
Regulation of pri-miRNA processing by a long noncoding RNA transcribed from an ultraconserved region
26
Molecular Cell Short Article Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Region Julia Liz, 1 Anna Portela, 1 Marta Soler, 1 Antonio Go ´ mez, 1 Hui Ling, 2 Gracjan Michlewski, 3 George A. Calin, 2 So ` nia Guil, 1, * and Manel Esteller 1,4,5, * 1 Cancer Epigenetics and Biology Program (PEBC), Bellvitge Biomedical Research Institute (IDIBELL), L’Hospitalet, 08908 Barcelona, Catalonia, Spain 2 Experimental Therapeutics & Cancer Genetics, MD Anderson Cancer Center, Texas State University, Houston, TX 77030, USA 3 Wellcome Trust Centre for Cell Biology, University of Edinburgh, King’s Buildings, Edinburgh, EH9 3JR, UK 4 School of Medicine, Department of Physiological Sciences II, University of Barcelona, 08036 Barcelona, Catalonia, Spain 5 Institucio Catalana de Recerca i Estudis Avanc ¸ ats (ICREA), 08010 Barcelona, Catalonia, Spain *Correspondence: [email protected] (S.G.), [email protected] (M.E.) http://dx.doi.org/10.1016/j.molcel.2014.05.005 SUMMARY Noncoding RNAs (ncRNAs) control cellular programs by affecting protein-coding genes, but evidence increasingly points to their involvement in a network of ncRNA-ncRNA interactions. Here, we show that a long ncRNA, Uc.283+A, controls pri-miRNA process- ing. Regulation requires complementarity between the lower stem region of the pri-miR-195 transcript and an ultraconserved sequence in Uc.283+A, which prevents pri-miRNA cleavage by Drosha. Mutation of the site in either RNA molecule uncouples regulation in vivo and in vitro. We propose a model in which lower-stem strand invasion by Uc.283+A impairs microprocessor recognition and efficient pri-miRNA cropping. In addition to identifying a case of RNA- directed regulation of miRNA biogenesis, our study reveals regulatory networks involving different ncRNA classes of importance in cancer. INTRODUCTION Global transcriptomic analysis has revealed a growing number of highly diverse noncoding RNAs (ncRNAs) with active roles in gene regulation. It has been estimated that only 2% of the genome is transcribed into protein-coding RNAs (Kapranov et al., 2007), so the largest fraction of the transcriptome corresponds to non- coding transcripts, which have traditionally been grouped into classes according to their length and then subclassified on the basis of their structural features (Costa, 2005; Djebali et al., 2012). Among the small noncoding RNAs, the abundant class of miRNAs (19–25 nt) comprises ncRNAs that fine-tune gene expression in eukaryotic organisms through inhibitory interac- tions based on partial complementarity with target mRNAs (Bar- tel, 2009; Thomas et al., 2010). miRNAs usually function in a combinatorial manner and regulate a large proportion of coding transcripts (Friedman et al., 2009) and are thereby implicated in a range of biological processes and often deregulated in cancer (reviewed in Garzon et al., 2009). miRNA biogenesis is a multilay- ered process that involves specific pathways to render mature, functional molecules (reviewed in Kim et al., 2009; Krol et al., 2010). The vast majority of miRNAs are transcribed by RNA Pol II, either from independent transcriptional units or embedding within the intronic or exonic sequence of a host gene, and their transcription may be subject to the same regulatory epigenetic processes of protein-coding genes (Lujambio et al., 2007; Saito et al., 2006). Transcription of miRNAs produces a long, capped, and polyadenylated primary molecule (the pri-miRNA) that folds into hairpins and is a substrate for the RNase III family enzyme Drosha that releases a 70 nt pre-miRNA that is exported to the cytoplasm and further cropped by Dicer. This cropping gen- erates a 20 bp duplex, from which usually only one strand, the mature miRNA, is incorporated into the RISC silencing complex. Regulation of miRNA processing at the posttranscriptional level is present at each step in this pathway. Several proteins have been described that regulate pri-miRNA and pre-miRNA pro- cessing by direct interaction with Drosha and Dicer or by binding to miRNA precursors (Winter et al., 2009). The other main group of ncRNAs refers to the long ncRNAs (lncRNAs, arbitrarily >200 nt), which are broadly defined as lack- ing protein-coding potential but otherwise often display mRNA- like properties, including multiexonic gene structures and poly(A) tails. Having tissue- and development-specific expression pat- terns, lncRNAs are associated with a variety of regulatory roles, including chromatin-related functions (Rinn et al., 2007), and regulation of transcription and splicing (Bernard et al., 2010; Kino et al., 2010). One intriguing class of lncRNAs comprises tran- scribed ultraconserved regions (T-UCRs), which originate from genomic elements of near-perfect evolutionary conservation in many mammalian genomes (Bejerano et al., 2004; Katzman et al., 2007) and are often located in cancer-associated regions. The expression of several T-UCRs is altered in cancer (Calin et al., 2007; Mestdagh et al., 2010), but despite growing evidence about the importance of T-UCRs in oncogenesis, few attempts have so far been made to address their molecular mechanism of action. We had previously taken a pharmacological and genomic approach to identify the T-UCR Uc.283+A as being subject to Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc. 1 Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re- gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Regulation of pri-miRNA processing by a long noncoding RNA transcribed from an ultraconserved region

Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re-gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005
Molecular Cell
Short Article
Regulation of pri-miRNA Processingby a Long Noncoding RNA Transcribedfrom an Ultraconserved RegionJulia Liz,1 Anna Portela,1 Marta Soler,1 Antonio Gomez,1 Hui Ling,2 Gracjan Michlewski,3 George A. Calin,2 Sonia Guil,1,*and Manel Esteller1,4,5,*1Cancer Epigenetics and Biology Program (PEBC), Bellvitge Biomedical Research Institute (IDIBELL), L’Hospitalet, 08908 Barcelona,
Catalonia, Spain2Experimental Therapeutics & Cancer Genetics, MD Anderson Cancer Center, Texas State University, Houston, TX 77030, USA3Wellcome Trust Centre for Cell Biology, University of Edinburgh, King’s Buildings, Edinburgh, EH9 3JR, UK4School of Medicine, Department of Physiological Sciences II, University of Barcelona, 08036 Barcelona, Catalonia, Spain5Institucio Catalana de Recerca i Estudis Avancats (ICREA), 08010 Barcelona, Catalonia, Spain*Correspondence: [email protected] (S.G.), [email protected] (M.E.)
http://dx.doi.org/10.1016/j.molcel.2014.05.005
SUMMARY
Noncoding RNAs (ncRNAs) control cellular programsby affecting protein-coding genes, but evidenceincreasingly points to their involvement in a networkof ncRNA-ncRNA interactions. Here, we show that along ncRNA,Uc.283+A, controls pri-miRNA process-ing. Regulation requires complementarity betweenthe lower stem region of the pri-miR-195 transcriptand an ultraconserved sequence in Uc.283+A, whichprevents pri-miRNA cleavage by Drosha. Mutation ofthe site in either RNA molecule uncouples regulationin vivo and in vitro. We propose a model in whichlower-stem strand invasion by Uc.283+A impairsmicroprocessor recognition and efficient pri-miRNAcropping. In addition to identifying a case of RNA-directed regulation of miRNA biogenesis, our studyreveals regulatory networks involving differentncRNA classes of importance in cancer.
INTRODUCTION
Global transcriptomic analysis has revealed a growing number of
highlydiversenoncodingRNAs (ncRNAs)withactive roles in gene
regulation. It has been estimated that only 2% of the genome is
transcribed into protein-coding RNAs (Kapranov et al., 2007),
so the largest fraction of the transcriptome corresponds to non-
coding transcripts, which have traditionally been grouped into
classes according to their length and then subclassified on the
basis of their structural features (Costa, 2005;Djebali et al., 2012).
Among the small noncoding RNAs, the abundant class of
miRNAs (19–25 nt) comprises ncRNAs that fine-tune gene
expression in eukaryotic organisms through inhibitory interac-
tions based on partial complementarity with target mRNAs (Bar-
tel, 2009; Thomas et al., 2010). miRNAs usually function in a
combinatorial manner and regulate a large proportion of coding
transcripts (Friedman et al., 2009) and are thereby implicated in a
range of biological processes and often deregulated in cancer
(reviewed in Garzon et al., 2009). miRNA biogenesis is a multilay-
ered process that involves specific pathways to render mature,
functional molecules (reviewed in Kim et al., 2009; Krol et al.,
2010). The vast majority of miRNAs are transcribed by RNA Pol
II, either from independent transcriptional units or embedding
within the intronic or exonic sequence of a host gene, and their
transcription may be subject to the same regulatory epigenetic
processes of protein-coding genes (Lujambio et al., 2007; Saito
et al., 2006). Transcription of miRNAs produces a long, capped,
and polyadenylated primary molecule (the pri-miRNA) that folds
into hairpins and is a substrate for the RNase III family enzyme
Drosha that releases a �70 nt pre-miRNA that is exported to
the cytoplasm and further cropped by Dicer. This cropping gen-
erates a �20 bp duplex, from which usually only one strand, the
mature miRNA, is incorporated into the RISC silencing complex.
Regulation of miRNA processing at the posttranscriptional level
is present at each step in this pathway. Several proteins have
been described that regulate pri-miRNA and pre-miRNA pro-
cessing by direct interaction with Drosha and Dicer or by binding
to miRNA precursors (Winter et al., 2009).
The other main group of ncRNAs refers to the long ncRNAs
(lncRNAs, arbitrarily >200 nt), which are broadly defined as lack-
ing protein-coding potential but otherwise often display mRNA-
like properties, includingmultiexonic gene structures and poly(A)
tails. Having tissue- and development-specific expression pat-
terns, lncRNAs are associated with a variety of regulatory roles,
including chromatin-related functions (Rinn et al., 2007), and
regulation of transcription and splicing (Bernard et al., 2010;
Kinoet al., 2010).One intriguing classof lncRNAscomprises tran-
scribed ultraconserved regions (T-UCRs), which originate from
genomic elements of near-perfect evolutionary conservation in
many mammalian genomes (Bejerano et al., 2004; Katzman
et al., 2007) and are often located in cancer-associated regions.
The expression of several T-UCRs is altered in cancer (Calin et al.,
2007;Mestdaghet al., 2010), but despite growingevidenceabout
the importance of T-UCRs in oncogenesis, few attempts have so
far been made to address their molecular mechanism of action.
We had previously taken a pharmacological and genomic
approach to identify the T-UCR Uc.283+A as being subject to
Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc. 1
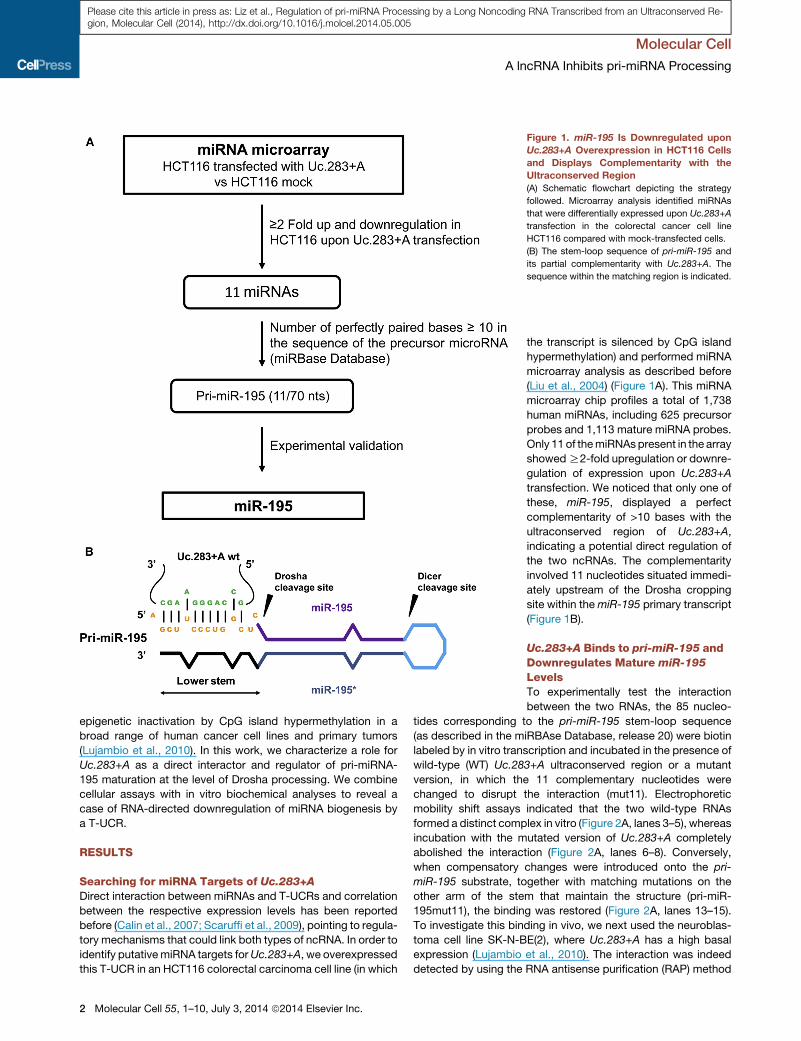
Figure 1. miR-195 Is Downregulated upon
Uc.283+A Overexpression in HCT116 Cells
and Displays Complementarity with the
Ultraconserved Region
(A) Schematic flowchart depicting the strategy
followed. Microarray analysis identified miRNAs
that were differentially expressed upon Uc.283+A
transfection in the colorectal cancer cell line
HCT116 compared with mock-transfected cells.
(B) The stem-loop sequence of pri-miR-195 and
its partial complementarity with Uc.283+A. The
sequence within the matching region is indicated.
Molecular Cell
A lncRNA Inhibits pri-miRNA Processing
Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re-gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005
epigenetic inactivation by CpG island hypermethylation in a
broad range of human cancer cell lines and primary tumors
(Lujambio et al., 2010). In this work, we characterize a role for
Uc.283+A as a direct interactor and regulator of pri-miRNA-
195 maturation at the level of Drosha processing. We combine
cellular assays with in vitro biochemical analyses to reveal a
case of RNA-directed downregulation of miRNA biogenesis by
a T-UCR.
RESULTS
Searching for miRNA Targets of Uc.283+ADirect interaction between miRNAs and T-UCRs and correlation
between the respective expression levels has been reported
before (Calin et al., 2007; Scaruffi et al., 2009), pointing to regula-
tory mechanisms that could link both types of ncRNA. In order to
identify putativemiRNA targets forUc.283+A, we overexpressed
this T-UCR in an HCT116 colorectal carcinoma cell line (in which
2 Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc.
the transcript is silenced by CpG island
hypermethylation) and performed miRNA
microarray analysis as described before
(Liu et al., 2004) (Figure 1A). This miRNA
microarray chip profiles a total of 1,738
human miRNAs, including 625 precursor
probes and 1,113 mature miRNA probes.
Only 11 of themiRNAspresent in the array
showedR2-fold upregulation or downre-
gulation of expression upon Uc.283+A
transfection. We noticed that only one of
these, miR-195, displayed a perfect
complementarity of >10 bases with the
ultraconserved region of Uc.283+A,
indicating a potential direct regulation of
the two ncRNAs. The complementarity
involved 11 nucleotides situated immedi-
ately upstream of the Drosha cropping
site within themiR-195 primary transcript
(Figure 1B).
Uc.283+A Binds to pri-miR-195 andDownregulates Mature miR-195
LevelsTo experimentally test the interaction
between the two RNAs, the 85 nucleo-
tides corresponding to the pri-miR-195 stem-loop sequence
(as described in the miRBAse Database, release 20) were biotin
labeled by in vitro transcription and incubated in the presence of
wild-type (WT) Uc.283+A ultraconserved region or a mutant
version, in which the 11 complementary nucleotides were
changed to disrupt the interaction (mut11). Electrophoretic
mobility shift assays indicated that the two wild-type RNAs
formed a distinct complex in vitro (Figure 2A, lanes 3–5), whereas
incubation with the mutated version of Uc.283+A completely
abolished the interaction (Figure 2A, lanes 6–8). Conversely,
when compensatory changes were introduced onto the pri-
miR-195 substrate, together with matching mutations on the
other arm of the stem that maintain the structure (pri-miR-
195mut11), the binding was restored (Figure 2A, lanes 13–15).
To investigate this binding in vivo, we next used the neuroblas-
toma cell line SK-N-BE(2), where Uc.283+A has a high basal
expression (Lujambio et al., 2010). The interaction was indeed
detected by using the RNA antisense purification (RAP) method

