
Noncoding Mutations of HGF Are Associated with Nonsyndromic Hearing Loss, DFNB39
Upload
independentCategory
view
0download
0
Antisense long noncoding RNAs regulate var geneactivation in the malaria parasitePlasmodium falciparumInbar Amit-Avrahama, Guy Poznera, Shiri Eshara, Yair Fastmana, Netanel Kolevzonb, Eylon Yavinb, and Ron Dzikowskia,1
aDepartment of Microbiology and Molecular Genetics, Institute for Medical Research Israel-Canada, The Kuvin Center for the Study of Infectious andTropical Diseases, The Hebrew University-Hadassah Medical School, Jerusalem 91120, Israel; and bInstitute for Drug Research, School of Pharmacy,Faculty of Medicine, The Hebrew University of Jerusalem, Jerusalem 91120, Israel
Edited by Thomas E. Wellems, National Institutes of Health, Bethesda, MD, and approved January 27, 2015 (received for review October 31, 2014)
The virulence of Plasmodium falciparum, the causative agent of thedeadliest form of human malaria, is attributed to its ability to evadehuman immunity through antigenic variation. These parasites alter-nate between expression of variable antigens, encoded bymembersof a multicopy gene family named var. Immune evasion throughantigenic variation depends on tight regulation of var gene expres-sion, ensuring that only a single var gene is expressed at a timewhile the rest of the family is maintained transcriptionally silent.Understanding how a single gene is chosen for activation is criticalfor understanding mutually exclusive expression but remains a mys-tery. Here, we show that antisense long noncoding RNAs (lncRNAs)initiating from var introns are associated with the single active vargene at the time in the cell cycle when the single var upstreampromoter is active. We demonstrate that these antisense transcriptsare incorporated into chromatin, and that expression of these anti-sense lncRNAs in trans triggers activation of a silent var gene ina sequence- and dose-dependent manner. On the other hand, in-terference with these lncRNAs using complement peptide nucleicacid molecules down-regulated the active var gene, erased the epi-genetic memory, and induced expression switching. Altogether, ourdata provide evidence that these antisense lncRNAs play a key rolein regulating var gene activation and mutually exclusive expression.
malaria | Plasmodium falciparum | var genes | noncoding RNA |exclusive expression
The most devastating form of human malaria is caused byPlasmodium falciparum. This protozoan parasite is estimated
to be responsible for the death of up to 1 million people eachyear, primarily pregnant woman and young children (1). Thisintracellular parasite replicates within the circulating blood of aninfected individual, and its virulence is attributed to its ability tomodify the surface of the infected RBCs (iRBCs) and to undergoantigenic variation (2). The main antigenic ligands responsiblefor the cytoadhesive properties of the iRBCs are variable surfaceproteins named P. falciparum erythrocyte membrane protein-1(PfEMP1) (3). These highly polymorphic proteins bind severalendothelial receptors, leading to sequestration of iRBCs, whichthen avoid clearance by the spleen. To avoid the antibody-mediated response against these antigens, the parasites haveevolved a tight mechanism of antigenic switches between ex-pression of different variants of PfEMP1 (4). PfEMP1s areencoded by the multicopy gene family named var, and each of the∼60 var genes in the parasite’s genome encodes a different variantof PfEMP1 (5). The var genes are expressed in a mutually exclusivemanner (i.e., the parasites express only a single var gene at a time,whereas the rest of the var gene family remains transcriptionallysilent) (6, 7). Thus, switches in expression to different var genes,each expressed in a mutually exclusive manner, enable the parasiteto evade immune attack and maintain long-term chronic infections.Changes in var gene expression and the resulting antigenic
variation appear to be controlled at the level of transcription (6)and do not require DNA rearrangements. Therefore, silencing
and activation of these genes are believed to be epigeneticallyregulated and were shown to involve processes of chromatinmodification, subnuclear localization, promoter/promoter inter-actions, and sterile RNAs (8).Each individual var gene contains the regulatory elements that
enable it to maintain a transcriptionally silent state, even whilean adjacent gene may be active, and to be “counted” by themutually exclusive expression mechanism. Apparently, func-tional antigen production is not required for mutually exclusiveexpression (9, 10), which, in turn, was found to be dependent onthe activity of two promoters found in each var gene, one up-stream of the coding region and responsible for production ofthe mRNA and the second within a conserved intron foundwithin each gene (11–13). Transcriptional activity of both varupstream and intron promoters is mediated by PolII (14), andwhereas the var upstream promoter gives rise to the mRNAcoding for PfEMP1 production, the bidirectional intron pro-moter gives rise to rather large sterile transcripts: about 2–2.5 kbin the forward orientation and 1.7 kb in the reverse orientation(15). These long noncoding RNAs (lncRNAs) do not have ORFs,and although they are capped, they are not polyadenylated, in-dicating that these sterile transcripts might be retained and func-tion in the nucleus. Indeed, var-intron sense lncRNAs have beenshown to be associated with chromatin (15). It has been postulatedthat the sense sterile transcripts of var introns might have a role inregulating epigenetic memory by imprinting var genes for silencing
Significance
How cells specifically express only a single gene among nu-merous equivalent copies within their genomes is one of theunsolved mysteries in the field of eukaryotic gene expression.The molecular mechanisms that underlie mutually exclusivegene expression are the key for understanding the virulenceof Plasmodium falciparum, the parasite responsible for thedeadliest form of human malaria. P. falciparum expresses itsprimary virulence determinants in a mutually exclusive mannerand evades human immune attack through switches in ex-pression between different variants of a large gene familynamed var. We found that var-specific antisense long non-coding RNA molecules incorporate into chromatin and de-termine how parasites select a single gene for expression whilethe rest of the family is maintained silenced.
Author contributions: I.A.-A. and R.D. designed research; I.A.-A., G.P., S.E., and R.D. per-formed research; Y.F., N.K., and E.Y. contributed new reagents/analytic tools; I.A.-A. andR.D. analyzed data; and I.A.-A. and R.D. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1420855112/-/DCSupplemental.
E982–E991 | PNAS | Published online February 17, 2015 www.pnas.org/cgi/doi/10.1073/pnas.1420855112
similar to chromosome X inactivation (Xist) in mammals anddosage compensation (Rox1 and Rox2) in Drosophila (15). Thereare several indications to support this hypothesis: First, the intronpromoter is transcriptionally active in the forward orientation atthe time in the cell cycle when var upstream promoters are silent(16); second, the sequence of the transcripts in this orientation issimilar among most of the var gene family because it is encoded bythe highly conserved second exon of var genes; and, third, thesetranscripts appear to be present in all var genes that were exam-ined, whereas var upstream promoters are subjected to mutuallyexclusive expression (5).Very little is known about the var-intron antisense lncRNAs,
and although some studies associated these transcripts with silentvar genes, others provided evidence associating them with activegenes (14, 15, 17–19). Nonetheless, their possible role in var generegulation remains unknown. Given the fact that the intronicantisense lncRNA complements the variable region of var genes,
we hypothesized that the antisense lncRNAs might be involvedin activation of a specific var gene. Here, we show that theseantisense lncRNAs are associated specifically with the singleactive var gene at the time of the cell cycle when var genes areactive. Moreover, we demonstrate that expression of thesetranscripts in trans induces var gene activation, and that inter-ference with these antisense transcripts leads to repression ofthe active gene, erases epigenetic memory, and induces ex-pression switching.
ResultsAntisense Noncoding RNAs Expressed by var Introns Are Associatedwith the Active var Gene. We initially observed that the levels ofvar transcripts measured by RT-quantitative PCR (qPCR) oncDNA produced with random primers, using a primer setdesigned to the 3′ of exon 1, are approximately double the levelsmeasured by a primer set designed to the 5′ of the same exon
0
2
4
6
8
02468
10121416
Rela
�ve
Copy
Num
ber
DC-J 'on'
02468
10121416
PFA
0015
cP
FD00
20c
PFD
1235
w/M
AL8
P1.2
07P
FF00
20c
PF0
8_01
41P
FI18
20w
PF1
1_00
08P
F11_
0521
PF1
3_00
03P
FF00
10w
PFF
1580
cP
F08_
0140
PFL
0020
wP
FA00
05w
PFA
0765
cP
FB00
10w
PFB
1055
c-B
SDP
FC00
05w
/PFC
1120
cP
FD00
05w
PFD
1245
cP
FE00
05w
PFF
1595
cM
al7P
1.21
2M
AL7
P1.
