
Novel method for the isolation and characterisation of the putative prostatic stem cell
11
Novel Method for the Isolation and Characterisation of the Putative Prostatic Stem Cell Rupesh I. Bhatt, 1,2 Michael D. Brown, 1 Claire A. Hart, 1 Paul Gilmore, 1,2 Vijay A.C. Ramani, 1,3 Nicholas J. George, 1,3 and Noel W. Clarke 1,2 * 1 Genito-Urinary Cancer Research Group, Cancer Research UK Paterson Institute for Cancer Research, Christie Hospital NHS Trust, Manchester, United Kingdom 2 Department of Urology, Salford Royal Hospital NHST, Salford, United Kingdom 3 Department of Urology, Withington Hospital, Nell Lane, Manchester, United Kingdom Received 8 January 2003; Revision Received 18 March 2003; Accepted 25 March 2003 Background: Prostate stem cells, responsible for the de- velopment, maturation, and function of the prostate, have been implicated in the aetiology of both benign prostate hyperplasia (BPH) and prostate cancer (CaP). However, research has been hampered by the lack of a definitive stem cell marker. We have adapted the protocol for dif- ferential Hoechst 33342 uptake by hemopoietic stem cells to enable isolation of putative stem cells from the pros- tate. Methods: Prostate epithelial cells isolated from prostate tissue obtained from patients with BPH after transurethral resection of the prostate were stained with Hoechst 33342. The Hoechst 33342 Red/Blue flow cytometry pro- file was then determined. Hoechst 33342 and Pyronin Y staining was used to determined the cell cycle status. Results: A verapamil-sensitive side population (SP) can be isolated from primary prostate tissue accounting for 1.38% 0.07% of prostate epithelial cells. Cell cycle analysis of this SP population revealed that the majority of SP cells are in either G 0 (12.38 0.31%) or G 1 (63.19 2.13%). Conclusions: The Hoechst 33342 dye efflux protocol can be adapted for the isolation of a SP from primary prostate tissue. Cytometry Part A 54A:89 –99, 2003. © 2003 Wiley-Liss, Inc. Key terms: prostate epithelial cells; Hoechst; stem cells; integrin 2 ; prostate cancer Stem cells are defined as clonogenic, pluripotent, self- renewing progenitor cells that can generate one or more specialised cell types (1–3). In common with many other tissues (4,5), the prostate is believed to harbour self- renewing or “stem” cells (6 –9). They are responsible for the development, maturation, and function of the prostate gland. These epithelial stem cells have been implicated in the aetiology of prostate cancer (10) and benign prostatic hyperplasia (BPH) (11). The proliferative compartment of normal prostate re- sides within the basal epithelial cell layer. Approximately 70% of proliferating cells are phenotypically basal; the remaining 30% lie within the luminal secretory epithelium (12,13). The ratio of basal cells-to-secretory luminal cells is between 1:3 and 1:6 in the human prostate (14). Controversy exists as to the exact nature and location of the prostatic stem cell, but experimental evidence sug- gests that it lies within the basal layer (15). Castration of male rats leads to a rapid involution of the prostate gland with only the basal epithelial cells surviving. Upon andro- gen replacement, the gland is restored to its normal func- tion (16). Only the differentiated luminal cells are affected by this androgen withdrawal, suggesting that regeneration of the gland must result from proliferation and differenti- ation of the surviving basal cells (10). Basal epithelial cells are capable of differentiating down various pathways and can replenish all layers within a castrated rat prostate after androgen replacement (16). Indeed basal cells retain the ability to undergo metaplasia and even undergo squamous differentiation in prostatic infarction and myoepithelial differentiation in sclerosing adenosis (9). Thus, because prostate basal epithelial cells make up the bulk of the proliferating cells and appear capable of differentiating down multiple lines, the postu- lated prostatic stem cells are most likely to reside in the basal compartment. Research continues to be hampered by the lack of a universally agreed prostatic stem cell marker. Many such markers have been proposed, including prostate stem cell *Correspondence to: Noel W Clarke, Genito-Urinary Cancer Research Group, Kay Kendall Laboratory, Paterson Institute for Cancer Research, Christie Hospital NHS Trust, Wilmslow Road, Manchester, M20 4BX, UK. E-mail: [email protected] Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/cyto.a.10058 © 2003 Wiley-Liss, Inc. Cytometry Part A 54A:89 –99 (2003)
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Novel method for the isolation and characterisation of the putative prostatic stem cell

Novel Method for the Isolation and Characterisation ofthe Putative Prostatic Stem Cell
Rupesh I. Bhatt,1,2 Michael D. Brown,1 Claire A. Hart,1 Paul Gilmore,1,2 Vijay A.C. Ramani,1,3
Nicholas J. George,1,3 and Noel W. Clarke1,2*1Genito-Urinary Cancer Research Group, Cancer Research UK Paterson Institute for Cancer Research,
Christie Hospital NHS Trust, Manchester, United Kingdom2Department of Urology, Salford Royal Hospital NHST, Salford, United Kingdom
3Department of Urology, Withington Hospital, Nell Lane, Manchester, United Kingdom
Received 8 January 2003; Revision Received 18 March 2003; Accepted 25 March 2003
Background: Prostate stem cells, responsible for the de-velopment, maturation, and function of the prostate, havebeen implicated in the aetiology of both benign prostatehyperplasia (BPH) and prostate cancer (CaP). However,research has been hampered by the lack of a definitivestem cell marker. We have adapted the protocol for dif-ferential Hoechst 33342 uptake by hemopoietic stem cellsto enable isolation of putative stem cells from the pros-tate.Methods: Prostate epithelial cells isolated from prostatetissue obtained from patients with BPH after transurethralresection of the prostate were stained with Hoechst33342. The Hoechst 33342 Red/Blue flow cytometry pro-file was then determined. Hoechst 33342 and Pyronin Ystaining was used to determined the cell cycle status.
Results: A verapamil-sensitive side population (SP) can beisolated from primary prostate tissue accounting for 1.38%� 0.07% of prostate epithelial cells. Cell cycle analysis ofthis SP population revealed that the majority of SP cells arein either G0 (12.38 � 0.31%) or G1 (63.19 � 2.13%).Conclusions: The Hoechst 33342 dye efflux protocol canbe adapted for the isolation of a SP from primary prostatetissue. Cytometry Part A 54A:89–99, 2003.© 2003 Wiley-Liss, Inc.
Key terms: prostate epithelial cells; Hoechst; stem cells;integrin �2; prostate cancer
Stem cells are defined as clonogenic, pluripotent, self-renewing progenitor cells that can generate one or morespecialised cell types (1–3). In common with many othertissues (4,5), the prostate is believed to harbour self-renewing or “stem” cells (6–9). They are responsible forthe development, maturation, and function of the prostategland. These epithelial stem cells have been implicated inthe aetiology of prostate cancer (10) and benign prostatichyperplasia (BPH) (11).
The proliferative compartment of normal prostate re-sides within the basal epithelial cell layer. Approximately70% of proliferating cells are phenotypically basal; theremaining 30% lie within the luminal secretory epithelium(12,13). The ratio of basal cells-to-secretory luminal cells isbetween 1:3 and 1:6 in the human prostate (14).
Controversy exists as to the exact nature and location ofthe prostatic stem cell, but experimental evidence sug-gests that it lies within the basal layer (15). Castration ofmale rats leads to a rapid involution of the prostate glandwith only the basal epithelial cells surviving. Upon andro-gen replacement, the gland is restored to its normal func-tion (16). Only the differentiated luminal cells are affectedby this androgen withdrawal, suggesting that regeneration
of the gland must result from proliferation and differenti-ation of the surviving basal cells (10).
Basal epithelial cells are capable of differentiating downvarious pathways and can replenish all layers within acastrated rat prostate after androgen replacement (16).Indeed basal cells retain the ability to undergo metaplasiaand even undergo squamous differentiation in prostaticinfarction and myoepithelial differentiation in sclerosingadenosis (9). Thus, because prostate basal epithelial cellsmake up the bulk of the proliferating cells and appearcapable of differentiating down multiple lines, the postu-lated prostatic stem cells are most likely to reside in thebasal compartment.
Research continues to be hampered by the lack of auniversally agreed prostatic stem cell marker. Many suchmarkers have been proposed, including prostate stem cell
*Correspondence to: Noel W Clarke, Genito-Urinary Cancer ResearchGroup, Kay Kendall Laboratory, Paterson Institute for Cancer Research,Christie Hospital NHS Trust, Wilmslow Road, Manchester, M20 4BX, UK.
