
Bosnians recruited to support U.S. military's fight against Ebola in West Africa
Upload
independentCategory
view
0download
0
Dendritic cells recruited to the lung shortly after intranasal delivery of
Mycobacterium bovis BCG drive the primary immune response
towards a type 1 cytokine production
MICHELINE LAGRANDERIE,* MARIE-ANNE NAHORI,y ANNE-MARIE BALAZUC,z HELENE KIEFER-BIASIZZO,zJOSE-ROBERTO LAPA E SILVA,§ GENEVIEVE MILON�, GILLES MARCHAL** & BORIS B. VARGAFTIGy
*Departement de Medecine Moleculaire, yUnite de Pharmacologie Cellulaire, zLaboratoire de Cytometrie
Analytique et Preparative, �Unite Immunophysiologie et Parasitisme Intracellulaire and **Laboratoire
de Reference des Mycobacteries, Institut Pasteur, Paris, France, and §Division of Pulmonary
Medicine, Hospital Universitario Clementino Fraga Filho, Federal University of Rio de
Janeiro, Rio de Janeiro, Brazil
SUMMARY
We showed in a previous study that the intranasal (i.n) delivery of bacille Calmette–Guerin (BCG)
to BP2 mice (H-2q) inhibits eosinophilia and bronchial hyperreactivity in a mouse model of asthma.
The present work has been performed to characterize the leucocyte lineages recruited to the lungs of
mice after i.n. delivery of BCG and potentially involved in the polarization of T lymphocytes. The
different antigen-presenting cells (APC) recruited to bronchoalveolar lavage (BAL) and to lung
tissue of mice shortly after the delivery of BCG were analysed in parallel as well as their capacity to
drive the immune response towards a T helper type 1 cytokine production. Alveolar macrophages
(AM) from the BAL were CD11cþ, F4/80þ and CD11b�, and in the lung tissue two major
populations of potential APC were detected: one CD11c�, F4/80þ, CD11bþ and I-Aq� was
identified as interstitial macrophages (IM) and a second expressing CD11cþ and I-Aqþ antigens,
negative for CD11b and F4/80 markers as leucocytic dendritic cells (DC). Freshly isolated DC up-
regulated CD11b and CD40 antigens after overnight culture, but remained negative for CD8aantigen, suggesting a myeloid origin. Lung DC which produced high amount of interleukin (IL)-12
were potent inducers of naive CD4þ T lymphocyte priming, as assessed by interferon-g (IFN-g)
production by these naive CD4þ T cells. Lung explants recovered long term after BCG delivery
produced sustained levels of IFN-g. Our results suggest that AM and particularly DC by secreting
IL-12 shortly after BCG delivery induce the long-term persistence of IFN-g-secreting T cells
percolating in BCG-loaded lung tissue.
INTRODUCTION
Allergic asthma involves airway allergen-induced inflamma-
tion, inflammatory leucocytes, eosinophils and sometimes neu-
trophils, being recruited to the airways as a consequence of the
presence and activation of T lymphocytes, which react with the
inhaled allergens in the respiratory tract. CD4þ T lymphocytes
play a major role in initiating allergic airway inflammation via
the production of T helper type 2 (Th2) cytokines, which trigger
the recruitment of eosinophils to the airways and, very likely,
their subsequent activation.1 It has been suggested that an
imbalance between Th2 and Th1 effectors drives the onset
and sustained pathogenesis of asthma.2,3 Interferon-g (IFN-g)
has been shown to inhibit the development of Th2 responses
both in vitro and in vivo.4,5
We and others have shown that the immunization of mice
with intracellular bacteria known to induce a strong Th1
immune response may counterbalance the allergen-induced
Th2 response and reduce eosinophilia and the associated bron-
chial hyperreactivity (BHR).6–9 The bacille Calmette–Guerin
(BCG)-loaded leucocyte lineages which direct the T cell polar-
ization after the intranasal (i.n.) delivery of BCG have not been
identified. For this reason, we explored, shortly after i.n.
delivery of BCG to the airways, the lineages and number of
leucocytes recruited to the lungs, their loading with BCG and
Immunology 2003 108 352–364
352 # 2003 Blackwell Publishing Ltd
Received 27 August 2002; revised 29 October 2002; accepted 20
December 2002.
Correspondence: Prof. Gilles Marchal, Laboratoire de Reference
des Mycobacteries, Institut Pasteur, 25 rue du Dr Roux, Paris 75015,
France. E-mail: [email protected]

the type of cytokines they induce when acting as antigen-
presenting cells (APC): i.e. as a source of immunogenic signals
for naive and for primed T lymphocytes.
T-cell immunity to live intracellular bacteria is triggered and
maintained by professional phagocytic cells, dendritic cells
(DC) and macrophages, via differential cytokine secretion
and membrane display of different costimulatory molecules
that act as immunogenic signals for T lymphocytes. In the
respiratory tract, alveolar macrophages (AM) are the first
leucocyte subset to clear inhaled soluble allergens or take up
particles including cellular microorganisms. They phagocytose
and secrete mediator molecules, some of which act directly on
the microorganisms within their vacuoles,10 whereas others,
such as interleukin (IL)-12, stimulate IFN-g production and
promote type 1 responses. DC, which also produce IL-12
following microbial stimuli, may preferentially direct the devel-
opment of Th1 cells.11 Immature DC phagocytose live myco-
bacteria in vitro12,13 and it has been recently demonstrated that
DC pulsed with mycobacteria in vitro can efficiently stimulate
mycobacteria-reactive T-cells primed in vivo.14 The inhalation
of bacteria or of soluble proteins results in the recruitment of DC
to the airway epithelium.15 Lung DC are equipped with pha-
gocytic receptors and have a rapid turnover rate, reflecting the
continuous sampling for antigen and migration of these cells to
the draining lymph nodes.16,17 These results suggest that the
AM and/or DC present before BCG delivery and/or recruited,
and the cytokines they produce during the first hours or days
following micro-organism delivery are crucial. AM and lung
DC may thus influence the outcome of the subsequent T-cell
dependent immune reactivity to other unrelated immunogenic
molecules. We therefore studied the time-dependent recruit-
ment of the different APC as well as their capacity to be loaded
with BCG and to secrete bioactive IL-12, in the lungs of mice
shortly after i.n. immunization with BCG. Our study shows that
DC recruited to the lungs during the first hours after BCG
delivery allow naive T cells to secrete IFN-g a Th1-type
cytokine known to play a pivotal role in protecting against
asthma.4,5 Moreover, by secreting IL-12 shortly after the BCG
delivery, lung DC induce the long-term persistence of IFN-gproduction by lung explants from 1 to 3 months after i.n.
delivery of BCG.
MATERIALS AND METHODS
Mice
Male BP2 (H-2q) mice at 6–7 weeks old were obtained from the
Centre d’Elevage Janvier (Le Genest, St Isle, France) and were
maintained in our animal facilities in specific pathogen free-
conditions.
Microorganisms and immunization
The Mycobacterium bovis BCG Pasteur vaccine strain 1173P2
was grown as dispersed bacilli in Beck–Proskauer medium
supplemented with 0�05% Triton WR 1339 (Sigma, St Louis,
MO) and 6% glucose. The bacteria were stored at �708 in
Beck–Proskauer medium supplemented with 0�05% Triton and
6% glycerol. The number of colony-forming units (CFU) per ml
was determined by plating suitable dilutions in phosphate-
buffered saline (PBS) on Middlebrook 7H10 agar medium
(Difco, Fisher Scientific Labs, Elancourt, France). The suspen-
sion was diluted in PBS just before its administration. Una-
naesthetized BP2 mice were immunized i.n. with 106 CFU of
BCG in a volume of 10 ml.
