
p21-Activated Kinase 5 (Pak5) Localizes to Mitochondria and Inhibits Apoptosis by Phosphorylating...
14
MOLECULAR AND CELLULAR BIOLOGY, Aug. 2003, p. 5526–5539 Vol. 23, No. 16 0270-7306/03/$08.000 DOI: 10.1128/MCB.23.16.5526–5539.2003 Copyright © 2003, American Society for Microbiology. All Rights Reserved. p21-Activated Kinase 5 (Pak5) Localizes to Mitochondria and Inhibits Apoptosis by Phosphorylating BAD Sophie Cotteret, Zahara M. Jaffer, Alexander Beeser, and Jonathan Chernoff* Fox Chase Cancer Center, Philadelphia, Pennsylvania Received 6 December 2002/Returned for modification 27 January 2003/Accepted 15 May 2003 Pak5 is the most recently identified and least understood member of the p21-activated kinase (Pak) family. This kinase is known to promote neurite outgrowth in vitro, but its localization, substrates, and effects on cell survival have not been reported. We show here that Pak5 has unique properties that distinguish it from all other members of the Pak family. First, Pak5, unlike Pak1, cannot complement an STE20 mutation in Sac- charomyces cerevisiae. Second, Pak5 binds to the GTPases Cdc42 and Rac, but these GTPases do not regulate Pak5 kinase activity, which is constitutive and stronger than any other Pak. Third, Pak5 prevents apoptosis induced by camptothecin and C2-ceramide by phosphorylating BAD on Ser-112 in a protein kinase A-inde- pendent manner and prevents the localization of BAD to mitochondria, thereby inhibiting the apopto- tic cascade that leads to apoptosis. Finally, we show that Pak5 itself is constitutively localized to mitochondria, and that this localization is independent of kinase activity or Cdc42 binding. These features make Pak5 unique among the Pak family and suggest that it plays an important role in apoptosis through BAD phosphorylation. The p21-activated kinases (Paks) are members of a growing class of Rac/Cdc42-associated Ser/Thr protein kinases, charac- terized by a highly conserved amino-terminal Cdc42/Rac inter- active binding (CRIB) domain and a carboxyl terminal kinase domain (2, 9, 10, 22, 35). Six human Paks, which fall into two subfamilies, have been identified. Group A Paks (Paks 1, 2, and 3) exhibit 80 to 90% sequence identity within their catalytic domains, whereas the recently identified group B Paks (Paks 4, 5, and 6), while highly related to each other, show only about 40 to 50% identity to the kinase domains of the group A Paks (9, 22). The group A Pak subfamily contains additional character- istic motifs, in particular an autoinhibitory domain located down- stream of the CRIB domain which inactivates the catalytic func- tion of these enzymes by intramolecular interaction with the kinase domain (6, 28, 30); this motif is absent in group B Paks. As effectors of Rho family GTPases, Paks play an important role in the regulation of morphology and motility by regulating the actin cytoskeleton (13, 15, 25, 29, 34, 36). While we know in reasonable detail how group A Paks are regulated and what their targets are, we know much less about the group B Paks. Group B Paks are much less closely related to one another than are group A Paks, especially in the regions outside the CRIB and kinase domains. Of the group B Paks, Pak4 is the best characterized. Unlike that of Paks 1, 2, and 3, the inter- action of Cdc42 with Pak4 is thought to regulate translocation of this kinase to the Golgi apparatus, rather than its activation (1). Pak4 is involved in reorganization of the actin cytoskel- eton, but only one likely substrate (LIM kinase) has been identified thus far (7). Pak6 interacts with the androgen and estrogen receptors and has been reported by one group to translocate into the nucleus with the androgen receptor in response to androgen (27, 41). Pak5 is expressed primarily in neuronal tissues and induces neurite-like projections when overexpressed in NIE-115 cells (8). These data suggest that the group B Paks have disparate cellular functions. Both group A and B Paks have been implicated in apoptosis, possessing either proapoptotic (Pak2) or antiapoptotic (Pak1 and Pak4) properties (18, 23, 31, 32, 38). Pak2 is cleaved during apoptosis, most likely by caspase 3; this cleavage leads to its activation, and the activated kinases in turn are thought to contribute to morphological and membrane changes that occur during apoptosis (26, 31, 40). In contrast Pak1 and Pak4, which are not cleaved by caspases, have been reported to protect cells from apoptosis (18, 31, 32, 38). The survival signal induced by Paks is thought to be related to phosphorylation of the apo- ptotic protein BAD on Ser-112 (Pak1 and Pak4) and Ser-136 (Pak1) (18, 31, 32, 38), though it is not known if these phos- phorylations are directly catalyzed by Paks in vivo. BAD is a proapoptotic member of the BCL-2 family that has been shown to translocate between cytosol and mitochondrial membrane- based partners such as BCL-2 or BCL-x L . This complex for- mation inhibits the ability of BCL-2 and BCL-x L to block the release of cytochrome c from mitochondria, a critical step in the activation of the downstream caspase protease cascade (16, 19, 20). BAD is rapidly phosphorylated on two serine residues in response to a survival factor, interleukin-3. Activated Akt has been shown to phosphorylate the proapoptotic BAD pro- tein on Ser-136, while a recent study identified mitochondrion- associated protein kinase A (PKA) as a BAD Ser-112-specific kinase (17). The phosphorylation of BAD results in its disso- ciation from complexes with BCL-2 and BCL-x L and its sub- sequent association with the cytosolic adapter protein 14-3-3 (10, 11). The uncomplexed BCL-x L is then capable of suppress- ing cell death responses by blocking the release of mitochon- drial cytochrome c (18). Here, we investigate the biochemical and functional prop- erties of Pak5, particularly with regard to its role in apoptosis. We show that Pak5 function differs from that of group A Paks such as Pak1, since it does not complement Ste20 and it is not activated by GTPases. We show that Pak5 induces resistance to * Corresponding author. Mailing address: Fox Chase Cancer Cen- ter, 7701 Burholme Ave, Philadelphia, PA 19111. Phone: (215) 728- 5319. Fax: (215) 728-3616. E-mail: [email protected]. 5526 on February 20, 2016 by guest http://mcb.asm.org/ Downloaded from
Transcript of p21-Activated Kinase 5 (Pak5) Localizes to Mitochondria and Inhibits Apoptosis by Phosphorylating...

MOLECULAR AND CELLULAR BIOLOGY, Aug. 2003, p. 5526–5539 Vol. 23, No. 160270-7306/03/$08.00�0 DOI: 10.1128/MCB.23.16.5526–5539.2003Copyright © 2003, American Society for Microbiology. All Rights Reserved.
p21-Activated Kinase 5 (Pak5) Localizes to Mitochondria andInhibits Apoptosis by Phosphorylating BAD
Sophie Cotteret, Zahara M. Jaffer, Alexander Beeser, and Jonathan Chernoff*Fox Chase Cancer Center, Philadelphia, Pennsylvania
Received 6 December 2002/Returned for modification 27 January 2003/Accepted 15 May 2003
Pak5 is the most recently identified and least understood member of the p21-activated kinase (Pak) family.This kinase is known to promote neurite outgrowth in vitro, but its localization, substrates, and effects on cellsurvival have not been reported. We show here that Pak5 has unique properties that distinguish it from allother members of the Pak family. First, Pak5, unlike Pak1, cannot complement an STE20 mutation in Sac-charomyces cerevisiae. Second, Pak5 binds to the GTPases Cdc42 and Rac, but these GTPases do not regulatePak5 kinase activity, which is constitutive and stronger than any other Pak. Third, Pak5 prevents apoptosisinduced by camptothecin and C2-ceramide by phosphorylating BAD on Ser-112 in a protein kinase A-inde-pendent manner and prevents the localization of BAD to mitochondria, thereby inhibiting the apopto-tic cascade that leads to apoptosis. Finally, we show that Pak5 itself is constitutively localized to mitochondria,and that this localization is independent of kinase activity or Cdc42 binding. These features make Pak5 uniqueamong the Pak family and suggest that it plays an important role in apoptosis through BAD phosphorylation.
The p21-activated kinases (Paks) are members of a growingclass of Rac/Cdc42-associated Ser/Thr protein kinases, charac-terized by a highly conserved amino-terminal Cdc42/Rac inter-active binding (CRIB) domain and a carboxyl terminal kinasedomain (2, 9, 10, 22, 35). Six human Paks, which fall into twosubfamilies, have been identified. Group A Paks (Paks 1, 2, and3) exhibit 80 to 90% sequence identity within their catalyticdomains, whereas the recently identified group B Paks (Paks 4,5, and 6), while highly related to each other, show only about 40to 50% identity to the kinase domains of the group A Paks (9,22). The group A Pak subfamily contains additional character-istic motifs, in particular an autoinhibitory domain located down-stream of the CRIB domain which inactivates the catalytic func-tion of these enzymes by intramolecular interaction with thekinase domain (6, 28, 30); this motif is absent in group B Paks.
