
Impact of a eucalyptus (Eucalyptus globulus Labill.) plantation on the nutrient content and dynamics...
23
Impact of a eucalyptus (Eucalyptus globulus Labill.) plantation on the nutrient content and dynamics of coarse particulate organic matter (CPOM) in a small stream Jon Molinero 1,2 & Jesus Pozo 1, 1 Lab de Ecologı´a, Fac de Ciencias, UPV/EHU, Apdo. 644, 48080 Bilbao, Spain 2 US Environmental Protection Agency, ERD/NERL, 960 College Station Road, Athens, GA 30605, USA (*Author for correspondence: E-mail: [email protected]) Received 2 June 2003; in revised form 18 February 2004; accepted 3 March 2004 Key words: organic matter, Eucalyptus globulus, eucalyptus plantations, forest streams, litterfall, Agu¨era watershed Abstract Litterfall inputs, benthic storage and the transport of coarse particulate organic matter (CPOM) were studied in two headwater streams, one flowing through a mixed deciduous forest and one through a plantation of Eucalyptus globulus. Vertical and lateral traps, transported CPOM and benthic CPOM were sampled monthly to biweekly and sorted into four categories: leaves, twigs and bark, fruits and flowers and debris. The litterfall inputs were about 20% lower at the eucalyptus site but this reduction was unevenly distributed among the litter categories. The reduction of the nitrogen and phosphorus inputs was larger (50%) than that of CPOM because of the low nutrient concentration of the CPOM at the eucalyptus site. Transported CPOM was also lower at the eucalyptus site. Although total CPOM inputs to the stream were reduced in the eucalyptus plantation, benthic storage of CPOM was 50% higher due to (1) high inputs of CPOM and low discharge during summer, (2) more twig and bark inputs, (3) eucalyptus leaves being retained more efficiently in the stream than deciduous leaves (4) a lower discharge, which may in part be attributable to eucalyptus-induced changes in the hydrological cycle. Increased retention balanced lower nitrogen and phosphorus content of CPOM, so benthic storage of nitrogen and phosphorus was similar at both sites. This work demonstrates that the timing, quality and quantity of inputs and benthic storage of CPOM in streams changes substantially because of the substitution of natural deciduous forest with eucalyptus plantation. Maintenance of buffer strips of natural vegetation may be the best way to protect ecological functioning of small, forested streams. Introduction Coarse particulate organic matter (CPOM) is the main energetic resource for aquatic communities in small streams flowing through forested areas (Fisher & Likens, 1973; Cummins et al., 1983; Webster et al., 1995). Its source is the vegetation located on the stream banks (Weigelhofer & Wa- ringer, 1994; Abelho & Grac¸ a, 1998; Larned, 2000) and it is retained by obstacles such as rocks and woody debris within the channel (Jones & Smock, 1991; Webster et al., 1994). As an ener- getic resource, CPOM supports a wide variety of stream organisms as it decomposes (Petersen et al., 1989; Grac¸a, 2001) and is also a source of dissolved organic matter (Ittekkot & Laane, 1991; Mulholland, 1997a). As a structural element, CPOM contributes to channel heteroge- neity and its aggregated distribution, either in riffles or pools, provides suitable habitat for Hydrobiologia 528: 143–165, 2004. Ó 2004 Kluwer Academic Publishers. Printed in the Netherlands. 143
Transcript of Impact of a eucalyptus (Eucalyptus globulus Labill.) plantation on the nutrient content and dynamics...

Impact of a eucalyptus (Eucalyptus globulus Labill.) plantation on the nutrient
content and dynamics of coarse particulate organic matter (CPOM) in a small
stream
Jon Molinero1,2 & Jesus Pozo1,�1Lab de Ecologıa, Fac de Ciencias, UPV/EHU, Apdo. 644, 48080 Bilbao, Spain2US Environmental Protection Agency, ERD/NERL, 960 College Station Road, Athens, GA 30605, USA(*Author for correspondence: E-mail: [email protected])
Received 2 June 2003; in revised form 18 February 2004; accepted 3 March 2004
Key words: organic matter, Eucalyptus globulus, eucalyptus plantations, forest streams, litterfall, Aguera watershed
Abstract
Litterfall inputs, benthic storage and the transport of coarse particulate organic matter (CPOM) werestudied in two headwater streams, one flowing through a mixed deciduous forest and one through aplantation of Eucalyptus globulus. Vertical and lateral traps, transported CPOM and benthic CPOM weresampled monthly to biweekly and sorted into four categories: leaves, twigs and bark, fruits and flowers anddebris. The litterfall inputs were about 20% lower at the eucalyptus site but this reduction was unevenlydistributed among the litter categories. The reduction of the nitrogen and phosphorus inputs was larger(50%) than that of CPOM because of the low nutrient concentration of the CPOM at the eucalyptus site.Transported CPOM was also lower at the eucalyptus site. Although total CPOM inputs to the stream werereduced in the eucalyptus plantation, benthic storage of CPOM was 50% higher due to (1) high inputs ofCPOM and low discharge during summer, (2) more twig and bark inputs, (3) eucalyptus leaves beingretained more efficiently in the stream than deciduous leaves (4) a lower discharge, which may in part beattributable to eucalyptus-induced changes in the hydrological cycle. Increased retention balanced lowernitrogen and phosphorus content of CPOM, so benthic storage of nitrogen and phosphorus was similar atboth sites. This work demonstrates that the timing, quality and quantity of inputs and benthic storage ofCPOM in streams changes substantially because of the substitution of natural deciduous forest witheucalyptus plantation. Maintenance of buffer strips of natural vegetation may be the best way to protectecological functioning of small, forested streams.
Introduction
Coarse particulate organic matter (CPOM) is themain energetic resource for aquatic communitiesin small streams flowing through forested areas(Fisher & Likens, 1973; Cummins et al., 1983;Webster et al., 1995). Its source is the vegetationlocated on the stream banks (Weigelhofer & Wa-ringer, 1994; Abelho & Graca, 1998; Larned,2000) and it is retained by obstacles such as rocksand woody debris within the channel (Jones &
Smock, 1991; Webster et al., 1994). As an ener-getic resource, CPOM supports a wide varietyof stream organisms as it decomposes (Petersenet al., 1989; Graca, 2001) and is also a sourceof dissolved organic matter (Ittekkot & Laane,1991; Mulholland, 1997a). As a structuralelement, CPOM contributes to channel heteroge-neity and its aggregated distribution, either inriffles or pools, provides suitable habitat for
Hydrobiologia 528: 143–165, 2004.� 2004 Kluwer Academic Publishers. Printed in the Netherlands. 143

macroinvertebrates (Reice, 1980; Dobson & Hil-drew, 1992).
Forestry plantations are best viewed as crops,generally mono-specific, managed to increase bio-mass production, as defined by Nambiar (1984).Forestry plantations usually involve the introduc-tion of tree species in areas out of their naturalrange. In the Iberian Peninsula, up to 33 exoticspecies have been recorded in afforestation plots,including 19 gymnosperms and 10 eucalyptusspecies (Ruiz de la Torre, 1990). The introductionof these species may have a large impact onstreams as multiple linkages exist among lotic andterrestrial ecosystems.
Eucalyptus is well accepted as a productivespecies by wood producers due to its very high rateof growth. For example, the area covered byeucalyptus plantations has tripled over the lastdecade in the province of Biscay, Spain (Ascasibar,1997). Its adaptability has favored the introduc-tion of eucalyptus to other Spanish regions, mainlyin the Northern and Southern Atlantic coast, butalso to Portugal where it is widespread (Graca,1993). Eucalyptus is used as productive species inmore than 80 countries, mostly in temperate andtropical areas (Bara et al., 1990). Secondly, there isalso concern about the use of this species in for-estry plantations, as eucalyptus plantations areknown to modify water and nutrient budget ofsoils, increase soil erosion and support a less di-verse community of plants and animals than nat-ural forests (Poore & Fries, 1986). Some studieshave also shown the potential of eucalyptus plan-tations to modify timing, quantity and quality oflitterfall (Abelho & Graca, 1996; Pozo et al.,1997b), benthic storage (Molinero & Pozo, 2002)and leaf litter processing (Canhoto & Graca, 1996;Pozo et al., 1998) in detritus based streams (for areview see Graca et al., 2002).
Nutrient input and availability for stream eco-systems are determined by watershed characteris-tics (e.g., geology, land uses, vegetation) and by awide variety of in-stream processes (Meyer et al.,1988). Although allochthonous organic matter, inthe form of plant litter, is the main energy sourcefor stream communities, few previous studies havequantified the amount of nitrogen and phosphorusassociated with organic matter inputs that entersthe stream (e.g., Benson & Pearson, 1993; Bret-schko & Moser, 1993; Pozo et al., 1997a). Nutrient
content of leaf litter is also important because itinfluences in-stream decomposition (Gessner &Chauvet, 1994), thus it controls the rate at whichleaf litter becomes an available energy source.Eucalyptus leaves have lower nitrogen and phos-phorus content than deciduous species (Pozo,1993), so afforestation with eucalyptus has thepotential to modify nutrient inputs associated withlitterfall inputs to streams.
In this paper, we compare the temporal dyna-mics of CPOM inputs, transport and benthicstorage in a stream that flows through naturaldeciduous forest to one that is surrounded by aneucalyptus plantation. Inputs of nitrogen andphosphorus associated with the CPOM and thestorage of these elements in the benthos are alsoquantified at both sites.
Study area
The Aguera watershed is located in the westernpart of Biscay, northern Spain, and covers about144 km2 (Fig. 1). The climate is temperate oceanicwith annual mean rainfall of 1414mm (n ¼ 10)
Figure 1. Map of the study area. Gauging station is located in
Guriezo (D – deciduous site; E – eucalyptus site).
144
and monthly mean temperature ranges from 9.4 �Cin January to 20.7 �C in August. Lithology is sili-ceous, except for a broad calcareous belt in themiddle of the watershed. This catchment has a lowpopulation density and is mainly occupied byagricultural land in the flat bottom of the valley,by forestry plantations in the high-sloped areas,and heath-lands and natural deciduous forestspredominate in the headwaters. Further detailsabout the Aguera watershed have been publishedelsewhere (Elosegui & Pozo, 1994).
Material and methods
Sampling sites
Two first order streams were selected (Fig. 1,Table 1): Salderrey stream, which drains a 184 ha
watershed and flows through natural deciduousforest and Jergueron stream, which drains an 83 hawatershed and flows through eucalyptus planta-tion. Both streams have narrow high-slopedchannels with steep banks.
Site D (deciduous site in Salderrey stream), anenclosed plot of 1742m2 that includes a 71mstream reach, has a tree density of0.103 individualsm)2. Though low in density, oak(Quercus robur L.) is the dominant species in thecanopy. Alder (Alnus glutinosa (L.) Gaetner) andchestnut (Castanea sativa Miller) also are presentin the forest canopy. Hazel (Corylus avellana L.),hawthorn (Crataegus monogyna Jacq.) and wych-elm (Ulmus glabra Hudson) are the most commonshrub species. Mean channel width along the studyreach is 3.5m and streambed substrate is domi-nated by bedrock with sparse patches of cobbles
Table 1. Description of the study sites (D – deciduous site; E – eucalyptus site; DBH – diameter at breast height)
Site D Site E
Stream order 1 1
Mean discharge (l s)1) 39.8 10.0
Drainage area (ha) 184 83
Channel length (m) 71 32
Channel width (m) 3.58 2.22
Channel slope (%) 14 18
Bed substrate (%)
Bedrock 30 0
Rocks (>100 cm) 7 27
Boulders (25.6–100 cm) 11 43
Cobbles (6.4–25.6 cm) 27 16
Gravels (0.2–6.4 cm) 22 12
Sand, clay (<0.2 cm) 2 3
Channel shadowing (%) 90 66
Vegetation Density (no./m2) Mean DBH (cm) Density (no./m2) Mean DBH (cm)
Alnus glutinosa 0.029 17 – –
Corylus avellana 0.046 6 – –
Castanea sativa 0.018 21 – –
Crataegus monogyna 0.004 7 – –
Eucalyptus globulus – – 0.147 13
Ulmus glabra 0.002 7 – –
Quercus robur 0.004 61 – –
Total 0.103 14 0.147 13
Data are presented only for trees over 2m height.
145