187
PF0
8_01
42M
AL8
P1.
220
PFI
0005
wP
FI18
30c
PF1
0_00
01P
F10_
0406
PF1
1_00
07P
FL00
05w
PFL
0935
cP
FL26
65c
MA
L13P
1.1
MA
L13P
1.35
6M
AL7
P1.
50P
F07_
0050
MA
L7P
1.55
PF0
8_01
06P
F08_
0103
PFL
1950
wP
FD06
30c/
PFD
0635
cP
FD10
05c
PFL
1955
w/P
FL19
70w
PFD
0615
cP
FD06
25c
PFD
0995
cP
FD09
95c/
PFD
1000
cP
FD10
15c-
3'P
FF08
45c
PF0
7_00
48P
F07_
0049
PF0
7_00
51M
AL7
P1.
56P
F08_
0107
PFL
1960
wP
FL00
30c
Rela
�ve
Copy
Num
ber DC-J 'off'
0
2
4
6
8
PFD
1015
c-3'
PFD
1015
c-5'
PFF
0845
c-3'
PFF
0845
c-5'
PFB
1055
c-B
SD
mRNA
an�sense
800bp
150bp
150bp
5’ upsBSD
3’ BSDArg
sense ncRNA
an�sense ncRNA
mRNA
Exon1 Exon 25’ 3’
PFB1055c-bsd (Ups B) var transgene BSD Exon 2
PFD1015c (Ups C) var gene
A
B
D
C
Ups-A Ups-A/B Ups-B Ups-B/C Ups-C Ups-E
an�sense ncRNA
BSD Exon 2
5’ upsBSD 3’ BSD
~800bp ~150bp
an�sense ncRNA
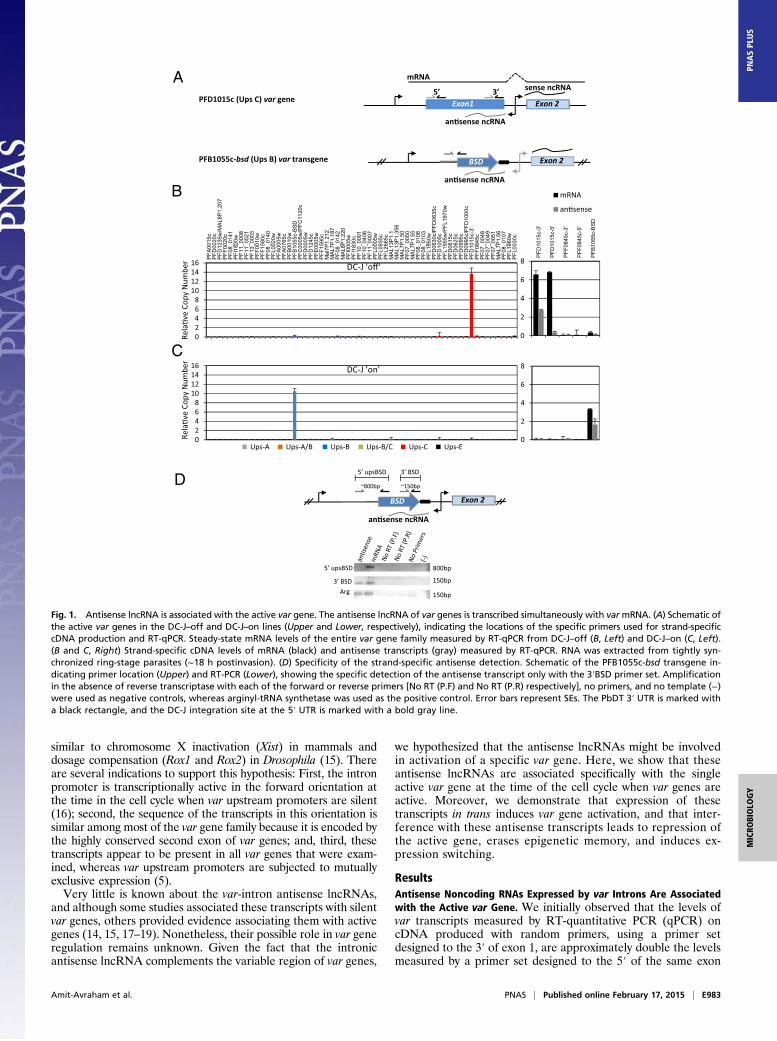
Fig. 1. Antisense lncRNA is associated with the active var gene. The antisense lncRNA of var genes is transcribed simultaneously with varmRNA. (A) Schematic ofthe active var genes in the DC-J–off and DC-J–on lines (Upper and Lower, respectively), indicating the locations of the specific primers used for strand-specificcDNA production and RT-qPCR. Steady-state mRNA levels of the entire var gene family measured by RT-qPCR from DC-J–off (B, Left) and DC-J–on (C, Left).(B and C, Right) Strand-specific cDNA levels of mRNA (black) and antisense transcripts (gray) measured by RT-qPCR. RNA was extracted from tightly syn-chronized ring-stage parasites (∼18 h postinvasion). (D) Specificity of the strand-specific antisense detection. Schematic of the PFB1055c-bsd transgene in-dicating primer location (Upper) and RT-PCR (Lower), showing the specific detection of the antisense transcript only with the 3′BSD primer set. Amplificationin the absence of reverse transcriptase with each of the forward or reverse primers [No RT (P.F) and No RT (P.R) respectively], no primers, and no template (−)were used as negative controls, whereas arginyl-tRNA synthetase was used as the positive control. Error bars represent SEs. The PbDT 3′ UTR is marked witha black rectangle, and the DC-J integration site at the 5′ UTR is marked with a bold gray line.
Amit-Avraham et al. PNAS | Published online February 17, 2015 | E983
MICRO
BIOLO
GY
PNASPL
US

(Fig. S1 A and B), although both primer sets had identical am-plification efficiency. We reasoned that because the promoterwithin the intron is bidirectional, the 3′ primer set could po-tentially detect both the var mRNA and the antisense lncRNAof the intron. We therefore hypothesized that the antisensetranscripts could be associated with the active var gene. To test ifvar-antisense transcripts are associated with the particular tran-scriptional status of an individual var gene, we initially used threeclonal populations of the NF54 line (C3, C350, and chondroitinsulfate A-selected), each predominantly expressing a differentvar upstream promoter (PFD1005c, PFB1055c, and PFL0030c,respectively) (Fig. S1C, Left). To differentiate between themRNA and the antisense lncRNA, we synthesized strand-spe-cific cDNAs from each clonal population (Materials and Meth-ods), and the levels of mRNA and intron antisense lncRNAswere measured from the predominantly active var gene and twosilent genes in each of these populations. In the active genes,both mRNA and antisense lncRNA were detected using theprimer set designed to amplify from the 3′ of exon 1. However,only mRNA, and not the antisense transcripts, was detectedwhen using the primer set designed to amplify at the 5′ of exon 1, aregion that the antisense transcript does not reach. In contrast,no mRNA or antisense lncRNA was detected from the silent vargenes in each of the clonal populations (Fig. S1C, Right). Toconfirm these observations, we used the DC-J transgenic parasiteline (9), which allows selecting for activation of a specific vargene (PFB1055c-bsd), thereby ensuring that the rest of thefamily is completely silent (Fig. 1A). We initially analyzed aclonal population of this line in which the transgene wassilent and another var gene (PFD1015c) was active (Fig. 1B, LeftUpper). We performed the same strand-specific cDNA synthesisas described above to differentiate between mRNA and theintronic antisense lncRNA. As in the NF54 WT clones, we coulddetect the antisense lncRNA only from the active gene, whereas notranscript was detected from the silent var genes (PFB1055c-bsdand PFF0845c) (Fig. 1B, Right Upper). As expected, the antisensetranscripts were only detected by primers designated to the 3′ ofexon 1. This parasite population was then grown under drugselection (2 μg/mL blasticidin S), thus forcing it to switch andexclusively express another var gene (PFB1055c-bsd) (Fig. 1C,Left Lower). In these parasites as well, the intronic antisenselncRNAs were detected exclusively from the active var genePFB1055c-bsd, and not from the silent var genes (Fig. 1C, RightLower). In the DC-J transgenic line, the long first exon ofPFB1055c was replaced with the short bsd cassette (1,350 bp).Therefore, to be able to distinguish between detection of mRNAand antisense, we used RT-PCR with a forward primer located atthe 5′ UTR of PFB1055c and a reverse primer located in the bsdcoding region. Both mRNA and antisense from the active vargene were detected using the 3′BSD primer set, whereas the5′upsBSD primer set could detect only the mRNA (Fig. 1D). Inaddition, these intronic antisense transcripts were only detectedfrom the active gene at the time in the cell cycle when the varupstream promoter is active. In both DC-J “on” and “off” drugselection, as well as in the NF54 clones C3 and G6, the antisensetranscripts were only detected in tightly synchronized parasites18 h postinvasion (hpi) and not in late stages (36 hpi) (Fig. S1D).These observations are consistent with recent RNA-sequencingdata from clonal parasite populations (17). Altogether, thesedata shows that var antisense lncRNAs are associated with thesingle active var gene and that these transcripts are detected inlate ring stages at the time when the single var upstream pro-moter is transcriptionally active.