E-mail: [email protected] online in Wiley InterScience (www.interscience.wiley.com).
DOI: 10.1002/cyto.a.10058
© 2003 Wiley-Liss, Inc. Cytometry Part A 54A:89–99 (2003)

antigen (PSCA) (17), prostate specific membrane antigen(PSMA) (18–20), and phosphoprotein p32 (21–23). How-ever, none are entirely tissue or location specific (24,25).
Integrins have been used to identify stem cells in skin(26) and testis (27). Epidermal stem cells express greaterlevels of integrin �2�1 than transit amplifying cells (TAP)(28). Epithelial cells within the human prostate expressintegrin �2�1, �3�1, and �6�4 (29–31). Integrin �2�1,restricted to prostatic basal cells (31), mediates adhesionof prostate epithelial cells to types I and IV collagens andto laminin I on the basement membrane (32). Putativestem cells have been shown to express higher levels of theintegrin �2 subunit than other cells in the basal layer, andthese integrin “bright” cells constitute approximately 1%of basal cells (33). Interest has focused on patterns ofprimary cytokeratin expression as markers for differentialepithelial phenotypes. Luminal cells express cytokeratins8 and 18 (34–37). In contrast, basal cells express mainlylarge molecular weight cytokeratins, namely CK4-7, 10,14, and 15 (38,39).
In addition to cell surface markers, cell cycle status maybe useful in the identification of the stem cell. Stem cellsare proliferatively quiescent, residing predominantly inthe G0 phase of the cell cycle (40,41). Therefore, cellcycle status may be critical in defining strategies for stemcell selection. It is now possible to distinguish and isolateviable cells in G0 or G1 phase using a combination of theDNA and RNA binding dyes Hoechst 33342 and Pyronin Y(42).
Haemopoietic stem cells (HSC) have been the subject ofconsiderable interest for decades and research has contin-ued into their characteristics. In 1961, Till and McCullochdemonstrated the existence of clonogenic bone marrowprecursors that give rise to multilineage haematopoieticcolonies in the spleen (1). Although this finding providedevidence for the existence of the HSC, their isolationremained a problem until the development of in vitro andin vivo quantitative assays for clonogenic human stem andprogenitor haematolymphoid cells (43). This allowed de-velopment of distinctive marker profiles for each progen-itor using monoclonal antibodies (44).
HSC can be identified by differential uptake of the vitaldye Hoechst 33342. This is a fluorescent dye that binds toDNA in live cells and, therefore, is an indicator of cellcycle as it relates to DNA content. It is actively taken up byall cells (excepting red blood cells) but is actively ef-fluxed, by means of the multidrug resistance protein(MDR1) gene product (p-glycoprotein), by primitive orstem cells (45). When analysed using flow cytometry asmall population of Hoechst “low” cells termed side pop-ulation (SP) cells can be defined. The SP profile is lostupon the addition of the MDR1 transporter inhibitor,verapamil. In vivo, these cells have been shown to beenriched 1,000-fold for HSC activity, which can protectmurine recipients from lethal irradiation at low cell dosesand contribute to both myeloid and lymphoid lineages(45). The cells have been found in multiple species, in-cluding murine, rhesus monkey, swine, and human bonemarrow.
Although other strategies for the isolation and enrich-ment of HSC exist (44,46–48), the Hoechst SP regionallows a single dye or manipulation to define a populationwith HSC activity that is strikingly small, homogeneous,easily identifiable and highly reproducible (45). Further-more, the Hoechst SP region allows independent verifica-tion of cell surface characteristics, cell cycle kinetics, andthe ability to collect and process cells in long-term in vitroand in vivo culture.
The aim of our study was to evaluate the Hoechst dyeefflux protocol for solid tissue using the prostate as anexample. We have modified the existing haematologicalHoechst protocol to isolate an SP population from humanprostatic epithelial cells (PEC), and subsequently investi-gated isolated SP cells with existing markers of stem cellphenotype, namely basal cytokeratins and integrin �2.
MATERIAL AND METHODSMaterials
All tissue culture medium and reagents were purchasedfrom Invitrogen (Paisley, UK), with the exception of pros-tate epithelial cell basal medium (Pr-EBM), Clonetics (Bio-Whittaker, Berkshire, UK). Bovine serum albumin (BSA),streptavidin PE-Cy5, propidium iodide, pan cytokeratin �(clone C-11), cytokeratin 8, cytokeratin 14 (clone CKB1),R(�)- verapamil hydrochloride, and metrizamide wasfrom Sigma-Aldridge (Poole, UK); foetal calf serum (FCS)was supplied by Sera Labs (Sussex, UK); and Worthingtoncollagenase type 1 and trypsin were from Lorne Labora-tories, Ltd. (Twyford, UK). Hoechst 33342 was acquiredfrom Molecular Probes (Eugene, Oregon, USA), Ber-EP4fluorescein isothiocyanate (FITC; epithelial antigen) andrabbit anti-mouse biotinylated (RAMBO) antibodies fromDAKO, Ltd. (Cambridge, UK), and streptavidin PE-Cy7conjugate was from Caltag Laboratories (Burlingame, CA,USA). Integrin �2 (CD49b) and CD45 FITC were suppliedby BD Pharmingen (Heidelberg, Germany).
Methods
Prostatic tissue collection and culture. Informedconsent was obtained before tissue collection. Men under-going transurethral resection of the prostate for bladderoutflow obstruction arising from benign disease werestudied. Prostate chips were obtained under sterile con-ditions. Each individual prostate chip was bisected withhalf being sent for histological analysis for diagnostic eval-uation and the remainder used for tissue culture. Prostateepithelial cell isolation and characterisation was carriedout by the method previously described (49). Briefly, byusing sterile forceps and scissors, the prostate chips werechopped and placed in collagenase type I at 200 U/ml inRPMI 1640 medium with 2% v/v FCS overnight on ashaking platform at 37°C. The digest was then brokendown further by shaking in 0.1% trypsin in PBS with 1%BSA and 1 mM ethylenediaminetetraacetic acid (EDTA) for15–20 min. The cell suspension was then washed threetimes in PBS with 1% BSA and 1 mM EDTA before resus-pending in RPMI 10% v/v FCS. Prostate epithelial cells
90 BHATT ET AL.
were separated from fibroblasts by differential centrifuga-tion (360 g, 1 min without braking). This process pro-duced a supernatant enriched for fibroblasts and a pelletenriched for epithelia. The epithelial cell suspension wasthen spun on a metrizamide gradient (1.079 g/ml), and thecells were isolated from the interface.
Ber-EP4/�2/CD45 labelling of prostate cells. Iso-lated prostate epithelial cells were labelled at room tem-perature with either anti-human integrin �2 monoclonalantibody or Ber-EP4 antibody (8 �g/ml in 1% BSA/PBS) for30 min before the addition of the secondary antibody,RAMBO (2.6 �g/ml in 1% BSA/PBS) for 30 min. Afterwashing with PBS, the cells were incubated for 20 min inthe dark with streptavidin PE-Cy7 (20 �g/ml). Sampleswere then dual labelled with CD45-FITC (1 �g/ml in 1%BSA/PBS) for 30 min.
Ber-EP4/�2 and Hoechst labelling for flow cytom-etry. Isolated prostate epithelial cells were labelled atroom temperature with anti-human integrin �2 monoclo-nal antibody (8 �g/ml in 1% BSA/PBS) for 30 min beforethe addition of the secondary antibody, RAMBO (2.6�g/ml in 1% BSA/PBS) for 30 min. After washing with PBS,the cells were incubated for 20 min in the dark withstreptavidin PE-Cy7 (20 �g/ml).
Hoechst staining was performed by using a modifica-tion of the protocol for HSC as described by (45). Briefly,epithelial cells were resuspended in Hoechst buffer(Hanks’ balanced salts solution, 10% FCS, 1% D-glucose,and 20mM HEPES) and warmed to 37°C. Hoechst 33342was then added to give a final concentration of 2 �M andthe cells incubated at 37°C for 2 h. Fifteen minutes beforethe end of incubation, the cells were labelled with mono-clonal anti-human Ber-EP4 directly conjugated to FITC (8�g/ml). The cells were then washed in ice-cold Hoechstbuffer before resuspending in ice-cold Hoechst buffer
containing propidium iodide (PI) at 20 ng/ml. Fluores-cence activated cell sorting analysis was performed imme-diately thereafter. To confirm active efflux of Hoechst33342 by means of the MDR1 transporter, verapamil hy-drochloride was added to a final concentration of 50 �Mto the control samples before the addition of Hoechst33342.
