





Mycobacterial Gene cuvA Is Required for Optimal Nutrient Utilization and Virulence
43
1 The mycobacterial gene cuvA/Rv1422 is required for optimal nutrient utilization and 1 virulence 2 Mushtaq Mir 1*+ , Sladjana Prisic 1+ , Choong-Min Kang 1** , Shichun Lun 2 , Haidan Guo 2 , Jeffrey P. 3 Murry 3*** , Eric J. Rubin 3 , Robert N. Husson 1# 4 5 1 Division of Infectious Diseases, Children’s Hospital, Harvard Medical School, Boston, MA, 6 02115 7 2 Center for Tuberculosis Research, Johns Hopkins University School of Medicine, Baltimore, 8 MD, 21231 9 3 Department of Immunology and Infectious Diseases, Harvard School of Public Health, Boston, 10 MA 02115 11 *Current address: Division of Genetics, Wadsworth Center, Center for Medical Science, New 12 York State Department of Health, Albany, NY 12201 13 **Current address: Department of Biological Sciences, California State University Stanislaus, 14 Turlock, CA 95382 15 ***Current address: Gilead Sciences, 333 Lakeside Drive, Foster City, CA 94404 16 + These authors contributed equally to this work. 17 Running title: Mycobacterial cuvA: nutrient utilization and virulence 18 Key words: Mycobacterium, cholesterol, bacterial cell pole, peptidoglycan synthesis, virulence, 19 cell wall, Rv1422 20 #Corresponding author information: 21 Robert N. Husson 22 Division of Infectious Diseases 23 Children’s Hospital Boston 24 300 Longwood Ave. 25 Boston, MA 02115 26 Tel 617-919-2900/Fax 617-730-0254 27 [email protected] 28 IAI Accepts, published online ahead of print on 21 July 2014 Infect. Immun. doi:10.1128/IAI.02207-14 Copyright © 2014, American Society for Microbiology. All Rights Reserved. on June 13, 2016 by guest http://iai.asm.org/ Downloaded from
Transcript of Mycobacterial Gene cuvA Is Required for Optimal Nutrient Utilization and Virulence

1
The mycobacterial gene cuvA/Rv1422 is required for optimal nutrient utilization and 1
virulence 2
Mushtaq Mir1*+, Sladjana Prisic1+, Choong-Min Kang1**, Shichun Lun2, Haidan Guo2, Jeffrey P. 3
Murry3***, Eric J. Rubin3, Robert N. Husson1# 4
5
1Division of Infectious Diseases, Children’s Hospital, Harvard Medical School, Boston, MA, 6
02115 7
2Center for Tuberculosis Research, Johns Hopkins University School of Medicine, Baltimore, 8
MD, 21231 9
3Department of Immunology and Infectious Diseases, Harvard School of Public Health, Boston, 10
MA 02115 11
*Current address: Division of Genetics, Wadsworth Center, Center for Medical Science, New 12
York State Department of Health, Albany, NY 12201 13
**Current address: Department of Biological Sciences, California State University Stanislaus, 14
Turlock, CA 95382 15
***Current address: Gilead Sciences, 333 Lakeside Drive, Foster City, CA 94404 16
+These authors contributed equally to this work. 17
Running title: Mycobacterial cuvA: nutrient utilization and virulence 18
Key words: Mycobacterium, cholesterol, bacterial cell pole, peptidoglycan synthesis, virulence, 19
cell wall, Rv1422 20
#Corresponding author information: 21 Robert N. Husson 22 Division of Infectious Diseases 23 Children’s Hospital Boston 24 300 Longwood Ave. 25 Boston, MA 02115 26 Tel 617-919-2900/Fax 617-730-0254 27 [email protected] 28
IAI Accepts, published online ahead of print on 21 July 2014Infect. Immun. doi:10.1128/IAI.02207-14Copyright © 2014, American Society for Microbiology. All Rights Reserved.
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

2
Abstract 29
To persist and cause disease in the host, Mycobacterium tuberculosis must adapt to its 30
environment during infection. These adaptations include changes in nutrient utilization and 31
alterations in growth rate. M tuberculosis Rv1422 is a conserved gene of unknown function that 32
was found in a genetic screen to interact with the mce4 cholesterol uptake locus. Rv1422 33
protein is phosphorylated by the M. tuberculosis Ser/Thr kinases PknA and PknB, which 34
regulate cell growth and cell wall synthesis. Bacillus subtilis strains lacking the Rv1422 35
homologue yvcK grow poorly on several carbon sources, and yvcK is required for proper 36
localization of peptidoglycan synthesis. Here, we show that M. smegmatis and M. tuberculosis 37
strains lacking Rv1422 have growth defects in minimal medium containing limiting amounts of 38
several different carbon sources. These strains also have morphologic abnormalities, including 39
shortened and bulging cells, findings that suggest a cell wall defect. In both mycobacterial 40
species, Rv1422 localizes uniquely to the growing cell pole, the site of peptidoglycan synthesis 41
in mycobacteria. M. tuberculosis ΔRv1422 is markedly attenuated for virulence in a mouse 42
infection model, where it elicits decreased inflammation in the lungs and shows impaired 43
bacterial persistence. These findings lead us to name this gene cuvA (carbon utilization and 44
virulence protein A) and suggest a model in which deletion of cuvA leads to changes in nutrient 45
uptake and/or metabolism that affect cell wall structure, morphology and virulence. Its role in 46
virulence suggests that CuvA may be a useful target for novel inhibitors of M. tuberculosis 47
during infection. 48
49
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

3
Introduction 50
Regulated pathways that allow physiologic adaptations in response to changes in the 51
environment are critical for the growth and survival of all bacteria. In the case of the human 52
pathogen Mycobacterium tuberculosis, where infection can persist for decades and cause 53
several different disease manifestations, the bacterium must adapt to multiple environments 54
during the prolonged course of infection. These environments include intracellular milieus within 55
host phagocytic cells such as neutrophils, macrophages and dendritic cells that may be part of 56
organized granulomas, as well as in extracellular environments such as occur in caseating 57
granulomas. In addition to mechanisms to defend against and modulate host antibacterial 58
activities, a critical adaptation of M. tuberculosis during infection is utilization of host lipids as a 59
primary carbon source (1-3). Another key bacterial adaptation during M. tuberculosis infection is 60
regulation of replication, which changes from rapid growth during the first few weeks of infection, 61
to markedly reduced bacterial replication following the onset of adaptive immunity (3, 4). 62
Gaining insight into bacterial adaptive mechanisms is critical for understanding tuberculosis 63
pathogenesis and may lead to novel therapeutic approaches that target these bacterial adaptive 64
processes. 65
M. tuberculosis Rv1422 encodes a protein of unknown function that has homologues in 66
other mycobacteria and across a broad range of bacterial phyla. Our interest in this protein as 67
potentially having an important role in bacterial adaptation during infection was motivated by 68
three observations. One is our previous finding that it is phosphorylated in M. tuberculosis cells 69
by the Ser/Thr protein kinases PknA and PknB. These kinases are essential for growth and 70
regulate cell division, peptidoglycan (PGN) synthesis and cell morphology (5-9), suggesting that 71
Rv1422 may play a role in one or more of these processes. Second, Rv1422 was identified in a 72
genome-wide transposon screen for genetic interactions with the mce1 and mce4 loci. The 73
mce4 locus encodes a cholesterol uptake system (2, 10) and the mce1 locus has been 74
implicated in the transport of other lipids (11), suggesting that the Rv1422 protein may function 75
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

4
in utilization of cholesterol or other lipids. In addition, an M. tuberculosis strain with transposon 76
insertion in Rv1422 was defective for replication in mouse spleens in an 8-week infection model, 77
suggesting that Rv1422 might also be important in M. tuberculosis virulence (12). 78
The orthologue of Rv1422 has been investigated in the model organism Bacillus subtilis, 79
where a null mutant of this gene, yvcK, was found to have partial growth defects on media 80
containing several different carbon sources, including tricarboxylic acid cycle intermediates (13). 81
On transfer to medium containing carbon sources in which this organism grew poorly, bulging 82
and lysis of cells were seen, indicating perturbation of PGN synthesis. Subsequently, YvcK was 83
found to be distributed in a circumferential pattern around the long axis of the cell in B. subtilis, 84
paralleling the distribution of PGN synthesis (14). Overexpression of MreB, a filament-forming 85
protein that is similarly distributed around the circumference of the cell (15), was shown to 86
suppress the carbon-source specific growth and morphology defects of the yvcK null mutant. In 87
addition, the PGN synthesis protein PBP1 was found to be mis-localized in the ΔyvcK strain in a 88
carbon-source-dependent manner (14). Thus, though its mechanism of action is not known, 89
YvcK appears to be required for proper localization of PGN synthesis and production of a 90
normal cell wall in B. subtilis. 91
These data from M. tuberculosis and B. subtilis led us to ask whether Rv1422 plays a 92
role in carbon source utilization, cell growth, and virulence in mycobacteria. Here we identify 93
growth defects on multiple carbon sources of deletion mutants in M. tuberculosis Rv1422 and 94
the M. smegmatis homologue MSMEG_3080 compared to wild type. Because of the importance 95
of cholesterol as a carbon source for M. tuberculosis during infection (2, 16, 17), we used 96
cholesterol as a representative carbon source for additional investigation of the function of 97
Rv1422. When grown on cholesterol, both the M. smegmatis and M. tuberculosis mutant strains 98
have morphologic abnormalities including bulging and shortened cell length, indicating a role for 99
Rv1422 in cell wall synthesis. We also show striking localization of Rv1422-gfp fusions 100
exclusively to the growing cell pole, the site of PGN synthesis in mycobacteria (8) and 101
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