and gravels. The riparian canopy extends well overthe stream channel which is fully shaded.
Site E (eucalyptus site in Jergueron stream), anenclosed plot of 393m2 that includes a 32m streamreach, is covered with eucalyptus (Eucalyptusglobulus Labill.), up to a density of 0.147 indi-vidualsm)2. Deciduous species grow very sparselyunder the eucalyptus canopy as small trees, usuallyless than 3m in height and 2 cm DBH. There isalso a dense growth of brambles (Rubus sp.) onboth stream banks, which represents the mostabundant species in the plantation understory.Mean channel width along the study reach is 2.2mand rocks and large boulders dominate thestreambed substrate. The riparian canopy in theeucalyptus plantation is more open than in thedeciduous forest.
Field and lab methods
Vertical inputs, litter falling directly into thestream, were measured with 0.21m2 litter traps (15at site D and 10 at site E) randomly placed on thebanks. Traps consisted of a wood frame support-ing a plastic bag (1-mm-mesh size) with theopening oriented upboard. Litterfall collected inthese traps was considered to be representative ofthe vertical inputs to the stream as the tree canopyover the channel and on the banks was similar.
Lateral inputs were measured with 50-cm-length litter traps (10 at site D and 6 at site E). Eachlateral trap consisted of a wood frame supporting aplastic bag (1mm-mesh size) with the openingfacing away from the channel, so they collected thelitter moving down slope towards the stream. In-stream transport was measured with drift nets (1-mm-mesh size) maintained at the inlet of the studyreaches for 4 h. Water passing through the drift netand stream discharge was measured after place-ment and before collection with a General Oceaniccurrent-meter. A gauging station, located nearGuriezo at the outlet of the Aguera basin, provideddaily discharge measurements (Fig. 1). At the studyreaches, five replicate benthic organic matter sam-ple were randomly collected with a Surber sampler(30 · 30 cm, 1-mm-mesh size) to determine benthicstorage of CPOM.
Litter traps and benthic CPOM were sampledmonthly to biweekly from October 1993 to March1996. Transported CPOM was sampled from
October 1993 to December 1994 with a similartime schedule.
In the lab, CPOM was air dried and sorted intofour categories: leaves, twigs and bark (<1cmdiameter), flower and fruits and debris (small frag-ments). Each category was oven dried at 70 �C for72h, weighed to the nearest 0.01 g and ground witha Culatti mill to pass a 1-mm-mesh size grid. Or-ganic matter content was determined by combustionin a furnace at 500 �C for 12h. All the transportedCPOM and litter trap samples were sorted intocategories. The composition of the benthic CPOMwas determined four times during 1995.
Analysis of nutrient content
All the transport and benthic CPOM samples wereanalyzed for nutrient content. Also, five verticaland five lateral traps were randomly selected fromeach site for chemical analysis. Up to 2 g of or-ganic matter from each category were frozen foranalysis of nitrogen and phosphorus content.Samples were defrosten and dried overnight at70 �C prior to analysis. Nitrogen content wasmeasured as total Kjeldahl nitrogen with a KjeltecAuto 1030 analyzer, after wet digestion of 100mgof sample with 5 ml of sulphuric acid (98%) andmercuric oxide as a catalyst in a block digester at420 �C for 40min. Phosphorus content was mea-sured on 200mg of sample, by mixed acid diges-tion (5ml of nitric acid, 60%+1ml of perchloricacid, 30%+0.5ml of sulphuric acid, 98%) at350 �C for 20min and followed by a colorimetricanalysis (molybdenum blue method) on the digest,following Allen et al. (1974).
We assumed that most of the nitrogen and phos-phorus contained in the CPOM was organic-boundand in analogy with the concept of CPOM, we havedesignated these fractions of nitrogen and phospho-rus as coarse particulate organic nitrogen (CPON)and coarse particulate organic phosphorus (CPOP).
Statistical analyses
Litterfall was compared by two-way ANOVA (si-te · time). Vertical traps (gm)2 day)1) and lateraltraps (gm)1 day)1) were tested independently fordifferences between the deciduous and eucalyptussites. These data were log-transformed previous tothe statistical analysis. Total litterfall was
146

estimated as the sum of the vertical and lateral in-puts. The standard error of the total litterfall wasestimated as the weighted mean standard error ofvertical and lateral inputs. Annual litterfall(gm)2 year)1) was calculated for two consecutiveyears starting in 22 September 1993. TransportCPOM concentrations (gm)3) were compared withthe Student’s t test. Benthic CPOM (gm)2) wascompared by two-way ANOVA (site · time), afterdata were log-transformed. The quantity of leaves,twigs and bark, flowers and fruits and debris in thebenthos was estimated by applying the percentageof each category obtained during 1995 to the meanstock of benthic CPOM. Turnover time (d) of thetotal CPOM and the four categories was estimatedas the ratio of benthic CPOM to litterfall. Nitrogenand phosphorus content (%) in each CPOM cate-gory were compared by two-way ANOVA (site
· category). Nitrogen and phosphorus content inthe total CPOM was compared by three-wayANOVA (site · time · input path) for the litterfall,by Student’s t test for the transported CPOM, andby two-way ANOVA (site · time) for the benthicCPOM. CPON and CPOP data were analyzed inthe same way as the CPOM data. Multiple com-parisons after ANOVAwere done with the Tukey’stest. All statistical analyses followed Zar (1984).
Results
Litter inputs
Litter inputs to the deciduous forest stream, or siteD, were higher than at the eucalyptus plantationsite, or site E, (Fig. 2a and b). Moreover, signifi-cant differences between the two streams were
Nov Feb May Aug Nov Feb May Aug Nov Feb
g A
FD
Mm
-2
0
200
400
600
g A
FD
M m
-3
0.00
0.05
0.10
0.15
g A
FD
M m
-2 d
-1
0
2
4
6
8
10
m3 s-1
0
30
60
90
120
Benthic CPOM
CPOM inputs
CPOM transport
1994 1995 19961993 1994 1995 19961993
CPOM inputs
CPOM transport
Benthic CPOM
(a)
(c)
(e) (f)
Site D Site E
(b)
(d)
Nov Feb May Aug Nov Feb May Aug Nov Feb
Figure 2. Input, transport and benthic storage of CPOM (mean±1 standard error). Discharge (m3 s)1) at the gauging station of
Guriezo is showed in the top of the graph (D – deciduous site on the left; E – eucalyptus site on the right).
147

