
Reactive oxygen species generation and antioxidant systems in plant mitochondria

Cross-References
▶ Potassium in Biological Systems
▶Metals and the Periodic Table
▶Thallium, Distribution in Animals
▶Thallium, Effects on Mitochondria
References
Berg T, Steinnes E (1997) Use of mosses (Hylocomiumsplendens and Pleurozium schreberi) as biomonitors of
heavy metal deposition: from relative to absolute deposition
values. Environ Poll 98(1):61–71
Djingova R, Ivanova J, Wagner G, Korhammer S,
Markert B (2001) Distribution of lanthanoids, Be, Bi,
Ga, Te, Tl, Th and U on the territory of Bulgaria using
Populus nigra ‘Italica’ as an indicator. Sci Total
Environ 280:85–91
Fraenzle S, Markert B (2002) The Biological System of the
Elements (BSE) – a brief introduction into historical and
applied aspects with special reference to “ecotoxicological
identity cards” for different elements (f.e. As and Sn).
Environ Pollut 120:27–45
Fraenzle S, Markert B, Wuenschmann S (2012) Introduction to
environmental engineering. Wiley-VCH, Weinheim
Fr€anzle S (2010) Chemical elements in plants and soil. Springer,
Berlin
Markert B (1992) Multi-element analysis in plants – analyt-
ical tools and biological questions. In: Adriano DC (ed)
Biogeochemistry of trace metals. Lewis, Boca Raton,
pp 401–428
Markert B (1994) The Biological System of the Elements (BSE)
for terrestrial plants (glycophytes). Sci Total Environ
155:221–228
Markert B (1996) Instrumental element and multielement anal-
ysis of plant samples – methods and applications. Wiley,
Chichester
Markert B, Breure A, Zechmeister H (2003) General
aspects and integrative approaches. In: Markert B,
Breue T, Zechmeister H (eds) Bioindicators &
Biomonitors. Priciples, Concepts & Applications.
Elsevier, pp 3–39
Markert B, Wuenschmann S, Fraenzle S, Figueiredo A,
Ribeiro AP, Wang M (2011) Bioindication of trace metals –
with special reference to megacities. Environ Pollut
159:1991–1995
Sasmaz A, Sen O, Kaya G, Yaman M, Sagiroglu A (2007)
Distribution of thallium in soil and plants growing in the
Keban mining district of Turkey and determined by ICP-
MS. Atomic Spectrosc 28(5):157–163
Schedlbauer OF, Heumann KG (2000) Biomethylation of
thallium by bacteria and first determination of biogenic
dimethylthallium in the ocean. Appl Organomet Chem
14:330–340
Wappelhorst O (1999) Charakterisierung atmosph€arischerDepositionen in der Euroregion Neiße durch ein
terrestrisches biomonitoring. PhD thesis, IHI Zittau
Wierzbicka M, Szarek-Łukaszewska G, Grodzinska K (2004)
Highly toxic thallium in plants from the vicinity of Olkusz
(Poland). Ecotoxicol Environ Saf 59(1):84–88
Wolterbeek HT, Garty J, Reis MA, Freitas MC
(2003) Biomonitors in use: lichens and metal air pollution.
In: Markert B, Breure AM, Zechmeister HG (eds)
Bioindicators and biomonitors, principles, concepts and
applications. Elsevier, Amsterdam, pp 377–419
Xiao T, Guha J, Boyle D, Liu CQ, Chen J (2004) Environmental
concerns related to high thallium levels in soils and thallium
uptake by plants in southwest Guizhou, China. Sci Total
Environ 318(1–3):223–244
Thallium, Effects on Mitochondria
Sergey M. Korotkov
Sechenov Institute of Evolutionary Physiology
and Biochemistry, The Russian Academy of Sciences,
St. Petersburg, Russia
Synonyms
Thallium: Thallium(I), Tl(I), Tl+; Effects: Biological
action, Influence; Mitochondria: Intracellular organ-
elles, Organoids
Definitions
Thallium: Thallium is a metal with atomic number of
81 and two oxidation states of +1 and +3. Thallium
belongs to the group of trace elements and is used in
the electronics industry, in the pharmaceutical industry
(with care), and in glass manufacturing. Thallium is
highly toxic and was used in rat poisons and
insecticides.
Mitochondria: Mitochondria are two-membrane
granular or prolate organelles of 0.5 mm size which
are located between the cytoplasm and nuclear mem-
branes of the majority eukaryotic cells of both auto-
troph (photosynthesizing plants) and heterotroph
(mushrooms and animal) organisms. The basic function
of mitochondria is oxidation of organic compounds
following use of energy, released at their disintegration,
in creation of both the proton gradient and electrochem-
ical potential on the inner mitochondrial membrane for
the purpose of ATP synthesis which occurs by the
mitochondrial H(+)-ATP-synthase.
