
A Babesia bovis gene syntenic to Theileria parva p67 is expressed in blood and tick stage parasites
Upload
kinhtecongnghiephanoiCategory
view
1download
0
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Molecular & Biochemical Parasitology 178 (2011) 40–45
Contents lists available at ScienceDirect
Molecular & Biochemical Parasitology
Short communication
Secretion of a new spherical body protein of Babesia bovis into the cytoplasm ofinfected erythrocytes
Mohamad Alaa Terkawia, Faasoa Junior Seuseua, Putut Eko-Wibowoa, Nguyen Xuan Huyena,Yuka Minodaa, Mahmoud AbouLailaa, Satoru Kawaib, Naoaki Yokoyamaa, Xuenan Xuana, Ikuo Igarashia,∗
a National Research Center for Protozoan Diseases, Obihiro University of Agriculture and Veterinary Medicine, Nishi 2-13, Inada-cho, Obihiro, Hokkaido 080-8555, Japanb Department of Tropical Medicine and Parasitology, Dokkyo Medical University, Mibu, Tochigi 321-0293, Japan
a r t i c l e i n f o
Article history:Received 10 September 2010Received in revised form 8 February 2011Accepted 14 February 2011Available online 13 March 2011
Keywords:Babesia bovisSpherical body protein 4SecretionErythrocytes
a b s t r a c t
A cDNA encoding a new Babesia bovis spherical body protein 4 (BbSBP-4) was reported to have no signifi-cant homology to other apicomplexan proteins or previously reported B. bovis spherical body proteins. Inthe present study, we further examined the molecular characteristics of BbSBP-4 including the expressionand cellular localization of the BbSBP-4. An anti-rBbSBP-4 mouse serum specifically reacted to a 41-kDanative protein B. bovis in Western blot analysis. The immunoelectron microscopic examination confirmedthe localization of BbSBP-4 in spherical bodies, but not in the nucleus, rhoptries, and micronemes. Inter-estingly, the protein was found to be localized not only in the spherical body of B. bovis but also inthe cytoplasm of infected erythrocytes (iRBC) at the later stage of parasite development. The confocallaser microscopic examination and Western blot analysis demonstrated the increased accumulation ofBbSBP-4 in the cytoplasm of iRBC and in the supernatant of cultivated B. bovis during the late develop-mental stage of the parasite. These results suggest that BbSBP-4 was secreted from spherical body intocytoplasm of iRBC during the late developmental stage of the parasite before the rupture of infectedRBC. Taken together, BbSBP-4 might play an important role as a secreted protein in the intracellulardevelopment and/or survival of B. bovis.
© 2011 Elsevier B.V. All rights reserved.
Babesia parasites are obligate intraerythrocytic protozoa,taxonomically classified in the phylum Apicomplexa, class Piro-plasmida, order Piroplasmida, which are capable of invading abroad range of vertebrate hosts, subsequently leading to seriouseconomic problems within the livestock industry throughout trop-ical and subtropical regions of the world [1]. The life cycle isrelatively similar among the order, and it requires a vertebratehost for the asexual stage as well as invertebrate hosts for thesexual stage. Babesia parasites initiate infection to mammalianhosts by sporozoites, which are transmitted through the bitesof infected ticks; subsequently, the merozoites invade and repli-cate within the infected erythrocytes (iRBC), eventually leading toacute babesiosis. Babesia bovis is the most economically importantspecies because of its impact on the cattle industry; it has abilityto modify the iRBC sequestered in microcapillaries of the kidneys,lungs, and brain, resulting in organ failure and systemic shock, lead-ing to death [1–3]. Cattle that have recovered from acute infectionbecome asymptomatic carriers, where the parasites persist in blood
∗ Corresponding author. Tel.: +81 155 49 5641; fax: +81 155 49 5643.E-mail address: [email protected] (I. Igarashi).
for years and recrudescence of parasitemia can occur at irregularintervals [3]. Although bovine babesiosis can be controlled withvaccination or treatment with antiparasitic drugs, the available vac-cines are live and have safety concerns, and many drugs have beenwithdrawn from the market due to safety or resistance problems[3]. Therefore, the development of new strategies, including vac-cination and chemotherapy, is highly required for controlling thedisease and reduce the hazards of infection.
Apicomplexan parasites are defined by their characteristic finestructures, including the apical complex, which consists of rhop-tries, micronemes, and dense granules. Spherical bodies are knownas unique organelles that present only in the babesial apical com-plex and are analogous to dense granules in other members of thegenera [4]. Proteins derived from those organelles are believed tohave critical functions, including host cell interaction and modi-fication for parasite invasion, growth, and egression [5,6]. Indeed,some of these proteins are released and incorporated into their hostcell environment during the invasion or post-invasion step, lead-ing to favorable conditions for the survival of the parasite [6]. Whilerhoptry and microneme proteins participate in the process of para-site attachment and invasion, spherical body proteins seem to playa role in the post-invasion step, aiding in parasite growth [5,6].
