
Electrospun Core-Sheath Bicomponent Nanostructures for Soft Tissue Engineering
Upload
independentCategory
view
2download
0
PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Kandan, A.]On: 17 September 2010Access details: Access Details: [subscription number 927030697]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Archives Of Phytopathology And Plant ProtectionPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713454295
Rapid suppression of defence enzymes and compounds by sheath blight(Rhizoctonia solani) and sheath rot (Sarocladium oryzae) toxins in rice cellsuspension culturesA. Kandana; R. Radja Commareb; R. Nandakumarc; G. Amuthad; A. Vijayasamundeeswarid; A.Ramanathand; T. Raguchanderd; P. Balasubramaniand; R. Samiyappand
a Agricultural Biotechnology Research Center, Academia Sinica, Nankang, Taipei 115, Taiwan, R.O.C. b
Institute of Plant and Microbial Biology, Academia Sinica, Nankang, Taiwan, R.O.C., Taipei 115 c
Department of Plant Pathology and Crop Physiology, Louisiana State University, Louisiana, USA d
Department of Plant Pathology, Centre for Plant Protection Studies, Tamil Nadu AgriculturalUniversity, Coimbatore, India
Online publication date: 16 September 2010
To cite this Article Kandan, A. , Commare, R. Radja , Nandakumar, R. , Amutha, G. , Vijayasamundeeswari, A. ,Ramanathan, A. , Raguchander, T. , Balasubramanian, P. and Samiyappan, R.(2010) 'Rapid suppression of defenceenzymes and compounds by sheath blight (Rhizoctonia solani) and sheath rot (Sarocladium oryzae) toxins in rice cellsuspension cultures', Archives Of Phytopathology And Plant Protection, 43: 14, 1407 — 1422To link to this Article: DOI: 10.1080/03235400802476658URL: http://dx.doi.org/10.1080/03235400802476658
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Rapid suppression of defence enzymes and compounds by sheath blight
(Rhizoctonia solani) and sheath rot (Sarocladium oryzae) toxins in rice cell
suspension cultures
A. Kandana*, R. Radja Commareb, R. Nandakumarc, G. Amuthad,A. Vijayasamundeeswarid, A. Ramanathand, T. Raguchanderd, P. Balasubramaniand andR. Samiyappand
aAgricultural Biotechnology Research Center, Academia Sinica, Nankang, Taipei 115, Taiwan,R.O.C.; bInstitute of Plant and Microbial Biology, Academia Sinica, Nankang, Taipei 115, Taiwan,R.O.C.; cDepartment of Plant Pathology and Crop Physiology, Louisiana State University, Louisiana,USA; dDepartment of Plant Pathology, Centre for Plant Protection Studies, Tamil Nadu AgriculturalUniversity, Coimbatore 641 003, India
(Received 5 September 2008; final version received 8 September 2008)
To understand the suppression mechanisms against disease resistance in rice, we tookadvantage of the fact that suspension cultured cells exhibit many of the defence responsesthat are characteristic of intact tissues. In this study we constitutively measured theRhizoctonia solani and Sarocladium oryzae toxins, induced and suppressed levels ofphenylalanine ammonia lyase, peroxidase, superoxide dismutase, phenols, catalase, b-1,3-glucanase and chitinase in rice suspension cultured cells. The addition of Rhizoctoniasolani and Sarocladium oryzae toxins separately in suspension cultured cells shows thesuppression of defence enzymes and compounds at 24 h and 48 h respectively exceptSOD. The rice cultivar IR50 delays the disease suppression effect when compared to theother cultivars viz., Pusa Basmati and Co 43. The PR proteins (namely b-1,3-glucanaseand chitinase) activities in rice suspension cultured cells were reduced during 48 h and72 h after the addition of Rhizoctonia solani toxin, whereas the activities were suppressedonly after 72 h when inoculated with Sarocladium oryzae toxin. Selective suppression ofthese defence enzymes and compounds by Rhizoctonia solani and Sarocladium oryzaetoxin shows that toxins play a major role during pathogenesis in rice cells.
Keywords: defence enzymes; rice; sheath blight; sheath rot; suspension cultures; toxins
Introduction
Plants have developed an impressive array of defence mechanisms and the genes inducedas part of the general defence response can be divided into several functional groups(Dixon et al. 1994). Generally, in vitro selection for disease resistance and the defencemechanisms of the cell are attractive approaches in host–pathogen systems wherepathotoxins are involved in pathogenesis. The use of suspension cultured cells can beexpected to have some advantage over intact plant tissues in order to study the mode ofaction of toxins because the suspension cultured cells are grown synchronously and can beuniformly treated with toxins. Moreover, the cultured cell systems have been utilised forconfirmation of defence mechanisms and also for selection of toxin tolerant cells
*Corresponding author. Email: [email protected]
Archives of Phytopathology and Plant Protection
Vol. 43, No. 14, 20 September 2010, 1407–1422
ISSN 0323-5408 print/ISSN 1477-2906 online
� 2010 Taylor & Francis
DOI: 10.1080/03235400802476658
http://www.informaworld.com
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

(Kodama et al. 1995). Recognition of pathogens triggers a large range of inducible defenceresponses. The induced mechanisms associated with the HR include strengthening of theplant cell wall by alterations in the synthesis of cell wall structural proteins (Lamb et al.1989). First the phenylpropanoid products play diverse roles in the response of plants todifferent biotic and abiotic stimuli. Second, the synthesis of peroxidases is induced. Thesedefence gene products may also contribute to the formation of a structural barrier topathogen invasion. Third, chitinase and b-1,3-glucanse enzyme synthesis occurs. Theseenzymes may either promote degradation of the cell walls of invading fungi which furtherstimulates the defence responses (Dietrich et al. 1990).