Thallium, Effects on Mitochondria 2193 T
T

Additional Definitions
Mitochondrial swelling: Mitochondrial swelling was
evaluated as a decrease in A540 at 20�C using a SF-46
spectrophotometer (LOMO, St. Petersburg, Russia).
Mitochondrial respiration: Mitochondrial respira-
tion (oxygen consumption rate) was measured polaro-
graphically by using LP-7 (Czechoslovakia) in a 1.5-ml
closed thermostatic chamber with magnetic stirring
at 26�C.Respiratory states: Respiratory states have been
defined by Chance and Williams (1956) according to
a protocol for oxygraphic experiments with isolated
mitochondria. State 3 is defined as the state after addi-
tion of ADP and state 4 as one after phosphorylation of
all ADP to ATP.
Mitochondrial membrane potential: Mitochondrial
membrane potential (DCmito) was evaluated as an
intensity of safranin fluorescence (arbitrary units) in
the mitochondrial suspension with magnetic stirring at
20�C using a Shimadzu RF-1501 spectrophoto-
fluorimeter (Shimadzu, Germany) at 485/590 nmwave-
length (excitation/emission).
Effects of Thallium on Cells and LivingOrganisms
Thallium is a highly toxic metal which belongs to
a group of trace elements. Human toxicity was found
in use of Tl compounds as a human depilatory (thal-
lium acetate), a component in manufacturing of optical
glasses (thallium oxide), a homicidal agent, and
a rodenticide (thallium sulfate) to kill rats, mice, and
other animals. After penetrating into an organism, Tl
damages cardiovascular, central nervous, and renal
systems as well as the gastrointestinal system and
skin, and it results in hair loss (Goel and Aggarwal
2007 and references herein). It has been shown that the
essence of the harmful effects of Tl+ on living organ-
isms lies in its ability both to easily penetrate the inner
mitochondrial membrane (Saris et al. 1981; Korotkov
et al. 2008 and #3 ref. herein) and to substitute K+ in
K+-dependent enzymes and biochemical processes
(Douglas et al. 1990). These effects of Tl+ resulted in
both the proximity of crystal-chemical radii of K+ and
Tl+, and easy thallium polarizability that allows Tl+
to form manifold chemical bonds with reactive groups
of molecules which constitute living organisms.
Experiments with rats, exposed to chronic thallium
intoxication in vivo, showed that Tl+ stimulated mas-
sive mitochondrial swelling which was followed by
disruption of mitochondrial and other intracellular
membranes of kidney, liver, brain, and other organs
(Korotkov et al. 2008, refs. of #9 and #10 herein). Tl+
has triggered apoptosis in Jurkat and PC12 cells
(Bragadin et al. 2003; Hanzel and Verstraeten 2009).
It was postulated earlier that Tl+ can stimulate release
of Ca2+ from intracellular compartments (Korotkov
et al. 2008, #9 ref. herein). Increase of cytoplasmic
concentration of Ca2+, Na+, or Pi and decrease of one
of K+ were both found in experiments with isolated rat
hepatocytes in medium containing TlCl. Interaction of
Tl+ with SH groups of mitochondrial and cellular
membranes, glutathione depletion, and the increased
production of reactive oxygen species can be among
other reasons of the thallium toxicity (Zierold 2000;
Korotkov et al. 2008, #9 ref. herein; Hanzel and
Verstraeten 2009). It was quite recently shown that
Tl+ could injure isolated rat hepatocytes that resulted
in hepatocyte proteolysis, in glutathione depletion, in
decline of the inner mitochondrial membrane potential
(DCmito), in ROS formation, and in lipid peroxidation
(Pourahmad et al. 2010).
Influence of Tl+ on Isolated Mitochondria
The study of swelling of isolated mitochondria in
nitrate media showed that the inner mitochondrial
membrane (IMM) is poorly penetrated by univalent
cations such as H+, K+, and Na+. However, visible
swelling of the mitochondria in TlNO3 medium
(Fig. 1) exposed substantial permeability of the mem-
brane to Tl+ (Saris et al. 1981; Korotkov et al. 2008).
Subsequent energization of the mitochondria stimu-
lated their massive contraction which occurred by
means of a Tl+/H+ exchange mechanism (Saris et al.