found when lateral and vertical inputs were testedindependently (Table 2). Litter inputs varied sea-sonally at site D where the highest CPOM inputs,between 2 and 8 gm)2 day)1, were observed fromOctober to February and the lowest,<2 gm)2 day)1, from March to June (Fig. 2a). Atsite E, litter inputs were less than 4 gm)2 day)1 anddid not show any seasonality (Fig. 2b).
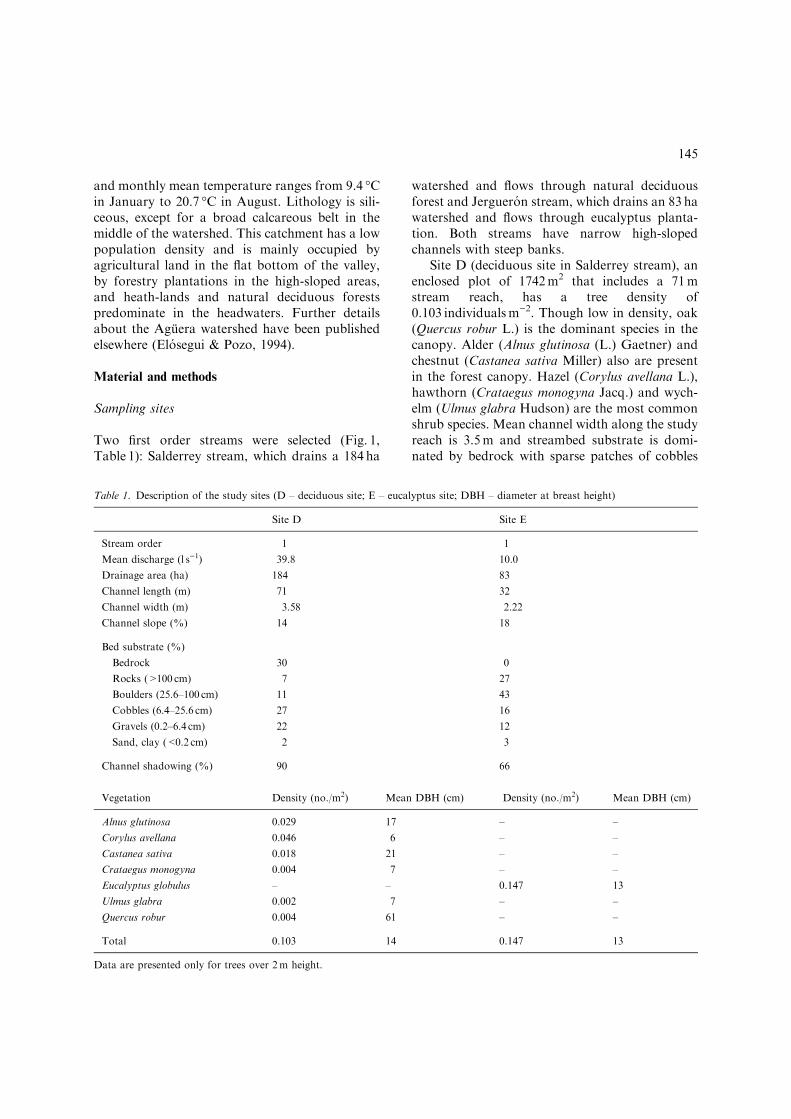
The composition of the litterfall also variedseasonally at site D (Fig. 3a). At this site, thepercentage of leaves in the CPOM inputs was highduring autumn and decreased substantially duringspring and summer while the percentage of debrisfollowed the inverse pattern. The composition ofthe litterfall showed a less marked seasonality atsite E (Fig. 3b). At this site, the percentage ofleaves was higher during the summer, but leaf in-puts entered the stream throughout the whole yearand rarely fell below 50% of the total CPOM in-puts. The percentage of twigs and bark alsoshowed a seasonal pattern at site E and its abun-dance was higher during autumn and winter. Fi-nally, the flowers and fruits category usually madea small contribution to CPOM inputs at both siteswith no defined temporal trend.
Annual CPOM inputs to the stream were lowerat the eucalyptus plantation with little interannualvariations when the two consecutive annual bud-gets were compared (Table 3). Reduction of totalCPOM inputs at site E averaged 20% in the2 years, but this reduction was unevenly distrib-uted among the litter categories. Actually, higherannual inputs of twigs and bark were observed inthe stream draining the eucalyptus plantation, sothe relative contribution of this category to thetotal CPOM doubled at this site.
The mechanism for CPOM input was alsomodified under the eucalyptus plantation. Verticalinputs of CPOM at site E averaged507.8 gm)2 year)1, only 10% less than the verticalinputs at site D (568.8 gm)2 year)1). However, thelateral inputs at site E averaged 53.8 g m)2 year)1,about 70% less than the lateral inputs at site D(159.8 gm)2 year)1). So, reduction of lateral in-puts at the eucalyptus site was responsible forabout 70% of the reduction of total inputs ofCPOM. Reduction of lateral inputs was due topresence of a dense shrub understory that trappedleaves and other materials before they reached thestream and because eucalyptus leaves were longer
and heavier than the leaves of most deciduousspecies, which further reduced their movement onthe banks.
Transported CPOM
CPOM transport at site D was higher than at site E(Student’s t, p<0.05, Table 4). When the littercategories were compared, there were no significantdifferences (Student’s t, p>0.05) between the twosites in the transport of flowers and fruits and twigsand bark. However, the leaves and the debris cate-gories showed higher values in the transport at siteD (Student’s t, p<0.05). At both sites, CPOMtransport showed no seasonal trend (Fig. 2c and d)or correlation with stream discharge (p>0.05). Atthe deciduous site, an intense rain during theDecember 1994 sampling led to transported CPOMover 2 gm)3. This datewas not included either in thegraph or in the statistical analysis as we consideredthat this single event was not enough to representthe effects of high flows on the transported CPOM.No transport occurred between July and Septemberat site E as the stream was almost dry.
The composition of the transported CPOM(Fig. 3c) varied seasonally at site D following thesame pattern that was observed in the CPOM in-puts. In contrast, there were no seasonal changesin the composition of the transported CPOM atsite E (Fig. 3d), which was probably reflecting theless marked seasonal changes of the CPOM cate-gories in the litterfall at this site.
Benthic standing stocks of CPOM
Mean benthic standing stock of CPOM was140.3 gm)2 at site E and 90.9 gm)2 at site D andthere were significant differences between the twosites (two-way ANOVA, p<0.001, Table 5).Temporal dynamics of CPOM standing stocksdiffered between the two streams (Fig. 2e and f). Atsite D, CPOM standing stocks peaked in autumnor winter, above 250 gm)2 but were about 25 gm)2
for most of the year (Fig. 2e). At site E, CPOMstanding stocks increased during the summer, upto 200 gm)2 but usually ranged from 10 to100 gm)2 (Fig. 2f).
Discharge largely influenced benthic CPOMstorage at both sites (Fig. 2e and f). Low discharges
148

Table 2. Two-way ANOVA (site· time) for lateral and vertical inputs of CPOM, CPON and CPOP to the streams
CPOM Lateral traps (gm)1 day)1) Vertical traps (gm)2 day)1)
Total
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time p<0.001 p<0.001
Leaves
Site p<0.001 E<D p<0.01 E<D
Time p<0.001 p<0.001
Site·Time p<0.001 p<0.001
Twigs and bark
Site n.s. n.s.
Time p<0.001 p<0.001
Site·Time n.s. p<0.001
Flowers and fruits
Site n.s. p<0.001 E<D
Time p<0.001 p<0.001
Site·Time n.s. p<0.001
Debris
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time p<0.05 p<0.001
CPON Lateral traps (gNm)1 day)1) Vertical traps (mgNm)2 day)1)
Total
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time p<0.001 p<0.001
Leaves
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time p<0.001 p<0.001
Twigs and bark
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time n. s. p<0.01
Flowers and fruits
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time p<0.001 p<0.001
Debris
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time p<0.001 p<0.001
Continued on p. 150
149

resulted in CPOM accumulation in the streamwhereas high flows resulted in CPOM export. Anegative correlation was observed between CPOMstanding stocks and monthly mean discharge atboth sites, but this relationship was stronger at theeucalyptus site (r2 ¼ 0.58, p<0.001) compared tothe deciduous site (r2 ¼ 0.12, p<0.05). At site E,CPOM standing stocks depended largely on dis-charge, as litterfall was almost constant through theyear. At the deciduous site, discharge and benthicCPOM correlation was weak and no accumulationof CPOM was observed during the summer eventhough discharge was low because low litterfall in-put prevented its accumulation. Large interannualdifferences were observed in the benthic CPOMstanding stocks. At the deciduous site, mean ben-thic CPOM standing stock during, 1994 was27.6 gm)2, but it increased to 146.1 gm)2 during1995. A similar trend was observed at the eucalyp-tus site. In this stream, mean benthic CPOMstanding stock was 85.6 gm)2 and increased to
230.1 gm)2 during 1995. Differences in the annualstorage of CPOM were related to the irregularprecipitation patterns during 1995; which led to lowflows and high CPOM retention in both streams(Fig. 2e and f).
In both sites, the composition of the benthicCPOM (Fig. 3e and f) followed the temporaltrends observed in the litterfall and in the trans-port. However, retention differed between the twosites and also across litter categories. Despite ofsimilar contribution of leaves to litterfall at bothsites (Table 3), the percentage of leaves in thebenthic CPOM at site E was higher than at site D(Table 5), so leaves were retained more efficientlyin the eucalyptus site than in the deciduous site.Also, the percentage of twigs and bark observed inthe benthic CPOM (Table 5) was higher than in theCPOM inputs (Table 3) and the transportedCPOM (Table 4), so this category was retainedmore efficiently in the benthos than the others atboth sites.
Table 2. (Continued)
CPOP Lateral traps (mg P m)1 d)1) Vertical traps (mg P m)2 d)1)
Total
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time p<0.001 p<0.001
Leaves
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.01
Site·Time p<0.001 p<0.001
Twigs and bark
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time n. s. p<0.01
Flowers and fruits
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time p<0.001 p<0.001
Debris
Site p<0.001 E<D p<0.001 E<D
Time p<0.001 p<0.001
Site·Time p<0.001 p<0.001
Data have been log-transformed (D-deciduous site; E-eucalyptus site; n.s.–not significant).
150
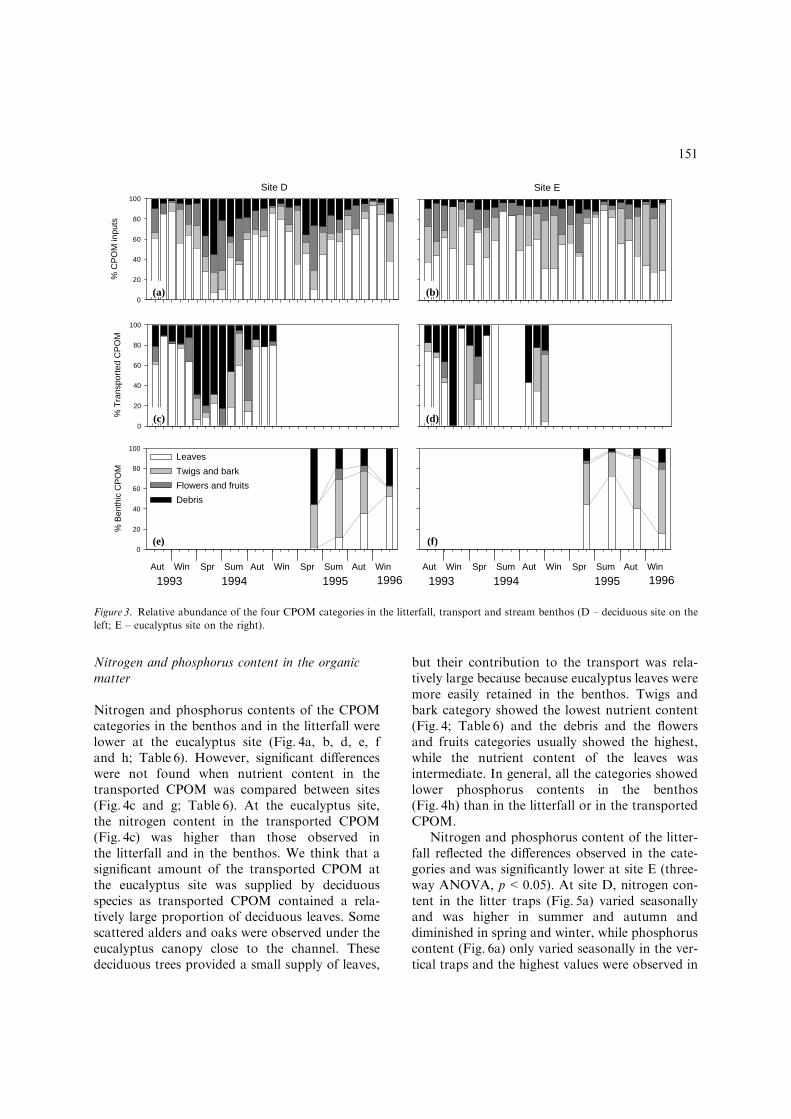
Nitrogen and phosphorus content in the organicmatter
Nitrogen and phosphorus contents of the CPOMcategories in the benthos and in the litterfall werelower at the eucalyptus site (Fig. 4a, b, d, e, fand h; Table 6). However, significant differenceswere not found when nutrient content in thetransported CPOM was compared between sites(Fig. 4c and g; Table 6). At the eucalyptus site,the nitrogen content in the transported CPOM(Fig. 4c) was higher than those observed inthe litterfall and in the benthos. We think that asignificant amount of the transported CPOM atthe eucalyptus site was supplied by deciduousspecies as transported CPOM contained a rela-tively large proportion of deciduous leaves. Somescattered alders and oaks were observed under theeucalyptus canopy close to the channel. Thesedeciduous trees provided a small supply of leaves,
but their contribution to the transport was rela-tively large because because eucalyptus leaves weremore easily retained in the benthos. Twigs andbark category showed the lowest nutrient content(Fig. 4; Table 6) and the debris and the flowersand fruits categories usually showed the highest,while the nutrient content of the leaves wasintermediate. In general, all the categories showedlower phosphorus contents in the benthos(Fig. 4h) than in the litterfall or in the transportedCPOM.
Nitrogen and phosphorus content of the litter-fall reflected the differences observed in the cate-gories and was significantly lower at site E (three-way ANOVA, p<0.05). At site D, nitrogen con-tent in the litter traps (Fig. 5a) varied seasonallyand was higher in summer and autumn anddiminished in spring and winter, while phosphoruscontent (Fig. 6a) only varied seasonally in the ver-tical traps and the highest values were observed in
Site D
% C
PO
M in
puts
0
20
40
60
80
100
Site E
% T
rans
port
ed C
PO
M
0
20
40
60
80
100
% B
enth
ic C
PO
M
0
20
40
60
80
100
Win Spr SumAut Win Spr SumAut WinAut
Leaves
Twigs and bark
Flowers and fruits
Debris
1993 1994 1995 1996
(a) (b)
(c) (d)
(e) (f)
Win Spr SumAut Win Spr SumAut WinAut
1993 1994 1995 1996
Figure 3. Relative abundance of the four CPOM categories in the litterfall, transport and stream benthos (D – deciduous site on the
left; E – eucalyptus site on the right).
151