Rice (Oryza sativa L.) is one of the most important cereal crops feeding three-quartersof the world’s population. One of the major limiting factors in rice production is damagethrough diseases caused by fungi, bacteria, viruses and other abiotic stresses. Among thefungal diseases, sheath blight caused by Rhizoctonia solani Kuhn and sheath rot caused bySarocladium oryzae W. Gams and D. Hawksw are the most serious diseases in all rice-growing countries (Sawada 1922; Chen and Chien 1964; Jimenez and Parizos 1975;Shajahan et al. 1977; Roy 1993; Mariappan 1996). Several fungal pathogens are able toproduce phytotoxins (Cutler 1993), varying in molecule size, chemical type and hostspecificity, but all modify the physiology of host cells. Suspension culture should alsoprove valuable for the identification and purification of individual defence enzymes, whichmay serve as indicators of disease resistance in plants (Popp et al. 1997). Considering theimportance of phenylpropanoid metabolism in plant defence mechanisms, it seemspossible that some phytotoxins might be capable of specifically suppressing the activationof this pathway in the host tissues. To investigate this possibility, we took advantage of thefact that rice suspension cultured cells that are characteristic of intact tissues where theinfluence of fungal toxins viz., Rhizoctonia solani toxin (RS toxin) and Sarocladium oryzaetoxin (SO toxin) on the suppression of defence mechanisms such as phenylalanineammonia-lyase (PAL), peroxidase, phenol, catalase, superoxide dismutase (Vidhyasekaranet al. 1992; Mohan et al. 1993; Mona 1994; Baker and Orlandi 1995; Shiraishi et al. 1995;Vurro and Ellis 1997), chitinase and b-1,3-glucanse (Graham and Graham 1991; Poppet al. 1997) were evaluated.
Materials and methods
Preparation of toxin
RS toxin
RS toxin from R. solani was purified as described earlier (Vidhyasekaran et al. 1997). R.solani isolates were grown on Richards’s broth (Fahmy 1923). The one-litre culture filtratewas collected after 21 days of incubation at room temperature. The filtrate was condensedto one tenth of its original volume by flash evaporation in vacuo at 508C. An equal volumeof methanol was added to the condensed culture filtrate and incubated overnight at 58C.The precipitate was removed by filtration and methanol was evaporated at 508C using avacuum flash evaporator. The water fraction was partitioned with equal volumes of ethylacetate, hexane and chloroform in sequence using a separating funnel. The partitionedwater fraction showing toxic action was condensed to half of its volume and subjected tocolumn chromatography (Pharmacia, USA) using Sephadex G-75 with water as eluant.Five mililitre fractions were collected, toxic fractions were pooled, evaporated to drynessin vacuo at 408C, weighed and re-dissolved in 5 ml of water. This partially purified toxinwas used in all further studies.
1408 A. Kandan et al.
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

SO toxin
SO toxin fromS. oryzaewas purified as described earlier (Hemalatha et al. 1999; Samiyappanet al. 2003). Richard’s broth (Fahmy 1923) was used for culturing the pathogen. Flasks withculture were incubated at 258C under stationary conditions for 20 days, then the culturefiltrates were harvested. After filtration, the culture fluid was concentrated to 10% of itsoriginal volume at 508C using a vacuum flask. The condensed culture fluid was treated withan equal volume of MeOH and kept overnight at 48C. Precipitates developed were removedby filtering throughWhatman No. 1 filter paper. Methanol was evaporated in vacuo and thewater fraction was partitioned with equal volumes of chloroform, ethylacetate, carbontetrachloride, hexane and ether in sequence using a separating funnel. The water fractionshowing the toxic activity was condensed to half of its volume, applied to Sephadex G-75column (2.5 6 60 cm Pharmacia, USA) and eluted with distilled water. Five mililitrefractions were collected and the toxic fractions were pooled.
Callus induction
The induction of callus and the initiation of cell suspension cultures were followed fromthe slightly modified protocols of Zhang et al. (1996). Rice cv. Pusa Basmati, IR 50 and Co43 were used for callus induction and further studies in all the experiments. The seeds werecollected from Paddy Breeding Station, Tamil Nadu Agricultural University, Coimbatore,India. Rice calli were produced from dehusked mature seeds. Dehusked seeds were surfacesterilised in 70% ethanol for 1 min, followed by 0.1% mercuric chloride for 8 min andrinsed with several changes of sterile distilled water. The seeds were plated on Petri dishescontaining MS (Murashige and Skoog 1962) medium supplemented with 2,4-D (2.0 mg/l)and incubated at 25 + 28C in darkness for four weeks. Then subclutured on fresh MSmedium supplemented with 2,4-D (2.5 mg/l), proline (500 mg/l), casein (300 mg/l) andkept under dark at 25 + 28C for 3–4 weeks. Phytagel 0.4% (w/v) was used as solidifyingagent instead of agar in all solid media used in this study.
Preparation of suspension cultured cells
Suspension-cultured cells were prepared by transferring 3 g of small, compact,embryogenic calli to 250 ml Erlenmeyer flask containing 50 ml of modified R2 medium(Ohira et al. 1973) supplemented with maltose (30 g/l) and 2,4-D (2 mg/l ). Flasks werekept on a rotary shaker at 125 rpm at 25 + 28C. Suspension-cultured cells weretransferred to fresh medium at weekly intervals.
Treatment of suspension-cultured cells
One mililitre of toxin (50 mg/ml glucose equivalents) was added to the suspension-culturedcells and incubated on a rotary shaker with 125 rpm at 25 + 28C. Suspension-culturedcells treated with 1 ml of sterile distilled water served as control. Cells were harvested atperiodic intervals and used for various analyses.
