
Intravenous total parenteral nutrition elevates oxidant production in critically ill patients
Upload
independentCategory
view
3download
0
EFFICACY OF ORAL AND PARENTERAL ROUTES OF
MYCOBACTERIUM BOVIS BACILLE CALMETTE-GUERIN
VACCINATION AGAINST EXPERIMENTAL BOVINE TUBERCULOSIS
IN WHITE-TAILED DEER (ODOCOILEUS VIRGINIANUS):
A FEASIBILITY STUDY
P. Nol,1,5,6 M. V. Palmer,2 W. R. Waters,2 F. E. Aldwell,3 B. M. Buddle,4 J. M. Triantis,5
L. M. Linke,5 G. E. Phillips,1 T. C. Thacker,2 J. C. Rhyan,1 M. R. Dunbar,1 and M. D. Salman5
1 USDA/APHIS National Wildlife Research Center, Fort Collins, Colorado 80521, USA2 USDA/ARS National Animal Disease Center, Ames, Iowa 50010, USA3 Department of Microbiology and Immunology, University of Otago, P.O. Box 56, Dunedin, New Zealand4 AgResearch, Hopkirk Research Institute, Private Bag 1108, Palmerston North 4442, New Zealand5 Department of Clinical Sciences, Colorado State University, Fort Collins, Colorado 80523, USA6 Corresponding author (email: [email protected])
ABSTRACT: We investigated the efficacy of oral and parenteral Mycobacterium bovis bacilleCalmette-Guerin Danish strain 1331 (BCG) in its ability to protect white-tailed deer (Odocoileusvirginianus) against disease caused by M. bovis infection. Twenty-two white-tailed deer weredivided into four groups. One group (n55) received 109 colony-forming units (cfu) BCG via alipid-formulated oral bait; one group (n55) received 109 cfu BCG in culture directly to theoropharynx, one group (n56) was vaccinated with 106 cfu BCG subcutaneously, and one groupserved as a control and received culture media directly to the oropharynx (n56). All animals werechallenged 3 mo after vaccination. Five months postchallenge the animals were examined forlesions. Results indicate that both oral forms of BCG and parenterally administerd BCG offeredsignificant protection against M. bovis challenge as compared to controls. This study suggests thatoral BCG vaccination may be a feasible means of controlling bovine tuberculosis in wild white-tailed deer populations.
Key words: BCG, bovine tuberculosis, Mycobacterium bovis, oral vaccination, vaccine,white-tailed deer.
INTRODUCTION
Bovine tuberculosis(BTb), caused byMycobacterium bovis, poses a seriouscontinual threat to the health and eco-nomic well-being of both livestock andhumans worldwide. In the developingworld, one third of the population lackseffective agricultural and food safetyprograms, leaving them at substantial riskfor zoonotic infection by M. bovis (Nelson,1999). In addition, wildlife reservoirs ofBTb are believed to play a very importantrole in the epidemiology of this disease inmany countries. In the United States, afree-ranging white-tailed deer (Odocoileusvirginianus) population located in north-eastern Michigan serves as such a reser-voir (Schmitt et al., 1997; Fitzgerald et al.,2000; Palmer et al., 1999; 2000; Waters etal., 2004b).
In 1994 M. bovis was isolated from a
hunter-killed white-tailed deer buck fromnorthern Michigan. Subsequent surveil-lance of the free-ranging herd verified anepidemic presence of the disease. Severalother wild mammalian species in the areahave also been found to be infected orexposed to M. bovis including cervids andcarnivores (Schmitt et al., 1997; Brunning-Fann et al., 2001). The self-sustainingpresence of bovine tuberculosis in thiswild deer population, and possibly otherwild populations, poses a significant chal-lenge to Michigan’s efforts to reestablishits BTb-free status in domestic livestock.The implementation of an oral vaccinationprogram, which would effectively reducedisease and shedding of M. bovis by deerand other affected species, would signifi-cantly aid efforts in eradicating BTb fromMichigan.
We assessed the efficacy of orallyadministered M. bovis bacille Calmette-
Journal of Wildlife Diseases, 44(2), 2008, pp. 247–259# Wildlife Disease Association 2008
247

Guerin Danish strain 1331 (BCG) inwhite-tailed deer against infection withvirulent M. bovis. Mycobacterium bovisbacille Calmette-Guerin is currently theonly vaccine licensed for use in humansagainst tuberculosis (Mycobacterium tu-berculosis), and there are currently noaccepted vaccination programs available tolivestock. In fact, BCG remains the goldstandard against which all experimentaltuberculosis vaccines are compared, anddespite intense efforts to develop a moreeffective vaccine, to date there are no newcommercially available tuberculosis vac-cines. Our study compared two types oforal BCG vaccination, as well as parenter-al BCG vaccination, in their ability toprotect white-tailed deer from diseasecaused by experimental infection of M.bovis.
MATERIALS AND METHODS
Animals
Thirty yearling female white-tailed deer wereobtained from four BTb-free deer farmsthroughout the state of Iowa, USA, and wereshipped to the USDA/ARS National AnimalDisease Center (NADC) in Ames, Iowa, USA,in January 2005 and housed in an outdoor penfacility. Prior to shipment the animals weresocialized as a group on one of the supplyingfarms for 3 wk. After delivery to NADC,animals were acclimated for 4 wk prior to startof the study. At time of challenge, deer weretransferred to a Biosafety level (BL)-3 animalbuilding. All deer were housed and cared foraccording to the Association for Assessmentand Accreditation for Laboratory Animal CareInternational and institutional guidelines. Thisstudy was restricted to using a maximum of 30animals due to space limitations.
Mycobacterium bovis BCG and M. bovischallenge strains
Mycobacterium bovis BCG Danish strain1331 in culture and in lipid-formulated pelletswere prepared at the University of Otago,Immune Solutions Ltd., as described inAldwell et al. (2003a, b). Vaccine doses weredetermined using standard enumeration tech-niques by serial dilution plate counting onMiddlebrook’s 7H11 media (Becton-Dickin-son, Cockysville, Maryland, USA). Mycobacte-rium bovis (strain 9839 Ames designation) was
grown to midlog phase on Middlebrook 7H9broth supplemented with 10% oleic acid-albumin-dextrose complex (OADC) (Difco,Detroit, Michigan, USA) plus 0.05% Tween80. Bacilli were harvested from culture mediaby pelleting the cells by centrifugation at 2,0003 G, washing twice with 1 ml of phosphate-buffered saline solution and diluting to theappropriate cell density in 2 ml of PBS. Thechallenge dose was determined as describedabove for vaccine doses.
Vaccination and challenge of animals
White-tailed deer were orally vaccinated(n58) with approximately 13109 cfu BCG vialipid-formulated bait (oral bait group). Thesedeer were each offered a 1 g, unflavored lipidpellet inside a piece of fresh apple and allowedto voluntarily eat the bait and apple so thatthey chewed the material before swallowing.Another group of eight deer were orallyvaccinated with BCG in liquid medium (oralliquid group). A 1-ml preparation of approx-imately 1.93108 cfu BCG in 7H9 broth wasadministered via 3-ml syringe and a 10 French25-cm sterile urinary catheter (Self-CathH,Mentor, Minneapolis, Minnesota, USA). Carewas taken to contact the back of the mouthwith the inoculum in order not to bypass thepharyngeal lymphoid tissue. Seven deer werevaccinated subcutaneously in the right caudalcervical area with a 1-ml preparation ofapproximately 3.43106 cfu BCG (parenteralgroup). Seven deer received 1 ml 7H9 brothorally directly into the back of the mouth viasyringe and catheter and served as the controlgroup. Control and vaccinated deer werehoused together in an outdoor paddock untilchallenge in an indoor BL3 facility.