Antisense lncRNA Is Incorporated into Chromatin. The var antisenselncRNA is transcribed by PolII (14), and although it is capped, itappears not to be polyadenylated, and thus is believed to remainin the nucleus (15). In recent years, it has been observed that
transcriptional enhancers can recruit PolII to specific loci andmediate transcription of lncRNAs, which were postulated to beinvolved in gene regulation, possibly through incorporation intochromatin structures and recruitment of transcription factorsand epigenetic modulators (20). Therefore, as a first step to in-vestigate the possibility that var antisense transcripts have a reg-ulatory role, we were interested to check if they are associatedwith chromatin. We performed ChIP using antibodies against thecore histone H3, and α–C-myc antibodies were used as negativecontrols. The precipitated RNA was used as a template forstrand-specific cDNA synthesis that enabled us to compare thelevel of enrichment of these antisense transcripts in active andsilent var genes. We used the same clones of the DC-J transgenicline mentioned above (Fig. 1) after drug selection, where thetransgene (PFB1055c-bsd) is active and all the remaining vargenes are silent (DC-J–on), and compared it with the clone thatgrew without drug and predominantly expressed PFD1015c(DC-J–off). An additional advantage of using this line is thatit enables us to distinguish between the PFB1055c-bsd mRNA,antisense lncRNA, and sense lncRNA by using different sets ofprimers (Fig. 2A). Following ChIP and RNA recovery of syn-chronized ring-stage parasites, the transcript levels were mea-sured by RT-qPCR and the level of enrichment was calculatedrelative to the values from the same parasite cultures beforeperforming ChIP (input). We found that in DC-J–on, the anti-sense of the active gene (PFB1055c-bsd) was significantlyenriched in the α-H3 ChIP fraction compared with the input,whereas the antisense transcript of the silent gene could not bedetected (Fig. 2 C and D, Upper). Similarly, in the DC-J–offline, where PFD1015c was predominantly expressed, its anti-sense transcript was significantly enriched in the ChIP fraction,whereas only low levels of antisense transcripts could be detectedfrom PFB1055c-bsd, which is mostly silenced in this line (Fig. 2 Cand D, Lower). In both lines, the sense noncoding transcripts ofthe intron, which are known to be incorporated into chromatin(15) and are expressed from numerous silent var genes, weresignificantly enriched in the α-H3 fraction as expected, indicatingsuccessful IP of these sterile transcripts. Similar to previousstudies, we could detect low levels of sense transcripts in earlystages even though these sense transcript are known to be highlytranscribed in late stages (15, 16, 21). No transcripts wereenriched when α–C-myc antibodies were used as negative con-trols for the IPs (Fig. 2 C and D), as expected, because chromatinwas selectively pulled down by α-H3 and not α–C-myc antibodies(Fig. 2B). Interestingly, detectable levels of mRNA of the se-lected transgene (PFB1055c-bsd) were precipitated with H3 anti-body, whereas no levels of mRNA transcripts were detected whenthe endogenous PFD1015c was active in the DC-J–off line.However, only the lncRNAs were enriched by ChIP, whereasmRNAs were not. These results suggest that the var antisenselncRNAs, which are associated with the single active var gene,are incorporated into chromatin.
Silent var Gene Is Activated by Expression of Its Specific AntisenseTranscript. The association of the var-intron–derived antisenselncRNA with the single active var gene and their interaction withchromatin led us to hypothesize that these particular steriletranscripts might play a role in var gene activation. To addressthis possibility, we transfected the DC-J–off line, where thetransgene PFB1055c-bsd is silent (and does not express antisensenoncoding RNA) and the endogenous var gene PFD1015c ispredominantly active, with a plasmid that expresses the specificantisense transcript of the silent var PFB1055c-bsd (Fig. 3). Wemonitored the expression of the entire var gene family usingRT-qPCR to determine if the expression of the antisenselncRNAs in trans could influence var gene expression. To ex-clude the possible effect of spontaneous switching events, thesepatterns were compared with the expression of the entire var
E984 | www.pnas.org/cgi/doi/10.1073/pnas.1420855112 Amit-Avraham et al.

gene family in DC-J–off parasites that were not transfected andgrown in parallel. In addition, we compared var expression withparasites that were transfected with a mock plasmid expressingluciferase instead of the sterile transcripts, as well as a plasmid inwhich the antisense transcript is fused to an unrelated 3′ UTR(Fig. 3A). Both parasite populations that were either not trans-fected or transfected with the mock plasmid had similar var ex-pression patterns, and both predominantly expressed PFD1015c(Fig. 3B, Upper and Middle Upper, respectively). However, this
expression pattern was altered in the DC-J–off parasites trans-fected with the plasmid expressing the PFB1055c intronic anti-sense transcripts. In this population, the transcript levels ofPFD1015c were down-regulated, whereas the transcript level ofPFB1055-bsd was up-regulated more than threefold without anyblasticidin selection (Fig. 3B, Middle Lower). These data suggestthat expression of particular var antisense lncRNAs in trans canalter their mRNA expression patterns and increase the activa-tion rate of a specific var gene. Interestingly, parasites that were
A
DC-J ‘on’ DC-J ‘off’α-H3 α-Cmyc α-H3 α-Cmyc
15kD
IP
α-H3
B
DC
DC-J ‘on’
DC-J ‘off’ mRNA
Exon1 Exon 23’
BSD Exon 2
mRNA
PFB1055c
PFD1015c
-150%
-100%
-50%
0%
50%
100%
150%
Enric
hmen
trel
a�ve
toin
put
P=0.037
P=0.002
-150%
-100%
-50%
0%
50%
100%
150%
PFD1
015c
-3'
PFB1
055c
-BSD
PFB1
055c
Exon
2
PFD1
015c
-3'
PFB1
055c
-BSD
PFB1
055c
Exon
2
α-H3 α-Cmyc
Enric
hmen
trel
a�ve
toin
put
P=0.001
P=0.0064
mRNA an�sense sense
0
1
2
3
4
5
6
7
Rela
�ve
Copy
Num
ber
0
1
2
3
4
5
6
7
PFD1
015c
-3'
PFB1
055c
-BSD
PFB1
055c
Exon
2
PFD1
015c
-3'
PFB1
055c
-BSD
PFB1
055c
Exon
2
PFD1
015c
-3'
PFB1
055c
-BSD
PFB1
055c
Exon
2
input α-H3 α-Cmyc
Rela
�ve
Copy
Num
ber
DC-J‘off’
DC-J‘on’
mRNA an�sense sense
Fig. 2. Antisense lncRNA is associated with chromatin. RNA immunoprecipitation of var transcripts was performed on tightly synchronized ring-stageparasites (∼18 h postinvasion) using antibodies against the core histone H3 or with antibodies against C-myc as a negative control. (A) Schematic of the activevar genes in each clone indicating the location of the specific primer sets that were used for strand-specific cDNA synthesis and RT-qPCR. The PbDT 3′ UTR ismarked with a black rectangle. (B) Western blot of the fractions that were immunoprecipitated with α-H3 and α–C-myc antibodies from DC-J–on and DC-J–offpopulations and detected by the α-H3 antibody. (C) Levels of sense and antisense var transcripts detected by RT-qPCR from the IP samples (α-H3 or α–C-myc)and non-IP samples (input). Different primer sets presented on top of each column were used differentially to detect mRNA (black), antisense (gray), andsense ncRNA (white). (D) Enrichment levels of var sense and antisense transcripts detected by RT-qPCR and calculated as the relative percentile of the levelsdetected in the IP samples (α-H3 or α–C-myc) relative to the input. The levels of strand-specific cDNA were measured by RT-qPCR relative to arginyl-tRNAsynthetase (PFL0900c). The active gene in each parasite line is marked with a red arrowhead. Each value is the average of two biological replicates and threetechnical triplicates. Error bars represent SEs. Samples differences were tested using the Student’s t test at P < 0.05.
