
Ensemble machine learning on gene expression data for cancer classification
Upload
independentCategory
view
0download
0
Overexpression of aldo-keto reductase 1C2 is associated withdisease progression in patients with prostatic cancer
Kuo-Hsuan Huang,1,2 Shiow-Her Chiou,3 Kuan-Chih Chow,4 Tze-Yi Lin,5 Hui-Wen Chang,5
I-Ping Chiang5 & Meng-Chih Lee1
1Institute of Medicine, Department of Family Medicine, Chung-Shan Medical University, Taichung, 2Section of Urology,
Department of Surgery, Erhlin Branch of Changhua Christian Hospital, Chang-Hua, 3Graduate Institute of Microbiology
and Public Health, 4Graduate Institute of Biomedical Sciences, National Chung Hsing University, and 5Department of
Pathology, China Medical University Hospital, Taichung, Taiwan
Date of submission 18 August 2009Accepted for publication 28 December 2009
Huang K-H, Chiou S-H, Chow K-C, Lin T-Y, Chang H-W, Chiang I-P & Lee M-C
(2010) Histopathology 57, 384–394
Overexpression of aldo-keto reductase 1C2 is associated with disease progression in patientswith prostatic cancer
Aims: Prostatic cancer is resistant to chemotherapy.Expression of aldo-keto reductase 1C (AKR1C) has beenassociated with drug resistance and disease progressionin several cancers. The aim was to investigate therelationship between AKR1C expression and diseaseprogression in prostatic cancer.Methods and results: From January 1996 to December2005, 86 pathological samples were collected frompatients with prostatic cancer. A tissue microarraycontaining 31 prostatic cancers from American patientswas used for comparison between Chinese and Amer-ican patients. Using immunohistochemistry, aldo-ketoreductase family 1, member C2 (AKR1C2) expressionwas assessed in tissue sections. The AKR1C2 wasdetermined by two-dimensional immunoblotting andDNA sequencing of reverse transcriptase-polymerase
chain reaction products. The relationship betweenAKR1C2 expression and clinicopathological variableswas statistically analysed. In vitro, the associationbetween AKR1C2 expression and drug resistance wasinvestigated in androgen-sensitive and androgen-insen-sitive prostatic cancer cells. DNA sequencing and two-dimensional immunoblotting showed that prostaticcancer expressed AKR1C2. It was overexpressed in 77of 86 (89.5%) Chinese and in 28 of 31 (90.3%)American samples. No difference was found in AKR1C2expression between Chinese and American prostaticcancer patients. In vitro, increased expression ofAKR1C2 and prostaglandin F2a correlated with cyto-protection against anticancer drugs and lycopene.Conclusion: Overexpression of AKR1C2 is associatedwith disease progression in prostatic cancer.
Keywords: aldo-keto reductase family 1 – member C2, disease progression, drug resistance, prostatic cancer
Abbreviations: 2-OHBT, 2-hydroxybutyrate; 5-FU, 5-fluorouracil; ADR, adriamycin; AIF, apoptosis-inducing factor;AKR, aldo-keto reductase; AR, androgen receptor; COX-2, cyclooxygenase-2; DDH, dihydrodiol dehydrogenase;EDTA, ethylenediamine tetraaceticacid; EIA, enzyme immunoassay; ELISA, enzyme-linked immunosorbent assay;HGF, hepatocyte growth factor; IEF, isoelectric focusing; IL, interleukin; IPG, immobilized pH gradient; MALDI-TOF,matrix-assisted laser desorption ⁄ ionization-time of flight; MTX, methotrexate; n-BT, n-sodium butyrate; NTPE, non-tumour prostatic epithelium; PAH, polycyclic aromatic hydrocarbons; PBS, phosphate-buffered saline; PG,prostaglandin; PMSF, phenylmethylsulphonylfluoride; RT-PCR, reverse transcriptase-polymerase chain reaction;siRNA, small inhibitory RNA
Address for correspondence: K-C Chow, Graduate Institute of Biomedical Sciences, National Chung Hsing University, 250 Kuo-Kuang Road,
Taichung 40227, Taiwan. e-mail: [email protected]
M-C Lee, Institute of Medicine, Chung-Shan Medical University, Taichung, Taiwan. e-mail: [email protected]
� 2010 Blackwell Publishing Limited.
Histopathology 2010, 57, 384–394. DOI: 10.1111/j.1365-2559.2010.03647.x

Introduction
Prostatic cancer is the second leading cause of cancer-related death in American men, accounting forabout 10% of cancer-related deaths in that popula-tion. The annual mortality rate of prostatic cancer is24.7 per 100 000 men.1 In Taiwan, the incidence ofprostatic cancer has increased threefold and themortality rate has increased fourfold in the past twodecades.2 In 2005, the annual mortality rate ofprostatic cancer was 8.6 per 100 000 men, whichaccounted for about 2.5% of all cancer-related deathsin Taiwan.2 Prostatectomy and androgen deprivationtreatment are adequate for most patients with early-stage disease. For those with locally advanced cancers,radiation therapy achieves effective control. Forpatients with more advanced disease, tumour cellsare resistant to radiation therapy and conventionalchemotherapeutic agents such as adriamycin (ADR),cisplatin, etoposide, 5-fluorouracil (5-FU), methotrex-ate (MTX), mitoxantrone, vincristine and vinblatine.3
The mechanism underlying this spontaneous resis-tance to anticancer drugs, however, is not wellunderstood.
Dihydrodiol dehydrogenase (DDH) 1 [also known asaldo-keto reductase family 1, member C1 (AKR1C1)]and DDH2 [also known as aldo-keto reductase family1, member C2 (AKR1C2)] have been shown to beoverexpressed in lung adenocarcinoma and in ethac-rynic acid-induced drug-resistant human colonic can-cer cells.4,5 Detection of DDH in daunorubicin-resistanthuman gastric cancer cells,6 cisplatin-resistant ovariancancer cells, advanced oesophageal squamous cellcarcinoma, human papillomavirus infection-relateduterine cervical cancers, and arsenic-contaminateddrinking water-associated bladder cancer has furtherindicated that overexpression of AKR1C could beassociated with resistance to chemotherapy as well asdisease progression, tumour recurrence, metastasis,and poor prognosis.7–10
Using reverse transcriptase-polymerase chain reac-tion (RT-PCR), Penning et al.11 showed that aldo-ketoreductase family 1, member C3 (AKR1C3) is abun-dantly expressed in the prostate. However, they did notcharacterize the cell type in the prostate or in theprostatic cancer that expressed AKR1C3. By transfect-ing AKR1C2 or AKR1C3 into androgen-insensitivePC3 cells, Wang et al.12 found that both enzymesmediate biosynthesis of prostaglandin (PG) F2a. Thesestudies suggested that expression of AKR1C might beassociated with prostatic carcinogenesis. Therefore, it isimportant to study whether AKR1C expression is ofclinical significance in patients with prostatic cancer.
In this study, we used an immunohistochemicalmethod and RT-PCR to examine expression of AKR1Cin prostatic cancer specimens. The isoform of AKR1Cwas determined by sequencing of RT-PCR DNA prod-ucts and two-dimensional immunoblotting assay. Thiswas predominantly AKR1C2. The relationship betweenclinicopathological variables and AKR1C2 expressionas well as the prognostic significance of AKR1C2expression in patients with primary untreated prostaticcancer was evaluated statistically. Expression ofAKR1C2 in vitro was investigated in androgen-sensitiveLNCaP cells and androgen-insensitive PC3 and DU145cells.