At 12 wk post-vaccination, all animals wereanesthetized with an intramuscular (IM)injection of ketamine hydrochloride (6 mg/kg) and xylazine (2 mg/kg), moved into a BL3facility also at NADC, and immediatelychallenged with a total of 228 cfu M. bovisstrain 9839 directly into the palatine tonsillarcrypts (114 cfu/tonsil) via pipette, the dosebeing divided between the two tonsils. Oralliquid and parenteral doses of BCG andchallenge dose of M. bovis were retrospec-tively determined via culture as describedabove. The oral bait dose, as estimated fromculture of bacilli prior to lipid formulation, wasapproximately 9.23108 cfu.
Mononuclear cell culture
Whole blood was collected by jugularvenipuncture. Peripheral blood mononuclearcells (PBMCs) were obtained from whole
248 JOURNAL OF WILDLIFE DISEASES, VOL. 44, NO. 2, APRIL 2008

blood collected in 60-cc syringes containing5 ml 2X acid citrate dextrose. PBMCs wereisolated from buffy coat fractions using aFicoll-Hypaque (Histopaque-1083, Sigma, St.Louis, Missouri, USA) technique followed by alyse and restore step as described (Waters etal., 2004a). The PBMCs were resuspended insupplemented RPMI 1640 (Sigma) and count-ed. The RPMI 1640 was supplemented with25 mM N-(2-hydroxyethyl) piperazine-N9-(2-ethanesulfonic acid) (HEPES) buffer,100 units/ml penicillin, 0.1 mg/ml streptomy-cin, 50 mM 2-mercaptoethanol (Sigma), and10% (v/v) fetal bovine sera (FBS; NationalVeterinary Services Laboratory [NVSL],Ames, Iowa, USA).
Lymphocyte blastogenesis
Lymphocyte blastogenesis assays were per-formed on days 0, 28, and 83 postvaccination.Mononuclear cells (53105 cells) were addedto wells of 96-well round-bottom microtiterplates (Falcon, Becton-Dickinson, LincolnPark, New Jersey, USA). Wells containedmedium plus 10 mg/ml M. bovis purifiedprotein derivative (PPDb, Prionics AG,Schlieren, Switzerland), 10 mg/ml Mycobacte-rium avium PPD (PPDa, Prionics AG), 10 mg/ml rESAT6:CFP10 (E:C, Waters et al.,2004a), 1 mg/ml pokeweed mitogen (PWM,Sigma), or medium alone (no stimulation).Each antigen treatment was run in triplicate.Cells were incubated for 6 days at 37 C in 5%CO2 in air, at which time each well waspulsed with 0.5 mCi methyl-[3H]thymidine(Amersham Life Science, Arlington Heights,Illinois, USA) in 50 ml RPMI. Then PBMCswere incubated for another 24 hr andharvested onto fiber filters with a 96-wellplate harvester (EG&G Wallace, Gaithers-burg, Maryland, USA), and the radioactivitylevels measured by liquid scintillation count-ing. Data are presented as stimulation indices(SI) where
SI~Mean counts=min (antigen� or mitogen� stimulated cultures)
Mean counts=min (nonstimulated cultures):
Interferon-c enzyme-linked immunosorbent assay
Interferon-c (IFN- c) responses to myco-bacterial antigens were measured at 277, 0,28, 49, and 83 days post-vaccination, and at 13,52, 88, and 119 days post-challenge (98, 137,173, and 204 days post-vaccination, respec-tively) using a commercial enzyme-linkedimmunosorbent assay (ELISA) kit (Cervi-gamTM, Prionics AG) modified as described
in Palmer et al. (2004b) to determine inter-feron production. Briefly, heparinized wholeblood was collected by jugular venipuncture,of which 1.5 ml was added to 24-well tissueculture plates (Falcon, Becton-Dickinson,Lincoln Park, New Jersey, USA). Treatmentsadministered included PBS (no stimulation),20 mg/ml PPDb, 20 mg/ml PPDa, 20 mg/mlPWM, or 10 mg/ml E:C. Plates were incubatedfor 48 hr at 37 C in a humidified chamber with5% CO2. Samples were centrifuged at 400 3G, and the plasma supernatants harvested andstored at 280 C until analysis using theCervigamTM ELISA according to the manu-facturer’s instructions. Optical density wasmeasured at 450 nm using an automatedELISA microplate reader (Molecular Devices,Menlo Park, California, USA). Concentrations(ng/ml) of IFN-c in test samples weredetermined by comparing absorbances of testsamples with absorbances of standards withina linear curve fit (Instat 2.0, GraphPadSoftware Inc., San Diego, California, USA).The IFN-c data (ng/ml) used for statisticalanalysis were derived from calculating thedifferences between response to antigen andresponse to no stimulation (PBS) (antigenstimulation (ng/ml)2no stimulation (ng/ml).
Post-mortem examination
Twenty weeks post-challenge, the deer wereanesthetized with ketamine hydrochloride(6 mg/kg) and xylazine (2 mg/kg) and eutha-nized by intravenous administration of sodiumpentobarbital (SleepawayH, Fort Dodge, Iowa,USA). Tissues collected for gross pathology,histopathology, culture, and polymerase chainreaction (PCR) were palatine tonsils, mandib-ular lymph nodes (LNs), parotid LNs, medialretropharyngeal LNs, right prescapular LN,tracheobronchial LN, mediastinal LNs, rightcranial lung lobe, liver, hepatic LN, andmesenteric LNs. In the case of bilateral LNs,excepting the prescapular LNs, portions ofboth right and left LNs were collected for bothculture and histopathology. All lung lobeswere carefully examined for lesions, and eachwas weighed individually for comparison withthose of the other deer in the study. A scoringsystem based on number and extent of grosslesions was applied to all LNs collected and alllung lobes for statistical comparison (adaptedfrom Vordermeier et al., 2002). For the lunglobes scoring was as follows: a score of ‘‘nolesions’’ meant there were no gross lesionsdetected. A score of ‘‘mild’’ representedtissues that contained any lesions fewer thanor equal to five gross lesions of less than10 mm in diameter. A score of ‘‘severe’’ was
NOL ET AL.—EFFICACY OF ORAL AND PARENTERAL BCG IN WHITE-TAILED DEER 249

assigned to tissues with greater than or equalto six gross lesions of less than 10 mm indiameter or any lesions larger than 10 mm indiameter. For the LNs scoring was as follows:a score of ‘‘no lesions’’ meant there were nogross lesions detected. A score of ‘‘mild’’represented tissues that contained one smallfocus 1–2 mm in diameter. A score of ‘‘severe’’was assigned to tissues with several small focior extensive necrosis. All tissues for cultureand PCR were stored in whirlpak bags at280 C until testing. Tissues for histopathologywere fixed in neutral-buffered 10% formalinand processed by routine paraffin embedmenttechniques. Sections were cut 5-mm thick,stained with hematoxylin and eosin, andexamined under light microscopy. Sectionscontaining microscopic lesions compatiblewith tuberculosis were subsequently stainedwith Ziehl-Neelsen for identification of acid-fast organisms. A scoring system was applied toall tissues collected for histopathology basedon the number and extent of microscopicgranulomatous lesions and was conducted asfollows: a score of ‘‘no lesions’’ meant therewere no granulomatous lesions detected; ascore of ‘‘mild’’ represented tissues that hadgranulomas containing macrophages, giantcells, lymphocytes, and neutrophils, but hadincomplete encapsulation and minimal to nonecrosis present; a score of ‘‘severe’’ was givento any granulomas that displayed encapsula-tion, caseous necrosis, and mineralization.