Amit-Avraham et al. PNAS | Published online February 17, 2015 | E985
MICRO
BIOLO
GY
PNASPL
US

transfected with episomes in which the antisense transcripts werefused to an unrelated 3′UTR showed similar activation levels of thePFB1055-bsd and down-regulation of PFD1015c (Fig. 3B, Lower).We noted that the level of steady-state antisense lncRNAs wasapproximately doubled when they were fused to the 3′ UTR. Alltransfected parasite populations carried similar copy numbers of theexpression vectors (Fig. 3B, Right).To verify that the up-regulation effect of the episomal anti-
sense is sequence-specific, the antisense expression vector wastransfected into a clonal NF54 population (A3) that predominantly
expresses PFF0845c. This WT line does not contain the sequencefound in the transgene PFB1055c-bsd, and thus does not share anysequence similarity with the episomal antisense transcripts. As wehypothesized, the antisense of the transgene PFB1055c-bsd hadno effect on var transcription in the WT NF54 population, in-dicating that the mechanism of activation is sequence-specific(Fig. S2).We tested whether this transcriptional alteration is indeed
a consequence of a switch in var gene expression by putting theseparasites under blasticidin pressure and measured their growth
0
5
10
15
20
25
30Ge
neCo
pyN
umbe
r0
1
2
3
4
5
Rela
�ve
Copy
Num
ber BbI1055Dh
0
1
2
3
4
5
PFA
0015
cP
FD00
20c
PFD
1235
w/M
AL8
P1.2
07P
FF00
20c
PF0
8_01
41P
FI18
20w
PF1
1_00
08P
F11_
0521
PF1
3_00
03P
FF00
10w
PFF
1580
cP
F08_
0140
PFL
0020
wP
FA00
05w
PFA
0765
cP
FB00
10w
PFB
1055
c-B
SDP
FC00
05w
/PFC
1120
cP
FD00
05w
PFD
1245
cP
FE00
05w
PFF
1595
cM
al7P
1.21
2M
AL7
P1.
187
PF0
8_01
42M
AL8
P1.
220
PFI
0005
wP
FI18
30c
PF1
0_00
01P
F10_
0406
PF1
1_00
07P
FL00
05w
PFL
0935
cP
FL26
65c
MA
L13P
1.1
MA
L13P
1.35
6M
AL7
P1.
50P
F07_
0050
MA
L7P
1.55
PF0
8_01
06P
F08_
0103
PFL
1950
wP
FD10
05c
PFD
0630
c/P
FD06
35c
PFL
1955
w/P
FL19
70w
PFD
0615
cP
FD06
25c
PFD
0995
cP
FD09
95c/
PFD
1000
cP
FD10
15c-
5'P
FF08
45c
PF0
7_00
48P
F07_
0049
PF0
7_00
51M
AL7
P1.
56P
F08_
0107
PFL
1960
wP
FL00
30c
Dhf
rLU
Cep
isom
al B
SD
Rela
�ve
Copy
Num
ber DC-J 'off'
15
14
0
5
10
15
20
25
30
Gene
Copy
Num
ber
0
5
10
15
20
25
30
Gene
Copy
Num
ber
0
1
2
3
4
5
6
PFB
1055
c-B
SDLU
ChD
HFR
epis
omal
BS
D
Gene
Copy
Num
ber
0
1
2
3
4
5
Rela
�ve
Copy
Num
ber hLI1055Dh
12
14
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
0 24 48 72
%Pa
rasit
emia
hours post treatment
DC-J onDC-J offDC-J off + hLIDhDC-J off + BbIDhDC-J off +3'BbIDhA3 +BbIDh
0
1
2
3
4
5
Rela
�ve
Copy
Num
ber 3'BbI1055Dh
CA
B
Ups-A Ups-A/B Ups-B Ups-B/C Ups-C Ups-E
DC-J
pBbI1055Dh
phLI1055Dh
PFB1055c 5’-UTR
BSD
PFB1055cintron
Exon 2
hDHFRLuc
hDHFRBSD
p3’BbI1055Dh hDHFRBSD3’
Fig. 3. Silent var gene is activated by expression of its specific antisense transcript. (A) Schematics of the PFB1055-bsd recombinant locus in the DC-J parasiteline (Upper); the plasmid pBbI1055Dh, which was used to express the specific antisense lncRNA of PFB1055-bsd (Middle Upper); the plasmid phLI1055Dh, whichexpresses an unrelated sequence (luciferase) that was used as a negative control (Middle Lower); and the plasmid p3′BbIDh, which expresses the antisenselncRNA of PFB1055-bsd fused to an unrelated 3′ UTR (Lower). The locations of the differential primers used for RT-qPCR are indicated by arrows. The PbDT 3′UTR is marked with a black rectangle. (B, Left) Steady-state mRNA levels of the entire var gene family measured by RT-qPCR from DC-J–off (Upper), DC-J–offtransfected with the control plasmid phLI1055Dh expressing an unrelated sequence (Middle Upper), DC-J–off transfected with the plasmid pBbI1055Dhexpressing the var-specific antisense transcript (Middle Lower), and DC-J–off transfected with the plasmid p3′BbI1055Dh expressing the var-specific antisensetranscript terminated by an unrelated 3′ UTR (Lower). RNA was extracted from tightly synchronized ring-stage parasites (∼18 h postinvasion). (B, Right)Plasmid copy number measured by qPCR of genomic DNA is presented. (C) Growth curves of parasite populations grown under selection with 2 μg/mLblasticidin. Growth curves were calculated for untransfected DC-J–on parasites that were preselected using blasticidin (brown); untransfected DC-J–off, wherethe bsd is predominantly silent (green); DC-J–off transfected with the mock plasmid phLI1055Dh (purple); DC-J–off transfected with the plasmid expressing thevar-specific antisense pBbI1055Dh (light blue); DC-J–off transfected with the plasmid expressing the var-specific antisense fused with the 3′ UTR p3′BbI1055Dh(gray); and WT NF54 clone (A3) transfected with the plasmid expressing the var-specific antisense pBbI1055Dh (orange). Error bars represent SEs.
E986 | www.pnas.org/cgi/doi/10.1073/pnas.1420855112 Amit-Avraham et al.
DC-J
pBbI5RDh hDHFRBSD
pBbRDh hDHFRBSD
0
1
2
3
4
5
6
Rela
�ve
Copy
Num
ber BbI5RDh
0
1
2
3
4
5
6
Rela
�ve
Copy
Num
ber BbRDh
0
5
10
15
PFB
1055
c-B
SDLU
ChD
HFR
epis
omal
BS
D
Gene
Copy
Num
ber
0
5
10
15
Gen
eCo
pyN
umbe
r
0
5
10
15
Gen
eCo
pyN
umbe
r
0
1
2
3
4
5
6
PFA
0015
cP
FD00
20c
PFD
1235
w/M
AL8
P1.2
07P
FF00
20c
PF0
8_01
41P
FI18
20w
PF1
1_00
08P
F11_
0521
PF1
3_00
03P
FF00
10w
PFF
1580
cP
F08_
0140
PFL
0020
wP
FA00
05w
PFA
0765
cP
FB00
10w
PFB
1055
c-B
SDP
FC00
05w
/PFC
1120
cP
FD00
05w
PFD
1245
cP
FE00
05w
PFF
1595
cM
al7P
1.21
2M
AL7
P1.
187
PF0
8_01
42M
AL8
P1.
220
PFI
0005
wP
FI18
30c
PF1
0_00
01P
F10_
0406
PF1
1_00
07P
FL00
05w
PFL
0935
cP
FL26
65c
MA
L13P
1.1
MA
L13P
1.35
6M
AL7
P1.