0166-6851/$ – see front matter © 2011 Elsevier B.V. All rights reserved.doi:10.1016/j.molbiopara.2011.02.006
Author's personal copy
M.A. Terkawi et al. / Molecular & Biochemical Parasitology 178 (2011) 40–45 41
Three proteins localized in the B. bovis spherical body organelleshave been previously identified and characterized. A gene encodingthe 77–80 kDa protein (BbSBP-1) was isolated from cDNA expres-sion library screened by naturally infected bovine sera. On theother hand, BbSBP-2 (225-kDa) and BbSBP-3 (135-kDa) were iso-lated using monoclonal antibodies raised against spherical bodies[7–9]. All of these proteins were found to localize to the cyto-plasmic face of the infected RBC membrane in a similar mannerto the dense granule proteins identified in Plasmodium spp. andToxoplasma gondii. Indeed, the dense granule proteins are releasedpost-invasion either into the cytoplasm of host cells or within aparasitophorous vacuole, aiding in stabilizing the parasite envi-ronment and facilitating the interaction between the parasites andthe erythrocyte membrane [5,6,10]. However, understanding thebiological functions of the proteins derived from these organelles,particularly those involved in B. bovis invasion, growth, and egres-sion, is undoubtedly necessary for the development of effectivechemotherapeutic agents or a protective vaccine.
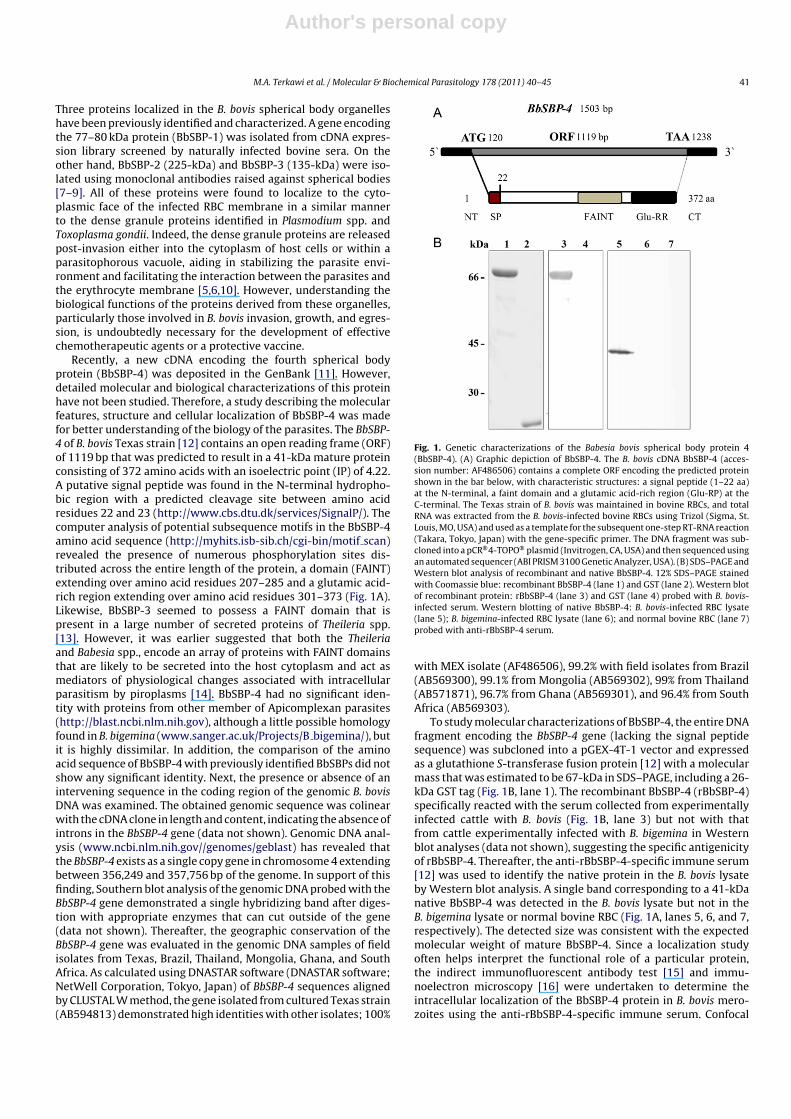
Recently, a new cDNA encoding the fourth spherical bodyprotein (BbSBP-4) was deposited in the GenBank [11]. However,detailed molecular and biological characterizations of this proteinhave not been studied. Therefore, a study describing the molecularfeatures, structure and cellular localization of BbSBP-4 was madefor better understanding of the biology of the parasites. The BbSBP-4 of B. bovis Texas strain [12] contains an open reading frame (ORF)of 1119 bp that was predicted to result in a 41-kDa mature proteinconsisting of 372 amino acids with an isoelectric point (IP) of 4.22.A putative signal peptide was found in the N-terminal hydropho-bic region with a predicted cleavage site between amino acidresidues 22 and 23 (http://www.cbs.dtu.dk/services/SignalP/). Thecomputer analysis of potential subsequence motifs in the BbSBP-4amino acid sequence (http://myhits.isb-sib.ch/cgi-bin/motif scan)revealed the presence of numerous phosphorylation sites dis-tributed across the entire length of the protein, a domain (FAINT)extending over amino acid residues 207–285 and a glutamic acid-rich region extending over amino acid residues 301–373 (Fig. 1A).Likewise, BbSBP-3 seemed to possess a FAINT domain that ispresent in a large number of secreted proteins of Theileria spp.