Materials and methods
prostatic cancer cell lines
AKR1C2 expression in vitro was evaluated in threeprostatic cancer cell lines (PC3, DU145 and LNCaP),which were obtained from American Type CultureCollection A (ATCC, Manassas, VA, USA). Cells weregrown at 37�C in a monolayer in RPMI1640 supple-mented with 10% fetal calf serum, 100 IU ⁄ ml penicil-lin, and 100 lg ⁄ ml streptomycin.
patients and tissue samples
From January 1996 to December 2005, 86 patho-logical samples were collected from patients withclinically diagnosed prostatic cancer. The pathologicalspecimens were collected before hormonal or radia-tion therapy. The Medical Ethics Committee of theChina Medical University Hospital (Taichung,Taiwan) approved the protocol, and written informedconsent to donate specimens was obtained from eachpatient. Gleason score, tumour stage and tumourgrade were determined in accordance with AmericanJoint Committee on Cancer guidelines.13 All patientshad undergone radical prostatectomy with bilateralpelvic lymph node dissection. The diagnosis of pros-tatic cancer was confirmed histopathologically.Patients with lymph node involvement or the pres-ence of tumour-positive margins underwent radiationtherapy with 72–78 Gy directed at the involvedareas. After treatment, all patients were routinelyfollowed up every 3–6 months on an out-patientbasis. Tumour recurrence or metastasis was diag-nosed based on elevated an prostate-specific antigenlevel (biochemical recurrence) and on evidence ofdisease seen on whole-body bone scans or computedtomography scans. A tissue microarray of tumoursamples from 31 American patients with prostatic
AKR1C2 expression in prostatic cancer 385
� 2010 Blackwell Publishing Ltd, Histopathology, 57, 384–394.

cancer (Ambion Inc., Austin, TX, USA) was used tocompare AKR1C2 expression between Chinese andAmerican patients.
immunohistochemistry and immunoblotting
Immunohistochemistry was performed on paraffinsections by an immunoperoxidase method.4,7–10 Fol-lowing incubation with monoclonal antibodies specificto AKR1C1 ⁄ 2, slides were treated with biotin-labelledgoat antimouse immunoglobulin and peroxidase-con-jugated streptavidin and then developed in 3-amino-9-ethylcarbazole. A crimson precipitate was identifiedas immunopositivity. For each immunohistochemicalrun, the non-tumour part of the prostatic tissue servedas the negative control. The immunoblotting assay wasperformed with the same antibodies.4,7–10 Proteinswere separated in a polyacrylamide gel and electro-transferred to a nitrocellulose membrane. The mem-brane was probed with antibodies to AKR1C1 ⁄ 2. Thesignal was visualized by exposing the membrane toan X-Omat film (Eastman Kodak, Rochester, NY,USA) with enhanced chemiluminescent reagent (NEN,Boston, MA, USA).
two-dimensional gel electrophoresis and
immunoblotting
The total cell lysate and tissue lysate of AKR1C+prostatic cancers were first separated by a tube-gelisoelectric focusing (IEF) system (Bio-Rad Laboratories,Inc., Hercules, CA, USA). The prostatic cancer cells orAKR1C+ prostatic cancer tissues were lysed in RIPAbuffer [20 mm Tris–HCl, pH 7.5, 150 mm NaCl, 1 mm
Na2 ethylenediamine tetraaceticacid (EDTA), 1 mm
ethylene glycol tetraaceticacid, 1% NP-40, 1% sodiumdeoxycholate, 2.5 mm sodium pyrophosphate, 1 mm
b-glycerophosphate, 1 mm Na3VO4, 1 lg ⁄ ml leupep-tin] at 4�C for 30 min. Following removal of theinsoluble product, the protein was precipitated withacetone at )20�C overnight and resuspended in thesample buffer [8 m urea, 2% Triton X-100, 40 mm
dithiothreitol, 2% immobilized pH gradient (IPG) buffer(GE Healthcare Life Sciences, Piscataway, NJ, USA)].Rehydration buffer was added to the protein mixture(100 lg) to bring the final volume to 125 ll in the IEFtray, and the solution was covered with an IPG strip.The IPG gel was rehydrated for 12 h and the proteinwas separated on an Ettan� IGPphor� 3 IsoelectricFocusing System (GE Healthcare Life Sciences). The pHranged from 3 to 10. After IEF separation, gels wereextracted from the tubes and equilibrated with sodiumdodecyl sulphate (SDS) buffer before SDS gel electro-
phoresis. One gel was processed for immunoblotting,and one gel was stained with silver nitrate.
rna extraction and reverse transcriptase-
polymerase chain reaction and preparation
of monoclonal antibodies
RNA extraction and gene amplification were performedas described previously.4,7–10 Briefly, cells and tissueswere pulverized in a grinder and then dissolved intoa buffer containing guanidinium. Total RNA wasextracted, precipitated in ethanol and resuspended indistilled water. Following measurement of RNA con-centration in a photometer, cDNA was synthesizedusing RT.4,7–10 An aliquot of cDNA was subjected to35 cycles of PCR using standard procedure denaturingat 94�C for 45 s, hybridizing at 56�C for 30 s andelongating at 72�C for 45 s. The primer sequences forAKR1C1 and AKR1C2 were 5¢-GTGTGAAGCTGAAT-GATGGTCA-3¢ (GenBank, ABO21654, nts 20–41) and5¢-TCTGATGCGCTGCTCATTGTAGCTC-3¢ (nts 834–810). The primer sequences for AKR1C3 were 5¢-TCC-AGAGGTTCCAAGAAGTAAAGCTTT-3¢ (BC001479.2,nts 134–160) and 5¢-TGGATAATTAGGGTGGCTAG-CAAA-3¢ (nts 1010–987). The primer sequences forAKR1C4 were 5¢-TCCAGAGGTTCCGAGGAACAGAGCT-3¢(NM001818.2, nts 111–135) and 5¢-AATGGATAA-TCAGGATGGTCCATA-3¢ (nts 989–966).
The amplified products were analysed on 1% agarosegel and visualized by ethidium bromide staining.Specificity of the 815 base-pair AKR1C2 fragmentswas determined by DNA sequencing (Perkin-Elmer,Foster City, CA, USA). The nucleotide sequences werematched to those in the GenBank database using ablast program (http://www.ncbi.nlm.nih.gov/blast).
The DNA sequence corresponding to the N-terminalfragment of AKR1C2 was amplified by primer se-quences containing Eco RI (sense) and Sal I (antisense)restriction sites, respectively. The primer sequenceswere 5¢-TCCGAATTCATGGACTGTGAAGCTGAATGA-TGGTCA-3¢ (Eco RI site is underlined) and 5¢-TTGTC-GACGTCACTTCTGATGCGCTGCTCATTGTAG-3¢ (Sal Isite is underlined).
The 813-bp cDNA of AKR1C2 was cloned into theexpression vector pET-32b+ (pET32+-AKR1C2; Pro-mega KK, Tokyo, Japan). Bacterial colonies containingthe pET32+-AKR1C2 were selected, and induced byisopropyl-b-d-thiogalactopyranoside to mass-produceAKR1C2. The recombinant protein was purified by anickel-affinity column, and used to immunize BALB ⁄ cmice. Sensitivity of antiserum (OD405 >0.6 at 1:3000dilutions) was measured by enzyme-linked immuno-sorbent assay (ELISA). Specificity of antibodies was
386 K-H Huang et al.
� 2010 Blackwell Publishing Ltd, Histopathology, 57, 384–394.