1981; Korotkov et al. 2008). Nonenergized mitochon-
dria in Tl acetate medium showed massive swelling
that was realized by means of Tl+/H+ exchange (Saris
et al. 1981; Korotkov et al. 2007). Further energization
of the mitochondria stimulated additional swelling due
to an electrophoretic uniport of Tl+ into the matrix
(Saris et al. 1981; Korotkov et al. 2007; Korotkov
et al. 2008, #3 ref. herein). The participation of the
mitochondrial KATP-dependent channel in the electro-
phoretic uniport of Tl+ in the matrix was demonstrated
T 2194 Thallium, Effects on Mitochondria

in both swelling and fluorescent experiments
(Wojtovich et al. 2010 and #45 ref. herein). Tl+
resulted in both the increase of state 4 respiration of
rat liver mitochondria (Fig. 2), and potent futile
cycling of Tl+ via the IMM (Bragadin et al. 2003;
Korotkov et al. 2007; Korotkov et al. 2008 and #3
ref. herein). Nonactin, a cyclic ionophore, and inor-
ganic phosphate (Saris et al. 1981; Korotkov et al.
2007; Korotkov et al. 2008 and #12 ref. herein) have
both facilitated transport of Tl+ into mitochondria and
increased both state 4 and swelling of mitochondria
(Figs. 1 and 2). With lower affinity to molecular
SH groups, Tl+ unlike bivalent heavy metals (Cd2+,
Zn2+, Hg2+, Cu2+, and Pb2+) has not inhibited state 3
or 2,4-dinitrophenol (DNP)-stimulated respiration
in medium containing TlNO3 or Tl acetate (Figs. 1
and 2) owing to the lack of inhibition of mitochondrial
respiratory enzymes by thallium (Korotkov et al. 2007;
Korotkov et al. 2008 and refs. of #3 and #10 herein;
Korotkov 2009).
Tl+ Increases the Permeability of the InnerMitochondrial Membrane to UnivalentCations (H+, K+, Na+, and Li+)
It was quite recently shown in experiments using
media containing nitrate salts of univalent cations
and TlNO3 (Korotkov 2009; Korotkov and Saris
2011) that Tl+ similar to the bivalent heavy metals
increased the permeability of the inner membrane of
rat liver mitochondria to univalent cations (H+, K+,
Na+, and Li+). The Tl+-induced increase of the perme-
ability (for more details, see Korotkov 2009 and
RLMRLM
RLM
RLM
SuccSucc
Succ Succ
25 25
50
50
75
75
0.052 min
0.05
2 min
ΔA540
ΔA540 ΔA540
ΔA540
50
75
75
0.1 0.1
3 min 3 min
0, 5, 1015
51015025
50
25
Pia b
c d
Thallium, Effects on Mitochondria, Fig. 1 Effects of Tl+ on
swelling of rat liver mitochondria in a 160 mOsm nitrate
medium. Mitochondria (1.5 mg protein/ml) were added to the
medium containing 5–75 mM TlNO3, as well as 5 mM Tris-NO3
(pH 7.3), 1 mM Tris-PO4 (b), 10�8 M nonactin (d), 4 mMrotenone, and 3 mg/ml of oligomycin. To obtain 160 mOsm,
sucrose is added to the medium. Additions of mitochondria
(RLM) and 5 mM succinate (Succ) are shown by arrows. Typ-ical traces for three different mitochondrial preparations are
