
Induction of Mycobacterial Resistance to Quinolone Class Antimicrobials
9
Induction of Mycobacterial Resistance to Quinolone Class Antimicrobials Muhammad Malik, a Kalyan Chavda, a Xilin Zhao, a,b Nirali Shah, a Syed Hussain, a Natalia Kurepina, a Barry N. Kreiswirth, a Robert J. Kerns, c and Karl Drlica a,b Public Health Research Institute, New Jersey Medical School, UMDNJ, Newark, New Jersey, USA a ; Department of Microbiology & Molecular Genetics, New Jersey Medical School, UMDNJ, Newark, New Jersey, USA b ; and University of Iowa, Division of Medicinal & Natural Products Chemistry, College of Pharmacy, Iowa City, Iowa, USA c An agar plate assay was developed for detecting the induction of drug-resistant mycobacterial mutants during exposure to inhib- itors of DNA gyrase. When Mycobacterium smegmatis on drug-containing agar, resistant colonies arose over a period of 2 weeks. A recA deficiency reduced mutant recovery, consistent with involvement of the SOS response in mutant induction. The C-8-me- thoxy compounds gatifloxacin and moxifloxacin allowed the recovery of fewer resistant mutants than either ciprofloxacin or levofloxacin when present at the same multiple of the MIC; a quinolone-like 8-methoxy-quinazoline-2,4-dione was more effec- tive at restricting the emergence of resistant mutants than its cognate fluoroquinolone. Thus, the structure of fluoroquinolone- like compounds affects mutant recovery. A spontaneous mutator mutant of M. smegmatis, obtained by growth in medium con- taining both isoniazid and rifampin, increased mutant induction during exposure to ciprofloxacin. Moreover, the mutator increased the size of spontaneous resistant mutant subpopulations, as detected by population analysis. Induction of ciprofloxa- cin resistance was also observed with Mycobacterium tuberculosis H37Rv. When measured with clinical isolates, no difference in mutant recovery was observed between multidrug-resistant (MDR) and pansusceptible isolates. This finding is consistent with at least some MDR isolates of M. tuberculosis lacking mutators detectable by the agar plate assay. Collectively, the data indicate that the use of fluoroquinolones against tuberculosis may induce resistance and that the choice of quinolone may be important for restricting the recovery of induced mutants. F luoroquinolones are broad-spectrum antimicrobials that are important for the treatment of multidrug-resistant (MDR) tu- berculosis (TB) (6, 37). Unfortunately, fluoroquinolone resis- tance is emerging, often in strains of Mycobacterium tuberculosis that are already MDR (62). When the resulting fluoroquinolone- resistant MDR mutants are also resistant to an injectable drug such as kanamycin, amikacin, or capreomycin, they are consid- ered to be extensively drug resistant (XDR) (12). At that point, treatment is still possible but obtaining a successful outcome is quite difficult (3, 19, 38). Thus, having a new, highly effective fluoroquinolone is desirable to halt the progression to XDR status. During the last decade, several new quinolones were developed for other Gram-positive bacteria, and two of these agents, moxifloxa- cin and gatifloxacin, are now being considered as additions to the anti-TB armamentarium (14, 39, 58). However, a well-known problem of fluoroquinolone action with other bacteria is the in- duction of the mutagenic SOS response (21, 42, 52). If this phe- nomenon extends to mycobacteria, the quinolones are expected to induce resistance to themselves and to other agents commonly employed. Thus, understanding and restricting the emergence of resistance during quinolone exposure are likely to be important. Work with Escherichia coli suggests that an agar plate assay can be used to detect the induction of resistant mutants during drug exposure (7, 8, 31). Induced mutants appear as colonies that grad- ually accumulate over a period of 10 to 14 days on fluoroquin- olone-containing agar; mutant subpopulations present prior to drug exposure appear as colonies within 1 to 2 days after plating. Mutant induction requires RecA and inducible LexA (activation of RecA promotes self-cleavage of LexA, the repressor of the SOS regulon (28). It also requires a large parental population, making the readout sensitive to the lethal action of quinolones. Some quinolone class compounds also suppress mutant growth, which will reduce the recovery of induced mutants. Thus, the agar plate assay is a composite test for several important quinolone activities that are likely to depend on drug structure. To determine whether the agar plate assay is suitable for my- cobacteria, we plated Mycobacterium smegmatis on quinolone- containing agar and measured colony accumulation over a 2-week period. To test for mutant induction, we blocked the induction of the mutagenic SOS response with a recA mutation and mea- sured the effect on the accumulation of ciprofloxacin-resistant colonies. The sensitivity of the assay to fluoroquinolone structure was then examined with four commercially available compounds, and assay sensitivity to mutator mutants was assessed with a spon- taneous mutator. When we applied the agar plate assay to M. tuberculosis using ciprofloxacin, a fluoroquinolone known to en- rich resistant mutants (65), mutant induction was readily ob- served. We expect the assay to be useful for comparing anti-TB agents and for assessing the mutator status of bacterial isolates. MATERIALS AND METHODS Bacterial strains and culture conditions. The M. smegmatis and M. tu- berculosis strains used in the study are listed in Tables 1 and 2, respectively. All mycobacteria were cultured in 7H9 liquid medium or on 7H10 agar plates, in both cases supplemented with 10% albumin-dextrose-catalase, Received 4 March 2012 Returned for modification 26 March 2012 Accepted 27 April 2012 Published ahead of print 7 May 2012 Address correspondence to Karl Drlica, [email protected]. Copyright © 2012, American Society for Microbiology. All Rights Reserved. doi:10.1128/AAC.00474-12 July 2012 Volume 56 Number 7 Antimicrobial Agents and Chemotherapy p. 3879 –3887 aac.asm.org 3879
Transcript of Induction of Mycobacterial Resistance to Quinolone Class Antimicrobials

Induction of Mycobacterial Resistance to Quinolone ClassAntimicrobials
Muhammad Malik,a Kalyan Chavda,a Xilin Zhao,a,b Nirali Shah,a Syed Hussain,a Natalia Kurepina,a Barry N. Kreiswirth,a
Robert J. Kerns,c and Karl Drlicaa,b
Public Health Research Institute, New Jersey Medical School, UMDNJ, Newark, New Jersey, USAa; Department of Microbiology & Molecular Genetics, New Jersey MedicalSchool, UMDNJ, Newark, New Jersey, USAb; and University of Iowa, Division of Medicinal & Natural Products Chemistry, College of Pharmacy, Iowa City, Iowa, USAc
An agar plate assay was developed for detecting the induction of drug-resistant mycobacterial mutants during exposure to inhib-itors of DNA gyrase. When Mycobacterium smegmatis on drug-containing agar, resistant colonies arose over a period of 2 weeks.A recA deficiency reduced mutant recovery, consistent with involvement of the SOS response in mutant induction. The C-8-me-thoxy compounds gatifloxacin and moxifloxacin allowed the recovery of fewer resistant mutants than either ciprofloxacin orlevofloxacin when present at the same multiple of the MIC; a quinolone-like 8-methoxy-quinazoline-2,4-dione was more effec-tive at restricting the emergence of resistant mutants than its cognate fluoroquinolone. Thus, the structure of fluoroquinolone-like compounds affects mutant recovery. A spontaneous mutator mutant of M. smegmatis, obtained by growth in medium con-taining both isoniazid and rifampin, increased mutant induction during exposure to ciprofloxacin. Moreover, the mutatorincreased the size of spontaneous resistant mutant subpopulations, as detected by population analysis. Induction of ciprofloxa-cin resistance was also observed with Mycobacterium tuberculosis H37Rv. When measured with clinical isolates, no difference inmutant recovery was observed between multidrug-resistant (MDR) and pansusceptible isolates. This finding is consistent with atleast some MDR isolates of M. tuberculosis lacking mutators detectable by the agar plate assay. Collectively, the data indicate thatthe use of fluoroquinolones against tuberculosis may induce resistance and that the choice of quinolone may be important forrestricting the recovery of induced mutants.
Fluoroquinolones are broad-spectrum antimicrobials that areimportant for the treatment of multidrug-resistant (MDR) tu-
berculosis (TB) (6, 37). Unfortunately, fluoroquinolone resis-tance is emerging, often in strains of Mycobacterium tuberculosisthat are already MDR (62). When the resulting fluoroquinolone-resistant MDR mutants are also resistant to an injectable drugsuch as kanamycin, amikacin, or capreomycin, they are consid-ered to be extensively drug resistant (XDR) (12). At that point,treatment is still possible but obtaining a successful outcome isquite difficult (3, 19, 38). Thus, having a new, highly effectivefluoroquinolone is desirable to halt the progression to XDR status.During the last decade, several new quinolones were developed forother Gram-positive bacteria, and two of these agents, moxifloxa-cin and gatifloxacin, are now being considered as additions to theanti-TB armamentarium (14, 39, 58). However, a well-knownproblem of fluoroquinolone action with other bacteria is the in-duction of the mutagenic SOS response (21, 42, 52). If this phe-nomenon extends to mycobacteria, the quinolones are expectedto induce resistance to themselves and to other agents commonlyemployed. Thus, understanding and restricting the emergence ofresistance during quinolone exposure are likely to be important.