5
demonstrate that the M.smegmatis deletion strain are hypersusceptbible to several β-lactam 102
antibiotics when grown on cholesterol, findings that link Rv1422 to cell growth and cell wall 103
synthesis. Consistent with these in vitro phenotypes, the virulence of the M. tuberculosis mutant 104
is markedly attenuated in a mouse infection model. Based on these results, we have named 105
this gene cuvA (carbon-source utilization and virulence protein A). 106
Materials and Methods 107
Strains, media, plasmids and primers 108
M. tuberculosis H37Rv and M. smegmatis mc2-155 (18) were used as wild type strains 109
and are the parental strains in which mutants were made. E. coli TOP10 (Invitrogen) was used 110
for cloning and was grown in LB broth. For routine growth, M. tuberculosis or M. smegmatis 111
were grown at 37o C in Middlebrook 7H9 liquid medium (Difco) supplemented with 0.5% 112
albumin, 0.2% glucose, 0.085% NaCl, 0.2% Glycerol and 0.05% Tween 80 (7H9-ADC-Tw). 113
Kanamycin (20 μg ml-1) or hygromycin (50 μg ml-1) was added to liquid or agar medium when 114
appropriate. Details of primers, plasmids and strains are shown in Table S1. 115
For growth of M. smegmatis on different carbon sources in liquid medium, cells were 116
grown in minimal medium (MM) (1 g/L KH2PO4, 2.5 g/L Na2HPO4, 0.5 g/liter asparagine, 0.5 g/L 117
MgSO4x7H2O, 0.5 mg/L CaCl2, 0.1 mg/L ZnSO4, 50 mg/L ferric ammonium citrate and 0.05% 118
Tyloxapol) (2), containing 0.01% (wt/vol) of glucose, glycerol, cholesterol, propionate, gluconate 119
or citrate. MM plus 0.2% glucose was used as a positive control for maximal growth. For 120
measuring growth on solid medium, cells grown in Middlebrook 7H9 liquid medium to an OD600 121
of 0.5 were washed in PBS containing 0.05% Tyloxapol and 10-fold serial dilutions in PBS were 122
spotted on agar plates containing MM as described above (without tyloxapol) plus 0.01% of the 123
carbon sources. MM plates containing 0.2% glucose were included as a control. Plates were 124
incubated at 37o C and photographed after one month at 37oC for M. tuberculosis and after 2 125
days for M. smegmatis. 126
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

6
For subcellular localization, CuvA was expressed as a carboxy-terminal GFP fusion. 127
cuvA was PCR-amplified from genomic DNA of M. tuberculosis H37RV using primers 128
Rv1422AE and Rv1422rPac1. gfp was PCR-amplified from pTracerCMV (Invitrogen) using 129
primers GFP3PacI and GFP4XK. Overlap PCR was then carried out using primers Rv1422AE 130
and GFP4XK giving rise to the fusion PCR product Rv1422-gfp. The Rv1422-gfp PCR product 131
was digested with AseI and XbaI (the AseI site in the Rv1422 gene was eliminated by overlap 132
PCR mutagenesis to create a silent mutation) and cloned into the integrating vector pMV306AC 133
3’ of the acetamidase promoter to obtain pMV306Ac-Rv1422-gfp (pMV306Ac-cuvA-gfp). 134
For localization of M. smegmatis PBP1 (encoded by MSMEG_6900), a C-terminal 135
MSMEG_6900-RFP fusion protein was expressed. The MSMEG6900 gene was amplified by 136
PCR from M. smegmatis genomic DNA using primers MSMEG6900-F and MSMEG6900-R. 137
Similarly rfp was PCR-amplified using primers Rfp_F and Rfp_R. Overlap PCR was then carried 138
out using the primers MSMEG6900-F and Rfp_R. The resulting fusion PCR product was cloned 139
into the NdeI and XbaI sites in pMV261Ac 3’ of the acetamidase promoter to obtain pMV261Ac-140
MSMEG6900-rfp. All recombinant clones used in this study were sequenced to rule out any 141
mutations. 142
Generation of M. smegmatis and M. tuberculosis cuvA deletion strains 143
To delete cuvA (MSMEG_3080) in M. smegmatis, a 1008 bp region (L-arm) 5’ to 144
MSMEG_3080 including 6 codons of MSMEG_3080 was PCR-amplified using primers 145
MSMEG3080-cond-1 and MSMEG3080-cond-2. Similarly a 1040 bp region (R-arm) 3’ to 146
MSMEG_3080 including 100 bp of the MSMEG_3080 gene was PCR-amplified using primers 147
MSMEG3080-cond-3 and MSMEG3080-cond-4. Overlap PCR was then carried out with primers 148
MSMEG3080-cond-1 and MSMEG3080-cond-4. The PCR product contained a PacI site 149
between the arms and was cloned into pRH1351 (19). A hygromycin resistance cassette was 150
introduced at the PacI site and the targeting construct thus obtained was electroporated into M. 151
smegmatis mc2155. Candidate deletion strains were obtained using a two-step double 152
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

7
counterselection method, as previously described (19, 20). PCR of genomic DNA from 153
candidate mutant strains, using primers annealing to flanking regions of MSMEG_3080, yielded 154
the expected 1.5 kb PCR product, confirming the replacement of MSMEG_3080 (cuvA) in the 155
genome by the hygromycin cassette (Fig. S1). For complementation of the M. smegmatis cuvA 156
deletion strain, cuvA was PCR-amplified using primers MSMEG3080-int-1 and MSMEG3080-157
int-2 and cloned under control of the tet promoter in pMind (21). This complemented strain was 158
grown in the presence of 5 ng/ml tetracycline to induce expression of cuvA. 159
The cuvA (Rv1422) gene was deleted in M. tuberculosis H37Rv genome using similar 160
methods. One kb regions 5’ and 3’ to Rv1422 were PCR-amplified and ligated to the 5’ and 3’ 161
ends respectively of a hygromycin-chloramphenicol cassette. The ligated DNA fragment was 162
introduced into a temperature-sensitive mycobacterial suicide vector harboring sacB and xylE 163
genes. The targeting vector was transformed into wild type M. tuberculosis H37RV and 164
candidate deletion strains were obtained by counterselection on sucrose-containing plates at 165
38oC. Deletion of cuvA was confirmed by PCR of genomic DNA using primers annealing to 166
flanking regions of the gene. The expected size of 2.1 kb resulting from the replacement of cuvA 167
by the hygromycin-chloramphenicol cassette confirmed the deletion of the gene (Fig. S1). The 168
complementing construct was obtained by cloning cuvA downstream of the hsp70 promoter in 169
the integrating vector pJEB402 (22). 170
Microplate Alamar Blue assay (MABA) for growth 171
M. smegmatis strains were shaken at 37oC in 7H9-ADC-Tw (without glycerol) until they reached 172
OD600 ~0.5. The cells were spun down, washed twice in PBS-Tx (phosphate buffered saline, 173
0.05% Tyloxapol) and diluted to calculated OD600 = 0.001 in MM alone or with added 0.2% 174
glucose, or each of the following added to 0.01%: glucose, cholesterol, glycerol, gluconate, 175
citrate or propionate. Tetracycline (5 ng/mL) was present in all cultures. 200 µl of each culture 176
was placed in a 96-well clear bottom plate and 20 µl filtered Alamar Blue (Life Technologies) 177
was added. Fluorescence (Excitation/Emission = 550nm/590nm) was measured every hour 178
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

8
from the bottom of the well in a plate reader (Tecan) with incubation at 37oC. The exponential 179
portion of the growth curves were analyzed by non-linear curve fitting to an exponential growth 180
model and the rate constants were compared by an extra sum of squares F test, using 181
Graphpad Prism 5.0 software. 182
MABA susceptibility determination 183
M. smegmatis cultures were prepared as for the MABA growth curves described above, 184
except that they were diluted to a calculated of OD600 of 0.005 in MM containing 0.01% 185
cholesterol. Glucose was added to half of the cultures, to obtain cultures with 0.01% cholesterol 186
plus 0.2% glucose; the other cultures contained 0.01% cholesterol only. To half of the 187
cholesterol alone cultures and half of the cholesterol plus glucose cultures, clavulanate was 188
added to a final concentration of 2.5 µg/ml. For each antibiotic tested, two-fold dilutions were 189
made and M. smegmatis cultures (190 µl) and antibiotics (10 µl) were mixed in clear 96-well 190
plates. After overnight (for glucose + cholesterol) or 2 days (for cholesterol only) incubation, 50 191
µl of 0.5X Alamar Blue reagent diluted in 10% Tween 80 was added. After overnight incubation 192
fluorescence (Ex/Em = 550nm/590nm) was measured. Growth inhibition was calculated using 193
the following formula: 1 100, where Fmin is the 194
fluorescence from wells without growth and Fmax is the fluorescence from control wells that did 195
not contain any antibiotic. The MIC90 was defined as the concentration of antibiotic that caused 196
90% growth inhibition (23). 197
M. tuberculosis H37Rv was tested for antibiotic susceptibility in a similar way, except 198
that cultures were pre-incubated in PBS-Tx for 2 days and were diluted to OD600 0.05, and the 199
clavulanate concentration was 7.5µg/ml. Plates were incubated for one week before Alamar 200
blue was added and analyzed as described above. The final concentration ranges of the 201
antibiotics tested were: rifampin (M. tuberculosis), 0.003-0.64 μg/ml; rifampin (M. smegmatis), 202
0.3-80 μg/ml; levofloxacin, 0.01-2.56 μg/ml; ampicillin, 4-1000 μg/ml; cefotaxime, 4-1000 μg/ml; 203
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

9
cephalothin, 2-500 μg/ml; meropenem, 0.2-50 μg/ml. For both M. smegmatis and M. 204
tuberculosis, each antibiotic was tested in at least three biological replicates with technical 205
duplicates for every experiment. MICs between strains were compared using the t-test in 206
Microsoft Excel. 207
Microscopy and fluorescent vancomycin staining 208
For cellular localization of CuvA the recombinant clone pMV306-pacet-Rv1422-gfp was 209
transformed into M. smegmatis strain mc2155 and in to an M. tuberculosis leucine/pantothenate 210
double auxotrophic strain (18, 24). The pMV306-pacet-gfp expressing GFP alone was 211
transformed into M. smegmatis as a control. The M. smegmatis strains were grown to an OD600 212
of 0.1 - 0.2 in 7H9 broth, induced with 0.2% acetamide for 6-8 hrs and visualized by microscopy. 213
The M. tuberculosis strains were grown to an OD600 of 0.5 and inoculated onto a layer of nutrient 214
agarose medium on microscope slides (agarose pads) made with Middlebrook 7H9 medium 215
containing glucose and 0.2% acetamide. Cells on these agarose pads were observed at 24 216
hours after growth. 217
For staining sites of peptidoglycan synthesis, Vancomycin-Alexa 568 (Molecular Probes) 218
was prepared and used as previously described (8). Vancomycin-Alexa568 was added to 219
cultures at OD600 of 0.1 - 0.2 to achieve a final concentration of 5 μg/ml and growth was 220
continued further for 2.5 h in the dark. The cells were washed, transferred onto a glass slide, air-221
dried, and a cover slip was mounted using Prolong Gold anti-fade reagent (Invitrogen). Cells 222
were observed using Zeiss AxioImager.Z2 microscope with 63x differential interference contrast 223
oil immersion objective. For fluorescence, green and red fluorescence filters were used. Images 224
were captured by a CoolSNAP HQ2 camera (Photometrics), acquired with AxiVision 4.8 225
software and processed by Adobe Photoshop CS5. 226
For examination of cell morphology, cells were grown in the indicated medium, pelleted 227
and fixed in 4% paraformaldehyde prepared in PBS for 2 hours. Ammonium chloride was then 228
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