presented. Concentrations of TlNO3 (mM) are shown on the
right of the traces (This material is reproduced with the courte-
ous permission of John Wiley & Sons, Inc. See Korotkov et al.
2008 and #6 ref. herein)
Thallium, Effects on Mitochondria 2195 T
T

a b
c d
e f
125
100
75
50
25
00
0
0 25 50 75
0 0
50
100
150
200
250
300
350
0
50
100
150
200
250
50
100
150
200
10 20 30
TI-acetate (mM)
ng a
tom
O/m
in /m
g of
pro
tein
ng a
tom
O/m
in /m
g of
pro
tein
ng O
/min
/mg
of p
rote
in
0
50
100
150
200
250
ng a
tom
O/m
in/m
g of
pro
tein
40 50 0 10 20 30
TI-acetate (mM)
40 50
0 10 20 30
TI-acetate (mM)
40 50
25 50
St 4 St 4 + DNP
St 4 + DNP
St 3
St 4
St 4 1
2
3
St 4 + DNP
St 3
2
2
2
2
1
1
1
1
2
1
TINO3 (mM)
TINO3 (mM)
ngat
om O
/min
/mg
of p
rote
in
150
200
250
300
100
50
0
ngat
om O
/min
/mg
of p
rote
in
75 100 125 0 25 50
TINO3 (mM)
75 100 125
Thallium, Effects on Mitochondria, Fig. 2 Effects of Tl+ on
oxygen consumption rates (ng atom O min/mg of protein) of rat
liver mitochondria in a 290 mOsm medium. Mitochondria
(1.5 mg protein/ml) were suspended in the medium containing
5 mM Tris-NO3 (pH 7.3) and 0–125 mM TlNO3 (a, b, e) or5 mM Tris acetate (pH 7.3) and 0–50 mM Tl acetate (c, d, f) aswell as 3 mM Mg(NO3)2 (c–f), 5 mM succinate, and 4 mMrotenone. Additionally the medium was supplemented with
3 mM Tris-PO4 (a–b [trace 2], and e–f) and 10�8 M nonactin
(c–d, f [trace 2]). To obtain 290 mOsm, sucrose was added to the
medium; 130 mM ADP or 30 mM DNP was added to the media
after 2 min recording of state 4 to induce state 3 or DNP-
stimulated respiration. Error bars were calculated by the Muller
formula from rates found for three different mitochondrial prep-
arations (see Korotkov 2009) (This material is reproduced with
the courteous permission of both John Wiley & Sons, Inc. (a–d,f) and Springer (e). See Korotkov et al. 2008 and #6 ref. herein;
Korotkov 2009)
T 2196 Thallium, Effects on Mitochondria

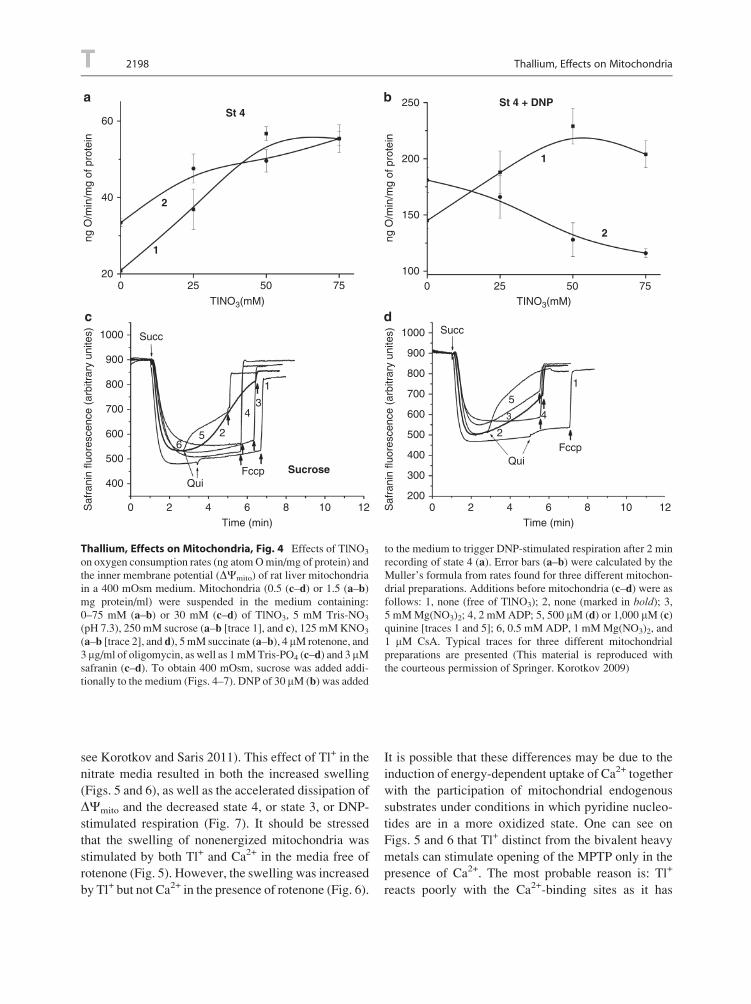
Korotkov and Saris 2011) was showed by the risen
swelling of both non-energized (Fig. 3) and energized
mitochondria, and by the accelerated dissipation of
DCmito (Fig. 4). Contraction of succinate-energized
mitochondria, swollen in the nitrate medium (Fig. 3d)
but not sucrose one (Fig. 3c), was inhibited by quinine,
which blocks mitochondrial K+/H+ exchange
(Korotkov 2009). The participation of the exchanger
in extruding the Tl+-induced excess concentration of
the univalent cations from the matrix was early
hypothesized (Saris et al. 1981; Korotkov et al.
2008). The Tl+-induced swelling of succinate-
energized mitochondria resulted in the decrease of
state 3 and 2,4-dinitrophenol (DNP)-stimulated respi-
ration (Fig. 4b) (more detail see Korotkov 2009).