Table 3. Annual inputs of CPOM, CPON and CPOP to the study streams (D – deciduous site; E – eucalyptus site)
Site D Site E
CPOM
(gm)2 year)1)
% of total
CPOM
Lateral
input as%
CPOM
(gm)2 y)1)
% of total
CPOM
Lateral
input as%
93–94
Total 759.3 19.5 517.4 7.7
Leaves 498.7 65.7 22.8 312.6 60.4 7.8
Twigs and bark 85.1 11.2 18.0 108.3 20.9 8.8
Flowers and fruits 108.9 14.3 6.0 63.7 12.3 4.4
Debris 66.7 8.8 18.2 32.9 6.4 10.1
94–95
Total 697.9 24.6 605.9 11.2
Leaves 476.1 68.2 30.3 359.4 59.3 10.1
Twigs and bark 67.4 9.7 12.2 146.9 24.3 10.1
Flowers and fruits 82.7 11.8 7.9 57.6 9.5 18.2
Debris 71.7 10.3 17.5 42.0 6.9 15.1
CPON
(gNm)2 y)1)
% of total
CPON
Lateral
input as%
CPON
(gNm)2 y)1)
% of total
CPON
Lateral
input as%
93–94
Total 11.99 24.4 4.02 11.2
Leaves 8.06 67.2 27.6 2.63 65.5 11.6
Twigs and bark 0.75 6.2 22.7 0.48 11.8 9.4
Flowers and fruits 1.75 14.6 8.8 0.47 11.8 7.2
Debris 1.43 12.0 26.8 0.44 10.9 15.4
94–95
Total 10.71 20.8 4.87 10.3
Leaves 7.36 68.7 24.8 3.14 64.6 8.5
Twigs and bark 0.64 6.0 12.9 0.65 13.4 7.0
Flowers and fruits 1.14 10.6 2.5 0.41 8.4 18.0
Debris 1.58 14.7 18.2 0.66 13.6 17.3
CPOP
(mgPm)2 year)1)
% of total
CPOP
Lateral
input as%
CPOP
(mgPm)2 year)1)
% of total
CPOP
Lateral
input as%
93–94
Total 402.4 22.1 141.1 11.6
Leaves 228.8 56.9 27.1 87.5 62.0 11.0
Twigs and bark 24.0 6.0 18.4 9.3 6.6 13.3
Flowers and fruits 91.5 22.8 9.9 27.8 19.7 8.6
Debris 57.9 14.4 23.0 16.6 11.8 18.7
94–95
Total 334.4 18.5 181.7 10.6
Leaves 187.0 55.9 25.3 104.9 57.7 7.7
Twigs and bark 24.6 7.3 11.0 19.9 11.0 8.6
Flowers and fruits 62.8 18.8 2.8 26.5 14.6 16.9
Debris 59.9 17.9 16.7 30.4 16.7 16.2
152

winter and spring. At site E, nutrient content wassimilar in the vertical and lateral traps and did notshow a defined temporal trend (Figs 5b and 6b).Nitrogen and phosphorus content in the trans-ported CPOMwas similar at both sites (Student’s t,p>0.05), with mean values about 1.7 and 0.05%(Figs 5c, d and 6c, d). Temporal variability in thenutrient content of the CPOM was related tochanges in the relative importance of the categories.Nutrient content was more variable at site D, wherethe composition of the litterfall varied seasonally,than at site E, where the abundance of the catego-ries showed a less marked seasonality. The increaseof nitrogen content in the litterfall at site D wasrelated to the presence of alder leaves in the traps.At both sites, high phosphorus content in coincidedwith the high percentages of debris in the litterfalland in the transported CPOM.
Nutrient content in the benthic CPOM at siteD was significantly higher than at site E (Figs. 5e, fand 6e, f, two-way ANOVA, p<0.05). Temporalvariability in the nutrient content was usuallysmall and significant differences (two-way ANO-VA, p<0.05) due to temporal variation only ap-
peared in the phosphorus content. A decrease inthe phosphorus content of the benthic CPOMalong the study period was observed at both sites.Nitrogen content at the eucalyptus site also de-creased slightly throughout the study period.
CPON and CPOP input, transport and benthicstorage
Nitrogen and phosphorus inputs to the streamswith the litterfall were lower at the eucalyptus site(Table 2). At site E, annual input of CPON wasabout 60% lower than at site D. CPOP annualinput was also reduced about 55% at site E com-pared to site D (Table 3). The large reduction inthe annual input of CPON and CPOP reflectedboth lower litterfall and lower nutrient contents inthe organic matter at the eucalyptus site and thedifferences between the two consecutive years ofthis study were small. About 55–65% of thenutrient inputs were associated with leaves(Table 3). Due to their high phosphorus content,the flower and fruits and the debris categoriestogether represented between 30 and 35% of the
Table 4. Discharge weighted mean concentrations of transported CPOM, CPON and CPOP in the study reaches (D – deciduous site; E
– eucalyptus site)
Site D Site E
CPOM (mgm)3) % of total CPOM CPOM (mgm)3) % of total CPOM
Total 55.2 22.7
Leaves 33.7 61.2 9.9 43.8
Twigs and bark 2.2 4.0 3.1 13.5
Flowers and fruits 3.4 6.2 2.7 11.9
Debris 15.8 28.7 7.0 30.7
CPON (mgNm)3) % of total CPON CPON (mgNm)3) % of total CPON
Total 1.08 0.35
Leaves 0.58 53.3 0.14 40.4
Twigs and bark 0.03 2.9 0.02 6.7
Flowers and fruits 0.10 9.1 0.02 5.4
Debris 0.38 34.7 0.17 47.4
CPOP (lgPm)3) % of total CPOP CPOP (lgPm)3) % of total CPOP
Total 36.00 10.69
Leaves 15.08 41.9 4.34 40.6
Twigs and bark 0.91 2.5 0.96 9.0
Flowers and fruits 5.33 14.8 1.02 9.6
Debris 14.67 40.8 4.36 40.8
153
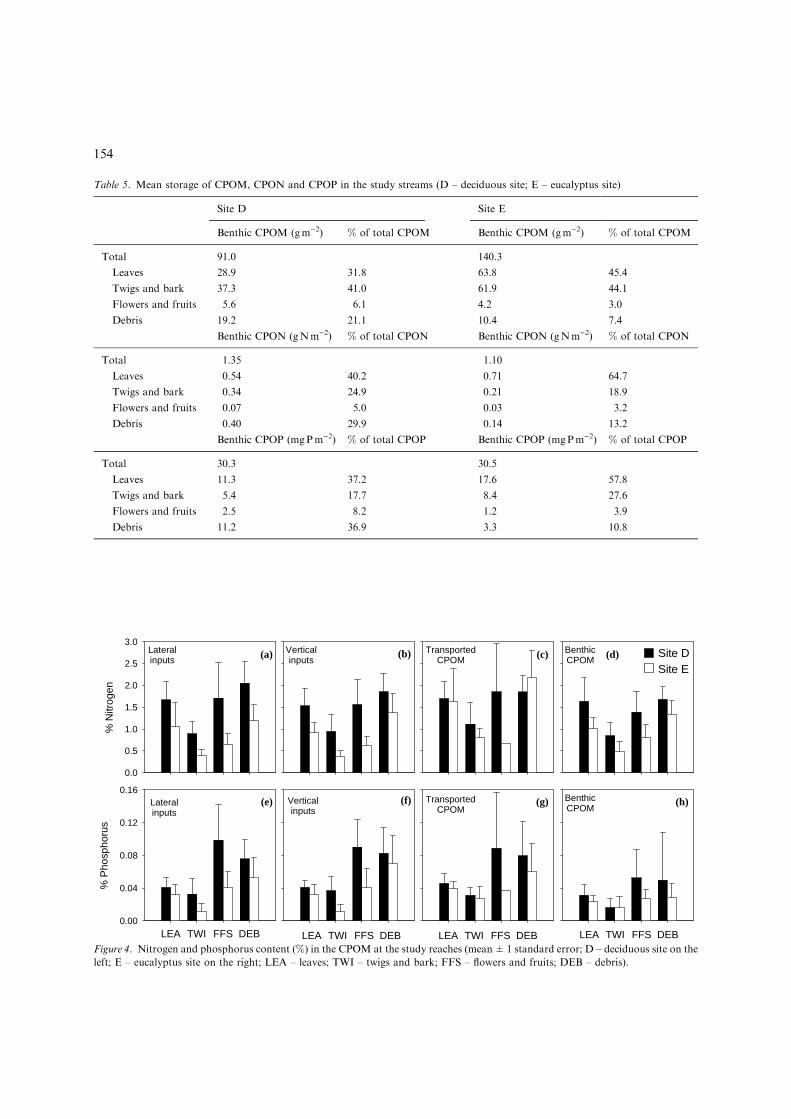
Table 5. Mean storage of CPOM, CPON and CPOP in the study streams (D – deciduous site; E – eucalyptus site)
Site D Site E
Benthic CPOM (gm)2) % of total CPOM Benthic CPOM (gm)2) % of total CPOM
Total 91.0 140.3
Leaves 28.9 31.8 63.8 45.4
Twigs and bark 37.3 41.0 61.9 44.1
Flowers and fruits 5.6 6.1 4.2 3.0
Debris 19.2 21.1 10.4 7.4
Benthic CPON (gNm)2) % of total CPON Benthic CPON (gNm)2) % of total CPON
Total 1.35 1.10
Leaves 0.54 40.2 0.71 64.7
Twigs and bark 0.34 24.9 0.21 18.9
Flowers and fruits 0.07 5.0 0.03 3.2
Debris 0.40 29.9 0.14 13.2
Benthic CPOP (mgPm)2) % of total CPOP Benthic CPOP (mgPm)2) % of total CPOP
Total 30.3 30.5
Leaves 11.3 37.2 17.6 57.8
Twigs and bark 5.4 17.7 8.4 27.6
Flowers and fruits 2.5 8.2 1.2 3.9
Debris 11.2 36.9 3.3 10.8
BenthicCPOM
Site DSite E
Lateralinputs
% N
itrog
en
0.0
0.5
1.0
1.5
2.0
2.5
3.0Verticalinputs
TransportedCPOM
Lateralinputs
LEA TWI FFS DEB
% P
hosp
horu
s
0.00
0.04
0.08
0.12
0.16Verticalinputs
TransportedCPOM
BenthicCPOM
(a) (c) (d)
(e) (g) (h)
(b)
(f)
LEA TWI FFS DEB LEA TWI FFS DEB LEA TWI FFS DEBFigure 4. Nitrogen and phosphorus content (%) in the CPOM at the study reaches (mean±1 standard error; D – deciduous site on the
left; E – eucalyptus site on the right; LEA – leaves; TWI – twigs and bark; FFS – flowers and fruits; DEB – debris).
154