Lung cell isolation for flow cytometry analysis
BP2 mice were anaesthetized intraperitoneally with urethane
(1�5 mg/g body weight) and the thoracic cavity was opened. The
trachea was cannulated and the lungs of each mouse were
washed eight times with 1 ml of saline each by gentle instilla-
tion. After bronchoalveolar lavages, mice were exsanguinated
via the abdominal aorta and their lungs were perfused via the
right ventricular cavity of the heart with 5 ml of saline (0�9%
NaCl). Lungs were aseptically removed, cut into small pieces
and digested for 30 min at 378 in RPMI-1640 medium (Ser-
omed, Munich, Germany) containing collagenase (2 mg/ml)
(Worthington Biochemical Corporation, Lakewood, NJ) and
DNAse I (1 mg/ml; Sigma). Enzyme activity was stopped by
adding 8 ml of RPMI supplemented with 10% fetal calf serum
(FCS). Single cell suspension of lung pieces were obtained by
pushing the digested tissues through cell-strainers (Falcon, BD,
Franklin Lakes, NJ). Cells isolated from bronchoalveolar lavage
(BAL) and lung tissue of 10 individual mice were washed and
resuspended in RPMI medium supplemented with 10% FCS.
Cells from each BAL and lung tissue were counted in haemo-
cytometer and thereafter pooled before stabilization with Sta-
bilCyte medium (BioErgonomics Inc., St Paul MN). Under
these conditions, cells from BAL and lung digests could be
stored at 48 for at least 4 weeks before labelling for flow
cytometric analysis.
In a second experiment in order to characterize more pre-
cisely lung DC, these cells were purified and accessory and
major histocompatibility complex (MHC) class II molecules
were analysed by flow cytometry on freshly isolated and on
overnight cultured lung DC. To purify lung DC, after BAL, cells
from digested lung tissue were suspended in RPMI–FCS sup-
plemented with 2 mM ethylenediaminetetraacetic acid (EDTA)
and passed through a 25-gauge needle to break up clumps. Cells
were counted and incubated for 15 min at 48 at the appropriate
ratios with magnetic activated cell sorting (MACS) CD11c
microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany).
The cells were washed, and diluted in 5 ml of PBS�0�5% FCS,
and CD11cþ cells were isolated by passing the antibody-coated
cell suspensions through a column on an AutoMACS magnetic
cell separator (Miltenyi). Freshly purified lung DC were stabi-
lized with StabilCyte medium or suspended in RPMI�10% FCS
and incubated overnight in tissue culture dishes. The non-
adherent cells and those that easily detached after the overnight
culture were harvested, washed and stabilized in StabilCyte
medium.
Monoclonal antibodies (mAb) for flow cytometric analysis
mAb reactive to CD11c (clone HL3, hamster immunoglobulin
G; IgG), CD11b (Mac-1, M1/70, rat IgG2a), Ly-6G-Gr1 (clone
RB6-8C5, rat IgG2b), CD8a (clone53-6.7, rat IgG2a), I-Aq
(clone KH116, mouse IgG2b), CD80 (B7-1, clone 16-10A1,
hamster IgG), CD86 (B7-2, clone GL1, rat IgG2a), CD40 (clone
3/23, rat IgG2a), were purchased from BD PharMingen (San
Diego, CA) as purified immunoglobulin or immunoglobulin
T priming function of lung DCs after BCG delivery 353
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364

directly conjugated to fluorescein isothiocyanate (FITC), phy-
coerythrin (PE), Cy-Chrome or biotin. Biotin-conjugated anti-
F4/80 (clone C1:A3-4, rat IgG-2b) and DEC-205 (clone NLDC-
145, rat IgG2a) were purchased from Caltag Laboratories,
Burlingame, CA. Streptavidin, anti-rat or anti-mouse IgG con-
jugated to FITC, PE or Cy-Chrome (Caltag Laboratories) were
used as secondary antibodies when necessary.
Flow cytometric analysis
Cells from the BAL and from digested lung tissues were
recovered 6, 24, 48, and 96 hr after the i.n. delivery of BCG
and fixed in StabilCyte medium. Before flow cytometry ana-
lysis, the cells were washed in PBS containing 5% FCS and
stained for 30 min at 48 with purified or directly conjugated
antibodies. If necessary, cells were then washed twice with
PBS–FCS and incubated with a secondary antibody. Isotype
controls were used for each antibody. The cells from all time
points were analysed on the same day on a FACScan flow
cytometer (BD, Oxford, UK). This procedure minimized varia-
bility in staining intensities. The number of cells from the BAL
expressing CD11cþ F4/80þ and negative for CD11b (AM) and
from lung digests expressing either F4/80þ CD11bþ and nega-
tive for CD11c (interstitial macrophages; IM) or CD11cþ and
negative for F4/80 and CD11b (DC) was calculated as: percen-
tage of each subset as determined by FACS analysis � number
of cells counted in the BAL or lung digest, and was expressed as
the mean � SEM.
To characterize more precisely the lung DCs, purified
CD11cþ cells freshly isolated from lung tissue or overnight
cultured in RPMI�10% FCS were analysed on the same day on
a FACScan flow cytometer and expression of accessory and
MHC class II molecules was compared to isotype controls.
Laser scanning image cytometry
To confirm the phenotype of lung dentritic cells and to evaluate
their capacity to achieve their maturation in vitro, purified
CD11cþ cells were cultured in Laboratory-Tek chambered cover-
glass for tissue culture (8 � 105 cells/chamber) (Nunc, Roskilde,
Denmark) for 1 or 24 hr in RPMI–FCS medium supplemented
with 10% granulocyte–macrophage colony-stimulating factor
(GM-CSF) medium. Cells were stained with anti-MHC class II
I-Aq mAb conjugated to PE and then double stained with anti-
B7-2 mAb conjugated to FITC. This technique allows visua-
lization of the same field with cells firstly monolabelled in red
(I-Aq PE-conjugated) and thereafter double labelled in red and
green (I-Aq PE-conjugated and B7-2 FITC-conjugated). The
visualization of cells was obtained with an argon ion laser
(Coherent 92-5, Palo Alto, CA) emitted beam at 488 nm, with
50 mW power reduced to 12�5 mW by an acousto-optic mod-
ulator. A 100� immersion oil objective lens allowed to obtain a
laser spot diameter of 0�2 mm. The effective power at the plane
of focus was determined using a field Master power meter with a
LM-2 detector head (Coherent). An image cytometer ACAS
570 (Meridian, Okemos, MI) was used for cell counting.
Immunohistochemical analysis of the lung
Mice which have received BCG i.n. 48 hr previously were
anaesthetized. BAL was performed and the lungs were inflated
via the trachea with 1 ml of 50% Optimum Cutting Temperature
(OCT; Sakura Finetek, Torrance, CA) in saline solution. One
lobe was placed in an Eppendorf tube and immediately frozen in
liquid nitrogen for cryostat sectioning. The frozen blocks were
stored at �808 prior to use. Sections (5 mm) were cut in a
cryostat at �208 and collected on glass slides coated with poly
L-lysine (Sigma). They were then fixed in chloroform–acetone
v/v (Merck) for 10 min, wrapped in plastic film and stored at
�208 prior to use. Consecutive sections of each block were
stained with the following purified or biotinylated mAb (all
from Pharmingen): CD11c (clone HL3, hamster IgG), I-Aq
(clone KH116, mouse IgG2b). The alkaline phosphatase–
anti-alkaline phosphatase (APAAP) staining procedure was
performed by incubation with rabbit immunoglobulin against
rat immunoglobulin (D455, Dako A/S, Glostrup, Denmark),
followed by rat APAAP antibody (D488, Dako) or alkaline
phosphatase-conjugated streptavidin AP antibody (D0396,
Dako) for the biotinylated primary antibodies. The reaction
was developed with the substrates Fast red TR (Sigma) and
naphthol AS MX phosphate (Sigma), and light haematoxylin
counter-staining. Sections were stained, coded and examined in
a blind fashion. At least 10 fields were analysed, and one
representative field is shown.