validated by the appearance of a 36-kDa detected onthe immunoblot of the lung cancer cell extract.Monoclonal antibodies were produced by a hybridomatechnique, and AKR1C-specific antibodies werescreened by the above-mentioned methods. The mono-clonal antibodies recognized both AKR1C1 andAKR1C2. Gene knockdown and immunoblotting of atwo-dimensional gel electrophoresis were used todifferentiate the AKR1C isoform that was expressed inthe prostatic cancer cells.
immunoprecip itation, gel electrophoresis and
protein analysis by matrix-assisted laser
desorption ⁄ ionization-time of fl ight
Total cell lysate was prepared by mixing 5 · 107
cells ⁄ 100 ll phosphate-buffered saline (PBS) with anequal volume of 2 · NP-40 lysis buffer [40 mm Tris–HCl, pH 7.6, 2 mm EDTA, 300 mm NaCl, 2% NP-40,and 2 mm phenylmethylsulphonylfluoride (PMSF)].The protein G sepharose� (Amersham BiosciencesAB, Uppsala, Sweden) was pre-washed before mixingwith 500 lg of total cell lysate. The reaction mixturewas incubated at 4�C for 60 min and centrifuged at800 g for 1 min. The supernatant was reacted with5 lg of purified monoclonal antibodies and 20 ll offresh protein G sepharose at 4�C for 18 h. The reactionmixture was centrifuged at 800 g for 1 min. Theprecipitate was washed with 1· PBS and dissolved inloading buffer (50 mm Tris, pH 6.8, 150 mm NaCl,1 mm disodium EDTA, 1 mm PMSF, 10% glycerol, 5%b-mercaptoethanol, 0.01% bromophenol blue and1% SDS). Electrophoresis was carried out on two 10%polyacrylamide gels with 4.5% stacking. One gel wasprocessed for immunoblotting, and the other gel wasstained with Coomassie blue.4 Protein bands on theCoomassie-stained gel, which corresponded to theimmunoblotting-positive bands, were extracted fromthe gel for identification by matrix-assisted laserdesorption ⁄ ionization-time of flight (MALDI-TOF) ona Voyager-DE� pro biospectrometry workstation(Applied Biosystems, Milpitas, CA, USA). Fragments ofpeptide fingerprints were matched with those onthe SwissProt database by MS-fit (ProteinProspector4.0.5, The Regents of the University of California).After electrophoresis, proteins on the first gel weretransferred to a nitrocellulose membrane for immuno-blotting.
enforced expression of akr 1 c 2
DNA sequences corresponding to the full-lengthAKR1C2 were amplified by primer sequences containing
Eco RI (sense) and Not I (antisense) restriction sites. Theprimer sequences were 5¢-TTGGAATTCATTATGGAT-TCGAAATACCAGTGT-3¢ (nts 23–43, BC_007024; theEco RI site is underlined and the initiation codon ishighlighted in bold) and 5¢-CTAGAGCGGCCGCTCCA-TGTTAATATTCATCAGAAAATGGA-3¢ (nts 994–973,BC_007024; the Not I site is underlined and the stopcodon is highlighted in bold). The 972-bp cDNA of thefull-length AKR1C2 was cloned into pcDNA�3.1 ⁄ myc-His (Invitrogen, Taipei, Taiwan) (pAKR1C2), which wasintroduced into cells by jetPEI� (Polyplus-transfectionInc., New York, NY, USA).
inhib it ion of akr 1 c 2 express ion by small
inhib itory rna
Inhibition of gene expression was performed using thesmall inhibitory (si) RNA method. The target sequencesfor AKR1C2 (underlined) were inserted into retroviralvector pSIREN-RetroQ (Clontech, Mountain View, CA,USA): 5¢-GATCCGATTGCAGATGGCAGTGTGTTCAAG-AGACACACTGCCATCTGCAATCTTTTTTCAGCGTG-3¢and 5¢-AATTCACGCGTAAAAAAGATTGCAGATGGC-AGTGTGTCTCTTGAACACACTGCCATCTGCAATCG-3¢with spacer (boldfaced).
cytotoxicity assay
Cells were seeded at 1000, 2500 and 5000 cells ⁄ well18 h prior to drug challenge. The cells were treatedcontinuously with various concentrations of drug for72 h. The control group was only treated with drugdiluent, PBS or dimethylsulphoxide. Culture mediawere replaced with PBS before addition of 10 ll ofWST-1 (BioVision, Mountain View, CA, USA) andincubation was continued for 4 h. Percent survival ofcells was quantified by comparing the number of viablecells in the experiment group with that in the controlgroup. All procedures were performed in triplicate.9
enzyme immunoassay
PGE2 and PGF2a levels were determined by enzymeimmunoassay (EIA) with a PGE2 or PGF2a HighSensitivity EIA Kit (Assay Designs, Inc., Ann Arbor,MI, USA). Absorbance at 405 nm was measured by anELISA reader (Sunrise�; TECAN, Grodig, Austria). Allsamples were run in duplicate.
slide evaluation
Non-tumour prostatic tissue served as the internalnegative control. Slides were evaluated by three
AKR1C2 expression in prostatic cancer 387
� 2010 Blackwell Publishing Ltd, Histopathology, 57, 384–394.
independent pathologists blinded to the clinicopatholog-ical variables. An immunoreactivity scoring system wasadapted for this study.14 A specimen was consideredstrongly positive if >25% of cancer cells were immuno-positive; negative if <25% were immunopositive.
statistical analysis
Associations between DDH expression and clinicopath-ological variables were analysed by chi-squared test.The chi-squared test for trend was used when corre-sponding variables exceeded two categories. Statisticalsignificance was set at P < 0.05. A t-test was used todetermine the statistical difference of PGE2 or PGF2aconcentrations between groups. Analysis of covarianceand the Mantel–Haenszel chi-squared procedure wereused to evaluate the statistical difference of cytotoxicitycurves among the various groups. All statistical anal-yses were performed by GraphPad Prism5 (Version X;GraphPad Software, Inc., La Jolla, CA, USA).
Results
characterization of monoclonal antibodies
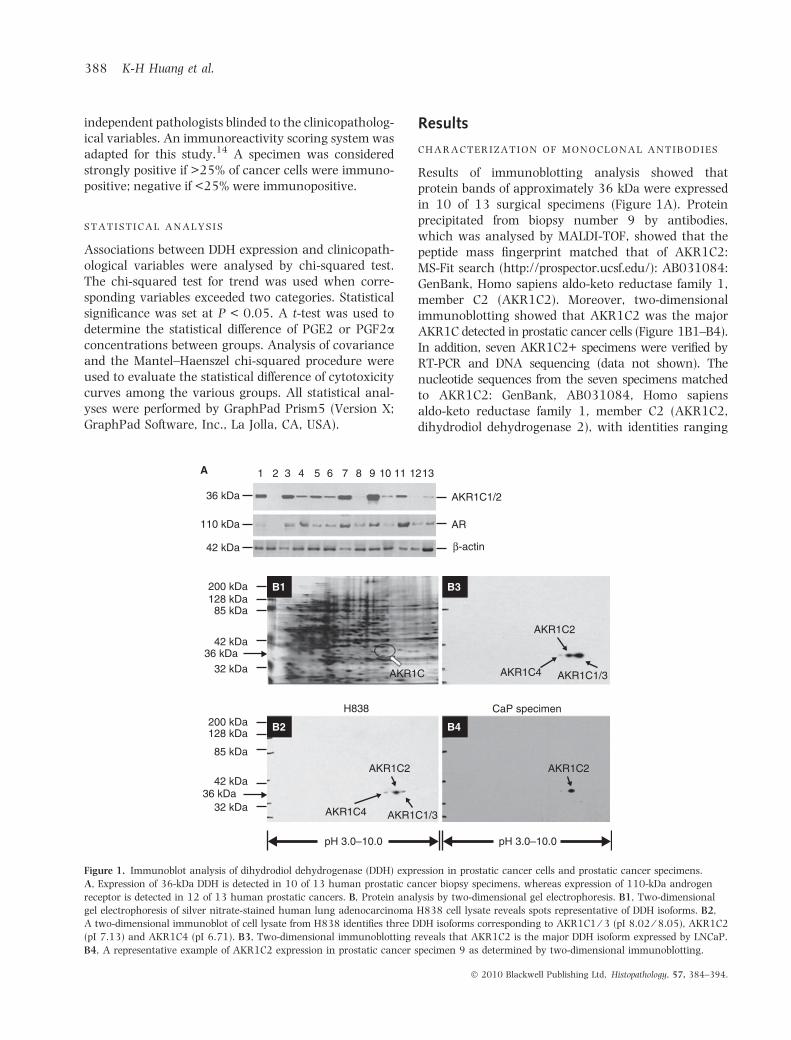
Results of immunoblotting analysis showed thatprotein bands of approximately 36 kDa were expressedin 10 of 13 surgical specimens (Figure 1A). Proteinprecipitated from biopsy number 9 by antibodies,which was analysed by MALDI-TOF, showed that thepeptide mass fingerprint matched that of AKR1C2:MS-Fit search (http://prospector.ucsf.edu/): AB031084:GenBank, Homo sapiens aldo-keto reductase family 1,member C2 (AKR1C2). Moreover, two-dimensionalimmunoblotting showed that AKR1C2 was the majorAKR1C detected in prostatic cancer cells (Figure 1B1–B4).In addition, seven AKR1C2+ specimens were verified byRT-PCR and DNA sequencing (data not shown). Thenucleotide sequences from the seven specimens matchedto AKR1C2: GenBank, AB031084, Homo sapiensaldo-keto reductase family 1, member C2 (AKR1C2,dihydrodiol dehydrogenase 2), with identities ranging
1 2 3 4 5 6 7 8 9 10 11 1213
36 kDa
110 kDa
42 kDa
200 kDa128 kDa85 kDa
42 kDa36 kDa
32 kDa
200 kDa128 kDa
85 kDa
42 kDa36 kDa
32 kDa
AKR1C1/2
AR
β-actin
AKR1C
AKR1C2
AKR1C4 AKR1C1/3
AKR1C2AKR1C2
AKR1C4 AKR1C1/3
H838 CaP specimen
pH 3.0–10.0 pH 3.0–10.0
A
B1 B3
B4B2
Figure 1. Immunoblot analysis of dihydrodiol dehydrogenase (DDH) expression in prostatic cancer cells and prostatic cancer specimens.