Work with Escherichia coli suggests that an agar plate assay canbe used to detect the induction of resistant mutants during drugexposure (7, 8, 31). Induced mutants appear as colonies that grad-ually accumulate over a period of 10 to 14 days on fluoroquin-olone-containing agar; mutant subpopulations present prior todrug exposure appear as colonies within 1 to 2 days after plating.Mutant induction requires RecA and inducible LexA (activationof RecA promotes self-cleavage of LexA, the repressor of the SOSregulon (28). It also requires a large parental population, makingthe readout sensitive to the lethal action of quinolones. Somequinolone class compounds also suppress mutant growth, which
will reduce the recovery of induced mutants. Thus, the agar plateassay is a composite test for several important quinolone activitiesthat are likely to depend on drug structure.
To determine whether the agar plate assay is suitable for my-cobacteria, we plated Mycobacterium smegmatis on quinolone-containing agar and measured colony accumulation over a 2-weekperiod. To test for mutant induction, we blocked the inductionof the mutagenic SOS response with a recA mutation and mea-sured the effect on the accumulation of ciprofloxacin-resistantcolonies. The sensitivity of the assay to fluoroquinolone structurewas then examined with four commercially available compounds,and assay sensitivity to mutator mutants was assessed with a spon-taneous mutator. When we applied the agar plate assay to M.tuberculosis using ciprofloxacin, a fluoroquinolone known to en-rich resistant mutants (65), mutant induction was readily ob-served. We expect the assay to be useful for comparing anti-TBagents and for assessing the mutator status of bacterial isolates.
MATERIALS AND METHODSBacterial strains and culture conditions. The M. smegmatis and M. tu-berculosis strains used in the study are listed in Tables 1 and 2, respectively.All mycobacteria were cultured in 7H9 liquid medium or on 7H10 agarplates, in both cases supplemented with 10% albumin-dextrose-catalase,
Received 4 March 2012 Returned for modification 26 March 2012Accepted 27 April 2012
Published ahead of print 7 May 2012
Address correspondence to Karl Drlica, [email protected].
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AAC.00474-12
July 2012 Volume 56 Number 7 Antimicrobial Agents and Chemotherapy p. 3879–3887 aac.asm.org 3879
0.2% glycerol, and 0.05% Tween 80 (18). Incubation was at 37°C; all M.tuberculosis work was conducted in a biosafety level 3 containment facility.
Chemicals and reagents. Ciprofloxacin and moxifloxacin were prod-ucts of Bayer Healthcare (West Haven, CT), and gatifloxacin was obtainedfrom Bristol-Myers Squibb (Princeton, NJ). PD160793 was a generous giftfrom John Domagala (Parke-Davis Division of Pfizer Chemical Co., AnnArbor, MI). Dione UING5-207 and the cognate fluoroquinolone UING5-249 were prepared as previously outlined (13). Levofloxacin, isoniazid,and rifampin were obtained from Sigma-Aldrich (St. Louis, MO). Fluo-roquinolones were dissolved in 1 N NaOH at 1/10 of the final volume, andthen sterile water was added to obtain a final concentration of 10 mg/ml.Dione UING5-207 was dissolved in dimethyl sulfoxide at 10 mg/ml. Iso-niazid was dissolved in water, and rifampin was dissolved in ethanol.Dilutions were prepared with sterile distilled water, and solutions werekept at �20°C for several weeks during testing.
Measurement of antimicrobial susceptibility. Inhibition of growth(MIC) was measured by agar or broth dilution. About 104 to 105 cellswere applied to a series of agar plates or broth cultures containingantimicrobials at various concentrations. M. smegmatis colonies werecounted after incubation for 3 days or turbid growth was detected after2 days. The MIC was taken as the minimal concentration that blockedthe growth of liquid cultures or inhibited the formation of colonies by99%, as determined from interpolation of log-log plots. MICs for M.tuberculosis were determined by the broth dilution method after incu-bation for 10 days.
Lethal activity was measured with liquid cultures of exponentiallygrowing M. smegmatis in which aliquots containing 5 � 108 CFU/ml wereincubated for 18 h in the presence of various concentrations of ciprofloxa-cin. The number of surviving cells was determined by dilution and platingon drug-free agar, followed by incubation at 37°C for 3 days. Percentsurvival was determined relative to that of an untreated control sampled atthe time of drug addition.
Detection of quinolone-induced, quinolone-resistant mutants. Theagar plate method previously described for E. coli (31) was used, with
slight modification. Briefly, concentrated quinolone solutions were addedto molten agar, and mycobacterial cells (104 to 108) were plated on solid-ified agar. The cumulative number of colonies forming during incubationat 37°C was determined at regular intervals.
Enrichment of mutator mutants. The presence of a mutator muta-tion was defined by the concurrent recovery of resistance to two drugs(single-step double mutants) under conditions in which the number ofbacteria in the culture was below that required for recovery of resistance toone drug alone times the number of bacteria required for the recovery ofresistance to the other drug. To obtain single-step double mutants, cul-tures of wild-type M. smegmatis (KD1163) were grown to mid-log phaseand diluted to generate sets of cultures (16 1-ml cultures) such that setscontained 3 � 105 CFU/ml, 1.3 � 106 CFU/ml, 5 � 106 CFU/ml, or 2 �107 CFU/ml. All cultures received both rifampin and isoniazid at 3 timesthe MIC (rifampin MIC, 1.5 �g/ml; isoniazid MIC, 6.25 �g/ml). After thecultures were incubated at 37°C for 10 days with shaking, those exhibitinggrowth (turbidity) were presumed to contain a mutator mutation. Singlecolonies were isolated and grown into cultures for analysis.
Population analysis profiles. Population analysis was performed aspreviously described (65). Briefly, a series of agar plates was prepared inwhich the concentration of the fluoroquinolone PD160793 varied over abroad range. Test strains of M. smegmatis, grown to stationary phase inliquid medium, were plated in amounts that allowed a small number ofcolonies to form during incubation for 3 days. To verify that the coloniesrepresented mutants, they were transferred to drug-free agar for a secondround of growth and then tested on agar containing the drug at the sameconcentration used initially for selection. All colonies grew when retestedon fluoroquinolone-containing agar.
Ethidium bromide-resistant mutants were identified by the ability toform colonies on agar containing 6.25 �g/ml ethidium bromide, whichwas equivalent to the MIC measured with cultures grown in liquid me-dium.
DNA isolation and nucleotide sequence determination. Cultures ofstationary-phase M. smegmatis were harvested by centrifugation, andDNA was extracted as described previously (65). Briefly, cells were treatedwith lysozyme, proteinase K, and sodium dodecyl sulfate; lysates wereextracted with mixtures of chloroform-isoamyl alcohol; and DNA wasprecipitated with alcohol. PCR was performed to amplify the quinoloneresistance-determining regions (QRDR) of gyrA and gyrB, and the nucle-otide sequences of those regions were determined. The oligonucleotidesused for amplification were 5=-ACG AAC TGT TCT CCA TCC TCA TGGGCG AAG-3= (forward primer), 5=-ACA TCT CCA TCG CCA ACG GAGTG-3= (reverse primer), and 5=-CAG CGC AGC TAC ATC GAC TACGC-3= (sequencing primer) for gyrA and 5=-CCG ACT GCC GTT CGACGG AT-3= (forward and sequencing primer) and 5=-CGG CCA TCAACA CGA TCT TG-3= (reverse primer) for gyrB.
TABLE 1 M. smegmatis strains used in this study
StrainRelevant genotypeor phenotype Source or reference
KD1163 Wild type (mc2155) Laboratory stock; S. ColeKD2859 KD1163 Inhr Rifr
mutatorThis worka
KD2860 KD1163 Inhr Rifr This workb
mc2155 (KD2045) recA� K. G. PapavinasasundaramHS42 (KD2046) mc2155 �recA K. G. Papavinasasundarama Obtained by selection of double resistance in a single step.b Obtained by stepwise selection.
TABLE 2 M. tuberculosis strains used in this study
Strain sSNP clustera RFLPb Resistancec Spoligotype octal coded PGGe Source
H37Rv VII H37Rv None 777777477760771 3 Laboratory strain, PHRIf TB Center10536 II KY R, I 000000000000771 1 Clinical isolate, PHRI TB Center10775 II W4 None 000000000003771 1 Clinical isolate, PHRI TB Center18996 IV H I, R, E, PZA 777776777760601 2 Clinical isolate, PHRI TB Center13571 IV H None 777776777760601 2 Clinical isolate, PHRI TB Center16644 V AH I, R, E, PZA, S 777776777760771 2 Clinical isolate, PHRI TB Center12850 V AH None 777776777760771 2 Clinical isolate, PHRI TB Centera sSNP, synonymous single nucleotide polymorphism. Isolates have been grouped based on closely related genotypes (1).b RFLP (restriction fragment length polymorphism) genotype method based on IS6110 (1).c Abbreviations: E, ethambutol; I, isoniazid; PZA, pyrazinamide; R, rifampin; S, streptomycin.d Spoligotyping is a PCR-based genotyping method focused on directly repeated nucleotide sequences (5).e PGG, principal genetic group (23, 57).f PHRI, Public Health Research Institute.
