Eimeria bovis-induced modulation of the host cell proteome at the meront I stage

ELSEVIER FEMS Immunology and Medical Microbiology 15 (1996) 213-222
IMMUNOLOGY AND MEDICAL MICROBIOLOGY
Chemical and ultrastructural investigations of Mycobacterium bovis BCG: implications for the
molecular structure of the mycobacterial cell envelope
Melvin E. Klegerman a3 * , Priscilla 0. Devadoss a, Jo& L. Garrido a*b, Hector R. Reyes b, Michael J. Groves a
a Institute for Tuberculosis Research, College of Pharmacy, University of Illinois at Chicago, 950 South Halsted Street (M/C 964), Chicago, IL 60607, USA
b Chemical Engineering Department, College of Engineering, Unitiersity of Illinois at Chicago, 850 South Clinton Street (M/C IlO), Chicago, IL 60607, USA
Received 17 January 1996; revised 28 May 1996; accepted 25 June 1996
Abstract
The mycobacterial cell wall visualized by transmission electron microscopy (TEM) of thin sections of resin-embedded specimens is generally believed to consist of an electron-dense peptidoglycan, an electron-transparent arabinogalactan- mycolate layer and an ele:ctron-dense outer layer (OL). In addition, a pseudocapsule known as the ‘electron-transparent zone’ (ETZ) has been observed after phagocytosis of mycobacteria by macrophages. TEM of thin sections of Mycobacterium
bouis BCG, Ticea substrain, revealed an OL bilayer, each of which measured 2-4 nm in diameter. The intermediate electron-transparent layer varied from 1 to about 250 nm in diameter and appears to be a previously observed oxygen-depen- dent amorphous integument that consists of hot water-extractable neutral polysaccharides, especially a recently characterized o-glucan, comprising about 12% of the dry cell weight. This and other recent studies of BCG have revealed cell-surface features that may provide a better understanding of the outer mycobacterial cell envelope.
Keywords: Mycobacterial cell wall structure; Transmission electron microscopy; Electron-transparent zone
1. Introduction
Current models of mycobacterial cell wall struc-
ture generally agree on a multi-layer scheme relating
transmission electron microscopy (TEM) of thin sec-
tions to the chemistry of the bacterial envelope [l-3].
* Corresponding author.
From the cytoplasm to the cell periphery, a unit
plasma membrane bilayer is followed by an
electron-transparent petiplasmic space, an electron-
dense peptidoglycan layer, an electron-transparent
middle wall and an electron-dense outer layer (OL). An electron-transparent zone (ETZ) of varying thick- ness that surrounds the cell beyond the OL has also been described by several investigators [4-71. The cell wall skeleton, consisting of a chemotype IV peptidoglycan (alternating p- 1 ,Clinked N-acetyl+
G928-8244/96/$15.00 Copyright 0 1996 Federation of European Microbiological Societies. Published by Elsevier Science B.V.
PII SO928-8244(96)00061-2

214 M.E. Klegerman et al./ FEMS Immunology and Medical Microbiology 15 ~19961213-222
glucosamine and D-muramic acid to which the tetra- peptide L-Ala-y-D-Glu-Dap-D-Ala is attached) [I] co- valently linked to a mycolylated arabinogalactan, spans the peptidoglycan and middle wall layers. The
OL is believed to contain charged polysaccharides, proteins and complex glycolipids with their apolar chains extending into the middle wall. The ETZ is believed to consist of either carbohydrate or myco- side B (phenolic glycolipids) in the case of M. bouis-BCG [1,4,7,8].
Previous studies indicated that an ETZ of consid- erable thickness could be identified in TEM of thin sections of BCG growing under normal in vitro conditions and that this structure corresponds to a secreted amorphous integument, visualized by scan-
ning electron microscopy @EM), that serves to bind the bacilli together [9]. We have carried out further ultrastructural studies of A4. bouis-BCG, Tice@ sub- strain, and chemical analyses of the separated integu- ment from both the Connaught and Tice BCG sub- strains that provide new information about the struc- ture and composition of the outer cell envelope of this mycobacterial species.