inputs of CPOP to these streams, though they wereonly a minor portion of the CPOM inputs.
Transported CPON and CPOP were about30% lower at the eucalyptus site (Table 4). Whenthe categories were tested individually, most ofthem showed significantly lower values in thetransport at the eucalyptus site (Student’s t,p<0.05). However, there were no significant dif-ferences between the two sites (Student’s t,p>0.05) in the amount of transported CPON andCPOP associated with the twigs and bark cate-gory. At both sites, a high proportion of thetransported CPON and CPOP (43–54%) wasassociated with the flowers and fruits and thedebris categories.
Despite a significant reduction in nutrient in-puts with litterfall, mean annual storage of benthicCPON and CPOP did not differ significantly be-tween the two sites (Table 5, two-way ANOVAp>0.05). The reduction of the nitrogen andphosphorus content of organic matter at theeucalyptus site was balanced with higher benthicstorage of CPOM, so no differences between thetwo sites were found in the benthic storage of
CPON and CPOP. The importance of the littercategories in the benthos varied as a function ofthe riparian vegetation (Table 5) and more CPONand CPOP was associated with leaves and less withdebris at the eucalyptus site. Also, the twigs andbark category, which represented close to 50% ofCPOM standing stocks in the streams, accountedfor a much lower percentage (18–25%) of thebenthic CPON and CPOP.
Turnover time
Turnover or residence time was calculated as theratio of CPOM inputs to benthic CPOM in days.Turnover time of total CPOM was about twice ashigh at the eucalyptus site compared to thedeciduous site (Table 7). This difference was due tothe higher residence time of the leaves at site E, asthe other CPOM categories showed similar turn-over time at both sites. Twigs and bark showed thelongest turnover time at both sites, while theflowers and fruits category showed the shortest.
Turnover of total CPON was similar to that oftotal CPOM at both sites. However, the turnover
Table 6. Two-way ANOVA (site· category) for the nitrogen and phosphorus concentration in the litterfall, transported CPOM and
benthic CPOM (D – deciduous site; E – eucalyptus site; Lea – leaves; Twi – twigs and bark; FFS – flowers and fruits; Deb – Debris; n.s.
– not significant)
Nitrogen (%) Phosphorus (%)
Litterfall, vertical traps
Site p<0.001 E<D p<0.001 E<D
Category p<0.001 Twi<FFSa Leaa<Deb p<0.001 Twi<Lea<FFS<Deb
Site·Category p<0.001 p<0.001
Litterfall, lateral traps
Site p<0.001 E<D p<0.001 E<D
Category p<0.001 Twi<FFSa Leaa<Deb p<0.001 Twi<Lea<Deb<FFS
Site·Category p<0.001 p<0.001
Transported CPOM*
Site n.s. n. s.
Category p<0.001 Twi<Leaa Deba p<0.001 Twia Leaa<Deb
Site·Category n.s. n.s.
Benthic CPOM
Site p<0.001 E<D p<0.05 E<D
Category p<0.001 Twi<FFSa Leaa,b Debb p<0.01 Twia Leaa,b Debb FFSb
Site·Category n. s. n. s.
There were no significant differences between categories with the same superscript.
*FFS not included because of lack of replicates.
155

time of the total CPOP was about one third shorterthan that of the total CPOM. Turnover of totalCPOP was shorter than that of CPOM and CPONbecause of the high percentage of phosphorus inthe litterfall that was associated with the flower andfruits category. Also, a smaller percentage ofphosphorus was associated with the twigs and barkcategory, which showed long turnover times. Theslight reduction in the phosphorus content of thematerials stored in the benthos when compared tothe litterfall also contributed to the overall shortturnover time of this element.
Discussion
Afforestation with eucalyptus and CPOM dynamic
Our data show that the loss of natural riparianvegetation and afforestation with eucalyptus
largely modifies CPOM dynamics in streams.Afforestation with eucalyptus modifies litterfallquantity and timing to streams. Streams drainingforested areas in temperate zones receive largeCPOM inputs from the terrestrial environmentduring late autumn or early winter (Chauvet &Jean-Louis, 1988; Abelho & Graca, 1998). How-ever, litter input to streams draining eucalyptusplantations shows little seasonality with only amajor peak during the summer (Abelho & Graca,1996). Also, the overall quantity of CPOMreaching the stream is reduced and an increase inthe proportion of twigs and bark entering thestream has been described (Pozo et al., 1997b).This alteration of the natural litterfall input regimealso modifies transport and storage of CPOMwithin streams.
In forested ecosystems, litterfall is the mainpath of nutrient return to the soil from the treecanopy (Cole & Rapp, 1981). Although, the
Nov Feb May Aug Nov Feb May Aug Nov Feb
% N
itrog
en
0
1
2
3
4
% N
itrog
en
0
1
2
3
4
% N
itrog
en
0
1
2
3
4
Lateral inputsVertical inputs
Benthic CPOM
CPOM inputs
CPOM transport
1994 1995 19961993
CPOM transport
Benthic CPOM
CPOM inputs(b)
Site D Site E
Nov Feb May Aug Nov Feb May Aug Nov Feb
1994 1995 19961993
(a)
(c)
(e) (f)
(d)
Figure 5. Nitrogen content (%) in the CPOM at the study reaches (mean±1 standard error; D – deciduous site on the left; E –
eucalyptus site on the right).
156

energetic role of leaflitter in streams is well rec-ognized, few works have evaluated the input ofnitrogen and phosphorus or other elements asso-ciated with these terrestrial CPOM inputs tostreams. Afforestation with E. globulus has also thepotential to modify nitrogen and phosphorus in-puts associated with litterfall to small streams(Table 8). Nutrient inputs with litterfall observedin some deciduous forests dominated by Quercusspecies ranges from 2.9 to 8.8 and from 161 to880mgPm)2 year)1. The high nitrogen inputs atsite D, about 2.2 gNm)2 year)1 more than thefollowing entry in table, is probably reflecting thepresence of alder, a nitrogen fixing species.Nitrogen inputs in natural eucalyptus forest – be-tween 0.5 and 7.0 gNm)2 year)1 — seem to beclose to the values found in deciduous forests, butphosphorus inputs, between 36 and237mgPm)2 year)1, are lower. Nutrient inputs ineucalyptus plantations show high variability —1.5–7.6 gNm)2 year)1and 82–170mgPm)2 year)1,
but nutrient inputs at site E and in otherE. globulus plantations are among the lowestreported values.
Lateral inputs – CPOM that enters the streamfrom the banks – must be estimated as it canrepresent as much as 70% of the total CPOM input(Cushing, 1988). At the eucalyptus site, a denseunderstory of shrubs develops as the clear canopyallows more light to on the forest floor. Also,eucalyptus leaves are heavier than deciduousleaves and their mobility on the stream bank is low(Pozo et al., 1994). Both differences in the struc-ture of vegetation cover and leaf characteristics area result of the eucalyptus plantation and contrib-ute to reduce the lateral inputs of CPOM.
Our data also show a large reduction in trans-ported CPOM in the stream flowing through theeucalyptus plantation. This reduction is explainedby the lower inputs and in-stream higher retentionof CPOM at site E. Nutrient content in trans-ported CPOM is similar at both sites, despite the
Nov Feb May Aug Nov Feb May Aug Nov Feb
% P
hosp
horu
s
0.00
0.04
0.08
0.12
0.16
% P
hosp
horu
s
0.00
0.04
0.08
0.12
0.16
% P
hosp
horu
s
0.00
0.04
0.08
0.12
0.16Lateral inputsVetical inputs
Benthic CPOM
CPOM inputs
CPOM transport
1994 1995 19961993
CPOM inputs
CPOM transport
Benthic CPOM
Site D Site E
Nov Feb May Aug Nov Feb May Aug Nov Feb
1994 1995 19961993
(a) (b)
(d)(c)
(e) (f)
Figure 6. Phosphorus content (%) in the CPOM at the study reaches (mean±1 standard error; D – deciduous site on the left; E –
eucalyptus site on the right).
157

large differences that are observed in the nutrientcontent of the litterfall. Although eucalyptusplantations are harvested every 13–15 years andthe regeneration of the natural riparian vegetationis limited, these small trees provide a surplus ofdeciduous leaves to the stream and can reduce theimpact of the eucalyptus plantation on the stream(Molinero & Pozo, 2002).
The dynamics of benthic storage of CPOMreflect the changes in timing and quality of litter-fall. Streams draining eucalyptus plantations showhigher CPOM storage than streams drainingdeciduous forests (Abelho & Graca, 1996; Molin-ero & Pozo, 2002). At the eucalyptus site, thetiming of litterfall leads to CPOM accumulation insummer and the larger percentage of twigs andbark in the litterfall can also increase CPOMretention as these woody materials are easily re-tained within the channel (Webster et al., 1994,1999) and act as subsequent retention devices forCPOM (Bilby & Likens, 1980; Diez et al., 2000).Otherwise, materials stored in streams that draineucalyptus plantations have lower nitrogen andphosphorus content than benthic CPOM instreams draining deciduous forest, but the benthic
storage of CPON and CPOP is similar in bothsystems because of the increase in the benthicstorage of CPOM in the streams impacted byeucalyptus (Molinero & Pozo, 2002).
The physical characteristics of the streams canalso explain the differences in CPOM storage andtransport. The smaller channel and the lower dis-charge at site E could lead to higher storage ofbenthic CPOM and lower CPOM transport andsince retention is a function of stream width anddischarge (Speaker et al., 1984). Also, 30% of thechannel in site D is exposed bedrock with lowretention potential. Therefore, it is difficult todistinguish if the higher storage and lower trans-port of CPOM at the eucalyptus site is a result ofthe differences in the characteristics of the streamsor the riparian vegetation. However, CPOMturnover time was affected by the presence of theeucalyptus plantation.
Turnover time of total CPOM was higher at theeucalyptus site, but among categories of CPOM,only the leaves differed between sites. If the phys-ical characteristics of the streams played animportant role in determining the differences in theCPOM retention, then an increase of the turnovertime of all the categories would be expected at theeucalyptus site. This observation supports thatdifferences in the turnover time of the leaves aredue to the timing of the inputs. Eucalyptus leavesenter the stream mainly during the summer (Pozoet al., 1997b), when they are retained due to lowflows. At the deciduous site, leaves enter thestream in late autumn when they are easily washeddownstream, as storms and high flows are morelikely to occur during this season. Over a relativelynarrow range of width-specific discharge values(ratio of discharge to mean stream width), CPOMretention is highly variable across CPOM types(Brookshire & Dwire, 2003). So, differences inphysical properties of leaves (eucalyptus leaves areheavier than deciduous leaves and have a longshape) can also influence turnover time of benthicCPOM at the eucalyptus site. Previous studieshave found differences in the breakdown rates ofeucalyptus and alder leaves (Pozo, 1993). How-ever, breakdown rates of eucalyptus are similar tothose of deciduous species such as oak and chest-nut (Molinero et al., 1996), which represent about80% of the leaf litter in site D (Pozo et al., 1997b).So, differences in leaf litter breakdown rates can-
Table 7. Turnover time (days) for the CPOM in the study
streams (D – deciduous site; E – eucalyptus site)
Site D Site E
CPOM
Total 46 91
Leaves 22 69
Twigs and bark 179 177
Flowers and fruits 21 25
Debris 101 102
CPON
Total 43 90
Leaves 26 90
Twigs and bark 178 136
Flowers and fruits 17 29
Debris 98 96
CPOP
Total 30 69
Leaves 20 67
Twigs and bark 81 210
Flowers and fruits 12 16
Debris 69 51
158