Assay of phenylalanine ammonia-lyase (EC. 4.5.1.5)
The PAL assay was carried out as per the method described by Brueske (1980). Suspension-cultured cells (200 mg) were extracted in 1.0 ml of 0.1 M sodium borate buffer (pH 7.0) and
Archives of Phytopathology and Plant Protection 1409
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

centrifuged at 10,000 g for 10 min at 48C, the supernatant served as the enzyme source. Theenzyme activity was expressed as mmol/min/g fresh weight of cultured cells.
Assay of peroxidase (EC.1.11.1.7)
Peroxidase activity was assayed using guaiacol as a hydrogen donor as previously reportedby Rathmell and Sequiera (1974). Suspension-cultured cells (200 mg) were extracted in1.0 ml of 0.1 M sodium phosphate buffer (pH 6.5) and centrifuged at 10,000 g for 10 minat 48C, the supernatant served as the enzyme source. The peroxidase activity was expressedas change in absorbance min/g of fresh weight of cultured cells.
Assay of phenols
The phenolic content was estimated as per the method of Zieslin and Benzaken (1993).Suspension-cultured cells (10 mg) were homogenised in 1 ml of 80% methanol and thismethanolic extract was utilised for phenol estimation. The phenol content was expressed inmg/g fresh weight of cultured cells.
Assay of catalase (EC. 1.11.1.6)
Catalase activity was assayed based on the decrease in H2O2 concentration was measuredas the decline in A240 during the first 10 min after initiation of the reaction (BeleidEl-Moshaty et al. 1993). Enzyme activity was expressed as change in absorbance min/gfresh weight of cultured cells.
Assay of superoxide dismutase (SOD) (EC. 1. 15. 1. 1)
Suspension-cultured cells (100 mg)were homogenised in a chilled pestle andmortar in 0.2 Mcitrate phosphate buffer (pH 6.5) containing 0.5%Triton X-100 (Sigma, USA), in 1:10 (w/v)ratio, centrifuged in the homogenate for 15 min at 20,000 g at 48C the supernatant wasused as the enzyme source. The assay mixture (3 ml) was prepared with 50 mM sodiumphosphate buffer (pH 7.8), 13 mM methionine, 75 mM Nitroblue tetrazolium (NBT),0.1 mM EDTA, 100 ml of the enzyme extract and 2 mM riboflavin. The assay was carriedout according to the protocol described earlier (Beleid El-Moshaty et al. 1993). The SODactivity was expressed as percentage inhibition of NBT photoreduction.
Assay of b-1,3-glucanase (EC.3.2.1.39)
The enzyme activity was colorimetrically assayed (Pan et al. 1991). Five hundredmilligrams of cultured cells were extracted with 0.05 M sodium citrate buffer (pH 5.0) at4C. The homogenate was centrifuged at 4C for 20 min at 10,000 rpm. The supernatantwas used as crude enzyme extract. The enzyme activity was expressed as g equivalents ofglucose/min/g fresh weight of cultured cells.
Assay of chitinase (EC.3.2.1.14)
The colorimetric assay of chitinase was carried out as per Boller and Mauch (1988). Fivehundred milligrams of cultured cells were extracted with 1 ml of 0.1 M sodium citratebuffer (pH 5.0) at 4C. The homogenate was centrifuged at 4C for 20 min at 10,000 rpm.
1410 A. Kandan et al.
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

The supernatant was used as crude enzyme extract. The enzyme activity was expressed asnmoles GlcNAc/equivalents/min/g fresh weight.
Statistical analysis
The data were analysed as completely randomised design (CRD) using the IRRI STATversion 92-1 program developed by the Biometrics Unit at the International Rice ResearchInstitute, the Philippines. The treatment means were compared by Duncan’s multiplerange test (DMRT) (Gomez and Gomez, 1984).
Results
Assay of PAL and PO
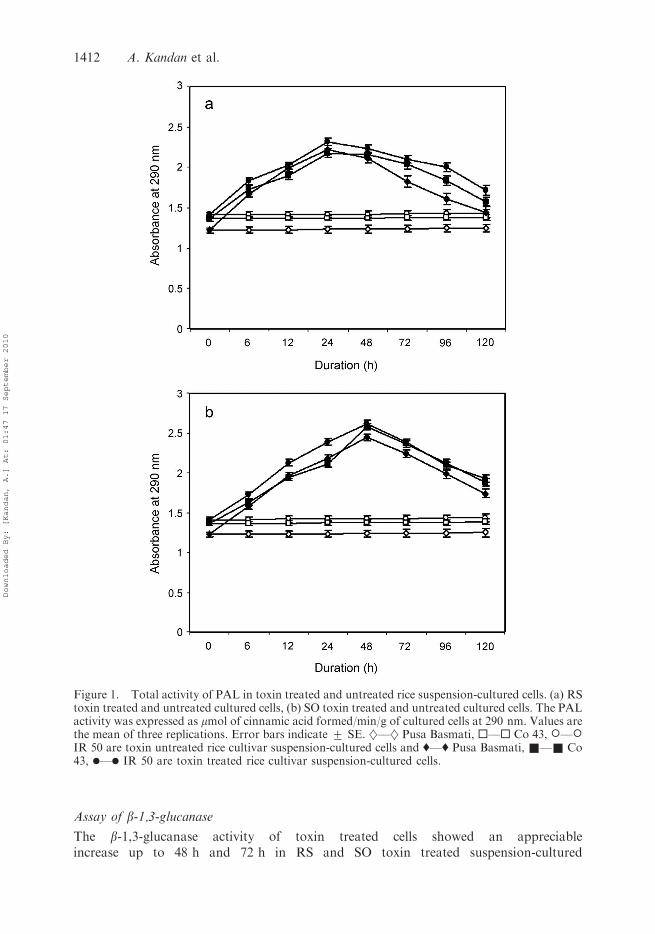
PAL and PO activities appeared to be stimulated by RS and SO toxin in all the ricecultivars. A significant difference in the PAL and PO activity was observed in toxin treatedsuspension-cultured cells compared to sterile distilled water treated suspension-culturedcells. After the addition of RS toxin PAL and PO activity showed twofold increase up to24 h in all the rice cultivars and then the effect was suppressed (Figures 1(a) and 2(a)),whereas the SO toxin increased the PAL and PO activity up to 48 h, and later thesedefence enzymes were decreased (Figures 1(b) and 2(b)).