A, Expression of 36-kDa DDH is detected in 10 of 13 human prostatic cancer biopsy specimens, whereas expression of 110-kDa androgen
receptor is detected in 12 of 13 human prostatic cancers. B, Protein analysis by two-dimensional gel electrophoresis. B1, Two-dimensional
gel electrophoresis of silver nitrate-stained human lung adenocarcinoma H838 cell lysate reveals spots representative of DDH isoforms. B2,
A two-dimensional immunoblot of cell lysate from H838 identifies three DDH isoforms corresponding to AKR1C1 ⁄ 3 (pI 8.02 ⁄ 8.05), AKR1C2
(pI 7.13) and AKR1C4 (pI 6.71). B3, Two-dimensional immunoblotting reveals that AKR1C2 is the major DDH isoform expressed by LNCaP.
B4, A representative example of AKR1C2 expression in prostatic cancer specimen 9 as determined by two-dimensional immunoblotting.
388 K-H Huang et al.
� 2010 Blackwell Publishing Ltd, Histopathology, 57, 384–394.
from 99.6% to 100%. No mutation was found inAKR1C2 from prostatic cancer specimens.
express ion of akr 1 c 2 and androgen receptor
in prostatic cancer
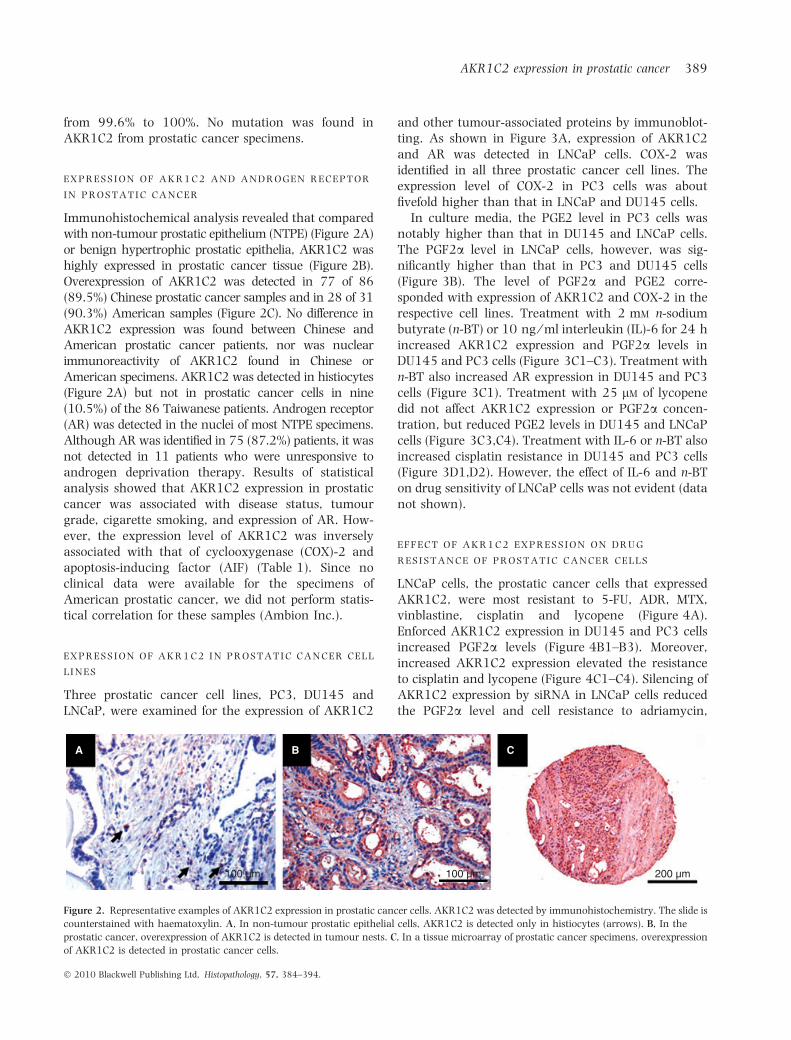
Immunohistochemical analysis revealed that comparedwith non-tumour prostatic epithelium (NTPE) (Figure 2A)or benign hypertrophic prostatic epithelia, AKR1C2 washighly expressed in prostatic cancer tissue (Figure 2B).Overexpression of AKR1C2 was detected in 77 of 86(89.5%) Chinese prostatic cancer samples and in 28 of 31(90.3%) American samples (Figure 2C). No difference inAKR1C2 expression was found between Chinese andAmerican prostatic cancer patients, nor was nuclearimmunoreactivity of AKR1C2 found in Chinese orAmerican specimens. AKR1C2 was detected in histiocytes(Figure 2A) but not in prostatic cancer cells in nine(10.5%) of the 86 Taiwanese patients. Androgen receptor(AR) was detected in the nuclei of most NTPE specimens.Although AR was identified in 75 (87.2%) patients, it wasnot detected in 11 patients who were unresponsive toandrogen deprivation therapy. Results of statisticalanalysis showed that AKR1C2 expression in prostaticcancer was associated with disease status, tumourgrade, cigarette smoking, and expression of AR. How-ever, the expression level of AKR1C2 was inverselyassociated with that of cyclooxygenase (COX)-2 andapoptosis-inducing factor (AIF) (Table 1). Since noclinical data were available for the specimens ofAmerican prostatic cancer, we did not perform statis-tical correlation for these samples (Ambion Inc.).