As effectors of Rho family GTPases, Paks play an importantrole in the regulation of morphology and motility by regulatingthe actin cytoskeleton (13, 15, 25, 29, 34, 36). While we knowin reasonable detail how group A Paks are regulated and whattheir targets are, we know much less about the group B Paks.Group B Paks are much less closely related to one anotherthan are group A Paks, especially in the regions outside theCRIB and kinase domains. Of the group B Paks, Pak4 is thebest characterized. Unlike that of Paks 1, 2, and 3, the inter-action of Cdc42 with Pak4 is thought to regulate translocationof this kinase to the Golgi apparatus, rather than its activation(1). Pak4 is involved in reorganization of the actin cytoskel-eton, but only one likely substrate (LIM kinase) has beenidentified thus far (7). Pak6 interacts with the androgen andestrogen receptors and has been reported by one group totranslocate into the nucleus with the androgen receptor inresponse to androgen (27, 41). Pak5 is expressed primarily inneuronal tissues and induces neurite-like projections when
overexpressed in NIE-115 cells (8). These data suggest that thegroup B Paks have disparate cellular functions.
Both group A and B Paks have been implicated in apoptosis,possessing either proapoptotic (Pak2) or antiapoptotic (Pak1and Pak4) properties (18, 23, 31, 32, 38). Pak2 is cleaved duringapoptosis, most likely by caspase 3; this cleavage leads to itsactivation, and the activated kinases in turn are thought tocontribute to morphological and membrane changes that occurduring apoptosis (26, 31, 40). In contrast Pak1 and Pak4, whichare not cleaved by caspases, have been reported to protect cellsfrom apoptosis (18, 31, 32, 38). The survival signal induced byPaks is thought to be related to phosphorylation of the apo-ptotic protein BAD on Ser-112 (Pak1 and Pak4) and Ser-136(Pak1) (18, 31, 32, 38), though it is not known if these phos-phorylations are directly catalyzed by Paks in vivo. BAD is aproapoptotic member of the BCL-2 family that has been shownto translocate between cytosol and mitochondrial membrane-based partners such as BCL-2 or BCL-xL. This complex for-mation inhibits the ability of BCL-2 and BCL-xL to block therelease of cytochrome c from mitochondria, a critical step inthe activation of the downstream caspase protease cascade (16,19, 20). BAD is rapidly phosphorylated on two serine residuesin response to a survival factor, interleukin-3. Activated Akthas been shown to phosphorylate the proapoptotic BAD pro-tein on Ser-136, while a recent study identified mitochondrion-associated protein kinase A (PKA) as a BAD Ser-112-specifickinase (17). The phosphorylation of BAD results in its disso-ciation from complexes with BCL-2 and BCL-xL and its sub-sequent association with the cytosolic adapter protein 14-3-3�(10, 11). The uncomplexed BCL-xL is then capable of suppress-ing cell death responses by blocking the release of mitochon-drial cytochrome c (18).
Here, we investigate the biochemical and functional prop-erties of Pak5, particularly with regard to its role in apoptosis.We show that Pak5 function differs from that of group A Pakssuch as Pak1, since it does not complement Ste20 and it is notactivated by GTPases. We show that Pak5 induces resistance to
* Corresponding author. Mailing address: Fox Chase Cancer Cen-ter, 7701 Burholme Ave, Philadelphia, PA 19111. Phone: (215) 728-5319. Fax: (215) 728-3616. E-mail: [email protected].
5526
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

apoptosis induced by two distinct stimuli: camptothecin (CA),a topoisomerase I inhibitor, and C2-ceramide (C2C), a well-known analog to a second messenger released from plasmamembrane during apoptosis. Expression of Pak5 inhibits PARPand caspase 3 cleavage and promotes BAD phosphorylation onSer-112, whereas the KD mutant does not protect against apo-ptosis. Pak5 phosphorylates BAD Ser-112 independently ofPKA, and also promotes the phosphorylation of Ser-136, mostlikely via activation of Akt. Moreover, Pak5 prevents BADtranslocation to mitochondria and more importantly, Pak5 lo-calizes to mitochondria, and this localization is independent ofkinase activity or the ability to bind Cdc42.
MATERIALS AND METHODS
Reagents and antibodies. CA and myelin basic protein (MBP) were purchasedfrom Sigma, and C2C, PKA myristoylated inhibitor, and polyclonal anticalnexinwere from Calbiochem. Mouse monoclonal anti-c-myc (sc-40, clone 9E10), rabbitpolyclonal anti-c-myc (clone A14), and anti-glutathione S-transferase (anti-GST)were from Santa Cruz Biotechnology. Mouse monoclonal antihemagglutinin(anti-HA) (12CA5J) was purchased from Babco. Rabbit polyclonal anti-BAD,anti-phospho-BAD to Ser-112, anti-BCL-xL, and anti-phospho-Akt to serine 473were from Cell Signaling Inc. Anti-phospho-Ser-136 was from Upstate Biotech-nologies. Mouse monoclonal anti-PARP antibody was from Pharmingen, andpolyclonal anti-caspase 3 antibody was from Oncogene Research. Monoclonalanti-COX IV was purchased from Molecular Probes, and anti-HSP60 and anti-BCL-2 were from Transduction Laboratories. Antitubulin antibody RB43 spe-cific for Saccharomyces cerevisiae was a gift from V. Guacci (Fox Chase CancerCenter, Philadelphia, Pa.). Polyclonal anti-Pak2 was purchased from Zymed.
Plasmids. Pak5 cDNA was obtained from the RIKEN cDNA collection andsubcloned in pCMV6 containing a Myc epitope tag (36). Kinase-dead (KD)(K478M) mutant of Pak5 (8) was constructed by site-directed mutagenesis(Stratagene QuikChange kit). These Pak5 variants along with the constitutivelyactive allele of Pak5 (PAK5 S573N) were cloned into the S. cerevisiae expressionvector pYes2 (Invitrogen). Additionally human Pak2 cloned into pYES2 was alsoused as a positive control. Plasmids encoding GST-tagged and HA-tagged Cdc42L61 and N17 and Rac1 L61 and N17 were constructed as described elsewhere(33). The Pak4-encoding vector was obtained from Audrey Minden (ColumbiaUniversity, New York, N.Y.). pEBG-BAD encoding for GST-BAD was pur-chased from Cell Signaling. pET vectors encoding His-tagged BAD, BADS112A, and BAD S136A and pcDNA-HA-BAD were a gift from Michael Green-berg (Harvard Medical School, Boston, Mass.). The BAD S112A/S136A doublemutant was obtained by site-directed mutagenesis. HIS-BAD proteins wereinducibly expressed in Escherichia coli strain BL21(DE3)pLysS and purified overNi-nitrilotriacetic acid resin (Qiagen).
Complementation analysis. An S. cerevisiae strain bearing a deletion in STE20(MATa ade2 his3-�1 leu2-�0 met15-�0 ura3-�0 ste20::KanMX) or the parentalisogenic strain BY4741 (Research Genetics) was transformed by a lithium ace-tate procedure (17) with a galactose-inducible expression vector, pYES2 bearingeither no insert or a cDNA encoding Pak-2, Pak5, Pak5 K478M (KD), or S573N(hyperactivated) mutants. Transformants were selected for growth on minimalmedium lacking uracil. To perform qualitative mating assays, the transformantswere serially diluted onto minimal raffinose plates lacking uracil (URA� plates).After the colonies had grown sufficiently the plates were replica plated ontoeither yeast nutrient broth (YNB) or Gal URA� plates and allowed to expressthe transgene for 5 h at 30°C. These plates were then replica plated onto YEPDplates that contained a lawn of the mating tester strain RSY16 (MAT� ade2-1 leu1-2lys2-1 trp5-20 ura1) (a gift from R. Strich, Fox Chase Cancer Center), and thecells were allowed to mate for 6 h before being replicated plated onto GN plates(0.17% YNB, 2% glucose, 0.5% ammonium sulfate). Mating efficiency wasscored by the formation of prototrophic strains on the GN plates.