10
added to a final concentration of 50 mM, cells were pelleted and then resuspended in PBS. 5 �l 229
of the cell suspension was spotted on a glass slide, which was air-dried, mounted with a 230
coverslip and observed using either a 63x or 100x differential interference contrast oil immersion 231
objective. Images were processed using Adobe Photoshop CS5. Measurement of cell length 232
was performed using ImageJ software and statistical analysis of cell length was performed with 233
one-way ANOVA and the nonparametric Kruskal-Wallis test using Graphpad Prism software. 234
For examination of live M. smegmatis, cells were grown overnight on agarose pads on glass 235
microscope slides. The cells were then directly observed on the agarose pads. 236
Transposon Mutagenesis 237
To identify mutations that suppress the cholesterol growth defect of the ΔcuvA strain of 238
M. tuberculosis, we performed transposon mutagenesis using the φMycoMarT7 phagemid (25, 239
26). Preparation of the phage stock, its titration and subsequent transduction of cuvA deletion 240
strain was carried out as described (27). Approximately 120,000 transductants were plated on 241
cholesterol-agar plates containing 20 μg/ml kanamycin and incubated at 37oC. After 5 weeks, 242
60 colonies were streaked on cholesterol-agar plates, of which 20 showed substantial growth. 243
Genomic DNA was isolated and three restriction enzymes: BamHI, SphI and SacII were used 244
individually to completely digest the genomic DNA. The recognition sites of these enzymes are 245
absent in the transposon. The fragmented DNA was purified and treated with T4 DNA ligase to 246
circularize the fragments. PCR amplification was then carried out using outward primers 247
annealing to the ends of transposon. The PCR products were analyzed by agarose gel 248
electrophoresis and strong intensity bands were extracted and sequenced to identify the site of 249
transposon insertion. 250
Mouse Infection experiments 251
Mouse infection experiments were performed at the NIAID-sponsored Tuberculosis 252
Animal Research and Gene Evaluation Task Force (TARGET). Mouse infection protocols were 253
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

11
approved by the Animal Care and Use Committee at Johns Hopkins School of Medicine, 254
Baltimore, MD and all experiments involving mice were carried out according to these protocols. 255
To determine the effect of cuvA deletion on growth of M. tuberculosis in vivo, wild type 256
H37Rv, the cuvA deletion strain (RH 478) and the ΔcuvA complemented strain (RH480), were 257
used to perform aerosol infection of BALB/c mice. Prior to infection, all M. tuberculosis stains 258
were confirmed to produce phthiocerol dimycolate and to have positive neutral red staining 259
(data not shown). For time to death experiments strains were grown to mid-log phase and 260
diluted to OD600 of 0.2 in 10 ml of 7H9 Middlebrook liquid medium supplemented with OADC. 16 261
BALB/c mice per strain were infected for 30 min in a Glas-col aerosol machine. At 1 day post 262
infection 3 mice per strain were sacrificed. Lungs from each mouse were homogenized and 263
dilutions of lung homogenate were plated on 7H11 antibiotic selective plates. CFU were 264
recorded after 3-4 weeks of incubation at 37oC. The remaining 12 mice infected with each strain 265
were monitored to record the days to death or severe illness for each mouse. Statistical analysis 266
of survival was performed with the Log Rank (Mantel-Cox) test using GraphPad Prism 5.0 267
software. 268
To determine the CFU of each strain in lung and spleen following infection, the three 269
strains were grown to an OD600 of 0.05 to 0.1 in Middlebrook 7H9 liquid medium supplemented 270
with OADC. 24 BALB/c mice per strain were infected as described above. Four mice per strain 271
were sacrificed at day 1, 14, 28, 56, 84 and 112 following infection. The lung and spleen of 272
each mouse was weighed and gross pathology pictures were taken. 3/4 of each lung and 273
spleen was homogenized and dilutions were plated on 7H11 selective plates. The CFU were 274
counted after 3-4 weeks at 37oC. The remaining 1/4 of each lung and spleen was stained with 275
hematoxylin and eosin for histopathology. CFU in lungs, lung weight and spleen weight for each 276
strain were compared at the indicated time points by one-way ANOVA with Tukey’s post-test 277
using GraphPad Prism 5.0 software 278
Results 279
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

12
CuvA is required for optimal growth and normal cell morphology on several carbon 280
sources 281
Mutant strains containing cuvA deletions were constructed by allelic exchange in both 282
the pathogen M. tuberculosis and the rapid-growing non-pathogen M. smegmatis (Fig. S1). A 283
complemented M. tuberculosis ΔcuvA strain was constructed by expressing cuvA under control 284
of the hsp70 promoter in the vector pJEB402 (22), to allow stable expression during prolonged 285
incubation in vitro and during infection experiments. The same construct was used to 286
complement the M. smegmatis mutant with M. tuberculosis cuvA. To complement the M. 287
smegmatis mutant with the M. smegmatis cuvA homologue MSMEG_3080, this gene was 288
expressed under control of a Tet repressor-regulated promoter, to allow inducible expression of 289
this gene (21). 290
When grown in Middlebrook 7H9 liquid medium containing 0.2% glucose, M. 291
tuberculosis ΔcuvA (Fig. 1A) and M. smegmatis ΔcuvA (Fig. S4A) grew at a similar rate and to 292
the same final density as the wild type parental strains. Based on genetic data linking Rv1422 to 293
the Mce4 cholesterol transport system (28), we initially examined growth of the deletion strains 294
in minimal medium (MM) with cholesterol added as a carbon source. In contrast to the results 295
obtained with glucose-containing 7H9 medium, we observed altered growth phenotypes for both 296
the M. smegmatis and M. tuberculosis ΔcuvA strains when they were grown on MM plus 297
cholesterol, though these growth defects were not identical in the two species. 298
For M. tuberculosis, our attempts to compare growth by A600 measurements of wild type, 299
ΔcuvA and complemented strains in liquid MM containing 0.01% cholesterol were limited by 300
marked clumping of all strains and by the medium being cloudy and forming precipitates. We 301
therefore used an alternative measure of growth, the microplate Alamar Blue assay (MABA), 302
which measures dye reduction in metabolically active growing cells. Data from this assay did not 303
show a clear growth defect of M. tuberculosis ΔcuvA in liquid medium containing cholesterol, 304
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

13
although all M. tuberculosis strains grew poorly and required a high inoculum to grow in this 305
medium, limiting interpretation of this result (Fig. S2). On solid medium, however, the M. 306
tuberculosis ΔcuvA strain had a clear growth defect compared to wild type (Fig. 1B). When 307
grown in Middlebrook 7H9 medium, washed and spotted onto agar plates, the wild type strain 308
grew and formed colonies on MM agar with either 0.01% cholesterol or 0.2% glucose, although 309
the growth on MM plus cholesterol was slower. The ΔcuvA strain grew more slowly than wild 310
type on MM plus 0.2% glucose and showed minimal growth on cholesterol, even after prolonged 311
incubation. These growth phenotypes were complemented by expression of cuvA in the M. 312
tuberculosis mutant. 313
The observation that the B. subtilis yvcK deletion had growth defects on multiple carbon 314
sources, led us to compare the growth of wild type M. tuberculosis to the cuvA deletion strain on 315
solid MM containing several other carbon sources. Each compound was added to a final 316
concentration of 0.01%, the concentration of cholesterol that has been used in mycobacterial 317
growth media because of its low aqueous solubility (2, 17). As shown in Figure 2, the M. 318
tuberculosis cuvA mutant strain grew much more slowly than wild type on each of these media, 319
as indicated by less dense growth and absence of visible growth at lower dilutions. In each case 320
the complemented strain showed growth similar to, though slightly less than that of the wild 321
type. These data indicate that the growth defect of the ΔcuvA strain is not specific, but rather is 322
a general defect in utilization of these several different carbon sources. 323
Because CuvA was shown to be phosphorylated in M. tuberculosis (5, 29), to determine 324
whether phosphorylation of CuvA is important for its function in nutrient utilization, 325
complementing constructs encoding CuvA in which the phosphoacceptor Thr residue was 326
replaced with either a non-phosphorylatable (Thr325Ala) or a phospho-mimetic (Thr325Glu) 327
amino acid were tested. Both of these mutant alleles also fully complemented the cholesterol 328
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