Tissue culture and polymerase chain reaction
We selected right cranial lung lobe, medi-astinal LN, and medial retropharyngeal LN forquantitative culture. Tonsil and mesentericLNs were selected for general culture. In aClass 3 biosafety cabinet, a small piece of eachtissue was removed and stored at 280 C forfresh tissue PCR analysis. All tissues were thenhomogenized using a blender (Oster, Shelton,Connecticut, USA) with 50 ml (LN) or 100 ml(lung) phenol red broth (NVSL). A 7.5-mlaliquot was removed and the remainingsample stored at 280 C for PCR analysis.The aliquot was then subjected to NaOHdecontamination as follows. Five milliliters of0.5 M NaOH was added to the 7.5-ml aliquotand allowed to incubate for 10 min. Then 10 NHCl was added dropwise until a yellow colorwas obtained followed by a dropwise additionof 1.0 N NaOH until a dusty rose color wasachieved. Samples for general culture werecentrifuged at 3,000 3 G for 15 min. Thesupernatant was discarded, and the remainingsediment was used to inoculate three tubes of7H10 media and three tubes of 7H11 media
(NVSL). Samples for quantitative culture werevortexed at high speed for 5 sec and serial-ly diluted at 1:10 dilutions in PBS out to 1026,vortexing for 5 sec between each dilution. Onehundred microliters of each dilution was usedto inoculate 7H11 agar plates (NVSL). Theundiluted sample was used to inoculate onetube of 7H10 media. All cultures were placedin an incubator at 37 C. Plates were kept inplastic bags to prevent the agar from dryingout. Samples were checked every 30 days, andfinal counts/assessments were made at60 days.
Samples that produced growth on 7H10 and7H11 media and considered suspect forMycobacterium spp. were picked and storedin 1 ml phosphate buffered saline at 280 Cfor follow-up PCR to confirm or rule out thepresence of M. bovis. Extraction of DNA fromcell culture samples was performed using theBioRad InstaGeneTMmatrix (Bio-Rad Labora-tories, Hercules, California, USA) according tothe manufacturer’s instructions with the fol-lowing modifications: Briefly, each sample wasvortexed, after which 200 ml of cell homoge-nized sample was transferred into a beadbeater tube (Sarstedt Inc., Newton, NorthCarolina, USA) containing 0.5 g of 0.5-mmzirconium/silica beads (Biospec Products Inc.,Bartlesville, Oklahoma, USA). Samples werebead beated using the MBB-8 Mini BeadBeater (Biospec Products) for 2 cycles(4,500 rpm) of 10 sec. After a quick spin,50 ml of bead-beated cells were transferred toa new tube, to which 200 ml of InstageneChelex matrix was added. Next, 14.2 ml ofproteinase K (Amresco Inc., Solon, Ohio,USA; 20 mg/ml) was added to each sample,which, after vortexing, was then incubated at60 C for 30 min. The proteinase K wasinactivated by heating the samples to 100 Cfor 10 min. Finally, the samples were centri-fuged at 11,000 3 G for 10 min, and theresulting supernatants containing DNA weresaved. For the purposes of PCR, dilutionswere made of the DNA samples ranging from1:10 to 1:10,000 depending on extent ofturbidity. For every extraction, one tubecontaining 200 ml of PBS was included halfwaybetween the total sample size and at the end ofthe extraction samples to serve as appropriatenegative extraction controls.
The samples were then analyzed for M.tuberculosis complex DNA using a PCRtargeting the IS6110 regions of M. tuberculosiscomplex (IS6110 PCR). The IS6110 PCR wasperformed using a PTC-100 MJ Thermalycler(MJ Research Inc., Waltham, Massachusetts,USA) for 50 cycles. The primers used in theIS6110 PCR were as follows: forward primer
250 JOURNAL OF WILDLIFE DISEASES, VOL. 44, NO. 2, APRIL 2008

(59CTCGTCCAGCGCCGCTTCGG39) andreverse primer (39CCTGCGAGCGTAGGCGTCGG59) (Operon Biotechnologies, Inc.,Huntsville, Alabama, USA). Each IS6110PCRreaction (25 mL) occurred in a 0.65 mL PCRtube (Sorenson BioScience Inc., Salt LakeCity, Utah, USA) and consisted of 1X Gen-eAmp Buffer II, 1.5 mM MgCl (AppliedBiosystems, Foster City, California, USA),2.5 mM of each deoxynucleoside triphosphate(Roche Applied Science, Indianapolis, Indi-ana, USA), 0.4 mM of each primer, 1.875 unitsof AmpliTaq Gold Polymerase (Applied Bio-systems), 10 mL template, and 6.625 mL PCR-grade water (Hyclone, Logan, Utah, USA)covered with a 30 mL layer of Chill-Out waxbuffer (Bio-Rad Laboratories). An amplifica-tion profile of 1) 94 C for 10 min, 2) 94 C for45 sec, 3) slope 222 degrees at 1 degree C/1 sec, 4) 72 C for 2.25 min, 5) slope +22 de-grees at 1.5 degrees C/1 sec, 6) repeat 2–5 for49 cycles, and 7) 72 C for 2.25 min wasfollowed by a final extension of 72 C for10 min.
A positive control (purified M. bovis DNA at0.5–5 fg/mL) and a negative control (water)were included in every experiment. Followingthe amplification protocol, 10 ml of the ampli-fication reactions were size fractionatedthrough 2.5% agarose (Amresco, Inc.) gels in1X TAE buffer (40 mM Tris acetate, 1 mMEDTA) (Amresco, Inc.). Gels were stained in0.001 mg/ml ethidium bromide (Amresco,Inc.) for 15 min, and products were thenvisualized using the BioRad Gel Doc EQSystem (Bio-Rad Laboratories) UV transillu-minator. Product size was 322 base pairs.
Medial retropharyngeal LN, mediastinalLN, and right cranial lung lobe–derived freshand homogenized tissues were also directlytested for M. tuberculosis complex by PCR.Frozen homogenates were placed at 4 Covernight to thaw. In a class 2 biosafetycabinet, approximately 1.3 ml homogenatewas removed from the bottom of the 50-mlpolypropylene centrifuge tube and transferredto a 2-ml polypropylene centrifuge tube.Samples were centrifuged for 10 min at11,000 3 G. Saving the resultant phenol brothsupernatant in a new tube, 100 mg of thetissue was weighed out and placed into a 2-mlbead beater tube containing 2.5 g of 2.5 mmand 0.5 g of 0.5 mm zirconia/silica beads, towhich 100 ml of the supernatant was returned.For every nine samples, a bead beater tubewas filled with 200 ml PBS to serve as negativecontrol. All tubes were quick spun and placedin a heat block for 10 min at 100 C. Thesamples were allowed to cool for 10 min.Tubes were then bead beated for three cycles
(4,500 rpm) of 30 sec. Samples were cooled inan ice block in between cycles to avoidoverheating. At this point samples wereconsidered safe to handle outside the biosafetycabinet. DNA extraction was then accom-plished using the Fermentas Genomic DNAPurification KitH according to manufacturer’sinstructions (Fermentas Life Sciences, Han-over, Maryland, USA). Following the manu-facturer’s recommended DNA ethanol(ETOH)/salt precipitation, one Breath-Ea-sierH tube membrane (ISC Bioexpress, Kays-ville, Utah, USA) was placed over each opentube to avoid cross-contamination and allowfor proper ETOH evaporation while incubat-ing at 65 C under a still-air hood. DNAsamples of the homogenized tissue wererehydrated in 100 ml of TE buffer (10 mMTris, 1 mM EDTA) (Amresco Inc.) anddiluted to 1:60 and 1:80 in PCR-grade waterfor PCR.