[13]. However, it was earlier suggested that both the Theileriaand Babesia spp., encode an array of proteins with FAINT domainsthat are likely to be secreted into the host cytoplasm and act asmediators of physiological changes associated with intracellularparasitism by piroplasms [14]. BbSBP-4 had no significant iden-tity with proteins from other member of Apicomplexan parasites(http://blast.ncbi.nlm.nih.gov), although a little possible homologyfound in B. bigemina (www.sanger.ac.uk/Projects/B bigemina/), butit is highly dissimilar. In addition, the comparison of the aminoacid sequence of BbSBP-4 with previously identified BbSBPs did notshow any significant identity. Next, the presence or absence of anintervening sequence in the coding region of the genomic B. bovisDNA was examined. The obtained genomic sequence was colinearwith the cDNA clone in length and content, indicating the absence ofintrons in the BbSBP-4 gene (data not shown). Genomic DNA anal-ysis (www.ncbi.nlm.nih.gov//genomes/geblast) has revealed thatthe BbSBP-4 exists as a single copy gene in chromosome 4 extendingbetween 356,249 and 357,756 bp of the genome. In support of thisfinding, Southern blot analysis of the genomic DNA probed with theBbSBP-4 gene demonstrated a single hybridizing band after diges-tion with appropriate enzymes that can cut outside of the gene(data not shown). Thereafter, the geographic conservation of theBbSBP-4 gene was evaluated in the genomic DNA samples of fieldisolates from Texas, Brazil, Thailand, Mongolia, Ghana, and SouthAfrica. As calculated using DNASTAR software (DNASTAR software;NetWell Corporation, Tokyo, Japan) of BbSBP-4 sequences alignedby CLUSTAL W method, the gene isolated from cultured Texas strain(AB594813) demonstrated high identities with other isolates; 100%
Fig. 1. Genetic characterizations of the Babesia bovis spherical body protein 4(BbSBP-4). (A) Graphic depiction of BbSBP-4. The B. bovis cDNA BbSBP-4 (acces-sion number: AF486506) contains a complete ORF encoding the predicted proteinshown in the bar below, with characteristic structures: a signal peptide (1–22 aa)at the N-terminal, a faint domain and a glutamic acid-rich region (Glu-RP) at theC-terminal. The Texas strain of B. bovis was maintained in bovine RBCs, and totalRNA was extracted from the B. bovis-infected bovine RBCs using Trizol (Sigma, St.Louis, MO, USA) and used as a template for the subsequent one-step RT-RNA reaction(Takara, Tokyo, Japan) with the gene-specific primer. The DNA fragment was sub-cloned into a pCR®4-TOPO® plasmid (Invitrogen, CA, USA) and then sequenced usingan automated sequencer (ABI PRISM 3100 Genetic Analyzer, USA). (B) SDS–PAGE andWestern blot analysis of recombinant and native BbSBP-4. 12% SDS–PAGE stainedwith Coomassie blue: recombinant BbSBP-4 (lane 1) and GST (lane 2). Western blotof recombinant protein: rBbSBP-4 (lane 3) and GST (lane 4) probed with B. bovis-infected serum. Western blotting of native BbSBP-4: B. bovis-infected RBC lysate(lane 5); B. bigemina-infected RBC lysate (lane 6); and normal bovine RBC (lane 7)probed with anti-rBbSBP-4 serum.
with MEX isolate (AF486506), 99.2% with field isolates from Brazil(AB569300), 99.1% from Mongolia (AB569302), 99% from Thailand(AB571871), 96.7% from Ghana (AB569301), and 96.4% from SouthAfrica (AB569303).