Flow cytometry isolation of the SP fraction. Flowcytometry was carried out using a Becton Dickinson FACSVantage SE flow cytometer. Hoechst 33342 was excitedwith an argon ion, ultraviolet-enhanced laser at 350 nm,and its fluorescence was measured with a 424/44 BP filter(Hoechst BLUE) and a 675DF20 BP optical filter (HoechstRED; Omega Optical, Brattleboro VT). A 640 LP dichroicmirror was used to separate the emission wavelengths. PIfluorescence was also measured through the 675DF20 BP(having been excited at 350 nm). A second argon ion laserwas used to excite the additional fluorochrome PE-Cy7 at488nM. PE-Cy7 was measured using a 787RDF40 (OmegaOptical) filter.
Prostate epithelial cells stained with the Hoechst dyewere kept ice cold during analysis on the flow cytometerby the use of a chilling apparatus. Live gates were estab-lished on forward versus side scatter and PI parameters, allPI-positive and dead cells were excluded. Cells understudy were sorted by positive labelling for Ber-EP4 FITCand/or integrin �2 streptavidin PE-Cy7 before being anal-ysed for Hoechst staining. These cells were then analysedusing the Hoechst BLUE versus RED profile, with BLUE onthe vertical axis and RED on the horizontal axis on a linearscale. A third gate was placed to select for the Hoechstlow SP tail.
Cell cycle characterisation of SP fraction. Sampleswere processed as in prostatic tissue collection: epithelialcells were isolated and live epithelial cells selected after
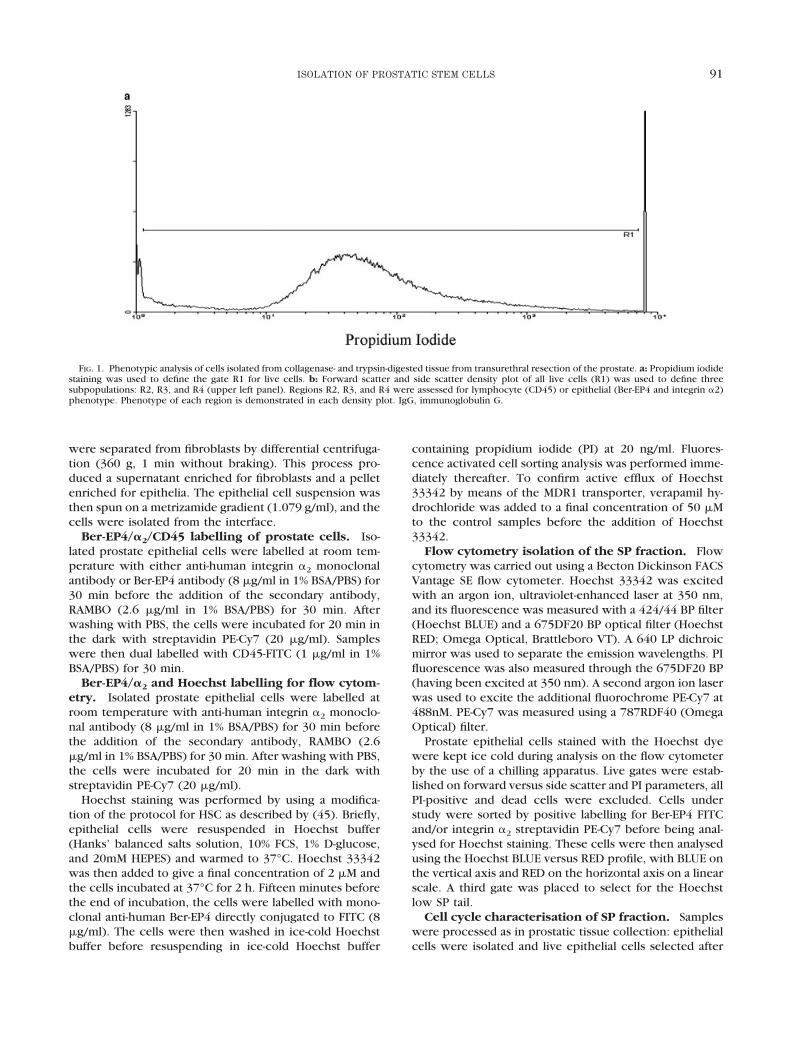
FIG. 1. Phenotypic analysis of cells isolated from collagenase- and trypsin-digested tissue from transurethral resection of the prostate. a: Propidium iodidestaining was used to define the gate R1 for live cells. b: Forward scatter and side scatter density plot of all live cells (R1) was used to define threesubpopulations: R2, R3, and R4 (upper left panel). Regions R2, R3, and R4 were assessed for lymphocyte (CD45) or epithelial (Ber-EP4 and integrin �2)phenotype. Phenotype of each region is demonstrated in each density plot. IgG, immunoglobulin G.
91ISOLATION OF PROSTATIC STEM CELLS
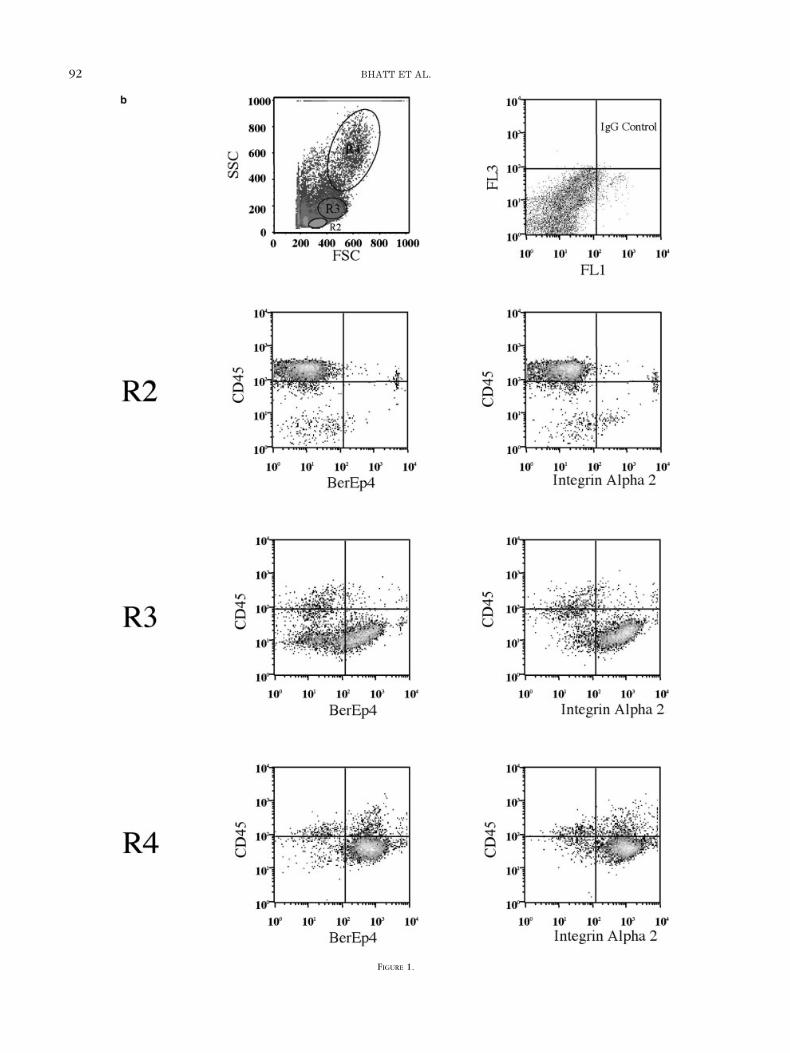
FIGURE 1.
92 BHATT ET AL.

processing through a Metrizamide gradient. All epithelialfractions were resuspended in Hoechst buffer andwarmed to 37°C. Hoechst 33342 was then added to givea concentration of 2 �M and incubated at 37°C for 45 min.Pyronin Y (250 ng/�l) was added to each tube, and thesamples were incubated for 45 min. Monoclonal anti-human Ber-EP4 FITC (8 �g/ml) was added as appropriate15 min before the end. After this, ice-cold Hoechst bufferwas added immediately and the samples were washedthen resuspended in ice-cold Hoechst buffer. The sampleswere analysed immediately by flow cytometry.