Protein isolation from S. cerevisiae and immunoprecipitation kinase assays.Fifty-milliliter cultures of the transformants were grown in Raff URA� medium.Galactose was added to 2% and the cells were allowed to express the transgenefor 5 h. At time zero and 5 h, 25-ml fractions were collected by centrifugation andfrozen on dry ice. The pellets were resuspended in 1.0 ml of Tris-EDTA anddivided as follows: 100 �l was transferred to an Eppendorf tube, collected bycentrifugation, and lysed by boiling in 1� sodium dodecyl sulfate-polyacrylamidegel electrophoresis (SDS-PAGE) sample buffer for 10 min. The remaining 900 �lwas collected by centrifugation and lysed with YPER buffer (Pierce) (5 ml/g of
cell pellet) for 25 min at room temperature. The lysates were centrifuged, andthe supernatant was transferred to an Eppendorf tube. The remaining pellet wasresuspended in 500 �l of radioimmunoprecipitation (RIPA) buffer and subjectedto four rounds of freeze-thaw in liquid nitrogen. The lysates were centrifuged for10 min at 14,000 rpm, and the supernatant was transferred to the tube containingthe YPER lysate supernatant. The combined lysates were used for immunopre-cipitation using the anti-Myc polyclonal antibody A14 or the anti-Pak2 specificantibody overnight at 4°C. Protein A/G was added for 2 h and the immunopre-cipitates were washed five times with lysis buffer (66% YPER buffer–33% RIPAbuffer), then three times with kinase assay buffer. Although the YPER-RIPA lysiswas effective, we could not immunologically detect expression of the transgenes inthe lysates or in the immunoprecipitations suggesting a problem of limited solubilityof the mammalian Paks in S. cerevisiae. Multiple extraction procedures failed toresolve this limitation. Kinase assays were performed as described below.
Cell culture, transfection and treatment. All cell lines were grown at 37°C in5% CO2 and cultured in Dulbecco’s modified Eagle’s medium for COS cells andDulbecco’s modified Eagle’s medium–F-12 for CHO cells containing 10% fetalbovine serum, and puromycin (2 �g/ml; Sigma) for CHO stable cell lines trans-fected with either empty vector (pCMV6, control cells), wild-type (WT) Pak5, orKD Pak5 K478M. HMN1 cells were kindly provided by R. Pittman (University ofPennsylvania, Philadelphia). Cells were transfected using Lipofectamine(Gibco), and lysates were collected 48 h after transfection. CHO stable cell linestransfected with pcDNA-HA-BAD were treated 24 h after transfection with CA(10 �M) or C2C (50 �M).
GST pull down assay. To perform Rac and Cdc42 binding assays, 50 �g ofpurified GST fusion proteins coupled to glutathione-Sepharose beads (Amer-sham Pharmacia Biotech) were incubated with buffer A (40 mM HEPES [pH7.4], NP-40 1%, 1 mM EDTA, 150 mM NaCl) supplemented with 10 mM EDTAfor 15 min at room temperature to release any nucleotide, washed with buffer A,incubated in buffer B (buffer A supplemented with 1 mM GTP-�S and 10 mMMgCl2) for 30 min at room temperature, and then washed with buffer B. Next,the proteins were incubated with 250 �l of Cos lysates for 3 h at 4°C. Precipitateswere washed three times with lysis buffer (described below). Bound proteins wereeluted in SDS sample buffer and subjected to immunoblotting with anti-Mycmonoclonal antibody 9E10.
Immunoblot and immunoprecipitation. Cos cells were transfected with appro-priate vectors. Cell extracts were obtained in lysis buffer (50 mM Tris [pH 8], 100mM NaCl, 1% NP-40, 0.1% SDS, 0.5% sodium deoxycholate, 0.5 mM phenyl-methylsulfonyl fluoride, aprotinin [1 �g/ml]), protein concentration was assessedusing bicinchoninic acid (Pierce) and equal amounts of proteins were separatedby SDS-PAGE, transferred to polyvinylidene difluoride membranes (ImmobilonP; Millipore Corp.). Blots were blocked 1 h in Tris-buffered saline–0.1% Tween20 supplemented with 5% nonfat milk, and incubated with primary antibodies foran hour or overnight at 4°C. After washing, blots were incubated with alkaline phos-phatase-conjugated secondary antibodies (Jackson Laboratories), which weredetected using the CPD Star reagent (New England Nuclear). The immunopre-cipitation assay was carried out as follows: whole-cell lysates were incubated withpolyclonal anti-Myc A14 overnight at 4°C and then 1 h at 4°C with protein Aagarose beads (Pierce). Beads were collected by centrifugation and washed withthe lysis buffer described above. Proteins were eluted by boiling in SDS samplebuffer and subjected to immunoblotting, and probed with anti-HA (12CA5J).
Protein kinase assay. Cos cells were transfected with appropriate expressionvectors. Transfected cells were collected in lysis buffer 48 h after transfection andequal amounts of the Myc-tagged proteins, as assayed by Western blotting, werethen immunoprecipitated with anti-Myc antibody A14 and protein A-agarose.After incubation, the immunoprecipitates were washed twice in lysis buffer andtwice in a kinase buffer (50 mM Tris [pH 7.5], 100 mM NaCl, 1 mM MnCl2, 10 mMMgCl2, 1 mM dithiothreitol) and then were incubated with 1.5 �g of MBP or His-tagged BAD recombinant proteins in the kinase buffer supplemented with 20 �MATP and 5 �Ci of [�-32P]ATP for 30 min at 30°C. The reaction was terminatedwith SDS-PAGE sample buffer, followed by SDS-PAGE and autoradiography.
Apoptosis assays. To estimate apoptosis of stably transfected cell lines, equalnumbers of cells were seeded in six-well plates. Cells were treated 16 h with CA(10 �M) or C2C (50 �M), and collected for flow cytometry or used to preparelysates. Cells were collected by trypsinization and, after combination with thefloating cells, were labeled with annexin V and propidium iodide according to themanufacturer’s recommendations (Clontech) for fluorescence-activated cell sort-ing analysis (Becton Dickinson). When the PKA inhibitor (PKI) was used, cellswere pretreated 30 min with 1 �M before inducers were added for 16 h.
Cellular fractionation. CHO cell lines were transfected with pcDNA-HA-BAD and after 24 h were treated for 16 h with CA (10 �M) or C2C (50 �M).Cellular fractionation was then performed as previously described (14). Briefly,cells were trypsinized and resuspended in isotonic mitochondrial buffer (MB)
VOL. 23, 2003 Pak5 LOCALIZES TO MITOCHONDRIA 5527
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

(210 mM mannitol, 70 mM sucrose, 1 mM EDTA, 10 mM HEPES [pH 7.5]supplemented with proteases inhibitors). Cells were broken by passages througha 25-gauge needle fitted on a syringe, and the suspension was centrifuged at 2,000� g at 4°C. Supernatants containing heavy membranes were subjected to a 13,000� g centrifugation at 4°C, the supernatant corresponding to the cytosolic fractionwas centrifuged at 100,000 � g for 30 min, and the pellet containing mitochon-dria was resuspended in MB and submitted to a 500 � g centrifugation. Theresultant supernatant was centrifuged 10 min at 10,000 � g to collect the mito-chondrial fraction. A further purification step was performed for Pak’s subcel-lular localization (24). The latter mitochondrial fraction was resuspended in MBand applied on a 1-to-2 M linear sucrose gradient and centrifuged 30 min at60,000 � g using an SW28 Beckman rotor. The mitochondria, which banded ata region corresponding to 1.5 M were collected and diluted in MB to 0.25 M andpelleted at 15,000 � g for 15 min. The mitochondrial pellets were resuspendedin MB. The S100 cytosolic fraction also generated the light microsomal pelletafter centrifugation, which was resuspended in buffer H (20 mM Tris-HCl, 1 mMEDTA, 1 mM EGTA, 6 mM -mercaptoethanol [pH 7.5], 1% SDS).
Immunofluorescence. HMN1 cells seeded on coverslips were transiently trans-fected with empty vector or vectors encoding Myc-tagged WT Pak5 or KD Pak5(K478M). HMN1 and CHO cells were stained 15 min at 37°C with a 5-ng/mlconcentration of the mitochondrion-specific dye MitoTracker Red CMXRos(Molecular Probes Inc.) 24 h after transfection and then fixed in 4% parafor-maldehyde. Cells were fixed, permeabilized in PBS–0.1% NP-40, and blocked 30min with 10% bovine serum albumin. Cells were stained for the presence of Pak5with rabbit polyclonal anti-Myc antibody (A14) and incubated with anti-rabbitimmunoglobulin G conjugated with fluorescein isothiocyanate (FITC) (JacksonLaboratories). Cells were then washed, mounted and photomicrographs wereobtained using a Bio-Rad radiant 2000 laser scanning confocal microscope witha Plan APO 60� water immersion objective (1.20 numerical aperture; NikonE800 Eclipse).