Malik et al.
3880 aac.asm.org Antimicrobial Agents and Chemotherapy
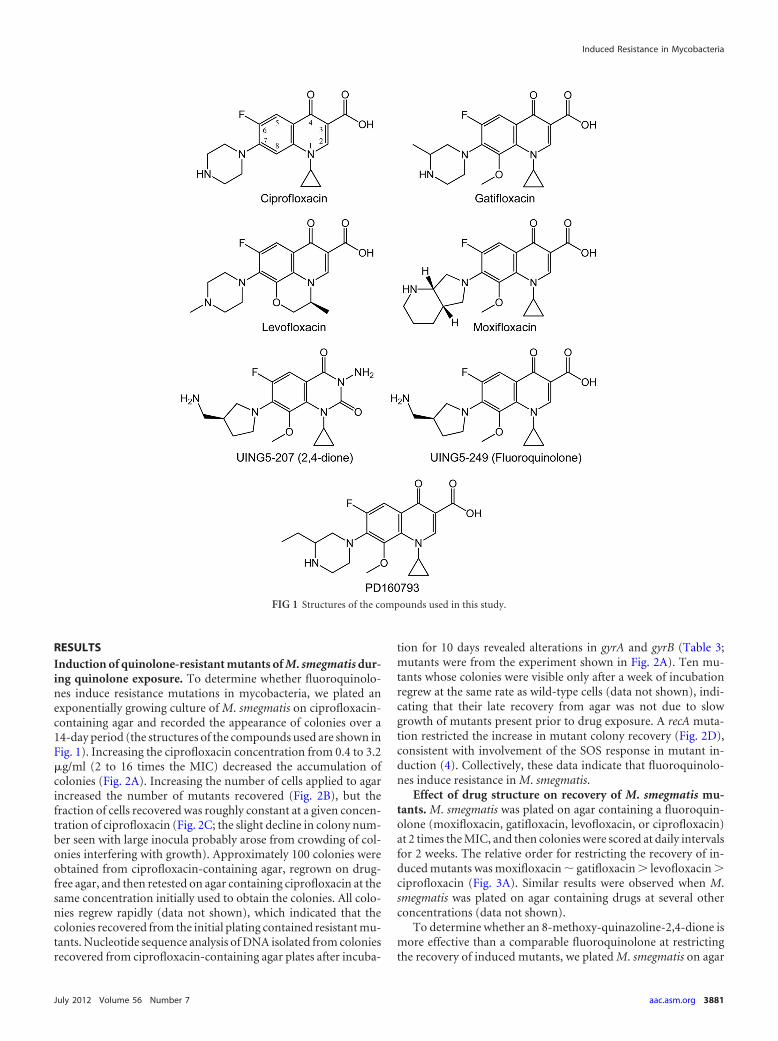
RESULTSInduction of quinolone-resistant mutants of M. smegmatis dur-ing quinolone exposure. To determine whether fluoroquinolo-nes induce resistance mutations in mycobacteria, we plated anexponentially growing culture of M. smegmatis on ciprofloxacin-containing agar and recorded the appearance of colonies over a14-day period (the structures of the compounds used are shown inFig. 1). Increasing the ciprofloxacin concentration from 0.4 to 3.2�g/ml (2 to 16 times the MIC) decreased the accumulation ofcolonies (Fig. 2A). Increasing the number of cells applied to agarincreased the number of mutants recovered (Fig. 2B), but thefraction of cells recovered was roughly constant at a given concen-tration of ciprofloxacin (Fig. 2C; the slight decline in colony num-ber seen with large inocula probably arose from crowding of col-onies interfering with growth). Approximately 100 colonies wereobtained from ciprofloxacin-containing agar, regrown on drug-free agar, and then retested on agar containing ciprofloxacin at thesame concentration initially used to obtain the colonies. All colo-nies regrew rapidly (data not shown), which indicated that thecolonies recovered from the initial plating contained resistant mu-tants. Nucleotide sequence analysis of DNA isolated from coloniesrecovered from ciprofloxacin-containing agar plates after incuba-
tion for 10 days revealed alterations in gyrA and gyrB (Table 3;mutants were from the experiment shown in Fig. 2A). Ten mu-tants whose colonies were visible only after a week of incubationregrew at the same rate as wild-type cells (data not shown), indi-cating that their late recovery from agar was not due to slowgrowth of mutants present prior to drug exposure. A recA muta-tion restricted the increase in mutant colony recovery (Fig. 2D),consistent with involvement of the SOS response in mutant in-duction (4). Collectively, these data indicate that fluoroquinolo-nes induce resistance in M. smegmatis.
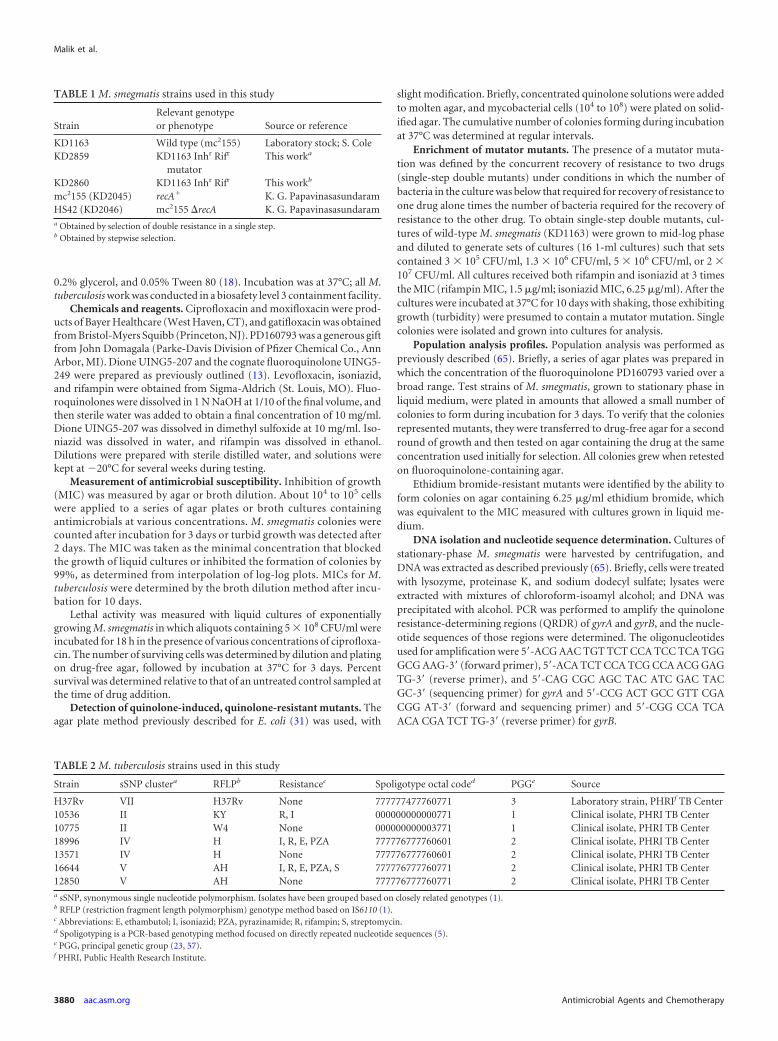
Effect of drug structure on recovery of M. smegmatis mu-tants. M. smegmatis was plated on agar containing a fluoroquin-olone (moxifloxacin, gatifloxacin, levofloxacin, or ciprofloxacin)at 2 times the MIC, and then colonies were scored at daily intervalsfor 2 weeks. The relative order for restricting the recovery of in-duced mutants was moxifloxacin � gatifloxacin � levofloxacin �ciprofloxacin (Fig. 3A). Similar results were observed when M.smegmatis was plated on agar containing drugs at several otherconcentrations (data not shown).
To determine whether an 8-methoxy-quinazoline-2,4-dione ismore effective than a comparable fluoroquinolone at restrictingthe recovery of induced mutants, we plated M. smegmatis on agar
FIG 1 Structures of the compounds used in this study.