14
growth defect (Fig. S3) suggesting that phosphorylation of Thr325 is not required for CuvA 329
function in cholesterol utilization under the conditions examined in these experiments. 330
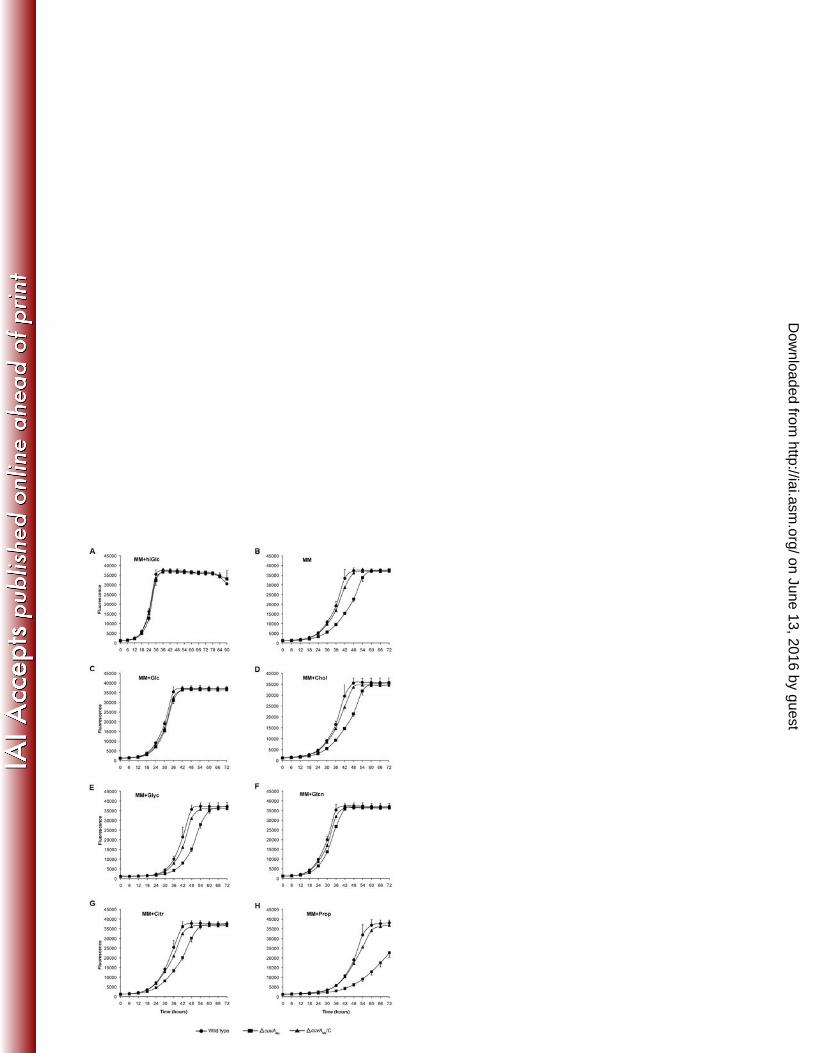
We then examined growth of the M. smegmatis ΔcuvA strain. This strain had substantial, 331
statistically significant growth defects observed using the MABA assay when grown in liquid MM 332
alone or with several carbon sources present at 0.01%, though growth was similar to wild type 333
in MM plus glucose or gluconate (Fig. 3). The mutant growth defects were well-complemented 334
in each case by MSMEG_3080 expressed in trans. Addition of 0.2% glucose both increased the 335
growth rate and eliminated the difference between the wild type and mutant strains grown on 336
cholesterol or propionate (data not shown), indicating that the growth defect is not likely to be 337
due to toxic effect of defective cholesterol catabolism as has been observed in other mutants 338
that cannot utilize cholesterol (16). When grown on solid medium, the ΔcuvA grew less than wild 339
type on MM alone or with added cholesterol, but grew as well as wild type on 0.2 or 0.01% 340
glucose (Fig S4B). These data show that the growth defects of M. smegmatis ΔcuvA, like that of 341
M. tuberculosis, are not specific. 342
To further investigate the growth defects of the cuvA mutant we undertook a series of 343
additional experiments. We first examined cell shape and observed that the growth defect of M. 344
smegmatis ΔcuvA grown in cholesterol liquid MM was accompanied by altered cell morphology, 345
with cells that were shorter than wild type and appeared to have thickened, wider cell poles (Fig 346
4A). Measurement of the cholesterol-grown cells demonstrated that the difference in cell length 347
was highly significant (Fig. 4B). These cell length and morphology defects were fully 348
complemented by the wild type cuvAMs allele. When M. smegmatis ΔcuvA was grown on solid 349
MM plus cholesterol, cell morphology was also affected, with many cells showing asymmetric 350
bulging (Fig. 4C). This phenotype is very similar to the ΔyvcK morphology phenotype in B. 351
subtilis strains grown on gluconate, where the bifunctional PGN synthesis enzyme PBP1 is mis-352
localized (14). This M. smegmatis ΔcuvA phenotype is also very similar to the morphologic 353
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

15
abnormalities seen in a PBP1 depletion strain of M. smegmatis, where PGN synthesis is directly 354
affected (30). 355
Because of the minimal growth of M. tuberculosis ΔcuvA on solid MM containing 356
cholesterol, there were too few cells to identify a morphologic phenotype of the ΔcuvA strain on 357
this medium. When ΔcuvA were grown to late stationary phase in liquid MM with 0.01% 358
cholesterol, however, we observed a bulging phenotype similar to that observed for M. 359
smegmatis (Fig. 4D). This phenotype was seen in 42% of ΔcuvA cells versus 11% of wild type 360
(P<0.0001 with over 200 cells analyzed per strain) and 12% of complemented cells. As was 361
seen for M. smegmatis, this M. tuberculosis phenotype was not seen for cells grown on MM plus 362
glucose. These morphologic phenotypes strongly suggest a defect in PGN synthesis or 363
structure in the ΔcuvA deletion strains. 364
CuvA localizes asymmetrically to the growing cell pole in mycobacteria 365
Peptidoglycan synthesis in mycobacteria and other actinomycetes is targeted to the 366
septum and asymmetrically to the growing cell pole (8, 31). The growth and morphology defects 367
we observed in the ΔcuvA strains, together with the data from B. subtilis indicating that YvcK 368
plays a role in regulating cell wall synthesis (14), led us to investigate the subcellular localization 369
of CuvA. We first determined that M. tuberculosis cuvA was a functional orthologue of the M. 370
smegmatis gene by showing that M. tuberculosis cuvA fully complements the cholesterol growth 371
defect of M. smegmatis ΔcuvA (Fig. S5). We then made a cuvAMt-gfp fusion and confirmed that 372
this construct also complemented the growth defect of the ΔcuvAMs strain in MM containing 373
0.01% cholesterol (Fig. S5). 374
Using this cuvAMt-gfp fusion, we found that CuvA-GFP strongly localizes to one pole of 375
most cells (Fig. 5), similar to the pattern we observed previously for Wag31 (DivIVA), an 376
essential protein that we showed to be required for polar localization of PGN synthesis in 377
mycobacteria (8). To determine the timing of CuvA localization and whether this protein is 378
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

16
targeted to the old or new pole, we performed time-lapse live cell microscopy of M. smegmatis 379
expressing CuvA-GFP growing on nutrient agarose containing MM with 0.01% cholesterol 380
overlaid on microscope slides. As shown in the supplemental movie, we found that CuvA-GFP 381
localized to the old pole in most cells. We also observed that cell growth occurred predominantly 382
at one pole of the mycobacterial cell as we and others have shown previously (8, 32), and that 383
CuvA consistently localized to this growing pole. This localization of CuvA to the growing cell 384
pole occurred when cells were grown on medium containing either glucose or cholesterol as the 385
carbon source. Similar to these observations in M. smegmatis, CuvA also localized to one pole 386
in M. tuberculosis (Fig. 6). Though we were not able to perform time-lapse microscopy with M. 387
tuberculosis, observation of cells that appear to have recently undergone cell division indicates 388
that CuvA also localizes to the old, actively growing cell pole in this species. A non-389
phosphorylatable form of CuvA, in which Thr325 was replaced with Ala, also localized to the cell 390
pole in M. tuberculosis (not shown). 391
Our finding that CuvA localizes to the growing cell pole led us to predict that CuvA and 392
new PGN synthesis would co-localize. Fluorescent Vancomycin (Van-Alexa568), which binds to 393
the terminal D-Ala-D-Ala of newly synthesized peptidoglycan precursors, can be used to identify 394
sites of active PGN synthesis in bacteria (31). Using Van-Alexa568, we observed that most M. 395
smegmatis cells grown in glucose-containing medium were stained predominantly at one pole, 396
consistent with our previous observations (8), though some cells had a weaker signal from the 397
other pole and a few had approximately equal staining of both poles (Fig. 5). CuvA-GFP 398
consistently localized to the pole that showed strong Van-Alexa568 staining, indicating that 399
CuvA is targeted to sites of active PGN synthesis at the elongating cell pole. Surprisingly, 400
however, whereas asymmetry of PGN synthesis was seen in nearly all cells grown in glucose, in 401
cells grown on cholesterol, strong bipolar staining with fluorescent vancomycin was seen in the 402
majority of cells (Fig. 5). Despite this bipolar Vancomycin staining, CuvA consistently localized 403
to one pole in cholesterol-grown cells, indicating that CuvA is not absolutely required for 404
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

17
localized PGN synthesis at the cell pole, but that it is consistently targeted to the old cell pole 405
when grown on either glucose or cholesterol. 406
Based on the findings in B. subtilis that the penicillin binding protein PBP1 is delocalized 407
in a yvck null mutant (14), we examined the localization of PBP1 in wild type M. smegmatis and 408
in the ΔcuvA strain. As previously reported (30), we found that a PBP1-RFP fusion protein 409
localizes to both cell poles and to the mid-cell in wild type M. smegmatis (Fig S6). In contrast to 410
the findings in B. subtilis, however, no differences were seen in PBP1-RFP localization in M. 411
smegmatis ΔcuvA compared to wild type, when grown on medium containing either glucose or 412
on cholesterol as the carbon source. Based on suppressor mutation results (see below) and its 413
identification as a probable substrate of PknB (6), we also examined the localization of PbpA. 414
As was observed with PBP1, the localization of PbpA was the same in wild type and the ΔcuvA 415
strain (data not shown). 416
417
The Ms-cuvA deletion strain is hyper-susceptible to β-lactam antibiotics and rifampin 418
Previous work had shown that M. smegmatis lacking the bifunctional penicillin binding 419
protein PBP1 is markedly more susceptible to several β-lactam antibiotics including ampicillin 420
and cephalothin (6 to >20 fold lower MIC), but shows little change in susceptibility to other 421
classes of antibiotics (33). In another study, a screen for M. smegmatis and M. tuberculosis 422
transposon mutants that are hypersusceptible to β-lactams identified several genes involved in 423
cell wall synthesis and cell division, including ponA2 (encoding PBP2), dapB and homologues of 424
DivIVA and DivIVC (34). Based on our data suggesting a role for CuvA in cell wall synthesis or 425
structure, we tested susceptibility of the M. tuberculosis and M. smegmatis deletion strains to 426
several antibiotics using the MABA assay. No significant differences in susceptibility of the M. 427
tuberculosis deletion strain compared to wild type were observed. Because of the high inoculum 428
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