express ion of akr 1 c 2 in prostatic cancer cell
lines
Three prostatic cancer cell lines, PC3, DU145 andLNCaP, were examined for the expression of AKR1C2
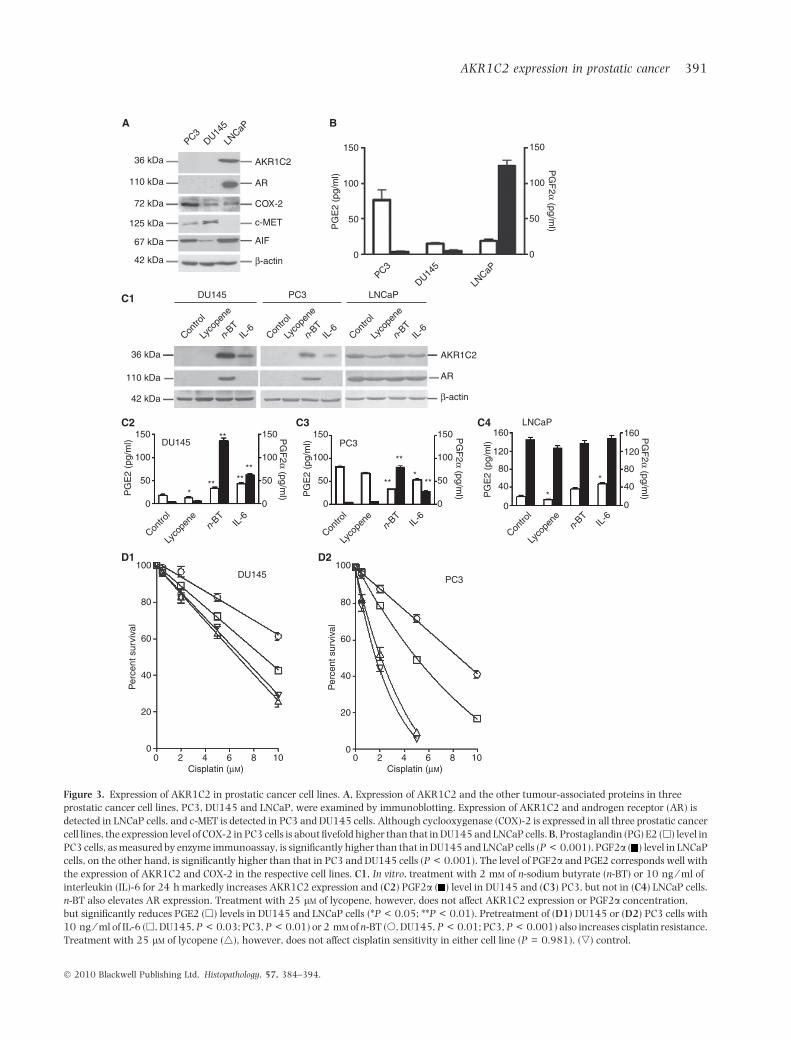
and other tumour-associated proteins by immunoblot-ting. As shown in Figure 3A, expression of AKR1C2and AR was detected in LNCaP cells. COX-2 wasidentified in all three prostatic cancer cell lines. Theexpression level of COX-2 in PC3 cells was aboutfivefold higher than that in LNCaP and DU145 cells.
In culture media, the PGE2 level in PC3 cells wasnotably higher than that in DU145 and LNCaP cells.The PGF2a level in LNCaP cells, however, was sig-nificantly higher than that in PC3 and DU145 cells(Figure 3B). The level of PGF2a and PGE2 corre-sponded with expression of AKR1C2 and COX-2 in therespective cell lines. Treatment with 2 mm n-sodiumbutyrate (n-BT) or 10 ng ⁄ ml interleukin (IL)-6 for 24 hincreased AKR1C2 expression and PGF2a levels inDU145 and PC3 cells (Figure 3C1–C3). Treatment withn-BT also increased AR expression in DU145 and PC3cells (Figure 3C1). Treatment with 25 lm of lycopenedid not affect AKR1C2 expression or PGF2a concen-tration, but reduced PGE2 levels in DU145 and LNCaPcells (Figure 3C3,C4). Treatment with IL-6 or n-BT alsoincreased cisplatin resistance in DU145 and PC3 cells(Figure 3D1,D2). However, the effect of IL-6 and n-BTon drug sensitivity of LNCaP cells was not evident (datanot shown).
effect of akr1 c 2 express ion on drug
resistance of prostatic cancer cells
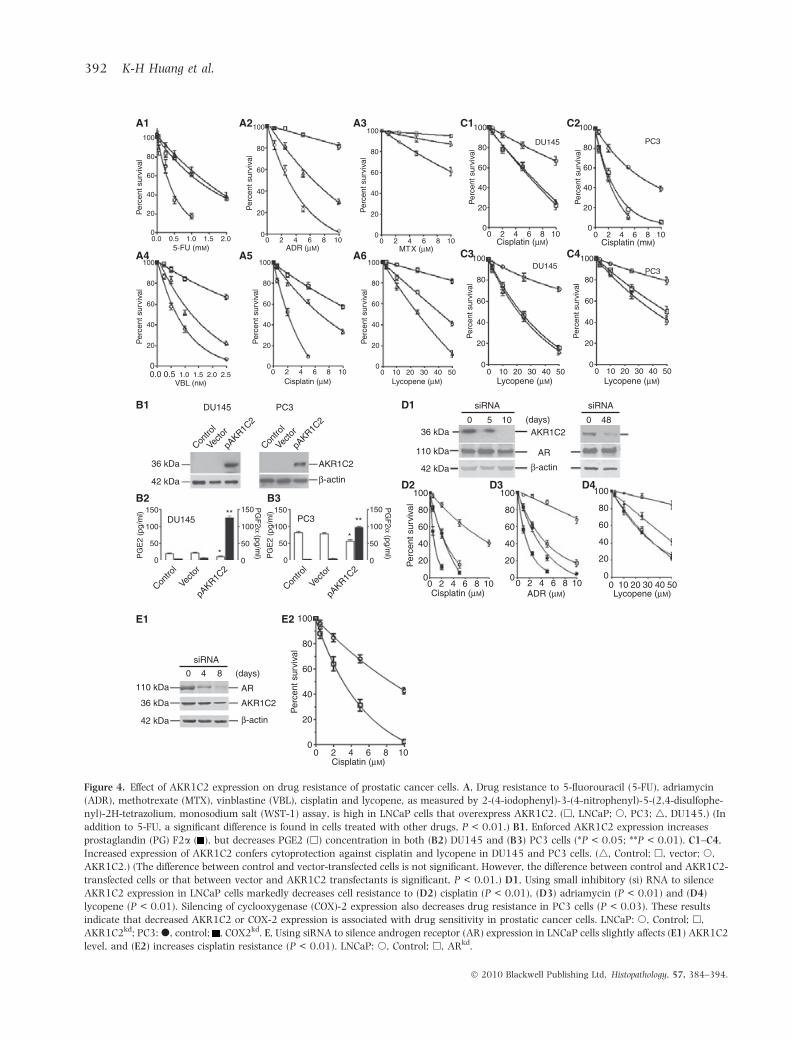
LNCaP cells, the prostatic cancer cells that expressedAKR1C2, were most resistant to 5-FU, ADR, MTX,vinblastine, cisplatin and lycopene (Figure 4A).Enforced AKR1C2 expression in DU145 and PC3 cellsincreased PGF2a levels (Figure 4B1–B3). Moreover,increased AKR1C2 expression elevated the resistanceto cisplatin and lycopene (Figure 4C1–C4). Silencing ofAKR1C2 expression by siRNA in LNCaP cells reducedthe PGF2a level and cell resistance to adriamycin,
A B C
100 µm 100 µm 200 µm
Figure 2. Representative examples of AKR1C2 expression in prostatic cancer cells. AKR1C2 was detected by immunohistochemistry. The slide is
counterstained with haematoxylin. A, In non-tumour prostatic epithelial cells, AKR1C2 is detected only in histiocytes (arrows). B, In the
prostatic cancer, overexpression of AKR1C2 is detected in tumour nests. C, In a tissue microarray of prostatic cancer specimens, overexpression
of AKR1C2 is detected in prostatic cancer cells.
AKR1C2 expression in prostatic cancer 389
� 2010 Blackwell Publishing Ltd, Histopathology, 57, 384–394.

cisplatin and lycopene (Figure 4D1–D4). DecreasedAKR1C2 expression did not influence AR level. Silenc-ing of AR expression, on the other hand, reducedAKR1C2 level and cisplatin resistance in LNCaP cells(Figure 4E1,E2). These results indicate that increasedAKR1C2 expression and PGF2a level are associatedwith drug resistance in prostatic cancer cells.