Labelling of living bacilli before the phagocytosis assay
To determine the phagocytic capacity of the different APCs,
fluorescent live BCG was prepared. Carboxyfluorescein-diace-
tate succinimidyl ester (CFDA; Molecular Probes Inc, Eugene,
OR) was used to label the bacteria. The native CFDA molecule
is not fluorescent. It is a non-polar molecule that spontaneously
penetrates the bacterial cell membrane. CFDA is then converted
to an anionic fluorescent molecule by intracellular esterases,
which are present and active in live bacteria. The succinimidyl
group makes possible the covalent amine binding of the mole-
cule to any amine residue in the vicinity. The stock solution of
CFDA (1 mg/ml in dimethyl sulphoxide; DMSO) was diluted
1:100 in PBS and 100 ml was added to 1 ml of BCG suspension
(108 CFU/ml). This suspension was incubated in the dark at
room temperature for 60 min Labelled bacteria were centri-
fuged for 15 min at 1600 g, washed twice and resuspended in
PBS. They were able to form colonies on 7H10 medium after
this treatment.
Phagocytosis assay
Alveolar macrophages were obtained from BAL 1, 6, 24 and
48 hr after the i.n. delivery of labelled BCG. The trachea was
cannulated and the lungs were washed as described above.
Lavage fluids from five mice were pooled and centrifuged at
48 for 5 min at 400 g. Cells were incubated 30 min at 48with PE
conjugated anti-Gr1 mAb (a neutrophil marker), and were
washed before incubation at the appropriate ratio for 15 min
at 48 with MACS anti-PE microbeads. The cells were then
washed, and diluted in 5 ml of PBS�0�5% FCS, by passing the
antibody-coated cell suspensions through a column on an
AutoMACS magnetic cell separator. Accordingly, BAL cells
were depleted from neutrophils (Gr1þ).
After BAL, cells from lung tissue were isolated as described
in lung cell isolation for flow cytometry, cells were then
suspended in RPMI–FCS supplemented with 2 mM EDTA
and passed through a 25-gauge needle to break up clumps.
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
354 M. Lagranderie et al.

Cells were counted and incubated at the appropriate ratios with
MACS CD11c or CD11b microbeads for 15 min at 48. They
were washed, and diluted in 5 ml of PBS�0�5% FCS, CD11cþ
or CD11bþ cells being isolated by passing the antibody-coated
cell suspensions through a column on an AutoMACS magnetic
cell separator. Cytospins were prepared by centrifugation of
200 ml containing around 105 cells isolated from BAL or lung
tissue 1, 6, 24 and 48 hr after the i.n. delivery of BCG. Cyto-
spins from BAL (AM) and from purified lung cells – CD11bþ
(IMs) or CD11cþ (DCs) – were stained with Diff-Quick (Baxter
Dade AG, Duedingen, Switzerland). Since non-specific fluor-
escence decreased during storage at 48, the labelled bacteria
were counted by fluorescence microscopy 15 days after stain-
ing. Moreover, the fluorescence due to labelled bacteria bound
to the surface of the cells but not ingested was excluded by
adding trypan blue to the preparations. The fluorescence of exter-
nally bound bacteria was quenched by trypan blue, whereas the
fluorescence of internalized bacteria, which were not in contact
with trypan blue, was not affected. We counted 200–400 cells
and the number of cells that were significantly phagocytic is
expressed as a percentage of the whole population studied.
IL-12 production by lung cells
At 6, 24, 48 and 96 hr after i.n. delivery of BCG, AMs were
harvested from BAL and then IM and DC were isolated from
digested lung tissue. Cells harvested from BAL of 10 mice were
pooled and depleted of neutrophils (Gr1þ) as described above. IM
and DC were isolated from digested lung tissue after incubation
withCD11borCD11cmicrobeadsandpassagethroughamagnetic
columnas describedabove.The purity of the different populations
sorted by the autoMACS system was confirmed by fluorescence-
activated cell sorting (FACS) analysis (90% purity). Purified
cells were cultured in RPMI�10% FCS at 378 and 5% CO2,
supernatants were harvested 24 hr later and assayed for immu-
noreactive IL-12 by enzyme-linked immunosorbent assay
(ELISA). mAb C15.6 (p40 þ p70) (Biosources, Camarillo,
CA) or 9A5 (p70) (Perbio Sciences, Erembodegem-Aalst,
Belgium) were used as the capture antibodies and the anti-
IL-12 mAb C17.8 as the secondary biotinylated antibody. The
binding of the secondary mAb was detected using peroxidase-
conjugated streptavidin (Biosources) with TMB as substrate
(KPL Laboratory, Gaithersburg, MD). Since immunoreactive
IL-12 p70 was not detected in our culture conditions, the results
obtained with IL12 p40 þ p70 correspond to the number of pg
of IL-12 p40/ml of supernatant.
Source and enrichment of naive and primed CD4þ
T lymphocytes
T cells were isolated from inguinal lymph nodes of non
immunized mice or from mice immunized subcutaneously
(s.c) at the base of the tail with 107 CFU of BCG 4 days before
recovery. Cells suspended in PBS�0�5% FCS and incubated at
the appropriate ratios with MACS CD4þ microbeads for 15 min
at 48 were washed, diluted in 5 ml of PBS-0�5% FCS and
separated by passing the antibody-coated cell suspension
through a column on an AutoMACS magnetic cell separator.
Naive and primed CD4þ T cells were counted and cocultured
with the different APC isolated from the lungs of naive or i.n.
immunized mice.
IFN-g and IL-5 production by T cells stimulated with IM,
AM and DC isolated from lungs
The stimulatory activity of lung AM, IM and DC isolated 24, 48
and 96 hr after the i.n. delivery of BCG as described above, was
assessed by coculturing these cells with CD4þ T cells from
naive mice or from mice immunized s.c. with BCG and isolated
as described above. CD4þ T cells were added at a ratio of 1 : 5
to the different APC populations separated from BAL and lungs
of naive or of i.n. immunized mice. The mixed APC and T cell
cultures were then incubated in triplicate in flat-bottomed 96-
well plates (Nunc) for 72 hr at 378 and 5% CO2. The inhibition
of IFN-g production by CD4þ T lymphocytes was monitored
after adding IL-12 p40 neutralizing mAb (clone C15.6) (10 mg/
ml) to some wells.
IFN-g was determined in the supernatants with a commercial
ELISA kit (Biosources). Briefly, 96-well plates (Nunc) were
coated with rabbit anti-mouse/-rat IFN-g polyclonal antibody,
incubated with supernatants and dilutions of rIFN-g standard. A
biotinylated anti-IFN-g antibody (RMMG-1) was added and
antibody binding was detected with steptavidin substrate, absor-
bance being measured at 490 nm. IL-5 was determined with an
immunometric assay: 96-well plates were coated with 10 mg/ml
rat anti-mouse IL-5 (TRFK-4), supernatants or dilutions of rIL-
5 standard were added, followed by an acetylcholinesterase-
labelled rat anti-mouse IL-5 antibody (TRFK-5) at 10 Ellman
units/ml. Absorbance was measured at 405 nm. Results for IFN-
g and IL-5 production are expressed in pg/ml.
IFN-g and IL-5 production by lung explants is a
long-term process after BCG delivery
To evaluate the ex vivo long-term production of IFN-g and IL-5
by lungs at 14, 28, 56, 90 and 116 days after the i.n. delivery of
BCG, lungs were perfused via the right ventricular cavity, they
were cut into small pieces (usually 15 pieces/lung). The pieces
were randomly collected and cultured at 378 and 5% CO2 in 24-
well plates (Nunc), each well containing five lung pieces and
1 ml of AIM V medium (Life Technologies) supplemented with
1% L-glutamine (Life Technologies) and gentamicin (10 mg/ml)
(Sigma). Supernatants from lung explants with medium alone or
stimulated with 1 mg/ml of soluble anti-CD3 mAb were har-
vested after 6 hr culture for IL-5 and after 24 hr culture for IFN-
g determinations. IFN-g and IL-5 contents were measured as
above described. Explants and total lung were weighted (dry
weight) and results are expressed in pg of IFN-g and IL-5
produced by total lung.
Statistical analysis
An unpaired Student’s t-test was used for all analysis. Values of
P < 0�05 were considered statistically significant.