Folin-ciocalteau assay
The RS toxin activates the phenolic content in suspension-cultured cells to a level ofthreefold in all the cultivars up to 24 h compared to control (Figure 3(a)). Similarly, asignificant increase was noticed in the SO toxin treated suspension-cultured cells up to48 h but later there was a gradual decrease in the phenolic content (Figure 3(b)). Therewas no appreciable increase in the phenolic content of sterile distilled water treatedsuspension-cultured cells.
Assay of catalase
A significant increase in catalase activity was noticed in RS and SO toxin treatedsuspension-cultured cells. Using the RS toxin, the cultivar IR 50 showed more catalaseactivity up to 24 h compared to the other cultivars and control, whereas in SO toxin all thecultivars showed high catalase activity up to 48 h compared to control. The catalaseactivity was high compared to RS toxin treated suspension-cultured cells (Figures 4(a) and(b)), but the catalase activity was suppressed after 24 h and 48 h in the RS and SO toxintreatment, respectively.
Superoxide dismutase (SOD) activity
SOD activity in suspension-cultured cells increased significantly from 6 h after thetreatment of suspension-cultured cells with RS and SO toxin. The SOD enzymeactivity gradually increased in the RS toxin treated suspension-cultured cells up to 72 hand up to 48 h in SO toxin treated suspension-cultured cells. Then the SOD activitygradually decreased in both the toxin treated suspension-cultured cells (Figures 5(a)and (b)).
Archives of Phytopathology and Plant Protection 1411
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010
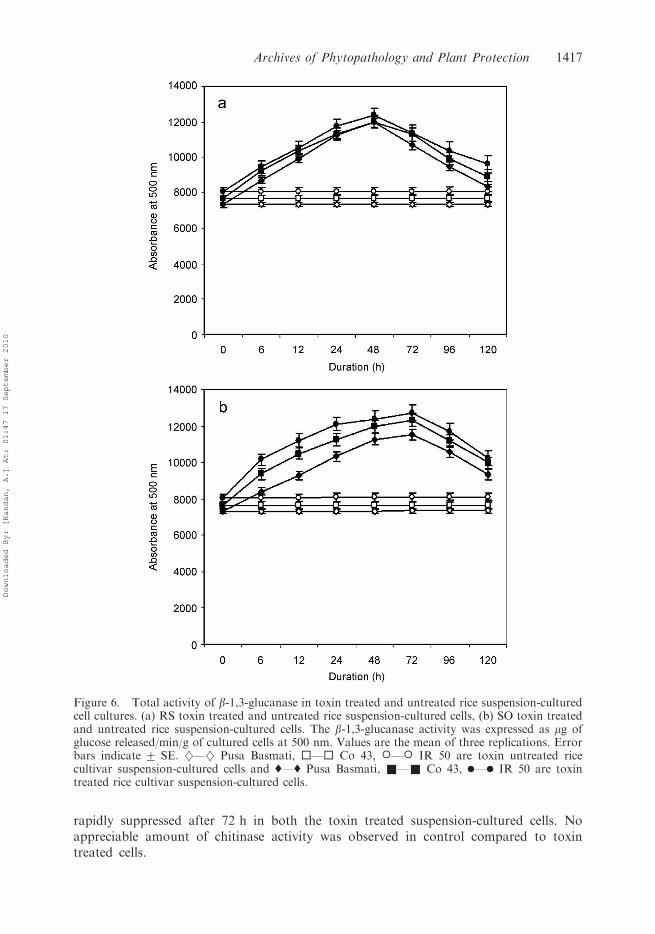
Assay of b-1,3-glucanase
The b-1,3-glucanase activity of toxin treated cells showed an appreciableincrease up to 48 h and 72 h in RS and SO toxin treated suspension-cultured
Figure 1. Total activity of PAL in toxin treated and untreated rice suspension-cultured cells. (a) RStoxin treated and untreated cultured cells, (b) SO toxin treated and untreated cultured cells. The PALactivity was expressed as mmol of cinnamic acid formed/min/g of cultured cells at 290 nm. Values arethe mean of three replications. Error bars indicate + SE. }––} Pusa Basmati, ¤––¤ Co 43, �––�IR 50 are toxin untreated rice cultivar suspension-cultured cells and '––' Pusa Basmati, &––& Co43, .––. IR 50 are toxin treated rice cultivar suspension-cultured cells.
1412 A. Kandan et al.
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

cells, respectively. Similarly the b-1,3-glucanase activity was later suppressed inboth the toxin treated suspension-cultured cells. The b-1,3-glucanase activitywas more in rice cv. IR 50 compared to the other cultivars in both RS and
Figure 2. Activity of PO in toxin treated and untreated rice suspension-cultured cells. (a) RS toxintreated and untreated rice suspension-cultured cells, (b) SO toxin treated and untreated ricesuspension-cultured cells. The PO activity was expressed as change in absorbance/min/g of culturedcells at 470 nm. Values are the mean of three replications. Error bars indicate + SE. }––} PusaBasmati, ¤––¤ Co 43, �––� IR 50 are toxin untreated rice cultivar cells and '––' Pusa Basmati,&––& Co 43, .––. IR 50 are toxin treated rice cultivar cells.