RESULTS
Complementation analysis of Pak5. Group A mammalianPaks are able to replace the function of Ste20p in S. cerevisiae(3, 4). The mammalian group B Paks, though clearly distinct instructure and biochemical properties from group A Paks (seebelow), are nevertheless as closely related as group A Paks toSte20p. It was therefore of interest to determine if group BPaks can functionally substitute for Ste20p. We transformed ahaploid STE20-null strain of S. cerevisiae with empty vector orvectors designed for inducible expression of human Pak2 orPak5 alleles. Immunoblot analysis revealed that the transgenesare expressed when cells are grown in galactose medium (Fig.1A). We then carried out immunocomplex-kinase assays toassess the activity of the Pak transgenes. As expected, Pak5S573N was far more active than either WT Pak5 or Pak2 (Fig.1B), and expression of this hyperactivated allele was cytostatic(Fig. 1C). This effect is similar to the effect of constitutivelyactive alleles of S. pombe Pak1 when expressed in S. cerevisiae(33, 39). A mating assay was then carried out to determinecomplementation (Fig. 1D). WT and STE20-null cells express-ing Pak2 are competent for mating, whereas cells expressing
any of the Pak5 alleles are not. These results indicate that thefunctions of human Pak2 and Pak5 are distinct.
Biochemical properties of Pak5. Sequence analysis indicatesthat Pak5 has a putative GBD/CRIB motif similar to that ofPak4. To determine whether Pak5 interacts with the GTPases,a pull-down assay was performed in which lysates from Coscells transfected with empty vector or a Myc-Pak5 or a Myc-Pak1 expression vectors were incubated with GST-Cdc42 L61(constitutively active) or -N17 (dominant-negative) or GST-Rac1 L61 or -N17. As shown in Fig. 2A, Pak5 interacts withboth Cdc42 and Rac1, suggesting that Pak5 is a target for theseGTPases. However, contrary to Pak1, but similar to Pak4 (1),Pak5 binds more strongly to Cdc42 than to Rac1.
To confirm these results in cells, COS cells were transfectedwith Pak5 or Pak1 alone or in combination with Cdc42 L61 orRac L61 and immunoprecipitations were performed on thelysates (Fig. 2B). As in the pull-down assay, the coprecipitationexperiments show that Pak5 binds to both GTPases, but with amarked preference for Cdc42. These results are consistent withthose reported by Dan et al. (7).
The group A Paks requires binding of GTPases to be acti-vated whereas Pak4 has been shown in vitro to autophosphor-ylate and to phosphorylate Histone H4 (5). We assessed Pak5kinase activity using MBP as a substrate (Fig. 2C). COS cellswere transfected with Myc-tagged Pak5 expression vector orMyc-tagged Pak1, used as a positive control. GTPases expres-sion vectors were also cotransfected. Equal amounts of Pak5and Pak1 were immunopurified from cell lysates and incubatedwith MBP and [�-32P]ATP in kinase buffer. Autophosphor-ylation of Pak5 and transphosphorylation of MBP in Fig. 2Cshows that Pak5 has a strong autophosphorylation activitycompared to Pak1 and phosphorylates MBP in the absenceof GTPases. On the contrary, Pak1 requires activation byGTPases in order to phosphorylate MBP, and does so atorders of magnitude less than Pak5 (Fig. 2C, inset).
Pak5 induces resistance to apoptosis induced by ceramideor CA. As many of the Paks have previously been shown toeither induce (Pak2, once cleaved by caspase 3) (26, 31, 40) orinhibit (Pak1 and Pak4) (18, 32, 38) apoptosis, we sought todetermine if Pak5 also plays a role in this process. In order toexamine the role of Pak5 in cell survival, CHO stable cell lineswere generated expressing either WT Pak5 or KD Pak5(K478M) (8) or empty vector (control). At least three inde-pendent stable clones were analyzed for each construct. Rep-resentative data for protein expression and kinase activity dataare shown (Fig. 3A). Consistent with the data from transiently
FIG. 1. Complementation analysis of Pak5. ste20-null or a control strain of S. cerevisiae was transformed with a galactose-inducible expressionvector bearing either no insert or a cDNA encoding Pak2, Myc-tagged WT Pak5, hyperactivated mutant (S573N), or K478 M (KD) mutant.Transformants were selected for growth in YNB URA� raffinose medium, galactose was added to 2% at time zero, and the cultures were inducedfor 5 h before harvesting. (A) Expression of the transgenes. Cells were lysed before and 5 h after induction, then assayed for protein expressionby immunoblotting with monoclonal anti-Myc 9E10 or anti-Pak2. Equal protein levels was assessed using an antitubulin antibody. (B) Kinase assay.Cultures tested in panel A and obtained after 5 h of induction were lysed and immunoprecipitated with a polyclonal anti-Myc antibody orpolyclonal anti-Pak2 and assayed for kinase activity as described in Materials and Methods. (C) Transformants were streaked onto YNB glucose,YNB raffinose, and YNB galactose URA� plates and incubated at 30°C before being photographed. (D) To perform semiquantitative matingassays, the transformants were serially diluted onto YNB raffinose URA� plates. After the colonies had grown sufficiently, the plates werereplicated onto YNB Gal URA� for 5 h to allow expression of the transgenes. The cells were then replica-plated onto YEPD plates containinga lawn of the mating tester strain, and allowed to mate for six hours before being replicated onto GN plates. Mating efficiency was scored by theformation of prototrophic strains on the GN plates.
5528 COTTERET ET AL. MOL. CELL. BIOL.
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

VOL. 23, 2003 Pak5 LOCALIZES TO MITOCHONDRIA 5529
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

FIG. 2. Biochemical properties of Pak5. (A) COS cells were transfected with equal amounts of expression vectors encoding WT Myc-taggedPak1 and Pak5 or empty vector. After transient expression, Myc-Pak1 and Myc-Pak5 were pulled down from whole-cell lysate (WCL) with GST-Cdc42 and GST-Rac loaded with GTP, Western blotting was then performed using an anti-Myc antibody. The expression level of Pak1 and Pak5in the lysates used for the pull-down is shown in the bottom panel, expression was detected by Western blotting using an anti-Myc antibody. (B) COScells were cotransfected with equal amounts of expression vectors encoding HA-tagged Cdc42 L61 or HA-Rac L61 or Myc-tagged Pak1 or -Pak5alone or plus HA-tagged Cdc42 L61 or HA-Rac L61. After transient expression, Myc-Pak1 and Myc-Pak5 were immunoprecipitated from totallysates with an anti-Myc antibody and protein A-agarose. Western blotting was then performed and the complexes were revealed with an anti-HAantibody. The bottom panel shows the expression of Pak1 and Pak5, revealed with an anti-Myc and Cdc42 and Rac, revealed with an anti-HA inthe WCL. (C) COS cells were transfected with expression vectors for Myc-tagged Pak1 or -Pak5 or cotransfected with Pak1 or Pak5 cDNAs andthe HA-tagged GTPases Cdc42 and Rac. Myc-tagged Pak1 and Pak5 were immunoprecipitated with an anti-Myc antibody and the kinase assaywas performed as described in Materials and Methods using [�-32P]ATP and MBP as a substrate. Reaction products were separated withSDS-PAGE and subjected to autoradiography. The bottom panel shows the expression levels of proteins in the WCL prior to immunoprecipitation.
5530 COTTERET ET AL. MOL. CELL. BIOL.
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

FIG. 3. Pak5 induces resistance to apoptosis. (A) Expression and activity of Pak5 in CHO stable cell lines. Stable CHO cell lines wereestablished to study apoptosis. Equal amounts of lysates from stable CHO cells containing empty vector, Myc-WT Pak5, or Myc-KD Pak5 wereanalyzed for the expression of Pak5 proteins by Western blotting using anti-Myc antibody. Kinase activity was assessed by performing a kinaseassay. Myc-tagged proteins were immunoprecipitated with anti-Myc and using MBP as a substrate. (B) Stable cell lines expressing Pak5 areresistant to apoptosis. Cells were treated with C2C (50 �M) and CA (10 �M) for 16 h and were collected and stained with annexin V and propidiumiodide. Apoptosis was assessed by flow cytometry. The percentages of total cells positive for annexin V are shown on the graph, and represent theaverage of three experiments. (C and D) Caspase 3 and PARP cleavage. Cells were treated as indicated above with C2C and CA. Cells were lysedand Western blotting was performed using anti-caspase 3 and anti-PARP antibodies. *, nonspecific band.
5531
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

FIG. 4. Pak5 phosphorylates BAD on serine 112 in vitro. (A) COS cells were transfected with empty vector or Myc-Pak5 or HA-Pak4. An equalamount of cell extracts were used for in vitro kinase assay. Myc-tagged proteins were immunoprecipitated with anti-Myc, the immunocomplexeswere incubated in the presence of [�-32P]ATP and equal amounts of recombinant His-tagged BAD (WT, S112A, S136A, and S112A/S136A) assubstrates as shown on the bottom panel. After SDS-PAGE, phosphorylated BAD was detected by autoradiography. (B) The kinase assay wasperformed using His-tagged WT BAD, and immunocomplexes from lysates of COS cells transfected with Myc-Pak5, HA-Pak4 and HA-Akt(activated mutant, del AA6-10). After Western blotting, phosphorylated BAD was detected with anti-phospho-Ser 112 and anti-phospho-Ser 136,while total BAD was detected with an anti-BAD antibody. Bottom panel shows the expression of the different proteins in the whole-cell lysate(WCL). * shows cross-reaction of anti-phospho-Ser 136 BAD with anti-HA immunoglobulin G light chain.