18
required for M. tuberculosis to grow in MM, however, there may be small differences in 429
susceptibility between strains that we could not detect with this assay (Table S1). 430
As shown in Table 1, however, we observed strikingly increased susceptibility of M. 431
smegmatis ΔcuvA to all four β-lactams tested when the strain was grown on cholesterol. For 432
ampicillin and the first generation cephalosporin cephalothin, the addition of the β-lactamase 433
inhibitor clavulanate was required to observe this increase in susceptibility, whereas for the third 434
generation cephalosporin cefotaxime and the carbapenem meropenem, which are less 435
susceptible to hydrolysis by β-lactamases, the increased susceptibility was evident without 436
addition of clavulanate (Table 1). This increased susceptibility phenotype was reversed in the 437
complemented strain. This hypersusceptibility was also reversed in each case by the addition of 438
0.2% glucose to the culture medium. The M. smegmatis mutant was not significantly more 439
susceptible to levofloxacin, isoniazid or gentamicin, none of which act on PGN synthesis. The 440
ΔcuvA strain was, however, significantly more susceptible to the RNA polymerase inhibitor 441
rifampin. While this finding raises the possibility of an effect of cuvA on RNA polymerase, an 442
indirect effect related to an altered cell wall in the mutant, such as increased uptake or 443
decreased efflux of rifampin, may account for this difference. 444
Suppressors of the CuvA cholesterol growth phenotype map to loci involved in cell wall 445
synthesis and transmembrane transport 446
Our phenotypic and localization data suggested that CuvA functions in utilization or 447
uptake of several carbon sources and is required for normal cell wall structure under nutrient-448
limited conditions. As another approach to gaining insight into the pathways in which CuvA 449
functions, we undertook a saturating transposon mutagenesis experiment to identify 450
suppressors of the M. tuberculosis ΔcuvA cholesterol growth defect. Genes in which two or 451
more insertions were obtained are shown in Table 2. A striking finding was the isolation of 452
several transductants with insertions in pbpA and rodA. These genes are involved in PGN 453
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

19
synthesis and cell shape control and are part of the operon containing pknA and pknB, genes 454
encoding essential Ser/Thr kinases that regulate these processes (5, 6, 8, 9, 35). These 455
suppressors thus indicate a link between the ΔcuvA growth phenotype and cell wall synthesis 456
and cell morphology, and are reminiscent of the suppression of the growth and morphology 457
defects of the B. subtilis yvcK mutant by deletion of ponA, the gene encoding PBP1 (14). We 458
also obtained three independent insertions in the forward orientation at the beginning of 459
Rv2683, and one in the 5’ end of the Rv2683 coding sequence in the same orientation. The 460
location and orientation of these insertions, suggest that increased expression of this operon via 461
read-through expression from the transposon may be suppressing the ΔcuvA cholesterol growth 462
phenotype. Rv2683 is the first gene in a three gene operon that encodes a transporter that was 463
recently shown to be essential for M. tuberculosis growth on cholesterol (36). This result 464
suggests that increased expression of this transporter may be suppressing the ΔcuvA growth 465
defect. Two additional insertions disrupt the response regulator of an uncharacterized two-466
component system. 467
CuvA is required for virulence and persistence in a mouse infection model 468
In the context of data indicating that M. tuberculosis strains lacking the Mce4 cholesterol 469
transport system are attenuated during infection and a transposon mutagenesis study 470
suggesting that Rv1422 is required for bacterial replication in mice following intravenous 471
infection (12, 37), the in vitro growth defects of the ΔcuvA strains led us to compare the 472
virulence of M. tuberculosis ΔcuvA to the parental wild type H37Rv in mouse infection 473
experiments. Balb/C mice were infected by the aerosol route with wild type H37Rv, ΔcuvA and 474
ΔcuvA complemented strains. Examination of colony forming units (CFUs) present in lungs at 475
serial time points demonstrated that the wild type and ΔcuvA strains replicated at a similar rate 476
to day 14-28, after which the number of mutant bacteria declined (Fig. 7A). In contrast, the wild 477
type persisted at a stable level until day 84, after which a small decline was seen. At days 56, 478
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

20
84 and 112 the difference between wild type and the cuvA strain was highly significant 479
(P<0.0001 to P<0.005). By the final time point at 112 days there was a 200-fold difference in 480
lung CFUs between these strains. The complemented strain showed an intermediate phenotype 481
with significantly more CFU in the lung than the ΔcuvA strain P<0.0001 to P<0.01), but fewer 482
CFU than the wild type (P<0.0001 to P<0.05). This persistence defect of the M. tuberculosis 483
cuvA mutant is similar to, but more pronounced than, the phenotype observed in an mce4 484
deletion strain, where decreased CFUs were seen at later stages of infection (28). In the spleen 485
the CFUs increased for all 3 strains to day 28, after which the wild type increased compared to 486
the mutant, though this phenotype was not well-complemented (Fig. S7). 487
Gross and microscopic pathology showed striking differences in the extent of disease in 488
the lungs from mice infected with the wild type and complemented strains compared to those 489
from ΔcuvA-infected mice. The lungs of wild type and complement-infected mice showed much 490
larger and more numerous tubercles, were significantly heavier (P<0.0001 to P<0.005 at days 491
54-112 for wild type vs. cuvA) and showed much more extensive cellular infiltrates on 492
histopathology (Fig. 7B-D). These phenotypes were well-complemented visually, and in the 493
case of lung weight statistically (P >0.05 at days 54-112 for wild type vs. complement). The 494
extent of disease in the spleen was also greater in wild type H37Rv-infected mice compared to 495
ΔcuvA-infected mice, though the difference was less striking (Fig. S7). 496
In separate time to death/severe morbidity experiments, mice were infected by aerosol, 497
four were sacrificed at day 1 to measure the inoculum (Fig. S7D) and the remaining mice were 498
monitored over time. This experiment also showed marked attenuation of the ΔcuvA strain. 50% 499
mortality for wild type H37Rv was reached at day 68, whereas by day 273 when the experiment 500
was ended, none of the mice infected with the ΔcuvA strain had died (P<0.0001) (Fig. 7E). 501
There was partial complementation, with 25% mortality of mice infected with the complemented 502
strain at day 84 (P=0.03 for ΔcuvA vs. complement). This incomplete complementation may 503
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

21
have resulted from constitutive high level cuvA expression in the complemented strain, which 504
we found to be more than 15-fold higher than expression in the wild type. We did not observe 505
polar effects on the expression of genes adjacent to Rv1422 in the cuvA mutant (Fig. S8). 506
507
Discussion 508
In this work, we have provided an initial characterization of the M. tuberculosis gene of 509
unknown function, Rv1422, and it M. smegmatis orthologue MS_3080, which we have named 510
cuvA based on altered carbon source utilization in M. smegmatis and M. tuberculosis cuvA 511
deletion strains and decreased virulence phenotypes of the M. tuberculosis deletion mutant. 512
This protein has homologues across a broad range of diverse bacterial phyla and its amino acid 513
sequence suggests that CuvA is distantly related to the unidentified protein family UPF0052 514
(38). The one characterized member of this family is CofD (LPPG: 2-phospho-L-lactate 515
transferase), a protein present in archaea and some bacteria that has been shown to be 516
involved in coenzyme F420 biosynthesis in Methanocaldococcus jannaschii (39, 40). At the 517
level of primary amino acid sequence, however, CuvA is not significantly similar to CofD. 518
Another M. tuberculosis protein, FbiA (Rv3261), is highly similar to CofD and has been shown to 519
be required for F420 biosynthesis in M. bovis BCG (41), indicating that FbiA is the functional 520
orthologue of CofD and that CuvA likely has a different activity. 521
If it does not function in F420 biosynthesis, what is the role of CuvA? A B. subtilis strain 522
with a deletion in the cuvA orthologue yvcK was shown to have growth defects on several 523
carbon sources and to be defective in localization of PBP1 and cell wall synthesis. In this work 524
we have identified several phenotypes of M. tuberculosis and M. smegmatis cuvA deletion 525
mutants that suggest that the CuvA protein is required for utilization of nutrients and for normal 526
cell wall synthesis. In both species, the cuvA mutants show defects in utilization of several 527
carbon sources, including cholesterol, which is important for M. tuberculosis during infection in 528
vivo. In the case of M. smegmatis, growth defects were apparent when cells were grown in 529
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

22
liquid MM or on agar MM. M. smegmatis ΔcuvA had growth defects on solid medium similar to 530
those seen in M. tuberculosis, with the exception that the M. smegmatis deletion strain grew 531
well on glucose but the M. tuberculosis strain did not. The basis of this difference in growth on 532
glucose will require further investigation, but might result from enhanced metabolic capacity 533
encoded in the much larger genome of the environmental organism M. smegmatis, or in the 534
more much more efficient uptake of glucose mediated via the MspA porin in M. smegmatis 535
compared to M. tuberculosis, which lacks this porin (42, 43). Growth defects of M. tuberculosis 536
ΔcuvA were not apparent in liquid medium. The very slow growth of M. tuberculosis in MM broth 537
and the requirement for a high inoculum for M. tuberculosis to grow in this medium may have 538
limited our ability to detect small differences. The mutant strains in both species had similar 539
morphologic abnormalities, with asymmetric bulging suggestive of a peptidoglycan defect. Also, 540
in both species CuvA localized to the growing cell pole, the site of peptidoglycan synthesis 541
required for cell elongation in actinomycetes including mycobacteria (8, 31). 542
A notable difference between the M. tuberculosis and M. smegmatis cuvA mutants was 543
the markedly increased susceptibility of the M. smegmatis, but not the M. tuberculosis strain, to 544
several cell-wall active antibiotics and to rifampin. Though differences in susceptibility of M. 545
tuberculosis ΔcuvA versus wild type to antibiotics may have been obscured by the poor growth 546
of M. tuberculosis in MM and the high inoculum needed to achieve growth, the magnitude of 547
differences seen in M. smegmatis, particularly for the cephalosporins, suggests that the 548
differences in this phenotype result from differences in the mycobacterial cell envelope in these 549
species. Taken together, the similarity of most phenotypes suggests that the function of CuvA is 550
conserved across these mycobacterial species. This interpretation is also supported by the 551
complementation of both the growth (Fig. S5) and antibiotic susceptibility (not shown) 552
phenotypes in M. smegmatis ΔcuvA by the cuvA gene of M. tuberculosis. 553
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