Induced Resistance in Mycobacteria
July 2012 Volume 56 Number 7 aac.asm.org 3881
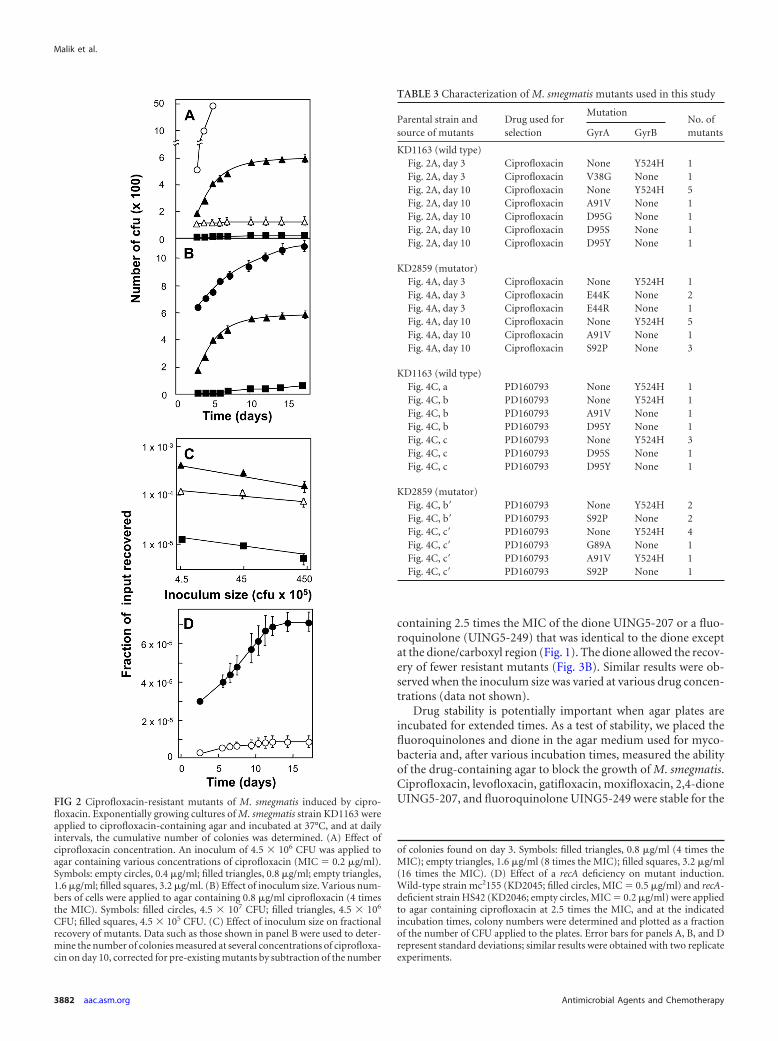
containing 2.5 times the MIC of the dione UING5-207 or a fluo-roquinolone (UING5-249) that was identical to the dione exceptat the dione/carboxyl region (Fig. 1). The dione allowed the recov-ery of fewer resistant mutants (Fig. 3B). Similar results were ob-served when the inoculum size was varied at various drug concen-trations (data not shown).
Drug stability is potentially important when agar plates areincubated for extended times. As a test of stability, we placed thefluoroquinolones and dione in the agar medium used for myco-bacteria and, after various incubation times, measured the abilityof the drug-containing agar to block the growth of M. smegmatis.Ciprofloxacin, levofloxacin, gatifloxacin, moxifloxacin, 2,4-dioneUING5-207, and fluoroquinolone UING5-249 were stable for theFIG 2 Ciprofloxacin-resistant mutants of M. smegmatis induced by cipro-
floxacin. Exponentially growing cultures of M. smegmatis strain KD1163 wereapplied to ciprofloxacin-containing agar and incubated at 37°C, and at dailyintervals, the cumulative number of colonies was determined. (A) Effect ofciprofloxacin concentration. An inoculum of 4.5 � 106 CFU was applied toagar containing various concentrations of ciprofloxacin (MIC � 0.2 �g/ml).Symbols: empty circles, 0.4 �g/ml; filled triangles, 0.8 �g/ml; empty triangles,1.6 �g/ml; filled squares, 3.2 �g/ml. (B) Effect of inoculum size. Various num-bers of cells were applied to agar containing 0.8 �g/ml ciprofloxacin (4 timesthe MIC). Symbols: filled circles, 4.5 � 107 CFU; filled triangles, 4.5 � 106
CFU; filled squares, 4.5 � 105 CFU. (C) Effect of inoculum size on fractionalrecovery of mutants. Data such as those shown in panel B were used to deter-mine the number of colonies measured at several concentrations of ciprofloxa-cin on day 10, corrected for pre-existing mutants by subtraction of the number
of colonies found on day 3. Symbols: filled triangles, 0.8 �g/ml (4 times theMIC); empty triangles, 1.6 �g/ml (8 times the MIC); filled squares, 3.2 �g/ml(16 times the MIC). (D) Effect of a recA deficiency on mutant induction.Wild-type strain mc2155 (KD2045; filled circles, MIC � 0.5 �g/ml) and recA-deficient strain HS42 (KD2046; empty circles, MIC � 0.2 �g/ml) were appliedto agar containing ciprofloxacin at 2.5 times the MIC, and at the indicatedincubation times, colony numbers were determined and plotted as a fractionof the number of CFU applied to the plates. Error bars for panels A, B, and Drepresent standard deviations; similar results were obtained with two replicateexperiments.
TABLE 3 Characterization of M. smegmatis mutants used in this study
Parental strain andsource of mutants
Drug used forselection
MutationNo. ofmutantsGyrA GyrB
KD1163 (wild type)Fig. 2A, day 3 Ciprofloxacin None Y524H 1Fig. 2A, day 3 Ciprofloxacin V38G None 1Fig. 2A, day 10 Ciprofloxacin None Y524H 5Fig. 2A, day 10 Ciprofloxacin A91V None 1Fig. 2A, day 10 Ciprofloxacin D95G None 1Fig. 2A, day 10 Ciprofloxacin D95S None 1Fig. 2A, day 10 Ciprofloxacin D95Y None 1
KD2859 (mutator)Fig. 4A, day 3 Ciprofloxacin None Y524H 1Fig. 4A, day 3 Ciprofloxacin E44K None 2Fig. 4A, day 3 Ciprofloxacin E44R None 1Fig. 4A, day 10 Ciprofloxacin None Y524H 5Fig. 4A, day 10 Ciprofloxacin A91V None 1Fig. 4A, day 10 Ciprofloxacin S92P None 3
KD1163 (wild type)Fig. 4C, a PD160793 None Y524H 1Fig. 4C, b PD160793 None Y524H 1Fig. 4C, b PD160793 A91V None 1Fig. 4C, b PD160793 D95Y None 1Fig. 4C, c PD160793 None Y524H 3Fig. 4C, c PD160793 D95S None 1Fig. 4C, c PD160793 D95Y None 1
KD2859 (mutator)Fig. 4C, b= PD160793 None Y524H 2Fig. 4C, b= PD160793 S92P None 2Fig. 4C, c= PD160793 None Y524H 4Fig. 4C, c= PD160793 G89A None 1Fig. 4C, c= PD160793 A91V Y524H 1Fig. 4C, c= PD160793 S92P None 1
Malik et al.
3882 aac.asm.org Antimicrobial Agents and Chemotherapy
duration of the experiment (Fig. 3C). These data indicate that thedifferences among the compounds described above are due todifferences in activity, not stability.
Enrichment of M. smegmatis mutator mutant. To determinewhether mycobacterial mutators can be detected by the agar plateassay, we first prepared spontaneous mutator strains by requiringgrowth of M. smegmatis in the presence of two antimicrobials(rifampin and isoniazid). Conditions were used in which resistantgrowth was unlikely to occur in liquid cultures unless a mutatorwas present to increase the frequency of rifampin and isoniazidresistance (see Materials and Methods for details). When sets ofcultures were inoculated with 3 � 105 or 1.3 � 106 CFU, no cul-ture (0/16 at each inoculum size) became turbid following incu-bation at 37°C. However, at an inoculum of 5 � 106 CFU, 7 of 16cultures exhibited growth; at 2 � 107 CFU, 16 of 16 cultures grew.The cell numbers at which growth occurred were below the num-ber needed to obtain isoniazid and rifampin resistance concur-rently in the absence of a mutator [(frif � finh) � (2 � 10�5) �(3 � 10�6) � 6 � 10�11, where f is the frequency of resistance](20, 50; data not shown).