Archives of Phytopathology and Plant Protection 1413
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

SO toxin treated suspension-cultured cells (Figures 6(a) and 6(b)). The b-1,3-glucanase activity was not found to a significant level in control compared to toxintreated cells.
Figure 3. Total phenol content in toxin treated and untreated rice suspension-cultured cells. (a) RStoxin treated and untreated rice suspension-cultured cells, (b) SO toxin treated and untreated ricesuspension-cultured cells. The phenol content was expressed as mg/g of cultured cells. Values are themean of three replications. Error bars indicate + SE. }––} Pusa Basmati, ¤––¤ Co 43, �––� IR50 are toxin untreated rice cultivar suspension-cultured cells and '––' Pusa Basmati, &––& Co 43,.––. IR 50 are toxin treated rice cultivar suspension-cultured cells.
1414 A. Kandan et al.
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

Chitinase assay
Chitinase activity was noticed to appreciable level in both RS and SO toxintreated suspension-cultured cells up to 72 h and nearly threefold increase of chitinase
Figure 4. Activity of catalase in toxin treated and untreated rice suspension-cultured cells. (a) RStoxin treated and untreated rice suspension-cultured cells, (b) SO toxin treated and untreated ricesuspension-cultured cells. The catalase activity was expressed as change in absorbance min/g ofcultured cells at 240 nm. Values are the mean of three replications. Error bars indicate + SE. }––}Pusa Basmati, ¤––¤ Co 43, �––� IR 50 are toxin untreated rice cultivar suspension-cultured cellsand '––' Pusa Basmati, &––& Co 43, .––. IR 50 are toxin treated rice cultivar suspension-cultured cells.
Archives of Phytopathology and Plant Protection 1415
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

activity was found in RS and SO toxin treated suspension-cultured cells. Thechitinase activity was higher in suspension-cultured cells after the addition of SOtoxin compared to RS toxin (Figures 7(a) and 7(b)). The chitinase activity was
Figure 5. Total activity of superoxide dismutase in toxin treated and untreated rice suspension-cultured cell cultures. (a) RS toxin treated and untreated rice suspension-cultured cells, (b) SO toxintreated and untreated rice suspension-cultured cells. Superoxide dismutase activity was expressed aspercentage inhibition of NBT photoreduction in the rice callus at 560 nm. Values are the mean ofthree replications. Error bars indicate + SE. }––} Pusa Basmati, ¤––¤ Co 43, �––� IR 50 aretoxin untreated rice cultivar suspension-cultured cells and '––' Pusa Basmati, &––& Co 43, .––.IR 50 are toxin treated rice cultivar suspension-cultured cells.
1416 A. Kandan et al.
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010
rapidly suppressed after 72 h in both the toxin treated suspension-cultured cells. Noappreciable amount of chitinase activity was observed in control compared to toxintreated cells.
Figure 6. Total activity of b-1,3-glucanase in toxin treated and untreated rice suspension-culturedcell cultures. (a) RS toxin treated and untreated rice suspension-cultured cells, (b) SO toxin treatedand untreated rice suspension-cultured cells. The b-1,3-glucanase activity was expressed as mg ofglucose released/min/g of cultured cells at 500 nm. Values are the mean of three replications. Errorbars indicate + SE. }––} Pusa Basmati, ¤––¤ Co 43, �––� IR 50 are toxin untreated ricecultivar suspension-cultured cells and '––' Pusa Basmati, &––& Co 43, .––. IR 50 are toxintreated rice cultivar suspension-cultured cells.
Archives of Phytopathology and Plant Protection 1417
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

Discussion
Our studies suggest that toxins suppress defence mechanisms of the plants and permitcolonisation of the pathogen. However, severity of disease depends on the succession ofinteractions between pathogen and plant. Generally, toxins are known as aggressins,
Figure 7. Total activity of chitinase in toxin treated and untreated rice suspension-cultured cells.(a) RS toxin treated and untreated rice suspension-cultured cells, (b) SO toxin treated and untreatedrice suspension-cultured cells. Activity chitinase was expressed as nmoles GlcNAc equivalents/min/gof fresh callus at 585 nm. Values are the mean of three replications. Error bars indicate + SE. }––}Pusa Basmati, ¤––¤ Co 43, �––� IR 50 are toxin untreated rice cultivar suspension-cultured cellsand '––' Pusa Basmati, &––& Co 43, .––. IR 50 are toxin treated rice cultivar suspension-cultured cells.