Thallium Induces the PermeabilityTransition in the Inner Membrane ofCa2+-Loaded Rat Liver Mitochondria
It is known that binding of Ca2+ with matrix calcium-
specific trigger sites, located near the adenine
nucleotide translocase (ANT), with following fall of
DCmito triggers opening of the mitochondrial perme-
ability transition pore (MPTP) in the inner membrane
which becomes permeable to molecules up to
1,500 kDa. The pore opening results in massive
mitochondrial swelling, lowering of the matrix con-
centration of Ca2+ and ATP, and dissipation of the
DCmito. If the Ca2+ sites are insufficiently saturated,
MPTPs are opened in the low conductance state
and small molecules up to 300 kDa or ions
(H+, K+, and Ca2+) may penetrate easily the IMM.
In the earlier times, it was believed that the MPTP
is formed by ANT, cyclophilin D (CyP-D),
and the voltage-dependent anion channel (VDAC).
In modern times, many researchers consider the
mitochondrial phosphate carrier and CyP-D to be
primary components of the MPTP, whereas ANT is
viewed as a regulatory part of the MPTP.
It was recently showed that Tl+ has induced opening
of the MPTP in Ca2+-loaded rat liver mitochondria
(CaRLM) energized by glutamate plus malate or suc-
cinate which are substrates of I and II respiratory
complexes, respectively (Figs. 5–7) (for more details,
RLM Succ
RLMSucc
RLM
Succ
0255075
0.03
0.1
2 min
2 min
ΔA540
ΔA5400.1
2 min
ΔA540
RLM Succ
0
25
50
750.03
2 min
ΔA540
0500
0150
5001000
a b
c d
Thallium, Effects on Mitochondria, Fig. 3 Effects of Tl+ on
swelling of rat liver mitochondria in a 400 mOsm medium.
Mitochondria (1.5 mg protein/ml) were added to the medium
containing 0–75 mM TlNO3 (a–b) or 75 mM TlNO3 and
50–1,000 mM quinine (c–d) as well as 5 mM Tris-NO3 (pH
7.3), 250 mM sucrose (a, c), 125 mM of KNO3 (b, d), 4 mMrotenone, and 3 mg/ml of oligomycin. To obtain 400 mOsm,
sucrose was added additionally to the medium. Numbers near
the traces show concentrations of TlNO3 (mM) or quinine (mM)
on panels A and B or C and D, correspondingly. Additions of
mitochondria (RLM) and 5 mM succinate (Succ) are shown by
arrows. Typical traces for three different mitochondrial prepa-
rations are presented (This material is reproduced with the cour-
teous permission of Springer. Korotkov 2009)
Thallium, Effects on Mitochondria 2197 T
T
see Korotkov and Saris 2011). This effect of Tl+ in the
nitrate media resulted in both the increased swelling
(Figs. 5 and 6), as well as the accelerated dissipation of
DCmito and the decreased state 4, or state 3, or DNP-
stimulated respiration (Fig. 7). It should be stressed
that the swelling of nonenergized mitochondria was
stimulated by both Tl+ and Ca2+ in the media free of
rotenone (Fig. 5). However, the swelling was increased
by Tl+ but not Ca2+ in the presence of rotenone (Fig. 6).
It is possible that these differences may be due to the
induction of energy-dependent uptake of Ca2+ together
with the participation of mitochondrial endogenous
substrates under conditions in which pyridine nucleo-
tides are in a more oxidized state. One can see on
Figs. 5 and 6 that Tl+ distinct from the bivalent heavy
metals can stimulate opening of the MPTP only in the
presence of Ca2+. The most probable reason is: Tl+
reacts poorly with the Ca2+-binding sites as it has
60
40
200 25 50 75
TINO3(mM) TINO3(mM)0 25 50 75
121086420200
300
400
500
600
700
800
900
1000
Time (min) Time (min)
6
4 43
3
1 1
2 25
5
121086420
400
500
600
700
800
900
1000 Succ
FccpQui
Fccp
Succ
Qui
Saf
rani
n flu
ores
cenc
e (a