Molecular Cell
A lncRNA Inhibits pri-miRNA Processing
Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re-gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005
(Engreitz et al., 2013), in which a pool of 124-nt-long antisense
probes designed against Uc.283+A were able to specifically
retrieve the cellular Uc.283+A RNA in association with the pri-
mary transcript ofmiR-195 (Figure 2B, left panel). No associated
DNA from themiR-195 locus could be recovered under the same
conditions, indicating that the interaction does not take place in
the chromatin context (Figure 2B, right panel). Pri-miRNAs are
nuclear species that undergo Drosha cleavage before being
exported to the cytoplasm. The full-length Uc.283+A transcript
is an unspliced, �2.2-kb-long RNA (Lujambio et al., 2010) that
is mainly retrieved from the polyA- fraction of total RNA (Fig-
ure S1A available online). In accordance with these observa-
tions, we detected high amounts of Uc.283+A in the nucleus of
SK-N-BE(2) cells, both by biochemical fractionation (Figure S1B)
and by RNA-FISH (Figure S1C). Overexpression in HCT116 cells
resulted in a similar pattern (Figures S1B and S1C).
Given the existence of this RNA:RNA-direct interaction, and to
confirm the microarray data, we measured miR-195 levels by
means of real-time qRT-PCR inwild-type HCT116 cells or in cells
overexpressing Uc.283+A. As seen in Figure 2C, transient over-
expression of Uc.283+A reduced mature miR-195 levels to less
than half the initial amount, confirming the effect of Uc.283+A on
miRNA biogenesis. This was affected through the complemen-
tary 11 nt described above, because neither cotransfection of
the wild-type Uc.283+Awith a 20-O-methyl-modified oligonucle-
otide that specifically blocked the site on Uc.283+A nor over-
expression of the mutant Uc.283+Amut11 had any impact on
miR-195 levels (Figure 2C). Next, we used the opposite
approach and carried out small hairpin RNA (shRNA)-mediated
knockdown of Uc.283+A in the SK-N-BE(2) cell line. Using two
different shRNAs, both nuclear and cytoplasmicUc.283+A levels
were reduced >60% (Figure S1D), and a concomitant �3- to 4-
fold increase in miR-195 levels was observed (Figure 2D).
Furthermore, to test the implications at the level of mRNA target
regulation, we used a luciferase-based reporter to assay miR-
195 activity (Figure 2E), with three target sites formiR-195 cloned
downstream of the renilla luciferase open reading frame. Co-
transfection with a plasmid that overexpressed miR-195,
induced a reduction in luciferase activity. By contrast, cotrans-
fection with Uc.283+A alleviated the repression of luciferase
activity nearly to the levels of the empty vector, whereas cotrans-
fection with its mutant version did not alter basal repression
(Figure 2E, left graph). Uc.283+Awas able to derepress the lucif-
erase reporter even in the context of enforced expression of pri-
miR-195, when all three plasmids were cotransfected (right
graph, black bars). Similar reporters with miR-9-1 target sites
and pri-miR-9-1 overexpression were unaffected by cotransfec-
tion with Uc.283+A (right graph, empty bars). Together, these
results suggest that Uc.283+A is able to specifically downregu-
late miR-195 levels and to counteract miR-195 repression of a
functional target in vivo, and that this effect requires the pres-
ence of the complementary 11 nt sequence.
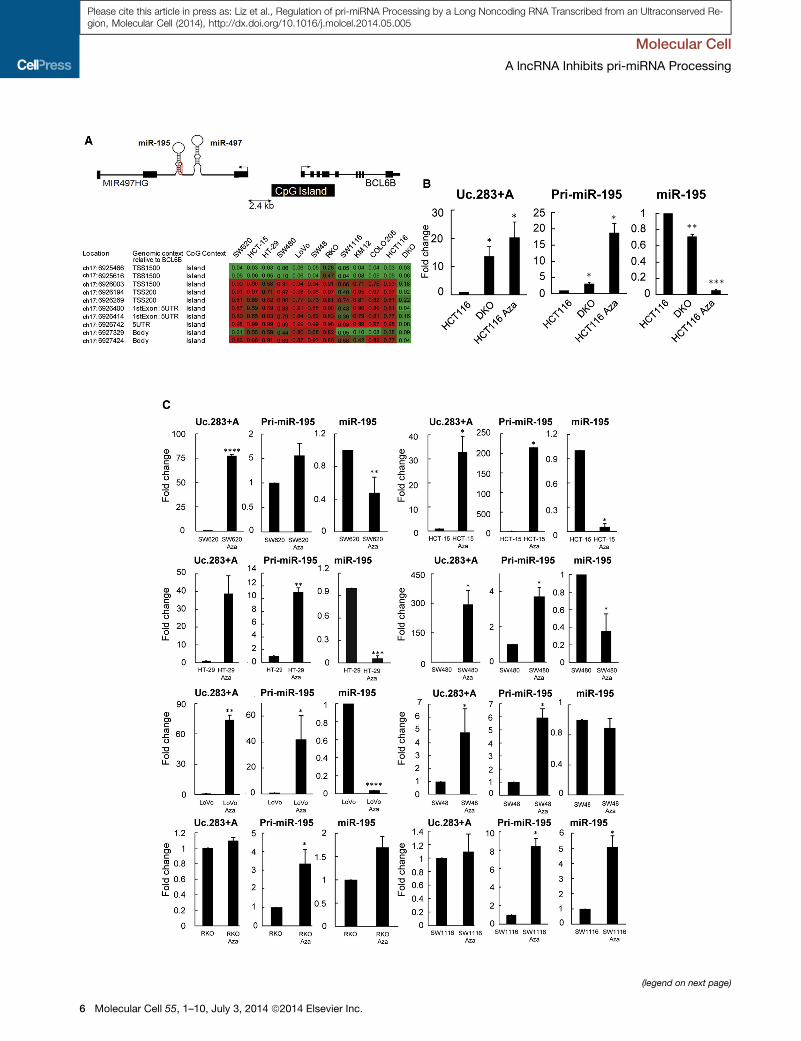
miR-195 Is Posttranscriptionally RegulatedTo investigate the mechanism by which Uc.283+A downregu-
lates miR-195 levels, we examined the transcriptional control
of its primary transcript. miR-195 resides, together with miR-
497, within the intron of a noncoding transcript (known as
MIR497HG, see upper diagram in Figure 3A), with a CpG island
present 2.4 kb upstream of the transcriptional start site. This
CpG island is hypermethylated in most human colon cancer
samples (Menigatti et al., 2013) and cell lines (Figure 3A, lower
panel), whereas the DKO cell line, which is derived from
HCT116 but is hypomorphic for the DNA methyltransferases
DNMT1 and DNMT3b, displays an unmethylated island. To test
the association between miR-195 transcription and the neigh-
boring CpG island methylation, we measured both the primary
transcript (pri-miR-195) and mature miR-195 levels by qRT-
PCR in HCT116 and DKO cell lines, as well as in HCT116 cells
treated with the demethylating agent 5-aza-20-deoxycytidine(Aza). In the hypermethylated HCT116 cell line, both pri-miR-
195 and miR-195 were expressed at low levels. However, as
seen in Figure 3B (middle panel) pri-miR-195 levels increased
in the DKO cell line and Aza-treated HCT116 cells. On the other
hand, maturemiR-195 levels decreased under both hypomethy-
lated conditions (Figure 3B, right panel), suggesting that some
posttranscriptional regulation occurs in this context. Intriguingly,
Uc.283+A transcript was upregulated in DKO and Aza-treated
HCT116 cells, concomitant with hypomethylation of its own
CpG island (Lujambio et al., 2010; Figure 3B, left panel). These
observations hinted at the regulation of miR-195 levels by
Uc.283+A at some point downstream of pri-miR-195 transcrip-
tion. To explore the correlation between Uc.283+A upregulation
and miR-195 posttranscriptional downregulation further, we
analyzed the expression levels of both genes in eight additional
wild-type or Aza-treated colon cancer cells that are normally
hypermethylated at both CpG islands (Figure 3C). Following
treatment with Aza, levels of CpG island methylation were
analyzed at bothUc.283+A andmiR-195 loci by bisulfite conver-
sion followed by PCR cloning and sequencing (Figure S2A). All
Aza-treated cell lines were hypomethylated at both loci, with
the exception of RKO and SW1116, which were hypomethylated
at the miR-195 locus but remained methylated at the Uc.283+A
locus under our Aza treatment conditions. Remarkably, when
both loci became hypomethylated, Uc.283+A and pri-miR-195
transcript levels increased relative to control cells, whereas
mature miR-195 levels dropped by between 2- and >10-fold in
Aza-treated cells. One exception was the SW48 cell line, which
showed the smallest increase in Uc.283+A transcript levels
and a statistically nonsignificant decrease in miR-195 levels.
Neither the RKO nor the SW1116 cell line exhibited any change
in Uc.283+A expression upon Aza treatment, and this was
concomitant with an increase in the level of expression of mature
miR-195 in parallel with an increase in pri-miR-195 transcription
levels. Taken together, these data confirm the association be-
tween Uc.283+A expression and posttranscriptional downregu-
lation of miR-195. This association is specific to miR-195,
because mature miR-497 from the same cluster was not down-
regulated upon Aza treatment (Figure S2B).
Uc.283+A Inhibits pri-miR-195 Processing by Droshathrough Direct RNA:RNA Interaction and Impairment ofDGCR8 BindingGiven the existence of a direct interaction between Uc.283+A
and pri-miR-195 and the posttranscriptional downregulation
observed for miR-195, we hypothesized that Uc.283+A could
Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc. 3
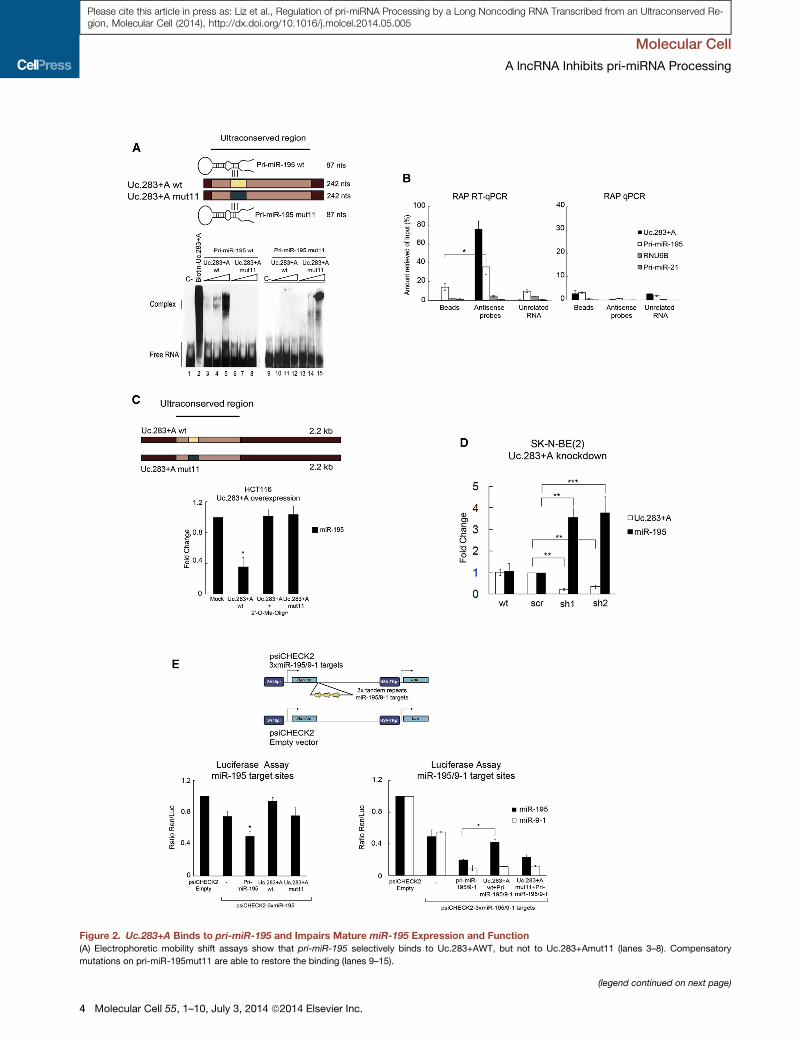
Figure 2. Uc.283+A Binds to pri-miR-195 and Impairs Mature miR-195 Expression and Function
(A) Electrophoretic mobility shift assays show that pri-miR-195 selectively binds to Uc.283+AWT, but not to Uc.283+Amut11 (lanes 3–8). Compensatory
mutations on pri-miR-195mut11 are able to restore the binding (lanes 9–15).
(legend continued on next page)
Molecular Cell
A lncRNA Inhibits pri-miRNA Processing
4 Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc.
Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re-gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005