2. Materials and methods
2.1. Transmission electron microscopy (TEM) 2.2. Scanning electron microscopy (SEM)
Tice@-substrain BCG was suspended in 0.02 M phosphate-buffered saline, pH 7.4 (PBS), after re-
Treated BCG samples and fractions were fixed in 3 ml of 2.5% glutaraldehyde in phosphate-buffered
constitution of freeze-dried vaccine and centrifugal washing or was harvested with a wire loop from colonies growing on Middlebrook 7H10 agar after
plating of undiluted vaccine. Heat-treated BCG was prepared by holding resuspended vaccine in a boiling water bath for 15 min.
Organisms were triple-fixed according to a modi- fication of the method of Kellenberger et al. [lo]. Successive fixation with 4% glutaraldehyde in 0.2 M sodium cacodylate buffer with 0.01% CaCl,, pH 7.2, 1% osmium tetroxide in Ryter-Kellenberger buffer, pH 6.0, and 0.5% uranyl acetate in Ryter-Kellen- berger buffer, pH 5.0, was followed by dehydration in ethanol (500/o, 70%, 80%, 95% and 2 X 100%) and propylene oxide (2 X ), embedding in Epon by the method of Luft [I l] and curing at 60°C for 24 h.
Ultrathin sections were cut with a diamond knife (Diatome, Fort Washington, PA) using an MT-6000 Sorvall ultramicrotome (Du Pont Co., Wilmington, DE), collected on coated copper grids and double- stained with 2% aqueous uranyl acetate and 0.5% alkaline lead citrate according to the methods of Watson [ 121 and Fahmy [13]. The sections were examined and photographed with a Philips EM 300 electron microscope (Philips Electronic Instruments, Mahwah, NJ) operated at 80 kV.
Fig. 1. Longitudinal section of a bacillus showing the outer layer (OL), the electron-translucent mycolate-rich layer (ML), inner
peptidoglycan layer (IL), periplasmic space (P) and plasma membrane (PM). X 79 800.

M.E. Klegeman et al./ FEMS Immunology and Medical Microbiology 15 (1996) 213-222 215
saline (PBS), pH 7.4, for 90 min. Suspended material was collected on 0.22 pm pore size Millipore filters (Millipore Corp., Bedford, MA) before dehydration through a graded ethanol series, critical point drying and sputter-coating with gold-palladium. Specimens
were visualized and photographed with a JEOL JSM-35C scanning electron microscope.
2.3. Integument separation, extraction and analysis
Samples of reconstituted freeze-dried Connaught- and Tice@-substrain 13CG vaccine (obtained from
Connaught Laboratories, North York, Ontario, Canada, and archived at the Institute for Tuberculo- sis Research, respectively) were grown in submerged Middlebrook 7H9 (Connaught) or Sauton (Tice) medium cultures for five weeks at 37°C. BCG har-
vested by centrifugation was treated with Protease type XIV (pronase) from Streptomyces griseus (Sigma Chemical Co., St. Louis, MO) at an enzyme concentration of 1 mg/ml for 60 min at ambient temperature, resulting in separation of the integu- ment from the intact BCG cells (verified by SEM), which retained their viability [14,15].
Fig. 2. Agar-grown BCG, revealing the OL, plasma membrane (PM), extracellular integument (I), membrane-like structures in different
orientations (S). Note the displaced OL (arrows), which eventually forms the extracellular integument. The double arrow indicates a young
lysed cell in which some membrane-like structures have remained intact. R = ribosomes, V = vesicles. X 63 840.

216 M.E. Klegennan et al./ FEM.7 hnmunology and Medical Microbiology 15 (1996) 213-222
Buoyant density centrifugation (500 X g at ambi- ent temperature in a swinging bucket rotor for 7 min) of the digest on Histopaque’-1077 (Sigma Chemical Co.) yielded cells devoid of integument in the pellet, while integumentary material in the presence of some cells was recovered from the supemate. Each frac- tion was washed twice with water and lyophilized prior to boiling water extraction [ 191 for 2 h under reflux, followed by filtration, concentration in a ro- tary evaporator, dialysis against water and lyophiliza- tion.