RESULTS
Recruitment of CD11cþ F4/80þ CD11b� (AM), CD11c�
F4/80þ CD11bþ (IM) and CD11cþ F4/80� CD11b� (DC)
to the lungs shortly after i.n. delivery of BCG
To characterize the recruitment of potential APC to BAL and
lungs shortly after the i.n. delivery of BCG (from 6 to 96 hr),
cells were harvested from the BAL fluid and from digested
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
T priming function of lung DCs after BCG delivery 355
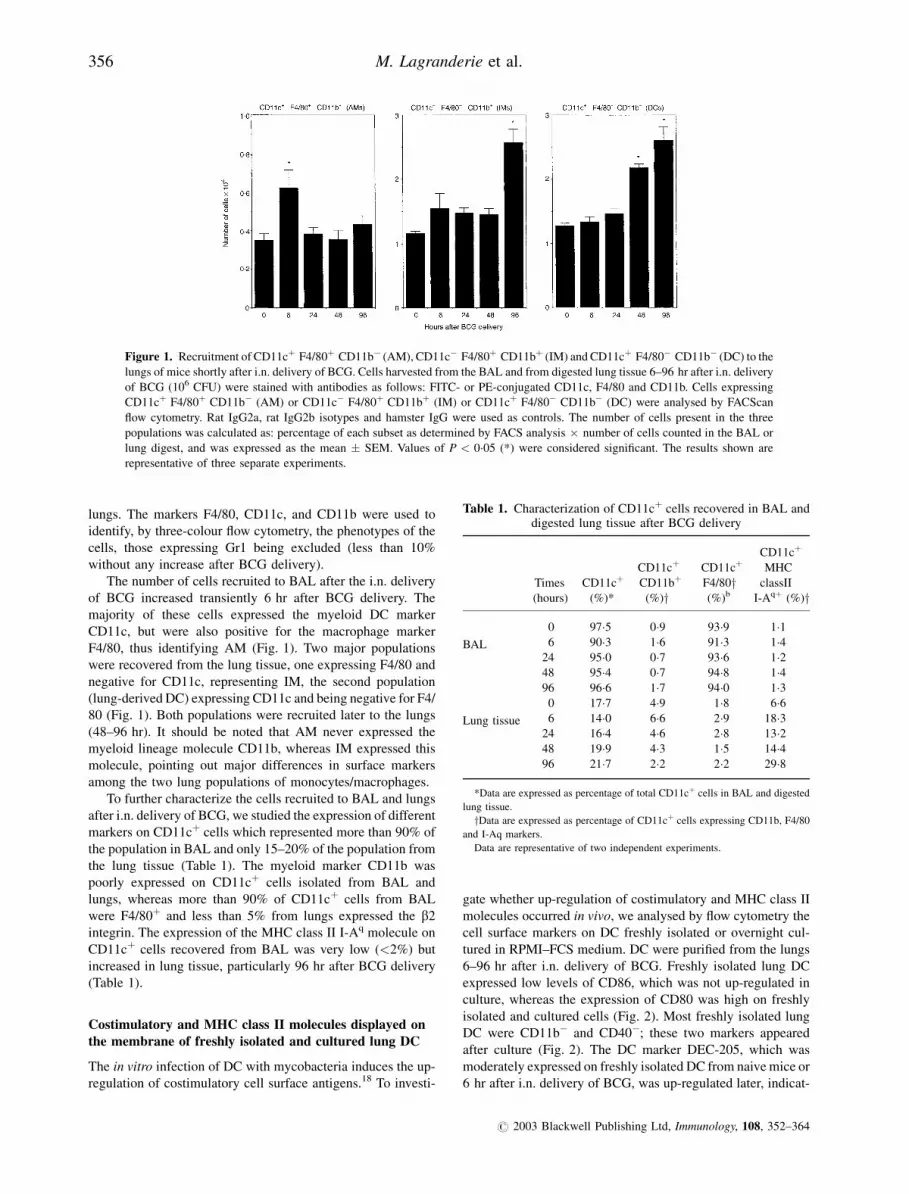
lungs. The markers F4/80, CD11c, and CD11b were used to
identify, by three-colour flow cytometry, the phenotypes of the
cells, those expressing Gr1 being excluded (less than 10%
without any increase after BCG delivery).
The number of cells recruited to BAL after the i.n. delivery
of BCG increased transiently 6 hr after BCG delivery. The
majority of these cells expressed the myeloid DC marker
CD11c, but were also positive for the macrophage marker
F4/80, thus identifying AM (Fig. 1). Two major populations
were recovered from the lung tissue, one expressing F4/80 and
negative for CD11c, representing IM, the second population
(lung-derived DC) expressing CD11c and being negative for F4/
80 (Fig. 1). Both populations were recruited later to the lungs
(48–96 hr). It should be noted that AM never expressed the
myeloid lineage molecule CD11b, whereas IM expressed this
molecule, pointing out major differences in surface markers
among the two lung populations of monocytes/macrophages.
To further characterize the cells recruited to BAL and lungs
after i.n. delivery of BCG, we studied the expression of different
markers on CD11cþ cells which represented more than 90% of
the population in BAL and only 15–20% of the population from
the lung tissue (Table 1). The myeloid marker CD11b was
poorly expressed on CD11cþ cells isolated from BAL and
lungs, whereas more than 90% of CD11cþ cells from BAL
were F4/80þ and less than 5% from lungs expressed the b2
integrin. The expression of the MHC class II I-Aq molecule on
CD11cþ cells recovered from BAL was very low (<2%) but
increased in lung tissue, particularly 96 hr after BCG delivery
(Table 1).
Costimulatory and MHC class II molecules displayed on
the membrane of freshly isolated and cultured lung DC
The in vitro infection of DC with mycobacteria induces the up-
regulation of costimulatory cell surface antigens.18 To investi-
gate whether up-regulation of costimulatory and MHC class II
molecules occurred in vivo, we analysed by flow cytometry the
cell surface markers on DC freshly isolated or overnight cul-
tured in RPMI–FCS medium. DC were purified from the lungs
6–96 hr after i.n. delivery of BCG. Freshly isolated lung DC
expressed low levels of CD86, which was not up-regulated in
culture, whereas the expression of CD80 was high on freshly
isolated and cultured cells (Fig. 2). Most freshly isolated lung
DC were CD11b� and CD40�; these two markers appeared
after culture (Fig. 2). The DC marker DEC-205, which was
moderately expressed on freshly isolated DC from naive mice or
6 hr after i.n. delivery of BCG, was up-regulated later, indicat-
Figure 1. Recruitment of CD11cþ F4/80þ CD11b� (AM), CD11c� F4/80þ CD11bþ (IM) and CD11cþ F4/80� CD11b� (DC) to the
lungs of mice shortly after i.n. delivery of BCG. Cells harvested from the BAL and from digested lung tissue 6–96 hr after i.n. delivery
of BCG (106 CFU) were stained with antibodies as follows: FITC- or PE-conjugated CD11c, F4/80 and CD11b. Cells expressing
CD11cþ F4/80þ CD11b� (AM) or CD11c� F4/80þ CD11bþ (IM) or CD11cþ F4/80� CD11b� (DC) were analysed by FACScan
flow cytometry. Rat IgG2a, rat IgG2b isotypes and hamster IgG were used as controls. The number of cells present in the three
populations was calculated as: percentage of each subset as determined by FACS analysis � number of cells counted in the BAL or
lung digest, and was expressed as the mean � SEM. Values of P < 0�05 (*) were considered significant. The results shown are
representative of three separate experiments.
Table 1. Characterization of CD11cþ cells recovered in BAL anddigested lung tissue after BCG delivery
Times
(hours)
CD11cþ
(%)*
CD11cþ
CD11bþ
(%)y
CD11cþ
F4/80y(%)b
CD11cþ
MHC
classII
I-Aqþ (%)y
BAL
0 97�5 0�9 93�9 1�16 90�3 1�6 91�3 1�4
24 95�0 0�7 93�6 1�248 95�4 0�7 94�8 1�496 96�6 1�7 94�0 1�3
Lung tissue
0 17�7 4�9 1�8 6�66 14�0 6�6 2�9 18�3
24 16�4 4�6 2�8 13�248 19�9 4�3 1�5 14�496 21�7 2�2 2�2 29�8
*Data are expressed as percentage of total CD11cþ cells in BAL and digested
lung tissue.
yData are expressed as percentage of CD11cþ cells expressing CD11b, F4/80
and I-Aq markers.