Molecular Cell
A lncRNA Inhibits pri-miRNA Processing
Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re-gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005
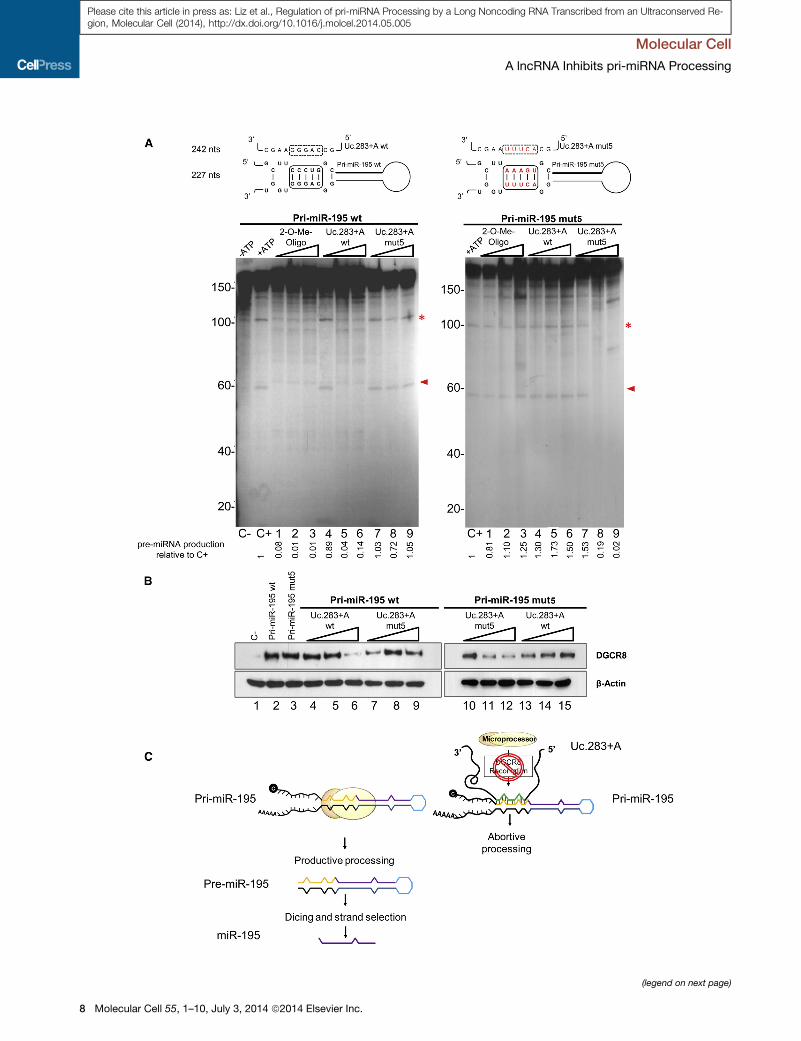
be directly involved in hindering Drosha-mediated processing of
pri-miR-195. To address this, we used the primary miRNA
sequence for miR-195 to set up in vitro processing assays (Fig-
ure 4A). The pri-miR-195 was processed in vitro in the presence
of HEK293T extracts to render a 60 nt product (Figure 4A, left
panel, lane 2). The reaction could be efficiently blocked by incu-
bation with a 20-O-methyl-oligonucleotide that specifically
matched the base of the pri-miR-195 stem, mimicking
Uc.283+A sequence (lanes 1–3). These stable oligos have
been used before to block pri-miRNA processing, particularly
through base-pairing to the loop region (Michlewski et al.,
2008) and were used here as a positive control for preventing
the reaction. Similarly, incubation with the wild-type Uc.283+A
was able to inhibit pri-miR-195 processing (lanes 4–6, see quan-
titation below the gel). This effect requires direct RNA-RNA base-
pairing, because a Uc.283+A version with five nucleotides of the
complementary region mutated had no effect on pri-miR-195
processing (Uc.283+Amut5, lanes 7–9). Conversely, when those
samemutations were introduced onto the pri-miR-195 substrate
(with compensatory mutations on the other arm of the stem that
maintain the structure), the mutant substrate (pri-miR-195mut5)
was efficiently processed independently of the presence of the
20-O-methyl-oligonucleotide or wild-type Uc.283+A (Figure 4A,
right panel, see diagram and lanes 1–6), whereas incubation
with the Uc.283+Amut5 that matched the changes in pri-miR-
195mut5 restored the inhibition (lanes 7–9).
The Drosha cleavage site of pri-miRNAs lies approximately 11
nt away from the stem-ssRNA junction, and this short region is
essential for correct recognition by the double-stranded RNA
(dsRNA)-binding protein DGCR8 and subsequent Drosha
recruitment and catalysis (Han et al., 2006). Because
Uc.283+A binds precisely along this region of pri-miR-195, we
next examined whether Uc.283+A functions through impairment
of DGCR8 anchoring. With that aim, we performed RNA affinity
purification to assess the degree of DGCR8 binding to pri-miR-
195 in the presence or absence ofUc.283+A. In vitro transcribed
wild-type pri-miR-195 or its mutated version were covalently
coupled to activated agarose beads and incubated in the
presence of HEK293T extracts. Analysis of the retained proteins
by western blot showed that DGCR8 was efficiently pulled
down by both RNAs, in accordance with their ability to be
efficiently processed (Figure 4B, lanes 2 and 3). By contrast,
incubation with increasing amounts of Uc.283+A reduced
DGCR8 binding to wild-type pri-miR-195WT, but not to the
(B) Left: qRT-PCR of the RNA captured in crosslinked SK-N-BE(2) cells using str
probes) or against the CELF2 mRNA (unrelated RNA). Right: DNA captured from t
enrichments represent means from three replicate experiments and are relative to
negative controls to assess binding specificity.
(C)miR-195 expression in transiently transfected HCT116 cells overexpressing fu
(D)miR-195 and Uc.283+A expression in SK-N-BE(2) upon Uc.283+A knockdown
(scr) or two shRNAs against Uc.283+A RNA (sh1, sh2). Levels were measured b
parison.
(E) Luciferase reporter assays to test miR-195 functionality upon Uc.283+A ove
reporter vector, pri-miR-195, or with either Uc.283+AWT or Uc.283+Amut11. Re
control vector (empty). Right graph: (black bars) HCT116 cells were cotransfec
together with Uc.283+A (WT or mut11 version). Empty bars represent data from a
overexpressing pri-miR-9-1 were used as controls.
For all panels, error bars represent SD. See also Figure S1.
mutant pri-miR-195mut5 (compare lanes 4–6 with 13–15), indi-
cating that the complementarity between the two RNAs hinders
DGCR8 recognition. This is confirmed by the fact that the mutant
Uc.283+Amut5 had no effect on wild-type pri-miR-195WT (lanes
7–9), while preventing DGCR8 binding to the mutant pri-miR-
195mut5 (lanes 10–12). Finally, the regulation was specific to
miR-195, because the other miRNA on the endogenous pri-
miRNA transcript,miR-497, was not affected byUc.283+A over-
expression in HCT116 cells (Figure S3A) or in in vitro processing
assays (Figure S3B).
Taken together, our results reveal a role for the T-UCR
Uc.283+A in controlling the posttranscriptional processing of
miR-195 at the level of Drosha cleavage. This is achieved
through a direct RNA-RNA interaction that prevents the efficient
recognition by DGCR8 and highlights the roles of noncoding
RNAs as important regulators of the transcriptome.
DISCUSSION
Given the structural similarity of mRNA and lncRNAs, the exis-
tence of natural miRNA binding sites along lncRNA sequences
was not entirely unexpected. The HITS-CLIP technique has
globally mapped the interactions between mature miRNAs and
lncRNAs (Chi et al., 2009). These interactions may represent a
mechanism of mutual regulation that could work in both direc-
tions. On the one hand, a number of lncRNAs have been identi-
fied as competing endogenous RNAs (ceRNAs) (Franco-Zorrilla
et al., 2007). In general, ceRNAs act as miRNA sponges (Ebert
et al., 2007) and reduce the activity of the target miRNAs without
essentially altering their biogenesis (Kallen et al., 2013; Cesana
et al., 2011). The recently characterized circular RNAs (circRNAs)
can also function asmiRNA sponges, as is the case for the brain-
specific ciRS-7 as an inhibitor ofmiR-7 (Hansen et al., 2013). On
the other hand, the miRNA-lncRNA interaction might not only
represent miRNA sequestration but also downregulation of the
target lncRNA, as has been shown in a zebrafish model (Jalali
et al., 2013).
The work presented here reveals another layer of regulation
between miRNAs and lncRNAs: to date, regulation of miRNA
biogenesis at the level of pri-miRNA to pre-miRNA Drosha cleav-
age had only dealt with protein factors that specifically bind to
distinct elements along the pri-miRNA (directly or through inter-
action with the Drosha-DGCR8 microprocessor complex) to
control miRNAmaturation (Guil andCaceres, 2007; Viswanathan
eptavidin beads alone (beads), with antisense probes to Uc.283+A (antisense
he same samples was amplified by quantitative real-time PCR. In both panels,
the input amount used per pull-down. Both RNU6B and pri-miR-21 are used as
ll-lengthUc.283+A. Levels were measured by qRT-PCR 72 hr posttransfection.
. Cells were transduced with lentiviral plasmids overexpressing control shRNA
y qRT-PCR after eight cell passages. Wild-type cells are also shown for com-
rexpression. Left graph: HCT116 cells were cotransfected with the indicated
lative firefly luciferase/renilla activity was determined and compared with the
ted with the indicated reporter plasmid and with either pri-miR-195 alone or
parallel experiment in which a reporter withmiR-9-1 target sites and a plasmid
Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc. 5
(legend on next page)
Molecular Cell
A lncRNA Inhibits pri-miRNA Processing
6 Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc.
Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re-gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005