For sugar analysis, extracts (1.5 mg) were hydro- lyzed in 2.0 M trifluoroacetic acid (TFA) at 121°C at 15 psi for 1 h, while enzymatic glucose analysis (Stanbio Laboratory, San Antonio, TX) was per- formed on samples that had been hydrolyzed in 0.1 N HCl at 121°C at 15 psi for 2 h, followed by
neutralization with NaOH. Sugar analysis was per- formed by HPLC with a Dionex DX 500 Chro- matography System (Dionex Corp., Sunnyvale, CA) [ 161 relative to monosaccharide standards. Colony- forming units of Histopaque fractions were deter- mined in a semi-solid medium by the method of Rosenthal et al. [17]. BCG viability was also deter- mined as cell-associated ATP extracted at 100°C for 3 min and measured by bioluminescence assay [ 181.
3. Results
The M. bouis cell wall clearly had a multi-layered appearance on TEM (Fig. 1). A unit plasma mem- brane bilayer of about 7 nm was separated from a relatively diffuse electron-dense peptidoglycan layer
Fig. 3. A transverse section of an agar-grown colony showing the OL covering the upper surface over several cells. Note the displaced OL
that becomes the extracellular integument. Double arrows indicate membrane-like structures in different orientations. X 40 750.

ME. Klegennan el al. / FEMS Immunology and Medical Microbiology 15 11996) 213-222 217
by a variable peripla:smic space of up to 10 mn thickness. A middle wall bilayer was easily discem- able and consisted of a lower electron-transparent layer of 3-5 nm thickness and an upper electron- translucent layer of 5-20 nm thickness. In single cells, a dark electron-dense OL was seen either as a single layer of about 5 nm thickness or a double layer that appeared similar to the plasma membrane. In BCG vaccine cell aggregates and the dorsal side of agar-grown colonie:s (Figs. 2-41, the outer layers were separated by a variable ETZ that attained a thickness of 250 nm or more and appeared to form a network among the cells. The upper OL was often covered with a variablle electron-dense accretion that exhibited ultrastructural features resembling ribo- somes and membrane fragments and was even ob- served to cover several cells (Fig. 4).
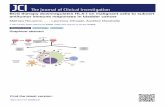
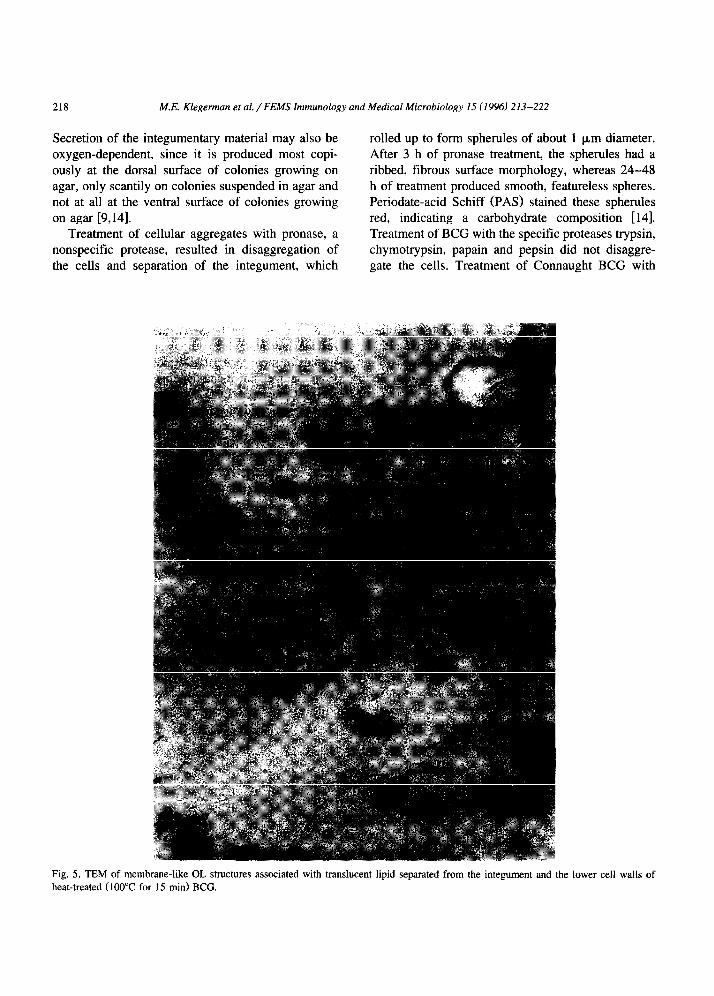
TEM examination of heat-treated BCG suspen- sions prepared under conditions consistent with ex- traction of a polysacch.aride-rich fraction [ 191 showed the apparent generation of insoluble lipid-OL com- plexes (Fig. 5). BCG cell walls after boiling ap- peared to consist only of peptidoglycan and middle wall layers (Fig. 61, suggesting that extracted mate- rial would reside mainly in the ETZ. This possibility was confirmed by extraction and analysis of the pronase-separated integument. CFU and ATP assays of fractions prepared from Connaught BCG indicated
that 50-62% of recovered viable cells remained in the Histopaque supemate with the integument. Since 65% of the BCG dry mass was consistently found in this fraction, the integument may account for 1 l-30% of the dry mass in this BCG substrain under the culture conditions employed. Tice-substrain BCG grown in Sauton medium and fractionated the same way yielded similar results.