rbitr
ary
unite
s)
Saf
rani
n flu
ores
cenc
e (a
rbitr
ary
unite
s)
100
150
200
250
ng O
/min
/mg
of p
rote
in
ng O
/min
/mg
of p
rote
in
St 4St 4 + DNPa
2
2
1
1
b
c
Sucrose
d
Thallium, Effects on Mitochondria, Fig. 4 Effects of TlNO3
on oxygen consumption rates (ng atomOmin/mg of protein) and
the inner membrane potential (DCmito) of rat liver mitochondria
in a 400 mOsm medium. Mitochondria (0.5 (c–d) or 1.5 (a–b)mg protein/ml) were suspended in the medium containing:
0–75 mM (a–b) or 30 mM (c–d) of TlNO3, 5 mM Tris-NO3
(pH 7.3), 250 mM sucrose (a–b [trace 1], and c), 125 mMKNO3
(a–b [trace 2], and d), 5 mM succinate (a–b), 4 mM rotenone, and
3 mg/ml of oligomycin, as well as 1mMTris-PO4 (c–d) and 3 mMsafranin (c–d). To obtain 400 mOsm, sucrose was added addi-
tionally to the medium (Figs. 4–7). DNP of 30 mM (b) was added
to the medium to trigger DNP-stimulated respiration after 2 min
recording of state 4 (a). Error bars (a–b) were calculated by the
Muller’s formula from rates found for three different mitochon-
drial preparations. Additions before mitochondria (c–d) were asfollows: 1, none (free of TlNO3); 2, none (marked in bold); 3,5 mMMg(NO3)2; 4, 2 mM ADP; 5, 500 mM (d) or 1,000 mM (c)quinine [traces 1 and 5]; 6, 0.5 mM ADP, 1 mMMg(NO3)2, and
1 mM CsA. Typical traces for three different mitochondrial
preparations are presented (This material is reproduced with
the courteous permission of Springer. Korotkov 2009)
T 2198 Thallium, Effects on Mitochondria

a single charge and shows comparatively low affinity
to molecular SH groups. The phenomenon of the sub-
strate specificity manifested in the fact that the total
concentrations of Ca2+ stimulated the maximum swell-
ing of mitochondria energized by glutamate plus
malate (Fig. 5), but not by succinate plus rotenone
(Fig. 6). The possible roles of Ca2+-binding sites,
located near the respiratory complex I, and the ANT
in inducing opening of the MPTP are discussed
(Korotkov and Saris 2011).
It was discovered that the Tl+-induced MPTP in
CaRLM was potentiated by inorganic phosphate and
diminished by the MPTP inhibitors (ADP, CsA, Mg2+,
Li+, rotenone, EGTA, and ruthenium red) (Korotkov
et al. 2008; Korotkov 2009; Korotkov and Saris 2011).
The Tl+-induced swelling of CaRLM, energized by
glutamate plus malate (Fig. 5) or succinate in the
presence of rotenone (Fig. 6), was reduced by ADP,
CsA, Mg2+, or ruthenium red, an inhibitor of the mito-
chondrial Ca2+ uniporter. The MPTP inhibitors
prevented both the decreased DNP-stimulated respira-
tion and the fall of DCmito in succinate-energized
CaRLM (Fig. 7). Maximal effect was found in the
simultaneous presence of ADP and CsA in the
RLM
RLMRLM
Mg+ADP+CsAADP+CsAMg+CsA
Mg+ADPADP
control
Mg+CsA Mg+ADP+CsA
ADP+CsACsA RR
control
MgADP
Mg+ADP
Mg
RR
CsA
2 min
0.1Δ A540
G + M RLM G + M
G + M
KNO3
KNO3
G + M
0 +100 0 +100
25 +100
75 + 0
50 + 50
50 + 100
75 + 25
75 + 5075 + 7575 + 100
25 +10075 + 0
75 + 25
50 + 50
75 + 5075 + 7575 +100
Sucrose
a b
dc
Sucrose
50 + 100
Thallium, Effects onMitochondria, Fig. 5 Effects of Tl+ and
Ca2+ on swelling of rat liver mitochondria in the presence of
glutamate and malate. Mitochondria (1.5 mg protein/ml) were
added to the 400 mOsm medium containing 0–75 mM (a–b) or50 mM (c–d) of TlNO3, 5 mM Tris-NO3 (pH 7.3), 0–100 mM(a–b) or 100 mM (c–d) of Ca2+, and 1 mg/ml of oligomycin, as
well as 250 mM sucrose (a) or 125 mM KNO3 (b). The numbers
on the right of the traces (a–b) show concentrations of TlNO3,
(mM) [in bold] or CaCl2 (mM) [in italics] in this medium.