Flow cytometry was performed using a modification ofthe method described above. Cells under study were se-lected by positive labelling for Ber-EP4 FITC before beinganalysed for Hoechst and Pyronin Y staining. These cellswere then analysed by plotting the Hoechst profile on thex-axis and Pyronin Y along the y-axis in a linear scale.
Cytokeratin phenotype studies. Samples were pro-cessed as above, divided into two fractions, and labelledwith either cytokeratin 8 or 14 indirectly conjugated toPE-Cy5. Samples were then dual labelled with Ber-EP4FITC and integrin �2 PE-CY7. Flow cytometry was per-formed as described and analysed on forward (FSC) andside (SSC) scatter.
Statistical Analysis
Statistical analysis was performed with the chi squaredtest. Fisher’s exact test was used when numbers analysedwere less than 5.
RESULTSIsolation and Characterisation of Prostate
Epithelial Cells
Cells isolated from prostate chips from men undergoingtransurethral resection of the prostate for bladder outflowobstruction arising from BPH were analysed by flow cy-tometry analysis to assess their phenotype. Live cells weregated (R1) on propidium iodide staining (Fig. 1a). A den-sity plot was then constructed with forward scatter plot-ted along the x-axis and side scatter along the y-axis.Density plots reveal three discrete populations defined asR2, R3, and R4 (Fig. 1b). Cells lying within region R2 wereextremely small, whereas the cells in R3 and R4 werelarger and more comparable with each other.
The forward and side scatter profile of the cells locatedwithin R2 has a similar profile to haemopoietic lympho-cytes from bone marrow. To establish the phenotype ofthe cells within each region, the isolated prostatic cellswere labelled with the lymphocyte marker anti-CD45 andthe epithelial marker Ber-EP4. Recent work by Collins etal. demonstrated that putative prostatic stem cells expressintegrin �2 on their surface (33). Therefore, each regionwas stained with integrin �2 to localise the region con-taining the putative stem cells.
R2 contains a population of lymphocytes as determinedby their low forward and side scatter profile, positivestaining for the lymphocyte marker CD45 and negativestaining for either the epithelial cell marker Ber-EP4 or
integrin �2. Regions R3 and R4 contain Ber-EP4–positiveepithelial populations. However, we were unable to iso-late a distinct population containing stem cells based onintegrin �2 labeling, as both R3 and R4 cell populationscontained cells that were integrin �2 positive (Fig. 1b).Because Ber-EP4 is highly conserved on epithelial cells,Ber-EP4 was used to select epithelial cell populations forfurther analysis, hence eliminating the lymphocyte popu-lation.
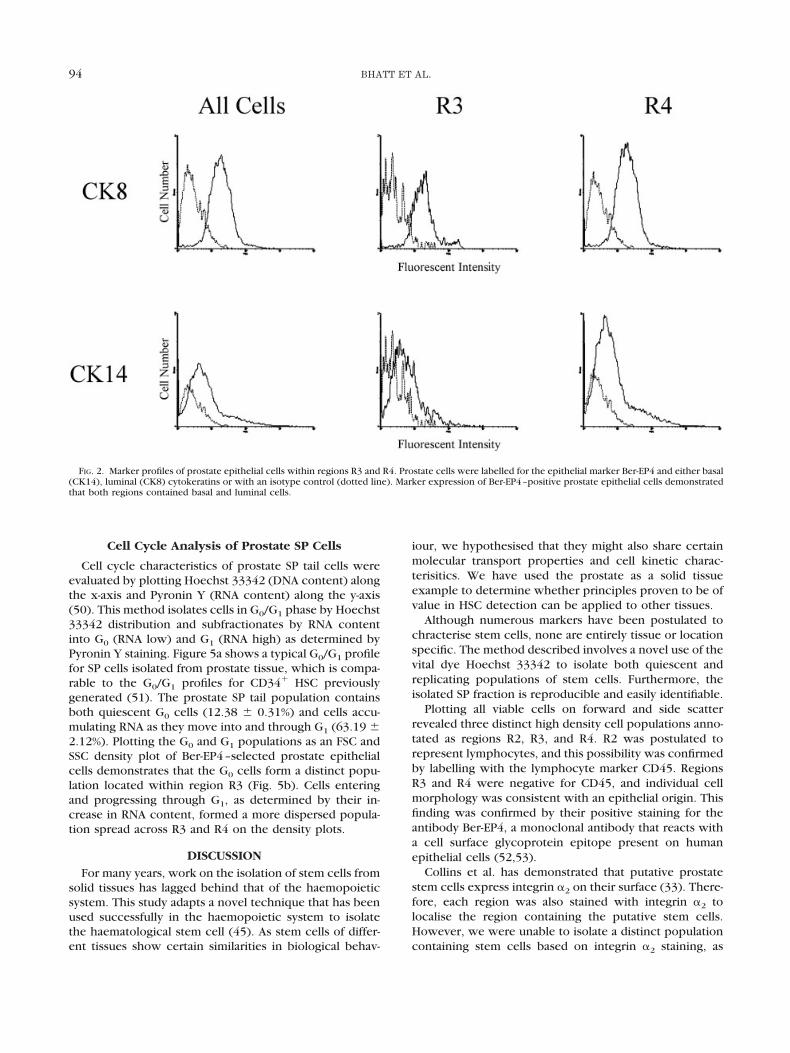
Phenotypic analysis of Ber-EP4–positive epithelial cellswithin regions R3 and R4 (Fig. 2) using basal (CK14) andluminal (CK8) cytokeratins demonstrated that both re-gions contained basal and luminal cells, with R3 beingslightly enriched for luminal cells (2:9 basal:luminal in R3compared with 1:3 in R4).
Isolation of Prostate Stem Cells UsingHoechst 33342
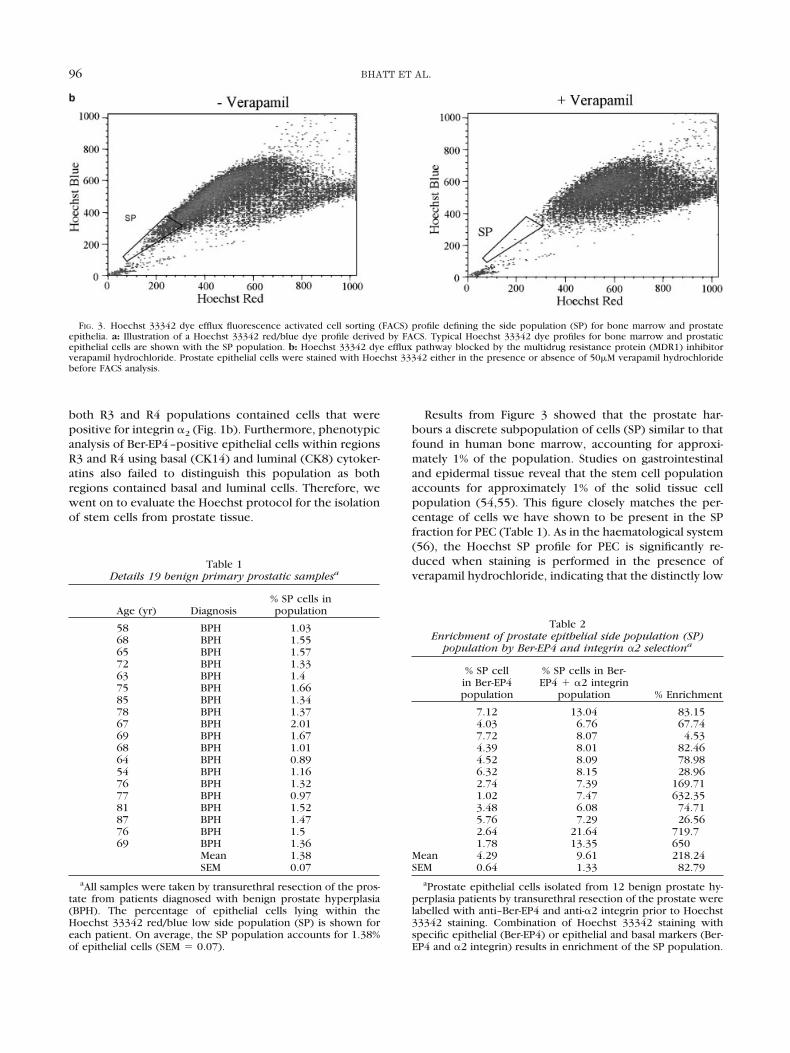
The differential uptake of the vital dye Hoechst 33342has been used to identify HSC (45). We sought to evaluatethe Hoechst 33342 dye efflux protocol for the identifica-tion of stem cells from prostate tissue. The Hoechst profilewas plotted in a similar way to that published previouslyfor haemopoietic cells (45) with Hoechst BLUE on thevertical axis and Hoechst RED on the horizontal. Figure 3acompares the predicted Hoechst 33342 profile with theprofiles generated by HSC isolated from human bone mar-row and Ber-EP4–positive prostatic epithelial cells derivedfrom a BPH sample. Both bone marrow and prostateepithelia generate a Hoechst 33342 profile with a distinctside population (SP). The prostate epithelial cell profilehas a greater concentration of cells within the SP popula-tion compared with the bone marrow SP population. Thisincreased number of cells within the side population wasobserved in all 19 prostate samples studied. Table 1 detailsthe percentage of epithelial cells lying within the Hoechstlow SP tail for each prostate sample. On average, theprostate SP accounts for 1.38 � 0.07% of epithelial cells(n � 19) compared with 0.14 � 0.09% (n � 4) of cellsisolated from bone marrow SP. Blocking the activeHoechst 33342 dye efflux pathway using verapamil hydro-chloride, a potent inhibitor of MDR1, significantly reducesthe SP population (Fig. 3b).