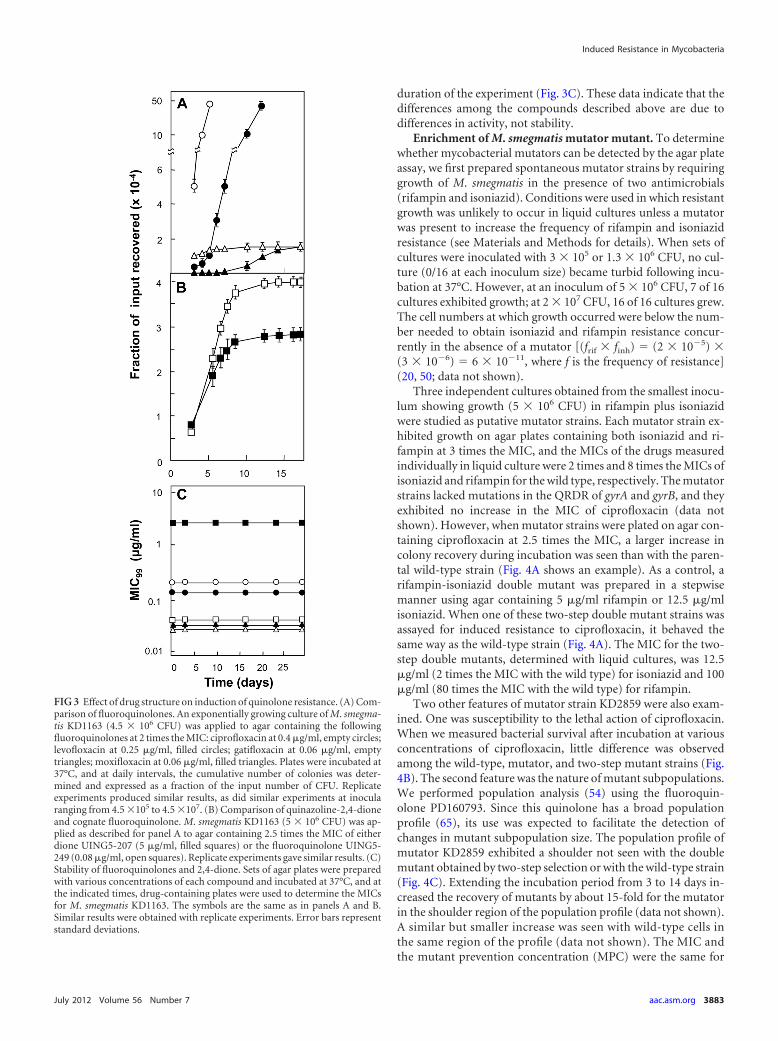
Three independent cultures obtained from the smallest inocu-lum showing growth (5 � 106 CFU) in rifampin plus isoniazidwere studied as putative mutator strains. Each mutator strain ex-hibited growth on agar plates containing both isoniazid and ri-fampin at 3 times the MIC, and the MICs of the drugs measuredindividually in liquid culture were 2 times and 8 times the MICs ofisoniazid and rifampin for the wild type, respectively. The mutatorstrains lacked mutations in the QRDR of gyrA and gyrB, and theyexhibited no increase in the MIC of ciprofloxacin (data notshown). However, when mutator strains were plated on agar con-taining ciprofloxacin at 2.5 times the MIC, a larger increase incolony recovery during incubation was seen than with the paren-tal wild-type strain (Fig. 4A shows an example). As a control, arifampin-isoniazid double mutant was prepared in a stepwisemanner using agar containing 5 �g/ml rifampin or 12.5 �g/mlisoniazid. When one of these two-step double mutant strains wasassayed for induced resistance to ciprofloxacin, it behaved thesame way as the wild-type strain (Fig. 4A). The MIC for the two-step double mutants, determined with liquid cultures, was 12.5�g/ml (2 times the MIC with the wild type) for isoniazid and 100�g/ml (80 times the MIC with the wild type) for rifampin.
Two other features of mutator strain KD2859 were also exam-ined. One was susceptibility to the lethal action of ciprofloxacin.When we measured bacterial survival after incubation at variousconcentrations of ciprofloxacin, little difference was observedamong the wild-type, mutator, and two-step mutant strains (Fig.4B). The second feature was the nature of mutant subpopulations.We performed population analysis (54) using the fluoroquin-olone PD160793. Since this quinolone has a broad populationprofile (65), its use was expected to facilitate the detection ofchanges in mutant subpopulation size. The population profile ofmutator KD2859 exhibited a shoulder not seen with the doublemutant obtained by two-step selection or with the wild-type strain(Fig. 4C). Extending the incubation period from 3 to 14 days in-creased the recovery of mutants by about 15-fold for the mutatorin the shoulder region of the population profile (data not shown).A similar but smaller increase was seen with wild-type cells inthe same region of the profile (data not shown). The MIC andthe mutant prevention concentration (MPC) were the same for
FIG 3 Effect of drug structure on induction of quinolone resistance. (A) Com-parison of fluoroquinolones. An exponentially growing culture of M. smegma-tis KD1163 (4.5 � 106 CFU) was applied to agar containing the followingfluoroquinolones at 2 times the MIC: ciprofloxacin at 0.4 �g/ml, empty circles;levofloxacin at 0.25 �g/ml, filled circles; gatifloxacin at 0.06 �g/ml, emptytriangles; moxifloxacin at 0.06 �g/ml, filled triangles. Plates were incubated at37°C, and at daily intervals, the cumulative number of colonies was deter-mined and expressed as a fraction of the input number of CFU. Replicateexperiments produced similar results, as did similar experiments at inocularanging from 4.5 �105 to 4.5 �107. (B) Comparison of quinazoline-2,4-dioneand cognate fluoroquinolone. M. smegmatis KD1163 (5 � 106 CFU) was ap-plied as described for panel A to agar containing 2.5 times the MIC of eitherdione UING5-207 (5 �g/ml, filled squares) or the fluoroquinolone UING5-249 (0.08 �g/ml, open squares). Replicate experiments gave similar results. (C)Stability of fluoroquinolones and 2,4-dione. Sets of agar plates were preparedwith various concentrations of each compound and incubated at 37°C, and atthe indicated times, drug-containing plates were used to determine the MICsfor M. smegmatis KD1163. The symbols are the same as in panels A and B.Similar results were obtained with replicate experiments. Error bars representstandard deviations.
Induced Resistance in Mycobacteria
July 2012 Volume 56 Number 7 aac.asm.org 3883
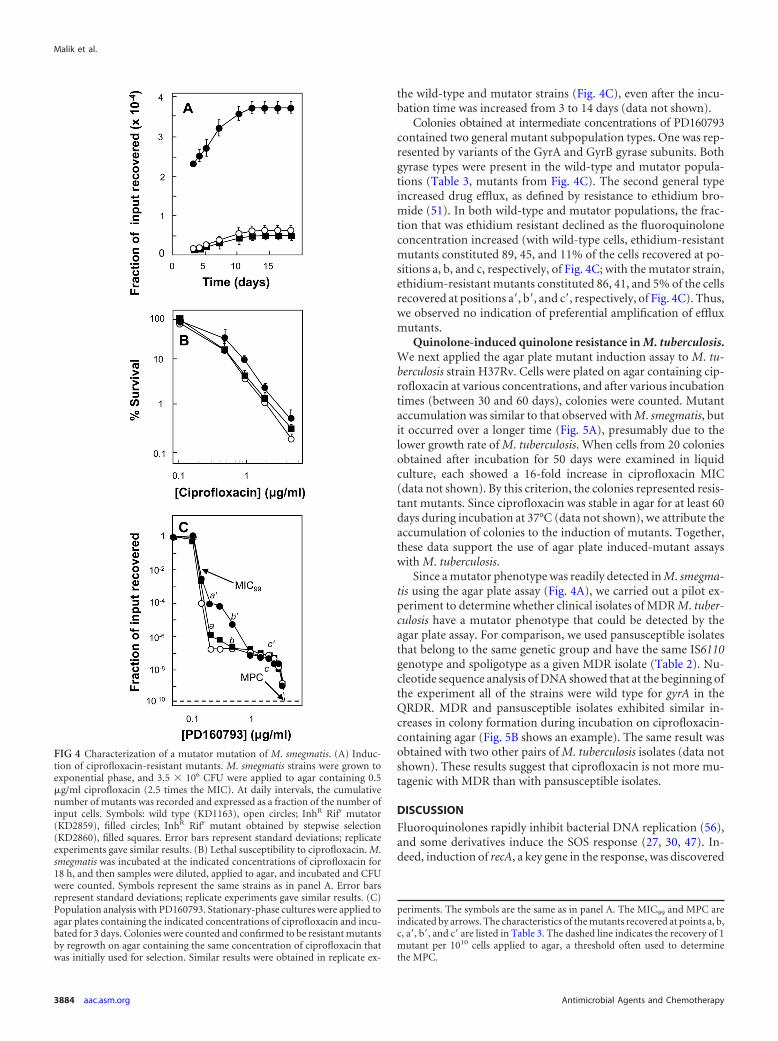
the wild-type and mutator strains (Fig. 4C), even after the incu-bation time was increased from 3 to 14 days (data not shown).