5532
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

transfected COS cells, WT Pak5 in CHO cells exhibits consid-erable basal activity, whereas the K478 M mutant is inactive.
To induce apoptosis, CHO cells were treated with two in-ducers: CA, a topoisomerase I inhibitor, and C2C, a well-knownanalog to a second messenger released from plasma membraneduring apoptosis. Cell survival was assessed with annexin Vstaining after a 16-h treatment with CA or C2C. As shown inFig. 3B, CA and C2C induce 35 and 50% apoptosis, respec-tively, in control cells, whereas no apoptosis is detectable inWT Pak5 CHO cells. In contrast, expression of KD Pak5 doesnot protect against apoptosis, and we observe a percentage ofapoptotic cells that is similar to the CHO cells when cells aretreated with CA, and to a lesser extent with C2C. These datashow that Pak5 kinase activity is responsible for the resistanceto apoptosis.
We examined PARP and caspase 3 cleavage, two biochem-ical markers of apoptosis, in the stable cell lines. After a 16-htreatment with C2C and CA, PARP and caspase 3 were cleavedin control and KD Pak5 cell lines (Fig. 3C and D), whereas WTPak5-expressing cell lines, which are resistant to apoptosis,retain full-length PARP and caspase 3 in both untreated andinduced cells. This result indicates that the apoptotic signalleading to the release of cytochrome c from the mitochon-drion, subsequently activating caspase 9 and caspase 3, isblocked downstream of the mitochondrion when Pak5 is ex-pressed.
Pak5 phosphorylates BAD. Activated Pak1 and Pak4 havebeen shown to phosphorylate BAD on Ser-112 and Ser-136and Ser-112, respectively (18, 32). We addressed whether BADis a target for Pak5, BAD being a key proapoptotic molecule.To analyze BAD phosphorylation by Pak5, COS cells weretransfected with empty vector, WT Pak5, KD Pak5 or WT Pak4as a positive control. In vitro kinase assay was carried out inwhich His-tagged recombinant WT BAD, S112A or S136Amutants or double mutant S112A/S136A proteins were used assubstrates for immunopurified Pak proteins (Fig. 4A). These
results indicate that WT Pak5 and Pak4 cannot phosphorylateBAD S112A mutant or the double mutant S112A/S136A, where-as they are able to phosphorylate WT BAD or the S136A mu-tant. Therefore, Pak5 phosphorylates BAD mainly on Ser-112,although BAD mutation on Ser-112 does not completely abol-ish its phosphorylation, indicating that Pak5 could target to alesser extent additional residues. In order to confirm this re-sult, we used phosphospecific antibodies directed against Ser-112 or Ser-136 to determine the status of the 112 and 136 sitesin an in vitro non radioactive kinase assay (Fig. 4B). These datashow that Pak5 phosphorylates BAD on Ser-112, in agreementwith the radioactive kinase activity assay.
Pak5 prevents BAD dephosphorylation during apoptosis.As BAD is phosphorylated by Pak5 in vitro, we determinedwhether Pak5 expression causes sustained BAD phosphoryla-tion in cells treated with apoptotic agents. CHO cell lines weretransfected with a vector encoding a GST-BAD protein. After a16-h treatment with CA (A) or C2C (B), a GST pull down wasperformed on lysates to retrieve GST-BAD. The expressionlevel of BAD was assessed by Western blotting with an anti-GST antibody and BAD phosphorylation status was assessedby Western blotting using phosphospecific antibodies. As shownin Fig. 5A and B, in treated cells, BAD is dephosphorylated onSer-112 in control cells as well as in cells expressing KD Pak5,whereas no dephosphorylation on Ser-112 is observed in cellsexpressing WT Pak5. Notably, BAD Ser-136 is also dephosphor-ylated in control and KD Pak5 treated cells, but not in WT Pak5cells. Because Pak5 does not phosphorylate BAD at Ser-136 invitro (Fig. 4), these results suggest that Pak5 influences BADphosphorylation by both direct and indirect mechanisms.
Analysis of Pak5-mediated BAD phosphorylation in cells.Although Pak5 phosphorylates BAD at Ser-112 in vitro, andmaintains BAD phosphorylation in cells treated with apoptoticagents, it is possible that Pak5 acts indirectly by activatingother kinases that phosphorylate BAD. As PKA has beenreported to catalyze BAD phosphorylation at Ser-112 (21), we
FIG. 5. Pak5 prevents BAD dephosphorylation in vivo. CHO control cells and stable cell lines expressing WT Pak5 or KD Pak5 weretransfected with pEBG-BAD and were treated with CA (10 �M) or C2C (50 �M) 24 h after transfection. Equal amounts of cell extracts wereanalyzed by Western blotting after pull-down assay of GST-BAD, using an anti-GST antibody to assess BAD expression level, and antibodiesdirected against phospho-Ser 112 and phospho-Ser 136 to assess BAD phosphorylation status.
VOL. 23, 2003 Pak5 LOCALIZES TO MITOCHONDRIA 5533
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

sought to determine if this kinase is required for Pak5-medi-ated BAD phosphorylation in cells. Control and Pak5-express-ing CHO cells were pretreated with a PKI and then treatedwith CA, and BAD phosphorylation on Ser-112 was assessedwith a phospho-specific antibody. As shown in Fig. 6A, incontrol cells, CA and PKI both induce BAD dephosphor-
ylation at Ser-112. In contrast, basal levels of BAD Ser-112phosphorylation are elevated in cells expressing Pak5, andneither CA nor PKI reduces Ser-112 phosphorylation. Identi-cal results were obtained using ceramide as the apoptotic stim-ulus (data not shown). These results show that Pak5 does notact through PKA to phosphorylate BAD at Ser-112.
We also assessed the role of Pak5 on Akt phosphorylation,since Akt has been reported to phosphorylate BAD at Ser-136(11, 12). Control cells, or cells expressing WT or KD Pak5 weretreated with vehicle, CA or C2C, and Akt activity was mea-sured by assessing phosphorylation on Akt Ser-473. Figure 6Bshows that both apoptotic inducers reduce Akt activity in CHOand KD Pak5-expressing cells. However, expression of WTPak5 prevents Akt dephosphorylation. These results suggestthat the Pak5-mediated phosphorylation of BAD at Ser-136may be mediated through Akt.
Pak5 prevents BAD translocation to mitochondria. Undernormal condition, phosphorylated BAD is sequestered by 14-3-3� in the cytosol (42). In response to apoptotic stimuli, BADis dephosphorylated and translocates to the mitochondrialmembrane where it binds to the antiapoptotic proteins BCL-2and BCL-xL (18). Having demonstrated that Pak5 phosphory-lates BAD on serine 112 and indirectly on serine 136, wedetermined if this activity of Pak5 was sufficient to maintainBAD in the cytosol and particularly if it prevents BAD trans-location to mitochondria, and therefore the apoptotic cascade.We transfected control, WT Pak5, and KD Pak5 stable celllines with a pcDNA-BAD plasmid, and treated cells with in-ducers, CA or C2C, in order to induce apoptosis, and subse-quent BAD translocation to mitochondria. We found that BADtranslocates to mitochondria in both control and KD Pak5 CHOcells, as shown by the presence of BAD in the mitochondrialfraction, whether apoptosis is induced by CA (Fig. 7A) or C2C
FIG. 6. Pak5 phosphorylates BAD independently of PKA and pre-vents Akt dephosphorylation on serine 473. (A) Stable cell lines weretransfected with pEBG-BAD and after 24 h were pretreated 30 minwith a specific myristoylated protein kinase A inhibitor (1 �M) beforeCA (10 �M) was added for 16 h. GST-BAD was then retrieved fromthe lysates and an anti-phospho-Ser-112 BAD antibody was used forthe Western blot to assess BAD phosphorylation status. (B) Stable celllines were treated for 16 h with CA (10 �M) or C2C (50 �M). Afterlysis, Western blotting was performed using an anti-phospho-Ser 473specific antibody.