Molecular Cell
A lncRNA Inhibits pri-miRNA Processing
Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re-gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005
et al., 2008; Trabucchi et al., 2009; Michlewski and Caceres,
2010, 2010; Kawahara and Mieda-Sato, 2012; Choudhury
et al., 2013). For instance, the terminal loop region has been
the focus of intense research in spite of being originally consid-
ered dispensable for microprocessor action, and many miRNAs
have evolutionarily conserved loops that may contain regulatory
regions (Michlewski et al., 2008). Instead, we have found an
example of negative regulation of pri-miRNA processing that de-
pends on direct RNA-RNA interaction between the lower stem
region of pri-miR-195 and a sequence present in the ultracon-
served region of the lncRNA Uc.283+A. As seen with the mutant
constructs, the regulation is not sequence dependent but re-
quires complementarity between the two RNA molecules. The
�11 nt lower stem region along pri-miRNA sequences between
Drosha cleavage site and the junctions with the surrounding sin-
gle-stranded RNA sequences (basal segments) is essential for
DGCR8 recognition of the characteristic structural features of
pri-miRNAs. The double strandedness of the lower stem and
the distance from the basal segments are both determinants of
the Drosha cleavage site (Han et al., 2006). We propose a model
in which disruption of the lower stem structure through
Uc.283+A strand invasion prevents optimal recognition of the
region by the dsRNA-binding protein DGCR8 and consequently
blocks efficient processing by Drosha (see diagram in Figure 4C).
We cannot rule out the possibility that, additionally, some protein
factors (such as helicases) may favor the stabilization of the pri-
miRNA-Uc.283+A interaction, and current efforts are directed
toward settling this point. Interestingly, the DEAD box RNA heli-
case domain-containing proteins p68/p72 selectively recruit pri-
miRNAs for efficient cleavage by Drosha (Fukuda et al., 2007).
Future work will assess how common this direct RNA:RNA regu-
lation is between different classes of ncRNAs and primary
miRNAmolecules. An in silico analysis of the number of potential
complementary regions between known T-UCRs and lncRNAs
and the stem-loop sequence of human pri-miRNAs is shown in
Table S1.
miR-195 has a variety of functions in diverse biological sys-
tems. Importantly, miR-195 regulates the proliferation of cancer
cell lines (Liu et al., 2010) and contributes to multidrug resistance
in head and neck squamous cell carcinoma cell lines (Dai et al.,
2011). For instance,miR-195 targets cyclin D1, CDK6, and E2F3
to suppress cell proliferation in human hepatocellular carcinoma
cells (Xu et al., 2009), whereas its overexpression promotes pro-
liferation in adult neural stem/progenitor cells, with no effect on
cyclin D1 expression (Liu et al., 2013), suggesting that the targets
ofmiR-195 and the effects ofmiR-195 on cell proliferationmay in
fact be context dependent. Our luciferase reporter assays
showed the impact of Uc.283+A expression on miR-195 func-
Figure 3. Promoter Demethylation atmiR-195 and Uc.283+A Loci Reve
transcriptional Downregulation
(A) Upper diagram: schematic representation of the genomic context of theMIR49
kb upstream of MIR497HG TSS. Lower diagram: methylation status of this CpG
lines.
(B) Expression analyses of pri-miR-195 and miR-195 by qRT-PCR.
(C) Expression analyses in a panel of colon cancer cell lines for Uc.283+A, pri-m
5-aza-20-deoxycytidine.Error bars represent SD. See also Figure S2.
tionality. It is of note that T-UCRs have been shown to be de-
regulated in neuroblastomas and have even been ascribed a
prognostic value (Mestdagh et al., 2010).
In summary, previous work had reported a range of mecha-
nisms that regulate miRNA biogenesis, involving protein-protein
and RNA-protein interactions, but our study characterizes a
regulatory control of miRNA processing based on RNA-RNA
interactions, highlighting the increasing complexity of ncRNA-
mediated regulatory networks.
EXPERIMENTAL PROCEDURES
Cell Lines, Culture Conditions, 5-aza-20-Deoxycytidine Treatment,
and Transfections
All cell lines were obtained from the American Type Culture and grown as
recommended. Treatments were as described in the Supplemental Experi-
mental Procedures.
Plasmid Constructions
See the Supplemental Experimental Procedures for details about the cloning
strategy and preparation of mutants.
RAP
RAP protocol was performed as described before (Engreitz et al., 2013).
Briefly, the Uc.283+A RNA was tiled with eight 124 nt antisense RNA probes
that had been biotinylated by in vitro transcription. The full-length Uc.283+A
was covered by these probes, with the exception of the region interacting
with pri-miRNA and the fragment corresponding to the amplicon in the RT-
PCR (to avoid interference in the qRT-PCR signal). SK-N-BE(2) cells were
crosslinked first with 2mM disuccinimidyl glutarate for 45min at room temper-
ature and then with 3% formaldehyde for 10min at 37�C. For each purification,100 ng of biotinylated probes were added to the precleared lysates, and the
mix was incubated at 45�C. The probes were then captured by streptavidin
beads, and the elutions for the associated RNA and DNA were performed.
As control, the same experiment was carried out in parallel with probes tiling
the unrelated CELF2 mRNA or with streptavidin beads without any probe.
Recovered RNA and DNA samples were analyzed by qRT-PCR and quantita-
tive real-time PCR, respectively, together with 1/10 dilution of the input
material.
RNA Isolation, Microarray Experiment, Real-Time qRT-PCR, and
Luciferase Reporter Assays
See the Supplemental Experimental Procedures for details about RNA isola-
tion, microarray experiment, real-time qRT-PCR, and luciferase reporter
assays.
In Vitro pri-miRNA Processing Assays
Pri-miR-195RNA substrates for in vitro processing assays were prepared from
DNA templates by standard in vitro transcription with T7 RNA Polymerase
(Roche) in the presence of [a-32P] ATP (PerkinElmer). RNA substrates for
Uc.283+A (WT andmut5) were obtained by in vitro transcription from linearized
DNA templates. Processing reactions were carried out with total HEK293T
extracts as previously described (Guil and Caceres, 2007).
als a Correlation between Uc.283+A Upregulation andmiR-195 Post-
7HG cluster. TheCpG island that governs pri-miR-195 expression is located 2.4
island according to Illumina 450K BeadChip array in several colon cancer cell
iR-195, and miR-195, before and after treatment with the demethylating agent
Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc. 7
(legend on next page)
Molecular Cell
A lncRNA Inhibits pri-miRNA Processing
8 Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc.
Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re-gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005