not explain the longer turnover time of the euca-lyptus leaves.
Differences in the turnover time of benthicCPOM observed in several forested streams ran-ged from 25 d to 512 days (Table 9). Turnover timetends to decrease as the size of the stream increasesand is likely a function of decreased retention inlarger streams. Turnover time observed in ourstreams was much shorter than in undisturbed first
order streams. Other authors have also found highturnover rates of CPOM in streams surrounded bydisturbed riparian vegetation (Larned, 2000) orwith variable discharge (Richardson, 1992). At ourstudy sites, the high-sloped channel and the flashyhidrology favor the export of CPOM. Since debrisdams play a major role in the retention of organicand inorganic materials (Smock et al., 1989), thelow quantity of coarse woody debris in both
Table 8. Annual inputs of nitrogen and phosphorus in several forest stands (Aus. – Australia; Bel. – Belgium; Spa. – Spain; USA –
United States of America; Eth. – Ethiopia; UK – United Kingdom; Ind. – India; Por. – Portugal)
Forest type gNm)2 year)1 mg P m)2 year)1 Source
Eucalyptus forests
Eucalyptus diversicolor, <20 years Australia Occ. (Aus.) 2.5 63 O’Connell & Menage (1982)
Eucalyptus diversicolor, >20 years Australia Occ. (Aus.) 5.6 113 O’Connell & Menage (1982)
Eucalyptus regnans, <20 years Victoria (Aus.) 5.3 210 Polglase & Attiwill (1992)
Eucalyptus regnans, >20 years Victoria (Aus.) 6.4 237 Polglase & Attiwill (1992)
Eucalyptus rossi, 100 years New Wales (Aus.) 2.9 140 Crockford & Richardson
(1998)
Eucalyptus clamaldulensis New Wales (Aus.) 1.1 67 Briggs & Maher (1983)
Eucalyptus gummifera New Wales (Aus.) 4.1 108 Lamb (1985)
Eucalyptus botryoides New Wales (Aus.) 7.0 223 Lamb (1985)
Eucalyptus cebra Queensland (Aus.) 0.8 122 McIvor (2001)
Eucalyptus drepanophylla Queensland (Aus.) 0.5 36 McIvor (2001)
Eucalyptus plantations
Eucalyptus tereticornis, 8 years Central Himalaya (Ind.) 7.6 170 Bargali & Singh (1991)
Eucalyptus globulus, 2–6 years Menagesha (Eth.) 2.1 120 Lisanework & Michelsen
(1994)
Eucalyptus globulus, 1–5 years Furadouro (Por.) 1.5 82 Madeira et al. (1995)
Eucalyptus globulus, 12 years Basque Country (Spa.) 4.0 144 This work
Deciduous forests
Quercus prinus Tenesse (USA) 3.4 240 Johnson & Henderson
(1989)
Quercus petraea Lunerburger Heide (Ger.) 4.4 384 Rode (1995)
Quercus robur Virelles (Bel.) 5.9 340 Duvigneaud & Denaeyer-
De Smet (1970)
Quercus robur Virelles (Bel.) 5.0 240 Duvigneaud & Denaeyer-
De Smet (1970)
Quercus robur Meathop (UK) 6.3 260 Data of Satchell (Cole &
Rapp, 1981)
Quercus petraea Salamanca (Spa.) 4.1 275 Gallardo et al. (1998)
Quercus macrocarpa Kansas (USA) 6.6 880 Killingbeck (1986)
Quercus robur Basque Country (Spa.) 8.8 293 This work
Some data represent mean values of different years or locations.
159

streams (Elosegi et al., 1999) promoted transportover retention. However, the effect of the coarsewoody debris is not clear in our sites as the euca-lyptus site retains more benthic CPOM even-though the amount of coarse woody debris is twothirds less than at the deciduous site (Elosegi et al.,1999). Thus, if the amount of woody debris in thestreams is low, the timing of the litterfall and thehydrological regime become more important incontrolling the CPOM dynamics.
The turnover time of the benthic CPON wassimilar to that of the CPOM, but the turnover timeof the CPOP was shorter than that of the CPOM.Much of the CPOP input at both sites was in theform of flowers and fruits, which showed the shortestturnover rate. Differences in the patterns of nitrogenand phosphorus content during the breakdownprocess have been also described. During thebreakdown of the leaf litter in streams, nitrogencontent remains constant or increases slightly butphosphorus is usually removed quickly from thedecaying leaves if dissolved nutrient levels are low, sothe N:P ratio of the leaves increases (Pozo, 1993;
Molinero et al., 1996). Rapid removal of phosphorusfrom the CPOM when it enters the stream can ex-plain the lower phosphorus contents of the materialsstored in the benthos and also contribute to shorterturnover time of benthic CPOP.
Impact of eucalyptus afforestation on the streamecosystem
Landscape level disturbances such as logging orvegetation changes result in changes in aquaticecosystems (Tuchman & King, 1993; Delong &Brusven, 1994; Whiles & Wallace, 1997; Benfield etal., 2001). Our data support that afforestation witheucalyptus leads to important changes in thedynamics of stream CPOM and changes both inthe amount and quality of CPOM can affectstream biota (Wallace et al., 1999; Graca, 2001).
Afforestation with eucalyptus reduces thenutrient content of the CPOM in the stream be-cause there is a shift towards materials with lownitrogen and phosphorus content, but also becausethere is a reduction of the inputs of nutrient rich
Table 9. Residence time (d) of CPOM in different forested streams
Stream
order
Mean
discharge (l s)1)
Benthic
CPOM (gm)2)
CPOM
Inputa (gm)2 y)1)
Turnover
time (d)
Source
Breitembach (Ger.) 1 26 621b 700 323 Marxen et al. (1997)
Buzzards Branch (USA) 1 – 739 527 512 Smock (1990)
Walker Branch (USA) 1 12 175 565 113 Mulholland (1997b)
Satellite Branch (USA) 1 2 526 629 305 Wallace et al. (1997)
Colliers Creek (USA) 1 – 983 961 373 Smock (1990)
Margaraca forest (Por.) 1 23 157 715 80 Abelho & Graca (1998)
Salderrey stream (Spa.) 1 40 91 729 46 This work
Jergueron stream (Spa.) 1 10 140 562 91 This work
Mayfly Creek (Can.) 2 – 32 255 46 Richardson (1992)
Spring Creek (Can.) 2 – 32 465 25 Richardson (1992)
Hugh White Creek (USA) 2 19 213 577 135 Webster et al. (1997)
Bear Brook (USA) 2 4 610 594 375 Findlay et al. (1997)
Kamiki stream (USA) 2 103 60 851 26 Larned (2000)
White Clay Creek (USA) 3 115 118 313 138 Newbold et al. (1997)
Keppel Creek (Aus.) 4 171 105 744 52 Treadwell et al. (1997)
Kings Creek (USA) 5 59 238 726 120 Gray (1997)
Ogeechee river (USA) 6 67 869 4363 73 Meyer et al. (1997)
Sites D (Salderrey) and E (Jergueron) are also included (Aus. – Australia; Ger. – Germany; USA – United States of America; Can. –
Canada; SA – South Africa; Por. – Portugal; Spa. – Spain).aOnly litterfall inputs. Transport was not included.bFine particulate organic matter was included.
160

materials such leaves and fruits and an increase ininputs of twigs and bark (Fig. 7). This change inthe nutrient content of CPOM entering the streamis important because organisms tend to assimilatethese elements at certain ratios (Frost et al., 2002).In streams, microorganisms that colonize organicsubstrates can use two sources of nutrients, thedecomposing material and the water column(Gessner & Chauvet, 1994). Thus, because thenutrient content in CPOM is lower in eucalyptusstreams, we hypothesize that decomposition instreams impacted by eucalyptus plantations ismore dependent on the availability of dissolvednutrients in the water column.
Previous work has proposed that eucalyptusplantations have a direct effect on the stream foodwebs because eucalyptus leaves have low nutrientcontent (Pozo, 1993). Also, Canhoto & Graca(1995) found that Tipula larvae could not surviveon a diet of eucalyptus leaves. However, it remainsunclear whether eucalyptus leaves were a poorfood source because of their low nutrient contentor for other reasons (e.g., presence of toxic com-pounds, physical characteristics of the leaves).The analysis of the impact of eucalyptus leaveson macroinvertebrate shredders is complexbecause their sensivity to changes in the foodsource may differ among species and because ourunderstanding of the nutrient requirements of thebenthic organisms is poor (Frost et al., 2002).
The breakdown rate of eucalyptus leaves islower than that of alder leaves (Pozo, 1993), be-cause eucalyptus leaves have a thick cuticle andorganic compounds that slow down the initialcolonization of the leaves by fungi (B€arlocher etal., 1995). However, the effect of slower decom-position on stream ecosystems is unclear becauseour data suggest that the leaves are retained for alonger time in streams draining eucalyptus plan-tations (Fig. 7) and because breakdown rates ofeucalyptus leaves are similar to those of otherdeciduous species (Molinero et al., 1996; Barlocher& Graca, 2002). Also, because of the multiplelinkages that exist between a stream and its wa-tershed, afforestation with eucalyptus can impactstreams in multiple ways (e.g., suppression ofcoarse woody debris inputs, leaching of toxicsubstances, changes in hydrology) and changes inthe leaf litter breakdown rates in streams drainingeucalyptus plantations may be not a result of the
suitability of eucalyptus leaves as a food source forshredders but because changes in the hydrological,chemical or physical characteristics of the streamlimit the populations of shredders.
Despite of the differences in the size of thewatershed, our data also suggest that less rainfall isavailable as stream flow at the eucalyptus site. Arough water budget was constructed for the studysites with the discharge measurements collected forthe transported CPOM calculations. BetweenOctober 1993 and October 1994, rainfall wasabout 1697mm in the Aguera watershed.
Figure 7. Impact of the eucalyptus plantation as a function of
CPOM inputs (gm)2 year)1), nutrient concentrations (%) in the
CPOM and residence time (d) of the CPOM in the stream (D –
deciduous site; E – eucalyptus site; LEA – leaves; TWI – twigs
and bark; FFS – flowers and fruits; DEB – debris).
161