50P
F07_
0050
MA
L7P
1.55
PF0
8_01
06P
F08_
0103
PFL
1950
wP
FD10
05c
PFD
0630
c/P
FD06
35c
PFL
1955
w/P
FL19
70w
PFD
0615
cP
FD06
25c
PFD
0995
cP
FD09
95c/
PFD
1000
cP
FD10
15c-
5'P
FF08
45c
PF0
7_00
48P
F07_
0049
PF0
7_00
51M
AL7
P1.
56P
F08_
0107
PFL
1960
wP
FL00
30c
Dhf
rLu
cep
isom
al B
SD
Rela
�ve
Copy
Num
ber DC-J 'off'
8
9
0
1
2
3
4
5
6
0 24 48 72
%Pa
rasit
emia
hours post treatment
DC-J onDC-J offDC-J off + BbRDhDC-J off + BbI5RDh
PFB1055c 5’-UTR
BSD
PFB1055cintron
Exon 2
CA
B
Ups-A Ups-A/B Ups-B Ups-B/C Ups-C Ups-E
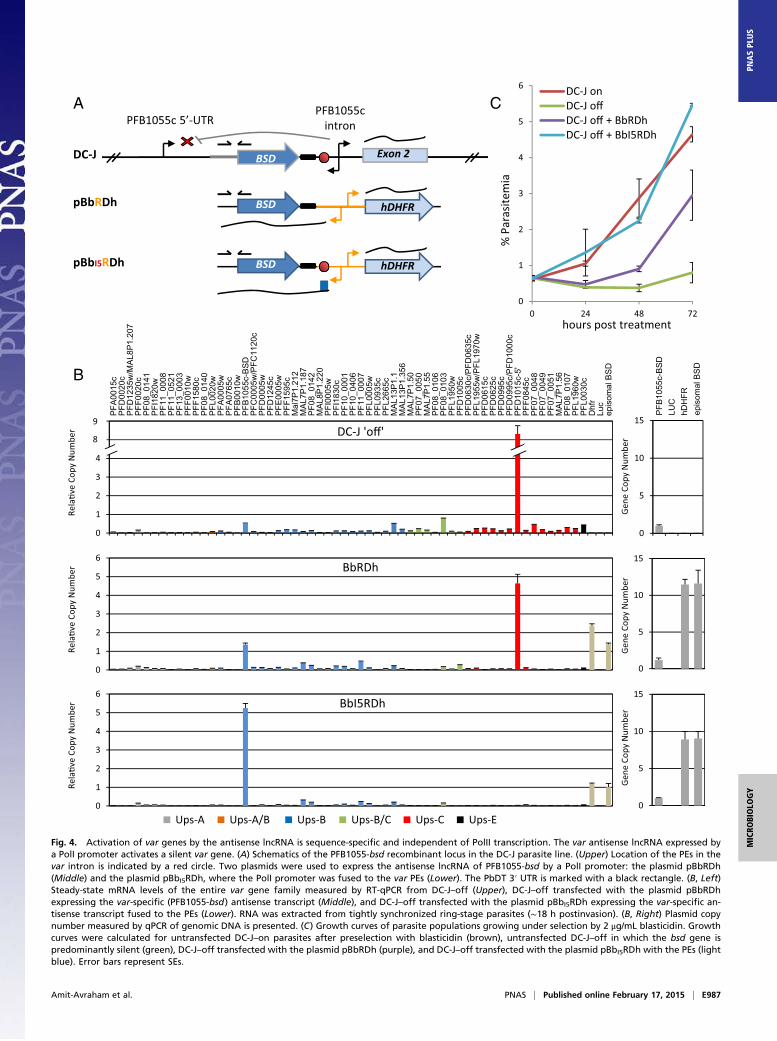
Fig. 4. Activation of var genes by the antisense lncRNA is sequence-specific and independent of PolII transcription. The var antisense lncRNA expressed bya PolI promoter activates a silent var gene. (A) Schematics of the PFB1055-bsd recombinant locus in the DC-J parasite line. (Upper) Location of the PEs in thevar intron is indicated by a red circle. Two plasmids were used to express the antisense lncRNA of PFB1055-bsd by a PolI promoter: the plasmid pBbRDh(Middle) and the plasmid pBbI5RDh, where the PolI promoter was fused to the var PEs (Lower). The PbDT 3′ UTR is marked with a black rectangle. (B, Left)Steady-state mRNA levels of the entire var gene family measured by RT-qPCR from DC-J–off (Upper), DC-J–off transfected with the plasmid pBbRDhexpressing the var-specific (PFB1055-bsd) antisense transcript (Middle), and DC-J–off transfected with the plasmid pBbI5RDh expressing the var-specific an-tisense transcript fused to the PEs (Lower). RNA was extracted from tightly synchronized ring-stage parasites (∼18 h postinvasion). (B, Right) Plasmid copynumber measured by qPCR of genomic DNA is presented. (C) Growth curves of parasite populations growing under selection by 2 μg/mL blasticidin. Growthcurves were calculated for untransfected DC-J–on parasites after preselection with blasticidin (brown), untransfected DC-J–off in which the bsd gene ispredominantly silent (green), DC-J–off transfected with the plasmid pBbRDh (purple), and DC-J–off transfected with the plasmid pBbI5RDh with the PEs (lightblue). Error bars represent SEs.
Amit-Avraham et al. PNAS | Published online February 17, 2015 | E987
MICRO
BIOLO
GY
PNASPL
US

rates. We assumed that the growth rate of parasites in which thePFB1055c-bsd transgene is activated, and therefore express bsd,will be faster than in parasites in which this gene is initially silent.As additional controls, we used DC-J–on parasites that had beenpreviously selected on 2 μg/mL blasticidin, activated PFB1055c-bsd, and therefore are blasticidin-resistant. In addition, we testedthe growth of blasticidin-sensitive NF54 parasites (clone A3)transfected with the vector expressing the PFB1055c-bsd anti-sense lncRNAs. We found that the growth rate of the DC-J–offparasites expressing the PFB1055c-bsd antisense lncRNAS fromeither the BI1055Dh or 3′BI1055Dh expression vector was higherthan in parasites that were not transfected (DC-J–off) or in par-asites that were transfected with the mock plasmid (DC-J–off +hLI1055Dh) (Fig. 3C). The WT NF54 parasites transfected withthe antisense expression vector (A3 + BI1055Dh) died afterblasticidin selection and never recovered (Fig. 3C). Similar up-regulation of a silent var gene was obtained when expressingthese antisense noncoding RNAs (ncRNAs) in another trans-genic line [B15C2 (9)] where var PFL0020w was integrated withbsd cassette that enables its activation in a mutually exclusivemanner (Fig. S3). Without bsd selection, these parasites undergonormal in vitro var switching. We identified a parasite populationwhere the PFL0020w-bsd transgene was silent (B15C2-G) andtransfected it with vectors expressing either the specific antisenselncRNAs (BbIYh) or a mock plasmid expressing an unrelatedsequence (hLIYh) (Fig. S3A). In these experiments as well, theexpression of the specific antisense transcripts activated theendogenous PFL0020c-bsd, whereas the mock plasmid had noeffect (Fig. S3 B and C). These results suggest that expression ofparticular var antisense ncRNAs in trans accelerates var gene ac-tivation, providing the first evidence, to our knowledge, of theinvolvement of these sterile transcripts in var gene regulation.
Activation of var Genes by the Antisense lncRNA Is Sequence-Specificand Is Independent of PolII Transcription. Both sense and antisensencRNAs derived from the promoter found within the var intronare transcribed by PolII (14). We were interested to test ifactivation of var genes by the antisense transcripts is PolII-dependent. Therefore, the PolII-dependent promoter in ourexpression vectors was replaced with a PolI-dependent pro-moter (rRNA promoter) so that the antisense of the transgenePFB1055c-bsd is transcribed by PolI (pBbRDh). We have pre-viously shown that var silencing is regulated by specific DNAelements [pairing elements (PEs)] found in each var upstreamregulatory region and intron (22). As a consequence of theirposition within each intron, these PEs are found also in each ofthe var antisense transcripts. Therefore, in one of our expressionvectors, we fused the sequence containing the var PE motif im-mediately after the PolI promoter (pBbI5RDh) (Fig. 4A). Theseplasmids were transfected into the DC-J–off clone, where thetransgene PFB1055c-bsd is silent (Fig. 4B, Upper). We foundthat the var antisense transcripts activated the PFB1055c-bsdeven when they are expressed by a PolI promoter. Moreover,when the PEs are added to these transcripts, the level of acti-vation increased as reflected in the increase in bsd mRNAtranscription level, as well as in the increase in the growth rateunder blasticidin selection (Fig. 4 B and C). Transfection of thesame parasite line with mock plasmids expressing unrelatedsequences by a PolI promoter, with or without the PEs, did notalter var gene activation (Fig. S4). These results indicate thatactivation of var genes by the antisense transcripts is sequence-specific but is independent of PolII transcription per se.