1418 A. Kandan et al.
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

which breach the host defence barrier. Toxins produced by the rice sheath blight pathogenR. solani and sheath rot pathogen S. oryzae induced typical symptoms in all rice cultivarstested which are susceptible to these pathogens (Vidhyasekaran et al. 1992, 1997;Mariappan 1996; Samiyappan et al. 2003). PAL is the first enzyme in thephenylpropanoid pathway, which leads to production of phytoalexins, phenolicsubstances and formation of lignin with the help of peroxidases (Kandan et al. 2002).By examining these defence enzymes by the addition of toxin may help to understand themolecular mechanisms of disease suppression against these pathogens. The RS and SOtoxin treated suspension-cultured rice cells showed an appreciable increase in PAL and POfrom 6 h to 24 h in RS toxin and up to 48 h in SO toxin, but gradually these enzymes weresuppressed which leads to the suppression of forthcoming defence enzymes. Our resultssupport the hypothesis originally proposed by Vidhyasekaran et al. (1992) thatHelminthosporium oryzae toxin suppresses the PAL and PO enzyme activity in riceplants. Vurro and Ellis (1997) reported that induction and suppression of PAL enzymeactivity was observed due to several fungal toxins. The next important defence mechanismagainst pathogen is brought about by the phenolic content of the host. Phenols play animportant role in determining resistance or susceptibility of a host to pathogen infection(Vidhyasekaran 1988). The accumulation of phenolic compounds due to infection bypathogens has been reported by several authors (Borah et al. 1978; Arora 1983;Vidhyasekaran et al. 1992). In our study, an increase in phenolic content was observed upto 24 h in RS toxin treated rice suspension-cultured cells and then suppressed to basallevel; similarly in SO toxin the treated suspension-cultured rice cells showed an increase inphenolic content up to 48 h and was then suppressed to basal level. This gradualsuppression of phenolic content in both toxin treated suspension-cultured rice cellsprovides evidence that the suppression of defence mechanism of the host against thepathogen leads to colonisation of the pathogen. Our results support the previous findingsof Kurosaki et al. (1987) that phenolic compounds like p-hydroxybenzoic acid, caffeic acidand ferulic acid increased in carrot cells in response to treatment with mycelial wallpreparation of Chaetomium globosum.
Catalase (scavenger) which converts H2O2 to H2O and O2 is found in most of the plantcells and plays a significant role in reducing high levels of H2O2. In our experiment, thecatalase increased up to 48 h and gradually suppressed due to toxin action. This reductionin the catalase activity in rice suspension-cultured cells provides evidence that a similarmechanism might be taking place in the host due to pathogen infection. The earlyreduction of catalase activity due to toxin treatment in the suspension-cultured cells ismainly due to rapid outburst of the active oxygen species particularly H2O2 (Gonner andSchlosser 1993; Milosevic and Slusarenko 1996). SOD is an important antioxidant, whichacts as scavenger of superoxide radicals that can cause extensive damage to the cellularcomponents and catalyze the dismutation of O2 and HO2 to H2O2. In the present studyrice suspension-cultured cells treated with RS and SO toxin showed an increase in SODactivity up to 72 h and 48 h, respectively, and later suppression was observed. The increasein SOD level up to a certain period may be due to stress created by the toxins. Theseresults are supported by the previous findings of Vera Estrella et al. (1993) of the activityof SOD against Cladosporium fulvum in tomato suspension cultured cells.
The pathogenesis-related (PR) proteins are host-encoded polypeptides that accumulateas a result of infection by pathogen. The biochemical functions of some groups of PRproteins have been established but all PR proteins possess antimicrobial activity (VanLoon et al. 1994; Brunner et al. 1995). The b-1,3-glucanase (PR-2) and chitinases (PR-3)have the potential to hydrolyse the b-1,3-glucan and chitin, respectively, which are the
Archives of Phytopathology and Plant Protection 1419
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

major components of fungal cell walls leading to direct inhibition of the growth of severalfungi in vitro (Leah et al. 1991). The b-1,3-glucanase (PR-2) activity increased up to 48 hand 72 h in RS and SO toxin treated suspension-cultured rice cells, respectively, but laterthis activity was suppressed which may be the reason for paving the way for pathogencolonisation in the host. These findings were in agreement with previous reports ofMazars et al. (1990) revealing stimulated and exhausted level of PR-2 activity in barley bytoxic glycoprotein from Rhynchosporium secalis. Madi and Katan (1998) showed increasedand decreased activity of PR-2 by Penicillium janczewskii on melon, while Anguelova et al.(1999) reported increased and decreased activity of PR-2 in wheat by leaf rust pathogen.The chitinase activity increased in suspension-cultured rice cells treated with both RS andSO toxin and later the activity was suppressed. Even though the increase in activity wastwo–three fold, the rapid suppression of the activity leads to pathogen colonisation in thehost. A similar observation was recorded by Popp et al. (1997) in pine due to Ophiostomaminus.
Many researchers have suggested that toxins may suppress defence mechanisms of theplants (Sheffer 1976; Hayami et al. 1980; Otani et al. 1980; Ouchi and Oku 1982), but thereare only a few published reports supporting the hypothesis (Tietjen and Suzuki 1984;Vidhyasekaran et al. 1992; Vurro and Ellis 1997). In this study, the activity of two toxin onsuspension-cultured rice cells clearly shows the suppression of various defence enzymes,including phenylpropanoid metabolism, PR-proteins etc. This toxic activity is postulated asunregulated metabolic process firstly exhausting the cells and secondly leading to starvationof the plant followed by these uncontrolled defence mechanisms induced by toxic moleculesresulting in death of the host at the final stage. In future, there is a possible attempt todevelop disease resistance plants against these disease by overexpression of this defence gene,namely PAL gene, chitinase gene, etc. Transgenic tobacco plants expressing the chitinasegene constitutively show a high degree of resistance against R. solani (Broglie et al. 1991).Thus, these defence genes which delay suppression by these pathotoxins may be highlyuseful to develop transgenic plants with disease resistance.
References
Anguelova VA, Westhuizen AJ, VanDerand Pretarious ZA. 1999. Intercellular proteins and b-1,3-glucanase activity associated with leaf resistance in wheat. Physiol Plantarum. 106:393–401.
Arora YK. 1983. Metabolic changes in mung bean (Vigna radiata) due to Rhizoctonia solani Kuhninfection. Plant Physiol Biochem. 10:40–45.
Baker CJ, Orlandi EW. 1995. Active oxygen in plant pathogenesis. Ann Rev Phytopathol.33:299–321.