In a class 2 biosafety cabinet, 100 mg offresh frozen tissue was weighed out from eachsample for DNA extraction. Tissues wereminced and placed in bead beater tubes,containing 2.5 g of 2.5 mm and 0.5 g of0.5 mm zirconia/silica beads, to which 100 mlof TE buffer was added. Tissues underwentbead beating homogenization with two pulsesof 30 sec at 4,500 rpm. Samples were cooledin an ice block in between cycles to avoidoverheating. The Epicentre MasterPureTM
DNA Purification Kit was used according tothe manufacturer’s instructions with somemodifications (Epicentre Biotechnologies, Ma-dison, Wisconsin, USA). Briefly, samples werequick spun, and 500 ml of T and C LysisSolution with 0.675 mg/ml proteinase K wasadded to each sample. Samples were incubat-ed for 60 min at 56 C and again for 10 min at100 C to inactivate the proteinase K andrender any remaining M. bovis nonviable.After the samples cooled, 250 ml of MPCProtein Precipitation Reagent was added, andsamples were centrifuged at 11,000 3 G for10 min at 4 C. The supernatant was thentreated with 750 ml of 100% isopropanol andcentrifuged for 15 min at 11,000 3 G at 4 C.The isopropanol was discarded, and the pelletwas washed two times with 75% ETOH. Anyremaining ETOH was allowed to evaporate byplacing a Breath-EasierH tube membrane overeach open tube as described above. DNAsamples of the fresh tissue were rehydrated in100 ml of TE buffer and diluted to 1:60 and1:80 for IS6110 PCR.
DNA from fresh and homogenized tissuesunderwent PCR analysis as described above.Any DNA samples deemed positive weresubjected to two repeat PCR analyses, and
NOL ET AL.—EFFICACY OF ORAL AND PARENTERAL BCG IN WHITE-TAILED DEER 251

corresponding tissue samples were subjectedto one additional extraction and PCR analysisto confirm presence of M. bovis. Unlikeculture, these assays do not reflect the viabilityof the mycobacteria, but serve as a sensitivemethod to test for evidence of infection atsome time during the experiment.
Statistical analysis
Fisher’s Exact Test (433; one-sided) wasused to compare all groups based on numberof animals with gross lesions (none, mild, andsevere) and microscopic lesions (none, mild,and severe) (Samuels and Witmer, 1999) (ProcFREQ; SAS 9.1, SAS Institute, Cary, NorthCarolina, USA). If an association existed,Fisher’s Exact Test (232; one-sided) wasagain used to compare the number of animalswith lesions within each vaccine group to thenumber of animals with lesions within thecontrol group or to each other based on nolesions versus lesions (mild and severe com-bined). Culture and PCR results were com-pared in the same manner (Fisher’s ExactTest; 232; one-sided) in order to determinedifferences in number of culture/PCR positiveanimals among groups. As this was a resource-intensive pilot study that was limited to a smallnumber of animals, statistical priority wasplaced on minimizing the chance of type 2error in order to detect differences if theywere biologically valid. It was determined thatusing 8 animals per group, at an alpha level of0.1, the test would have a power of 0.8 indetecting a difference when the proportion ofanimals without lesions in the vaccine groupsexceeded the proportion of animals withoutlesions in the control group by 0.5 (SASInstitute, 2006). Therefore, differences deter-mined by the one-sided Fisher’s Exact Test tohave P #0.1 were considered significant.
We modeled IFN-c and lymphocyte blasto-genesis response variables as functions oftreatment group, time point, and group3timepoint interaction. We coupled this fixed-effects model structure with three alternativecovariance structures to account for randomeffects of individual deer, effects of repeatedmeasures within individuals, and unequalvariances among treatment groups using theGLIMMIX procedure of SAS (Littell et al.,2006; SAS Institute 2006). The most generalcovariance structure estimated separate vari-ance components and autoregressive correla-tion coefficients by treatment group. Modelswith reduced covariance structures includedone model estimating a common variancecomponent (random effect of individuals)and a common autoregressive correlation
among treatment groups, and one modelestimating a common variance component(i.e., no treatment-group covariance effect,no repeated measures effect). We used aninformation theoretic approach to select themost parsimonious form of covariance struc-ture for each response variable (Burnham andAnderson, 2002; Littell et al., 2006). For theselected models, we considered fixed effects assignificant for P#0.1.
RESULTS
Animals
Because of illnesses and injuries, eightanimals were removed at various timesthroughout the study. Twenty-two animalsremained in the study at the time of post-mortem examination. Of these 22 animals,five were in the oral bait group, five werein the oral liquid group, six were in theparenteral group, and six were in thecontrol group.
IFN-c and lymphocyte proliferative responses tomycobacterial antigens
Overall there was no detectable inter-action between group and sampling timeregarding IFN-c responses to PPDb orE:C (P50.25 and 0.39, respectively; datanot shown). There was a significantinteraction detected regarding IFN-c re-sponses to PPDa, although this interactionis based on responses obtained at only onetime point (52 days post-challenge;137 days post-vaccination; data notshown). Least squares means estimatesof vaccinates were compared to those ofthe control group as well. At 28 days post-vaccination and at 13 days post-challenge(98 days post-vaccination) the parenteralgroup produced significantly greater levelsof response to PPDb relative to thecontrol group (P50.09 and P50.057,respectively). At 52 days post-challenge(137 days post-vaccination) the parenteralgroup exhibited increased reactivity toPPDa in relation to the control group(P50.062), whereas the oral group exhib-ited decreased reactivity to PPDa(P50.0025). The control group showedgreater reactivity to PPDb and E:C than
252 JOURNAL OF WILDLIFE DISEASES, VOL. 44, NO. 2, APRIL 2008
all three vaccine groups at 52 days post-challenge (P,0.096) (data not shown).
There were no detectable differences inlymphocyte blastogenic responses to anyof the M. bovis antigens after vaccinationbetween vaccine groups and the controlgroup individually or over time (P$0.57;data not shown).
Post-mortem examination
When present, gross or microscopiclesions consistent with tuberculous lesionswere found in the right cranial lung lobeand mediastinal LNs. Lesions were notconsistently observed in any other tissues,including medial retropharyngeal LNs,although they were present. Therefore,only the right cranial lung lobe andmediastinal LN were suitable for statisticalanalysis.
The number of animals sustaining grosslesions in the right cranial lung lobe andmediastinal LNs in the vaccine groups was
significantly lower than in the controlgroup (oral bait—0/5; oral liquid—1/5;parenteral—1/6; control—6/6 for craniallung lobe; oral bait—0/5; oral liquid—0/5;parenteral—1/6; control—6/6 for medias-tinal LN; P,0.02) (Table 1). No differ-ences could be detected among thevaccine groups regarding the number oflesions in either right cranial lung lobe ormediastinal LN (P51.0).