Dose-Dependent Transcriptional Activation of Endogenous var Genesby Gradual Overexpression of Their Specific var Antisense lncRNA. Toexplore the possibility that the antisense ncRNAs can activate vargenes in a dose-dependent manner, we took advantage of a par-asite line that contains several copies of var-luciferase (var-luc)
transgenes, which were integrated into the genome as four con-catameric repeats [D10 (23)]. In each of these four var-luc trans-genes, luciferase expression is controlled by the var upstreampromoters, but because these var upstream promoters are pairedwith an intron, they are silent by default and express no luciferase.In addition to these paired var upstream promoters, this parasiteline contains a var upstream promoter that is unpaired with anintron, and thus constitutively expresses low levels of luciferase(Fig. S5A, Upper). This parasite line was then transfected witha plasmid that expresses the specific antisense of the transgenes(pLhIBb) or a mock plasmid expressing an unrelated sequence(phGIBb) (Fig. S5A). The use of blasticidin as a selectable markerenables fine-tuned selection of parasites that carry increasingnumbers of the expression vectors, and thus enables one to expressincreasing levels of any gene of interest as previously shown (21,24). Following selection on 2 μg/mL blasticidin, the cultures weresplit and exposed to increasing concentrations of 6 and 10 μg/mLblasticidin. As expected, both new lines expressed increasing levelsof either the episomal antisense (luc) or the unrelated sequence(GFP) (Fig. S5B) from the pLhIBb and phGIBb plasmids, re-spectively. In each of these transfected lines, the activity of theendogenous var-luc was monitored by RT-qPCR (Fig. S5C) and byquantifying protein activity (Fig. S5D). We found that as we in-creased expression of the var-luc antisense lncRNAs, the levels ofluciferase from the endogenous var-luc had increased as well ina dose-dependent manner both at the level of steady-state mRNAand luciferase activity (Fig. S5 C and D, dark gray columns).However, increased expression of the unrelated GFP sequencehad no effect on var-luc activation (Fig. S5 C and D, light graycolumns). Overall, our data show that a cluster of multicopy vargenes can be gradually activated by fine-tuned overexpression oftheir specific antisense lncRNAs, providing evidence for their rolein var transcriptional activation.
Interference with the Antisense lncRNAs Down-Regulates the Activevar Gene, Erases the Epigenetic Memory, and Induces var GeneSwitching. After demonstrating that silent var genes could beactivated by expressing their specific antisense lncRNAs, wewere interested to test if interference with the antisenselncRNAs of the active gene would down-regulate it. We recentlyshowed that peptide nucleic acids (PNAs) could be used todown-regulate gene expression by hybridizing to mRNAs (25).We designed a PNA molecule that specifically complementsa sequence of 17 bp found specifically in the antisense lncRNAof the active var gene (PFD1015c) in the DC-J–off parasite line.This parasite culture was split and incubated with 10 μM com-plementary PNA or scrambled PNA that contains the samenucleotides, which were shuffled and thus contained no sequencesimilarity to the P. falciparum genome. An additional culture wasgrown in parallel without any PNA treatment. We found that thepopulation that was incubated with the scrambled PNA pre-dominantly expressed PFD1015c similar to the population cul-tured in parallel that was not incubated with PNA (Fig. S6A,Upper and Middle). However, in the population that was in-cubated with the PNA molecules that complement the antisenseof PFD1015c, this gene was significantly down-regulated (Fig.S6A, Lower) and expressed steady-state mRNA levels that were∼70% lower than the parasites that were not incubated withPNA or those parasites incubated with the scrambled PNA(Fig. S6B).It has been shown previously that the slow switch rate of in-
ternal var genes, such as PFD1015c (26), is maintained throughtight regulation of epigenetic memory (27) and that this memoryis maintained by active transcription of the active gene (24). Wedecided to test if down-regulation of a telomeric var gene thathas a higher switch rate would erase the epigenetic memory andcause antigenic switching. We therefore designed PNAs thatcomplement the antisense of the telomeric active var gene
E988 | www.pnas.org/cgi/doi/10.1073/pnas.1420855112 Amit-Avraham et al.

(PFD1055c-bsd) in the DC-J–on line and compared var tran-scription patterns in cultures that were incubated with the specificPNA with those cultures that were not incubated or were in-cubated with the scrambled PNA. We found that incubation withthe scrambled PNA had no effect on var transcription andPFB1055c-bsd was predominantly expressed similar to the cul-ture that was not incubated with PNA (Fig. 5A, Upper andMiddle). However, the active gene (PFB1055c-bsd) was signifi-cantly down-regulated (∼70%) in the culture that was incubatedwith the specific PNAs that complement its antisense lncRNAs(Fig. 5 A, Lower and B). Furthermore, by down-regulating thetranscription levels of this gene, we erased the epigeneticmemory that imprinted this gene as the active gene and inducedswitching to activation of other var genes in the population,
which became transcriptionally heterogeneous. Altogether, thisset of experiments indicates that interference with the antisenselncRNAs down-regulates the active var gene, erases the epige-netic memory, and induces var gene switching.
DiscussionThe lncRNAs were shown to play an important role in epigeneticregulation of gene transcription in several eukaryotic systems(28). However, although ncRNAs were predicted to be abundantin Plasmodium (29–34), their function remained unknown. Here,we provide evidence that antisense lncRNAs are involved in vargene activation. These transcripts are specifically associated withthe active gene and are detected from late ring stages parallelto the time during the intraerythrocytic development when the
0
1
2
3
4
Rela
�ve
Copy
Num
ber DC-J 'on' α-PFB1055c-bsd an�sense PNA
0
1
2
3
4
Rela
�ve
Copy
Num
ber DC-J 'on' scrambled PNA
0
1
2
3
4
PFA
0015
cP
FD00
20c
PFD
1235
w/M
AL8
P1.
207
PFF
0020
cP
F08_
0141
PFI
1820
wP
F11_
0008
PF1
1_05
21P
F13_
0003
PFF
0010
wP
FF15
80c
PF0
8_01
40P
FL00
20w
PFA
0005
wP
FA07
65c
PFB
0010
wP
FB10
55c-
BS
DP
FC00
05w
/PFC
1120
cP
FD00
05w
PFD
1245
cP
FE00
05w
PFF
1595
cM
al7P
1.21
2M
AL7
P1.
187
PF0
8_01
42M
AL8
P1.
220
PFI
0005
wP
FI18
30c
PF1
0_00
01P
F10_
0406
PF1
1_00
07P
FL00
05w
PFL
0935
cP
FL26
65c
MA
L13P
1.1
MA
L13P
1.35
6M
AL7
P1.
50P
F07_
0050
MA
L7P
1.55
PF0
8_01
06P
F08_
0103
PFL
1950
wP
FD06
30c/
PFD
0635
cP
FD10
05c
PFL
1955
w/P
FL19
70w
PFD
0615
cP
FD06
25c
PFD
0995
cP
FD09
95c/
PFD
1000
cP
FD10
15c
PFF
0845
cP
F07_
0048
PF0
7_00
49P
F07_
0051
MA
L7P
1.56
PF0
8_01
07P
FL19
60w
PFL
0030
c
Rela
�ve
Copy
Num
ber DC-J 'on' no PNA
A
BUps-A Ups-A/B Ups-B Ups-B/C Ups-C Ups-E
0%
20%
40%
60%
80%
100%
Scrambled α-PFB1055c-bsdan�sense
Fig. 5. Interference with the antisense lncRNAs down-regulates the active var gene, erases epigenetic memory, and induces var gene switching. PNAscomplementary to the antisense transcript of the active var gene (PFB1055c-bsd) in the DC-J–on parasite population (growing without selection) were used toblock its activity by hybridization. (A) Steady-state mRNA levels of the entire var gene family measured by RT-qPCR from DC-J–on with no PNA treatment(Upper), DC-J–on incubated with 10 μM scrambled PNA (Middle), and DC-J–on incubated with 10 μM specific α–PFB1055c-bsd antisense PNA (Lower). RNA wasextracted from tightly synchronized ring-stage parasites (∼18 h postinvasion). (B) Levels of down-regulation of PFB1055c-bsd were calculated as the ratiobetween the levels of steady-state mRNA in PNA-treated parasites relative to nontreated parasites. These levels are presented as a percentage of down-regulation. All values presented are the average of three biological replicates and three technical triplicates. Error bars represent SEs.