Hot-water extraction of both the Connaught and Tice BCG Histopaque supemates yielded about 30% dry residue, while similar extraction of the Con- naught cell pellet, which was ascertained to be de- void of integument by SEM, yielded no detectable residue, indicating that all hot-water extractable com- pounds of more than 10 kDa molecular mass derive from the integument and the upper OL. Sugar analy- sis of Connaught BCG integument extract hydro- lyzates indicated that about half of the extracted material was polysaccharide, consisting of 70-75% D-glucose, 11% D-galactose and 12-18% D-mannose.
4. Discussion
Scanning electron microscope @EM) studies of BCG have demonstrated that the organisms produce an amorphous integument that physically aggregates the cells and apparently corresponds to the ETZ [9].
Fig. 4. A sectional view of the integument showing its membrane-like composition (arrows). X 40750.
218 M.E. Klegerman et al. / FEMS Immunology and Medical Microbiology 15 (1996) 213-222
Secretion of the integumentary material may also be oxygen-dependent, since it is produced most copi- ously at the dorsal surface of colonies growing on agar, only scantily on colonies suspended in agar and not at all at the ventral surface of colonies growing on agar [9,14].
Treatment of cellular aggregates with pronase, a nonspecific protease, resulted in disaggregation of the cells and separation of the integument, which
rolled up to form spherules of about 1 pm diameter. After 3 h of pronase treatment, the spherules had a ribbed, fibrous surface morphology, whereas 24-48 h of treatment produced smooth, featureless spheres. Periodate-acid Schiff (PAS) stained these spherules red, indicating a carbohydrate composition [14]. Treatment of BCG with the specific proteases trypsin, chymotrypsin, papain and pepsin did not disaggre- gate the cells. Treatment of Connaught BCG with
Fig. 5. TEM of membrane-like OL structures associated with translucent lipid separated from the integument and the lower cell walls of
heat-treated (100°C for 15 min) BCG.
M.E. Klegennan et al. / FEMS Immunology and Medical Microbiology 15 (1996) 213-222 219
pronase for 1 h in the study reported here liberated bacilli from the integument (as visualized by SEM),
which remained flat and extended. Determination of 13CG surface charge character-
istics to a depth of 4 nm [20] indicated that about half the negative charge, which predominates at neu- tral pH, may be attributed to carboxylate groups, while one-third is due to the charged oxygens of phosphodiesters and one-sixth is due to strong acidic groups such as sulfate.. Standardless quantitative x-ray microanalysis confirmed the presence of phosphorus
and sulfur within 10 nm of the cell surface [20]. The entire positive charge, which predominates below pH 3, is attributable to amino groups. Protease treat- ments established that these groups are completely contained within proteins, but that few of the carbox- ylate groups are. Pronase abolished half the negative charge, indicating that most of the cell-surface car- boxylate groups are located within an anionic polypeptide that does not contain basic or aromatic residues [21] and somehow attaches the integument to the OL.