Discussion
Our results have shown that AKR1C2 is the majorisoform of AKR1C, which is overexpressed in patients
with prostatic cancer. Expression of AKR1C2 is asso-ciated with disease status and tumour grade as wellas expression of AR and cigarette smoking. In vitro,AKR1C2 expression is associated with an increasedPGF2a level as well as resistance to anticancer drugsand lycopene in prostatic cancer cells. These data areconsistent with our prior studies in lung, oesophageal,ovarian, uterine cervical and bladder cancers.4,7–10
Cigarette smoking is frequently associated withcarcinogenesis, disease manifestation, tissue invasionand metastasis of prostatic cancer.15,16 Plaskon et al.17
suggested that while the content of cadmium in
Table 1. Association ofAKR1C2 expression withclinicopathologicalparameters in patients withprostatic cancer in Taiwan
Clinicopathological parameters(number of patients)
Expression of AKR1C2
P-valueLow (n = 9) High (n = 77)
Age (48–83 years) 70.4 ± 3.8 66.1 ± 4.5 0.120*
Disease statusLocalized (stage A or B) (n = 54) 3 51 0.010†
Advanced (Stage C or D) (n = 21) 2 19
Undetermined (n = 11) 4 7
Tumour gradeGleason <7 or well or moderatelydifferentiated (n = 59)
7 52 0.025†
Gleason ‡7 or poorlydifferentiated (n = 22)
0 22
Undetermined (n = 5) 2 3
Cigarette smokingSmoker (n = 65) 3 62 0.006‡
Non-smoker (n = 21) 6 15
Expression of androgen receptorHigh (n = 75) 2 73 <0.001‡
Low (n = 11) 7 4
Expression of cyclooxygenase-2High (n = 21) 5 16 0.036‡
Low (n = 65) 4 61
Expression of AIFHigh (n = 23) 8 15 <0.001‡
Low (n = 63) 1 62
A smoker was defined as a person who smoked more than 20 packs per year.
*Two-sided P-value determined by Mann–Whitney test.
†Two-sided P-value determined by chi-squared test for trend.
‡Two-sided P-value determined by chi-squared test (Fisher’s exact test was used when thenumber in one of the cells was £ 5).
AIF, apoptosis-inducing factor.
390 K-H Huang et al.
� 2010 Blackwell Publishing Ltd, Histopathology, 57, 384–394.
A
C1
D1
36 kDa AKR1C2
150
PG
E2
(pg/
ml)
100
50
0
DU145 PC3
100
80
60
40
Per
cent
sur
viva
l
20
0
100
80
60
40
Per
cent
sur
viva
l
20
00 2 4 6
Cisplatin (μM)8 10 0 2 4 6
Cisplatin (μM)8 10
150
PG
F2α
(pg/ml)
100
50
0
AR
COX-2
c-MET
AIF
β-actin
AKR1C2
AR
β-actin
PC3
Contro
l
Lyco
pene
n-BT
IL-6
Contro
l
Lyco
pene
n-BT
IL-6
Contro
l
Lyco
pene
n-BT
IL-6
PC3
PC3
DU145
DU145
LNCaP
DU145 LNCaPLN
CaP
110 kDa
72 kDa
36 kDa
110 kDa
42 kDa
125 kDa
67 kDa
42 kDa
D2
C2150 **
****
** ***
*
******
DU145 PC3
LNCaP
PG
E2
(pg/
ml)
PG
E2
(pg/
ml)
PG
E2
(pg/
ml)
PG
F2α
(pg/ml)
PG
F2α
(pg/ml)
PG
F2α
(pg/ml)
100
50
0
150
100
50
0
150
100
50
0
150 160
120
80
40
0
160
120
80
40
0
100
50
0
Contro
l
Lyco
pene
n-BT
IL-6
Contro
l
Lyco
pene
n-BT
IL-6
Contro
l
Lyco
pene
n-BT
IL-6
C3 C4
B
Figure 3. Expression of AKR1C2 in prostatic cancer cell lines. A, Expression of AKR1C2 and the other tumour-associated proteins in three
prostatic cancer cell lines, PC3, DU145 and LNCaP, were examined by immunoblotting. Expression of AKR1C2 and androgen receptor (AR) is
detected in LNCaP cells, and c-MET is detected in PC3 and DU145 cells. Although cyclooxygenase (COX)-2 is expressed in all three prostatic cancer
cell lines, the expression level of COX-2 in PC3 cells is about fivefold higher than that in DU145 and LNCaP cells. B, Prostaglandin (PG) E2 (h) level in
PC3 cells, as measured by enzyme immunoassay, is significantly higher than that in DU145 and LNCaP cells (P < 0.001). PGF2a ( ) level in LNCaP
cells, on the other hand, is significantly higher than that in PC3 and DU145 cells (P < 0.001). The level of PGF2a and PGE2 corresponds well with
the expression of AKR1C2 and COX-2 in the respective cell lines. C1, In vitro, treatment with 2 mm of n-sodium butyrate (n-BT) or 10 ng ⁄ ml of
interleukin (IL)-6 for 24 h markedly increases AKR1C2 expression and (C2) PGF2a ( ) level in DU145 and (C3) PC3, but not in (C4) LNCaP cells.
n-BT also elevates AR expression. Treatment with 25 lm of lycopene, however, does not affect AKR1C2 expression or PGF2a concentration,
but significantly reduces PGE2 (h) levels in DU145 and LNCaP cells (*P < 0.05; **P < 0.01). Pretreatment of (D1) DU145 or (D2) PC3 cells with
10 ng ⁄ ml of IL-6 (h, DU145, P < 0.03; PC3, P < 0.01) or 2 mm of n-BT (s, DU145, P < 0.01; PC3, P < 0.001) also increases cisplatin resistance.
Treatment with 25 lm of lycopene (4), however, does not affect cisplatin sensitivity in either cell line (P = 0.981). (,) control.
AKR1C2 expression in prostatic cancer 391
� 2010 Blackwell Publishing Ltd, Histopathology, 57, 384–394.