Amit-Avraham et al. PNAS | Published online February 17, 2015 | E989
MICRO
BIOLO
GY
PNASPL
US

mRNA-producing var upstream promoter is active. In contrast tothe mRNA, which is translated into PfEMP1, we show that theseantisense lncRNAs are associated with chromatin. We also showthat expression of particular var antisense lncRNAs in transactivates the corresponding endogenous var gene in a sequence-and dose-dependent manner. Last, we present evidence that in-terference with these antisense lncRNAs leads to transcriptionalrepression of an endogenous active var gene, which triggers vargene switching. Thus far, the promoter found in var introns hasbeen implicated in var silencing and considered to act as a silencerof upstream var promoter (11–13, 35). The data presented heresuggest that the bidirectional promoter within var intron may actas a silencer when expressing the sense lncRNAs and as an en-hancer when expressing the antisense transcripts. It is still unclearwhat determines the timing and orientation of the intron promoteractivity, and perhaps it involves some alternative transcriptionstart site (TSS) found in the intron promoter (15), which could beassociated with different promoter orientation. Recent data pro-vided evidence for the mechanism of var gene silencing and epi-genetic memory linking PolII transcription of sense lncRNA torecruitment of a var-specific heterochromatin marker (17, 36).The mechanisms by which PolII transcription in the antisenseorientation is involved in var gene activation are unknown;nonetheless, our results indicate that these antisense transcriptsactivate var genes independent of PolII transcription.The best-studied examples for epigenetic regulation by ncRNAs
include Xist in mammals (37–39) and Rox1 and Rox2 in Drosophila(40, 41), where ncRNAs incorporated into chromatin structuresinitiate a cascade of events that epigenetically mark these partic-ular loci for either silent heterochromatin or active euchromatin.Recent studies have shown that lncRNAs are associated withenhancer regions and have important roles in activatingtranscription of neighboring genes (42–46). These lncRNAswere proposed to incorporate into chromatin and to playa role in recruiting transcription factors and chromatin modu-lators required for gene activation (20). In mouse cortical neu-rons, PolII transcription of enhancer ncRNAs was positivelycorrelated with the level of RNA synthesis at nearby genes,suggesting that long-distance interactions between enhancersand promoters are necessary for activation (43). In addition, inHOXA activation, it was shown that HOTTIP (HOXA transcriptat the distal tip) lncRNA functions in cis on its target genes bymodifying chromatin structure and coordinates activation by re-cruiting specific chromatin-modifying factors to a specific genomelocation (45, 47). The incorporation of var antisense lncRNAs intochromatin and their function in a sequence-dependent mannercould indicate that similar mechanisms are involved in var geneactivation. These antisense lncRNAs may contribute to remodelingof chromatin conformation and recruitment of activation factors tothe locus of the active var gene. In addition, var antisense tran-scripts may contribute to the maintenance of epigenetic memory byrecruiting chromatin-modifying enzymes that drive specific histonemarks, which epigenetically imprint the active locus for many cells’cycles and ensure a very slow switch rate. Furthermore, var si-lencing by the intron promoter was shown to be mediated byinsulator-like PEs that bind specific nuclear proteins (22). The
intronic PEs are found upstream to the intronic TSS; therefore,the antisense transcripts contain the complementary sequenceof the PEs. The data presented here indicate that the presenceof the PE sequence increases the level of activation by the an-tisense transcripts. It is possible that when incorporating intochromatin, these transcripts could hybridize to the cDNA se-quence and interfere with the binding of nuclear proteins to thePEs. Preventing binding of proteins associated with silencingmight influence chromatin conformation and change it from com-pact heterochromatin into loose euchromatin that enables a singlevar gene to be accessible for the transcriptional machinery (Fig. S7).Our findings that antisense lncRNAs are involved in var gene
activation open new avenues to explore mutually exclusive expres-sion of var genes, particularly how a single var gene is chosen to beactive and what mechanisms are involved in the epigenetic memory.
Materials and MethodsDetails of plasmid construction andmethodologies that have been previouslydescribed are provided in SI Materials and Methods.
Extraction of DNA, RNA, cDNA Synthesis, and RT-qPCR. Extraction of genomic DNAand RNA and cDNA synthesis were performed as described (9, 24). Strand-specificcDNAs from mRNA and sense transcripts were synthesized by using specificreverse (R) primers, whereas cDNA from antisense transcripts was synthesizedusing forward (F) primers for specific var genes as detailed in each figure andlisted in Table S1. The cDNA of the control genes was synthesized with both Fand R primers specific to the following: arginyl-tRNA synthetase (PFL0900c),seryl-tRNA synthetase (PF07_0073), actin (PFL2215), and glutaminyl-tRNA syn-thetase (PF13_0170). For each cDNA synthesis, we have included three negativecontrols: without reverse transcriptase (no reverse transcriptase-F and no reversetranscriptase-R, respectively) and without primers. All primers used are listed inTable S1. RT-qPCR was performed as described (48). Additional primers used inthis study are listed in Table S1. All values presented are relative copy num-bers to the housekeeping gene arginyl-tRNA synthetase (PFL0900c). Each valueis the average of at least two biological replicates and three technical triplicates.
RNA Immunoprecipitation. RNA immunoprecipitation was done as described(49) using α-H3 antibody (Ab1791; Abcam) bound to Protein-G Sepharosebeads (Santa Cruz Biotechnology). Beads bound to α–C-myc antibody(Santa Cruz Biotechnology) or no antibody were used as negative con-trols. Bound RNA was collected in TRIzol LS Reagent (Life Technologies)and purified on a PureLink column (Invitrogen).
PNA Synthesis and Treatment. PNA sequences, equipped with a C-terminaloctalysine moiety, were synthesized as described (25). PNA sequences aredetailed in Table S2. DC-J–on and DC-J–off parasite lines were tightly syn-chronized as described above. Late-stage parasites were recovered andplaced back in culture. Twenty-two hours after synchronization, the drugselection from the DC-J–on parasite line was removed and each parasite linewas divided into three cultures and incubated for an additional 48 h with10 μM PNA designed to complement the antisense lncRNA of the activegene, 10 μM scrambled PNA, and no PNA treatment. Parasites from eachculture were then collected for RT-qPCR analysis as described above.
ACKNOWLEDGMENTS. We thank Dr. Yoav Smith for his help in performingthe statistical analysis and Dr. Adina Heinberg for critical reading of themanuscript. This work was supported partially by Israeli Academy for ScienceGrant 141/13 and continued to be funded by European Research Council(erc.europa.eu) Consolidator Grant 615412 (to R.D.). I.A.-A. was supportedby the Abisch–Frenkel Foundation.
1. WHO (2010) World Malaria Report 2010 (World Health Organization, Geneva).2. Miller LH, Baruch DI, Marsh K, Doumbo OK (2002) The pathogenic basis of malaria.
Nature 415(6872):673–679.3. Baruch DI, Gormely JA, Ma C, Howard RJ, Pasloske BL (1996) Plasmodium falciparum
erythrocyte membrane protein 1 is a parasitized erythrocyte receptor for adherenceto CD36, thrombospondin, and intercellular adhesion molecule 1. Proc Natl Acad SciUSA 93(8):3497–3502.
4. Smith JD, et al. (1995) Switches in expression of Plasmodium falciparum var genescorrelate with changes in antigenic and cytoadherent phenotypes of infected eryth-rocytes. Cell 82(1):101–110.
5. Su XZ, et al. (1995) The large diverse gene family var encodes proteins involved incytoadherence and antigenic variation of Plasmodium falciparum-infected eryth-rocytes. Cell 82(1):89–100.
6. Scherf A, et al. (1998) Antigenic variation in malaria: in situ switching, relaxed andmutually exclusive transcription of var genes during intra-erythrocytic developmentin Plasmodium falciparum. EMBO J 17(18):5418–5426.
7. Chen Q, et al. (1998) Developmental selection of var gene expression in Plasmodiumfalciparum. Nature 394(6691):392–395.