23
The growth defect of the cuvA deletion strains was not specific to a single carbon 554
source. Growth rates of the cuvA mutants on each of the carbon sources tested, with the 555
exception of propionate, were comparable to or faster than on MM alone, in which asparagine is 556
the primary available energy source. Growth of M. smegmatis ΔcuvA on MM plus propionate, 557
however, was slower than on MM alone. Mycobacterial strains with defects in either the 558
glyoxalate cycle (icl1, icl2 double null mutant) or methylcitrate cycle (prpC, prpD double null 559
mutant) are unable to grow on cholesterol or propionate (44, 45). Though there is some 560
similarity in phenotypes of the ΔcuvA strains and the strains disrupted in glyoxalate or 561
methylcitrate pathways, there are important differences that suggest that CuvA does not 562
participate directly in these pathways. First, the M. tuberculosis strains with defects in the 563
glyoxylate or methylcitrate pathways show minimal growth on cholesterol or propionate in liquid 564
MM (17, 45, 46). In contrast the M. tuberculosis ΔcuvA strain did not have a significant defect in 565
liquid cholesterol medium and the M. smegmatis ΔcuvA strain grew more slowly but did achieve 566
sustained growth in MM plus cholesterol and MM plus propionate. Second, the M. smegmatis 567
ΔcuvA cholesterol growth defect was fully reversed by the addition of glucose, whereas the 568
growth defects of the icl1-2 and prpC-D M. tuberculosis strains were not reversed by glycerol. In 569
addition, the growth inhibitory effect of propionate on the M. smegmatis cuvA mutant was not 570
reversed by addition vitamin B12, in contrast to the prpC-D and icl1,2 mutants in which B12 571
addition allows cells to grow on propionate via utilization of the B12-dependent methylmalonyl 572
CoA pathway (17, 45). Thus, cuvA does not appear to be required for the glyoxylate or 573
methylcitrate pathways. 574
As noted above, our data indicate that the cuvA deletion mutant growth defect is not 575
specific and that CuvA is not an enzyme required for cholesterol catabolism. This inference is 576
consistent with a recent study using genetic approaches and metabolic profiling to map the 577
pathway of cholesterol catabolism in M. tuberculosis, in which Rv1422 was not identified as 578
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

24
being required for any step in cholesterol degradation (17). These data, together with rescue of 579
the growth defect by addition of glucose also suggest that the growth defect is not the result of 580
accumulation of a toxic metabolite from catabolism of the several carbon sources on which the 581
ΔcuvA strain grew less well than wild type. Though a metabolic defect may explain the cuvA 582
growth phenotypes, these data, together with the genetic interaction of CuvA with both the mce-583
4 cholesterol transport locus and with the mce-1 locus that appears to be involved in turnover of 584
mycolic acids (11), suggest a possible alternative mechanism, i.e. that CuvA is required for 585
nutrient uptake into the mycobacterial cell. One mechanism for a broad effect on uptake of 586
many different nutrients would be alterations in the cell envelope in the ΔcuvA strains that affect 587
cell permeability or transporter functions. 588
Several of our results indicate that the mutant strains do have an altered cell envelope 589
and suggest that CuvA has a role in the structure and/or function of the mycobacterial cell wall. 590
First, in addition to decreased growth of the M. tuberculosis and M. smegmatis ΔcuvA mutants 591
on MM containing several carbon sources, these cells have an abnormal morphology, showing 592
shortened, bulging cells. This phenotype is strongly suggestive of an effect on cell wall integrity. 593
This altered morphology is highly similar to the previously described phenotype of an M. 594
smegmatis ponA depletion mutant, and similar to, though less severe than a wag31 depletion 595
mutant (8, 30); in both of these mutants, PGN synthesis is disrupted. The phenotype is also very 596
similar to the morphology of the B. subtilis yvcK deletion strain grown on gluconate, in which the 597
PGN biosynthetic enzyme PBP1 is mis-localized (8, 14, 30). The marked increase in 598
susceptibility of the M. smegmatis ΔcuvA strain to several β-lactam antibiotics, which parallels 599
the phenotype observed in a ponA depletion strain, also suggests that the cell wall is altered in 600
this strain. The localization of CuvA to the growing cell pole, the site of new PGN biosynthesis in 601
mycobacteria, is also consistent with a role for this gene in PGN synthesis, though our data 602
show that CuvA is not essential for this function. 603
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

25
Suppressor mutagenesis can be a valuable means to gain insight into the pathway in 604
which a gene of unknown function is active. The suppressor mutagenesis results we obtained 605
provide further support a role for CuvA in cell wall synthesis and/or transport of nutrients. The 606
inactivating insertions present in pbpA and rodA that restore growth of M. tuberculosis ΔcuvA on 607
cholesterol likely affect the structure of the cell wall. This alteration may allow uptake of 608
cholesterol (and other nutrients) in the ΔcuvA strain in which cholesterol uptake is otherwise 609
impaired. In addition, for the insertions in Rv2683, their location at the 5’ end of the operon and 610
the orientation of all insertions in the direction of transcription of the operon suggest that these 611
may be gain of function mutations resulting from increased expression of genes in the operon 612
from the transposon promoter. Rv2683 is the first gene in a three gene operon that encodes a 613
transporter that was recently shown to be essential for M. tuberculosis growth on cholesterol 614
(36), suggesting that these insertions may enhance uptake of cholesterol and other compounds 615
into the mycobacterial cell. 616
The markedly decreased virulence of the cuvA deletion mutant in a mouse model of 617
tuberculosis suggests a critical role for this protein during infection. This attenuation was 618
especially pronounced in the lung, where both gross tubercle formation and microscopic 619
inflammation were markedly decreased in the ΔcuvA strain, and where CFU of the mutant 620
decreased significantly during the course of infection. Though M. tuberculosis appears to utilize 621
cholesterol throughout infection in mice (16), the small difference in lung CFU early during 622
infection and the progressive decline in CFU of the ΔcuvA strain that we observed during the 623
chronic phase parallels the persistence defect seen in an M. tuberculosis strain lacking the 624
Mce4 cholesterol transport system (2, 28). A second prior mouse infection study also showed 625
an mce4 deletion strain to be attenuated (37). In this study very small differences in lung CFU 626
were seen, along with decreased lung pathology and significantly increased survival of mice 627
infected with the mce4 strain. Compared to these studies, the decrease in lung CFU of the cuvA 628
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

26
mutant in our infection experiment was more pronounced and the survival of the cuvA mutant 629
was more prolonged, even though we used a more susceptible mouse strain (Balb/C) than the 630
previous studies (c57BL/6). In the context of these prior data and the in vitro data presented 631
here, this result suggests that compromised utilization of cholesterol and other nutrients in the 632
ΔcuvA strain during infection likely contribute to the attenuation of the mutant. This interpretation 633
is also consistent with the genetic interaction study that first linked cuvA (Rv1422) to mce4 (28), 634
though other effects of the cuvA deletion, such as a defective cell wall, may also contribute to 635
decreased virulence. 636
The presence of CuvA homologues encoded in the genomes of a broad range of 637
bacteria suggests a highly conserved function for this protein. Though CuvA is required for 638
optimal nutrient utilization by mycobacteria, as for B. subtilis YvcK, the exact function of M. 639
tuberculosis CuvA remains to be defined at the molecular level. Our data, taken together with 640
the B. subtilis YvcK results, however, indicate a role for CuvA that links cell wall synthesis and 641
nutrient utilization. For M. tuberculosis, the severe attenuation of M. tuberculosis virulence and 642
persistence in the deletion strain suggest that CuvA may be a valuable target for development 643
of novel therapeutics to treat tuberculosis disease and/or latent tuberculosis infection. 644
645
646
647
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

27
Acknowledgments 648
We thank members of the Husson laboratory for helpful discussions. This work was supported 649
by NIH grants AI059702 and AI099204 to RNH and by the NIH TARGET consortium contract 650
(N01-AI30036) to William Bishai, Johns Hopkins University. Microscopy resources were 651
provided by the Children’s Hospital Intellectual and Developmental Disabilities Research Center 652
(IDDRC) Cellular Imaging Core (NIH-P30-HD-1865) and New England Regional Center of 653
Excellence in Biodefense and Emerging Infectious Diseases live-cell imaging core (NIH grant 654
AI0571). All authors declare that they have no conflicts of interest.655
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

28
REFERENCES 656
1. McKinney JD, Honer zu Bentrup K, Munoz-Elias EJ, Miczak A, Chen B, 657 Chan WT, Swenson D, Sacchettini JC, Jacobs WR, Jr., Russell DG. 2000. 658 Persistence of Mycobacterium tuberculosis in macrophages and mice requires 659 the glyoxylate shunt enzyme isocitrate lyase. Nature 406:735-738. 660
2. Pandey AK, Sassetti CM. 2008. Mycobacterial persistence requires the 661 utilization of host cholesterol. Proc Natl Acad Sci U S A 105:4376-4380. 662
3. Russell DG, VanderVen BC, Lee W, Abramovitch RB, Kim MJ, Homolka S, 663 Niemann S, Rohde KH. 2010. Mycobacterium tuberculosis wears what it eats. 664 Cell Host & Microbe 8:68-76. 665
4. North RJ, Jung Y-J. 2004. Immunity to tuberculosis. Annu Rev Immunol 22:599-666 623. 667
5. Kang CM, Abbott DW, Park ST, Dascher CC, Cantley LC, Husson RN. 2005. 668 The Mycobacterium tuberculosis serine/threonine kinases PknA and PknB: 669 substrate identification and regulation of cell shape. Genes Dev 19:1692-1704. 670
6. Dasgupta A, Datta P, Kundu M, Basu J. 2006. The serine/threonine kinase 671 PknB of Mycobacterium tuberculosis phosphorylates PBPA, a penicillin-binding 672 protein required for cell division. Microbiology 152:493-504. 673
7. Sureka K, Hossain T, Mukherjee P, Chatterjee P, Datta P, Kundu M, Basu J. 674 2010. Novel role of phosphorylation-dependent interaction between FtsZ and 675 FipA in mycobacterial cell division. PLoS ONE 5:e8590. 676
8. Kang CM, Nyayapathy S, Lee JY, Suh JW, Husson RN. 2008. Wag31, a 677 homologue of the cell division protein DivIVA, regulates growth, morphology and 678 polar cell wall synthesis in mycobacteria. Microbiology 154:725-735. 679
9. Gee CL, Papavinasasundaram KG, Blair SR, Baer CE, Falick AM, King DS, 680 Griffin JE, Venghatakrishnan H, Zukauskas A, Wei JR, Dhiman RK, Crick 681 DC, Rubin EJ, Sassetti CM, Alber T. 2012. A phosphorylated pseudokinase 682 complex controls cell wall synthesis in mycobacteria. Science signaling 5:ra7. 683
10. Mohn WW, van der Geize R, Stewart GR, Okamoto S, Liu J, Dijkhuizen L, 684 Eltis LD. 2008. The actinobacterial mce4 locus encodes a steroid transporter. J 685 Biol Chem 283:35368-35374. 686
11. Forrellad MA, McNeil M, Santangelo MdlP, Blanco FC, García E, Klepp LI, 687 Huff J, Niederweis M, Jackson M, Bigi F. 2014. Role of the Mce1 transporter in 688 the lipid homeostasis of Mycobacterium tuberculosis. Tuberculosis (Edinb) 689 94:170-177. 690
12. Sassetti CM, Rubin EJ. 2003. Genetic requirements for mycobacterial survival 691 during infection. Proc Natl Acad Sci U S A 100:12989-12994. 692
13. Gorke B, Foulquier E, Galinier A. 2005. YvcK of Bacillus subtilis is required for 693 a normal cell shape and for growth on Krebs cycle intermediates and substrates 694 of the pentose phosphate pathway. Microbiology 151:3777-3791. 695
14. Foulquier E, Pompeo F, Bernadac A, Espinosa L, Galinier A. 2011. The YvcK 696 protein is required for morphogenesis via localization of PBP1 under 697 gluconeogenic growth conditions in Bacillus subtilis. Molecular Microbiology 698 80:309-318. 699
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

