics
and Biology 27 : 134–145.
Wessels JGH, A! sgiersdo! ttir SA, Birkenkamp KU, de VriesOMH, Lugones LG, Scheer JMJ, Schuren FHJ, SchuursTA, van Wetter MA, Wo$ sten HAB. 1995. Genetic regulation
of emergent growth in Schizophyllum commune. Canadian
Journal of Botany 73 : S273–S281.
Wong KKY, Fortin JA. 1989. A Petri dish technique for the
aseptic synthesis of ectomycorrhizae. Canadian Journal of
Botany 67 : 1713–1716.
Wo$ sten HAB, Richter M, Willey JM. 1999. Structural
problems involved in emergence of microbial aerial hyphae.
Fungal Genetics and Biology 27 : 153–160.
Young N, Ashford AE. 1992. Changes during development in
the permeability of sclerotia of Sclerotinia minor to an apoplastic
tracer. Protoplasma 167 : 205–214.
Young N, Ashford AE. 1996. The effects of rind damage and
regeneration on permeability of the apoplast in sclerotia of
Sclerotinia minor. New Phytologist 134 : 13–24.
Zimmermann HM, Steudle E. 1998. Apoplastic transport
across young maize roots : effect of the exodermis. Planta 206 :
7–19.
Copyright © 2022 FDOKUMEN




