Fig. 6: TEM of heat-treated BCG cells showing mesosomal vesicles that were released from the cells. Note
peptidoglycan layer and the middle wall.
the retainment of the

220 M.E. Klegerman et al./ FEMS Immunology and Medical Microbiology 15 (1996) 213-222
This study supports the hypothesis that the ETZ is solubilized by boiling of BCG, since TEM of heat- treated Tice BCG showed no evidence of the ETZ, while disrupted OL structures could be seen and the cell wall of the remaining cells only extended through the upper middle wall. Water-soluble material, termed PSl, extracted under these conditions for 2 h consisted of at least 50% neutral polysaccharides,
including D-glucans, D-mannans and D-arabino-D- mannans. Lipids comprised no more than ten percent of the extract and contained esterified palmitic and
stearic acids. Only a trace of lipoarabinomannan (LAM) was found in this fraction [19]. The remain- ing PSl material consisted mainly of variable amounts of protein.
Others have noted the variable thickness of the ETZ, extending up to 300nm, but have believed it to consist mainly of mycosides (phenolic glycolipids in M. tuberculosis and M. bouis) [22-241. Identifying
the ETZ as a ‘capsule’ or ‘capsule-like structure’, Rastogi et al. [3] contended that the OL is situated below the capsule in M. &urn, but TEM observa- tions of M. bovis BCG reported here and previously
[9,25] clearly showed the OL above the ETZ as the outermost layer. Rastogi et al. [3] also surmised that the capsule may consist of polymethylated saccha- ride moieties of the lipopolysaccharides anchored in the electron-transparent layer of the middle wall.
An OL bilayer or surface pseudomembrane had been described by Chapman et al. [26] and Imaeda et al. [27] for M. lepraemurium and M. leprae, respec- tively, but formation of the ETZ between the outer layers may not have been noted previously. The
evidence summarized here indicates that the elec- tron-dense OL may consist of anionic amphipathic molecules with their lipid moieties extending both between the bilayers and into the middle wall.
The phosphate at the cell surface (i.e. in the upper OL) may be due to phosphoinositolmannoside (PIM), lipomannan (LM), lipoarabinomannan (LAM) and other phosphoglycolipids, while the strong acid groups could be provided by sulfolipids. The acidic polypeptidyl carboxy functions may be indicative of poly-L-glutamic acid previously found in the cell walls of BCG and M. tuberculosis, but not other mycobacterial species [28,29]. The release of the integument and the upper OL by pronase treatment of BCG implies that the polyglutamate somehow
connects the two OL layers, despite formation of the ETZ between them. This possibility was supported by the observation that Protease type XVII-B from Staphylococcus aureus, strain V8, which specifically cleaves peptide bonds on the carboxyl side of aspar- tic and glutamic acid residues, appeared to release Connaught bacilli from the integument (visualized with SEM; unpublished). The lower OL appeared intact on TEM photos of pronase-treated BCG (not
shown). The composition of PSl, a boiling water extract
of BCG [19], suggests that the ETZ is largely com-
posed of neutral polysaccharides, especially a D- glucan and a D-arabino-D-mannan, that are not linked to lipids or phosphate. Recently, Orgalo-MagnC et al. 1301 reported that the ‘capsular material’ of M. tu- berculosis contains appreciable quantities of a D- glucan, a D-arabino-D-mannan and a D-mannan. The glucan, a glycogen with o-1,6 branches, was identi- cal in structure to a previously characterized D-glucan found in the culture filtrate of M. tuberculosis [31]. The structure of this glucan is similar to that found for the D-glucan of PSl, a dextran with + 4)a-D- Glcp[o-D-Glcp(1 + 6)] branches [32], which may be a common feature of both glucans.
Numerous investigations have indicated that the outer mycobacterial cell envelope varies greatly among species. While the pseudocapsule of M. bovis and M. tuberculosis appears to be primarily carbo- hydrate in nature, that of M. auium and M. kansasii has been shown to be predominantly lipidic, consist- ing mainly of mycosides C and phenolic glycolipids,
respectively [3,33]. Ortalo-MagnC et al. [34] have also demonstrated that surface-exposed lipids vary widely in nature among mycobacterial species. Fur- ther characterization of the ETZ, capsular/pseudo- capsular or integumentary material (all of which appear to be the same entity) of different mycobacte- rial species may provide important new insights into the pathogenicity and virulence factors of these or- ganisms.