29
15. Garner EC, Bernard R, Wang W, Zhuang X, Rudner DZ, Mitchison T. 2011. 700 Coupled, circumferential motions of the cell wall synthesis machinery and MreB 701 filaments in B. subtilis. Science 333:222-225. 702
16. Chang JC, Miner MD, Pandey AK, Gill WP, Harik NS, Sassetti CM, Sherman 703 DR. 2009. igr Genes and Mycobacterium tuberculosis cholesterol metabolism. J 704 Bacteriol 191:5232-5239. 705
17. Griffin JE, Pandey AK, Gilmore SA, Mizrahi V, McKinney JD, Bertozzi CR, 706 Sassetti CM. 2012. Cholesterol Catabolism by Mycobacterium tuberculosis 707 Requires Transcriptional and Metabolic Adaptations. Chemistry & Biology 708 19:218-227. 709
18. Snapper SB, Melton RE, Mustafa S, Kieser T, Jacobs WR, Jr. 1990. Isolation 710 and characterization of efficient plasmid transformation mutants of 711 Mycobacterium smegmatis. Mol Microbiol 4:1911-1919. 712
19. Raman S, Hazra R, Dascher CC, Husson RN. 2004. Transcription regulation by 713 the Mycobacterium tuberculosis alternative sigma factor SigD and its role in 714 virulence. J Bacteriol 186:6605-6616. 715
20. Pelicic V, Jackson M, Reyrat J-M, Jacobs WJ, Gicquel B, Guilhot C. 1997. 716 Efficient allelic exchange and transposon mutagenesis in Mycobacterium 717 tuberculosis. Proc Natl Acad Sci USA 94:10955-10960. 718
21. Blokpoel MC, Murphy HN, O'Toole R, Wiles S, Runn ES, Stewart GR, Young 719 DB, Robertson BD. 2005. Tetracycline-inducible gene regulation in 720 mycobacteria. Nucleic Acids Res 33:e22. 721
22. Guinn KM, Hickey MJ, Mathur SK, Zakel KL, Grotzke JE, Lewinsohn DM, 722 Smith S, Sherman DR. 2004. Individual RD1-region genes are required for 723 export of ESAT-6/CFP-10 and for virulence of Mycobacterium tuberculosis. Mol 724 Microbiol 51:359-370. 725
23. Collins L, Franzblau SG. 1997. Microplate alamar blue assay versus BACTEC 726 460 system for high-throughput screening of compounds against Mycobacterium 727 tuberculosis and Mycobacterium avium. Antimicrob Agents Chemother 41:1004-728 1009. 729
24. Sampson SL, Dascher CC, Sambandamurthy VK, Russell RG, Jacobs WR, 730 Bloom BR, Hondalus MK. 2004. Protection elicited by a double leucine and 731 pantothenate auxotroph of Mycobacterium tuberculosis in guinea pigs. Infect 732 immun 72:3031-3037. 733
25. Rubin EJ, Akerley BJ, Novik VN, Lampe DJ, Husson RN, Mekalanos JJ. 734 1999. In vivo transposition of mariner-based elements in enteric bacteria and 735 mycobacteria. Proc Natl Acad Sci USA 96:1645-1650. 736
26. Sassetti CM, Boyd DH, Rubin EJ. 2001. Comprehensive identification of 737 conditionally essential genes in mycobacteria. Proc Natl Acad of Sci U S A 738 98:12712-12717. 739
27. Murry JP, Sassetti CM, Lane JM, Xie Z, Rubin EJ. 2008. Transposon site 740 hybridization in Mycobacterium tuberculosis. Methods in Molecular Biology 741 416:45-59. 742
28. Joshi SM, Pandey AK, Capite N, Fortune SM, Rubin EJ, Sassetti CM. 2006. 743 Characterization of mycobacterial virulence genes through genetic interaction 744 mapping. Proc Natl Acad Sci U S A 103:11760-11765. 745
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

30
29. Prisic S, Dankwa S, Schwartz D, Chou MF, Locasale JW, Kang CM, Bemis 746 G, Church GM, Steen H, Husson RN. 2010. Extensive phosphorylation with 747 overlapping specificity by Mycobacterium tuberculosis serine/threonine protein 748 kinases. Proc Natl Acad Sci U S A 107:7521-7526. 749
30. Hett EC, Chao MC, Rubin EJ. 2010. Interaction and modulation of two 750 antagonistic cell wall enzymes of mycobacteria. PLoS Pathogens 6:e1001020. 751
31. Daniel RA, Errington J. 2003. Control of cell morphogenesis in bacteria: two 752 distinct ways to make a rod-shaped cell. Cell 113:767-776. 753
32. Aldridge BB, Fernández-Suárez M, Heller D, Ambravaneswaran V, Irimia D, 754 Toner M, Fortune SM. 2012. Asymmetry and aging of mycobacterial cells lead 755 to variable growth and antibiotic susceptibility. Science 335:100-104. 756
33. Billman-Jacobe H, Haites RE, Coppel RL. 1999. Characterization of a 757 Mycobacterium smegmatis mutant lacking penicillin binding protein 1. Antimicrob 758 Agents Chemother 43:3011-3013. 759
34. Flores AR, Parsons LM, Pavelka MS. 2005. Characterization of novel 760 Mycobacterium tuberculosis and Mycobacterium smegmatis mutants 761 hypersusceptible to beta-lactam antibiotics. J. Bacteriol. 187:1892-1900. 762
35. Parikh A, Verma SK, Khan S, Prakash B, Nandicoori VK. 2009. PknB-763 mediated phosphorylation of a novel substrate, N-acetylglucosamine-1-764 phosphate uridyltransferase, modulates its acetyltransferase activity. J Mol Biol 765 386:451-464. 766
36. Griffin JE, Gawronski JD, DeJesus MA, Ioerger TR, Akerley BJ, Sassetti 767 CM. 2011. High-Resolution Phenotypic Profiling Defines Genes Essential for 768 Mycobacterial Growth and Cholesterol Catabolism. PLoS Pathogens 769 7:e1002251. 770
37. Senaratne RH, Sidders B, Sequeira P, Saunders G, Dunphy K, Marjanovic 771 O, Reader JR, Lima P, Chan S, Kendall S, McFadden J, Riley LW. 2008. 772 Mycobacterium tuberculosis strains disrupted in mce3 and mce4 operons are 773 attenuated in mice. J Med Microbiol 57:164-170. 774
38. Jain E, Bairoch A, Duvaud S, Phan I, Redaschi N, Suzek B, Martin M, 775 McGarvey P, Gasteiger E. 2009. Infrastructure for the life sciences: design and 776 implementation of the UniProt website. BMC Bioinformatics 10:136. 777
39. Forouhar F, Abashidze M, Xu H, Grochowski L, Seetharaman J, Hussain M, 778 Kuzin A, Chen Y, Zhou W, Xiao R, Acton T, Montelione G, Galinier A, White 779 R, Tong L. 2008. Molecular insights into the biosynthesis of the F420 coenzyme. 780 J Biol Chem 283:11832-11840. 781
40. Graupner M, Xu H, White R. 2002. Characterization of the 2-phospho-L-lactate 782 transferase enzyme involved in coenzyme F(420) biosynthesis in Methanococcus 783 jannaschii. Biochemistry 41:3754-3761. 784
41. Choi KP, Bair TB, Bae YM, Daniels L. 2001. Use of transposon Tn5367 785 mutagenesis and a nitroimidazopyran-based selection system to demonstrate a 786 requirement for fbiA and fbiB in coenzyme F(420) biosynthesis by 787 Mycobacterium bovis BCG. J Bacteriol 183:7058-7066. 788
42. Stephan J, Bender J, Wolschendorf F, Hoffmann C, Roth E, Mailänder C, 789 Engelhardt H, Niederweis M. 2005. The growth rate of Mycobacterium 790
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