Additions before mitochondria are indicated on the right of the
traces (c–d): none (control); 1 mM CsA (CsA); 0.5 mM ADP
(ADP); 3 mM Mg2+ (Mg); 7 mM RR (RR). Injections of mito-
chondria (RLM) and 5 mM of glutamate and malate (G + M) are
shown by arrows. Typical traces for three different mitochon-
drial preparations are presented (This material is reproduced
with the courteous permission of Springer. Korotkov and
Saris 2011)
Thallium, Effects on Mitochondria 2199 T
T

media (Figs. 5–7). It was suggested that swelling of
energized mitochondria in the nitrate media containing
Ca2+ and Tl+ may be caused by opening of CsA-
inhibited and ADP-dependent pores in the IMM (for
more details, see Korotkov and Saris 2011). The Tl+-
induced swelling was inhibited by both Li+ (Korotkov
and Saris 2011) and Mg2+ (Figs. 5–7) that is possibly
related to the involvement of Ca2+ sites on the outer
surface of the IMM in opening the MPTP. The
decrease of state 4 and DNP-stimulated respiration of
CaRLM (Fig. 7) is related to the increased swelling of
the mitochondria (Fig. 6) that may be associated with
the reduced activity of the respiratory enzymes
because the mitochondrial structure can be disturbed
by the more massive swelling of CaRLM (Korotkov
and Saris 2011). Comparing experiments with the
sucrose and KNO3 media (Figs. 5 and 6), one can
conclude that Tl+, similar to Cd2+ and Ca2+, induces
opening of theMPTP less actively in the high conduction
states and furthermore, that this effect of Tl+ occurs in the
presence of the substrates of respiratory complex
I (glutamate plus malate) and II (succinate). Thus, it
RLM RLMSucca b
c dRLM
Succ
ADP(2) ADP+CsAMg+CsA
CsA
Mgcontrol
MgCsA
Mg+CsA
Mg+ADP+CsAADP+CsAADP(2)Mg+ADPnone CaADP
control
0.1
2 min
Δ A540
ADPnone Ca
Succ
RLMSucc
0 +10025 +100 50 +10050 + 50
75 + 25
75 + 0
75 + 50
75 + 7575 + 100
0 +10025 +10050 + 50
75 + 25
75 + 50
75 + 75
50 + 100
75 + 100
75 + 075 + 0
Sucrose
Sucrose
KNO3
KNO3
Thallium, Effects onMitochondria, Fig. 6 Effects of Tl+ and
Ca2+ on swelling of rat liver mitochondria in the presence of
succinate. The medium, additions, and designations are as
shown in the Fig. 5. Exceptions from Fig. 5 are that 5 mM
succinate and 2 mM rotenone were used there instead of 5 mM
of glutamate plus malate; 75 mM TlNO3 was used in experi-
ments with the MPTP inhibitors (c–d). Typical traces for threedifferent mitochondrial preparations are presented (This mate-
rial is reproduced with the courteous permission of Springer.
Korotkov and Saris 2011)
T 2200 Thallium, Effects on Mitochondria

57
5776
53
RLMa
c d
bRLM
DNP
KNO3
KNO3
Sucrose
Sucrose
SuccSucc
DNP
Mg+ADP+Cs
Mg+ADP+Cs
Mg+ADP
Mg+ADP
Mg
MgADP
ADP
400 ngatom O
2 min
0 2 4 6 8 10 12
Saf
rani
n flu
ores
cenc
e (a
rbitr
ary
unite
s)
Saf
rani
n flu
ores
cenc
e (a
rbitr
ary
unite
s)
400
600
800
1000
531
2
4FCCP
FCCP
Time (min)
200
400
600
800
1000
1
4
2
3 5
0 2 4 6 8 10 12
Time (min)
control
control
CsA
CsA
Pi
Pi
Ca2+
Ca2+
Ca2+
Ca2+
53
10994
139
5254
54
138
175
211
177
223
67
113 (190)
(208)
(211)
(181)
152(162)
(138)
64
(169)
(76)71
71
65
60
49
(49)
(49)
47
{21}
{145}
4877
80
9055
45
62
62(63)
(75)
97
17
3877
4472
{28}
{190}
64
59
8361
61
5399
5185
65
53
45
79
54
113
105
4373
5757
56
79
58
124
56(60)
(122)
(122)
(124)
(110)(121)
(94)
(83)
(133)
(51)(54)
(55)
(59)
(63)
(66)
(54)(41)
59
(57)
(57)
65(66)
(57)
(57)
(60)
(59)
(55)
57(52)
Thallium, Effects on Mitochondria, Fig. 7 Effects of TlNO3
and Ca2+ on oxygen consumption rates (ng atom O min/mg of
protein) and the inner membrane potential (DCmito) of rat liver
mitochondria in a 400 mOsm medium. The medium, additions,
and designations are as in the Fig. 4. Exception from Fig. 4 is
that additions of trace 5 on Fig. 7 are the same ones of trace 6 on
Fig. 4. The rates are presented as numbers placed above exper-
imental traces. Numbers in parentheses were obtained from
experiments with Ca2+-free media. The numbers in braces
were calculated from experiments with the Ca2+-free media,
where 75 mM TlNO3 was substituted by 150 mM sucrose
(Korotkov 2009). Accessorial additions of 30 mM (d) and
50 mM (c), or 100 mM (a–b) [traces 2–5] of Ca2+ (Ca2+) are
correspondingly shown by short (c–d) or long bold (a–b)arrows. Typical traces for three different mitochondrial prepa-
rations are presented (This material is reproduced with the cour-
teous permission of Springer. Korotkov and Saris 2011)
Thallium, Effects on Mitochondria 2201 T
T

can be postulated that opening of the MPTP can play an
important role in the development of toxic processes
during thallium intoxication in living organisms
(Bragadin et al. 2003; Korotkov et al. 2008; Korotkov
2009; Pourahmad et al. 2010; Korotkov and Saris 2011).