To study molecular characterizations of BbSBP-4, the entire DNAfragment encoding the BbSBP-4 gene (lacking the signal peptidesequence) was subcloned into a pGEX-4T-1 vector and expressedas a glutathione S-transferase fusion protein [12] with a molecularmass that was estimated to be 67-kDa in SDS–PAGE, including a 26-kDa GST tag (Fig. 1B, lane 1). The recombinant BbSBP-4 (rBbSBP-4)specifically reacted with the serum collected from experimentallyinfected cattle with B. bovis (Fig. 1B, lane 3) but not with thatfrom cattle experimentally infected with B. bigemina in Westernblot analyses (data not shown), suggesting the specific antigenicityof rBbSBP-4. Thereafter, the anti-rBbSBP-4-specific immune serum[12] was used to identify the native protein in the B. bovis lysateby Western blot analysis. A single band corresponding to a 41-kDanative BbSBP-4 was detected in the B. bovis lysate but not in theB. bigemina lysate or normal bovine RBC (Fig. 1A, lanes 5, 6, and 7,respectively). The detected size was consistent with the expectedmolecular weight of mature BbSBP-4. Since a localization studyoften helps interpret the functional role of a particular protein,the indirect immunofluorescent antibody test [15] and immu-noelectron microscopy [16] were undertaken to determine theintracellular localization of the BbSBP-4 protein in B. bovis mero-zoites using the anti-rBbSBP-4-specific immune serum. Confocal
Author's personal copy
42 M.A. Terkawi et al. / Molecular & Biochemical Parasitology 178 (2011) 40–45
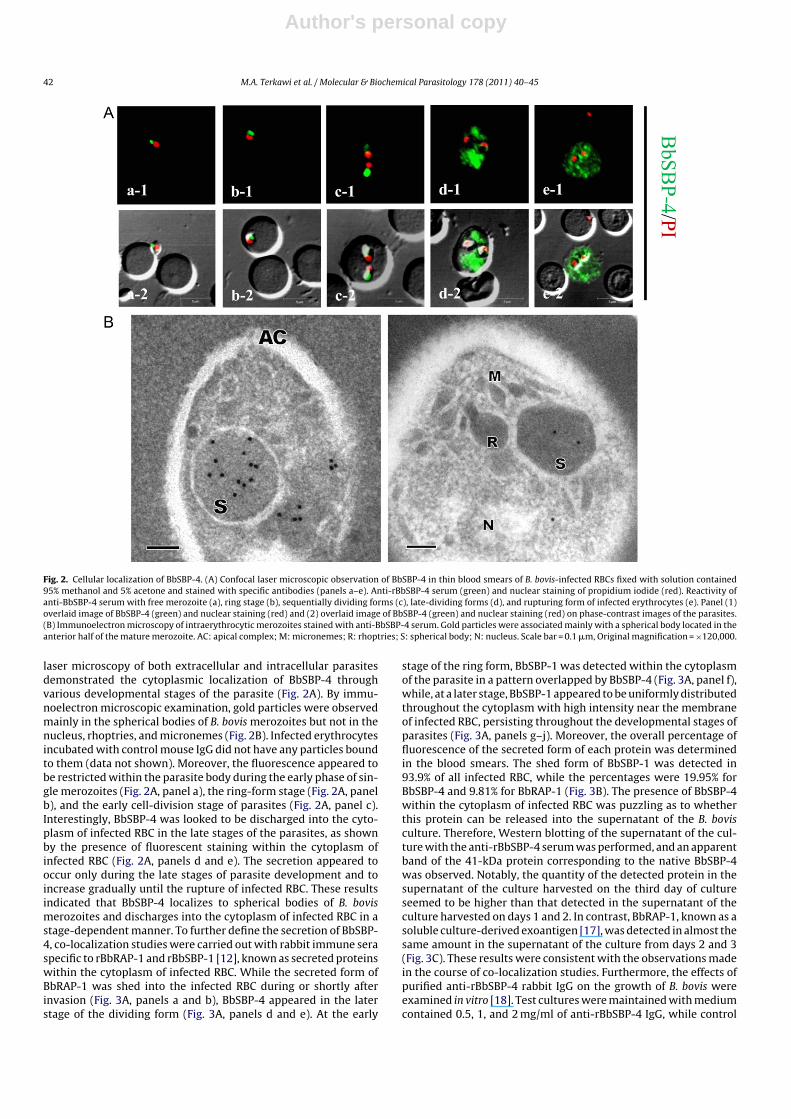
Fig. 2. Cellular localization of BbSBP-4. (A) Confocal laser microscopic observation of BbSBP-4 in thin blood smears of B. bovis-infected RBCs fixed with solution contained95% methanol and 5% acetone and stained with specific antibodies (panels a–e). Anti-rBbSBP-4 serum (green) and nuclear staining of propidium iodide (red). Reactivity ofanti-BbSBP-4 serum with free merozoite (a), ring stage (b), sequentially dividing forms (c), late-dividing forms (d), and rupturing form of infected erythrocytes (e). Panel (1)overlaid image of BbSBP-4 (green) and nuclear staining (red) and (2) overlaid image of BbSBP-4 (green) and nuclear staining (red) on phase-contrast images of the parasites.(B) Immunoelectron microscopy of intraerythrocytic merozoites stained with anti-BbSBP-4 serum. Gold particles were associated mainly with a spherical body located in theanterior half of the mature merozoite. AC: apical complex; M: micronemes; R: rhoptries; S: spherical body; N: nucleus. Scale bar = 0.1 �m, Original magnification = ×120,000.