31
smegmatis depends on sufficient porin-mediated influx of nutrients. Mol Microbiol 791 58:714-730. 792
43. Siroy A, Mailaender C, Harder D, Koerber S, Wolschendorf F, Danilchanka 793 O, Wang Y, Heinz C, Niederweis M. 2008. Rv1698 of Mycobacterium 794 tuberculosis represents a new class of channel-forming outer membrane 795 proteins. J Biol Chem 283:17827-17837. 796
44. Munoz-Elias EJ, McKinney JD. 2005. Mycobacterium tuberculosis isocitrate 797 lyases 1 and 2 are jointly required for in vivo growth and virulence. Nat Med 798 11:638-644. 799
45. Savvi S, Warner DF, Kana BD, Mckinney JD, Mizrahi V, Dawes SS. 2008. 800 Functional characterization of a vitamin B12-dependent methylmalonyl pathway 801 in Mycobacterium tuberculosis: implications for propionate metabolism during 802 growth on fatty acids. J. Bacteriol. 190:3886-3895. 803
46. Eoh H, Rhee KY. 2014. Methylcitrate cycle defines the bactericidal essentiality of 804 isocitrate lyase for survival of Mycobacterium tuberculosis on fatty acids. Proc 805 Natl Acad Sci U S A 111:4976-4981. 806
807 808
809
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

32
810
Figure legends 811
Figure 1. Growth of M. tuberculosis strains in liquid and solid media. A. Wild type M. 812
tuberculosis H37Rv, ΔcuvAMt and complemented ΔcuvA (ΔcuvAMt/C) strains were grown in M-813
ADC-Tw liquid medium with 0.2% glucose, and OD600 was measured daily. B. Growth of the 814
same strains on agar plates containing MM plus 0.2% glucose or MM plus 0.01% cholesterol. 815
Serial 10-fold dilutions were spotted, incubated at 37o C and photographs were taken after one 816
month of incubation. In both panels, data shown are from one experiment that was repeated at 817
least twice with similar results. Glc, 0.2% glucose; Chol, 0.01% cholesterol. 818
819
Figure 2. Growth of M. tuberculosis strains on MM agar containing added carbon sources. Serial 820
10-fold dilutions of wild type M. tuberculosis H37Rv, ΔcuvAMt and complemented ΔcuvA 821
(ΔcuvAMt/C) strains were spotted on MM agar alone or containing the indicated compounds 822
added to a concentration of 0.01%, incubated at 37o C and photographs were taken after one 823
month of incubation. Data shown are from one experiment that was repeated at least twice with 824
similar results. Glc, glucose; Glyc, glycerol; Glcn, gluconate; Citr, citrate; Prop, propionate. 825
826
827
Figure 3. Growth of M. smegmatis strains on different carbon sources. M. smegmatis mc2-155 828
(WT); ΔcuvAMs, and complemented ΔcuvA (∆cuvAMs/C) were diluted to OD600=0.001 and grown 829
in liquid MM alone or with added carbon sources and analyzed using the MABA performed with 830
technical duplicates. Except for hiGlc, which contained 0.2% glucose, all compounds were 831
added to a concentration of 0.01%. The exponential growth phase of ΔcuvAMs was significantly 832
slower than wild type or ∆cuvAMs/C in MM alone (P<0.0001), MM plus cholesterol (P<0.0001), 833
MM plus glycerol (P<0.0001), MM plus citrate (P<0.005) and MM plus propionate ((P<0.0001). 834
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

33
The growth rates were not significantly different (P> 0.05) when grown on 0.2% glucose, 0.01% 835
glucose or gluconate. Glc, glucose; Glyc, glycerol ; Citr, citrate;. Chol, cholesterol; Glcn, 836
gluconate; Prop, propionate. Error bars are +/- 1 SD. 837
838
Figure 4. Morphology of M. smegmatis and M. tuberculosis cells. A. M. smegmatis mc2-155 839
(Wild type); ΔcuvAMs, and complemented ΔcuvA (∆cuvAMs/C) were grown in MM plus 0.01% 840
cholesterol, harvested after 24h and examined by light microscopy. Bar = 1 μm. B. Length 841
distribution of M. smegmatis cells obtained after 24h of growth in MM plus 0.01% cholesterol, 842
determined by measuring over 200 cells of each strain, analyzed using imageJ software. The 843
horizontal line is mean cell length. C. Cells of each M. smegmatis strain were grown overnight 844
on agarose pads made with MM plus 0.01% cholesterol and photographed. D. Wild type M. 845
tuberculosis H37Rv, ΔcuvAMt and complemented ΔcuvA (ΔcuvAMt/C) strains were grown to late 846
stationary phase in MM liquid medium plus 0.01% cholesterol and examined by light microscopy 847
using a 100x oil immersion objective. Arrowheads in panels C and D indicate areas of abnormal 848
cell morphology. 849
850
851
Fig. 5. Co-localization of CuvA and nascent peptidoglycan synthesis in M. smegmatis. Wild 852
type M. smegmatis mc2-155 cells expressing GFP alone or GFP fused to CuvAMt were grown M-853
ADC-TW containing 0.2% glucose or MM containing 0.01% cholesterol. Fluorescent 854
Vancomycin labeled with Alexa-568 was used to bind to newly synthesized peptidoglycan. 855
Fluorescence and DIC microscopy images were obtained to determine the localization of 856
CuvAMt-GFP and nascent peptidoglycan synthesis. Bar = 1 μM. 857
858
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

34
Fig. 6. Localization of CuvA in M. tuberculosis. An M. tuberculosis leucine/pantothenate 859
auxotroph strain (24) expressing CuvAMt-GFP or unfused GFP were grown on agarose pads 860
containing M-ADC medium with 0.2% glucose, and acetamide to induce expression of cuvAMt-861
gfp. Fluorescence and DIC microscopy images were obtained as described in the Methods 862
section after 24 hours induction to determine the localization of CuvAMt-GFP. Bar = 1 μM. 863
864
Fig. 7. Mouse infection experiments. Balb/C mice were infected by aerosol with wild type M. 865
tuberculosis wild type H37Rv, ΔcuvAMt or the ΔcuvA complemented strain (cuvAMt/C). In 866
separate experiments mice were monitored for morbidity/mortality or sacrificed at serial time 867
points for pathologic analysis and measurement of bacterial burden. A. CFUs in lungs from day 868
0 to day 112. Data are mean from four mice at each time point. Lungs from mice infected with 869
wild type had significantly more CFU in the lung than those infected with the ΔcuvA strain at 870
days 56, 84 and 112 (P<0.0001 to P<0.005). Mice infected with the complemented strain had 871
significantly more CFUs in the lung than those infected with the ΔcuvA strain at these time 872
points (P<0.0001 to P<0.01), but fewer CFUs than those infected with the wild type (P<0.0001 873
to P<0.05). Error bars = 1 S.D. B. Microscopic pathology of sections stained with hematoxylin 874
and eosin on day 84. C. Gross pathology of lungs on day 84. D. Lung weight from day 0 to day 875
112. Data are mean from four mice at each time point. Lungs of mice infected with the wild type 876
or complemented strain were significantly heavier than those infected with the ΔcuvA strain 877
(P<0.0001 to P<0.005 at days 54-112). Error bars = 1 S.D. E. Survival curve of mice infected 878
with the three strains of M. tuberculosis. Mortality was significantly lower in mice infected with 879
the ΔcuvA strain compared to those infected with wild type (P<0.0001) or the complemented 880
strain (P<0.05). Mortality in the complement-infected mice was also significantly lower than 881
those infected with wild type (P<0.0001). 882
883
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

35
Table 1. Antibiotic Susceptibility of M. smegmatis strains 884 885
A Chol. Chol.+Glc Chol.+CLAV Chol.+Glc+CLAV
AMP
WT 375±137 250±0 63±0 52±16
ΔcuvA 229±51 250±0 5±2 31±0
Δ/C 375±137 250±0 31±0 47±38
CEF
WT >1000* >1000* >1000* >1000*
ΔcuvA 15.6±0.0 >1000* 7.3±0.8 >1000*
Δ/C >1000* >1000* >1000* >1000*
MER
WT 40.6±17.4
28.1±14.6
16.7±6.5 16.7±6.5
ΔcuvA 5.9±3.1 17.2±8.7 3.1±1.7 6.3±0.0
Δ/C 28.1±14.6
28.1±14.6
12.5±0.0 12.5±0.0
CEPH
WT >500* >500* >500* >500*
ΔcuvA >500* >500* 36.5±12.7 >500*
Δ/C >500* >500* >500* >500*
Data are minimal inhibitory concentration in μg/ml +/- 1 standard deviation. AMP, ampicillin; CEF, 886 cefotaxime; MER, meropenem; CEPH, cephalothin; RIF, rifampin; LEV, levofloxacin; INH, isoniazid; GEN, 887 gentamicin; CLAV, clavulanic acid. Chol, 0.01% cholesterol; Glc, 0.2% glucose. Shaded boxes indicate 888 results for ΔcuvA that are significantly different from wild type (P<0.05 and >2-fold difference). *Highest 889 concentration tested. 890
B Chol. Chol.+Glc
RIF
WT 26.7±10.3 16.3±5.2
ΔcuvA 2.3±1.9 15.6±6.2
Δ/C 17.5±5.0 16.7±5.2
LEV
WT 0.58±0.13 0.58±0.13
ΔcuvA 0.32±0.00 0.32±0.00
Δ/C 0.32±0.00 0.30±0.05
INH
WT >32* >32*
ΔcuvA >32* >32*
Δ/C >32* >32*
GEN
WT 0.08±0.02 0.08±0.02
ΔcuvA 0.08±0.03 0.08±0.02
Δ/C 0.08±0.02 0.07±0.03
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

36
Table 2. Transposon insertions that suppress the cholesterol growth defect of M. 891 tuberculosis ΔcuvA 892
Genes in which two or more independent insertions were obtained are included in the Table. f and r in 893 parentheses indicate the forward or reverse orientation of the kanamycin marker in the transposon with 894 respect to the direction of transcription of the gene disrupted. The number preceding f or r indicates the 895 number of independent transposon insertions obtained at each site. 896
897
Gene disrupted
Insertion sites and number of insertions
Probable function of gene/operon
rodA +755 (2r), +1291 (1r) Cell shape/peptidoglycan synthesis, Ser/Thr phosphorylation
pbpA +142 (1r), +232 (1r), +250 (1f), +270 (1f), +541 (1f)
Peptidoglycan synthesis/cell shape, Ser/Thr phosphorylation
Rv2683 -1 (3f), +191 (1f) Conserved hypothetical regulatory protein/transmembrane transport
trcR +681 (1f), +732 (1r) Response regulator/TrcRS two component system
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from

Wild type cuvAMt cuvAMt/C
A
B
C
D
on June 13, 2016 by guesthttp://iai.asm
.org/D
ownloaded from




