Beleid El-Moshaty FL, Pike SM, Novacky AJ, Sehgal OP. 1993. Lipid peroxidation and superoxideproduction in cowpea (Vigna unguiculata) leaves infected with tobacco ringspot virus or southernbean mosaic virus. Physiol Mol Plant Pathol. 43:109–119.
Boller T, Mauch F. 1988. Colorimetric assay for chitinase. Meth Enzymol. 161:430–435.Borah RC, Bajaj KL, Bhatia IS. 1978. Biochemical changes in tea leaves after infection with red rust
Cephaleuros parasiticus Phytopath Z. 93:208–217.Broglie K, Chet I, Holliday M, Cressman R, Biddle P, Knowlton S, Mauvais CJ, Broglie R. 1991.
Transgenic plants with enhanced resistance to the fungal pathogen Rhizoctonia solani. Science.254:1194–1197.
Brueske CH. 1980. Phenylalanine ammonia-lyase in tomato roots infected and resistant to the rootknot nematode, Meloidogyne incognita. Physiol Plant Pathol. 16:409–414.
Brunner F, Baillieuli F, Geneter I, Geoffroy P, Kauffmann S, Fritig B. 1995. The hypersensitivereaction in tobacco as a model to study the regulation of synthesis and the biological activities ofPR proteins. Abstracts of 4th International workshop on Pathogenesis-related proteins in Plants:Biology and Biotechnological Potential; Kloster Isree, Germany.
1420 A. Kandan et al.
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

Chen CC, Chien CC. 1964. Some observations on the outbreak of rice sheath rot disease. J TaiwanAgric Res. 11:1–19.
Cutler HG. 1993. Phytotoxins of microbial origin. In: Keeler RF, Tu AT, editors. Handbookof natural toxins: Toxicology of plant and fungal compounds. New York: Marcel Dekker.p. 411–438.
Dietrich A, Mayer JE, Hahlbrock K. 1990. Fungal elicitor triggers rapid, transient andspecific protein phosphorylation in parsley cell suspension cultures. J Biol Chem. 265:6360–6368.
Dixon RA, Harrison MJ, Lamb CJ. 1994. Early events in the activation of plant defence responses.Ann Rev Phytopathol. 32:479–501.
Fahmy T. 1923. The production by Fusarium solani of a toxic excretory substance capable of causingwilting of plants. Phytopathology. 13:543–550.
Gomez KA, Gomez AA. 1984. Statistical procedure for Agricultural Research. New York: JohnWiley and Sons. p. 320–356.
Gonner MV, Schlosser E. 1993. Oxidative stress in interactions between Avena sativa L. andDreschlera spp. Physiol Mol Plant Pathol. 42:221–234.
Graham TL, Graham MY. 1991. Cellular coordination of molecular responses in plant disease. MolPlant-Micro Interact. 4:415–422.
Hayami C, Otani H, Nishimura S, Kohmoto K. 1980. Induced resistance in pear leaves by sporegermination, fluids of non-pathogens to Alternaria alternata, Japanese pear pathotype, andsuppression of the induction by AK-toxin. Ann Phytopathol Soc Jpn. 45:536–537.
Hemalatha RG, Jebaraj S, Raja JVJ, Raguchander T, Ramanathan A, Samiyappan R,Balasubramanian P. 1999. Employing a crude toxin preparation from Sarocladium oryzae as amolecular sieve to select sheath rot-resistant somaclones of rice. J Plant Biochem Biotech.8:75–80.
Jimenez SA, Parizos SC. 1975. Some observations on the outbreak of rice sheath rot disease.J Taiwan Agric Res. 11:1–19.
Kandan A, Radja commare R, Nandakumar R, Raguchander T, Ramiah M, Samiyappan R. 2002.Induction of Phenylpropanoid metabolism by Pseudomonas fluorescens against tomato spottedwilt virus in tomato. Folia Microbiol. 47:121–129.
Kodama M, Inoue K, Otani H, Kohmoto K. 1995. Cultivar specific and non-specific responsees intomato cell cultures to AL-toxin from Alternaria alternata tomato pathotype. Ann PhytopatholSoc Jpn. 81:582–585.
Kurosaki F, Tashino N, Nishi A. 1987. Induction, purification and possible function of chitinase incultured carrot cells. Physiol Mol Plant Pathol. 31:201–210.
Lamb C, Lawton M, Dron M, Dixon R. 1989. Signals and transduction mechanisms for activationof plant defences against microbial attack. Cell. 56:215–224.
Leah R, Tommerup H, Svendsen I, Mundy J. 1991. Biochemcial and molecular characterisation ofthree barley seed proteins with antifungal properties. J Biol Chem. 266:1564–1573.
Madi L, Katan J. 1998. Penicillium janczewskii and its metabolites, applied to leaves elicit systemicacquired resistance to stem rot caused by Rhizoctonia solani. Physiol Mol Plant Pathol.53:163–175.
Mariappan C. 1996. Studies on induction of antifungal proteins in rice with special reference toinfection by the Sheath rot pathogen Sarocladium oryzae Saw. M.Sc thesis, Coimbatore (India):Tamil Nadu Agricultural University.
Mazars C, Lafitte C, Marquet PY, Rossignol M, Auriol P. 1990. Elicitor-like activity of the toxicglycoprotein isolated from Rhynchosporium secalis (OUD.) Davis culture filtrates. Plant Sci.69:11–17.
Milosevic N, Slusarenko AJ. 1996. Active oxygen metabolism and lignification in the hypersensitiveresponse in bean. Physiol Mol Plant Pathol. 49:143–158.