Colonies obtained at intermediate concentrations of PD160793contained two general mutant subpopulation types. One was rep-resented by variants of the GyrA and GyrB gyrase subunits. Bothgyrase types were present in the wild-type and mutator popula-tions (Table 3, mutants from Fig. 4C). The second general typeincreased drug efflux, as defined by resistance to ethidium bro-mide (51). In both wild-type and mutator populations, the frac-tion that was ethidium resistant declined as the fluoroquinoloneconcentration increased (with wild-type cells, ethidium-resistantmutants constituted 89, 45, and 11% of the cells recovered at po-sitions a, b, and c, respectively, of Fig. 4C; with the mutator strain,ethidium-resistant mutants constituted 86, 41, and 5% of the cellsrecovered at positions a=, b=, and c=, respectively, of Fig. 4C). Thus,we observed no indication of preferential amplification of effluxmutants.
Quinolone-induced quinolone resistance in M. tuberculosis.We next applied the agar plate mutant induction assay to M. tu-berculosis strain H37Rv. Cells were plated on agar containing cip-rofloxacin at various concentrations, and after various incubationtimes (between 30 and 60 days), colonies were counted. Mutantaccumulation was similar to that observed with M. smegmatis, butit occurred over a longer time (Fig. 5A), presumably due to thelower growth rate of M. tuberculosis. When cells from 20 coloniesobtained after incubation for 50 days were examined in liquidculture, each showed a 16-fold increase in ciprofloxacin MIC(data not shown). By this criterion, the colonies represented resis-tant mutants. Since ciprofloxacin was stable in agar for at least 60days during incubation at 37°C (data not shown), we attribute theaccumulation of colonies to the induction of mutants. Together,these data support the use of agar plate induced-mutant assayswith M. tuberculosis.
Since a mutator phenotype was readily detected in M. smegma-tis using the agar plate assay (Fig. 4A), we carried out a pilot ex-periment to determine whether clinical isolates of MDR M. tuber-culosis have a mutator phenotype that could be detected by theagar plate assay. For comparison, we used pansusceptible isolatesthat belong to the same genetic group and have the same IS6110genotype and spoligotype as a given MDR isolate (Table 2). Nu-cleotide sequence analysis of DNA showed that at the beginning ofthe experiment all of the strains were wild type for gyrA in theQRDR. MDR and pansusceptible isolates exhibited similar in-creases in colony formation during incubation on ciprofloxacin-containing agar (Fig. 5B shows an example). The same result wasobtained with two other pairs of M. tuberculosis isolates (data notshown). These results suggest that ciprofloxacin is not more mu-tagenic with MDR than with pansusceptible isolates.
DISCUSSION
Fluoroquinolones rapidly inhibit bacterial DNA replication (56),and some derivatives induce the SOS response (27, 30, 47). In-deed, induction of recA, a key gene in the response, was discovered
FIG 4 Characterization of a mutator mutation of M. smegmatis. (A) Induc-tion of ciprofloxacin-resistant mutants. M. smegmatis strains were grown toexponential phase, and 3.5 � 106 CFU were applied to agar containing 0.5�g/ml ciprofloxacin (2.5 times the MIC). At daily intervals, the cumulativenumber of mutants was recorded and expressed as a fraction of the number ofinput cells. Symbols: wild type (KD1163), open circles; InhR Rifr mutator(KD2859), filled circles; InhR Rifr mutant obtained by stepwise selection(KD2860), filled squares. Error bars represent standard deviations; replicateexperiments gave similar results. (B) Lethal susceptibility to ciprofloxacin. M.smegmatis was incubated at the indicated concentrations of ciprofloxacin for18 h, and then samples were diluted, applied to agar, and incubated and CFUwere counted. Symbols represent the same strains as in panel A. Error barsrepresent standard deviations; replicate experiments gave similar results. (C)Population analysis with PD160793. Stationary-phase cultures were applied toagar plates containing the indicated concentrations of ciprofloxacin and incu-bated for 3 days. Colonies were counted and confirmed to be resistant mutantsby regrowth on agar containing the same concentration of ciprofloxacin thatwas initially used for selection. Similar results were obtained in replicate ex-
periments. The symbols are the same as in panel A. The MIC99 and MPC areindicated by arrows. The characteristics of the mutants recovered at points a, b,c, a=, b=, and c= are listed in Table 3. The dashed line indicates the recovery of 1mutant per 1010 cells applied to agar, a threshold often used to determinethe MPC.
Malik et al.
3884 aac.asm.org Antimicrobial Agents and Chemotherapy
in part by treating cells with nalidixic acid (16, 17), the prototypeof this antimicrobial class. Since error-prone repair occurs duringthe SOS response (55), the quinolones are generally considered tobe mutagenic (15, 31, 34, 36, 46, 49). Differences in the inductionof mutants among structural variants of the fluoroquinoloneswere easily seen with M. smegmatis. When the compounds werepresent at comparable concentrations (2 to 3 times the MIC),gatifloxacin and moxifloxacin allowed the recovery of fewer in-duced mutants than levofloxacin, which in turn was more restric-tive than ciprofloxacin (Fig. 3A). A quinazoline-2,4-dione thatlacks critical structural features of the quinolone pharmacophorewas more restrictive than the cognate fluoroquinolone (Fig. 3B).Thus, the agar plate mutant accumulation assay can be used toevaluate quinolones and related compounds for the ability to gen-erate resistance during drug exposure.
The agar plate assay tests the combined effects of at least threefactors: survival of the parental population, induction of the SOSresponse, and growth of resistant mutants. Since the size of theparental population affects whether a new mutation will occur,bactericidal activity is important. Quinolones are “concentration-
dependent killers”; consequently, higher concentrations are morerestrictive for mutant induction due to lethal activity (Fig. 2A).Likewise, aspects of drug structure that increase lethal activity willrestrict the induction and recovery of resistant mutants. With myco-bacteria, appending a C-8-methoxy group to the quinolone structureimproves lethal activity against both wild-type and resistant cells (9,32), which explains why gatifloxacin and moxifloxacin treatment re-sulted in the recovery of fewer mutants (Fig. 3A).
Quinolone-mediated DNA damage induces the SOS responsethrough the activation of RecA (28), and increased expression ofRecA has been taken as a marker of induction (48). Inductionoccurs over a broad range of quinolone concentrations (16, 48),with maximal induction correlating with the MIC (48). Twosources of DNA damage are likely to be involved. One probablyarises directly from the formation of drug-gyrase/topoisomeraseIV-DNA complexes blocking DNA replication (43, 48). The otherderives from the cascade of reactive oxygen species that is as-sociated with lethal antimicrobials (22, 59). The final step, ac-cumulation of hydroxyl radical, damages DNA and would ex-plain how ampicillin can induce the SOS response (35). Wenote that some very bactericidal fluoroquinolones kill cellswithout a contribution from hydroxyl radical (60). Additionalwork is needed to fully define the role of drug structure in theinduction of the SOS regulon.
Drug structure also affects the propensity of a given compoundto selectively restrict the growth of resistant mutants. We havetermed this propensity antimutant activity, which is defined as theratio of the MIC for mutant cells to the MIC for wild-type cells (13,33). If mutant growth is restricted, resistance is less likely toemerge. The presence of an 8-methoxy group on fluoroquinolo-nes contributes to increased antimutant activity, as does replace-ment of the 3-carboxylate group of quinolones with the dionemotif in quinazoline-2,4-diones (Fig. 3) (13, 33). X-ray structuresof drug-topoisomerase-DNA complexes (2, 24, 25, 61) suggestthat the preferential activity of quinazoline-2,4-diones with gyrAmutants may be due to (i) the dione binding at the DNA nick sitebeing farther from the common resistance residues and (ii) bounddione not requiring magnesium coordination with aspartic acid95 (M. smegmatis numbering system [24, 33]). The latter reducesthe effect of substitutions at position 95 on the MIC. Thus, findingthat the 2,4-dione restricts the recovery of induced mutants onagar plates is consistent with structural analyses. Since the lethalactivity of 2,4-diones is similar to that of cognate fluoroquinolo-nes with E. coli when concentrations are normalized to the MIC(13, 33), lethal activity may contribute little to dione UING5-207allowing the recovery of fewer induced mutants than the cognatefluoroquinolone.
Mutator strains exhibit an elevated mutation frequency andare a significant factor in the emergence of resistance during in-fection with some Gram-negative bacteria (29, 44, 45, 63). Herewe obtained spontaneous mutators in cultures of M. smegmatis byforcing growth in the presence of both rifampin and isoniazid. Theresulting cells exhibited elevated induction of ciprofloxacin resis-tance (Fig. 4A). The population profiles of mutator strains ofPseudomonas aeruginosa show an overall shift to a higher quino-lone concentration and an increase in the mutant subpopulationsize (39). With M. smegmatis, only part of the population profilewas shifted (Fig. 4C). Thus, the effects of mutators can vary. Nu-cleotide sequence analysis revealed alterations in M. smegmatisgyrA and gyrB at these intermediate fluoroquinolone concentra-
FIG 5 Ciprofloxacin-resistant mutants of M. tuberculosis induced by cipro-floxacin. (A) Exponentially growing cultures of M. tuberculosis H37Rv (1.5 �107 CFU) were applied to ciprofloxacin-containing agar and incubated at37°C. At the indicated times, the number of colonies was determined. Symbols:filled triangles, 0.3 �g/ml (1.5 times the MIC); filled circles, 0.4 �g/ml (2 timesthe MIC); open circles, 0.5 �g/ml (2.5 times the MIC); filled squares, 0.6 �g/ml(3 times the MIC); open squares, 0.8 �g/ml (4 times the MIC). (B) Inductionof ciprofloxacin-resistant mutants by pansusceptible and MDR clinical iso-lates. Cultures (2.5 � 106 CFU) of a pansusceptible isolate (strain 12850, filledsquares) and an MDR isolate (strain 16644, empty squares) were applied toagar containing ciprofloxacin at 2 times the MIC and incubated at 37°C. At theindicated times, the cumulative number of colonies was determined and ex-pressed as a fraction of the number of input cells. Similar results were obtainedwith two other pairs of pansusceptible and MDR isolates (strains 10775 and10536 and strains 13571 and 18996). Error bars represent standard deviations;replicate experiments gave similar results.