However, discharge was only 34% (584mm) of therainfall input at the eucalyptus site, meanwhile 60%(1006) of the rainfall contributed to stream flow atthe deciduous site. Modification of the naturalvegetation leads to changes in the hydrologic budgetof small watersheds (Waring & Sohleninger, 1984;Garman & Moring, 1991; Heede, 1991; Smith,1992). Moreover, eucalyptus is thought to modifythe hydrological budget in soils due to a hightranspiration rates (Poore & Fries, 1986). Both asmaller watershed and loss of soil water due to hightranspiration help to explain the lower dischargeand CPOM transport observed in the eucalyptusplantation. We identify watershed hydrology as akey issue to be addressed in junction with the studyof leaf litter breakdown and CPOM dynamics.
Our data show that afforestation with euca-lyptus has the potential to alter the CPOMdynamics in small streams and that this impact isrelated to differences in the timing and the nutrientcontent of CPOM inputs. However, the lack ofreplication limits the conclusiveness of our study.Eucalyptus plantations can vary in tree density,soil preparation, management of the understoryvegetation and harvest cycle among others char-acteristics. Also, local and regional differences inthe impact of eucalyptus plantation on streams canbe expected due to variation in climate and soiland stream characteristics. Although the impor-tance of these factors in determining the impact ofeucalyptus plantations cannot be inferred fromthis study, we think that we have shown enoughevidence to state that loss of the natural riparianvegetation due to afforestation with eucalyptuslargely impacts CPOM dynamics in streams. Thus,the maintenance of buffer strips of natural vege-tation may be the best way to protect these smallstreams from the changes due to land use modifi-cation in the watershed. These buffer strips canprovide a surplus of deciduous leaves to the streamand reduce the presence of eucalyptus litter withinthe channel (Pozo et al., 1994; Molinero & Pozo,2002). Also, if managed properly, they will becomea source of coarse woody debris allowing furtherrestoration of the stream ecological functioning.
Acknowledgements
This project has been funded by the DGICYT(PB92-0459) and the University of the Basque
Country (UPV 118.310-EA043/93). Jon Molineroreceived a fellowship from the Program forTraining of Research Staff (Department of Edu-cation, Universities and Research, Basque Gov-ernment) and is a NRC research associate at theUS Environmental Protection Agency. Z. Arteaga,A. Elosegi, A. Basaguren, J. Dıez, J. M. Gonzalez,E. Gonzalez, A. Lopez de Alda, A. Lopez deLuzuriaga and P. Riano also collaborated in thefield and laboratory work. Authors want to thankR. Burke, J. Richardson and two anonymousreviewers for improving the manuscript.
References
Abelho, M. & M. A. S. Graca, 1996. Effects of eucalyptus
afforestation on leaf litter dynamics and macroinvertebrate
community structure of streams in Central Portugal.
Hydrobiologia 324: 195–204.
Abelho, M. & M. A. S. Graca, 1998. Litter in a first-order
stream of a temperate deciduous forest (Margaraca Forest,
central Portugal). Hydrobiologia 386: 147–152.
Allen, S. E., H. M. Grimshaw, J. A. Parkinson & C. Quarmby,
1974. Chemical Analysis of Ecological Materials. Blackwell
Scientific Publications, Oxford.
Ascasibar, J., 1997. La situacion forestal de cara al siglo XXI.
Revista Internacional de Estudios Vascos 42: 309–323.
Bara, S., J. L. Montero & A. Rigueiro, 1990. Sobre el eucalipto.
Asociacion para el progreso forestal, Madrid, Spain.
Bargali, S. S. & S. P. Singh, 1991. Aspects of productivity and
nutrient cycling in an 8-year-old Eucalyptus plantation in a
moist plain area adjacent to central Himalaya, India.
Canadian Journal of Forest Research 21: 1362–1365.
Barlocher, F. & M. A. S. Graca, 2002. Exotic riparian vegeta-
tion lowers fungal diversity but not leaf decomposition in
Portuguese sterams. Freshwater Biology 47: 1123–1135.
Barlocher, F., C. Canhoto & M. A. S. Graca, 1995. Fungal
colonization of alder and eucalypt leaves in two streams in
Central Portugal. Archive fur Hydrobiologie 233: 457–470.
Benfield, E. F., J. R. Webster, J. L. Tank & J. J. Hutchens,
2001. Long-term patterns in leaf breakdown in streams in
response to watershed logging. Internationale Revue der
gesamten Hydrobiologie 86: 467–474.
Benson, L. J. & R. G. Pearson, 1993. Litter inputs to a tropical
Australian rainforest stream. Australian Journal of Ecology
18: 377–383.
Bilby, R. E. & G. E. Likens, 1980. Importance of organic debris
dams in the structure and function of stream ecosystems.
Ecology 61: 1107–1113.
Bretschko, G. & H. Moser, 1993. Transport and retention of
matter in riparian ecotones. Hydrobiologia 251: 95–101.
Briggs, S. V. & M. T. Maher, 1983. Litter fall and leaf
decomposition in a river red gum (Eucalyptus calmaldulensis)
swamp. Australian Journal of Botany 31: 307–316.
Brookshire, E. N. J. & K. A. Dwire, 2003. Controls on patterns
of coarse organic particle retention in headwater streams.
162

Journal of the North American Benthological Society 22:
17–34.
Canhoto, C. & M. A. S. Graca, 1995. Food value of introduced
eucalypt leaves for a Mediterranean stream detritivore Tip-
ula lateralis. Freshwater Biology 34: 209–214.
Canhoto, C. & M. A. S. Graca, 1996. Decomposition of
Eucalyptus globulus leaves and three native leaf species (Al-
nus glutinosa, Castanea sativa and Quercus faginea) in a
Portuguese low order stream. Hydrobiologia 333: 79–85.
Chauvet, E. & A. M. Jean-Louis, 1988. Production de litiere de
la rypisylve de la Garonne et apport au fleuve. Acta Oeco-
logica 9: 265–279.
Cole, D. W. & M. Rapp, 1981. Elemental cycling in forest
ecosystems. In Reichle, D. E. (ed.), Dynamic Properties of
Forest Ecosystems. Cambridge University Press, Cambridge:
341–409.
Crockford, R. H. & P. Richardson, 1998. Litterfall, litter and
associated chemistry in a dry sclerophyll eucalypt forest and
a pine plantation in south-eastern Australia: 2. Nutrient
recycling by litter, throughfall and stemflow. Hydrological
Processes 12: 385–400.
Cummins, K. W., J. R. Sedell, F. J. Swanson, G. W. Minshall,
S. G. Fisher, C. E. Cushing, R. C. Petersen & R. L. Vannote,
1983. Organic matter budgets for stream ecosystems: prob-
lems in their evaluation. In Barnes, J. R. & G. W. Minshall
(eds), Stream Ecology: Application and Testing of General
Ecological Theory. Plenum Press, New York: 299–353.
Cushing, C. E., 1988. Allochthonous detritus inputs to a small,
cold desert spring-stream. Verhandlungen der Internationa-
len Vereinigung fur theoretische und angewandte Limnolo-
gie 23: 1107–1113.
Delong, M. D. & M. A. Brusven, 1994. Allochthonous input of
organic matter from different riparian habitats of an agri-
culturally impacted stream. Environmental Management 18:
59–71.
Dıez, J. R., S. Larranaga, A. Elosegi & J. Pozo, 2000. Effect of
removal of wood on streambed stability and retention of
organic matter. Journal of the North American Bentholog-
ical Society 19: 621–632.
Dobson, M. & A. G. Hildrew, 1992. A test of resource limita-
tion among shredding detritivores in low order stream in
southern England. Journal of Animal Ecology 61: 69–77.
Duvigneaud, P. & S. Denaeyer-De Smet, 1970. Biological cy-
cling of minerals in temperate deciduous forests. In Jacobs,
J., O. L. Lange, J. S. Olson & W. Wieser (eds), Ecological
Studies 1: Analysis of Temperate Forest Ecosystems.
Springer-Verlag, Berlin (Germany): 199–225.
Elosegui, A. & J. Pozo, 1994. Spatial versus temporal variability
in the physical and chemical characteristics of the Ag€uera
stream (Northern Spain). Acta Oecologica 15: 543–559.
Elosegi, A., J. R. Dıez & J. Pozo, 1999. Abundance, charac-
teristics, and movement of woody debris in four Basque
streams. Archive fur Hydrobiologie 144: 455–471.
Findlay, S., G. E. Likens, L. Hedin, S. G. Fisher & W. H.
McDowell, 1997. Organic matter dynamics in Bear Brook,
Hubbard Brook experimental forest, New Hampshire, USA.
In Webster, J. R. & J. L. Meyer (eds), Stream Organic
Matter Budgets: 43–46. Journal of the North American
Benthological Society 16: 3–161.
Fisher, S. G. & G. E. Likens, 1973. Energy flow in Bear Brook,
New Hampshire: an integrative approach to stream ecosys-
tem metabolism. Ecological Monographs 43: 421–439.
Frost, P. C., R. S. Stelzer, G. A. Lamberti & J. J. Elser, 2002.
Ecological stoichiometry of trophic interactions in the ben-
thos: understanding the role of C:N:P ratios in lentic and
lotic habitats. Journal of the North American Benthological
Society 21: 525–528.
Gallardo, J. F., A. Martın & I. Santa Regina, 1998. Nutrient
cycling in deciduous forest ecosystems of the Sierra de Gata
mountains: aboveground litter production and potential
nutrient return. Annales des Sciences Forestieres 55: 749–
769.
Garman, G. C. & J. R. Moring, 1991. Initial effects of defor-
estation on physical characteristics of a boreal river. Hyd-
robiologia 209: 29–37.
Gessner, M. O. & E. Chauvet, 1994. Importance of stream
microfungi in controlling breakdown rates of leaf litter.
Ecology 75: 1807–1817.
Graca, M. A. S., 1993. Patterns and processes in detritus-based
streams systems. Limnologica 23: 107–114.
Graca, M. A. S., 2001. The role of invertebrates on leaf litter de
composition in streams – a review. Internationale Revue der
gesamten Hydrobiologie 86: 389–393.
Graca, M.A.S., J. Pozo, C. Canhoto & A. Elosegi, 2002. Effects
of Eucalyptus plantations on detritus, decomposers, and
detritivores in streams. The ScientificWorld Journal 2: 1173–
1185.
Gray, L. J., 1997. Organic matter dynamics in Kings Creek,
Konza Prairie, Kansas, USA. In Webster, J. R. & J. L.
Meyer (eds), Stream Organic Matter Budgets: 50–54. Journal
of the North American Benthological Society 16: 3–161.
Heede, B. H., 1991. Response of a stream in disequlibrium to
timber harvest. Environmental Management 15: 251–255.
Ittekot, V. & R. W. P. M. Laane, 1991. Fate of riverine par-
ticulate matter. In Degens, E. T., S. Kempe & J. E. Richey
(eds), Biogeochemistry of Major World Rivers. John Wiley
and Sons Ltd., New York: 233–243.
Jones, J. B. & L. A. Smock, 1991. Transport and retention of
particulate organic matter in two low-gradient headwater
streams. Journal of the North American Benthological
Society 10: 115–126.
Johnson, D. W. & G. S. Henderson, 1989. Terrestrial nutrient
cycling. In Johnson, D. W. & R. I. Van Hook (eds), Analysis
of Biochemical Cycling Processes in Walker Branch
Watershed. Springer-Verlag, New York: 233–300.
Killingbeck, K. T., 1986. Litterfall dynamics and element use
efficiency in a Kansas gallery forest. American Midland
Naturalist 116: 180–189.
Lamb, R. J., 1985. Litter fall and nutrient turnover in two
eucalypt woodlands. Australian Journal of Botany 33: 1–14.
Larned, S. T., 2000. Dynamic of coarse riparian detritus in a
Hawaiian stream ecosystem: a comparison of drought and
post-drought conditions. Journal of the North American
Benthological Society 19: 215–234.
Lisanework, N. & A. Michelsen, 1994. Litterfall and nutrient
release by decomposition in three plantations compared with
a natural forest in the Ethiopian higland. Forest Ecology and
Management 65: 149–164.
163