FIG. 7. Pak5 prevents BAD translocation to mitochondria. CHO control cells and stable cell lines expressing WT Pak5 and KD Pak5 weretransfected with HA-BAD and 24 h later treated with CA or C2C for 16 h. Cells were collected by trypsinization and a cellular fractionation wasperformed to isolate mitochondria as described in Materials and Methods. Equal amount of proteins were separated by SDS-PAGE. BAD wasdetected by Western blotting using an anti-HA antibody. Equal loading of the mitochondrial pellet was assessed with an anti-COX IV antibody.
5534 COTTERET ET AL. MOL. CELL. BIOL.
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

FIG. 8. Pak5 is present in the mitochondrial fraction. A cellular fractionation was performed on CHO stable cell lines as described in Materialsand Methods to assess Pak5 presence in the cellular compartments. (A) Equal amount of proteins from cytosolic (Cy) S100, light microsomes (LM)and mitochondrial (M) fractions were loaded on a gel, and Western blotting was performed using anti-Myc antibody. The mitochondrial fractionpurity was assessed using an anti-COX IV and anticalnexin antibodies. The fractions were also subjected to immunoblotting with anti-BAD,anti-BCL-2 and anti-BCL-xL antibodies. (B) CHO cells expressing vector alone, WT Pak5, or KD Pak5 were treated with CA (10 �M) or C2C (50�M) for 16 h. A cellular fractionation was then performed. Equal amount of proteins from cytosolic (Cy) S100, light microsomes (LM) andmitochondrial (M) fractions were loaded on a gel, and Western blotting was performed using anti-Myc antibody. The mitochondrial fraction puritywas assessed using an anti-COX IV and anticalnexin antibodies. (C) CHO cells were transfected with pCMV6, HA-Pak4 or HA-Pak6, and asubcellular fractionation was performed 48 h after transfection as described above.
VOL. 23, 2003 Pak5 LOCALIZES TO MITOCHONDRIA 5535
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

(Fig. 7B). In contrast, no translocation occurs in the WT Pak5cell lines, as suggested by the absence of BAD in the mito-chondrial fraction of these cells. This is consistent with thephosphorylation of serine 112 by Pak5 and shows that Pak5, byits phosphorylation activity, can prevent BAD translocation tomitochondria.
Pak5 localizes to mitochondria. As Pak5 stimulates BADphosphorylation and prevents its translocation to mitochon-
dria, we investigated whether Pak5 acts in the cytosol or onmitochondria. We assessed Pak5 localization by performingcellular fractionation. As shown in Fig. 8A, we found that bothWT and KD Pak5 are found exclusively in the mitochondrialfraction. Immunoblots with specific marker antibodies (COXIV for mitochondria, calnexin for endoplasmic reticulum/lightmicrosomes) show that the subcellular fractions are reasonablypure. Nuclear fractions are not shown because they are con-
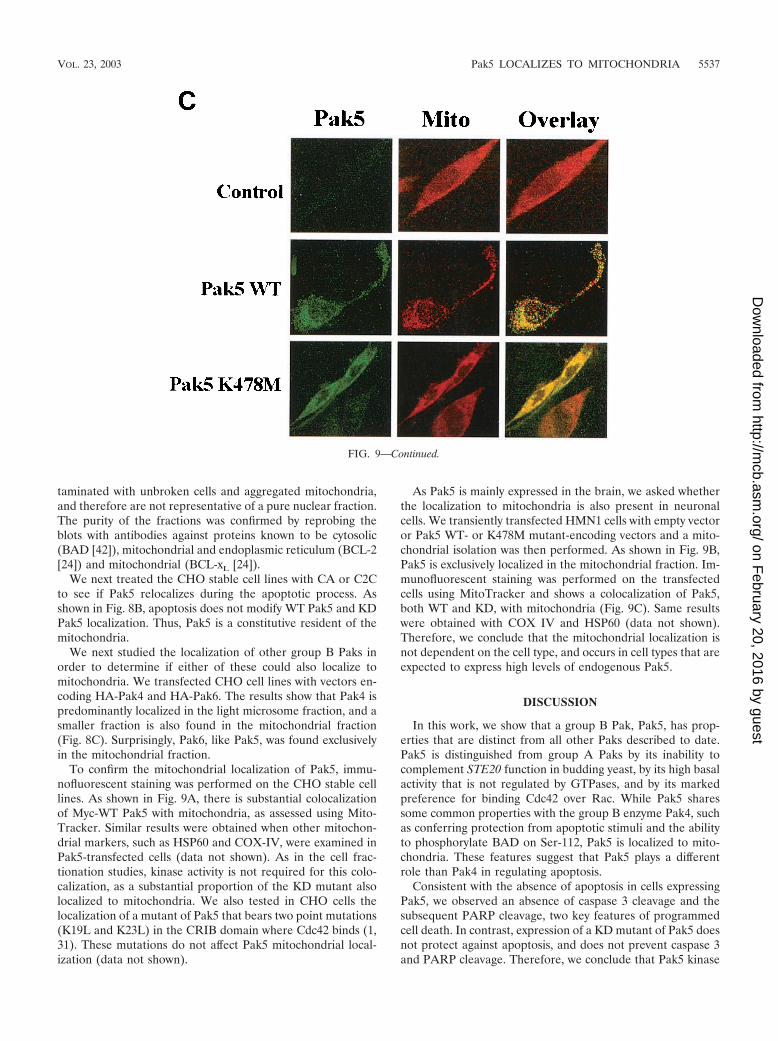
FIG. 9. Pak5 colocalizes with mitochondria. CHO stable cell lines (A) expressing WT Pak5 or KD Pak5 were labeled as described in Materialsand Methods. The fields shown were analyzed independently by digital confocal fluorescence microscopy at the appropriate wavelength for FITC(Pak5) and MitoTracker Red CMXRos (Mito), and the two images were overlaid (Overlay). (B) HMN1 cells were transfected with the emptyvector (control) or with a vector carrying WT Pak5 or Pak5 K478M cDNA. A cellular fractionation was then performed 48 h after transfectionsas described in Materials and Methods to assess Pak5 presence in the cellular compartments. Equal amount of proteins from cytosolic (Cy) S100,light microsomes (LM) and mitochondrial (M) fractions were loaded on a gel, and Western blotting was performed using anti-Myc antibody. Themitochondrial fraction was assessed using anti-COX IV and anticalnexin antibodies. (C) HMN1 cells were transfected with empty vector or Myc-tagged WT Pak5 or KD Pak5 were stained as described in Materials and Methods. The fields shown were analyzed independently by fluorescencemicroscopy at the appropriate wavelength for FITC (Pak5) and MitoTracker Red CMXRos (Mito), and the two images were overlaid (Overlay).
5536 COTTERET ET AL. MOL. CELL. BIOL.
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from
taminated with unbroken cells and aggregated mitochondria,and therefore are not representative of a pure nuclear fraction.The purity of the fractions was confirmed by reprobing theblots with antibodies against proteins known to be cytosolic(BAD [42]), mitochondrial and endoplasmic reticulum (BCL-2[24]) and mitochondrial (BCL-xL [24]).
We next treated the CHO stable cell lines with CA or C2Cto see if Pak5 relocalizes during the apoptotic process. Asshown in Fig. 8B, apoptosis does not modify WT Pak5 and KDPak5 localization. Thus, Pak5 is a constitutive resident of themitochondria.
We next studied the localization of other group B Paks inorder to determine if either of these could also localize tomitochondria. We transfected CHO cell lines with vectors en-coding HA-Pak4 and HA-Pak6. The results show that Pak4 ispredominantly localized in the light microsome fraction, and asmaller fraction is also found in the mitochondrial fraction(Fig. 8C). Surprisingly, Pak6, like Pak5, was found exclusivelyin the mitochondrial fraction.
To confirm the mitochondrial localization of Pak5, immu-nofluorescent staining was performed on the CHO stable celllines. As shown in Fig. 9A, there is substantial colocalizationof Myc-WT Pak5 with mitochondria, as assessed using Mito-Tracker. Similar results were obtained when other mitochon-drial markers, such as HSP60 and COX-IV, were examined inPak5-transfected cells (data not shown). As in the cell frac-tionation studies, kinase activity is not required for this colo-calization, as a substantial proportion of the KD mutant alsolocalized to mitochondria. We also tested in CHO cells thelocalization of a mutant of Pak5 that bears two point mutations(K19L and K23L) in the CRIB domain where Cdc42 binds (1,31). These mutations do not affect Pak5 mitochondrial local-ization (data not shown).
As Pak5 is mainly expressed in the brain, we asked whetherthe localization to mitochondria is also present in neuronalcells. We transiently transfected HMN1 cells with empty vectoror Pak5 WT- or K478M mutant-encoding vectors and a mito-chondrial isolation was then performed. As shown in Fig. 9B,Pak5 is exclusively localized in the mitochondrial fraction. Im-munofluorescent staining was performed on the transfectedcells using MitoTracker and shows a colocalization of Pak5,both WT and KD, with mitochondria (Fig. 9C). Same resultswere obtained with COX IV and HSP60 (data not shown).Therefore, we conclude that the mitochondrial localization isnot dependent on the cell type, and occurs in cell types that areexpected to express high levels of endogenous Pak5.