Induced Resistance in Mycobacteria
July 2012 Volume 56 Number 7 aac.asm.org 3885

tions (Table 3). A more extensive analysis is required to determinewhether particular gyrase mutants are enriched. Ethidium bro-mide resistance, which was taken as a marker for mutants havingelevated efflux, showed no difference in frequency between muta-tor and wild-type populations.
Effects of mutators can be interpreted within the framework ofthe mutant selection window hypothesis (10, 64), which postu-lates that resistant mutant subpopulations are enriched by drugtreatment within a specific drug concentration range (roughly be-tween the MIC and the MPC, where the MPC is the MIC of theleast susceptible mutant subpopulation). With short incubationtimes, population profiles are expected to reflect mutant sub-populations present prior to drug exposure. In a study of P.aeruginosa, mutator mutants exhibited increased MICs, shouldersin the population profiles, and increased MPCs (39). Thus, themutant selection window for a P. aeruginosa mutator is populatedby a larger subpopulation of resistant mutants, and the MICs for atleast some of the mutants are elevated. In the present study withM. smegmatis, the subpopulation size was increased (shoulder inthe profile shown in Fig. 4C) without changes in the susceptibilityof the least susceptible mutant subpopulation (without changes in theMPC). Increasing the incubation time to allow the scoring of inducedmutants also increased only the subpopulation size and not the MPC(data not shown). These data are consistent with the M. smegmatismutator generating more of the same mutant subpopulations thatwere present in the wild-type population (Table 3).
Fluoroquinolones also induce resistance in M. tuberculosis(Fig. 5). No mutator phenotype was observed when we applied theassay to three MDR clinical isolates. Previous work reported thatclinical isolates of the M. tuberculosis W-Beijing lineage containalterations in orthologues of mutator genes of other bacteria (11,26). However, no evidence of mutator activity was detected. Otherwork showed that the DnaE2 polymerase is induced by UV irra-diation and that the absence of DnaE2 lowers the UV-mediatedmutation frequency (4). Moreover, DnaE2 is important for theacquisition of rifampin resistance in infected mice. If the DnaE2polymerase is a mutator, a dnaE2 deficiency should reduce theaccumulation of mutants in the agar plate assay used in Fig. 5,especially following UV treatment.
In conclusion, to minimize the induction of new resistantmutants and to restrict the amplification of pre-existing mu-tant subpopulations, we suggest that TB treatment regimenskeep quinolone concentrations high and avoid agents that arenot highly effective, such as ciprofloxacin, ofloxacin, and levo-floxacin. Keeping activity high is important even when otheractive antimicrobials are present, since pharmacokinetic dif-ferences can create times at which the equivalent of mono-therapy occurs. Whether mutators in M. tuberculosis can beenriched by the methods used with M. smegmatis is not known.In preliminary work with Mycobacterium bovis BCG, we failedto recover mutators at cell densities 1,000 times higher thanthose used for M. smegmatis. Thus, the genome of M. bovis, andpresumably that of M. tuberculosis, may be exceptionally stable.Such a conclusion for M. tuberculosis is supported by (i) thepaucity of synonymous (silent) mutations in structural genes(40, 41, 57) and (ii) the failure of sequential isolates obtainedduring the development of MDR TB to show mutations otherthan those in the resistance genes (53).
ACKNOWLEDGMENTS
We thank Marila Gennaro and Bo Shopsin for critical comments on themanuscript.
This work was supported by NIH grants AI73491 and AI087671.
REFERENCES1. Barnes P, Cave M. 2003. Molecular epidemiology of tuberculosis. N.
Engl. J. Med. 349:1149 –1156.2. Bax B, et al. 2010. Type IIA topoisomerase inhibition by a new class of
antibacterial agents. Nature 466:935–940.3. Bonilla C, et al. 2008. Management of extensively drug-resistant tuber-
culosis in Peru: cure is possible. PLoS One 3:e2957.4. Boshoff H, Reed M, Barry C, Mizrahi V. 2003. DnaE2 polymerase
contributes to in vivo survival and the emergence of drug resistance inMycobacterium tuberculosis. Cell 113:183–193.
5. Brudey K, et al. 2006. Mycobacterium tuberculosis complex genetic diver-sity: mining the fourth international spoligotyping database (SpolDB4)for classification, population genetics and epidemiology. BMC Microbiol.6:23.
6. Caminero J, Sotgiu G, Zumla A, Migliori G. 2010. Best drug treatmentfor multidrug-resistant and extensively drug-resistant tuberculosis. Lan-cet Infect. Dis. 10:621– 629.
7. Cirz R, et al. 2005. Inhibition of mutation and combating the evolution ofantibiotic resistance. PLoS Biol. 3:e176.
8. Cirz R, Romesberg F. 2006. Induction and inhibition of ciprofloxacinresistance-conferring mutations in hypermutator bacteria. Antimicrob.Agents Chemother. 50:220 –225.
9. Dong Y, Xu C, Zhao X, Domagala J, Drlica K. 1998. Fluoroquinoloneaction against mycobacteria: effects of C8 substituents on bacterialgrowth, survival, and resistance. Antimicrob. Agents Chemother. 42:2978 –2984.
10. Drlica K, Zhao X. 2007. Mutant selection window hypothesis updated.Clin. Infect. Dis. 44:681– 688.
11. Ebrahimi-Rad M, et al. 2003. Mutations in putative mutator genes ofMycobacterium tuberculosis strains of the W-Beijing family. Emerg. Infect.Dis. 9:838 – 845.
12. Gandhi NR, et al. 2006. Extensively drug-resistant tuberculosis as a causeof death in patients co-infected with tuberculosis and HIV in a rural areaof South Africa. Lancet 368:1575–1580.
13. German N, Malik M, Rosen J, Drlica K, Kerns R. 2008. Use of gyraseresistance mutants to guide selection of 8-methoxy-quinazoline-2,4-diones. Antimicrob. Agents Chemother. 52:3915–3921.
14. Ginsberg A. 2010. Drugs in development for tuberculosis. Drugs 70:2201–2214.
15. Gocke E. 1991. Mechanism of quinolone mutagenicity in bacteria. Mutat.Res. 248:135–143.
16. Gudas LJ, Pardee AB. 1976. DNA synthesis inhibition and the inductionof protein X in Escherichia coli. J. Mol. Biol. 101:459 – 477.
17. Inouye M, Pardee A. 1970. Changes of membrane proteins and theirrelation to deoxyribonucleic acid synthesis and cell division of Escherichiacoli. J. Biol. Chem. 245:5813–5819.
18. Jacobs WR, et al. 1991. Genetic systems in mycobacteria. Methods En-zymol. 204:537–555.
19. Jacobson K, Tierney D, Jeon C, Mitnick C, Murray M. 2010. Treatmentoutcomes among patients with extensively drug-resistant tuberculosis:systematic review and meta-analysis. Clin. Infect. Dis. 51:6 –14.
20. Karunakaran P, Davies J. 2000. Genetic antagonism and hypermutabilityin Mycobacterium smegmatis. J. Bacteriol. 182:3331–3335.
21. Kim S, et al. 1997. Multiple pathways for SOS-induced mutagenesis inEscherichia coli: an overexpression of dinB/dinP results in strongly enhanc-ing mutagenesis in the absence of any exogenous treatment to damageDNA. Proc. Natl. Acad. Sci. U. S. A. 94:13792–13797.
22. Kohanski M, Dwyer D, Hayete B, Lawrence C, Collins J. 2007. Acommon mechanism of cellular death induced by bactericidal antibiotics.Cell 130:797– 810.
23. Kong Y, et al. 2007. Association between Mycobacterium tuberculosisBeijing/W lineage strain infection and extrathoracic tuberculosis: insightsfrom epidemiologic and clinical characterization of the three principalgenetic groups of M. tuberculosis clinical isolates. J. Clin. Microbiol. 45:409 – 414.