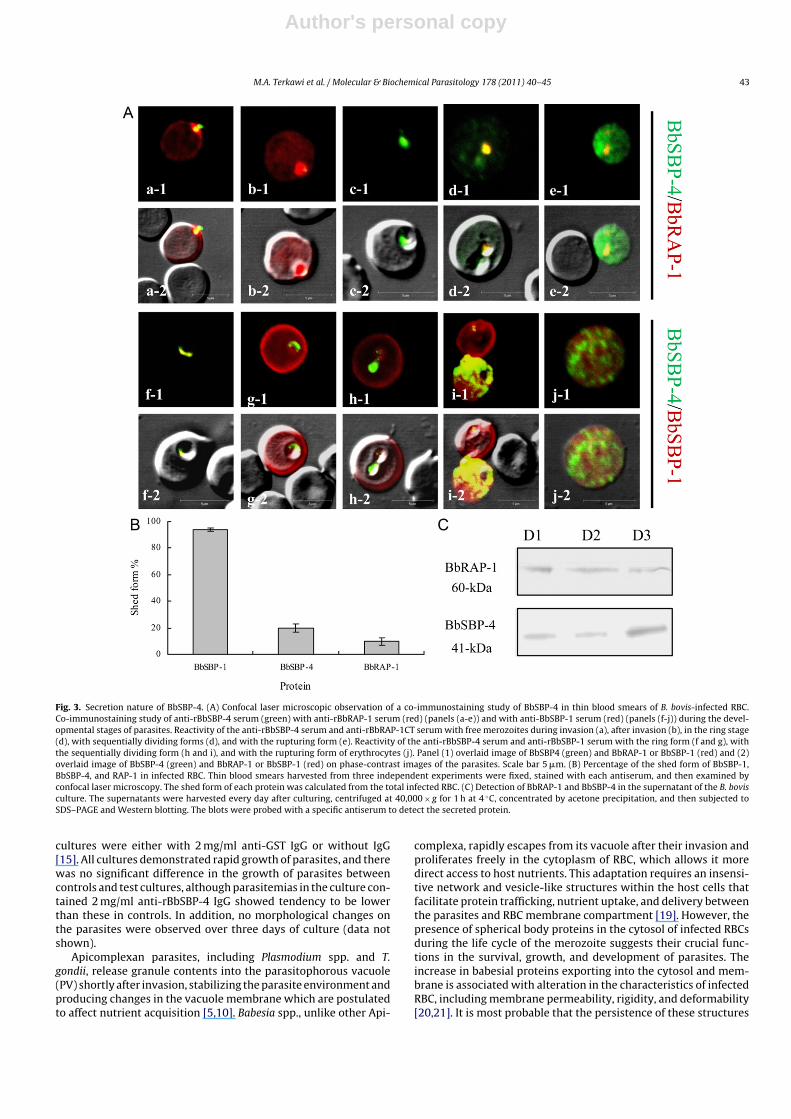
laser microscopy of both extracellular and intracellular parasitesdemonstrated the cytoplasmic localization of BbSBP-4 throughvarious developmental stages of the parasite (Fig. 2A). By immu-noelectron microscopic examination, gold particles were observedmainly in the spherical bodies of B. bovis merozoites but not in thenucleus, rhoptries, and micronemes (Fig. 2B). Infected erythrocytesincubated with control mouse IgG did not have any particles boundto them (data not shown). Moreover, the fluorescence appeared tobe restricted within the parasite body during the early phase of sin-gle merozoites (Fig. 2A, panel a), the ring-form stage (Fig. 2A, panelb), and the early cell-division stage of parasites (Fig. 2A, panel c).Interestingly, BbSBP-4 was looked to be discharged into the cyto-plasm of infected RBC in the late stages of the parasites, as shownby the presence of fluorescent staining within the cytoplasm ofinfected RBC (Fig. 2A, panels d and e). The secretion appeared tooccur only during the late stages of parasite development and toincrease gradually until the rupture of infected RBC. These resultsindicated that BbSBP-4 localizes to spherical bodies of B. bovismerozoites and discharges into the cytoplasm of infected RBC in astage-dependent manner. To further define the secretion of BbSBP-4, co-localization studies were carried out with rabbit immune seraspecific to rBbRAP-1 and rBbSBP-1 [12], known as secreted proteinswithin the cytoplasm of infected RBC. While the secreted form ofBbRAP-1 was shed into the infected RBC during or shortly afterinvasion (Fig. 3A, panels a and b), BbSBP-4 appeared in the laterstage of the dividing form (Fig. 3A, panels d and e). At the early
stage of the ring form, BbSBP-1 was detected within the cytoplasmof the parasite in a pattern overlapped by BbSBP-4 (Fig. 3A, panel f),while, at a later stage, BbSBP-1 appeared to be uniformly distributedthroughout the cytoplasm with high intensity near the membraneof infected RBC, persisting throughout the developmental stages ofparasites (Fig. 3A, panels g–j). Moreover, the overall percentage offluorescence of the secreted form of each protein was determinedin the blood smears. The shed form of BbSBP-1 was detected in93.9% of all infected RBC, while the percentages were 19.95% forBbSBP-4 and 9.81% for BbRAP-1 (Fig. 3B). The presence of BbSBP-4within the cytoplasm of infected RBC was puzzling as to whetherthis protein can be released into the supernatant of the B. bovisculture. Therefore, Western blotting of the supernatant of the cul-ture with the anti-rBbSBP-4 serum was performed, and an apparentband of the 41-kDa protein corresponding to the native BbSBP-4was observed. Notably, the quantity of the detected protein in thesupernatant of the culture harvested on the third day of cultureseemed to be higher than that detected in the supernatant of theculture harvested on days 1 and 2. In contrast, BbRAP-1, known as asoluble culture-derived exoantigen [17], was detected in almost thesame amount in the supernatant of the culture from days 2 and 3(Fig. 3C). These results were consistent with the observations madein the course of co-localization studies. Furthermore, the effects ofpurified anti-rBbSBP-4 rabbit IgG on the growth of B. bovis wereexamined in vitro [18]. Test cultures were maintained with mediumcontained 0.5, 1, and 2 mg/ml of anti-rBbSBP-4 IgG, while control
Author's personal copy
M.A. Terkawi et al. / Molecular & Biochemical Parasitology 178 (2011) 40–45 43