DISCUSSION
In this work, we show that a group B Pak, Pak5, has prop-erties that are distinct from all other Paks described to date.Pak5 is distinguished from group A Paks by its inability tocomplement STE20 function in budding yeast, by its high basalactivity that is not regulated by GTPases, and by its markedpreference for binding Cdc42 over Rac. While Pak5 sharessome common properties with the group B enzyme Pak4, suchas conferring protection from apoptotic stimuli and the abilityto phosphorylate BAD on Ser-112, Pak5 is localized to mito-chondria. These features suggest that Pak5 plays a differentrole than Pak4 in regulating apoptosis.
Consistent with the absence of apoptosis in cells expressingPak5, we observed an absence of caspase 3 cleavage and thesubsequent PARP cleavage, two key features of programmedcell death. In contrast, expression of a KD mutant of Pak5 doesnot protect against apoptosis, and does not prevent caspase 3and PARP cleavage. Therefore, we conclude that Pak5 kinase
FIG. 9—Continued.
VOL. 23, 2003 Pak5 LOCALIZES TO MITOCHONDRIA 5537
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

activity is responsible for resistance to apoptosis. Interestingly,activation of the group A enzyme, Pak2, is associated withproapoptotic rather than antiapoptotic effects. One key differ-ence between proapoptotic kinases such as Pak2 and antiapo-ptotic kinases such as Pak1, Pak4, and Pak5 is that the formercontains a caspase site and is sensitive to cleavage by caspase 3(26, 31), whereas the latter lack such sites and are not knownto be cleaved. Possibly, the generation of a separated kinasedomain, such as occurs following Pak2 cleavage, results inrelocalization and acquisition of new substrates that contributeto cell death. Therefore, we suspect that the N terminal portionof Pak5 regulates the localization and substrate specificity ofthis kinase, and is required for its pro-survival effects.
One of the mechanisms by which Pak5 induces resistance toapoptosis is the phosphorylation of BAD. We show that Pak5phosphorylates BAD in vitro and in vivo, as it has also beenobserved for Pak1 and Pak4 (18, 31, 32, 38). These phosphor-ylations occur at Ser-112, and perhaps, to a lesser extent, onadditional Ser residues. While Pak5 may have additional tar-gets that are relevant to its antiapoptotic effects, the phosphor-ylation of BAD is likely to be germane in this setting. AlthoughPak5 can phosphorylate BAD in vitro and induce BAD phos-phorylation in cells, there are several other kinases that alsoexhibit these properties. To date, PKA has been identified as amajor BAD Ser-112 kinase, while Akt has been reported to bea major Ser-136 kinase. Our data suggest that the Pak5-stim-ulated phosphorylation of BAD at Ser-112 is direct, as Pak5acts on this site in vitro and inhibitors of PKA do not affect theability of Pak5 to maintain BAD Ser-112 phosphorylation incells treated with inducers of apoptosis. In contrast, the Pak5-stimulated phosphorylation of BAD at Ser-136 may be medi-ated through Akt, as Pak5 expression is associated with ele-vated activity of Akt. These results suggest that Pak5 and PKAare independent mediators of BAD phosphorylation (Fig. 10).They also suggest that Pak5 signaling differs from Pak1 whichappears to act downstream of Akt (38).
The phosphorylation of BAD by Pak5 is likely to occur at themitochondrion, as Pak5 appears to be a constitutive resident ofthis organelle, and BAD translocates to mitochondria follow-ing apoptotic stimuli. The predominantly mitochondrial local-ization of Pak5 is independent of cell type, as we were able tosee that same colocalization of Pak5 in CHO, HMN1 and NIH-3T3 cells (data not shown). Pak5 is not recruited to mitochon-dria specifically during apoptosis, and its residence there isindependent of kinase activity or its ability to bind Cdc42. Inthis respect, Pak5 resembles the mitochondrion-anchored PKA,although we do not know if Pak5 is anchored to the mitochon-drion directly or bound to a mitochondrion-anchored protein.
To date, localization of other group B Pak isoforms has notbeen studied in detail; therefore, we also assessed Pak4 andPak6 localization. Surprisingly, we found that Pak6, and, to alesser extent, Pak4, also localized to mitochondria. The bulk ofPak4, however, is detected in the light microsome fraction (Fig.8C), in agreement with the findings of Abo et al. (1), whoshowed that Pak4 translocates to Golgi apparatus when coex-pressed with Cdc42. Pak6 was previously shown to be cytosolicand to shuttle to the nucleus when coexpressed with androgenand estrogen receptors (27), but has not been previously re-ported to localize to mitochondria. Our data suggest that mito-chondrial localization is a common feature of group B Paks.
Mammalian group B Paks appear to localize to many dis-tinct cellular compartments, unlike group A Paks such as Pak1,which are found mostly in areas of active actin dynamics, suchas leading edge lamellipodia (37) or the group B X-Pak5 fromXenopus, which has been found in association with microtu-bules (5). The mammalian group B Paks are in general lesssimilar to one another than are the group A Paks, and it islikely that this diverged sequence reflects diverged cellularfunction. While we do not yet know what domain on Pak5targets the mitochondrion, the N terminus is probably in-volved. Paks 4, 5, and 6 differ substantially in this noncatalyticN-terminal domain, whereas the C-terminal kinase domainsare very similar. Moreover preliminary data from our labshows that the isolated N terminus of Pak5 localizes to mito-chondria. Whether the Cdc42 binding domain, which is locatedat the extreme N terminus of the protein, is involved is cur-rently unknown but appears unlikely as Cdc42 is not known tolocalize to this organelle.
Pak5 expression is restricted to the brain and overexpressionof this kinase has been shown to induce neurite outgrowth inN1E-115 cells (8). It is likely that such kinase activity is tightlycontrolled in central nervous system, as 95% of neurons un-dergo apoptosis during embryogenesis. How is this kinase reg-ulated? Pak5 has unusually high basal activity compared toother group A or group B Paks. While Pak5 binds Cdc42, thisinteraction has no effect on kinase activity in vitro. This findingsuggests that the binding to Cdc42 could have a role other thandirect activation. However, it is also possible that there areendogenous inhibitors of Pak5 that regulate its activity underphysiological conditions. Provided that adequate sera can be
FIG. 10. Model of Pak5 in cell survival signaling.Pak5 is localizedto mitochondria, either directly or via interactions with a tetheringprotein. There, it encounters BAD, which it phosphorylates at Ser-112.The PKA complex (comprising a catalytic subunit, a regulatory subunit[RII], and a mitochondrial-tethering subunit [AKAP]) independentlytargets the Ser-112 site. Pak5 also induces, through an unknown mech-anism, the activation of Akt, which phosphorylates BAD at Ser-136.Phosphorylated BAD complexes with 14-3-3� in the cytosol.
5538 COTTERET ET AL. MOL. CELL. BIOL.
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

raised to the endogenous protein, we plan to examine theseand related issues in the near future.
ACKNOWLEDGMENTS
We thank Audrey Minden, Michael Greenberg, Randall Pittman,and Randy Strich for their generous donation of reagents and EdnaCukierman and Jonathan Boyd for assistance in imaging.
This work was supported by grants from the National Institutes ofHealth (RO1 GM-54168 and CA-09035-27) and the American CancerSociety (CB-189), as well as CORE Grant CA-06927 and an appro-priation from the Commonwealth of Pennsylvania.
REFERENCES
1. Abo, A., J. Qu, M. S. Cammarano, C. Dan, A. Fritsch, V. Baud, B. Belisle,and A. Minden. 1998. PAK4, a novel effector for Cdc42Hs, is implicated inthe reorganization of the actin cytoskeleton and in the formation of filopo-dia. EMBO J. 17:6527–6540.
2. Bagrodia, S., and R. A. Cerione. 1999. PAK to the future. Trends Cell Biol.9:350–355.
3. Bagrodia, S., S. Taylor, C. L. Creasy, J. Chernoff, and R. A. Cerione. 1995.Identification of a murine p21Cdc42/Rac activated protein kinase (PAK).J. Biol. Chem. 270:22731–22738.
4. Brown, J. L., L. Stowers, M. Maer, J. Trejo, S. Coughlin, and J. Chant. 1996.Human Ste20 homologue hPak1 links GTPases to the JNK MAP kinasepathway. Curr. Biol. 6:598–605.
5. Cau, J., S. Faure, M. Comps, C. Delsert, and N. Morin. 2001. A novelp21-activated kinase binds the actin and microtubule networks and inducesmicrotubule stabilization. J. Cell Biol. 155:1029–1042.