Molecular Cell
A lncRNA Inhibits pri-miRNA Processing
Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re-gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005
Electrophoretic Mobility Assays and RNA Pull-Down
For the electrophoreticmobility shift assay, the RNA substrate for thewild-type
or mutant miR-195 stem-loop sequence (87 nt) was obtained and labeled by
random biotinylation during the in vitro transcription using 0.25 mM biotin-
16-UTP (Roche) from linearized DNA templates. For RNA pull-downs, in vitro
transcribed RNAs were covalently linked to adipic acid dihydrazide agarose
beads and incubated with increasing amounts of Uc.283+A RNA. Affinity pu-
rification of DGCR8 protein was performed with HEK293T nuclear extracts.
See the Supplemental Experimental Procedures for additional details on
protocols and methods used.
ACCESSION NUMBERS
The Gene Expression Omnibus accession number for the miRNA microarray
data reported in this paper is GSE56841.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures,
three figures, and two tables and can be found with this article online at
http://dx.doi.org/10.1016/j.molcel.2014.05.005.
ACKNOWLEDGMENTS
This work was supported by a European Research Council Advanced Grant
(#268626 – EPINORC project) (to M.E), the Ministerio de Ciencia e Innovacion
(MICINN) (SAF2011-22803 to M.E.; SAF2011-22895 to S.G.), Fundacion
Ramon Areces, the Cellex Foundation, the Health and Science Departments
of the Catalan Government (Generalitat de Catalunya), and Fondo de Investi-
gaciones Sanitarias (PI08-1345) (to M.E.). G.M. is a recipient of a Medical
Research Council Career Development Award (G10000564). S.G. is funded
by the Ramon y Cajal Research Program (MICINN). M.E. is an ICREA Research
Professor.
Received: December 10, 2013
Revised: March 17, 2014
Accepted: May 1, 2014
Published: June 5, 2014
REFERENCES
Bartel, D.P. (2009). MicroRNAs: target recognition and regulatory functions.
Cell 136, 215–233.
Bejerano, G., Pheasant, M., Makunin, I., Stephen, S., Kent, W.J., Mattick, J.S.,
and Haussler, D. (2004). Ultraconserved elements in the human genome.
Science 304, 1321–1325.
Bernard, D., Prasanth, K.V., Tripathi, V., Colasse, S., Nakamura, T., Xuan, Z.,
Zhang, M.Q., Sedel, F., Jourdren, L., Coulpier, F., et al. (2010). A long
nuclear-retained non-coding RNA regulates synaptogenesis by modulating
gene expression. EMBO J. 29, 3082–3093.
Calin, G.A., Liu, C.G., Ferracin, M., Hyslop, T., Spizzo, R., Sevignani, C.,
Fabbri, M., Cimmino, A., Lee, E.J., Wojcik, S.E., et al. (2007). Ultraconserved
Figure 4. Uc.283+A Binding to pri-miR-195 Prevents Drosha Cropping
(A) The diagrams above show the 5 positions (boxed) within the 11 nt complement
introduced are indicated in red. For pri-miR-195mut5, compensatorymutationswe
in vitro microprocessing assays show that pri-miR-195 processing is affected by
pri-miR-195mut5 processing is affected only when Uc.283+Amut5 is added. In b
was used as a control. The red arrow indicates the pre-miRNA hairpin product, an
of pre-miRNA produced was determined by densitometry and calculated relative
(B) RNA pull-down. RNA transcripts (pri-miR-195wild-type ormutant forms) were
(wild-type or mutant), in the presence of nuclear extract from HEK293T cells. Th
(C) Schematic depicting our proposed model for Uc.283+A disruption of DGCR8
See also Figure S3.
regions encoding ncRNAs are altered in human leukemias and carcinomas.
Cancer Cell 12, 215–229.
Cesana,M., Cacchiarelli, D., Legnini, I., Santini, T., Sthandier, O., Chinappi, M.,
Tramontano, A., and Bozzoni, I. (2011). A long noncoding RNA controls muscle
differentiation by functioning as a competing endogenous RNA. Cell 147,
358–369.
Chi, S.W., Zang, J.B., Mele, A., and Darnell, R.B. (2009). Argonaute HITS-CLIP
decodes microRNA-mRNA interaction maps. Nature 460, 479–486.
Choudhury, N.R., de Lima Alves, F., de Andres-Aguayo, L., Graf, T., Caceres,
J.F., Rappsilber, J., andMichlewski, G. (2013). Tissue-specific control of brain-
enriched miR-7 biogenesis. Genes Dev. 27, 24–38.
Costa, F.F. (2005). Non-coding RNAs: new players in eukaryotic biology. Gene
357, 83–94.
Dai, Y., Xie, C.H., Neis, J.P., Fan, C.Y., Vural, E., and Spring, P.M. (2011).
MicroRNA expression profiles of head and neck squamous cell carcinoma
with docetaxel-induced multidrug resistance. Head Neck 33, 786–791.
Djebali, S., Davis, C.A., Merkel, A., Dobin, A., Lassmann, T., Mortazavi, A.,
Tanzer, A., Lagarde, J., Lin, W., Schlesinger, F., et al. (2012). Landscape of
transcription in human cells. Nature 489, 101–108.
Ebert, M.S., Neilson, J.R., and Sharp, P.A. (2007). MicroRNA sponges:
competitive inhibitors of small RNAs in mammalian cells. Nat. Methods 4,
721–726.
Engreitz, J.M., Pandya-Jones, A., McDonel, P., Shishkin, A., Sirokman, K.,
Surka, C., Kadri, S., Xing, J., Goren, A., Lander, E.S., et al. (2013). The Xist
lncRNA exploits three-dimensional genome architecture to spread across
the X chromosome. Science 341, 1237973.
Franco-Zorrilla, J.M., Valli, A., Todesco, M., Mateos, I., Puga, M.I., Rubio-
Somoza, I., Leyva, A., Weigel, D., Garcıa, J.A., and Paz-Ares, J. (2007).
Target mimicry provides a newmechanism for regulation of microRNA activity.
Nat. Genet. 39, 1033–1037.
Friedman, R.C., Farh, K.K., Burge, C.B., andBartel, D.P. (2009). Mostmamma-
lian mRNAs are conserved targets of microRNAs. Genome Res. 19, 92–105.
Fukuda, T., Yamagata, K., Fujiyama, S., Matsumoto, T., Koshida, I.,
Yoshimura, K., Mihara, M., Naitou, M., Endoh, H., Nakamura, T., et al.
(2007). DEAD-box RNA helicase subunits of the Drosha complex are required
for processing of rRNA and a subset of microRNAs. Nat. Cell Biol. 9, 604–611.
Garzon, R., Calin, G.A., and Croce, C.M. (2009). MicroRNAs in cancer. Annu.
Rev. Med. 60, 167–179.
Guil, S., and Caceres, J.F. (2007). The multifunctional RNA-binding protein
hnRNP A1 is required for processing of miR-18a. Nat. Struct. Mol. Biol. 14,
591–596.
Han, J., Lee, Y., Yeom, K.H., Nam, J.W., Heo, I., Rhee, J.K., Sohn, S.Y., Cho,
Y., Zhang, B.T., and Kim, V.N. (2006). Molecular basis for the recognition of
primary microRNAs by the Drosha-DGCR8 complex. Cell 125, 887–901.
Hansen, T.B., Jensen, T.I., Clausen, B.H., Bramsen, J.B., Finsen, B.,
Damgaard, C.K., and Kjems, J. (2013). Natural RNA circles function as efficient
microRNA sponges. Nature 495, 384–388.
Jalali, S., Bhartiya, D., Lalwani, M.K., Sivasubbu, S., and Scaria, V. (2013).
Systematic transcriptome wide analysis of lncRNA-miRNA interactions.
PLoS ONE 8, e53823.
by Impairment of DGCR8 Binding
ary region that aremutated in pri-miR-195mut5 or Uc.283+Amut5. The changes
re also introduced in the 30 armof the stem tomaintain the structure. Left panel:
the addition of Uc.283+AWT, but not of Uc.283+Amut5. Right panel: similarly,
oth panels, the same 20-O-Me-oligo that mimics the Uc.283+AWT binding site
d the red asterisk corresponds to intermediate cleavage products. The amount
to the C+ lane.
coupled to agarose beads and incubated with increasing amounts ofUc.283+A
e relative amount of DGCR8 in each case was analyzed by western blot.
recognition.
Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc. 9

Molecular Cell
A lncRNA Inhibits pri-miRNA Processing
Please cite this article in press as: Liz et al., Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed from an Ultraconserved Re-gion, Molecular Cell (2014), http://dx.doi.org/10.1016/j.molcel.2014.05.005
Kallen, A.N., Zhou, X.B., Xu, J., Qiao, C., Ma, J., Yan, L., Lu, L., Liu, C., Yi, J.S.,
Zhang, H., et al. (2013). The imprinted H19 lncRNA antagonizes let-7
microRNAs. Mol. Cell 52, 101–112.
Kapranov, P., Cheng, J., Dike, S., Nix, D.A., Duttagupta, R., Willingham, A.T.,
Stadler, P.F., Hertel, J., Hackermuller, J., Hofacker, I.L., et al. (2007). RNA
maps reveal new RNA classes and a possible function for pervasive transcrip-
tion. Science 316, 1484–1488.
Katzman, S., Kern, A.D., Bejerano, G., Fewell, G., Fulton, L., Wilson, R.K.,
Salama, S.R., and Haussler, D. (2007). Human genome ultraconserved
elements are ultraselected. Science 317, 915.
Kawahara, Y., and Mieda-Sato, A. (2012). TDP-43 promotes microRNA
biogenesis as a component of the Drosha and Dicer complexes. Proc. Natl.
Acad. Sci. USA 109, 3347–3352.
Kim, V.N., Han, J., and Siomi, M.C. (2009). Biogenesis of small RNAs in
animals. Nat. Rev. Mol. Cell Biol. 10, 126–139.
Kino, T., Hurt, D.E., Ichijo, T., Nader, N., and Chrousos, G.P. (2010). Noncoding
RNA gas5 is a growth arrest- and starvation-associated repressor of the gluco-
corticoid receptor. Sci. Signal. 3, ra8.
Krol, J., Loedige, I., and Filipowicz, W. (2010). The widespread regulation of
microRNA biogenesis, function and decay. Nat. Rev. Genet. 11, 597–610.
Liu, C.G., Calin, G.A., Meloon, B., Gamliel, N., Sevignani, C., Ferracin, M.,
Dumitru, C.D., Shimizu, M., Zupo, S., Dono, M., et al. (2004). An oligonucleo-
tide microchip for genome-wide microRNA profiling in human and mouse
tissues. Proc. Natl. Acad. Sci. USA 101, 9740–9744.
Liu, L., Chen, L., Xu, Y., Li, R., and Du, X. (2010). microRNA-195 promotes
apoptosis and suppresses tumorigenicity of human colorectal cancer cells.
Biochem. Biophys. Res. Commun. 400, 236–240.
Liu, C., Teng, Z.Q., McQuate, A.L., Jobe, E.M., Christ, C.C., von Hoyningen-
Huene, S.J., Reyes, M.D., Polich, E.D., Xing, Y., Li, Y., et al. (2013). An epige-
netic feedback regulatory loop involving microRNA-195 and MBD1 governs
neural stem cell differentiation. PloS One 8, e51436.
Lujambio, A., Ropero, S., Ballestar, E., Fraga, M.F., Cerrato, C., Setien, F.,
Casado, S., Suarez-Gauthier, A., Sanchez-Cespedes, M., Git, A., et al.
(2007). Genetic unmasking of an epigenetically silenced microRNA in human
cancer cells. Cancer Res. 67, 1424–1429.
Lujambio, A., Portela, A., Liz, J., Melo, S.A., Rossi, S., Spizzo, R., Croce, C.M.,
Calin, G.A., and Esteller, M. (2010). CpG island hypermethylation-associated
silencing of non-coding RNAs transcribed from ultraconserved regions in
human cancer. Oncogene 29, 6390–6401.
10 Molecular Cell 55, 1–10, July 3, 2014 ª2014 Elsevier Inc.
Menigatti, M., Staiano, T., Manser, C.N., Bauerfeind, P., Komljenovic, A.,
Robinson, M., Jiricny, J., Buffoli, F., and Marra, G. (2013). Epigenetic silencing
of monoallelically methylated miRNA loci in precancerous colorectal lesions.
Oncogenesis 2, e56.
Mestdagh, P., Fredlund, E., Pattyn, F., Rihani, A., Van Maerken, T., Vermeulen,
J., Kumps, C., Menten, B., De Preter, K., Schramm, A., et al. (2010). An integra-
tive genomics screen uncovers ncRNA T-UCR functions in neuroblastoma
tumours. Oncogene 29, 3583–3592.
Michlewski, G., and Caceres, J.F. (2010). Antagonistic role of hnRNP A1 and
KSRP in the regulation of let-7a biogenesis. Nat. Struct. Mol. Biol. 17, 1011–
1018.
Michlewski, G., Guil, S., Semple, C.A., and Caceres, J.F. (2008).
Posttranscriptional regulation of miRNAs harboring conserved terminal loops.
Mol. Cell 32, 383–393.
Rinn, J.L., Kertesz, M., Wang, J.K., Squazzo, S.L., Xu, X., Brugmann, S.A.,
Goodnough, L.H., Helms, J.A., Farnham, P.J., Segal, E., and Chang, H.Y.
(2007). Functional demarcation of active and silent chromatin domains in
human HOX loci by noncoding RNAs. Cell 129, 1311–1323.
Saito, Y., Liang, G., Egger, G., Friedman, J.M., Chuang, J.C., Coetzee, G.A.,
and Jones, P.A. (2006). Specific activation of microRNA-127 with downregula-
tion of the proto-oncogene BCL6 by chromatin-modifying drugs in human
cancer cells. Cancer Cell 9, 435–443.
Scaruffi, P., Stigliani, S., Moretti, S., Coco, S., De Vecchi, C., Valdora, F.,
Garaventa, A., Bonassi, S., and Tonini, G.P. (2009). Transcribed-ultra
conserved region expression is associated with outcome in high-risk neuro-
blastoma. BMC Cancer 9, 441.
Thomas, M., Lieberman, J., and Lal, A. (2010). Desperately seeking microRNA
targets. Nat. Struct. Mol. Biol. 17, 1169–1174.
Trabucchi, M., Briata, P., Garcia-Mayoral, M., Haase, A.D., Filipowicz, W.,
Ramos, A., Gherzi, R., and Rosenfeld, M.G. (2009). The RNA-binding protein
KSRP promotes the biogenesis of a subset of microRNAs. Nature 459,
1010–1014.
Viswanathan, S.R., Daley, G.Q., and Gregory, R.I. (2008). Selective blockade
of microRNA processing by Lin28. Science 320, 97–100.
Winter, J., Jung, S., Keller, S., Gregory, R.I., and Diederichs, S. (2009). Many
roads to maturity: microRNA biogenesis pathways and their regulation. Nat.
Cell Biol. 11, 228–234.
Xu, T., Zhu, Y., Xiong, Y., Ge, Y.Y., Yun, J.P., and Zhuang, S.M. (2009).
MicroRNA-195 suppresses tumorigenicity and regulates G1/S transition of
human hepatocellular carcinoma cells. Hepatology 50, 113–121.