Fig. 3. Secretion nature of BbSBP-4. (A) Confocal laser microscopic observation of a co-immunostaining study of BbSBP-4 in thin blood smears of B. bovis-infected RBC.Co-immunostaining study of anti-rBbSBP-4 serum (green) with anti-rBbRAP-1 serum (red) (panels (a-e)) and with anti-BbSBP-1 serum (red) (panels (f-j)) during the devel-opmental stages of parasites. Reactivity of the anti-rBbSBP-4 serum and anti-rBbRAP-1CT serum with free merozoites during invasion (a), after invasion (b), in the ring stage(d), with sequentially dividing forms (d), and with the rupturing form (e). Reactivity of the anti-rBbSBP-4 serum and anti-rBbSBP-1 serum with the ring form (f and g), withthe sequentially dividing form (h and i), and with the rupturing form of erythrocytes (j). Panel (1) overlaid image of BbSBP4 (green) and BbRAP-1 or BbSBP-1 (red) and (2)overlaid image of BbSBP-4 (green) and BbRAP-1 or BbSBP-1 (red) on phase-contrast images of the parasites. Scale bar 5 �m. (B) Percentage of the shed form of BbSBP-1,BbSBP-4, and RAP-1 in infected RBC. Thin blood smears harvested from three independent experiments were fixed, stained with each antiserum, and then examined byconfocal laser microscopy. The shed form of each protein was calculated from the total infected RBC. (C) Detection of BbRAP-1 and BbSBP-4 in the supernatant of the B. bovisculture. The supernatants were harvested every day after culturing, centrifuged at 40,000 × g for 1 h at 4 ◦C, concentrated by acetone precipitation, and then subjected toSDS–PAGE and Western blotting. The blots were probed with a specific antiserum to detect the secreted protein.
cultures were either with 2 mg/ml anti-GST IgG or without IgG[15]. All cultures demonstrated rapid growth of parasites, and therewas no significant difference in the growth of parasites betweencontrols and test cultures, although parasitemias in the culture con-tained 2 mg/ml anti-rBbSBP-4 IgG showed tendency to be lowerthan these in controls. In addition, no morphological changes onthe parasites were observed over three days of culture (data notshown).
Apicomplexan parasites, including Plasmodium spp. and T.gondii, release granule contents into the parasitophorous vacuole(PV) shortly after invasion, stabilizing the parasite environment andproducing changes in the vacuole membrane which are postulatedto affect nutrient acquisition [5,10]. Babesia spp., unlike other Api-
complexa, rapidly escapes from its vacuole after their invasion andproliferates freely in the cytoplasm of RBC, which allows it moredirect access to host nutrients. This adaptation requires an insensi-tive network and vesicle-like structures within the host cells thatfacilitate protein trafficking, nutrient uptake, and delivery betweenthe parasites and RBC membrane compartment [19]. However, thepresence of spherical body proteins in the cytosol of infected RBCsduring the life cycle of the merozoite suggests their crucial func-tions in the survival, growth, and development of parasites. Theincrease in babesial proteins exporting into the cytosol and mem-brane is associated with alteration in the characteristics of infectedRBC, including membrane permeability, rigidity, and deformability[20,21]. It is most probable that the persistence of these structures

Author's personal copy
44 M.A. Terkawi et al. / Molecular & Biochemical Parasitology 178 (2011) 40–45
within the cytoskeleton of infected RBC maintains the mechanicalintegrity and stability of the host cells during their intracel-lular replication, preventing RBC lysis and consequent parasiterelease.