Data are representative of two independent experiments.
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
356 M. Lagranderie et al.

Figure 2. Flow cytometric analysis of cell surface phenotype of purified lung DC freshly isolated or overnight cultured in RPMI
medium. Lung DC were purified from 6 to 96 hr after i.n. delivery of BCG and stained for MHC class II I-Aq and costimulatory
molecule (CD80, CD86, CD40) and DEC205, CD11b, CD8a expression. The thin open histograms represent isotype control mAb, grey
histograms freshly isolated cells and bold open histograms overnight cultured cells. Data are representative of three separate
experiments.
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
T priming function of lung DCs after BCG delivery 357

ing a more mature phenotype of the DC. The level of expression
of DEC-205 increased in cultured cells only when DC were
isolated from mice that received BCG i.n. The expression of the
lymphoid-related CD8a marker was very low and did not
change in culture. Following overnight culture, CD11cþ cells
expressed CD11b and CD40 antigens and remained negative for
CD8a antigen, suggesting that they were myeloid-derived
(Fig. 2). A large percentage of DC expressed moderate levels
of I-Aq MHC class II and B7-2 molecules even after culture,
suggesting that DCs present and/or recruited to the lung after
i.n. delivery of BCG display an immature phenotype.
We further evaluated if purified lung DC recovered 48 hr
after the i.n. delivery of BCG, would achieve in vitro differ-
entiation if cultured in medium supplemented with GM-CSF. A
laser scanning image cytometry was used to compare MHC
Class II I-Aq and B7-2 antigen expression on freshly isolated
DC and after culture in presence of GM-CSF. This technique
allows visualization of the cells of a same field either unstained
(Fig. 3a, d), stained with anti-MHC class II I-Aq–PE conjugated
mAb (Fig. 3b, e) and double-stained with I-Aq–PE and B7-2–
FITC conjugated mAb (Fig. 3c, f). Forty-eight hours after the
i.n. delivery of BCG, freshly isolated DCs loaded or not with
labelled BCG expressed different intensity in the expression of
I-Aq (Fig. 3b, c), B7-2 not being expressed (Fig. 3c). After
24 hr culture in presence of GM-CSF, DC displayed a more
mature phenotype, expressing higher levels of I-Aq (Fig. 3e, f)
and doubly expressing I-Aq and B7-2 antigens. Indeed, the
majority of cells after 24 hr of culture in presence of GM-CSF
were double-labelled in red and green (Fig. 3f).
Identification and morphology of lung DCs after the i.n.
delivery of BCG
Lung tissue sections were stained with anti-CD11c and MHC
class II I-Aq mAb. The recruitment of positive cells was more
intense in the peribronchiolar zone, even though DC were also
present in the whole lung tissue (Fig. 4a, b). After BAL, DC
were purified from the lung tissue, most cells presenting large
nuclei, abundant cytoplasm and small cytoplasmic projections
(Fig. 4c), with few cells displaying very long cytoplasmic
processes (Fig. 4d). After in vivo phagocytosis, unlabelled
BCG induced weak non-specific fluorescence (Fig. 4e),
whereas CFDA-labelled BCG, visualized by fluorescence
microscopy, were intensely fluorescent (Fig. 4f). The release
of labelled bacterial constituents accounts for the abundant
fluorescent labelling of a few DC.
Phagocytic activity of lung IM, AM and DC
The capacity of different APC to phagocytose mycobacteria is
well documented in vitro12,13,18 and recently it was shown that
both macrophages and DC recovered from the spleen phago-
cytose BCG upon i.v. injection of very high doses (108 CFU).19
However, the phagocytic capacity of the potential APC (IM,
AM, DC) recruited to the lungs after i.n. delivery of BCG
remained to be explored. Therefore, AM were harvested from
the BAL 1, 6, 24 and 48 hr after the i.n. delivery of labelled
BCG. Following exsanguination and lung perfusion with saline,
IM and DC were isolated from the digested lungs. Cytospins
Figure 3. Laser scanning image cytometry on lung-derived DC freshly isolated or overnight cultured with RPMI medium
supplemented with GM-CSF. Lung DC were purified 48 hr after i.n. delivery of CFDA-label BCG, cells were cultured for 1 h (a,
b, c) or 24 hr (d, e, f) with GM-CSF. The laser scanning image cytometry allows visualization of the cells of same field with cells either
unstained (a, d), monolabelled in red (I-Aq conjugated to PE) (b, e) and then double labelled in red and green (I-Aq and B7-2 conjugated,
respectively, to PE and FITC) (c, f). CD11c DCs freshly isolated expressed moderate levels of I-Aq molecule (b), the expression of B7-2
was not observed (c). After overnight culture with GM-CSF the majority of the cells were double-labelled in red and green expressing I-
Aq and B7-2 molecules (f). Figures are representative of two separate experiments.
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
358 M. Lagranderie et al.

were stained with Diff-Quick and vizualized under a fluores-
cence microscope, positive cells (more than two fluorescent
bacilli) being counted. Cytospins obtained 1 hr after the i.n.
delivery of labelled BCG showed numerous AM containing
fluorescent bacilli. No fluorescent bacteria were detected in
DC isolated from lung tissue at this early time point. At 6 hr,
the percentage of AM showing fluorescent bacilli remained
high and few DC (<20%) were loaded with fluorescent bacilli.
At 24 and 48 hr, the percentage of AM and DC showing
fluorescent particles was comparable (40–50%). No fluorescent
Figure 4. Identification and morphology of lung DC after the i.n. delivery of BCG.Frozen lung tissue sections were stained with anti-I-
Aq (a) or anti-CD11c mAb (b). Positive cells in red were easily detected in the peribronchiolar area and at a lower extent in the lung
tissue. Original magnification �20. The tissue sections shown are representative of 10 individual sections. All the CD11c enriched
leucocytes purified from the lung tissue after i.n. delivery of BCG presented large nuclei and abundant cytoplasm with small
cytoplasmic projections, the typical morphology of lung DC (c). A few cells had very long cytoplasmic processes, as shown in this
higher magnification (d). Lung DCs purified after i.n delivery of unlabelled BCG (e) or CFDA-labelled BCG (f) were analysed by
fluorescence microscopy after cytospin and Diff-Quick staining. Labelled bacteria are strongly fluorescent as compared to non labelled
bacteria.
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
T priming function of lung DCs after BCG delivery 359

bacilli were detected at any time in IM from mice immu-
nized i.n. with labelled BCG (Fig. 5a). These results show
that after its i.n. delivery, BCG is phagocytosed by AM and
by DC in the lung tissue, and that IM are unable to phagocytose
bacilli.
IL-12 production by IM, AM and DC shortly after the i.n.
BCG delivery
IL-12 is a potent inducer of T-lymphocyte differentiation and
initiates the development of the Th1 phenotype of naive T cells.
We therefore analysed the capacity of isolated APC from naive
and immunized mice to produce IL-12. AM and DC isolated
from naive mice produced low amounts of IL-12 and 6 hr after
BCG delivery there is no increase of IL-12 production by these
APC harvested, respectively, from the BAL and the lung tissue.
At 24, 48 and 96 hr after the in vivo infection and in absence of
additional ex vivo antigen stimulation, DC produced large
amounts of IL-12 p40 (Fig. 5b). It has to be noted that the
production of IL-12 by AM increased slightly 24 and 48 hr after
delivery of BCG; this could not be caused by DC contamination
because in our experiment the majority of AM producing IL-12
are F4/80þ and this marker is poorly expressed on lung DC. IM,
which failed to phagocytose BCG in vivo, also failed to produce
IL-12 under our experimental conditions (Fig. 5a, b). No
immunoreactive IL-12 p70 was detected in our experimental
conditions.