Molecular Cell, Volume 55
Supplemental Information
Regulation of pri-miRNA Processing by a Long Noncoding RNA Transcribed
from an Ultraconserved Region
Julia Liz, Anna Portela, Marta Soler, Antonio Gómez, Hui Ling, Gracjan Michlewski,
George A. Calin, Sònia Guil, and Manel Esteller
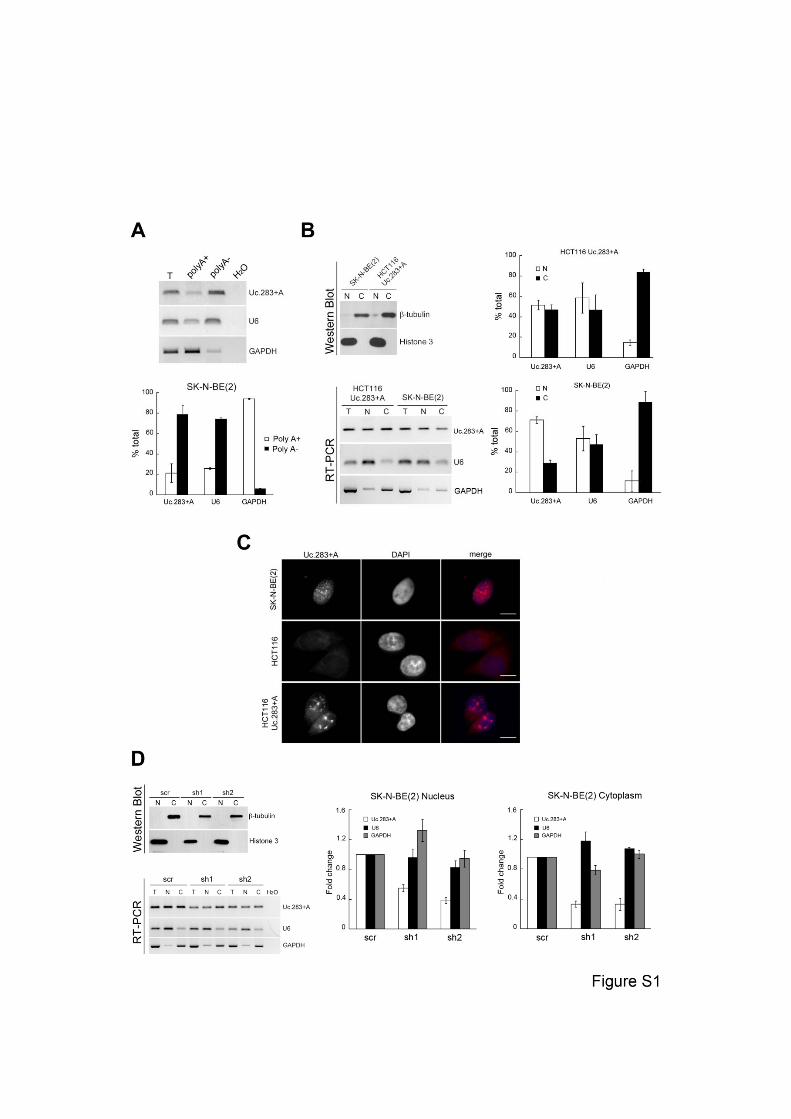

Figure S1. Uc.283+A is a polyA-, nuclear RNA. (A) polyA+/polyA- partition of SK-N-
BE(2) transcripts. Each fraction was amplified by semi-quantitative RT-PCR (upper
panel) or RT-qPCR (lower panel). (T, total RNA). (B) Nuclear/cytoplasmic fractionation
of wild-type SK-N-BE(2) cells and HCT116 overexpressing full-length Uc.283+A RNA.
Upper gel, protein samples were subject to Western Blot to assess fraction purity. RNA
from the same samples was analyzed by semi-quantitative PCR (lower gel) or
quantitative RT-PCR (graphs on the right). (C) RNA-FISH targeting Uc.283+A RNA in
wild-type SK-N-BE(2) cells (which display high expression) (upper panel), wild-type
HCT116 cells (low expression) (middle panel) or HCT116 cells overexpressing
Uc.283+A (lower panel). Scale bars, 10μm. (D) Nuclear/cytoplasmic fractionation of
control (scr) or Uc.283+A-depleted (sh1, sh2) SK-N-BE(2) cells. Western Blot to assess
fraction purity and semiquantitative RT-PCR from each fraction are shown on the left.
Quantitative RT-PCR analyses shown on the right indicate depletion of both the
nuclear and the cytoplasmic Uc.283+A transcripts relative to the levels for each
compartment in the control cells. Error bars represent standard deviation
Related to Figure 2.