The numbers of animals with micro-scopic lesions observed in the right craniallung lobe and mediastinal LN of the oralliquid and oral bait groups were signifi-cantly lower than in the control group(oral bait—0/5, oral liquid—0/5, control—6/6 for cranial lung lobe; oral bait—0/5,oral liquid—0/5, control—6/6 for medias-tinal LN; P50.002). In the parenteralgroup, there were fewer animals havinghistologic lesions in the right cranial lunglobe compared to the control group(parenteral—1/6, control—6/6; P50.03).
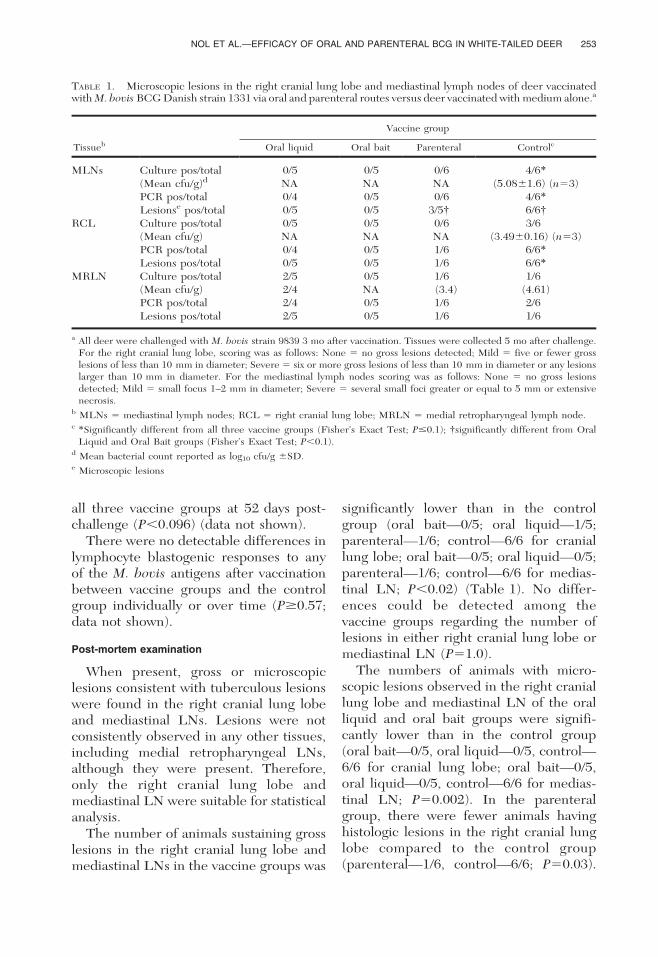
TABLE 1. Microscopic lesions in the right cranial lung lobe and mediastinal lymph nodes of deer vaccinatedwith M. bovis BCG Danish strain 1331 via oral and parenteral routes versus deer vaccinated with medium alone.a
Tissueb
Vaccine group
Oral liquid Oral bait Parenteral Controlc
MLNs Culture pos/total 0/5 0/5 0/6 4/6*
(Mean cfu/g)d NA NA NA (5.0861.6) (n53)PCR pos/total 0/4 0/5 0/6 4/6*
Lesionse pos/total 0/5 0/5 3/5{ 6/6{RCL Culture pos/total 0/5 0/5 0/6 3/6
(Mean cfu/g) NA NA NA (3.4960.16) (n53)PCR pos/total 0/4 0/5 1/6 6/6*
Lesions pos/total 0/5 0/5 1/6 6/6*
MRLN Culture pos/total 2/5 0/5 1/6 1/6(Mean cfu/g) 2/4 NA (3.4) (4.61)PCR pos/total 2/4 0/5 1/6 2/6Lesions pos/total 2/5 0/5 1/6 1/6
a All deer were challenged with M. bovis strain 9839 3 mo after vaccination. Tissues were collected 5 mo after challenge.For the right cranial lung lobe, scoring was as follows: None 5 no gross lesions detected; Mild 5 five or fewer grosslesions of less than 10 mm in diameter; Severe 5 six or more gross lesions of less than 10 mm in diameter or any lesionslarger than 10 mm in diameter. For the mediastinal lymph nodes scoring was as follows: None 5 no gross lesionsdetected; Mild 5 small focus 1–2 mm in diameter; Severe 5 several small foci greater or equal to 5 mm or extensivenecrosis.
b MLNs 5 mediastinal lymph nodes; RCL 5 right cranial lung lobe; MRLN 5 medial retropharyngeal lymph node.c *Significantly different from all three vaccine groups (Fisher’s Exact Test; P#0.1); {significantly different from Oral
Liquid and Oral Bait groups (Fisher’s Exact Test; P,0.1).d Mean bacterial count reported as log10 cfu/g 6SD.e Microscopic lesions
NOL ET AL.—EFFICACY OF ORAL AND PARENTERAL BCG IN WHITE-TAILED DEER 253

However, regarding the mediastinal LN,the parenteral group did not appear todiffer from the control group (parenter-al—3/5, control—6/6; P50.18), but diddiffer from both oral groups, having moreanimals with microscopic lesions in thattissue compared to the other vaccinegroups (P50.08; Tables 2 and 3). Nodifferences could be detected among thevaccine groups regarding the number ofmicroscopic lesions in the right craniallung lobe (P50.13). Microscopic lesions inthe medial retropharyngeal LNs werefound in two animals from the oral liquidgroup, one animal from the parenteralgroup, and one animal from the controlgroup. Again, these data were not sub-jected to statistical analysis and aresummarized in Table 3.
Presence of M. tuberculosis complex in tissues
When looking at all five tissues cultured(right cranial lung, mediastinal LNs,medial retropharyngeal LNs, tonsil, mes-enteric LNs), the oral bait and parenteralgroups, but not the oral liquid group, hadsignificantly fewer culture positive animalsthan did the control group in which all sixanimals were positive (oral bait—0/5 pos.,P50.002; parenteral—2/6 pos., P50.03;oral liquid—3/5 pos., P50.18). No signif-icant differences were noted among thevaccine groups (P50.18). When looking atdifferences in individual tissues (Table 3),there were fewer animals in the oralliquid, oral bait, and parenteral groupsthat were culture positive for M. tubercu-losis complex in the mediastinal LNs thanin the control group (oral liquid—0/5, oral
TABLE 2. Number of deer that are positive for culture, PCR, or microscopic lesions in selected tissues overtotal number of animals tested in the treatment group.a
Vaccine group
Right cranial lung lobe Mediastinal lymph nodes
None Mild Severe None Mild Severe
Oral liquid (n55) 5 0 0 5 0 0Oral bait (n55) 4 1 0 5 0 0Parenteral (n56) 5 0 1 5 1 0Control (n56) 0 0 6b 0 0 6b
a All deer were challenged with M. bovis strain 9839, 3 mo after vaccination. Tissues were collected 5 mo after challenge.None 5 no granulomatous lesions detected; Mild 5 presence of granulomas containing macrophages, giant cells,lymphocytes, and neutrophils, but having incomplete encapsulation and minimal to no necrosis present; Severe 5
presence of granulomas displaying encapsulation, caseous necrosis, and mineralization.b Significantly different from all three vaccine groups (Fisher’s Exact Test; P,0.1).