IFN-c production by T cells stimulated with
IM, AM and DC
We next investigated the stimulatory capacity of lung APC
isolated shortly after the i.n. delivery of BCG. IM, AM and DC
were isolated as described above from 24 to 96 h after i.n. BCG
delivery. These APC were overlaid with syngeneic naive or
immune CD4þ T cells isolated from the inguinal lymph nodes
and purified with a MACS separator. After 72 hr of coculture,
supernatants were assayed for IFN-g content. Co-cultures of
AM with naive CD4þ T cells produced low levels of IFN-g. DC
purified from lung tissue strongly initiated IFN-g secretion by
naive CD4þ T lymphocytes, particularly when they were iso-
lated 48 hr after i.n. delivery of BCG (Fig. 6a). When AM were
used to stimulate immune CD4þ T cells, IFN-g production was
increased, but not to the same extent as when DC were used as
APC (P < 0�05; Fig. 6b). The levels of IFN-g produced by
CD4þ T cells closely correlated with IL-12 produced by APC.
IM, which failed to phagocytose BCG and to produce IL-12, did
not induce IFN-g production by CD4þ T cells (Fig. 6a, b). The
lymph node cells from immunized mice did not produce IFN-gin the absence of APC stimulation, similarly APC from mice
that had not been immunized with BCG failed to stimulate IFN-
g production by CD4þ T cells (data not shown). Finally, the
addition of IL-12 p40 neutralizing mAb to APC/CD4þ T
lymphocyte cocultures prevented IFN-g production (Fig. 6c).
As expected, the stimulation of APC/CD4þ T cell cocultures
with soluble anti-CD3 mAb enhanced IFN-g production what-
ever the source of APC (AMs or DC) but this production was
higher when DC, rather than AM, were used as APC (data not
shown). The production of IL-5 at the limit of detection
(<20 pg/ml) when APC/CD4þ T cells were cocultured without
restimulation was significantly enhanced in the presence of anti-
CD3 (data not shown).
Production of IFN-c and IL-5 by lung explants after
stimulation in vitro with anti-CD3 mAb
Lung explants were prepared from 14 to 116 days after i.n.
delivery of BCG and the ex vivo production of IFN-g and IL-5
was compared to that of lung explants prepared from control
mice. Initially, a time course for the production of cytokines by
those explants stimulated ex vivo by anti-CD3 mAb was studied
and the secretion of IL-5 and IFN-g peaked, respectively, after 6
and 24 hr culture. These intervals were used for monitoring IL-5
and IFN-g production by the lung explants. As shown in
Fig. 7(a), the lung explants collected from 28 to 90 days
after BCG delivery clearly secreted higher amounts of IFN-gthan lung explants from control mice (P < 0�05). The produc-
tion of IFN-g peaked at 28 days (>7000 pg/lung) but remained
at a substantial levels until 90 days after i.n. delivery of BCG
(�3000 pg/lung; Fig. 7a). As expected, BP2 mice with a
Th2 background produced high levels of IL-5 (Fig. 7b); the
large amounts of IFN-g produced by explants from BCG
immunized mice did not inhibit the production of IL-5. This
ex vivo technique for the study of cytokine secretion allowed
Figure 5. Phagocytic capacity and IL-12 p40 production of AM, IM
and DC isolated from BAL and lungs of mice shortly after i.n. delivery
of BCG. (a) BCG bacilli were labelled with CFDA, and 1, 6, 24 and
48 hr after their i.n. delivery, AM were obtained by bronchoalveolar
lavage and IM and DC were isolated from digested lung tissue, followed
by incubation with MACS CD11b or CD11c microbeads and passage
through a column of an AutoMACS separator. Cytospins of purified
AM, IM and DC were stained with Diff-Quick and the bacilli taken up
by phagocytosis were visualized by fluorescence microscopy. Two
hundred to 400 cells were counted and the number of cells that were
phagocytic (more than two bacilli) is expressed as a percentage of the
whole population studied. The experiment is representative of three
separate experiments. (b) AM, IM and DC purified from naive mice and
6, 24, 48 and 96 hr after BCG delivery, were cultured for 24 hr and
supernatants were assayed by ELISA for of IL-12 p40 production. IM
were not able to produce IL-12 p40 and DC produced significantly
higher amounts of IL-12 than AM (P < 0�05). The experiment shown is
representative of three separate experiments. Results are expressed as
mean � SEM.
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
360 M. Lagranderie et al.

demonstration of the fact that, for the first time, IFN-g can be
produced in situ after i.n. delivery of BCG, whereas in earlier
studies IFN-g was not found in BAL and only evidenced after in
vitro restimulation of cells isolated from spleen or lymph
nodes.6,9
DISCUSSION
This study was undertaken to identify, shortly after the delivery
of BCG to the airways, which APC are recruited and their role in
Th1/Th2 regulation of immune responses, a key element of
these experiments involving the study of the in vivo T-cell
priming properties of DC. FACS analysis showed that mono-
cyte/macrophages recruited to the BAL (AM) and those
recruited to the lungs (IM) expressed different markers and
played different roles. The majority of cells recovered from the
BAL expresses the cell surface marker CD11c, which has been
described as a specific marker for mouse DC.20 These cells also
expressed the macrophage marker F4/80, which has been shown
in some subsets of lung or spleen DC.13,21 However, cells
present in the BAL very shortly after i.n. delivery of BCG
had the capacity to phagocytose bacilli, but were unable to
stimulate naive T cells, identifying them as AM rather than DC.
Recently, after administrating fluorescent beads into the mouse
airway, Byersdorfer et al.22 identified cells displaying CD11cþ
and F4/80þ in the BAL fluid, and speculated that they represent
a subset similar to the Langerhans cells. However, their capacity
to stimulate naive CD4þ T cells was not studied; our present
results demonstrate that only CD11cþ isolated from lung tissue
have the capacity to stimulate these T cells. AM expressing
CD11c and F4/80 markers and negative for the CD11b marker
represent a unique subset in the airways, by contrast to IM
recovered from the lung tissue, which never expressed the
CD11c marker and were CD11bþ F4/80þ. Moreover, AM
are well positioned in the alveolar area to take up bacilli rapidly;
Figure 6. IFN-g production by CD4þ T cells stimulated with APC (IM, AM or DC) isolated from the lungs of mice, shortly after i.n.
delivery of BCG. Purified CD4þ T cells from lymph nodes of naive or BCG-immunized mice were cocultured for 3 days with AM, IM
or DC isolated 24, 48 and 96 hr after i.n. delivery of BCG from BAL and lungs, respectively. A 1 : 5 ratio of APC/CD4þ T lymphocytes
was used. Supernatants were assayed by ELISA for IFN-g content. When APC were cocultured with CD4þ T cells from naive mice,
IMs induced no IFN-g production, AM very low levels, whereas DC allowed the production of high amounts of IFN-g by these CD4þ T
cells (a). When APC were cocultured with immune CD4þ T cells isolated from BCG-immunized mice, the IFN-g production was found
only if CD4þ T cells were stimulated with AM or DC. AM were less efficient than DC for initiating the production of IFN-g by CD4þ T
cells (P < 0�05), whereas IM failed to stimulate the production of IFN-g (b). The addition of anti-IL-12 p40 mAb completely abrogated
the production of IFN-g (c). The experiment shown is representative of two separate experiments. Results are expressed as mean �SEM.
Figure 7. Long-term production of IFN-g and IL-5 by lung explants
exposed to anti-CD3 mAb. Lung explants were prepared from 14 to
116 days after i.n. delivery of BCG. After stimulation with soluble anti-
CD3 mAb, the secretion of IL-5 and IFN-g peaked, respectively, at 6 and
24 hr culture period. These intervals were used for monitoring IL-5 and
IFN-g production by the explants. From 28 to 90 days the production of
IFN-g was significantly higher in BCG-immunized than in control mice
(P < 0�05) (a). There were no significant differences in the production
of IL-5 among the two groups of mice (b). Results are expressed as mean
� SEM.