Figure S2. CpG island methylation analysis in colon cancer cell lines following
Aza treatment. (A) CpG dinucleotides on Uc.283+A (left) and miR-195 (right) CpG
islands are represented as short vertical lines. Eight single clones are represented for
each sample. Black squares represent methylated cytosines and white ones,
unmethylated.
(B) Expression analysis in a panel of colon cancer cell lines for pri-miR-497 and miR-
497, before and after treatment with the demethylating agent 5-aza-2′-deoxycytidine.
Error bars represent standard deviation.
Related to Figure 3.
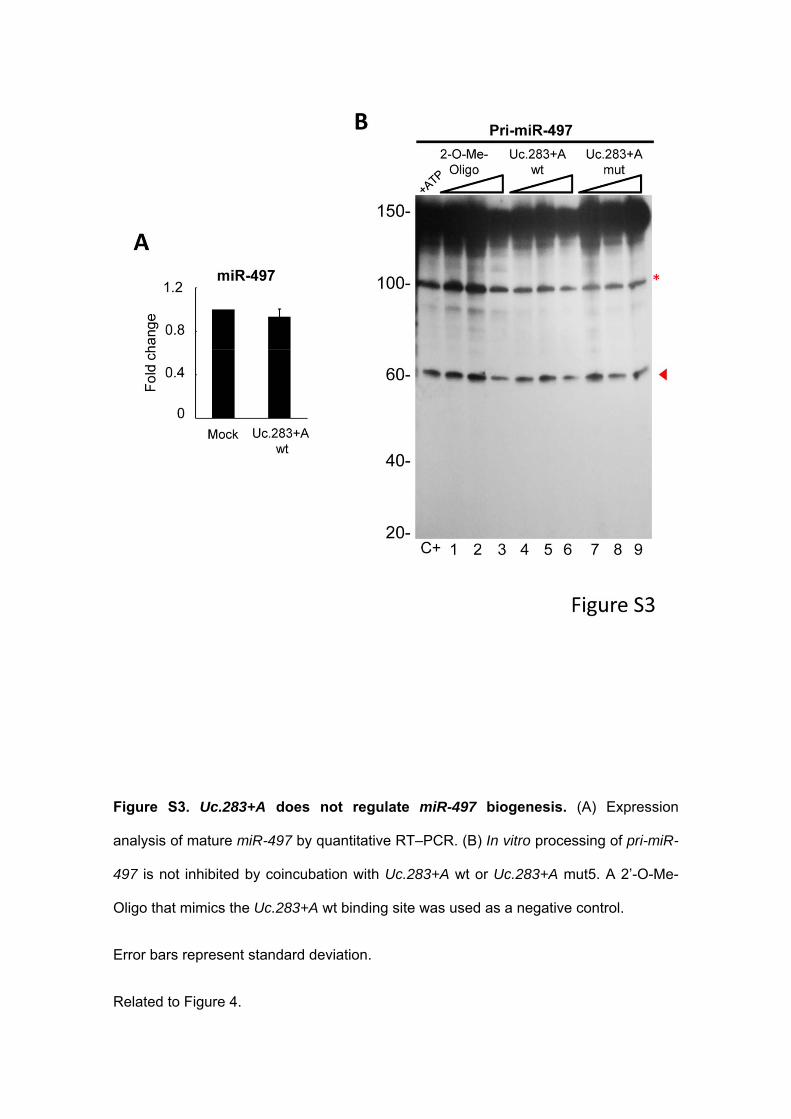
Figure S3. Uc.283+A does not regulate miR-497 biogenesis. (A) Expression
analysis of mature miR-497 by quantitative RT–PCR. (B) In vitro processing of pri-miR-
497 is not inhibited by coincubation with Uc.283+A wt or Uc.283+A mut5. A 2’-O-Me-
Oligo that mimics the Uc.283+A wt binding site was used as a negative control.
Error bars represent standard deviation.
Related to Figure 4.
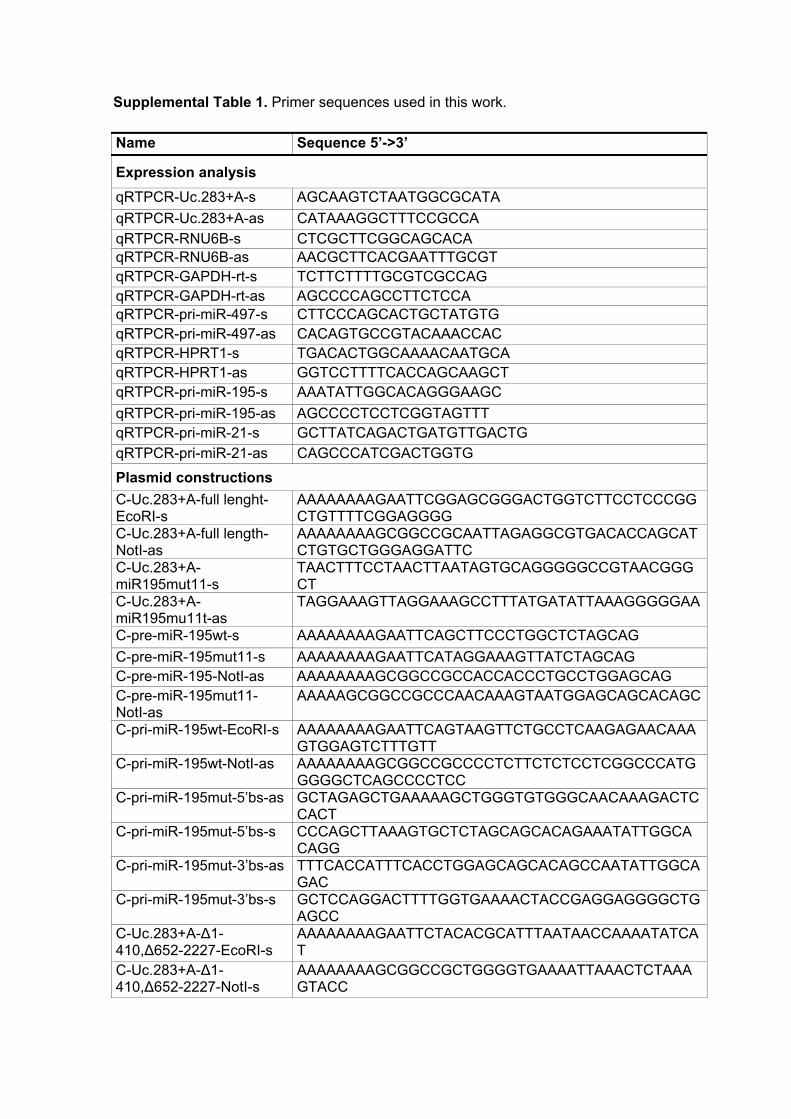
Supplemental Table 1. Primer sequences used in this work.
Name Sequence 5’->3’
Expression analysis
qRTPCR-Uc.283+A-s AGCAAGTCTAATGGCGCATA
qRTPCR-Uc.283+A-as CATAAAGGCTTTCCGCCA qRTPCR-RNU6B-s CTCGCTTCGGCAGCACA qRTPCR-RNU6B-as AACGCTTCACGAATTTGCGT qRTPCR-GAPDH-rt-s TCTTCTTTTGCGTCGCCAG qRTPCR-GAPDH-rt-as AGCCCCAGCCTTCTCCA qRTPCR-pri-miR-497-s CTTCCCAGCACTGCTATGTG qRTPCR-pri-miR-497-as CACAGTGCCGTACAAACCAC qRTPCR-HPRT1-s TGACACTGGCAAAACAATGCA qRTPCR-HPRT1-as GGTCCTTTTCACCAGCAAGCT qRTPCR-pri-miR-195-s AAATATTGGCACAGGGAAGC
qRTPCR-pri-miR-195-as AGCCCCTCCTCGGTAGTTT qRTPCR-pri-miR-21-s GCTTATCAGACTGATGTTGACTG qRTPCR-pri-miR-21-as CAGCCCATCGACTGGTG
Plasmid constructions
C-Uc.283+A-full lenght-EcoRI-s
AAAAAAAAGAATTCGGAGCGGGACTGGTCTTCCTCCCGGCTGTTTTCGGAGGGG
C-Uc.283+A-full length-NotI-as
AAAAAAAAGCGGCCGCAATTAGAGGCGTGACACCAGCATCTGTGCTGGGAGGATTC
C-Uc.283+A-miR195mut11-s
TAACTTTCCTAACTTAATAGTGCAGGGGGCCGTAACGGGCT
C-Uc.283+A-miR195mu11t-as
TAGGAAAGTTAGGAAAGCCTTTATGATATTAAAGGGGGAA
C-pre-miR-195wt-s AAAAAAAAGAATTCAGCTTCCCTGGCTCTAGCAG
C-pre-miR-195mut11-s AAAAAAAAGAATTCATAGGAAAGTTATCTAGCAG C-pre-miR-195-NotI-as AAAAAAAAGCGGCCGCCACCACCCTGCCTGGAGCAG C-pre-miR-195mut11-NotI-as
AAAAAGCGGCCGCCCAACAAAGTAATGGAGCAGCACAGC
C-pri-miR-195wt-EcoRI-s AAAAAAAAGAATTCAGTAAGTTCTGCCTCAAGAGAACAAAGTGGAGTCTTTGTT
C-pri-miR-195wt-NotI-as AAAAAAAAGCGGCCGCCCCTCTTCTCTCCTCGGCCCATGGGGGCTCAGCCCCTCC
C-pri-miR-195mut-5’bs-as GCTAGAGCTGAAAAAGCTGGGTGTGGGCAACAAAGACTCCACT
C-pri-miR-195mut-5’bs-s CCCAGCTTAAAGTGCTCTAGCAGCACAGAAATATTGGCACAGG
C-pri-miR-195mut-3’bs-as TTTCACCATTTCACCTGGAGCAGCACAGCCAATATTGGCAGAC
C-pri-miR-195mut-3’bs-s GCTCCAGGACTTTTGGTGAAAACTACCGAGGAGGGGCTGAGCC
C-Uc.283+A-∆1-410,∆652-2227-EcoRI-s
AAAAAAAAGAATTCTACACGCATTTAATAACCAAAATATCAT
C-Uc.283+A-∆1-410,∆652-2227-NotI-s
AAAAAAAAGCGGCCGCTGGGGTGAAAATTAAACTCTAAAGTACC
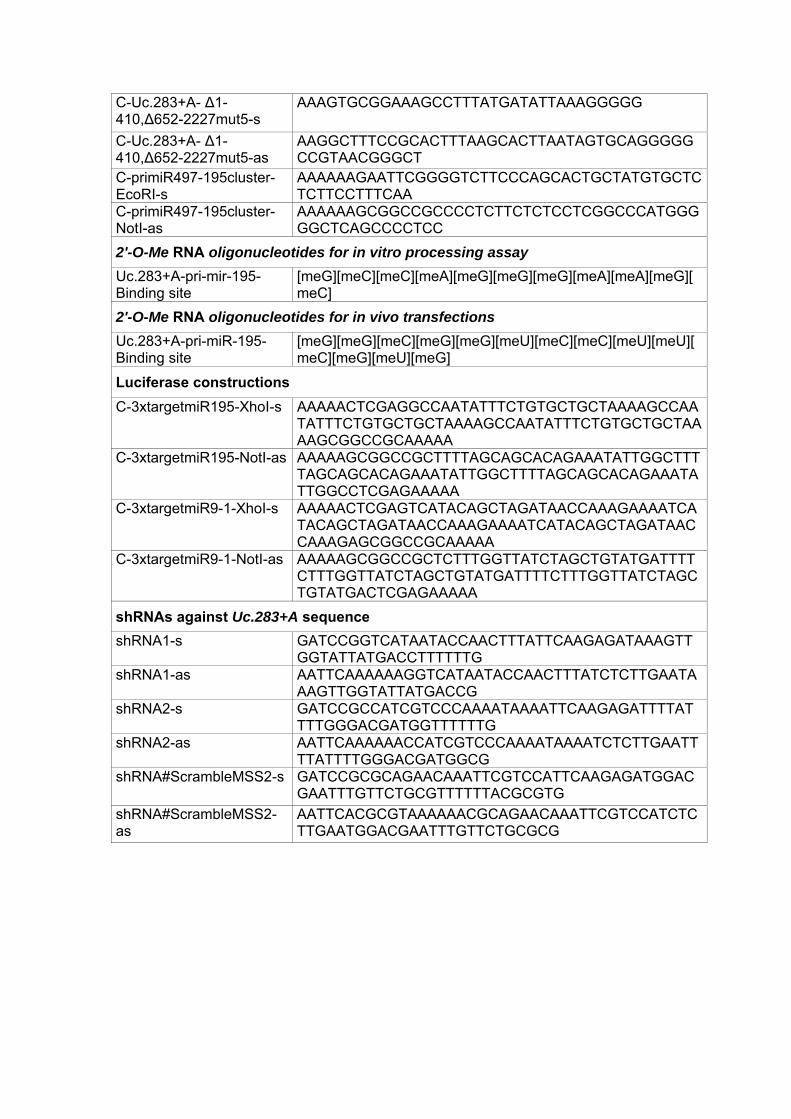
C-Uc.283+A- ∆1-410,∆652-2227mut5-s
AAAGTGCGGAAAGCCTTTATGATATTAAAGGGGG
C-Uc.283+A- ∆1-410,∆652-2227mut5-as
AAGGCTTTCCGCACTTTAAGCACTTAATAGTGCAGGGGGCCGTAACGGGCT
C-primiR497-195cluster-EcoRI-s
AAAAAAGAATTCGGGGTCTTCCCAGCACTGCTATGTGCTCTCTTCCTTTCAA
C-primiR497-195cluster-NotI-as
AAAAAAGCGGCCGCCCCTCTTCTCTCCTCGGCCCATGGGGGCTCAGCCCCTCC
2'-O-Me RNA oligonucleotides for in vitro processing assay
Uc.283+A-pri-mir-195-Binding site
[meG][meC][meC][meA][meG][meG][meG][meA][meA][meG][meC]
2'-O-Me RNA oligonucleotides for in vivo transfections
Uc.283+A-pri-miR-195-Binding site
[meG][meG][meC][meG][meG][meU][meC][meC][meU][meU][meC][meG][meU][meG]
Luciferase constructions
C-3xtargetmiR195-XhoI-s AAAAACTCGAGGCCAATATTTCTGTGCTGCTAAAAGCCAATATTTCTGTGCTGCTAAAAGCCAATATTTCTGTGCTGCTAAAAGCGGCCGCAAAAA
C-3xtargetmiR195-NotI-as AAAAAGCGGCCGCTTTTAGCAGCACAGAAATATTGGCTTTTAGCAGCACAGAAATATTGGCTTTTAGCAGCACAGAAATATTGGCCTCGAGAAAAA
C-3xtargetmiR9-1-XhoI-s AAAAACTCGAGTCATACAGCTAGATAACCAAAGAAAATCATACAGCTAGATAACCAAAGAAAATCATACAGCTAGATAACCAAAGAGCGGCCGCAAAAA
C-3xtargetmiR9-1-NotI-as AAAAAGCGGCCGCTCTTTGGTTATCTAGCTGTATGATTTTCTTTGGTTATCTAGCTGTATGATTTTCTTTGGTTATCTAGCTGTATGACTCGAGAAAAA
shRNAs against Uc.283+A sequence
shRNA1-s GATCCGGTCATAATACCAACTTTATTCAAGAGATAAAGTTGGTATTATGACCTTTTTTG
shRNA1-as AATTCAAAAAAGGTCATAATACCAACTTTATCTCTTGAATAAAGTTGGTATTATGACCG
shRNA2-s GATCCGCCATCGTCCCAAAATAAAATTCAAGAGATTTTATTTTGGGACGATGGTTTTTTG
shRNA2-as AATTCAAAAAACCATCGTCCCAAAATAAAATCTCTTGAATTTTATTTTGGGACGATGGCG
shRNA#ScrambleMSS2-s GATCCGCGCAGAACAAATTCGTCCATTCAAGAGATGGACGAATTTGTTCTGCGTTTTTTACGCGTG
shRNA#ScrambleMSS2-as
AATTCACGCGTAAAAAACGCAGAACAAATTCGTCCATCTCTTGAATGGACGAATTTGTTCTGCGCG
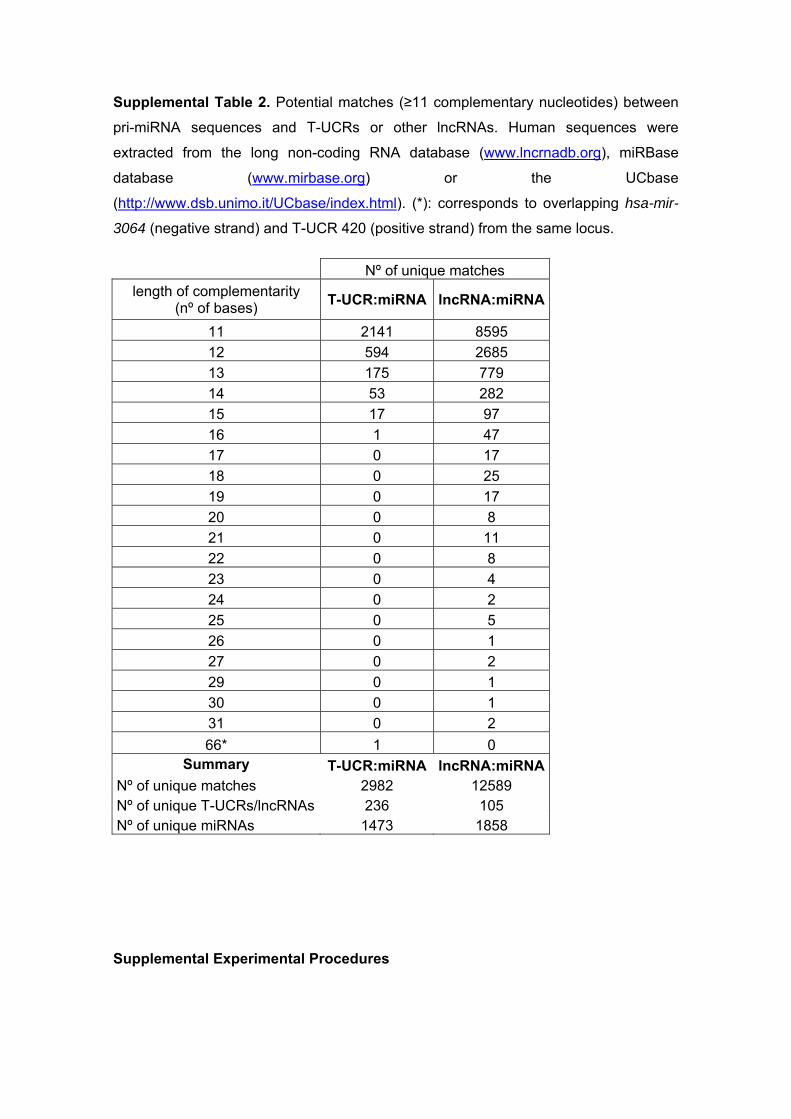
Supplemental Table 2. Potential matches (≥11 complementary nucleotides) between
pri-miRNA sequences and T-UCRs or other lncRNAs. Human sequences were
extracted from the long non-coding RNA database (www.lncrnadb.org), miRBase
database (www.mirbase.org) or the UCbase
(http://www.dsb.unimo.it/UCbase/index.html). (*): corresponds to overlapping hsa-mir-
3064 (negative strand) and T-UCR 420 (positive strand) from the same locus.
Nº of unique matches
length of complementarity (nº of bases)
T-UCR:miRNA lncRNA:miRNA
11 2141 8595
12 594 2685
13 175 779
14 53 282
15 17 97
16 1 47
17 0 17
18 0 25
19 0 17
20 0 8
21 0 11
22 0 8
23 0 4
24 0 2
25 0 5
26 0 1
27 0 2
29 0 1
30 0 1
31 0 2
66* 1 0 Summary T-UCR:miRNA lncRNA:miRNA
Nº of unique matches 2982 12589 Nº of unique T-UCRs/lncRNAs 236 105 Nº of unique miRNAs 1473 1858
Supplemental Experimental Procedures