Madeira, M., M. C. Araujo & J. S. Pereira, 1995. Effects of
water and nutrient supply on amount and on nutrient con-
centration of litterfall and forest floor litter in Eucalyptus
globulus plantations. Plant and Soil 168–169: 287–295.
Marxen, J., H. Schmidt & D. M. Fiebig, 1997. Organic matter
dynamics in the Breitenbach, Germany. In Webster, J. R. &
J. L. Meyer (eds), Stream Organic Matter Budgets: 28–32.
Journal of the North American Benthological Society 16:
3–161.
McIvor, J. G., 2001. Litterfall from trees in semiarid woodlands
of north-east Queensland. Austral Ecology 26: 150–155.
Meyer, J. L., W. H. McDowell, T. L. Bott, J. W. Elwood, C.
Ishizaki, J. M. Melack, B. L. Peckarsky, B. J. Peterson & P.
A. Rublee, 1988. Elemental dynamics in streams. Journal of
the North American Benthological Society 7: 410–
432.
Meyer, J. L., A. C. Benke, R. T. Edwards & J. B. Wallace,
1997. Organic matter dynamics in the Ogeechee river, a
blackwater river in Georgia, USA. In Webster, J. R. & J.
L. Meyer (eds), Stream Organic Matter Budgets: 82–87.
Journal of the North American Benthological Society 16:
3–161.
Molinero, J. & J. Pozo, 2002. Impact of eucalyptus plantations
on the benthic storage of coarse particulate organic matter,
nitrogen and phosphorus in small streams. Verhandlungen
der Internationalen Vereinigung fur theoretische und ange-
wandte Limnologie 28: 540–544.
Molinero, J., J. Pozo & E. Gonzalez, 1996. Litter breakdown in
streams of the Aguera catchmen: influence of dissolved
nutrients and land use. Freshwater Biology 36: 745–
756.
Mulholland, P. J., 1997a. Dissolved organic matter concentra-
tion and flux in streams. In Webster, J. R. & J. L. Meyer
(eds), Stream Organic Matter Budgets: 131–141. Journal of
the North American Benthological Society 16: 3–161.
Mulholland, P. J., 1997b. Organic matter dynamics in the West
Fork of Walker Branch, Tennesse, USA. In Webster, J. R. &
J. L. Meyer (eds), Stream Organic Matter Budgets: 61–67.
Journal of the North American Benthological Society 16: 3–
161.
Nambiar, E. K. S., 1984. Plantation forests: their scope and a
perspective on plantation nutrition. In Bowen, G. D. & E. K.
S. Nambiar (eds), Nutrition on Plantation Forests. Aca-
demic Press, London: 1–15.
Newbold, J. D., T. L. Bott, L. A. Kaplan, B. W. Sweeney & R.
L. Vannote, 1997. Organic matter dynamics in White Clay
Creek, Pennsilvania, USA. In Webster, J. R. & J. L. Meyer
(eds), Stream Organic Matter Budgets: 46–50. Journal of the
North American Benthological Society 16: 3–161.
O’Connell, A. M. & P. M. A. Menage, 1982. Litter fall and
nutrient cycling in karri (Eucalyptus diversicolor F. Muell.)
forest in relation to stand age. Australian Journal of Ecology
7: 49–62.
Petersen, R. C., K. W. Cummins & G. M. Ward, 1989.
Microbial and animal processing of detritus in a woodland
stream. Ecological Monographs 59: 21–39.
Polglase, P. J. & P. M. Attiwill, 1992. Nitrogen and phosphorus
cycling in relation to stand age of Eucalyptus regnans F.
Muell. Plant and Soil 142: 157–166.
Poore, D. & C. Fries, 1986. Efectos ecologicos de los eucaliptos.
Technical report, FAO.
Pozo, J., 1993. Leaf litter processing of alder and eucalyptus in
the Aguera stream system (North Spain) I. Chemical chan-
ges. Archive fur Hydrobiologie 127: 299–317.
Pozo, J., A. Basaguren & A. Elosegui, 1994. Transported and
benthic coarse particulate organic matter in the Aguera
stream (North Spain). Verhandlungen der Internationalen
Vereinigung fur theoretische und angewandte Limnologie
25: 1723–1726.
Pozo, J., E. Gonzalez, J. Dıez & A. Elosegi, 1997a. Leaf litter
budgets in two contrasting forested streams. Limnetica 13:
77–84.
Pozo, J., E. Gonzalez, J. R. Dıez, J. Molinero & A. Elosegui,
1997b. Inputs of particulate organic matter to streams with
different riparian vegetation. Journal of the North American
Benthological Society 16: 602–611.
Pozo, J., A. Basaguren, A. Elosegui, J. Molinero, E. Fabre & E.
Chauvet, 1998. Afforestation with Eucalyptus globulus and
leaf litter decomposition in streams of northern Spain.
Hydrobiologia 373/374: 101–109.
Reice, S. R., 1980. The role of sustratum in benthic macroin-
vertebrate microdistribution and litter decomposition in a
woodland stream. Ecology 61: 580–590.
Richardson, J. S., 1992. Coarse particulate detritus dynamics in
small, montane streams of southwestern British Columbia.
Canadian Journal of Fisheries and Aquatic Sciences 49: 337–
346.
Rode, M. W., 1995. Aboveground nutrient cycling and forest
development on poor sandy soil. Plant and Soil 168–169:
337–343.
Ruiz de la Torre, J., 1990. Distribucion y caracterısticas de las
masas forestales espanolas. Ecologıa, fuera de serie 1: 11–30.
Smith, C. M., 1992. Riparian afforestation effects on water
yields and water quality in pasture catchments. Journal of
Environmental Quality 21: 237–245.
Smock, L. A., 1990. Spatial and temporal variation in organic
matter storage in low-gradient, headwater streams. Archive
fur Hydrobiologie 118: 169–184.
Smock, L. A., G. M. Metzler & J. E. Gladden, 1989. Role of
debris dams in the structure and functioning of low-gradient
headwater streams. Ecology 70: 764–775.
Speaker, R., K. Moore & S. Gregory, 1984. Analysis of the
process of retention of organic matter in stream ecosystems.
Verhandlungen der Internationalen Vereinigung fur theo-
retische und angewandte Limnologie 22: 1835–1841.
Treadwell, S. A., I. C. Campbell & R. T. Edwards, 1997. Or-
ganic matter dynamics in Keppel Creek, southeastern Aus-
tralia. In Webster, J. R. & J. L. Meyer (eds), Stream Organic
Matter Budgets: 58–61. Journal of the North American
Benthological Society 16: 3–161.
Tuchman, N. C. & R. H. King, 1993. Changes in mechanisms
of summer detritus processing between wooded and agri-
cultural sites in a Michigan headwater stream. Hydrobiolo-
gia 268: 115–127.
Wallace, J. B., T. F. Cuffney, S. L. Eggert & M. R. Whiles,
1997. Stream organic matter inputs, storage and export for
Satellite Branch at Coweeta Hydrologic Laboratory, North
Carolina, USA. In Webster, J. R. & J. L. Meyer (eds),
164

Stream Organic Matter Budgets: 67–74. Journal of the
North American Benthological Society 16: 3–161.
Wallace, J. B., S. L. Eggert, J. L. Meyer & J. R. Webster, 1999.
Effects of resource limitation on a detrital-based ecosystem.
Ecological Monographs 69: 409–442.
Waring, R. H. & W. H. Sohlesinger, 1984. Forest ecosystems:
concepts and management. Academic Press, Orlando, Flor-
ida, USA.
Webster, J. R., A. P. Covich, J. L. Tank & T. V. Crockett, 1994.
Retention of coarse organic particles in streams in the
southern Appalachian mountains. Journal of the North
American Benthological Society 13: 140–150.
Webster, J. R., J. B. Wallace & E. F. Benfield, 1995. Organic
processes in streams of the eastern United States. In Cush-
ing, C. E., K. W. Cummins & G. W. Minshall (eds), River
and Stream Ecosystems. Ecosystems of the World, 22.
Elsevier, Amsterdam: 117–187.
Webster, J. R., J. L. Meyer, J. B. Wallace & E. F. Benfield,
1997. Organic matter dynamics in Hugh White Creek,
Coweeta Hidrologic Laboratory, North Carolina, USA. In
Webster, J. R. & J. L. Meyer (eds), Stream Organic Matter
Budgets: 74–78. Journal of the North American Bentholog-
ical Society 16: 3–161.
Webster, J. R., E. F. Benfield, T. P. Ehrman, M. A. Schaeffer, J.
L. Tank, J. J. Hutchens & D. J. D’Agelo, 1999. What hap-
pens to allochthonous material that falls into streams? A
synthesis of new and published information from Coweeta.
Freshwater Biology 41: 687–705.
Weigelhofer, G., & J. A. Waringer, 1994. Allochtonous input of
coarse particulate organic matter (CPOM) in a first to four
order Australian forest stream. Internationale Revue der
gesamten Hydrobiologie 79: 461–471.
Whiles, M. R. & J. B. Wallace, 1997. Leaf litter decomposition
and macroinvertebrate communities in headwater streams
draining pine and hardwood catchments. Hydrobiologia
353: 107–119.
Zar, J. H., 1984. Biostatistical Analysis. Prentice-Hall, Engle-
wood Cliffs, New Jersey, USA.
165




