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
T priming function of lung DCs after BCG delivery 361

IM located in the lung tissue failed to do so, and in a previous
study9 we have shown that BCG could be recovered in the AM
26 weeks after i.n. administration.
The third subset of potential APC recovered from the lung
shortly after i.n. delivery of BCG expressing CD11c marker and
negative for the F4/80 marker might represent DC. The influx of
DC into the lungs after i.n. administration of BCG is consistent
with the concept that the airway DC network extension is
dependent upon inflammatory stimuli.15 In vitro studies on
the growth of DC from precursors suggest that DC cultured
in GM-CSF þ IL-4 are specialized for antigen uptake but are
ineffective as activators of T cells.23 However, it is not known
precisely how applicable this in vitro model is to DC popula-
tions in vivo, and particularly to those located in subepithelial
and interstitial compartments, such as lung DC. This study
therefore sought to characterize in detail the phenotype of lung
DC isolated from the perfused lung tissue and to determine to
what degree their function can be modulated by BCG, a
potential promoter of DC maturation and stimulator of Th1
immune responses. To do so, we used histological, flow and
scanning image cytometric approaches and thus characterized
more precisely the DC recruited to the lungs of mice. Forty-
eight hr after BCG delivery, cells expressing CD11c and MHC
class II markers were detected essentially in the peribronchiolar
area of lung sections; the FACS analysis revealed DC expressing
similar markers. Purified lung DC freshly isolated expressed
moderate levels of membrane I-Aq MHC class II and CD86 (B7-
2) molecules, which are considered to be markers of a mature
phenotype, when abundantly expressed.24,25 In accordance with
our results, in a recent study26 lung DC collected after the
intratracheal instillation of macromolecules showed the same
immature phenotype with low-level expression of B7-2 mole-
cule. The elevated expression of I-A and B7-2 molecules is the
hallmark of a mature phenotype of DC,24,25 which were not up-
regulated after 24 hr culture in RPMI medium alone. However,
when purified CD11c cells were cultured in presence of GM-
CSF, the majority of the cells displayed a more mature pheno-
type, indicating that DC freshly isolated from lung tissue can
achieve their maturation in vitro at least in the presence of GM-
CSF. Epidermis-derived Langerhans cells, which presented
similarities to lung DC,27 initiate their maturation process when
cultured with GM-CSF.28 In vivo, at the steady state, lung DC
remain relatively immature but, upon contact with immunogenic
molecules, they evolve towards a mature state. DC maturation
induced their migration out of the immunogen-exposed site into
the interstitial afferent lymphatics, towards the T-cell area of
regional lymph nodes.29 After 24 hr of culture, lung DC up-
regulated the costimulatory molecule CD40 and the myeloid
CD11b marker, suggesting that they are myeloid-derived DC.
In the spleen, three populations of DC have been described:
CD4� CD8aþ DEC205þ CD11b�; CD4þ CD8a� DEC205�
CD11bþ and CD4� CD8a� DEC205� CD11bþ.30 Our lineage
phenotypic analysis showed a population of CD8a�
DEC205þ CD11b� when the expression of these markers
was analysed on freshly isolated DC. After culture, this popula-
tion expressed DEC205þ and CD11bþand remained negative
for the CD8a expression. Lung DC thus do not correspond to a
strict lymphoid or myeloid lineage as it was described in the
spleen.30 Moreover, altogether the procedures used for cell
isolation, for evaluating the phagocytic activity and the devel-
opmental state of the DC must all be considered in analysing
lung DC subtypes. In a recent study22 the phenotype of lung DC
was evaluated only on cells taking fluorescent beads, while in
another report, lung DC phenotyping was performed on a FITC-
bead-negative population.31 In our study, we analysed the
phenotype of DC (CD11cþ) isolated from lung tissue after
i.n. delivery of BCG, without discrimination between cells
loaded or not with BCG. However, this lung-recovered popula-
tion of DC displaying an immature phenotype is highly spe-
cialized for uptake of mycobacteria as shown in vivo in our
study and in vitro by others.12,32
It has been shown that M. tuberculosis-infected DC among
DC generated from mouse bone marrow progenitors are more
potent APC than macrophages,18 and similar results were
recently obtained in vivo with spleen DC following i.v. admin-
istration of M. bovis BCG.19 The functional activity of AM and
lung DC after i.n. delivery of BCG, particularly in terms of their
ability to activate type 1- or type 2-mediated T-cell responses,
still remains to be explored. IL-12 plays a key role in the control
of mycobacterial infection33 stimulating IFN-g production by
natural killer and T cells, leading to the development of a type 1
immune response.34 In this study, in vivo BCG-loaded lung DC
produced more IL-12 p40 than did AM, and the failure to detect
IL-12 p70 in our model may be due to the production of
much less immunoreactive IL-12 p70 than of IL-12 p40.
However, it has been shown that M. tuberculosis-infected DC
present among DC generated from mouse bone marrow pro-
genitors produced IL-12 p70.18 These differences may result
from the complexities of the in vivo interactions of lung DC with
BCG bacilli, as compared to DC generated in vitro and infected
with M. tuberculosis. Mycobacteria-free interstitial macro-
phages did not secrete IL-12 p40, suggesting that the phagocytic
uptake of BCG bacilli is an essential step for triggering IL-12
secretion.
The activation of naive T cells requires two signals provided
by APC. The first signal is delivered through the T-cell receptor
upon engagement of MHC molecules loaded with appropriate
peptides. The second signal involves cross-linking of CD28 and
other receptors on the T cell by costimulatory molecules such as
B7-1, B7-2, CD40 expressed and up-regulated by the APC.
Upon antigen stimulation, CD4þ T cells can be subdivided into
Th1 or Th2 cells. Recent study suggest that the decision to
differentiate into Th1 or Th2 cells occurs shortly after stimula-
tion of naive CD4þ T cells by antigen-pulsed DC.35 The DC
recruited to the lungs early post-BCG delivery promote the
activation of effectors dominated by type 1 immune cytokines,
as shown by short-term production of IFN-g by naive and
immune CD4þ T cells and ex vivo long-term production by
lung explants. Thus, early events occurring after i.n. delivery of
BCG may play a key role in regulating the balance between
Th1- and Th2- mediated immune responses. The activation of
Th1 rather than of Th2 responses by AM and DC after BCG
delivery shown in this study, probably accounts for the protec-
tive effect observed on airway allergen challenge in other
studies.6,9 Thus, despite the Th2-dominant background of
BP2 mice, APC may contribute to the development of Th1
response shortly after i.n. delivery of BCG: probably because
intracellular BCG acts as a strong stimulus on AM and DC,
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
362 M. Lagranderie et al.

inducing the high IFN-g production by activated CD4þ T cells.
No IL-5 associated with the Th2-type response was detected
when CD4þ T cells isolated from secondary lymphoid organs
were co-cultured with lung DC; however, after anti-CD3 sti-
mulation, the production of IL-5 is clearly enhanced (data not
shown). Likewise, upon anti-CD3 stimulation, T-cells of the
lung explants produced high amounts of IL-5; however, differ-
ences among naive mice and those receiving BCG i.n. were not
observed. Nevertheless, by influencing the initiation of a Th1
immune response, through IL-12 production, AM, and parti-
cularly DC, recruited to the lungs of mice at the early stage after
i.n. BCG delivery, induced long-term IFN-g production by CD3
cells present in lung explants. AM and DC once loaded with
BCG are indeed expected to shape an immune response which
could protect against those complex stimuli which result in
allergen-driven asthma.
ACKNOWLEDGMENTS
We thank Stephanie Riveron for technical assistance, and Mai Lebastard
for provinding us with the GM-CSF medium.
This work was supported by Procter & Gamble.
REFERENCES
1 Robinson DS, Hamid Q, Ying S et al. Predominant Th2-like
bronchoalveolar lavage T-lymphocyte population in atopic asthma.
N Engl J Med 1992; 326:298–304.
2 Kline JN, Hunninghake GW. T-lymphocyte dysregulation in asthma.
Proc Soc Exp Biol Med 1994; 207:243–53.