Enrichment for the basal integrin �2 was studied bydifferential staining of the tail and body of the Hoechstprofile with Ber-EP4 and integrin �2. The ratio of epithelialcells (Ber-EP4 alone) to basal cells (Ber-EP4 and integrin�2) was then calculated. Table 2 details 12 samples anddocuments this enrichment of basal cells within the tail(mean enrichment � 218%, P � 0.005).
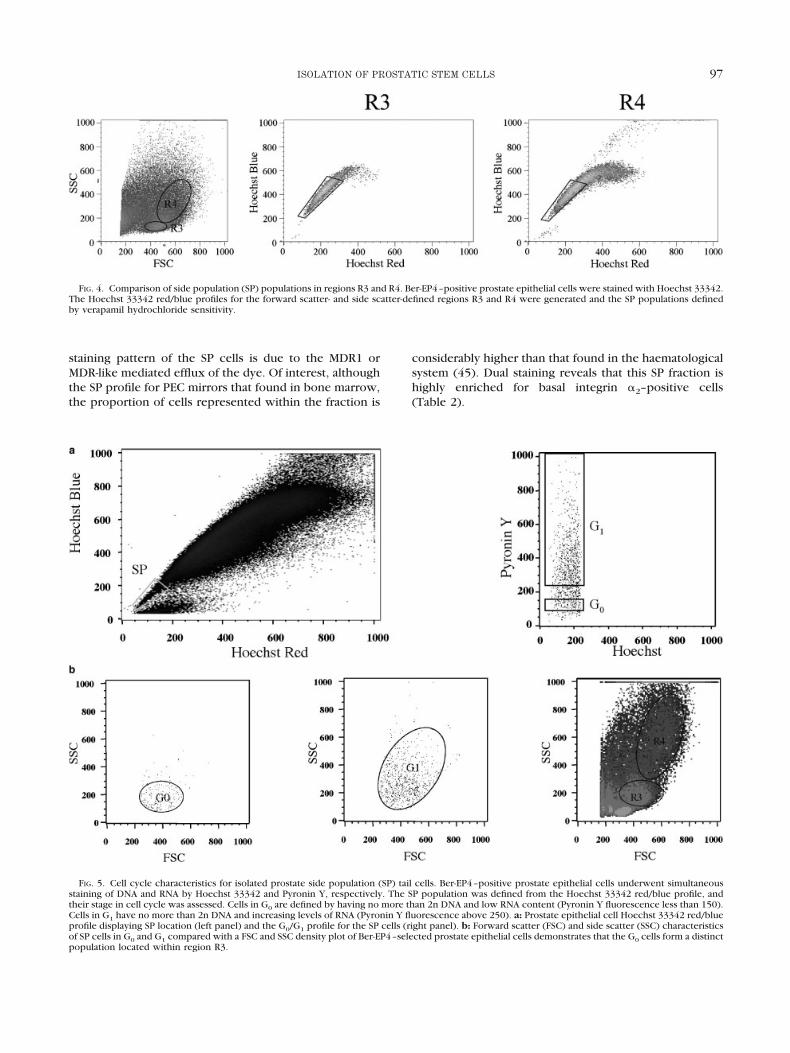
Hoechst 33342 profiles for regions R3 and R4 definedby FSC and SSC density plots of prostate Ber-EP4–positiveepithelial cells were generated and the SP tail populationscompared (Fig. 4). R3 is enriched for SP cells containing44.5% � 4.4% SP cells compared with 29% � 1.34% SPcells in R4 (n � 3).
93ISOLATION OF PROSTATIC STEM CELLS
Cell Cycle Analysis of Prostate SP Cells
Cell cycle characteristics of prostate SP tail cells wereevaluated by plotting Hoechst 33342 (DNA content) alongthe x-axis and Pyronin Y (RNA content) along the y-axis(50). This method isolates cells in G0/G1 phase by Hoechst33342 distribution and subfractionates by RNA contentinto G0 (RNA low) and G1 (RNA high) as determined byPyronin Y staining. Figure 5a shows a typical G0/G1 profilefor SP cells isolated from prostate tissue, which is compa-rable to the G0/G1 profiles for CD34� HSC previouslygenerated (51). The prostate SP tail population containsboth quiescent G0 cells (12.38 � 0.31%) and cells accu-mulating RNA as they move into and through G1 (63.19 �2.12%). Plotting the G0 and G1 populations as an FSC andSSC density plot of Ber-EP4–selected prostate epithelialcells demonstrates that the G0 cells form a distinct popu-lation located within region R3 (Fig. 5b). Cells enteringand progressing through G1, as determined by their in-crease in RNA content, formed a more dispersed popula-tion spread across R3 and R4 on the density plots.
DISCUSSIONFor many years, work on the isolation of stem cells from
solid tissues has lagged behind that of the haemopoieticsystem. This study adapts a novel technique that has beenused successfully in the haemopoietic system to isolatethe haematological stem cell (45). As stem cells of differ-ent tissues show certain similarities in biological behav-
iour, we hypothesised that they might also share certainmolecular transport properties and cell kinetic charac-terisitics. We have used the prostate as a solid tissueexample to determine whether principles proven to be ofvalue in HSC detection can be applied to other tissues.
Although numerous markers have been postulated tochracterise stem cells, none are entirely tissue or locationspecific. The method described involves a novel use of thevital dye Hoechst 33342 to isolate both quiescent andreplicating populations of stem cells. Furthermore, theisolated SP fraction is reproducible and easily identifiable.
Plotting all viable cells on forward and side scatterrevealed three distinct high density cell populations anno-tated as regions R2, R3, and R4. R2 was postulated torepresent lymphocytes, and this possibility was confirmedby labelling with the lymphocyte marker CD45. RegionsR3 and R4 were negative for CD45, and individual cellmorphology was consistent with an epithelial origin. Thisfinding was confirmed by their positive staining for theantibody Ber-EP4, a monoclonal antibody that reacts witha cell surface glycoprotein epitope present on humanepithelial cells (52,53).
Collins et al. has demonstrated that putative prostatestem cells express integrin �2 on their surface (33). There-fore, each region was also stained with integrin �2 tolocalise the region containing the putative stem cells.However, we were unable to isolate a distinct populationcontaining stem cells based on integrin �2 staining, as
FIG. 2. Marker profiles of prostate epithelial cells within regions R3 and R4. Prostate cells were labelled for the epithelial marker Ber-EP4 and either basal(CK14), luminal (CK8) cytokeratins or with an isotype control (dotted line). Marker expression of Ber-EP4–positive prostate epithelial cells demonstratedthat both regions contained basal and luminal cells.