TABLE 3. Gross lesions in the right cranial lung lobe and mediastinal lymph nodes of deer vaccinated withM. bovis BCG Danish strain 1331 via oral and parenteral routes versus deer vaccinated with medium alone.a
Vaccine group
Right cranial lung lobe Mediastinal lymph nodes
None Mild Severe None Mild Severe
Oral liquid (n55) 5 0 0 5 0 0Oral bait (n55) 5 0 0 5 0 0Parenteral (n56) 5 0 1 2 1 2b
Control (n56) 0 0 6c 0 0 6b
a Deer were vaccinated with M. bovis BCG Danish strain 1331 via oral and parenteral routes or received medium alone.All deer were challenged with M. bovis strain 9839, 3 mo after vaccination. Tissues were collected 5 mo after challenge.
b Significantly different from Oral Liquid and Oral Bait groups (Fisher’s Exact Test; P,0.1).c Significantly different from all three vaccine groups (Fisher’s Exact Test; P#0.1).
254 JOURNAL OF WILDLIFE DISEASES, VOL. 44, NO. 2, APRIL 2008

bait—0/5, parenteral—0/6, control—4/6;P#0.045). Regarding the right craniallung, there were differences, althoughnot significant differences, detected be-tween the vaccine groups and the controlgroup (oral liquid—0/5, oral bait—0/5,parenteral—0/6, control—3/6; P50.18).No differences were observed among thevaccine groups either. No differenceswere seen in the medial retropharyngealLN cultures (Table 3). Quantitative cul-ture results supported the above findingsthat M. bovis was more readily culturedout of mediastinal LNs and right craniallung lobe collected from unvaccinatedanimals than in tissues from vaccinatedanimals. These results are reported inTable 3; however, they were not subjectedto statistical analysis due to small samplesize.
When PCR was applied directly totissues and tissue homogenates, we ob-served similar trends but greater differ-ences among the groups as compared toculture regarding presence of M. tubercu-losis complex in right cranial lung lobe andmediastinal LNs (Table 3). The number ofanimals testing PCR positive for M.tuberculosis complex in mediastinal LNsin the three vaccine groups were signifi-cantly fewer than in the control group(oral liquid—0/4, oral bait—0/5, parenter-al—0/6, control—4/6; P#0.008). As well,there were significantly fewer animalstesting PCR positive in the right craniallung lobe in the three vaccine groups thanin the control group (oral liquid—0/4, oralbait—0/5, parenteral—1/6, control—6/6;P#0.072). Again there were no differenc-es noted among the vaccine groups(P50.28). Culture, PCR, and histopatho-logic results in the three tissues aresummarized in Table 3.
DISCUSSION
The present study indicates that oralBCG Danish 1331 is effective in protect-ing white-tailed deer against diseasecaused by experimental M. bovis infection.
Orally vaccinated deer had fewer tuber-culous lesions, both gross and microscop-ic, than did control deer 5 mo afterchallenge. To the authors’ knowledge, thisis the first study reporting efficacy of oralBCG in this species in experimentalsettings. Waters et al. (2004b) evaluatedcellular immune responses in white-taileddeer vaccinated subcutaneously with BCGPasteur, and demonstrated strong IFN-cand proliferative responses to PPDb,suggesting that BCG may be protectiveagainst M. bovis infection in this species.Challenge with M. bovis was not per-formed in this study. Miller and others(1999) orally administered a recombinantBCG expressing Borrelia burgdorferi OspA to white-tailed deer and detected bothcellular and humoral immune response toM. bovis antigen, evidence that BCG wastaken up via oral route. However, poten-tial efficacy of the vaccine against M. bovisinfection was not investigated, as the solepurpose of this study was to test thefeasibility of BCG as an oral deliverysystem for nonmycobacterial antigens. Ina New Zealand study investigating BCG(Pasteur 1173P2) in red deer (Cervuselaphus), researchers vaccinated animalsdirectly in the tonsilar crypt (Griffin et al.,1999). Similar to oral BCG in white-taileddeer, the New Zealand scientists demon-strated that one dose of 2.53106 cfu BCGinto the tonsilar crypt of red deer protect-ed 8 of 10 animals from disease 21 wkafter M. bovis challenge, and 5 of 10animals were culture negative. Red deerthat were vaccinated subcutaneously withthe same single dose of BCG experiencedcomparable protection, also similar to thewhite-tailed deer in this study. It shouldbe noted that this and subsequent studiesalso demonstrated that red deer vaccinat-ed with two subcutaneous doses of BCG4–8 wk apart fared better than eithersubcutaneous or intratonsilar single-dosegroups after challenge (Griffin et al.,2006).
The results of the present experimentindicate that white-tailed deer can be
NOL ET AL.—EFFICACY OF ORAL AND PARENTERAL BCG IN WHITE-TAILED DEER 255

vaccinated orally using BCG incorporatedin a lipid-formulated bait. This oral baithas successfully induced protectionagainst M. bovis and M. tuberculosisinfection in a number of species, includinglaboratory mice (Aldwell et al., 2003),brushtail possum (Trichosurus vulpecula;Aldwell et al., 2003a), European badger(Meles meles; S. Lesellier, pers. comm.),and domestic cattle (Buddle et al., 2005).Buddle and others (2005) first demon-strated that ruminants can be vaccinatedusing this method. In their study, calvesfed 109 cfu BCG (Pasteur 1173P2) via oralbait and challenged 21 wk after vaccina-tion had significantly lower lung and LNlesion scores than did nonvaccinatedcattle. The current findings are consistentwith this previous study and suggest thatwhite-tailed deer are a good candidate fororal vaccination programs to control bo-vine tuberculosis in the field.
This study examined the level of pro-tection provided by oral BCG in white-tailed deer 5 mo post-challenge. Althoughthe level of protection was significantbased on analyses of lung and mediastinalLNs, these data do not suggest that a lackof lesions equates to sterile immunity. Afew animals still had evidence of infection,based on histologic lesions or positivedetection of M. bovis in medial retropha-ryngeal LNs and occasionally in othertissues as well (data not shown). For thisreason it was important to test tissues inparallel via histopathology, PCR, andculture, as culture for Mycobacteria isquite difficult and is therefore relativelyinsensitive as an independent test. Itwould be of great interest to evaluatedisease progression, if any, in this speciesin subsequent vaccine trials through ex-tending the time between challenge andpostmortem examination. Griffin and oth-ers (2006) addressed this question in reddeer and found that animals vaccinatedsubcutaneously 8 wk apart remained re-fractory to disease up to 52 wk, despite thefact that they harbored low numbers of M.bovis in their tissues. Animals with stable,
nonprogressive lesions are less likely toplay an active role in disease transmission(Griffin et al., 2006). Therefore, vaccinesthat do not produce universal sterileimmunity but do significantly limit diseasestill have great potential to control en-demic tuberculosis in wildlife.
The predominant sites of mucosaluptake of oral BCG in ruminants arecurrently unknown. It has been shown inrabbits that mycobacteria can cross themucosal barrier through M cells present ingut-associated lymphoid tissues, namely,Peyer’s patches (Fujimura, 1986; Tizzard,2004). In Lagranderie et al. (2000), micevaccinated orally with 23109 cfu BCGtransiently harbored BCG in both thesubmaxillary glands and Peyer’s patchesimmediately post-vaccination. However,only the periglandular LNs in the sub-maxillary region maintained high numbersof bacilli 84 days after administration ofBCG when compared to the mesentericLNs, suggesting that primary uptakeoccurs in the pharyngeal region, wherenasopharyngeal lymphoid tissues are pre-sent. In contrast, mice which received thesame dose intragastrically maintainedhigher numbers of bacilli in the mesen-teric LNs. Aldwell and others (2005a,2006) inoculated mice with 5.13107 cfulipid-microencapsulated BCG and foundthat the major site of bacterial replicationwas the mesenteric LNs, indicating thatthe lipid encapsulation reduced uptake inthe pharyngeal region and protected theattenuated bacillus from destruction in thestomach. However, Dorer et al. (2007)showed BCG to be present in cervical andmesenteric LNs and Peyer’s patches ofmice 8 wk after receiving 1–23107 cfulipid-encapsulated BCG. Similar studies inother species corroborate these findings,although they all utilize a monogastricmodel. It is therefore of vital importanceto investigate the potential impact of thecomplex ruminant gastrointestinal systemupon both protected and nonprotectedBCG in future studies.