3 Krug N, Frew AJ. The Th2 cell in asthma: initial expectations yet to
be realized. Clin Exp Allergy 1997; 27:142–50.
4 Gajewski T, Fitch F. Anti-proliferative effect of IFN-gamma in
immune regulation. Part I. IFN-gamma inhibits the proliferation
of Th2 but not Th1 murine helper T lymphocyte clones. J Immunol
1988; 140:4245–52.
5 Lack G, Bradey KL, Hamelmann E, Renz H, Loader J, Leung OJ,
Larsen G, Gelfand GW. Nebulized IFN-g inhibits the development of
secondary allergic responses in mice. J Immunol 1996; 157:1432–9.
6 Erb KJ, Holloway JW, Sobeck A, Moll H, Le Gros G. Infection of
mice with Mycobacterium bovis-bacillus Calmette–Guerin (BCG)
suppresses allergen-induced airway eosinophilia. J Exp Med 1998;
187:561–9.
7 Herz U, Gerhold K, Gruber C, Braun A, Wahn U, Renz H, Paul K.
BCG infection suppresses allergic sensitization and development of
increased airway reactivity in an animal model. J Allergy Clin
Immunol 1998; 102:867–74.
8 Yeung VP, Gieni RS, Umetsu DT, DeKruyff RH. Heat killed Listeria
monocytogenes as an adjuvant converts established Th2-dominated
immune responses into Th1-dominated responses. J Immunol 1998;
161:4146–52.
9 Nahori M-A, Lagranderie M, Lefort J et al. Effects of Mycobacter-
ium bovis BCG on the development of allergic inflammation and
bronchial hyperresponsiveness in hyper-IgE BP2 mice vaccinated as
newborns. Vaccine 2001; 19:1484–95.
10 Nathan CF, Hibbs Jr. JB. Role of nitric oxide synthesis in macro-
phage antimicrobial activity. Curr Opin Immunol 1991; 3:65–70.
11 Reis e Souza C, Hieny S, Scharton-Kersten T, Jankovic D, Charest
H, Germain RN, Sher A. In vivo microbial stimulation induces rapid
CD40 ligand-independent production of interleukin 12 by dendritic
cells and their redistribution to T cell areas. J Exp Med 1997;
186:1819–29.
12 Inaba K, Inaba M, Naito M, Steinman RM. Dendritic cell progeni-
tors phagocytose particulates, including bacillus Calmette–Guerin
organisms, and sensitize mice to mycobacterial antigens in vivo. J
Exp Med 1993; 178:479–88.
13 Gonzales-Juarrero M, Orme IM. Characterization of murine lung
dendritic cells infected with Mycobacterium tuberculosis. Infect
Immun 2001; 69:1127–33.
14 Demangel C, Bean AGD, Martin E, Feng CG, Kamath AT, Britton
WJ. Protection against aerosol Mycobacterium tuberculosis infec-
tion using Mycobacterium bovis bacillus Calmette–Guerin-infected
dendritic cells. Eur J Immunol 1999; 29:1972–9.
15 McWilliam AS, Napoli S, Marsh AM et al. Dendritic cells are
recruited into the airway epithelium during the inflammatory res-
ponse to a broad spectrum of stimuli. J Exp Med 1996; 184:
2429–32.
16 Holt PG, Haining S, Nelson DJ, Sedgwick JD. Origin and seady-
state turnover of class II MHC-bearing dendritic cells in the epithe-
lium of the conducting airways. J Immunol 1994; 153:256–61.
17 Lambrecht BN, Prins J-B, Hoogsteden HC. Lung dendritic cells and
host immunity to infection. Eur Respir J 2001; 18:692–704.
18 Bodnar KA, Serbina NV, Flynn JA. Fate of Mycobacterium tuber-
culosis within murine dendritic cells. Inf Imm 2001; 69:800–9.
19 Jiao X, Lo-Man R, Guermonprez Pet al. Dentritic cells are host cells
for mycobacteria in vivo that trigger innate and acquired immunity. J
Immunol 2002; 168:1294–301.
20 Metlay JP, Witmer-Pack MD, Agger R, Crowley MT, Lawless D,
Steinman RM. The distinct leukocyte integrins of mouse spleen
dendritic cells as identified with new hamster monoclonal antibo-
dies. J Exp Med 1990; 171:1753–71.
21 Leenen PJ, Radosevic K, Voerman JS, Salomon B, van Rooijen N,
Klatzmann D, van Ewijk W. Heterogeneity of spleen mouse den-
dritic cells: in vivo phagocytic activity, expression of macrophage
markers, and subpopulation turnover. J Immunol 1998; 160:
2166–73.
22 Byersdorfer CA, Chaplin DD. Visualization of early APC/T cell
interactions in the mouse lung following intranasal challenge. J
Immunol 2001; 167:6756–64.
23 Sallusto F, Lanzavecchia A. Efficient presentation of soluble antigen
by cultured human dendritic cells is maintained by granulocyte/
macrophage colony-stimulating factor plus interleukin 4 and down
regulated by tumor necrosis factor alpha. J Exp Med 1994;
179:1109–18.
24 Pierre P, Shannon TJ, Gatti E, Hull M, Meltzer J, Mizra A, Inaba K,
Steinman RM, Mellman I. Developmental regulation of MHC class
II transport in mouse dendritic cells. Nature 1997; 388:787–92.
25 Inaba K, Witmer-Pack M, Inaba M et al. The tissue distribution of
the B7-2 costimulator in mice: abundant expression on dendritic cells
in situ and during maturation in vitro. J Exp Med 1994; 180:1849–60.
26 Vermaelen KY, Carro-Muino I, Lambrecht BN, Pauwels RA. Spe-
cific migratory dentritic cells rapidly transport antigen from the
airways to the thoracic lymph nodes. J Exp Med 2001; 193:51–60.
27 Pollard AM, Lipscomb MF. Characterization of murine lung den-
dritic cells: similarities to Langerhans cells and thymic dendritic
cells. J Exp Med 1990; 172:159–67.
28 Witmer-Pack MD, Valinsky J, Olivier W, Steinman RM. Quantita-
tion of surface antigens on cultured murine epidermal Langerhans
cells: rapid and selective increase in the level of surface MHC
products. J Invest Dermatol 1988; 90:387–94.
29 Cyster JG. Chemokines and cell migration in secondary lymphoid
organs. Science 1999; 286:2098–102.
30 Shortman K. Dendritic cells: multiple subtypes, multiple origins,
multiple functions. Immunol Cell Biol 2000; 78:161–5.
31 Masten BJ, Yates JL, Pollard Koga AM, Lipscomb MF. Character-
ization of accessory molecules in murine lung dendritic cell func-
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
T priming function of lung DCs after BCG delivery 363

tion. Roles for Cd80, Cd86, Cd54, and CD40L. Am J Respir Cell
Mol Biol 1997; 16:335–42.
32 Henderson RA, Watkins SC, Flynn JA. Activation of human den-
dritic cells following infection with Mycobacterium tuberculosis. J
Immunol 1997; 159:635–43.
33 DeJong R, Altare F, Haagen IA et al. Severe mycobacterial and
Salmonella infections in interleukin-12 receptor-deficient patients.
Science 1998; 280:1435–8.
34 Macatonia SE, Hosken NA, Litton M et al. Dendritic cells produce
IL-12 and direct the development of Th1 cells from naive CD4þ T
cells. J Immunol 1995; 154:5071–9.
35 Tollener KM, Luther SA, Sze DM, Choy RK, Taylor DR, MacLen-
nan ICM, Acha-Orbea H. T helper 1 (Th1) and Th2 characteristics
start to develop during T cell priming and are associated with an
immediate ability to induce immunoglobulin class switching. J Exp
Med 1998; 187:1193–204.
# 2003 Blackwell Publishing Ltd, Immunology, 108, 352–364
364 M. Lagranderie et al.
Copyright © 2022 FDOKUMEN




