DU145 PC3
Contro
l
Vecto
r
pAKR1C
2
Contro
l
Vecto
r
pAKR1C
2
Contro
l
Vecto
r
pAKR1C
2
Contro
l
Vecto
r
pAKR1C
2
36 kDa
42 kDa
AKR1C2
β-actin
150DU145
PG
E2
(pg/
ml)
100
50
0
150
PG
E2
(pg/
ml)
100
50
0
PG
F2α
(pg/ml)
150****
*
*
100
50
0
PG
F2α
(pg/ml)
150
100
50
0
PC3
ADR (μM) Lycopene (μM)
36 kDa
siRNA siRNA
0 0(days)5 10 48
110 kDa
42 kDa
AKR1C2
AR
β-actin
100
80
60
Per
cent
sur
viva
l
40
20
00 2 4 6 8 10
100
80
60
40
20
0
100
80
60
40
20
00 10 20 30 40 500 2 4 6 8 10
Cisplatin (μM)
100
80
60
Per
cent
sur
viva
l
40
20
00.0 0.5
5-FU (mM) ADR (μM)1.0 1.5 2.0
100
80
60
Per
cent
sur
viva
l
40
20
0
100
80
60
Per
cent
sur
viva
l
40
20
0
100
80
60
Per
cent
sur
viva
l
40
20
00 10 20 30 40 500.0 0.5
VBL (nM)1.0 1.5 2.0 2.5
100
80
60
Per
cent
sur
viva
l
40
20
00 2 4 6 8 10
Cisplatin (μM) Lycopene (μM)0 2 4 6 8 10
MTX (μM)0 2 4 6 8 10
100
80
60
Per
cent
sur
viva
l
40
20
0
100
80
60
Per
cent
sur
viva
l
40
20
0
DU145
DU145
PC3
PC3
0 2 4 6 8 10Cisplatin (μM)
Lycopene (μM)
100
80
60
40
20
00 10 20 30 40 50
100
80
60
Per
cent
sur
viva
l
Per
cent
sur
viva
l
Lycopene (μM)
100
80
60
40
20
00 10 20 30 40 50
Per
cent
sur
viva
l
40
20
00
2 4 6 8 10Cisplatin (mM)
siRNA
0 (days)4 8
100
80
60
Per
cent
sur
viva
l
40
20
00 2 4 6 8 10
Cisplatin (μM)
36 kDa
110 kDa
42 kDa
AKR1C2
AR
β-actin
A1 A2 A3
A4 A5 A6
C1 C2
C3 C4
B1
B2 B3
E1 E2
D1
D2 D3 D4
Figure 4. Effect of AKR1C2 expression on drug resistance of prostatic cancer cells. A, Drug resistance to 5-fluorouracil (5-FU), adriamycin
(ADR), methotrexate (MTX), vinblastine (VBL), cisplatin and lycopene, as measured by 2-(4-iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophe-
nyl)-2H-tetrazolium, monosodium salt (WST-1) assay, is high in LNCaP cells that overexpress AKR1C2. (h, LNCaP; s, PC3; 4, DU145.) (In
addition to 5-FU, a significant difference is found in cells treated with other drugs, P < 0.01.) B1, Enforced AKR1C2 expression increases
prostaglandin (PG) F2a ( ), but decreases PGE2 (h) concentration in both (B2) DU145 and (B3) PC3 cells (*P < 0.05; **P < 0.01). C1–C4,
Increased expression of AKR1C2 confers cytoprotection against cisplatin and lycopene in DU145 and PC3 cells. (4, Control; h, vector; s,
AKR1C2.) (The difference between control and vector-transfected cells is not significant. However, the difference between control and AKR1C2-
transfected cells or that between vector and AKR1C2 transfectants is significant, P < 0.01.) D1, Using small inhibitory (si) RNA to silence
AKR1C2 expression in LNCaP cells markedly decreases cell resistance to (D2) cisplatin (P < 0.01), (D3) adriamycin (P < 0.01) and (D4)
lycopene (P < 0.01). Silencing of cyclooxygenase (COX)-2 expression also decreases drug resistance in PC3 cells (P < 0.03). These results
indicate that decreased AKR1C2 or COX-2 expression is associated with drug sensitivity in prostatic cancer cells. LNCaP: s, Control; h,
AKR1C2kd; PC3: d, control; , COX2kd. E, Using siRNA to silence androgen receptor (AR) expression in LNCaP cells slightly affects (E1) AKR1C2
level, and (E2) increases cisplatin resistance (P < 0.01). LNCaP: s, Control; h, ARkd.
392 K-H Huang et al.
� 2010 Blackwell Publishing Ltd, Histopathology, 57, 384–394.

cigarette smoke was responsible for prostate carcino-genesis, cigarette smoking-induced elevation of testos-terone accounted for the progression of prostaticcancer. Based on a statistical correlation betweentumour progression and overexpression of AKR1C2,Chiang et al.18 indicated that cigarette smoke inducedAKR1C2 expression in cancer cells. Their finding wassupported by data from Lou et al.19 that phase IIinducers, derivatives of polycyclic aromatic hydrocar-bons (PAH) in cigarette smoke, induced AKR1C2 andAKR1C1, but not AKR1C3, expression by binding toantioxidant response element and NF-E2-related factor-2. Rizner et al.20 found that AKR1C2, which washighly expressed in prostatic cancer, was not detectedin NTPE. We have confirmed their findings by immu-nohistochemistry, two-dimensional gel immunoblot-ting and DNA sequence analyses. Moreover, our resultshave shown that the level of AR affected drugresistance and expression of AKR1C2, which was up-regulated in vitro by IL-6 and n-sodium butyrate.21,22
The level of AKR1C2, on the other hand, did not affectexpression of AR in prostatic cancer cells.
In fact, the serum level of n-BT, a short-chain fattyacid by-product of colonic fermentation of dietary fibresby anaerobic bacteria, frequently increased in cancerpatients, and n-BT induced IL-6 expression in tumourcells.21 Our results suggest a relationship between hostimmune surveillance, tumour cell growth, and intes-tinal flora, which are symbiotic with the host only if thehost immune surveillance is capable of managingbacterial growth balance in the lower gut. The serumbutyrate level could, therefore, reflect the balance ofintestinal flora and the competence of a patient’simmune surveillance. Recently, a metabolic study bySreekumar et al.23 has shown that the 2-hydroxybu-tyrate (2-OHBT) level is correlated with prostaticcancer progression and metastasis. Since the glutathi-one level in cancer patients decreases with diseaseprogression,11 increased serum 2-OHBT must be fromother sources, such as hydroxylation of n-BT. Althoughconversion to 2-OHBT remains to be determined, theirdata support our results that patients with advanced-stage cancer, in particular those who have developedcachexia, often present with severe weight loss as wellas elevation of urinary butyrate and serum IL-6,21 andthe persistently increased serum butyrate level is from acontinuous source, e.g. intestinal flora.21,22
It is therefore worth noting that increased serum IL-6is frequently detected in patients with late-stage pros-tatic cancer, and that serum IL-6 levels serve as asurvival biomarker among these patients.24 By showingthat AKR1C2 is expressed in AR+ LNCaP, but not in IL-6-secreting PC3 or DU145 cells, our data suggested that
AKR1C2 expression is directly associated with the ARlevel in prostatic cancer. Cigarette smoking also inducesAKR1C2 expression, which in turn catabolyses PAHinto o-quinone derivatives to damage macromolecules,such as DNA, RNA and proteins, and to introduce localinflammation.25 By showing that NS398, a COX-2inhibitor, wogonin, an anti-inflammatory ingredientfrom a Chinese herb (Scutellaria baicalensis), and 4-hydroxyacetophenone inhibits endogenous PGF2a syn-thase activity of AKR1C2,12,18,26 our data support thefact that chronic local inflammation is associated withprostatic carcinogenesis,11,12,27 and, possibly, diseaseprogression. The impact of AKR1C2 and the direct effectof PGF2a on the manifestations of prostatic cancer,however, are yet to be determined.
Based on the similarity in chemical structure betweenPAH derivatives and anticancer drugs, e.g. daunorubi-cin, doxorubicin and mitoxantrone, we have proposedthat DDH plays a role in drug inactivation, and henceincreases drug resistance in tumour cells.4–10 Althoughno direct drug inactivation activity was found inAKR1C2, its expression was frequently associated withmicrosomal epoxide hydrolase, which directly inacti-vates daunorubicin and doxorubicin.28 The results ofsiRNA studies, considered together with those of ectopicgene expression, clearly indicate that the AKR1C2level is associated with increased resistance to antime-tabolites, anthracyclines, DNA alkylating agents, andlycopene in patients with prostatic cancer. Recently,Chiang has shown that in AKR1C2+ cells, hypoxiaincreases PGF2a biosynthesis to activate expression ofhepatocyte growth factor (HGF) and IL-8. These datacorrespond well with those presented by Chen et al. thatcigarette smoking increased HGF expression inAKR1C2+ lung adenocarcinoma cells.4,29 Moreover,overexpression of HGF reduced AIF expression and thedrug sensitivity of lung cancer cells.18,29 By showingthat expression of AKR1C2 inversely correlated withthat of AIF in pathological specimens of prostaticcancer, our data support their observations. AlthoughIL-8 does not directly mediate tumour cell migration orinvasion, it does increase expression of vascular endo-thelial growth factor29 to induce angiogenesis andincrease expression of matrix metalloproteinases tofacilitate tumour invasion and metastasis.30 However,since only three prostatice cancer cell lines wereavailable, and only one cell line expressed both ARand AKR1C2, more in vitro data are required to confirmour observations.