94 BHATT ET AL.
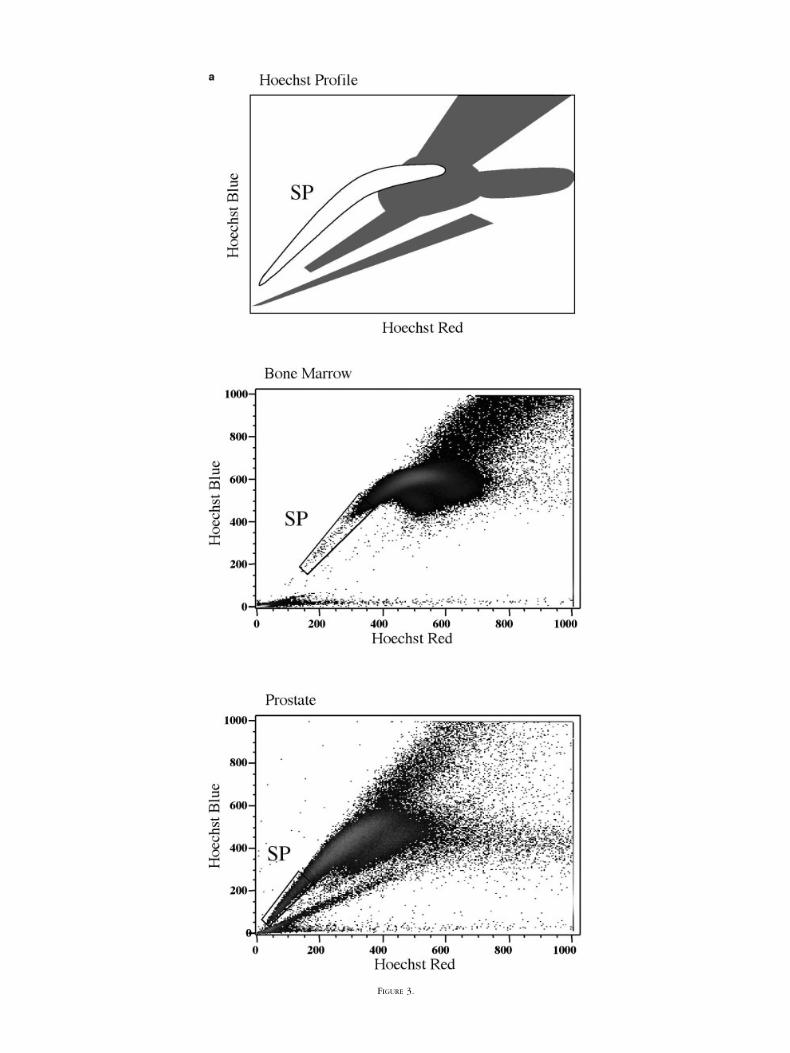
FIGURE 3.
both R3 and R4 populations contained cells that werepositive for integrin �2 (Fig. 1b). Furthermore, phenotypicanalysis of Ber-EP4–positive epithelial cells within regionsR3 and R4 using basal (CK14) and luminal (CK8) cytoker-atins also failed to distinguish this population as bothregions contained basal and luminal cells. Therefore, wewent on to evaluate the Hoechst protocol for the isolationof stem cells from prostate tissue.
Results from Figure 3 showed that the prostate har-bours a discrete subpopulation of cells (SP) similar to thatfound in human bone marrow, accounting for approxi-mately 1% of the population. Studies on gastrointestinaland epidermal tissue reveal that the stem cell populationaccounts for approximately 1% of the solid tissue cellpopulation (54,55). This figure closely matches the per-centage of cells we have shown to be present in the SPfraction for PEC (Table 1). As in the haematological system(56), the Hoechst SP profile for PEC is significantly re-duced when staining is performed in the presence ofverapamil hydrochloride, indicating that the distinctly low
Table 1Details 19 benign primary prostatic samplesa
Age (yr) Diagnosis% SP cells inpopulation
58 BPH 1.0368 BPH 1.5565 BPH 1.5772 BPH 1.3363 BPH 1.475 BPH 1.6685 BPH 1.3478 BPH 1.3767 BPH 2.0169 BPH 1.6768 BPH 1.0164 BPH 0.8954 BPH 1.1676 BPH 1.3277 BPH 0.9781 BPH 1.5287 BPH 1.4776 BPH 1.569 BPH 1.36
Mean 1.38SEM 0.07
aAll samples were taken by transurethral resection of the pros-tate from patients diagnosed with benign prostate hyperplasia(BPH). The percentage of epithelial cells lying within theHoechst 33342 red/blue low side population (SP) is shown foreach patient. On average, the SP population accounts for 1.38%of epithelial cells (SEM � 0.07).
Table 2Enrichment of prostate epithelial side population (SP)
population by Ber-EP4 and integrin �2 selectiona
% SP cellin Ber-EP4population
% SP cells in Ber-EP4 � �2 integrin
population % Enrichment
7.12 13.04 83.154.03 6.76 67.747.72 8.07 4.534.39 8.01 82.464.52 8.09 78.986.32 8.15 28.962.74 7.39 169.711.02 7.47 632.353.48 6.08 74.715.76 7.29 26.562.64 21.64 719.71.78 13.35 650
Mean 4.29 9.61 218.24SEM 0.64 1.33 82.79
aProstate epithelial cells isolated from 12 benign prostate hy-perplasia patients by transurethral resection of the prostate werelabelled with anti–Ber-EP4 and anti-�2 integrin prior to Hoechst33342 staining. Combination of Hoechst 33342 staining withspecific epithelial (Ber-EP4) or epithelial and basal markers (Ber-EP4 and �2 integrin) results in enrichment of the SP population.
FIG. 3. Hoechst 33342 dye efflux fluorescence activated cell sorting (FACS) profile defining the side population (SP) for bone marrow and prostateepithelia. a: Illustration of a Hoechst 33342 red/blue dye profile derived by FACS. Typical Hoechst 33342 dye profiles for bone marrow and prostaticepithelial cells are shown with the SP population. b: Hoechst 33342 dye efflux pathway blocked by the multidrug resistance protein (MDR1) inhibitorverapamil hydrochloride. Prostate epithelial cells were stained with Hoechst 33342 either in the presence or absence of 50�M verapamil hydrochloridebefore FACS analysis.
96 BHATT ET AL.
staining pattern of the SP cells is due to the MDR1 orMDR-like mediated efflux of the dye. Of interest, althoughthe SP profile for PEC mirrors that found in bone marrow,the proportion of cells represented within the fraction is
considerably higher than that found in the haematologicalsystem (45). Dual staining reveals that this SP fraction ishighly enriched for basal integrin �2–positive cells(Table 2).
FIG. 4. Comparison of side population (SP) populations in regions R3 and R4. Ber-EP4–positive prostate epithelial cells were stained with Hoechst 33342.The Hoechst 33342 red/blue profiles for the forward scatter- and side scatter-defined regions R3 and R4 were generated and the SP populations definedby verapamil hydrochloride sensitivity.
FIG. 5. Cell cycle characteristics for isolated prostate side population (SP) tail cells. Ber-EP4–positive prostate epithelial cells underwent simultaneousstaining of DNA and RNA by Hoechst 33342 and Pyronin Y, respectively. The SP population was defined from the Hoechst 33342 red/blue profile, andtheir stage in cell cycle was assessed. Cells in G0 are defined by having no more than 2n DNA and low RNA content (Pyronin Y fluorescence less than 150).Cells in G1 have no more than 2n DNA and increasing levels of RNA (Pyronin Y fluorescence above 250). a: Prostate epithelial cell Hoechst 33342 red/blueprofile displaying SP location (left panel) and the G0/G1 profile for the SP cells (right panel). b: Forward scatter (FSC) and side scatter (SSC) characteristicsof SP cells in G0 and G1 compared with a FSC and SSC density plot of Ber-EP4–selected prostate epithelial cells demonstrates that the G0 cells form a distinctpopulation located within region R3.
97ISOLATION OF PROSTATIC STEM CELLS

As with the HSC (57), cell cycle characterisation ofprostate SP tail cells reveals that the SP tail populationcontains both quiescent G0 cells and cells accumulatingRNA as they move into and through G1. G0 cells form adiscrete population located within region R3, whereasthose entering and progressing through G1 formed a moredispersed population spread across R3 and R4 on thedensity plots.