Cell lines, culture conditions, 5-aza-2′-deoxycytidine treatment and transfections
HCT116 and DKO cells were a gift from Dr. Bert Vogelstein (Johns Hopkins Kimmel
Comprehensive Cancer Center, Baltimore, MD, USA). All other cell lines were obtained
from the ATCC (American Type Culture Collection, Manassas, VA, USA) and grown as
recommended. Cell lines were treated with 2μM 5-aza-2′-deoxycytidine (Sigma-
Aldrich) for 72 h in order to achieve DNA demethylation. For transient transfections, 20
millions of cells were electroporated in 0.8 ml PBS 1x with 40 g of each plasmid
(described below) at 950 F, and 250V in 4mm electroporation cuvettes (Gene Pulser
Xcell Electroporation System, BioRad). For lipid-based transfections, 2’-O-Me-Oligo
(Sigma-Aldrich) (650nM) and 10 g of pCDNA3.1_Uc.283+A(+) were mixed with
Lipofectamine 2000 (Invitrogen) and incubated with cells for 72 h. Knock-down of
Uc.283+A was achieved by lentivirus-mediated overexpression of shRNA-containing
plasmids. Briefly, HEK293T cells were transfected with pLVX-shRNA2-constructs plus
packaging plasmids with jetPRIME® (Polyplus-transfection) according to the
manufacturer’s recommendations. 48 h post-transfection, the supernatant containing
viral particles was used for infection of target cell lines. RNA was collected and
analyzed after at least 8 cell passages post-infection.
Plasmids and site-directed mutagenesis
For overexpression experiments, the previously cloned wild type Uc.283+A was used
(Lujambio, 2010). This vector was also used as a template to perform site-directed
mutagenesis for the pri-miR-195 binding site. Briefly, the primers for consecutive
mutagenesis based on Phusion PCR were C-uc283+A-full lenght-EcoRI with C-
uc283+A-miR195mut-as (PCR 1) and C-uc283+A-miR195mut-s with C-uc283+A-full
length-NotI (PCR 2). The overlapping PCR fragments were mixed and amplified using
C-uc283+A-full length-EcoRI and C-uc283+A-full length-NotI primers and finally cloned
into pCDNA3.1(+). For knock-down experiments, shRNA1 and shRNA2 target the 5’
GGTCATAATACCAACTTTATT3’ and 5’GCCATCGTCCCAAAATAAAAT3’ sequences,

respectively, of Uc.283+A RNA, and were expressed by cloning oligos shRNA1-s and
shRNA1-as or shRNA2-s and shRNA2-as into the BamHI and EcoRI sites of vector
pLVX-shRNA2 (Clontech). Supplemental Table I contains the full list of oligos used for
cloning.
For the in vitro pri-miRNA processing assays, short forms of wild type and mutant
Uc.283+A were used. C-uc283+A-∆1-410, ∆652-2227-EcoRI and C-uc283+A-∆1-410,
∆652-2227-NotI primers were used with the parental plasmid as a template. The short
mutant version was generated as described before, with the internal primers C-
uc283+A-∆1-410, ∆652-2227miR195mut-s and C-uc283+A-∆1-410, ∆652-
2227miR195mut-as. Moreover, the constructs that serve as template for the in vitro
transcription, were cloned directly from genomic DNA (in the case of pri-miR-195 wild
type) and, for the pri-miR-195 mut5 substrate, by using primers that introduce the
pertinent mutation. See Supplementary Table 1 for primer sequences.
For the electrophoretic mobility shift assay, the hsa-miR-195 stem-loop sequence (85
nts) was obtained from miRBase Database. The mut11 version was derived from the
wild-type sequence by site-directed mutagenesis with primers C-pre-miR-195mut11-s
and C-pre-miR-195mut11-NotI-as. Wild type and mutant forms for Uc.283+A binding
site, were generated from genomic DNA and next cloned into pCDNA3.1(+).
RNA isolation and miRNA expression microarray analysis
Total RNA was isolated from all cancer cell lines by Trizol extraction according to the
manufacturer's instructions (Invitrogen) and treated with Turbo DNA-free (Ambion).
Microarray experiments, target preparation and array hybridization were performed as
described elsewhere (Liu et al, 2004). This hybridization was carried out on the second
version of miRNA-chip (MDACC miRNA Expression Bioarray 19k v5, based on
miRBase Release 15).

Hybridization signals were detected by biotin binding of a streptavidin-Alexa647
conjugate (one-color signal) using a GenePix 4000B scanner (Axon Instruments).
Images were quantified using the GenePix Pro-6.0 (Axon Instruments).
Quantitative reverse transcription PCR
TaqMan MiRNA assays (Applied Biosystems) were used to quantify the level of mature
miRNAs. Each reverse transcriptase (RT) reaction contained 10 ng of purified RNA.
Real-time PCR reactions for each miRNA (10 μl of final volume) were performed in
triplicate. Reactions were incubated in Applied Biosystems 7900HT Fast Real Time
PCR in 384-well plates. Real-time PCR experiments were performed in triplicate. The
2-(∆∆Ct) method (Livak and Schmittgen, 2001) was used to determine relative
quantitative levels of each miRNAs. RNU6B was used to normalize the data.
For pri-miRNA expression analysis, 1 µg of treated RNA was retrotranscribed using
ThermoScript™ RT–PCR System (Invitrogen). All the real-time PCR reactions were
carried out in triplicate on Applied Biosystems 7900HT Fast Real Time PCR System
using 20 ng of cDNA and 1 µl of Specific Taqman Pri-microRNA Assay per reaction
(Applied Biosystems), in a final volume of 10 μl in a 384-well plate. Data was
normalized to GAPDH expression.
PolyA+/polyA- RNA selection and subcellular fractionation
PolyA+ and polyA- RNAs were separated using the Dynabeads® mRNA Purification Kit
(Life Technologies, #61006), using three rounds of selection. RNAs were then subject
to qRT-PCR. Enrichment was assessed using GAPDH or U6 RNAs as controls and
normalizing relative to the percentage of RNA from each fraction used in the reverse
transcription reaction. Nuclear/cytoplasmic fractionation was performed with the
PARIS™ kit (Life Technologies, #AM1921). Equal amounts of protein from each
fraction were analysed by Western Blot to assess purity by blotting against β-tubulin
(Abcam, #ab21058 (dilution 1/10,000)) or Histone 3 (Abcam, #ab7191 (dilution

1/5000)). Equal amounts of RNA from each fraction were subject to qRT-PCR and the
results were normalized taking into account the total amount of RNA recovered from
each fraction.
RNA-Fluorescence in situ hybridization (RNA-FISH)
A pool of 44 probes tiling the Uc.283+A RNA coupled to TAMRA reporter dye were
designed following Stellaris® RNA FISH Probe designer (Biosearch Technologies).
Cell fixation, permeabilization and hybridization to probe were performed following
Stellaris® FISH Protocols for adherent cells. Image acquisition was carried out on a
Zeiss Axio Observer Z1 Apotome fluorescence microscope.
RNA antisense purification (RAP)
RAP protocol was performed as described before (Engreitz et al, 2013). Briefly, the
Uc.2839+A RNA was tiled with eight 124-nucleotides antisense RNA probes that had
been biotinylated by in vitro transcription. The full-length Uc.283+A was covered by
these probes, with the exception of the region interacting with pri-miRNA and the
fragment corresponding to the amplicon in the RT-PCR (to avoid interference in the RT-
qPCR signal). SK-N-BE(2) cells were cross-linked first with 2mM DSG for 45min at
room temperature and then with 3% formaldehyde for 10min at 37ºC. For each
purification, 100 ng of biotinylated probes were added to the precleared lysates and the
mix was incubated at 45ºC. The probes were then captured by streptavidin beads, and
the elutions for the associated RNA and DNA were performed. As control, the same
experiment was carried out in parallel with probes tiling the unrelated CELF2 mRNA or
with streptavidin beads without any probe. Recovered RNA and DNA samples were
analyzed by RT-qPCR together with 1/10 dilution of the input material.
Luciferase reporter vectors

All reporters were constructed based on psiCHECK™-2 (Promega) that uses Renilla
luciferase as the primary reporter gene. Sequences of interest are inserted into a
multiple cloning region (MCR) located downstream of the Renilla translational stop
codon. Reporter plasmids 3xmiR-195targets or 3xmiR-9-1targets were generated with
oligonucleotides that hold three tandem repeats of the reverse complementary
sequence of mature miR-195 or miR-9-1, respectively (primers used are listed on
Supplemental Table 1). This system also contains a constitutively expressed firefly
luciferase gene in order to normalize the transfection efficiency. Dual-Luciferase
Reporter Assay System (DLR Assay System, Promega) was used for measurements.
Briefly, each transfection was carried out with 10 µg of pCDNA3.1(+)_primiR-195 (or
pri-miR-9-1), pCDNA3.1(+)_Uc.283+Awt, pCDNA3.1(+)_Uc.283+Amut11 and 10 µg of
each Reporter construct. Total amount of transfected DNA was maintained constant
across transfections. After 24h of transfection, cells were collected and the assay was
performed following the manufacturer's instructions. Each luciferase assay was
analyzed based on the ratio Renilla/Firefly.
In vitro pri-miRNA processing assay.
Pri-miR-195 RNA substrates for in vitro processing assays were prepared from DNA
templates by standard in vitro transcription with T7 RNA Polymerase (Roche) in the
presence of [α-32P] ATP (PerkinElmer). RNA substrates for Uc.283+A (wt and mut5)
were obtained by in vitro transcription from linearized DNA templates. Processing
reactions were carried out with total HEK293T extracts as previously described (Guil
and Cáceres, 2007). Briefly, each pri-miRNA was incubated with increasing amounts
(0.21, 0.42 and 0.83 μM) of Uc.283+A (wt or mutant form) or 100, 200 and 500 mM of a
2’-O-methyl-oligonucleotide matching the 11 nucleotides of Uc.283+A that are
complementary to pri-miR-195. The RNA mixture was preheated at 65ºC for 2 min and
cooled to 30ºC. The nuclear extract was then added and the reaction mixture incubated

for a further 90 min at 30ºC. RNAs were phenol-extracted, precipitated and loaded onto
an 8% denaturing polyacrylamide gel.
RNA pull-down
In vitro-transcribed RNAs were covalently linked to adipic acid dihydrazide agarose
beads, as described before (Caputi et al, 1999). Each reaction contained 80 pmol of
pri-miR-195 (wild type or mutant) and increasing amounts (80, 160 and 320 pmol) of
Uc.283+A. Affinity purification of DGCR8 protein was performed with HEK293T nuclear
extracts, and the unbound fraction was blotted with an anti-β actin antibody as a
loading control. The antibodies used in western blot were anti-DGCR8 (# AB90579,
Abcam) and anti-β actin (#A3854, Sigma-Aldrich).
Electrophoretic mobility shift assay
The RNA substrate for the wild-type or mutant miR-195 stem-loop sequence (87 nt)
were obtained and labeled by random biotinylation during the in vitro transcription using
0.25 mM biotin-16-UTP (Roche) from linearized DNA templates (pcDNA3.1(+)_premiR-
195 and pcDNA3.1(+)_premiR-195mut11). Uc.283+A RNA substrates (wt and mutant)
were synthesized by in vitro transcription from linearized DNA templates. Binding
reactions were carried out in 1x binding buffer (20 mM Tris-HCl, pH 8.0, 1 mM DTT, 1
mM MgCl2, 20 mM KCl, 10 mM Na2HPO4-NaH2PO4, pH 8.0) with the biotin-labeled
RNAs alone or in the presence of increasing amounts (0.5-6.25 pmol) of unlabeled
Uc.283+A (wt or mutant). Each RNA mixture was preheated at 70ºC for 5 min and then
cooled and maintained at 30ºC for 20 min. All reactions were then immediately loaded
onto a native 5% polyacrylamide gel, transferred to a nylon membrane and developed
using Ambion’s Brightstar BioDetect Nonisotopic Detection Kit system.
Statistical Evaluation

All statistical analysis were performed using GraphPad Prism Software 5.04. For
comparisons, a Student’s t-test was performed as appropriate. Any P value less than
0.05 was designated with (*); ** P ≤ 0.01, *** P ≤ 0.001 and **** P ≤ 0.0001.
Supplemental References
Caputi, M., Mayeda, A., Krainer, A.R., and Zahler, A.M. (1999). hnRNP A/B proteins
are required for inhibition of HIV-1 pre-mRNA splicing. EMBO J 18, 4060-4067.
Livak, K.J. and Schmittgen, T.D. (2001). Analysis of relative gene expression data
using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25,
402-408.




