The dramatic changes in the infected RBCs caused by theparasite-derived proteins are not only in their biochemical prop-erties but also in the shape and morphology, as seen, for example,in the membrane protrusions (ridges) of B. bovis-infected RBC[19]. These structure alterations contribute to the sequestration ofinfected RBC in organ capillaries, where the variant erythrocyte sur-face antigen (VESA1) on the surface of the ridge structure serves asan RBCs ligand for cytoadhesion. This phenomenon is important forthe survival of the parasites, allowing the infected RBC to adhere tohost endothelial cells of microcapillaries and subsequently escapefrom destruction in the spleen [19,22]. Therefore, the potentialfunction of BbSBPs might include the stabilization of the parasiteenvironment, alteration of the membrane permeability requiredfor nutrient acquisition, and formation of the ridge structure. Dif-ferently from the previously reported BbSBPs, which were detectedwithin the cytoplasmic face of the infected RBC membrane shortlyafter parasite invasion [7–9] BbSBP-4 appeared to be dischargedinto the cytoplasm of RBC at a later stage and to increase graduallyuntil the rupture of RBCs. This characteristic, coupled with the highconservation of BbSBP-4 among different isolates and the absenceof homology in B. bigemina, suggests a potential role in parasitevirulence; probably, BbSBP-4 contributes to the formation of theridge structure. Ridge structures are completely absent from thesurface of B. bigemina-infected RBCs, a species closely related to B.bovis that causes a much less severe disease with lower mortal-ity [23]. Elucidating the precise function of BbSBPs as a modulatorof host RBCs is crucial for a better understanding of the virulenceand pathogenesis of parasites and beneficial for developing newtherapeutic strategies to combat this important disease. Furtherstudy including investigation of the possibility of BbSBP-4 to inter-act with the vesicles of RBCs and other member of SBPs might bevery necessary as step toward understanding of the function ofthis molecule. The failure of anti-BbSBP-4 IgG to inhibit the growthof the parasites suggests that BbSBP-4 is not involved in invasionof B. bovis into RBCs. The inability of purified IgG to neutralizethe free merozoite in vitro is most probably due to the inacces-sibility of the antibody to BbSBP-4 found within the cytoplasm offree merozoites at early stage of invasion. In contrast, antiserumto dense granule proteins of a variety of Apicomplexan parasitesinhibits the growth of the parasites in vitro [10,24]. These inhibitoryeffects appear to reflect interference of the antibodies with eventsoccurring during or shortly after invasion. Even in B. bovis, sev-eral studies have shown the inhibitory effects of antiserum raisedagainst molecules involved in the invasion process; such moleculesare usually derived from cell membrane, micronemes or rhoptries[6]. After the entry of the parasites inside the RBCs, the antibod-ies are less effective and the degeneration of parasites inside theerythrocyte is due to the interaction with the soluble mediatorsof cell-mediated immunity [25]. However, the secretion nature ofBbSBP-4 at the late stage of infection and its consequent releasefrom RBCs might make this molecule more accessible to a hostimmune cells, this attribute is very important for subunit vaccinecandidate and diagnostic antigen [26]. A recent study has shownthat BbSBP-4 is highly antigenic during the infection and has thebest diagnostic performance in detection antibody to B. bovis infec-tion in cattle as compared to other antigens derived from cellmembrane, micronemes or rhoptries [12].
In summary, we have identified and characterized a unique 41-kDa protein that has no significant identity to other Apicomplexanparasites and is highly conserved among field isolates from differ-ently geographic regions. The protein was localized to the sphericalbody of B. bovis and was discharged into the cytoplasm of infected
RBC at a later stage of parasite development. These findings areexpected to provide a basis for future work to exploit the functionof spherical body protein 4 in the development and survival of theseparasites.
Acknowledgments
This work was supported by a grant from the Global COE Pro-gram (J02) and a Grant-in-Aid for Scientific Research (A-22248035),both from the Ministry of Education, Culture, Sports, Science, andTechnology, Japan, and a program for the Promotion of BasicResearch Activities for Innovative Bioscience (PROBRAIN), anda cooperative research grant (22-23 joint-6) from the NationalResearch Center for Protozoan Diseases, Obihiro University of Agri-culture and Veterinary Medicine. The first author was supported bya research grant fellowship from the Japanese Society for the Pro-motion of Science (JSPS) for young scientists. The second, third andfourth authors were supported by Japan International CooperationAgency (JICA).
References
[1] Kuttler KL. Worldwide impact of babesiosis. In: Ristic M, editor. Babesiosis ofdomestic animals. Boca Raton, FL, USA: CRC Press, Inc.; 1988. p. 1–22.
[2] McCosker PJ. The global importance of babesiosis. In: Ristic M, Kreier JP, editors.Babesiosis. New York: Academic Press; 1981. p. 1–24.
[3] Bock R, Jackson L, De Vos A, Jorgensen W. Babesiosis of cattle. Parasitology2004;129:247–69.
[4] Potgieter FT, Els HJ. The fine structure of intra-erythrocytic stages of Babesiabovis merozoite surface-exposed epitopes. Infect Immun 1977;59:3340–2.