Cellular populations are highly ordered within epithe-lial tissues. They contain a small stem cell population thatharbours the capacity for extended or prolonged growth,giving rise to progeny with more limited proliferativecapacity and greater differentiation (58–60). There existsa discrete population of cells derived directly from thestem cell that are either second-order stem cells or cellswith limited proliferative capacity, often termed TAP.These daughter cells are responsible for maintaining tissuebalance (58,61). In the gastrointestinal system, cell typesare organised whereby stem cells, TAP, and mature termi-nally differentiated cells occupy discrete locations withinthe tissue, often forming stratified layers (59). A similarhierarchical arrangement has been postulated for the pros-tate (10,11), but controversy remains regarding the exactnature and location of these TAP.
Foster and Ke regard the pluripotent stem cell as a basalcell that can differentiate further into a basal epithelial cell(and then into a luminal cell) or can itself differentiatedown the neuroendocrine lineage and form a neuroendo-crine cell (9). These basal epithelial cells, therefore, wouldrepresent a transit amplifying cell population that stillresides within the basal layer. Xue et al. detail a stem cell,lying within the basal layer, that differentiates into a basalepithelial cell (62). Differentiation from this cell leads toan intermediate cell (TAP) and then after to both luminalepithelial and neuroendocrine cells. Xue et al. postulatethat BPH and CaP derive secondary to an epigeneticchange within the intermediate cell (62). Evidence is ac-cumulating for the presence of such an intermediate ortransit amplifying cell within the prostate with the iden-tification of cells that coexpress both basal and luminalcytokeratins (36,63). Our results suggest the presence of atransit amplifying population, with SP tail cells in regionsR3 and R4 representing prime candidates with a part basalphenotype and a high proportion of cells in G1. Thesecells may be identified by the expression of integrin �2 inthe basal layer, and they retain properties enabling themto efflux Hoechst 33342.
Phenotype studies were performed by cytokeratin stain-ing of isolated epithelial cells. Dual labelling with basaland luminal cytokeratins and the basal integrin �2 placethe SP cell within the basal layer, but there are at least twotypes of Hoechst low cells as defined by their cellularmorphology and size (regions R3 and R4). We postulatethat cells in R3, which are small, express basal cytokera-tins, are in the quiescent cell cycle phase (G0), and fall tothe extreme left of the Hoechst profile (very low), repre-sent the putative prostatic stem cells. As mentionedabove, we also postulate that SP tail cells in R3 and R4represent an intermediate or transit amplifying cell popu-
lation; the majority of these cells are in the G1 phase of thecell cycle and dual labelling studies reveal that they ex-press an intermediate cytokeratin characteristic and alsoshow enrichment for integrin �2. Integrin �2, therefore, ispresent on the putative stem cell and on the transit am-plifying cell.
Future work will be required to characterise the SP cellfurther with respect to long-term cell culture. In vivostudies may be beneficial in this regard, comparing stemcell markers and using the Hoechst technique to confirmor reject individual marker expression for the stem cell.
In conclusion, we have shown that it is possible toisolate SP cells from primary human prostatic tissue in away that is similar to that previously identified from hae-mopoietic cells, liver, lung, muscle, and brain tissue.These SP cells account for approximately 1% of the wholecell population and demonstrate enrichment for the basalcell marker integrin �2. The SP cells reveal a preponder-ance of quiescent G0 cells within the small epithelial cellpopulation and can be located to a region within theintegrin �2–positive basal cell layer. The above propertiesare consistent with a stem cell phenotype.
ACKNOWLEDGMENTThe authors thank the British Urological Foundation
and the Reverend Hugh Mackay Fellowship in Urology(RCS Edinburgh) for their support.
LITERATURE CITED1. Till J, McCulloch EA. A direct measurement of the radiation sensitivity
of normal mouse bone marrow cells. Radiat Res 1961;14:1419.2. Becker A, McCulloch E, Till J. Cytological demonstration of the clonal
nature of spleen colonies from transplanted mouse cells. Nature1963;197:452.
3. Potten CS, Lajtha LG. Stem cells versus stem lines. Ann N Y Acad Sci1982;397:49–61.
4. Potten CS, Loeffler M. Stem cells: attributes, cycles, spirals, pitfallsand uncertainties. Lessons for and from the crypt. Development1990;110:1001–1020.
5. Watt FM. Epidermal stem cells: markers, patterning and the control ofstem cell fate. Philos Trans R Soc Lond B Biol Sci 1998;353:831–837.
6. Dermer GB. Basal cell proliferation in benign prostatic hyperplasia.Cancer 1978;41:1857–1862.
7. Bazer GT. Basal cell proliferation and differentiation in regenerationof the rat ventral prostate. Invest Urol 1980;17:470–473.
8. Merchant DJ, Clarke SM, Ives K, Harris S. Primary explant culture: anin vitro model of the human prostate. Prostate 1983;4:523–542.
9. Foster CS, Ke Y. Stem cells in prostatic epithelia. Int J Exp Pathol1997;78:311–329.
10. Bonkhoff H, Remberger K. Differentiation pathways and histogeneticaspects of normal and abnormal prostatic growth: a stem cell model.Prostate 1996;28:98–106.
11. Isaacs JT, Coffey DS. Etiology and disease process of benign prostatichyperplasia. Prostate Suppl 1989;2:33–50.
12. Bonkhoff H, Wernert N, Dhom G, Remberger K. Relation of endo-crine-paracrine cells to cell proliferation in normal, hyperplastic, andneoplastic human prostate. Prostate 1991;19:91–98.
13. Bonkhoff H, Stein U, Remberger K. Endocrine-paracrine cell types inthe prostate and prostatic adenocarcinoma are postmitotic cells. HumPathol 1995;26:167–170.
14. De Marzo AM, Nelson WG, Meeker AK, Coffey DS. Stem cell featuresof benign and malignant prostate epithelial cells. J Urol 1998;160(Pt2):2381–2392.
15. Bonkhoff H, Stein U, Remberger K. The proliferative function of basalcells in the normal and hyperplastic human prostate. Prostate 1994;24:114–118.
16. English HF, Kyprianou N, Isaacs JT. Relationship between DNA frag-mentation and apoptosis in the programmed cell death in the ratprostate following castration. Prostate 1989;15:233–250.
98 BHATT ET AL.

17. Reiter RE, Gu Z, Watabe T, Thomas G, Szigeti K, Davis E, Wahl M,Nisitani S, Yamashiro J, Le Beau MM, Loda M, Witte ON. Prostate stemcell antigen: a cell surface marker overexpressed in prostate cancer.Proc Natl Acad Sci U S A 1998;95:1735–1740.
18. Horoszewicz JS, Murphy GP. An assessment of the current use ofhuman interferons in therapy of urological cancers. J Urol 1989;142:1173–1180.
19. Israeli RS, Powell CT, Corr JG, Fair WR, Heston WD. Expression of theprostate-specific membrane antigen. Cancer Res 1994;54:1807–1811.
20. Silver DA, Pellicer I, Fair WR, Heston WD, Cordon Cardo C. Prostate-specific membrane antigen expression in normal and malignant hu-man tissues. Clin Cancer Res 1997;3:81–85.
21. Walensky LD, Coffey DS, Chen TH, Wu TC, Pasternack GR. A novelM(r) 32,000 nuclear phosphoprotein is selectively expressed in cellscompetent for self-renewal. Cancer Res 1993;53:4720–4726.
22. Kadkol SS, Brody JR, Epstein JI, Kuhajda FP, Pasternack GR. Novelnuclear phosphoprotein pp32 is highly expressed in intermediate-and high-grade prostate cancer. Prostate 1998;34:231–237.
23. Brody JR, Kadkol SS, Mahmoud MA, Rebel JM, Pasternack GR. Iden-tification of sequences required for inhibition of oncogene- mediatedtransformation by pp32. J Biol Chem 1999;274:20053–20055.
24. Liu H, Moy P, Kim S, Xia Y, Rajasekaran A, Navarro V, Knudsen B,Bander NH. Monoclonal antibodies to the extracellular domain ofprostate-specific membrane antigen also react with tumor vascularendothelium. Cancer Res 1997;57:3629–3634.
25. Chang SS, Reuter VE, Heston WD, Bander NH, Grauer LS, Gaudin PB.Five different anti-prostate-specific membrane antigen (PSMA) anti-bodies confirm PSMA expression in tumor-associated neovasculature.Cancer Res 1999;59:3192–3198.