Cross-References
▶Ca2+
▶ Sodium-Hydrogen Exchangers, Structure and
Function in Human Health and Disease
▶Tobacco
References
Bragadin M, Toninello A, Bindoli A et al (2003) Thallium
induces apoptosis in Jurkat cells. Ann N Y Acad Sci
1010:283–291
Chance B, Williams GR (1956) The respiratory chain and
oxidative phosphorylation. Adv Enzymol 17:65–134
Douglas KT, Bunni MA, Baindur SR (1990) Thallium in bio-
chemistry. Int J Biochem 22:429–438
Goel A, Aggarwal P (2007) Pesticide poisoning. Natl Med
J India 20:182–191
Hanzel CE, Verstraeten SV (2009) Tl(I) and Tl(III) activate both
mitochondrial and extrinsic pathways of apoptosis in rat
pheochromocytoma (PC12) cells. Toxicol Appl Pharmacol
236:59–70
Korotkov SM, Glazunov VV, Yagodina OV (2007) Increase
in toxic effects of Tl+ on isolated rat liver mitochondria
in the presence of nonactin. J Biochem Mol Toxicol
21:81–91
Korotkov SM (2009) Effects of Tl+ on ion permeability,
membrane potential and respiration of isolated rat liver
mitochondria. J Bioenerg Biomembr 41:277–287
Korotkov SM, Saris NE (2011) Influence of Tl+ on
mitochondrial permeability transition pore in Ca2+-
loaded Rat liver mitochondria. J Bioenerg Biomembr
43:149–162
Korotkov SM, Emel’yanova LV, Yagodina OV (2008)
Inorganic phosphate stimulates the toxic effects of Tl+
in rat liver mitochondria. J Biochem Mol Toxicol
22:148–157
Pourahmad J, Eskandari MR, Daraei B (2010) A comparison of
hepatocyte cytotoxic mechanisms for thallium (I) and thal-
lium (III). Environ Toxicol 25:456–467
Saris NE, Skulskii IA, Savina MV et al (1981) Mechanism of
mitochondrial transport of thallous ions. J Bioenerg Biomembr
13:51–59
Wojtovich AP, Williams DM, Karcz MK et al (2010) A novel
mitochondrial K(ATP) channel assay. Circ Res 106:
1190–1196
Zierold K (2000) Heavy metal cytotoxicity studied by electron
probe X-ray microanalysis of cultured rat hepatocytes.
Toxicol In Vitro 14:557–563
Thallium, Physical and ChemicalProperties
Fathi Habashi
Department of Mining, Metallurgical, and Materials
Engineering, Laval University, Quebec City, Canada
Thallium is a silvery white metal, considered a less
typical metal because when it loses its outermost elec-
trons it will not achieve the inert gas electronic
structure. It occurs as a trace element in pyrite, and is
extracted as a by-product of roasting this mineral for
the production of sulfuric acid. Thallium can also be
obtained from the smelting of lead and zinc sulfide
ores. The fresh surface of the metal has a bluish-white
luster. In many of its physical properties, thallium
resembles its neighbor, lead. It exists in two allotropic
forms: a-thallium, hexagonal close-packed, stable at
room temperature; and b-thallium, body-centered
cubic, stable above 226�C. A volume increase of
3.23% takes place on solidification.
Thallium and its compounds are extremely
toxic. The odorless and tasteless thallium sulfate
was once widely used as rat poison and ant killer.
Thallium (I) sulfide’s electrical conductivity
changes with exposure to infrared light therefore
making this compound useful in photoresistors.
Thallium selenide has been used in a bolometer
for infrared detection.
Physical Properties
Atomic number 81
Atomic weight 204.38
Relative abundance in Earth’s crust, % 3 � 10�5
Density, g cm�3 11.85
Melting point, �C 303
Boiling point, �C 1,473
Atomic radius, pm 170
Heat of fusion, kJ mol�1 4.14
Heat of vaporization, kJ mol�1 165
Molar heat capacity, J mol�1 K�1 26.32
Electrical resistivity at 20�C, mO m 0.18
Thermal conductivity, W m�1 K�1 46.1
Thermal expansion at 25�C, mm m�1 K�1 29.9
Young’s modulus, GPa 8
(continued)
T 2202 Thallium, Physical and Chemical Properties
Copyright © 2022 FDOKUMEN




