Acknowledgements
We wish to thank the Electron Microscope Facil- ity of the Research Resources Center, University of

M. E. Klegerman et al. / FEMS Immunology and Medical Microbiology 15 (1996) 213-222 221
Illinois at Chicago, which provided equipment and assistance necessary to carry out these investigations.
References
[II McNeil, M.R. and Brennan, P.J. (1991) Structure, function
and biogenesis of the cell envelope of mycobacteria in
relation to bacterial physiology, pathogenesis and drug resis-
tance; some thoughts and possibilities arising from recent
structural information. Res. Microbial. 142, 451-463.
[2] Minnikin, D.E. (1991) Chemical principles in the organiza-
tion of lipid components in the mycobacterial cell envelope.
Res. Microbial. 142, 423-427.
[3] Rastogi, N., Hellio, R. and David, H.L. (1991) A new insight
into the mycobacterial cell envelope architecture by the
localization of antigens in ultrathin sections. Zbl. Bakt. 275,
287-302.
[4] Kolbel, H.K. (1984) ELlectron microscopy. In: The Mycobac-
teria. A Sourcebook, Part A (Kubica, G.P. and Wayne, LG.,
Eds.), pp. 249-300. Marcel Dekker, Inc., New York.
[5] Ryter, A., Frehel, C., Rastogi, N. and David, H.L. (1984)
Macrophage interaction with mycobacteria including M. lep-
roe. Acta Leprol. 2, 211-226.
[61 Rastogi, N., Potar, MC. and David, H.L. (1987) Intracellular
growth of pathogenic mycobacteria in the continuous murine
macrophage cell line J-774: ultrastructure and drug-suscept-
ibility studies. Curr. Microbial. 16, 79-92.
[7] Rastogi, N. and David, H.L. (1988) Mechanisms of patho-
genicity in mycobacteria. Biochimie 70, 1101-l 120.
181 Chatterjee, D., Bozic, C.M., Knisley, C., Cho, S. and Bren-
nan, P.J. (1989) Phenolic glycolipids of Mycobacterium bovis: new structures and synthesis of a corresponding
seroreactive neoglycoprotein. Infect. Immun. 57, 322-330.
[9] Devadoss, P., Klegerman, M.E. and Groves, M.J. (1991)
Surface morphology of Mycobacterium bouis BCG: relation
to mechanisms of cellular aggregation. Microbios 65, 11 l-
125.
[lo] Kellenberger, E., Ryter, A. and Sechaud, .I. (1958) Electron
microscope study of DNA-containing plasms. II. Vegetative
and mature phage DNA as compared with normal bacterial
nucleoids in different physiological states, J. Biophys.
B&hem. Cytol. 4, 671-678.
[ill Luft, J.H. (1961) Improvements in epoxy resin embedding
methods. J. Biophys. Biochem. Cytol. 9, 409-414.
1121 Watson, M.L. (1958) Staining of tissue sections for electron
microscopy with heavy metals. J. Biophys. Biochem. Cytol.
4,475-478.
[13] Fahmy, A. (1967) An extemporaneous lead citrate stain for
electron microscopy. Proc. 25th Ann. Conf. Electron Mi-
crosc. Sot. Am., pp. 148-149.
[14] Devadoss, P.O., Groves, M.J. and Klegerman, M.E. (1991)
Electron microscopy of the integument secreted around cells
of BCG vaccine, Tice Thl substrain. Biomed. Lett. 46, 8 l-90.
[15] Klegerman, M.E. and Groves, M.J. (1992) Process for the
I161
r171
[IsI
[191
PO1
Pll
enzymatic dispersal of Mycobacterium bock BCG. U.S.
Patent No. 5 091308.
Wang, R., Klegerman, M.E. and Groves, M.J. (1996) Analy-
sis of antineoplastic polysaccharides from Mycobacterium bovis BCG vaccine by high performance anion exchange
chromatography with pulsed amperometric detection. J. Liq.
Chrom. Rel. Technol., in press.
Rosenthal, S.R., Kenniebrew, A.H. and Raisys, N. (1962) A
modified semi-solid medium and technique for determining
viability of BCG. Acta Tuberc. Pneumol. Stand. 42, 167-
172.
Shi, M., Klegerman, M.E. and Groves, M.J. (1989) Viability
of freeze-dried Tice’“-substrain BCG by bioluminescent
measurement of adenosine triphosphate. Microbios 59, 145-
155.