6. Chong, C., L. Tan, L. Lim, and E. Manser. 2001. The mechanism of PAKactivation. Autophosphorylation events in both regulatory and kinase do-mains control activity. J. Biol. Chem. 276:17347–17353.
7. Dan, C., A. Kelly, O. Bernard, and A. Minden. 2001. Cytoskeletal changesregulated by the PAK4 serine/threonine kinase are mediated by LIM kinase1 and cofilin. J. Biol. Chem. 276:32115–32121.
8. Dan, C., N. Nath, M. Liberto, and A. Minden. 2002. PAK5, a new brain-specific kinase, promotes neurite outgrowth in N1E-115 cells. Mol. Cell.Biol. 22:567–577.
9. Dan, I., N. M. Watanabe, and A. Kusumi. 2001. The Ste20 group kinases asregulators of MAP kinase cascades. Trends Cell Biol. 11:220–230.
10. Daniels, R. H., and G. M. Bokoch. 1999. p21-activated protein kinase: acrucial component of morphological signaling? Trends Biochem. 24:350–355.
11. Datta, S. R., H. Dudek, X. Tao, S. Masters, H. Fu, Y. Gotoh, and M. E.Greenberg. 1997. Akt phosphorylation of BAD couples survival signals to thecell-intrinsic death machinery. Cell 91:231–241.
12. del Peso, L., M. Gonzalez-Garcia, C. Page, R. Herrera, and G. Nunez. 1997.Interleukin-3-induced phosphorylation of BAD through the protein kinaseAkt. Science 278:687–689.
13. Edwards, D. C., L. C. Sanders, G. M. Bokoch, and G. N. Gill. 1999. Activa-tion of LIM-kinase by Pak1 couples Rac/Cdc42 GTPase signalling to actincytoskeletal dynamics. Nat. Cell Biol. 1:253–259.
14. Eskes, R., S. Desagher, B. Antonsson, and J. C. Martinou. 2000. Bid inducesthe oligomerization and insertion of Bax into the outer mitochondrial mem-brane. Mol. Cell. Biol. 20:929–935.
15. Frost, J. A., A. Khokhlatchev, S. Stippec, M. A. White, and M. H. Cobb. 1998.Differential effects of PAK1-activating mutations reveal activity-dependentand -independent effects on cytoskeletal regulation. J. Biol. Chem. 273:28191–28198.
16. Gajewski, T. F., and C. B. Thompson. 1996. On a BAD influence. Cell87:589–592.
17. Gietz, D., A. St. Jean, R. A. Woods, and R. H. Schiestl. 1992. Improvedmethod for high efficiency transformation of intact yeast cells. Nucleic AcidsRes. 20:1425.
18. Gnesutta, N., J. Qu, and A. Minden. 2001. The serine/threonine kinasePAK4 prevents caspase activation and protects cells from apoptosis. J. Biol.Chem. 276:14414–14419.
19. Golstein, P. 1997. Controlling cell death. Science 275:1081–1082.20. Green, D., and J. C. Reed. 1998. Mitochondria and apoptosis. Science 281:
1309–1312.21. Harada, H., B. Becknell, M. Wilm, M. Mann, L. J. Huang, S. S. Taylor, J. D.
Scott, and S. J. Korsmeyer. 1999. Phosphorylation and inactivation of BADby mitochondria-anchored protein kinase A. Mol. Cell 3:413–422.
22. Jaffer, Z. M., and J. Chernoff. 2002. p21-activated kinases: three more jointhe Pak. Int. J. Biochem. Cell Biol. 34:713–717.
23. Jakobi, R., E. Moertl, and M. A. Koeppel. 2001. p21-activated protein kinasegamma-PAK suppresses programmed cell death of BALB3T3 fibroblasts.J. Biol. Chem. 276:16624–16634.
24. Kaufmann, T., S. Schlipf, J. Sanz, K. Neubert, R. Stein, and C. Borner. 2003.Characterization of the signal that directs Bcl-XL, but not Bcl-2, to themitochondrial outer membrane. J. Cell Biol. 160:53–64.
25. Kiosses, W. B., R. H. Daniels, C. Otey, G. M. Bokoch, and M. A. Schwartz.1999. A role for p21-activated kinase in endothelial cell migration. J. CellBiol. 147:831–844.
26. Lee, N., H. MacDonald, C. Reinhard, R. Halenbeck, A. Roulston, T. Shi, andL. T. Williams. 1997. Activation of hPAK65 by caspase cleavage inducessome of the morphological and biochemical changes of apoptosis. Proc. Natl.Acad. Sci. USA 94:13642–13647.
27. Lee, S. R., S. M. Ramos, A. Ko, D. Masiello, K. D. Swanson, M. L. Lu, andS. P. Balk. 2002. AR and ER interaction with a p21-activated kinase (PAK6).Mol. Endocrinol. 16:85–99.
28. Lei, M., W. Lu, W. Meng, M.-C. Parrini, M. J. Eck, B. J. Mayer, and S. C.Harrison. 2000. Structure of PAK1 in an autoinhibited conformation revealsa multistage activation switch. Cell 102:387–397.
29. Manser, E., H.-Y. Huang, T.-H. Loo, X. Q. Chen, T. Leung, and L. Lim. 1997.Expression of constitutively active a-Pak reveals effects of the kinase on actinand focal complexes. Mol. Cell. Biol. 17:1129–1143.
30. Morreale, A., M. Venkatesan, H. R. Mott, D. Owen, D. Nietlispach, P. N.Lowe, and E. D. Laue. 2000. Structure of Cdc42 bound to the GTPasebinding domain of PAK. Nat. Struct. Biol. 7:384–388.
31. Rudel, T., and G. M. Bokoch. 1997. Membrane and morphological changesin apoptotic cells regulated by caspase-mediated activation of PAK2. Science276:1571–1574.
32. Schurmann, A., A. F. Mooney, L. C. Sanders, M. A. Sells, H.-G. Wang, J. C.Reed, and G. M. Bokoch. 2000. p21-activated kinase 1 (PAK1) phosphor-ylates the death agonist BAD and protects cells from apoptosis. Mol. Cell.Biol. 20:453–461.
33. Sells, M. A., J. T. Barratt, J. Caviston, S. Ottilie, E. Leberer, and J. Chernoff.1998. Characterization of Pak2p, a pleckstrin homology domain-containing,p21-activated protein kinase from fission yeast. J. Biol. Chem. 273:18490–18498.
34. Sells, M. A., J. T. Boyd, and J. Chernoff. 1999. p21-activated kinase 1 (Pak1)regulates cell motility in mammalian fibroblasts. J. Cell Biol. 145:837–849.
35. Sells, M. A., and J. Chernoff. 1997. Emerging from the Pak: the p21-activatedprotein kinase family. Trends Cell. Biol. 7:162–167.
36. Sells, M. A., U. G. Knaus, S. Bagrodia, D. M. Ambrose, G. M. Bokoch, andJ. Chernoff. 1997. Human p21-activated kinase (Pak1) regulates actin orga-nization in mammalian cells. Curr. Biol. 7:202–210.
37. Sells, M. A., A. Pfaff, and J. Chernoff. 2000. Temporal and spatial distribu-tion of activated Pak1 in fibroblasts. J. Cell Biol. 151:1449–1457.
38. Tang, Y., H. Zhou, A. Chen, R. N. Pittman, and J. Field. 2000. The Aktproto-oncogene links Ras to Pak and cell survival signals. J. Biol. Chem.275:9106–9109.
39. Tu, H., and M. Wigler. 1999. Genetic evidence for Pak1 autoinhibition andits release by Cdc42. Mol. Cell. Biol. 19:602–611.
40. Walter, B. N., Z. Huang, R. Jakobi, P. T. Tuazon, E. S. Alnemri, G. Litwack,and J. A. Traugh. 1998. Cleavage and activation of p21-activated proteinkinase gamma-PAK by CPP32 (caspase 3). Effects of autophosphorylationon activity. J. Biol. Chem. 273:28733–28739.
41. Yang, F., X. Li, M. Sharma, M. Zarnegar, and Z. Lim. 2001. Androgenreceptor specifically interacts with a novel p21-activated kinase, PAK6.J. Biol. Chem. 276:15345–15353.
42. Zha, J., H. Harada, E. Yang, J. Jockel, and S. J. Korsmeyer. 1996. Serinephosphorylation of death agonist BAD in response to survival factor resultsin binding to 14–3–3 not BCL-X(L). Cell 87:619–628.
VOL. 23, 2003 Pak5 LOCALIZES TO MITOCHONDRIA 5539
on February 20, 2016 by guest
http://mcb.asm
.org/D
ownloaded from




