256 JOURNAL OF WILDLIFE DISEASES, VOL. 44, NO. 2, APRIL 2008

Increases in IFN-c production andproliferation by lymphocytes in the pres-ence of M. bovis antigens can be used asindicators that an appropriate cellularimmune response to BCG vaccinationhas occurred. In infected deer, however,robust IFN-c responses can imply thepresence of more severe disease. Thackeret al. (2006) found that PBMC fromwhite-tailed deer with severe pathologydue to M. bovis infection had greater IFN-c mRNA expression to PPDb and E:Cearly on in the infection than did animalswith pathology limited to the head LNs.As infection progressed the IFN-c re-sponses of the low-pathology group in-creased to those of the high-pathologygroup. Although the deer in this studyresponded well to vaccination, we couldnot have predicted this based on our IFN-c or lymphoproliferative data prior tochallenge. We also did not observe im-portant differences in IFN-c productionbetween controls and vaccinates afterinfection. It is interesting that the groupthat received subcutaneous BCG vaccina-tion displayed a degree of vacillation inIFN-c responses not seen in any of theother groups over the course of the study.Although this may be due to the differ-ence in the character of the immuneresponse to BCG and M. bovis by thisgroup, there may be other factors in-volved, such as the influence of exposureto antigens of M. avium. An importantaspect to consider is that, at multiple timepoints, samples from up to 20% of theanimals did not show an IFN-c responseto any antigen, based on the CervigamELISA, including the pokeweed mitogencontrol. Similar results were reported in astudy by Waters et al. (in press) evaluatingCervigam as a diagnostic tool in variousdeer species, wherein only 44% of white-tailed deer had responses to PWM, callinginto question the validity of this test forthis species. In addition, the IFN-c andlymphocyte responses observed in indi-vidual deer within groups were extremelyvariable. It is quite likely that, had our
sample sizes been larger, we would haveobtained data more consistent with thosereported in other studies.
Experiments involving large, highlystressed species under BL-3 conditionsare typically limited by space and cost.Power and significance levels need to becarefully considered prior to carrying outsuch a study and analyzing the results.Vaccine studies with low sample sizes areparticularly susceptible to producing out-comes that apparently fail to detectdifferences between or among treatmentgroups when there actually is a treatmenteffect. It is therefore justified to increasethe alpha level and hence choose a higherP value in order to increase the power ofthe test and avoid type 2 error that rendersotherwise valuable data meaningless in apractical sense. The purpose of this pilotstudy was to investigate the feasibility ofpursuing further research with oral BCGin white-tailed deer. Despite the fact thatour numbers were very small, we were stillable to clearly show that BCG does appearto be effective in protecting deer fromdisease.
In conclusion, the use of BCG for oralvaccination of white-tailed deer showsgreat potential in controlling diseasecaused by M. bovis infection. Oral vacci-nation significantly slowed pathologic pro-gression of disease in our animals over a 5-mo period and thus is a promisingcandidate for use in field. Oral vaccinationof wildlife is a management tool that hasbeen successfully implemented in Europeto control rabies in red fox (Vulpes vulpes;Brochier et al., 1989) and is currentlybeing used in the United States to managethe disease in raccoons (Procyon lotor).Oral BCG vaccination of wildlife reser-voirs of bovine tuberculosis is beingextensively researched for application inmany countries, including New Zealand,United Kingdom, Ireland, United States,and South Africa. Successful mucosaldelivery (intranasal or intraconjunctival)of BCG to a wild population of brushtailpossums in New Zealand has already been
NOL ET AL.—EFFICACY OF ORAL AND PARENTERAL BCG IN WHITE-TAILED DEER 257

demonstrated by Corner et al. (2002). Theresults of this study provide strong supportfor further research that could ultimatelylead to safe and successful field vaccina-tion of white-tailed deer and eventualeradication of bovine tuberculosis in theUnited States.
ACKNOWLEDGMENTS
The authors would like to thank thefollowing people for their contributions to thisstudy: J. Pollock, B. Olthoff, M. Howard, P.Lasley, R. Lyon, M. Lambeth, C. Fitzpatrick,and K. Deines for their technical assistance; T.Krausman, D. Ewing, J. Steffan, T. Holtz, andR. Whipple for animal care. Our appreciationalso goes out to R. McLean and K. Fagerstonefor their support on this project. Funding forthis study was obtained from USDA/ARS,USDA/APHIS Veterinary Services, USDA/APHIS Science Fellow Program, USDA/APHIS Wildlife Services, and Colorado StateUniversity’s Program of Economically Impor-tant Infectious Animal Diseases through spe-cial grant from USDA:CSEERS.
LITERATURE CITED
ALDWELL, F. E., D. L. KEEN, N. A. PARLANE, M. A.SKINNER, G. W. DE LISLE, AND B. M. BUDDLE.2003a. Oral vaccination with Mycobacteriumbovis BCG in a lipid formulation inducesresistance to pulmonary tuberculosis in brushtailpossums. Vaccine 22: 70–76.
———, I. G. TUCKER, G. W. DE LISLE, AND B. M.BUDDLE. 2003b. Oral delivery of Mycobacteriumbovis BCG in a lipid formulation inducesresistance to pulmonary tuberculosis in mice.Infection and Immunity 71: 101–108.
———, M. A. BAIRD, C. E. FITZPATRICK, A. D.MCLELLAN., M. L. CROSS, M. R. LAMBETH, AND G.S. BUCHAN. 2005a. Oral vaccination of mice withlipid-encapsulated Mycobacterium bovis BCG:Anatomical sites of bacterial replication andimmune activity. Immunology and Cell Biology83: 549–553.
———, L. BRANDT, C. FITZPATRICK, AND I. M. ORME.2005b. Mice fed lipid-encapsulated Mycobacte-rium bovis BCG are protected against aerosolchallenge with Mycobacterium tuberculosis.Infection and Immunity 73: 1903–1905.
———, M. L. CROSS, C. E. FITZPATRICK, M. R.LAMBETH, G. W. DE LISLE, AND B. M. BUDDLE.2006. Oral delivery of lipid-encapsulated Myco-bacterium bovis BCG extends survival of thebacillus in vivo and induces a long-termprotective immune response against tuberculo-sis. Vaccine 24: 2071–2078.
BROCHIER, B., J. BLANCOU, I. THOMAS, B. LANGUET, M.ARTOIS, M. P. KIENY, J. P. LECOCQ, F. COSTY, P.DESMETTRE, AND G. CHAPPUIS. 1989. Use ofrecombinant vaccinia-rabies glycoprotein virusfor oral vaccination of wildlife against rabies:Innocuity to several non-target bait consumingspecies. Journal of Wildlife Diseases 25: 540–547.