Lou, Y., Klegerman, M.E., Muhammad, AS., Dai, X. and
Groves, M.J. (1994) Initial characterization of an antineoplas-
tic, polysaccharide-rich extract of Mycobacterium bouis BCG, Tice@ substrain. Anticancer Res. 14, 1469-1476.
Kristensen, S., Tian, Y., Klegerman, M.E. and Groves, M.J.
(1992) Origins of BCG surface charge: effect of ionic strength
and chemical modifications on zeta potential of Mycobac- terium bovis BCG, Tice’” substrain, cells. Microbios 70,
185-198.
Dixon, M. and Webb, E.C. (1964) Enzymes, 2nd ed., pp.
226-239. Academic Press, New York.
[22] Draper, P. and Rees. R.J.W. (1973) The nature of the elec-
tron-transparent zone that surrounds Mycobacterium leprae- mu&m inside host cells. J. Gen. Microbial. 77, 79-87.
[23] Rastogi, N. and Hellio, R. (1990) Evidence that the capsule
around mycobacteria grown in axenic media contains my-
cobacterial antigens: implications of the cell envelope archi-
tecture. FEMS Microbial. Lett. 70, 161-166.
[24] Paul, T.R. and Beveridge, T.J. (1990) Ultrastructure of my-
cobacterial surfaces by freeze-substitution. Zbl. Bakt. 279,
450-457.
[25] Devadoss, P.O., Groves, M.J. and Klegerman, M.E. (1990)
Transmission electron microscopy of the mature and coc-
coidal forms of Mycobacietium bovis found in Tice” sub-
strain BCG vaccine. Microbios Lett. 45, 7-21.
[26] Chapman, G.B., Hanks, J.H. and Wallace, J.H. (1958) Elec-
tron microscope study of the disposition and fine structure of
Mycobacterium lepraemurium in mouse spleen. J. Bacterial.
77, 205-211.
[27] Imaeda, T., Convit, J. and Lapenta, P. (1964) Electron
microscopic study of borderline leprosy. Int. J. Leprosy 31, 389-417.
[28] Wietzerbin-Falszpan, J., Das, B.C., Gros, C., Petit, J.F. Led-
erer E (1973) The amino acids of the cell wall of Mycobac- terium tuberculosis var. bovis, strain BCG. Presence of a
poly(L-glutamic acid). Eur. J. Biochem. 32, 525-532.
[29] Phiet, P.H., Wietzerbin, J., Zissman, E., Petit, J.F. and Lederer, E. (1976) Analysis of the cell wall of five strains of
Mycobacferium tuberculosis BCG and of an attenuated hu-
man strain, W 115. Infect. Immun. 13, 677-681.
[30] Ortalo-Magnt, A., DuPont, M.A., Lemassu, A., Andersen,
A.B., Gounon P. and Daff6, M. (1995) Molecular composi-

222 M.E. Klegerman et al. / FEMS Immunology and Medical Microbiology 15 (1996) 213-222
tion of the outermost capsular material of the tubercle bacil-
lus. Microbiology 141, 1609-1620.
[31] Lemassu, A. and Daff6, M. (1994) Structural features of the
exocellular polysaccharides of Mycobacferium tuberculosis. B&hem. .I. 297, 351-357.
[32] Wang, R., Klegerman, M.E., Marsden, I., Sinnott, M. and
Groves, M.J. (1995) An anti-neoplastic glycan isolated from
Mycobacterium bouis (BCG vaccine). B&hem. J. 3 11, 867-
872.
[33] Paul, T.R. and Beveridge, T.L. (1994) Preservation of sur-
face lipids and determination of ultrastructure of Mycobac- terium kansasii by freeze-substitution. Infect. Immun. 62,
1542-1550.
[34] Ortalo-Magne, A., Lemassu, A., Laneelle, M.A., Bardou, F.,
Silve, G., Gounon, P., Marchal, G. and Daffe, M. (1996)
Identification of the surface-exposed lipids on the cell en-
velopes of Mycobacferium tuberculosis and other mycobac-
terial species. J. Bacterial. 178, 456-461.
Copyright © 2022 FDOKUMEN