[5] Preiser P, Kaviratne M, Khan S, Bannister L, Jarra W. The apical organelles ofmalaria merozoites: host cell selection, invasion, host immunity and immuneevasion. Microbes Infect 2000;12:1461–77.
[6] Yokoyama N, Okamura M, Igarashi I. Erythrocyte invasion by Babesia para-sites: current advances in the elucidation of the molecular interactions betweenthe protozoan ligands and host receptors in the invasion stage. Vet Parasitol2006;138:22–32.
[7] Ruef BJ, Dowling SC, Conley PG, et al. A unique Babesia bovis spherical bodyprotein is conserved among geographic isolates and localizes to the infectederythrocyte membrane. Mol Biochem Parasitol 2000;105:1–12.
[8] Jasmer DP, Reduker DW, Perryman LE, McGuire TC. A Babesia bovis 225-kilodalton protein located on the cytoplasmic side of the erythrocytemembrane has sequence similarity with a region of glycogen phosphorylase.Mol Biochem Parasitol 1992;52(2):263–9.
[9] Dowling SC, Perryman LE, Jasmer DP. A Babesia bovis 225-kilodalton sphericalbody protein: localization to the cytoplasmic face of infected erythrocytes aftermerozoite invasion. Infect Immun 1996;64:2618–26.
[10] Cesbron-Delauw MF. Dense-granule organelles of Toxoplasma gondii: their rolein the host–parasite relationship. Parasitol Today 1994;10:293–6.
[11] De Vries E, Corton C, Harris B, Cornelissen AW, Berriman M. Expressed sequencetag (EST) analysis of the erythrocytic stages of Babesia bovis. Vet Parasitol2006;138:61–74.
[12] Terkawi MA, Huyen NX, Wibowo PE, et al. Spherical body protein 4 is a newserological antigen for the global detection of Babesia bovis infection in cattle.Clin Vaccine Immunol 2011;18(2):337–42.
[13] Pain A, Renauld H, Berriman M, et al. Genome of the host-cell trans-forming parasite Theileria annulata compared with T. parva. Science2005;309(5731):131–3.
[14] Anantharaman V, Iyer LM, Balaji S, Aravind L. Adhesion molecules and othersecreted host-interaction determinants in Apicomplexa: insights from com-parative genomics. Int Rev Cytol 2007;262:1–74.
[15] Terkawi MA, Aboge G, Jia H, et al. Molecular and immunological characteri-zation of Babesia gibsoni and Babesia microti heat shock protein-70. ParasiteImmunol 2009;31(6):328–40.
[16] Aikawa M. Localization of protective malarial antigens by immunoelectronmicroscopy. Mem Inst Oswaldo Cruz Rio J 1986;81:159–66.
[17] Johnson WC, Perryman LE, Goff WL. Babesia bovis: Identification of immun-odominant merozoite surface proteins in soluble culture-derived exoantigen.Parasitol Res 1997;83:776–80.
[18] Yokoyama N, Suthisak B, Hirata H, et al. Cellular localization of Babesia bovismerozoite rhoptry-associated protein 1 and its erythrocyte-binding activity.Infect Immun 2002;70:5822–6.
[19] Gohil S, Kats LM, Sturm A, Cooke BM. Recent insights into alteration of red bloodcells by Babesia bovis: Moovin’ forward. Trends Parasitol 2010;26(12):591–9.
[20] Gero AM, Wood AM, Hogue DL, Upston JM. Effect of diamide on nucleosideand glucose transport in Plasmodium falciparum- and Babesia bovis-infectederythrocytes. Mol Biochem Parasitol 1991;44:195–6.
[21] Upston JM, Gero AM. Increased glucose permeability in Babesia bovis-infectederythrocytes. Int J Parasitol 1990;20:69–76.

Author's personal copy
M.A. Terkawi et al. / Molecular & Biochemical Parasitology 178 (2011) 40–45 45
[22] O’Connor RM, Long JA, Allred DR. Cytoadherence of Babesia bovis-infected ery-throcytes to bovine brain capillary endothelial cells provides an in vitro modelfor sequestration. Infect Immun 1999;67(8):3921–8.
[23] Hutchings CL, Li A, Fernandez KM, et al. New insights into the altered adhesiveand mechanical properties of red blood cells parasitized by Babesia bovis. MolMicrobiol 2007;65(4):1092–5.
[24] Augustine PC, Jenkins MC, Dubey JP. Effect of polyclonal antisera developedagainst dense granule-associated Neospora caninum proteins on cell invasion
and development in vitro by N. caninum tachyzoites. Parasitology 1999;119(Pt5):441–5.
[25] Homer MJ, Aguilar-Delfin SR, Telford PJ, Krause, Persing DH. Babesiosis. ClinMicrobiol 2000;13(3):451–69.
[26] Homer MJ, Lodes MJ, Reynolds LD, et al. Identification and characteriza-tion of putative secreted antigens from Babesia microti. J Clin Microbiol2003;41(2):723–9.
Copyright © 2022 FDOKUMEN




