BRUNING-FANN, C. S., S. M. SCHMITT, S. D. FITZGER-
ALD, J. S. FIERKE, P. D. FRIEDRICH, J. B.KANEENE, K. A. CLARKE, K. L. BUTLER, J. B.PAYEUR, D. L. WHIPPLE, T. M. COOLEY, J. M.MILLER, AND D. P. MUZO. 2001. Tuberculosis infree-ranging carnivores from Michigan. Journalof Wildlife Diseases 37: 58–64.
BUDDLE, B. M., F. E. ALDWELL, M. A. SKINNER, G. W.DE LISLE, M. DENIS, H. M. VORDERMEIER, R. G.HEWINSON, AND D. N. WEDLOCK. 2005. Effect oforal vaccination of cattle with lipid-formulatedBCG on immune responses and protectionagainst bovine tuberculosis. Vaccine 23: 3581–3589.
BURNHAM, K. P., AND D. R. ANDERSON. 2002. Modelselection and multimodel inference: A practicalinformation-theoretic approach. 2nd Edition.Springer-Verlag, New York, New York, 488 pp.
CORNER, L. A., S. NORTON, B. M. BUDDLE, AND R. S.MORRIS. 2002. The efficacy of bacille Calmette-Guerin vaccine in wild brushtail possums(Trichosurus vulpecula). Research in VeterinaryScience 73: 145–152.
DORER, D. E., W. CZEPLUCH, M. R. LAMBETH, A. C.DUNN, C. REITINGER, F. E. ALDWELL, AND A. D.MCLELLAN. 2007. Lymphatic tracing and T cellresponses following oral vaccination with liveMycobacterium bovis (BCG). Cellular Microbi-ology 9: 544–553.
FITZGERALD, S. D., J. B. KANEENE, K. L. BUTLER, C.K. A. CLARKE, J. S. FIERKE, S. M. SCHMITT, S.BRUNING-FANN, R. R. MITCHELL, D. E. BERRY,AND J. B. PAYEUR. 2000. Comparison of postmor-tem techniques for the detection of Mycobacte-rium bovis in white-tailed deer (Odocoileusvirginianus). Journal of Veterinary DiagnosticInvestigation 12: 322–327.
FUJIMURA, Y. 1986. Functional morphology of micro-fold cells (M cells) in Peyer’s patches: Phagocy-tosis and transport of BCG by M cells into rabbitPeyer’s patches. Gastroenterology Japan 21:325–335.
GRIFFIN, J. F. T., C. G. MACKINTOSH., L. SLOBBE, A. J.THOMSON, AND G. S. BUCHAN. 1999. Vaccineprotocols to optimize the protective efficacyof BCG. Tubercle and Lung Disease 79:135–143.
———, C. G. MACKINTOSH, AND C. R. RODGERS.2006. Factors influencing the protective efficacyof a BCG homologous prime-boost vaccinationregime against tuberculosis. Vaccine 24: 835–845.
258 JOURNAL OF WILDLIFE DISEASES, VOL. 44, NO. 2, APRIL 2008

LAGRANDERIE, M., P. CHAVAROT, A. M. BALAZUC, AND
G. MARCHAL. 2002. Immunogenecity and pro-tective capacity of Mycobacterium bovis BCGafter oral or intragastric administration in mice.Vaccine 18: 1186–1195.
LITTELL, R. C., G. A. MILLIKEN, W. W. STROUP, R. D.WOLFINGER, AND O. SCHABENBERGER. 2006. SASSystem for mixed models. 2nd Edition. SASInstitute, Cary, North Carolina, 814 pp.
MILLER, L. A., B. E. JOHNS, D. J. ELIAS, AND G. J.KILLIAN. 1999. Oral vaccination of white-taileddeer using a recombinant bacillus Calmette-Guerin vaccine expressing the Borrelia burgdor-feri outer surface protein A: Prospects forimmunocontraception. American Journal of Re-productive Immunology 41: 279–285.
NELSON, A. M. 1999. The cost of disease eradication:Smallpox and bovine tuberculosis. Annals of theNew York Academy of Science 894: 83–91.
PALMER, M. V., D. L. WHIPPLE, AND S. C. OLSEN. 1999.Development of a model of natural infection withMycobacterium bovis in white-tailed deer. Jour-nal of Wildlife Diseases 35: 450–457.
———, ———, J. B. PAYEUR, D. P. ALT, K. J. ESCH,C. S. BRUNING-FANN, AND J. B. KANEENE. 2000.Naturally occurring tuberculosis in white-taileddeer. Journal of the American Veterinary Med-ical Association 12: 1921–1924.
SAMUELS, M. L., AND J. A. WITMER. 1999. Statistics forthe life sciences. Prentice-Hall, Upper SaddleRiver, New Jersey, pp. 683.
SAS INSTITUTE. 2006. The GLIMMIX procedure. June.www.sas.com/apps/demosdownloads/sasstatglimmix_PROD_sysdep.jsf?packageID500353/. Accessed27 June 2006.
SCHMITT, S. M., S. D. FITZGERALD, T. M. COOLEY, C.S. BRUNING-FANN, L. SULLIVAN, D. BERRY, T.CARLSON, R. B. MINNIS, J. B. PAYEUR, AND J.SIKARSKIE. 1997. Bovine tuberculosis in free-
ranging white-tailed deer from Michigan. Jour-nal of Wildlife Diseases 33: 749–758.
THACKER, T. C., M. V. PALMER, AND W. R. WATERS.2006. Correlation of cytokine gene expressionwith pathology in white-tailed deer (Odocoileusvirginianus) infected with Mycobacterium bovis.Clinical and Vaccine Immunology 13: 640–647.
TIZZARD, I. 2004. Veterinary immunology: An intro-duction. 7th Edition. Saunders, Elsevier Scienc-es, Philadelphia, Pennsylvania, pp. 494.
VORDERMEIER, H. M., M. A. CHAMBERS, P. J. COCKLE,A. O. WHELAN, J. SIMMONS, AND R. G. HEWINSON.2002. Correlation of ESAT-6 specific gammainterferon production with pathology in cattlefollowing Mycobacterium bovis BCG vaccinationagainst experimental bovine tuberculosis. Infec-tion and Immunity 70: 3026–3032.
WATERS, W. R., B. J. NONNECKE, M. V. PALMER, S.ROBBE-AUSTERMANN, J. P. BANNANTINE, J. R.STABEL, D. L. WHIPPLE, J. B. PAYEUR, D. M.ESTES, J. E. PITZER, AND F. C. MINION. 2004a.Use of recombinant ESAT-6:CFP-10 fusionprotein for differentiation of infections of cattleby Mycobacterium bovis and by M. avium subsp.avium and M. avium subsp. paratuberculosis.Clinical and Diagnostic Laboratory Immunolol-ogy 11: 729–735.
———, M. V. PSLMER, T. C. THACKER, K. ORLOSKI, P.NOL, N. P. HARRINGTON, S. C. OLSEN, AND B. J.NONNECKE. In press. Tuberculosis diagnosisusing the CervigamTM assay: Blood culture andstimulation conditions. Veterinary Record.
———, ———, D. L. WHIPPLE, R. E. SLAUGHTER,AND S. L. JONES. 2004b. Immune responses ofwhite-tailed deer (Odocoileus virginianus) toMycobacterium bovis BCG vaccination. Journalof Wildlife Diseases 40: 66–78.
Received for publication 1 June 2007.
NOL ET AL.—EFFICACY OF ORAL AND PARENTERAL BCG IN WHITE-TAILED DEER 259
Copyright © 2022 FDOKUMEN




