8. Dzikowski R, Deitsch KW (2009) Genetics of antigenic variation in Plasmodium falci-parum. Curr Genet 55(2):103–110.
9. Dzikowski R, Frank M, Deitsch K (2006) Mutually exclusive expression of virulence genes bymalaria parasites is regulated independently of antigen production. PLoS Pathog 2(3):e22.
10. Voss TS, et al. (2006) A var gene promoter controls allelic exclusion of virulence genesin Plasmodium falciparum malaria. Nature 439(7079):1004–1008.
11. Calderwood MS, Gannoun-Zaki L, Wellems TE, Deitsch KW (2003) Plasmodium falci-parum var genes are regulated by two regions with separate promoters, one
E990 | www.pnas.org/cgi/doi/10.1073/pnas.1420855112 Amit-Avraham et al.

upstream of the coding region and a second within the intron. J Biol Chem278(36):34125–34132.
12. Gannoun-Zaki L, Jost A, Mu J, Deitsch KW, Wellems TE (2005) A silenced Plasmodiumfalciparum var promoter can be activated in vivo through spontaneous deletion ofa silencing element in the intron. Eukaryot Cell 4(2):490–492.
13. Dzikowski R, et al. (2007) Mechanisms underlying mutually exclusive expression ofvirulence genes by malaria parasites. EMBO Rep 8(10):959–965.
14. Kyes S, et al. (2007) Plasmodium falciparum var gene expression is developmentallycontrolled at the level of RNA polymerase II-mediated transcription initiation. MolMicrobiol 63(4):1237–1247.
15. Epp C, Li F, Howitt CA, Chookajorn T, Deitsch KW (2009) Chromatin associated senseand antisense noncoding RNAs are transcribed from the var gene family of virulencegenes of the malaria parasite Plasmodium falciparum. RNA 15(1):116–127.
16. Kyes SA, et al. (2003) A well-conserved Plasmodium falciparum var gene shows anunusual stage-specific transcript pattern. Mol Microbiol 48(5):1339–1348.
17. Jiang L, et al. (2013) PfSETvs methylation of histone H3K36 represses virulence genesin Plasmodium falciparum. Nature 499(7457):223–227.
18. Zhang Q, et al. (2014) Exonuclease-mediated degradation of nascent RNA silencesgenes linked to severe malaria. Nature 513(7518):431–435.
19. Ralph SA, et al. (2005) Transcriptome analysis of antigenic variation in Plasmodiumfalciparum—var silencing is not dependent on antisense RNA. Genome Biol 6(11):R93.
20. Ong CT, Corces VG (2011) Enhancer function: New insights into the regulation oftissue-specific gene expression. Nat Rev Genet 12(4):283–293.
21. Epp C, Raskolnikov D, Deitsch KW (2008) A regulatable transgene expression systemfor cultured Plasmodium falciparum parasites. Malar J 7:86.
22. Avraham I, Schreier J, Dzikowski R (2012) Insulator-like pairing elements regulatesilencing and mutually exclusive expression in the malaria parasite Plasmodium fal-ciparum. Proc Natl Acad Sci USA 109(52):E3678–E3686.
23. Frank M, et al. (2006) Strict pairing of var promoters and introns is required for vargene silencing in the malaria parasite Plasmodium falciparum. J Biol Chem 281(15):9942–9952.
24. Dzikowski R, Deitsch KW (2008) Active transcription is required for maintenance ofepigenetic memory in the malaria parasite Plasmodium falciparum. J Mol Biol 382(2):288–297.
25. Kolevzon N, Nasereddin A, Naik S, Yavin E, Dzikowski R (2014) Use of peptide nucleicacids to manipulate gene expression in the malaria parasite Plasmodium falciparum.PLoS ONE 9(1):e86802.
26. Frank M, Dzikowski R, Amulic B, Deitsch K (2007) Variable switching rates of malariavirulence genes are associated with chromosomal position. Mol Microbiol 64(6):1486–1498.
27. Chookajorn T, et al. (2007) Epigenetic memory at malaria virulence genes. Proc NatlAcad Sci USA 104(3):899–902.
28. Mercer TR, Dinger ME, Mattick JS (2009) Long non-coding RNAs: Insights into func-tions. Nat Rev Genet 10(3):155–159.
29. Mourier T, et al. (2008) Genome-wide discovery and verification of novel structuredRNAs in Plasmodium falciparum. Genome Res 18(2):281–292.
30. Gunasekera AM, et al. (2004) Widespread distribution of antisense transcripts in thePlasmodium falciparum genome. Mol Biochem Parasitol 136(1):35–42.
31. Patankar S, Munasinghe A, Shoaibi A, Cummings LM, Wirth DF (2001) Serial analysisof gene expression in Plasmodium falciparum reveals the global expression profile oferythrocytic stages and the presence of anti-sense transcripts in the malarial parasite.Mol Biol Cell 12(10):3114–3125.
32. Li F, Sonbuchner L, Kyes SA, Epp C, Deitsch KW (2008) Nuclear non-coding RNAs aretranscribed from the centromeres of Plasmodium falciparum and are associated withcentromeric chromatin. J Biol Chem 283(9):5692–5698.
33. Broadbent KM, et al. (2011) A global transcriptional analysis of Plasmodium falcipa-rum malaria reveals a novel family of telomere-associated lncRNAs. Genome Biol12(6):R56.
34. Sierra-Miranda M, et al. (2012) Two long non-coding RNAs generated from sub-telomeric regions accumulate in a novel perinuclear compartment in Plasmodiumfalciparum. Mol Biochem Parasitol 185(1):36–47.
35. Deitsch KW, Calderwood MS, Wellems TE (2001) Malaria. Cooperative silencing ele-ments in var genes. Nature 412(6850):875–876.
36. Ukaegbu UE, et al. (2014) Recruitment of PfSET2 by RNA polymerase II to variantantigen encoding loci contributes to antigenic variation in P. falciparum. PLoS Pathog10(1):e1003854.
37. Rasmussen TP, Wutz AP, Pehrson JR, Jaenisch RR (2001) Expression of Xist RNA issufficient to initiate macrochromatin body formation. Chromosoma 110(6):411–420.
38. Lee JT, Davidow LS, Warshawsky D (1999) Tsix, a gene antisense to Xist at theX-inactivation centre. Nat Genet 21(4):400–404.
39. Jeon Y, Sarma K, Lee JT (2012) New and Xisting regulatory mechanisms of X chro-mosome inactivation. Curr Opin Genet Dev 22(2):62–71.
40. Meller VH, et al. (2000) Ordered assembly of roX RNAs into MSL complexes on thedosage-compensated X chromosome in Drosophila. Curr Biol 10(3):136–143.
41. Park Y, Kelley RL, Oh H, Kuroda MI, Meller VH (2002) Extent of chromatin spreadingdetermined by roX RNA recruitment of MSL proteins. Science 298(5598):1620–1623.
42. Ørom UA, et al. (2010) Long noncoding RNAs with enhancer-like function in humancells. Cell 143(1):46–58.
43. Kim TK, et al. (2010) Widespread transcription at neuronal activity-regulated en-hancers. Nature 465(7295):182–187.
44. De Santa F, et al. (2010) A large fraction of extragenic RNA pol II transcription sitesoverlap enhancers. PLoS Biol 8(5):e1000384.
45. Wang KC, et al. (2011) A long noncoding RNA maintains active chromatin to co-ordinate homeotic gene expression. Nature 472(7341):120–124.
46. Ørom UA, Shiekhattar R (2011) Long non-coding RNAs and enhancers. Curr OpinGenet Dev 21(2):194–198.
47. Tsai MC, et al. (2010) Long noncoding RNA as modular scaffold of histone modifi-cation complexes. Science 329(5992):689–693.
48. Fastman Y, Noble R, Recker M, Dzikowski R (2012) Erasing the epigenetic memory andbeginning to switch—The onset of antigenic switching of var genes in Plasmodiumfalciparum. PLoS ONE 7(3):e34168.
49. Mair GR, et al. (2006) Regulation of sexual development of Plasmodium bytranslational repression. Science 313(5787):667–669.
Amit-Avraham et al. PNAS | Published online February 17, 2015 | E991
MICRO
BIOLO
GY
PNASPL
US
Copyright © 2022 FDOKUMEN




