Mohan R, Bajeeis AM, Kolattukudy PE. 1993. Induction to tomato anionic peroxidasegene activation (tap I) by wounding in transgenic tobacco and activatioin of tap 1/GUS andtap 2/GUS chimeric gene fusions in transgenic tobacco by wounding and pathogen attack. PlantMol Biol. 21:341–354.
Mona CM. 1994. Active oxygen species in plant defence against pathogens. Plant Physiol. 105:467–472.
Murashige T, Skoog F. 1962. A revised medium for a rapid growth and bioassay with tobacco tissueculture. Physiol Plantarum. 15:473–479.
Archives of Phytopathology and Plant Protection 1421
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010

Ohira K, Ojima K, Fijiwara A. 1973. Studies on the nutrition of rice cell culture I. A simple, definedmedium for rapid growth in suspension culture. Plant cell Physiol. 14:1113–1121.
Otani H, Hayami C, Yoshimura Y, Nishimura S, Kohmoto K. 1980. Different responses of heat-shocked host tissues to Alternaria alternata Japanese pear pathotype and its toxin. AnnPhytopathol Soc Jpn. 46:386–387.
Ouchi S, Oku H. 1982. Physiolgical basis of susceptibility induced by pathogens. In: Asada Y,Bushnell WR, Ouchi S, Vance CP, editors. Plant Infection: The Physiological and BiochemicalBasis. Tokyo: Japan Scientific Press. Berlin: Springer-Verlag. p. 117–136.
Pan Q, Te YS, Kuc J. 1991. A technique for detection of chitinases, b,-1,3 glucanases and proteinpatterns after single separation using PAGE or isoelectic focussing. Phytopathology. 81:970–974.
Popp MP, Lesney MS, Davis JM. 1997. Defence responses elicited in pine cell suspension cultures.Plant Cell Tissue Org Cult. 47:199–206.
Rathmell WG, Sequiera L. 1974. Soluble peroxidase in fluid from the intercellular spaces of tobaccoleaves. Plant Physiol. 53:317–318.
Roy AK. 1993. Sheath blight of rice. Indian Phytopathol. 46:197–205.Samiyappan R, Amutha G, Kandan A, Nandakumar R, Babu S, Vijayasamundeeswari A,
Radjacommare R, Ramanathan A, Balasubramanian P. 2003. Purification and partialcharacterisation of a phytotoxin produced by Sarocladium oryzae, the rice sheath rot pathogen.Arch. Phytopathol Plant Prot. 36:247–256.
Sawada K. 1992. Descriptive catalogue of Formosan fungi II. Rep: Govt. Res. Inst. Dep. AgricFormosa. 2:136.
Shajahan AKM, Harahap Z, Rush MC. 1977. Sheath rot of rice caused by Acrocylindrium oryzaeSaw. in Louisiana. Plant Dis Report 61:307–310.
Sheffer RP. 1976. Host-specific toxins in relation to pathogenesis and disease resistance. In: HeitfussR, Williams PH, editors. Encyclopedia of Plant Physiology, New Series, Vol. I, PhysiologicalPlant Pathology. Berlin: Springer-Verlag. p. 247–269.
Shiraishi T, Yamada T, Nicholson RL, Kuroh H. 1995. Phenylalanine ammonia lyase in barley:activity enhancement in response to Erysipe graminis f. sp. hordei (race 1) a pathogen, andErysiphe pisi, a nonpathogen. Physiol Mol Plant Pathol. 46:153–162.
Tietjen KG, Suzuki U. 1984. Induction and suppression of phytoalexin biosynthesis in cultured cellsof safflower, Carthamus tinctorius L., by metabolites of Alternaria carthami Choudhury. ArchBiochem Biophys. 229:136–144.
Van Loon LC, Pierpoint WS, Boller T, Conejero V. 1994. Recommendation for naming plantpathogenesis-related proteins. Plant Mol Biol Report. 12:245–264.
Vera-Estrella R, Blumwald E, Higgins VJ. 1993. Non-specific glycopeptide elicitors of Cladosporiumfulvum L. evidence for involvement of active oxygen species in elicitor-induced effects on tomatocell suspensions. Physiol Mol Plant Pathol. 42:9–22.
Vidhyasekaran P. 1988. Physiology of disease resistance in plants. Vol. II. Boca Raton (FL): CRCPress, Inc. p. 125–130.
Vidhyasekaran P, Borromeo ES, Mew TW. 1992. Helminthosporium oryzae toxin suppressesphenol metabolism in rice plants and aids pathogen colonisation. Physiol Mol Plant Pathol.41:307–315.
Vidhyasekaran P, RubyPonmalar T, Samiyappan R, Velazhahan R, Vimala R, Ramanathan R,Paranidharan V, Muthukrishnan S. 1997. Host-Specific toxin production by Rhizoctonia solani,the rice sheath blight pathogen. Phytopathology. 87:1258–1263.
Vurro M, Ellis BE. 1997. Effect of fungal toxins on induction of phenylalanine ammonia-lyaseactivity in elicited cultures of hybrid poplar. Plant Sci. 126:29–38.
Zhang S, Lili C, Rongda Qu, Phillippe M, Roger B, Fauquet C. 1996. Regeneration of fertiletransgenic indica (group 1) rice plants following microprojectile transformation of embryogenicsuspension culture cells. Plant cell Report 15:465–469.
Zieslin N, Ben-Zaken R. 1993. Peroxidase activity and presence of phenolic substances in pedunclesof rose flowers. Plant Physiol Biochem. 31:333–339.
1422 A. Kandan et al.
Downloaded By: [Kandan, A.] At: 01:47 17 September 2010
Copyright © 2022 FDOKUMEN




