26. Li A, Simmons PJ, Kaur P. Identification and isolation of candidatehuman keratinocyte stem cells based on cell surface phenotype. ProcNatl Acad Sci U S A 1998;95:3902–3907.
27. Shinohara T, Avarbock MR, Brinster RL. beta1- and alpha6-integrin aresurface markers on mouse spermatogonial stem cells. Proc Natl AcadSci U S A 1999;96:5504–5509.
28. Jones PH, Harper S, Watt FM. Stem cell patterning and fate in humanepidermis. Cell 1995;80:83–93.
29. Bonkhoff H, Stein U, Remberger K. Differential expression of alpha 6and alpha 2 very late antigen integrins in the normal, hyperplastic,and neoplastic prostate: simultaneous demonstration of cell surfacereceptors and their extracellular ligands. Hum Pathol 1993;24:243–248.
30. Nagle RB, Knox JD, Wolf C, Bowden GT, Cress AE. Adhesion mole-cules, extracellular matrix, and proteases in prostate carcinoma. J CellBiochem Suppl 1994;19:232–237.
31. Knox JD, Cress AE, Clark V, Manriquez L, Affinito KS, Dalkin BL,Nagle RB. Differential expression of extracellular matrix moleculesand the alpha 6-integrins in the normal and neoplastic prostate. Am JPathol 1994;145:167–174.
32. Lang SH, Clarke NW, George NJ, Testa NG. Primary prostatic epithe-lial cell binding to human bone marrow stroma and the role ofalpha2beta1 integrin. Clin Exp Metastasis 1997;15:218–227.
33. Collins AT, Habib FK, Maitland NJ, Neal DE. Identification and isola-tion of human prostate epithelial stem cells based on alpha(2)beta(1)-integrin expression. J Cell Sci 2001;114(Pt 21):3865–3872.
34. Wernert N, Seitz G, Achtstatter T. Immunohistochemical investiga-tion of different cytokeratins and vimentin in the prostate from thefetal period up to adulthood and in prostate carcinoma. Pathol ResPract 1987;182:617–626.
35. Achtstatter T, Moll R, Moore B, Franke WW. Cytokeratin polypeptidepatterns of different epithelia of the human male urogenital tract:immunofluorescence and gel electrophoretic studies. J HistochemCytochem 1985;33:415–426.
36. Verhagen AP, Aalders TW, Ramaekers FC, Debruyne FM, Schalken JA.Differential expression of keratins in the basal and luminal compart-ments of rat prostatic epithelium during degeneration and regenera-tion. Prostate 1988;13:25–38.
37. Sherwood ER, Berg LA, Mitchell NJ, McNeal JE, Kozlowski JM, Lee C.Differential cytokeratin expression in normal, hyperplastic and ma-lignant epithelial cells from human prostate. J Urol 1990;143:167–171.
38. Okada H, Tsubura A, Okamura A, Senzaki H, Naka Y, Komatz Y, MoriiS. Keratin profiles in normal/hyperplastic prostates and prostate car-cinoma. Virchows Arch A Pathol Anat Histopathol 1992;421:157–161.
39. Cussenot O, Berthon P, Cochand-Priollet B, Maitland NJ, Le Duc A.
Immunocytochemical comparison of cultured normal epithelial pros-tatic cells with prostatic tissue sections. Exp Cell Res 1994;214:83–92.
40. Lajtha LG. Stem cell concepts. Differentiation 1979;14:23–34.41. Fleming WH, Alpern EJ, Uchida N, Ikuta K, Spangrude GJ, Weissman
IL. Functional heterogeneity is associated with the cell cycle status ofmurine hematopoietic stem cells. J Cell Biol 1993;122:897–902.
42. Shapiro HM. Flow cytometric estimation of DNA and RNA content inintact cells stained with Hoechst 33342 and pyronin Y. Cytometry1981;2:143–150.
43. Baum CM, Weissman IL, Tsukamoto AS, Buckle AM, Peault B. Isola-tion of a candidate human hematopoietic stem-cell population. ProcNatl Acad Sci U S A 1992;89:2804–2808.
44. Spangrude GJ, Heimfeld S, Weissman IL. Purification and character-ization of mouse hematopoietic stem cells. Science 1988;241:58–62.
45. Goodell MA, Brose K, Paradis G, Conner AS, Mulligan RC. Isolationand functional properties of murine hematopoietic stem cells that arereplicating in vivo. J Exp Med 1996;183:1797–1806.
46. Bertoncello I, Hodgson GS, Bradley TR. Multiparameter analysis oftransplantable hemopoietic stem cells: I. The separation and enrich-ment of stem cells homing to marrow and spleen on the basis ofrhodamine-123 fluorescence. Exp Hematol 1985;13:999–1006.
47. Pallavicini MG, Summers LJ, Dean PN, Gray JW. Enrichment of mu-rine hemopoietic clonogenic cells by multivariate analyses and sort-ing. Exp Hematol 1985;13:1173–1181.
48. Szilvassy SJ, Lansdorp PM, Humphries RK, Eaves AC, Eaves CJ. Isola-tion in a single step of a highly enriched murine hematopoietic stemcell population with competitive long-term repopulating ability.Blood 1989;74:930–939.
49. Lang SH, Clarke NW, George NJ, Allen TD, Testa NG. Interaction ofprostate epithelial cells from benign and malignant tumor tissue withbone-marrow stroma. Prostate 1998;34:203–213.
50. Ladd AC, Pyatt R, Gothot A, Rice S, McMahel J, Traycoff CM, Srour EF.Orderly process of sequential cytokine stimulation is required foractivation and maximal proliferation of primitive human bone mar-row CD34� hematopoietic progenitor cells residing in G0. Blood1997;90:658–668.
51. Summers YJ, Heyworth CM, de Wynter EA, Chang J, Testa NG. Cordblood G(0) CD34� cells have a thousand-fold higher capacity forgenerating progenitors in vitro than G(1) CD34� cells. Stem Cells2001;19:505–513.
52. Latza U, Niedobitek G, Schwarting R, Nekarda H, Stein H. Ber-EP4:new monoclonal antibody which distinguishes epithelia from me-sothelial. J Clin Pathol 1990;43:213–219.
53. Gaffey MJ, Mills SE, Swanson PE, Zarbo RJ, Shah AR, Wick MR.Immunoreactivity for BER-EP4 in adenocarcinomas, adenomatoid tu-mors, and malignant mesotheliomas. Am J Surg Pathol 1992;16:593–599.
54. Potten CS. Stem cells in gastrointestinal epithelium: numbers, char-acteristics and death. Philos Trans R Soc Lond B Biol Sci 1998;353:821–830.
55. Watt FM. Terminal differentiation of epidermal keratinocytes. CurrOpin Cell Biol 1989;1:1107–1115.
56. Goodell MA, Rosenzweig M, Kim H, Marks DF, DeMaria M, Paradis G,Grupp SA, Sieff CA, Mulligan RC, Johnson RP. Dye efflux studiessuggest that hematopoietic stem cells expressing low or undetectablelevels of CD34 antigen exist in multiple species. Nat Med 1997;3:1337–1345.
57. Huttmann A, Liu SL, Boyd AW, Li CL. Functional heterogeneity withinrhodamine123(lo) Hoechst33342(lo/sp) primitive hemopoietic stemcells revealed by pyronin Y. Exp Hematol 2001;29:1109–1116.
58. Potten CS. Cell replacement in epidermis (keratopoiesis) via discreteunits of proliferation. Int Rev Cytol 1981;69:271–318.
59. Potten CS, Morris RJ. Epithelial stem cells in vivo. J Cell Sci Suppl1988;10:45–62.
60. Potten CS, Booth C, Pritchard DM. The intestinal epithelial stem cell:the mucosal governor. Int J Exp Pathol 1997;78:219–243.
61. Hall PA, Watt FM. Stem cells: the generation and maintenance ofcellular diversity. Development 1989;106:619–633.
62. Xue Y, Smedts F, Verhofstad A, Debruyne F, de la Rosette J, SchalkenJ. Cell kinetics of prostate exocrine and neuroendocrine epitheliumand their differential interrelationship: new perspectives. ProstateSuppl 1998;8:62–73.
63. Robinson EJ, Neal DE, Collins AT. Basal cells are progenitors ofluminal cells in primary cultures of differentiating human prostaticepithelium. Prostate 1998;37:149–160.
99ISOLATION OF PROSTATIC STEM CELLS




