In this study, we provide a clue for the spontaneousresistance of prostatic cancer to anticancer drugs andlycopene. We have shown that AKR1C2 expression inprostatic cancer is associated with drug resistance,
AKR1C2 expression in prostatic cancer 393
� 2010 Blackwell Publishing Ltd, Histopathology, 57, 384–394.

which in turn reflects poor prognosis. In vitro, we haveprovided evidence to support the clinical correlationbetween AR and AKR1C2, in which resistance toanticancer drugs in prostatic cancer could be mediatedby the enzyme overexpression. AR expression couldthen act as a biomarker for sensitivity to androgentherapy, androgen ablation and chemotherapeuticregimens. In conclusion, our results show that overex-pression of AKR1C2 is associated with drug resistanceand, possibly, disease progression in prostatic cancer.
Acknowledgements
This study was supported in part by a grant fromthe Comprehensive Academic Promotion Projects(NCHU975014).
References
1. Jemal A, Siegel R, Ward E, Murray T, Xu J, Thun MJ. Cancer
statistics, 2007. CA Cancer J. Clin. 2007; 57; 43–66.
2. Annual reports of the Department of Health, the Executive Yuan,
Republic of China, 2006.
3. Simoneau AR. Treatment- and disease-related complications of
prostate cancer. Rev. Urol. 2006; 8; S56–S67.
4. Hsu NY, Ho HC, Chow KC et al. Overexpression of dihydrodiol
dehydrogenase as a prognostic marker of non-small cell lung
cancer. Cancer Res. 2001; 61; 2727–2731.
5. Shen H, Kauvar L, Tew KD. Importance of glutathione and asso-
ciated enzymes in drug response. Oncol. Res. 1997; 9; 295–302.
6. Ax W, Soldan M, Koch L, Maser E. Development of daunorubicin
resistance in tumour cells by induction of carbonyl reduction.
Biochem. Pharmacol. 2000; 59; 293–300.
7. Deng HB, Parekh HK, Chow KC, Simpkins H. Increased expres-
sion of dihydrodiol dehydrogenase induces resistance to cisplatin
in human ovarian carcinoma cells. J. Biol. Chem. 2002; 277;
15035–15043.
8. Wang LS, Chow KC, Wu YC, Hsu NY, Lin TY. Inverse expressions
of dihydrodiol dehydrogenase and glutathione-S-transferase in
patients with esophageal squamous cell carcinoma. Int. J. Cancer
2004; 111; 246–251.
9. Ueda M, Hung YC, Chen JT et al. Infection of human papilloma-
virus and overexpression of dihydrodiol dehydrogenase in uterine
cervical cancer. Gynecol. Oncol. 2006; 102; 173–181.
10. Tai HL, Lin TS, Chiou SH et al. Overexpression of aldo-keto
reductase 1C2 as a high-risk factor in bladder cancer. Oncol. Rep.
2007; 17; 305–311.
11. Penning TM, Burczynski ME, Jez JM et al. Human 3alpha-
hydroxysteroid dehydrogenase isoforms (AKR1C1-AKR1C4) of
the aldo-keto reductase superfamily: functional plasticity and
tissue distribution reveals roles in the inactivation and formation of
male and female sex hormones. Biochem. J. 2000; 351; 67–77.
12. Wang S, Yang Q, Fung KM, Lin HK. AKR1C2 and AKR1C3
mediated prostaglandin D2 metabolism augments the PI3K ⁄ Akt
proliferative signaling pathway in human prostate cancer cells.
Mol. Cell. Endocrinol. 2008; 289; 60–66.
13. Greene FL, Page DL, Fleaming ID et al. American Joint Committee
on Cancer, Manual for staging cancer, 6th edn. New York, NY:
Springer, 2002.
14. Remmele W, Schicketanz KH. Immunohistochemical determina-
tion of estrogen and progesterone receptor content in human
breast cancer. Computer-assisted image analysis (QIC score) vs.
subjective grading (IRS). Pathol. Res. Pract. 1993; 189; 862–866.
15. Coughlin SS, Neaton JD, Sengupta A. Cigarette smoking as a
predictor of death from prostate cancer in 348,874 men screened
for the Multiple Risk Factor Intervention Trial. Am. J. Epidemiol.
1996; 143; 1002–1106.
16. Roberts WW, Platz EA, Walsh PC. Association of cigarette
smoking with extraprostatic prostate cancer in young men.
J. Urol. 2003; 169; 512–516.
17. Plaskon LA, Penson DF, Vaughan TL, Stanford JL. Cigarette
smoking and risk of prostate cancer in middle-aged men. Cancer
Epidemiol. Biomarkers Prev. 2003; 12; 604–609.
18. Chiang YY. Hepatocyte growth factor induces hypoxia-related
interleukin-8 expression in lung adenocarcinoma cells. Mol.
Carcinog. 2009; 48; 662–670.
19. Lou H, Du S, Ji Q, Stolz A. Induction of AKR1C2 by phase II
inducers: identification of a distal consensus antioxidant response
element regulated by NRF2. Mol. Pharmacol. 2006; 69; 1662–
1672.
20. Rizner TL, Lin HK, Peehl DM, Steckelbroeck S, Bauman DR,
Penning TM. Human type 3 3alpha-hydroxysteroid dehydroge-
nase (aldo-keto reductase 1C2) and androgen metabolism in
prostate cells. Endocrinology 2003; 144; 2922–2932.
21. Chow KC, Chiou SH, Ho SP et al. The elevated serum interleukin-6
correlates with the increased serum butyrate level in patients with
nasopharyngeal carcinoma. Oncol. Rep. 2003; 10; 813–819.
22. Kim J, Park H, Im JY, Choi WS, Kim HS. Sodium butyrate regulates
androgen receptor expression and cell cycle arrest in human
prostate cancer cells. Anticancer Res. 2007; 27; 3285–3292.
23. Sreekumar A, Poisson LM, Rajendiran TM et al. Metabolomic
profiles delineate potential role for sarcosine in prostate cancer
progression. Nature 2009; 457; 910–914.
24. Twillie DA, Eisenberger MA, Carducci MA, Hseih WS,
Kim WY, Simons JW. Interleukin-6: a candidate mediator of
human prostate cancer morbidity. Urology 1995; 45; 542–
549.
25. Burczynski ME, Lin HK, Penning TM. Isoform-specific induction
of a human aldo-keto reductase by polycyclic aromatic hydro-
carbons (PAHs), electrophiles, and oxidative stress: implications
for the alternative pathway of PAH activation catalyzed by
human dihydrodiol dehydrogenase. Cancer Res. 1999; 59; 607–
614.
26. Wang HW, Lin CP, Chiu JH et al. Reversal of inflammation-
associated dihydrodiol dehydrogenases (AKR1C1 and AKR1C2)
overexpression and drug resistance in nonsmall cell lung cancer
cells by wogonin and chrysin. Int. J. Cancer 2007; 120; 2019–
2027.
27. De Marzo AM, Platz EA, Sutcliffe S et al. Inflammation in prostate
carcinogenesis. Nat. Rev. Cancer 2007; 7; 256–269.
28. Lin TS, Huang HH, Fan YH, Chiou SH, Chow KC. Genetic
polymorphism and gene expression of microsomal epoxide hydro-
lase in non-small cell lung cancer. Oncol. Rep. 2007; 17; 565–572.
29. Chen JT, Lin TS, Chow KC et al. Cigarette smoking induces
overexpression of hepatocyte growth factor in type II pneumo-
cytes and lung cancer cells. Am. J. Respir. Cell Mol. Biol. 2006;
34; 264–273.
30. Lin TS, Chiou SH, Wang LS et al. Expression spectra of matrix
metalloproteinases in metastatic non-small cell lung cancer.
Oncol. Rep. 2004; 12; 717–723.
394 K-H Huang et al.
� 2010 Blackwell Publishing Ltd, Histopathology, 57, 384–394.
Copyright © 2022 FDOKUMEN




