24. Laponogov I, et al. 2010. Structural basis of gate-DNA breakage andresealing by type II topoisomerases. PLoS One 5:e11338.
Malik et al.
3886 aac.asm.org Antimicrobial Agents and Chemotherapy

25. Laponogov I, et al. 2009. Structural insight into the quinolone-DNAcleavage complex of type IIA topoisomerases. Nat. Struct. Mol. Biol. 16:667– 669.
26. Lari N, Rindi L, Bonanni D, Tortolli E, Garzelli C. 2006. Mutations inmutT genes of Mycobacterium tuberculosis isolates of Beijing genotype. J.Med. Microbiol. 55:599 – 603.
27. Lewin C, Howard B, Ratcliffe N, Smith J. 1989. 4-Quinolones and theSOS response. J. Med. Microbiol. 29:139 –144.
28. Little J. 1993. LexA cleavage and other self-processing reactions. J. Bacte-riol. 175:4943– 4950.
29. Maciá M, et al. 2005. Hypermutation is a key factor in development ofmultiple-antimicrobial resistance in Pseudomonas aeruginosa strains caus-ing chronic lung infections. Antimicrob. Agents Chemother. 49:3382–3386.
30. Maguin E, Lutkenhaus J, D’Ari R. 1986. Reversibility of SOS-associateddivision inhibition in Escherichia coli. J. Bacteriol. 166:733–738.
31. Malik M, Hoatam G, Chavda K, Kerns R, Drlica K. 2010. Novelapproach for comparing abilities of quinolones to restrict the emergenceof resistant mutants during quinolone exposure. Antimicrob. Agents Che-mother. 54:149 –156.
32. Malik M, et al. 2005. Lethality of quinolones against Mycobacteriumsmegmatis in the presence or absence of chloramphenicol. Antimicrob.Agents Chemother. 49:2008 –2014.
33. Malik M, et al. 2011. Fluoroquinolone and quinazolinedione activity withwild-type and gyrase mutants of Mycobacterium smegmatis. Antimicrob.Agents Chemother. 55:2335–2343.
34. Mamber S, Kolek B, Brookshire K, Bonner D, Fung-Tomc J. 1993.Activity of quinolones in the Ames Salmonella TA102 mutagenicity testand other bacterial genotoxicity assays. Antimicrob. Agents Chemother.37:213–217.
35. Miller C, et al. 2004. SOS response induction by -lactams and bacterialdefense against antibiotic lethality. Science 305:1629 –1631.
36. Miller JH, Low KB. 1984. Specificity of mutagenesis resulting from theinduction of the SOS system in the absence of mutagenic treatment. Cell37:675– 682.
37. Mitnick C, Castro K, Harrington M, Sacks L, Burman W. 2007. Ran-domized trials to optimize treatment of multidrug-resistant tuberculosis.PLoS Med. 4:e292.
38. Mitnick C, et al. 2008. Comprehensive treatment of extensively drug-resistant tuberculosis. N. Engl. J. Med. 359:563–574.
39. Morero N, Monti M, Argaraña CE. 2011. Effect of ciprofloxacin concen-tration on the frequency and nature of resistant mutants selected fromPseudomonas aeruginosa mutS and mutT hypermutators. Antimicrob.Agents Chemother. 55:3668 –3676.
40. Musser J. 2001. Single nucleotide polymorphisms in Mycobacterium tu-berculosis structural genes. Emerg. Infect. Dis. 7:486 – 488.
41. Musser J, Amin A, Ramaswamy S. 2000. Negligible genetic diversity ofMycobacterium tuberculosis host immune system protein targets: evidenceof limited selective pressure. Genetics 155:7–16.
42. Napolitano R, Janel-Bintz R, Wagner J, Fuchs R. 2000. All three SOS-inducible DNA polymerases (Pol II, Pol IV and Pol V) are involved ininduced mutagenesis. EMBO J. 19:6259 – 6265.
43. Oishi M, Smith CL, Friefeld B. 1979. Molecular events and moleculesthat lead to induction of prophage and SOS functions. Cold Spring HarborSymp. Quant. Biol. 43:897–907.
44. Oliver A, Canton R, Campo P, Baquero F, Blazquez J. 2000. Highfrequency of hypermutable Pseudomonas aeruginosa in cystic fibrosis lunginfection. Science 288:1251–1253.
45. Oliver A, Levin B, Juan C, Baquero F, Blazquez J. 2004. Hypermutationand the preexistence of antibiotic-resistant Pseudomonas aeruginosa mu-
tants: implications for susceptibility testing and treatment of chronic in-fections. Antimicrob. Agents Chemother. 48:4226 – 4233.
46. Phillips I. 1987. Bacterial mutagenicity and the 4-quinolones. J. Antimi-crob. Chemother. 20:771–773.
47. Piddock LJV, Wise R. 1987. Induction of the SOS response in Escherichiacoli by 4-quinolone antimicrobial agents. FEMS Microbiol. Lett. 41(3):289 –294.
48. Piddock LJ, Walters RN, Diver JM. 1990. Correlation of quinolone MICand inhibition of DNA, RNA, and protein synthesis and induction of theSOS response in Escherichia coli. Antimicrob. Agents Chemother. 34:2331–2336.
49. Power E, Phillips I. 1993. Correlation between umuC induction andSalmonella mutagenicity assay for quinolone antimicrobial agents. FEMSMicrobiol. Lett. 112:251–254.
50. Quan S, Venter H, Dabbs E. 1997. Ribosylative inactivation of rifampinby Mycobacterium smegmatis is a principle contributor to its low suscep-tibility to this antibiotic. Antimicrob. Agents Chemother. 41:2456 –2460.
51. Rodrigues L, Ramos J, Couto I, Amaral L, Viveiros M. 2011. Ethidiumbromide transport across Mycobacterium smegmatis cell-wall: correlationwith antibiotic resistance. BMC Microbiol. 11:35.
52. Sanders L, Rockel A, Lu H, Wozniak D, Sutton M. 2006. Role ofPseudomonas aeruginosa dinB-encoded DNA polymerase IV in mutagen-esis. J. Bacteriol. 188:8573– 8585.
53. Saunders NJ, et al. 2011. Deep resequencing of serial sputum isolates ofMycobacterium tuberculosis during therapeutic failure due to poor com-pliance reveals stepwise mutation of key resistance genes on an otherwisestable genetic background. J. Infect. 62:212–217.
54. Sieradzki K, Roberts R, Haber S, Tomasz A. 1999. The development ofvancomycin resistance in a patient with methicillin-resistant Staphylococ-cus aureus infection. N. Engl. J. Med. 340:517–523.
55. Smith B, Walker G. 1998. Mutagenesis and more: umuDC and the Esch-erichia coli SOS response. Genetics 148:1599 –1610.
56. Snyder M, Drlica K. 1979. DNA gyrase on the bacterial chromosome:DNA cleavage induced by oxolinic acid. J. Mol. Biol. 131:287–302.
57. Sreevatsan S, et al. 1997. Restricted structural gene polymorphism in theMycobacterium tuberculosis complex indicates evolutionary recent globaldissemination. Proc. Natl. Acad. Sci. U. S. A. 94:9869 –9874.
58. Wang J, et al. 2010. Adding moxifloxacin is associated with a shorter timeto culture conversion in pulmonary tuberculosis. Int. J. Tuberc. Lung Dis.14:65–71.
59. Wang X, Zhao X. 2009. Contribution of oxidative damage to antimicro-bial lethality. Antimicrob. Agents Chemother. 53:1395–1402.
60. Wang X, Zhao X, Malik M, Drlica K. 2010. Contribution of reactiveoxygen species to pathways of quinolone-mediated bacterial cell death. J.Antimicrob. Chemother. 65:520 –524.
61. Wohlkonig A, et al. 2010. Structural basis of quinolone inhibition of typeIIA topoisomerases and target-mediated resistance. Nat. Struct. Mol. Biol.17:1152–1153.
62. World Health Organization. 2010. Multidrug and extensively drug-resistant TB (M/XDR-TB) 2010 global report on surveillance andresponse. World Health Organization, Geneva, Switzerland. http://whqlibdoc.who.int/publications/2010/9789241599191_eng.pdf.
63. Yang H, Wolff E, Kim M, Diep A, Miller J. 2004. Identification ofmutator genes and mutational pathways in Escherichia coli using a multi-copy cloning approach. Mol. Microbiol. 53:283–295.
64. Zhao X, Drlica K. 2001. Restricting the selection of antibiotic-resistantmutants: a general strategy derived from fluoroquinolone studies. Clin.Infect. Dis. 33(Suppl 3):S147–S156.
65. Zhou J, et al. 2000. Selection of antibiotic resistant bacterial mutants:allelic diversity among fluoroquinolone-resistant mutations. J. Infect. Dis.182:517–525.
Induced Resistance in Mycobacteria
July 2012 Volume 56 Number 7 aac.asm.org 3887




















